
Research Collection
Doctoral Thesis
Engineering chondrogenic micro-environments for tissueengineering applications
Author(s): Mhanna, Rami
Publication Date: 2013
Permanent Link: https://doi.org/10.3929/ethz-a-009921642
Rights / License: In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

DISS. ETH NO. 21108
Engineering Chondrogenic Micro-
Environments for Tissue Engineering Applications
DISSERTATION
Submitted to
ETH ZURICH
for the degree of
DOCTOR OF SCIENCES
by
Rami Mhanna
M. Eng., Biomedical Eng., The University of Melbourne
born May 5th, 1982
Lebanese
Accepted on the recommendation of
Prof. Dr. Marcy Zenobi-Wong, examiner Dr. Daniel Eberli, co-examiner
Prof. Dr. Ivan Martin, co-examiner
2013


To My Parents


Often you shall think your road impassable, somber and companionless.
Have will and plod along; and round each curve you shall find a new companion.
Mikhail Naimy


Acknowledgments
I would like to express my deep gratitude to the people that helped realizing
this thesis and turning the PhD dream into reality.
A special thank you to Marcy Zenobi-Wong, my supervisor, for being always
there, supporting and responding to any question I had so quickly. I never had to worry
of sending you an abstract a few days before the deadline, I knew you would respond
with all corrections only a few hours after. Thank you very much for the support in the
last few months of my thesis in which I felt really in need for. I would also like to thank
you for your patience on my questions, initial limited cell lab experience and for
accepting my stubbornness in our meetings. Thank you for the dinners and BBQs at
your place, the lovely Spanish dinner, group retreat I still remember the pancakes. I
would like to express my deep gratitude to Janos Vörös for accepting me as a PhD
student. Thank you Janos for creating such a stimulating research and social
environment, your lab is so attractive that people do not want to leave it. Thank you
for your great input to most of the raised questions and to your subjective productive
advices. Thank you for joining the coffee, beer, soccer and volleyball games. I would
like to express my sincere appreciation to Prof. Ivan Martin and Dr. Daniel Eberli for
accepting to be my co-referees. It is a great honor for me that you review my thesis.
I would like to thank all my collaborators and colleagues in the Find and Bind
consortium, in particular Dr. Matthias Schnabelrauch and Dr. Jana Becher who have
provided me with a variety of materials that were essential for my thesis. I would like
to thank Dr. Markus Rottmar for the work with staurosporine, Dr. Felix Theiss for
believing in my stretching chamber, Dr. Li Zhang and Famin Qiu for the collaboration in
drug delivery using their magnetic helices. I would also like to thank Nadiia Kondratiuk
for her help in the immunostaining, Dr. Eva Beurer for her help with the contact angle
measurements and Celine Gandar for her help with the Qgel experiments.
I would like to thank all the people from CERL and LBB who have helped
realizing this thesis. Thank you Ece, you have been a great support in a large part of
this thesis. Thank you Gemma, your input and biology experience was very valuable in
several aspects of the thesis. Thank you Chris for your amazing images and for the

great time in New Orleans, and the Houston conference was great with you and
Gemma. Thank you Mischa for helping me with the mechanical testing. Thanks Orane
for your advices, discussions, beers, shishas and snowboarding lessons. Thanks Marta
for hosting me in Goteborg, dinners, beers and the visit to Lebanon. Thanks Alex
Larmagnac for the discussions and going out watching the Champions League, going
out with you was always fun. Thanks Bernd for all the nice times we spent going out
and more beers. Thanks Tatiana, it was always so much fun hanging out with you.
Thanks Esther, Dario, Elsa, Deborah, Pablo, Alex Tanno, Mike, Raphael Zahn and
Grüter, Blandine, Geraldine, Pascal, Sophie, Benjamin, Nicolas, Takumi, Andreas
Binkert and Dahlin, Kaori, Ana, Peter, Anette, Gosia, Alfredo, Claudio, Max, Anne,
Martin, Bebeka, Prayanka, Victoria, Tomaso, Harald, Laszlo, Juliane, you have all been
so much fun to work with and I can definitely remember happy moments with each
and every one of you. Special thanks to my students, Philippe, Aditya and Queralt
whose input was totally essential in this thesis. I enjoyed a lot working with all of you
each in a special way. Philippe, I still remember from you “life is always good”. Addy
you were so enthusiastic hard working and fun. Queralt, you were so enthusiastic, hard
working and fun for a bit longer time than Addy. Addy and Queralt, you made the last
year of my thesis so much productive but also so much fun, I am very glad to have met
you and worked with you. I would like to thank Leena Jaatinen for being my soul mate
during a big part of my PhD. Thank you for the great times we spent together, for the
amazing Saint Feliu conference, I will never forget that week. Thank you for all the
advices, support and for being there whenever I needed. Thank you for pushing me
into cycling, I think I will do it for the rest of my life, thanks for the snowboarding
lessons and all the amazing times we had.

Abstract
Mature articular cartilage has very limited ability to self-repair after injury or
disease. In autologous chondrocyte implantation procedures (ACI), chondrocytes are
cultured on tissue culture plastic where they undergo dedifferentiation assuming a
fibroblast-like phenotype and gene expression profile. Dedifferentiated chondrocytes
re-express cartilage specific genes when cultured in materials like alginate hydrogels
which induce a round morphology. The cellular microenvironment plays a crucial role
in directing proliferation, adhesion, metabolic and catabolic activities of cells. Cells
interact with their microenvironment via membrane receptors that recognize changes
in the extracellular matrix (ECM), oxygen levels, mechanical stimuli and small
molecules. The aim of this thesis was to engineer 2D and 3D microenvironments which
will improve conditions for cartilage tissue engineering applications such as ACI. To
achieve this goal we applied the layer-by-layer technique to optimize chondrocyte
culture conditions in 2D substrates and prevent dedifferentiation. We then worked on
designing biomimetic microenvironments by incorporating cartilage-specific molecules
into 3D scaffolds. The scaffolds developed in this thesis could restore the cartilage
phenotype of dedifferentiated chondrocytes or induce chondrogenic differentiation of
stem cells while enhancing cell proliferation. Finally, we investigated the
microenvironment conditions (mechanical load, oxygen tension and stiffness) which
promote expression of superficial zone protein (SZP), a key protein in cartilage
lubrication.
With the use of the layer-by-layer technique we were able to prepare natural
ECM-based films made of type I collagen (Col1)/chondroitin sulfate (CS) or
Col1/Heparin (HN) on stretchable polydimethylsiloxane (PDMS) substrates. The
Col1/CS films were stable in media while Col1/HN films were not. Preliminary studies
showed that chondrocytes did not restore the expression of cartilage markers when
grown on these films which indicates that a 3D environment is probably more
important to maintain the cartilage phenotype than the interaction with ECM
molecules.

Given the results of the 2D system, the focus of the thesis shifted to engineered
3D microenvironments. We observed that sulfation of alginate induced cell
proliferation while maintaining the chondrogenic phenotype of encapsulated
chondrocytes. Remarkably, alginate sulfate hydrogels showed a cartilage-like, opaque
appearance after 5 weeks in culture whereas the unmodified alginate samples
remained translucent. The problems associated with limited availability of healthy
early passage chondrocytes drives the work towards using human mesenchymal stem
cells (hMSCs) for repair. We therefore studied the possibility of inducing
chondrogenesis of hMSCs using a bio-inspired hydrogel. Interactions of MSCs with the
triple helical collagen mimetic, GPC(GPP)5-GFOGER-(GPP)5GPC-NH2, and the
fibronectin adhesion peptide, RGD, were studied in degradable or non-degradable PEG
gels. GFOGER-modified degradable gels induced the highest cell proliferation and were
the most chondrogenic of the investigated conditions. Finally we studied the in-vitro
conditions which promote expression of the superficial zone marker SZP. We observed
that bovine chondrocytes in monolayer showed a drastic decrease in SZP expression,
similar in trend to the commonly reported downregulation of Col2. Chondrocytes
embedded in alginate beads for 4 days re-expressed SZP but not Col2. Cyclic
mechanical strain and normoxic conditions upregulated SZP expression whereas Col2
expression was upregulated only in alginate beads under hypoxic conditions. This work
suggests that the distribution of environmental signals in the different zones of
cartilage in-vivo is responsible for the layer-specific distribution of matrix proteins
throughout the thickness of articular cartilage.
To conclude, the cell microenvironment can be engineered to induce cell
proliferation, maintain the cartilage phenotype of chondrocytes, regulate expression of
cartilage zonal markers and promote chondrogenic differentiation of stem cells. The
results of this thesis provide insight into several crucial aspects of the
microenvironment and should lead the way to the discovery and application of novel
promising materials, such as alginate sulfate derivatives, which could transform
current treatment strategies for repair and regeneration of cartilage lesions.

Riassunto
La cartilagine articolare matura ha una capacità molto limitata di
autorinnovamento in seguito a lesioni o malattie. Nelle procedure d’impianto di
condrociti autologhi (ACI), i condrociti sono solitamente coltivati in fiasche di plastica
per colture cellulari in cui vanno incontro a de-differenziamento, assumendo un
fenotipo e un profilo di espressione genica, tipici dei fibroblasti. I condrociti de-
differenziati ri-esprimono geni specifici della cartilagine quando coltivati in materiali
come idrogel di alginato che inducono una morfologia rotondeggiante. Il
microambiente cellulare gioca un ruolo cruciale nel dirigere proliferazione, adesione, e
attività metaboliche e cataboliche delle cellule. Queste ultime interagiscono infatti con
il microambiente circostante attraverso recettori di membrana che riconoscono
cambiamenti nella matrice extracellulare (ECM), nei livelli di ossigeno, stimoli
meccanici e piccole molecole. Lo scopo di questa tesi è stato quello di ingegnerizzare
microambienti in 2D e 3D che potessero migliorare le condizioni per applicazioni
d’ingegneria tissutale della cartilagine come l’ACI. Per raggiungere tale obiettivo,
abbiamo innanzitutto utilizzato la tecnica "layer-by-layer" che ha permesso di
ottimizzare le condizioni di coltura dei condrociti in substrati 2D e al tempo stesso
prevenirne il de-differenziamento. In seguito abbiamo lavorato alla progettazione di
materiali bio-mimetici in cui molecole specifiche della cartilagine fossero incorporate
in scaffold 3D. Gli scaffold sviluppati in questa tesi potrebbero essere utili per
ristabilire il fenotipo cartilagineo dei condrociti de-differenziati o indurre il
differenziamento condrogenico di cellule staminali, aumentando al tempo stesso la
proliferazione cellulare. Infine abbiamo investigato le condizioni del microambiente
(carico meccanico, tensione di ossigeno e rigidezza) che promuovessero l’espressione
della "proteina della zona superficiale" (SZP), una proteina chiave nella lubrificazione
della cartilagine.
Con l’uso della tecnica "layer-by-layer" abbiamo preparato dei film basati sulla
matrice extracellulare naturale e contenenti collagene di tipo I (Col1)/condroitin
solfato (CS) o Col1/eparina (HN) su substrati estensibili di polidimetilsilossano (PDMS).
Tra i due tipi di film, solo quelli contenenti Col1/CS si sono rivelati stabili nel terreno di

coltura. Studi preliminari hanno dimostrato che i condrociti non erano capaci di
ripristinare l’espressione di marcatori della cartilagine se cresciuti su questi film. Tal
evidenza sperimentale indica che un ambiente tridimensionale è probabilmente più
importante dell’interazione con le molecole della matrice extracellulare, al fine di
mantenere il fenotipo cartilagineo.
Dati i risultati del sistema bidimensionale, abbiamo rivolto l’attenzione verso
l’ingegnerizzazione di microambienti in 3D. Abbiamo osservato che la solfatazione di
alginato promuoveva proliferazione cellulare, mantenendo al tempo stesso il fenotipo
condrogenico di condrociti incapsulati. Sorprendentemente gli idrogel di alginato
solfato assumevano un aspetto opaco simile alla cartilagine dopo 5 settimane in
coltura mentre i campioni di alginato non modificato rimanevano traslucidi. I problemi
associati alla disponibilità limitata di condrociti sani a passaggio precoce indirizza il
lavoro verso l’utilizzo di cellule staminali mesenchimali umane (hMSCs) per la
rigenerazione. Di conseguenza abbiamo studiato la possibilità di indurre la
condrogenesi di cellule hMSCs usando un idrogel bio-ispirato. In particolare, abbiamo
investigato le interazioni di hMSCs con un peptide che mima il collagene a tripla elica,
GPC(GPP)5-GFOGER-(GPP)5GPC-NH2, e con un peptide di adesione tipico della
fibronectina, RGD, all’interno di gel di polietilenglicole (PEG), degradabili e non. Tra le
condizioni prese in esame, i gel contenenti il peptide GFOGER e degradabili si sono
rilevati i migliori sia per l’induzione di proliferazione cellulare che per la condrogenesi.
Infine, abbiamo condotto uno studio per identificare le condizioni in vitro che
promuovessero l’espressione di SZP, il marcatore della zona superficiale della
cartilagine. A tale proposito, abbiamo osservato che i condrociti bovini cresciuti come
mono-strato mostravano una notevole diminuzione nell’espressione di SZP, con una
tendenza simile alla riduzione di Col2 comunemente riportata. Condrociti incorporati
in perline di alginato per 4 giorni ri-esprimevano SZP ma non Col2. Una sollecitazione
meccanica ciclica e condizioni di normossia inducevano l’espressione di SZP mentre
l’espressione di Col2 risultava aumentata solo in perline di alginato mantenute in
ipossia. Questo lavoro suggerisce che la distribuzione strato-specifica di proteine della
matrice attraverso lo spessore della cartilagine articolare.

In conclusione, il microambiente cellulare può essere ingegnerizzato per
indurre proliferazione cellulare, mantenere il fenotipo cartilagineo, regolare
l’espressione di marcatori specifici delle diverse zone della cartilagine e promuovere il
differenziamento condrogenico delle cellule staminali. I risultati di questa tesi hanno
permesso la comprensione di diversi aspetti cruciali del microambiente e aprono la
strada alla scoperta e l’applicazionedi nuovi materiali ideali per l’ingegneria tissutale
della cartilagine come l’alginato solfato.


Contents
1 Tissue Engineering & Articular Cartilage ................................................... 1
1.1 The structure of articular cartilage ........................................................ 2
1.2 Cartilage disease, injury and management ............................................ 4
1.2.1 Non-surgical approaches (focus on chondroitin sulfate) .................. 5
1.2.2 Surgical approaches .......................................................................... 6
1.3 Autologous chondrocyte implantation (ACI) ......................................... 7
1.3.1 History and development of ACI ....................................................... 7
1.3.2 The limitations of ACI ....................................................................... 8
1.4 3D culture systems ................................................................................ 9
1.4.1 Preventing dedifferentiation in monolayer cultures....................... 10
1.4.2 Biomimetic systems ........................................................................ 11
1.4.3 Mechanical stimulation .................................................................. 12
2 Scope of the thesis ................................................................................ 15
3 Materials and methods .......................................................................... 19
3.1 Materials ............................................................................................. 19
3.2 Instruments ......................................................................................... 21
3.3 Protocols .............................................................................................. 24
4 Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates ................................................................................ 35
4.1 Current monolayer culturing techniques ............................................. 35
4.2 Build-up of Col1/CS and Col1/HN films on PDMS ................................ 37
4.3 Effect of substrate on film build-up ..................................................... 41
4.4 Film topography .................................................................................. 42

4.5 Film thickness and stability .................................................................. 44
4.6 Assessment of cell adhesion and integrin mediated spreading ........... 46
4.7 Chapter summary ................................................................................ 51
5 Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers ... 53
5.1 Improving chondrogenic performance of chondrocytes in 3D may be
achieved using biomimetic materials. ................................................... 53
5.2 Preparation and characterization of sulfated alginate ......................... 55
5.3 Morphology of chondrocytes encapsulated in sulfated alginate (DSs =
0.8) ........................................................................................................ 57
5.4 Assessment of cell proliferation within the hydrogels ......................... 59
5.5 Decoupling stiffness from cell spreading ............................................. 60
5.6 RhoA and integrin signalling of chondrocytes in alginate sulfate ........ 62
5.7 Cyclin D1 expression is upregulated in alginate sulfate samples ......... 64
5.8 Expression of cartilage markers for dedifferentiated chondrocytes within
alginate sulfate hydrogels ...................................................................... 65
5.9 Immunohistological staining and gross appearance of alginate sulfate
hydrogels ............................................................................................... 66
5.10 Chapter summary ............................................................................ 69
6 GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells ......... 71
6.1 Chondrogenic differentiation and the microenvironment ................... 71
6.2 Hydrogel modification did not affect mechanical properties .............. 73
6.3 Cell viability ......................................................................................... 74
6.4 Cell morphology .................................................................................. 75
6.5 Cell proliferation was highest in RGD and GFOGER degradable hydrogels
............................................................................................................. 77

Contents
6.6 Gene expression .................................................................................. 78
6.7 GAG production in peptide-modified gels ........................................... 79
6.8 Histology and immunostaining ............................................................ 80
6.9 Chapter summary ................................................................................ 84
7 Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression ........................................................................ 85
7.1 Chondrocyte dedifferentiation and superficial zone protein ............... 85
7.2 Both SZP and Col2 undergo dedifferentiation during serial passaging 87
7.3 Redifferentiation of serially passaged primary chondrocytes .............. 88
7.4 Design and evaluation of a 3D tension and compression chamber
compatible with the strex strain machine ............................................. 90
7.5 Application of cyclic mechanical strain to 2D and 3D constructs ......... 92
7.6 Effect of mechanical strain on SZP expression ..................................... 94
7.7 Effect of oxygen tension on expression of SZP .................................... 95
7.8 Cell morphology and the expression of SZP ......................................... 98
7.9 Chapter summary .............................................................................. 105
8 Conclusions and outlook ...................................................................... 107
9 References .......................................................................................... 109
Curriculum Vitae .................................................................................................. 125



List of Figures and Tables
Figure 1. 1 | Articular cartilage structure and composition ......................................................................... 4
Figure 1. 2. | ACI procedure as described in Brittberg et al. 1994 ............................................................... 8
Figure 1. 3. | Schematic representation of the build-up of ECM films and application of load on seeded
cells. ............................................................................................................................................................ 11
Table 3. 1. | Genes used in qRT-PCR analysis ............................................................................................ 23
Figure 4. 1. | Buid-up of Col1/CS multilayer ............................................................................................... 38
Figure 4. 2. | Buid-up of Col1/HN multilayer .............................................................................................. 39
Figure 4. 3. | Buid-up exponential (Col1/HN) vs. linear (Col1/CS) ............................................................. 40
Figure 4. 4. | Build-up of Col1/CS on gold .................................................................................................. 42
Figure 4. 5. | Morphology of Col1/CS and Col1/HN films ........................................................................... 43
Figure 4. 6. | Films’ thickness and stability. ............................................................................................... 45
Figure 4. 7. | Morphology of bovine chondrocytes seeded ECM films ...................................................... 48
Figure 4. 8. | Integrin mediated cell spreading .......................................................................................... 50
Figure 5. 1. | Reaction scheme for the synthesis of alginate sulfate ......................................................... 56
Figure 5. 2. | Morphology of cells encapsulated in alginate and alginate sulfate ...................................... 58
Figure 5. 3 | Cell proliferation in gels of various alginate sulfate (AlgSulf) contents ................................. 60
Figure 5. 4 | Mechanical properties of alginate sulfate (AlgSulf) hydrogels and effect on cell proliferation
Figure 5. 5. | Integrin mediated cell spreading and proliferation within alginate sulfate (AlgSulf)
hydrogels and RhoA activity. ...................................................................................................................... 63
Figure 5. 6. | Cyclin D1 gene expression. ................................................................................................... 65
Figure 5. 7. | Expression of relevant cartilage markers. ............................................................................. 66
Figure 5. 8. | Gross appearance and immunostaining of matrix molecules. .............................................. 67
Figure 6. 1. | Compressive moduli of the hydrogels.. ................................................................................ 74
Figure 6. 2. | Cell viability in the hydrogels.. .............................................................................................. 75
Figure 6. 3. | hMSCs cultured for 21 days in chondrogenic media in modified PEG gels prior to fixation
and staining for nuclei (DAPI, blue) and actin filaments (phalloidin, red).. ................................................ 76
Figure 6. 4. | Assessment of cell proliferaiton............................................................................................ 77
Figure 6. 5. | Gene expression of relevant cartilage markers .................................................................... 79
Figure 6. 6. | GAG/DNA quantification in the various conditions .............................................................. 80
Figure 6. 7. | Alcian blue staining of hMSC cultured in pellets and in PEG gels of different compositions.
hMSC cultured in pellets with control medium .......................................................................................... 81
Figure 6. 8. | Type II collagen immunostaining of hMSC cultured in pellets and in PEG gels of different
compositions .............................................................................................................................................. 82
Figure 7. 1. | Effect of chondrocyte dedifferentiation on gene expression of SZP and type II collagen. ... 87
Figure 7. 2. | Redifferentiation of serially passaged chondrocytes ............................................................ 89
Figure 7. 3. | Design for the 3D molds A and the final 3D chamber B. ....................................................... 91

List of Figures
Table 7. 1. | Actual strains measured in the alginate gel and the corresponding strain setting in the
STREX device. .............................................................................................................................................. 92
Figure 7. 4. | Design for application of 3D cyclic strain. ............................................................................. 93
Figure 7. 5. | The effect of mechanical strain on SZP mRNA levels was measured by qRT-PCR ................ 95
Figure 7. 6. | The effect of oxygen tension on SZP and Col2 mRNA levels was quantified by qRT-PCR. .... 97
Figure 7. 7. |Morphology of chondrocytes cultured in 2D and 3D substrates. .......................................... 99
Figure 7. 8. | Effect of cell morphology and dimensionality on SZP gene expression .............................. 100
Figure 7. 9. | Effect of RGD sequences on morphology and SZP gene expression. .................................. 102
Figure 7. 10. | Morphology and gene expression of SZP for chondrocytes cultured in 3D MMP sensitive
PEG hydrogels with and without RGD peptides ....................................................................................... 104

Chapter 1
1
1 Tissue Engineering & Articular Cartilage
Tissue engineering is an interdisciplinary field that utilizes cells, biomaterials,
biochemical (e.g. growth factors) and physical (e.g. mechanical loading) signals, as well
as their combinations to generate tissue-like structures [1]. The goal of tissue
engineering is to provide biological substitutes that can maintain, restore or improve
the function of damaged tissues [2]. Skin grafts were the first tissue-based therapies
and introduction of techniques to preserve cells and tissues enabled allograft skin
banking [3-5]. However, limited donor availability and rejection of the grafts by the
immune system drove the concept of in-vitro grown tissues. Although the first tissue
engineered skin products were introduced in the late 1970s and early 1980s giving rise
to modern tissue engineering, the term “tissue engineering” was only coined in 1987
[6-9]. The success of engineering skin grafts boosted interest in applying similar
concepts to other tissues and organs [10]. However, the relatively simple structure, the
limited vascular demands of skin and the ease of growing keratinocytes in-vitro are not
common to most tissues. The dream of regenerating tissues in-vitro faced major
hurdles associated with the engineering of complex 3-dimentional vascularized
multicellular tissues.
Cartilage, an avascular unicellular tissue, was the next target for tissue
engineering which generated early success. The high prevalence of osteoarthritic
disease as well as cartilage injuries incurred by athletes create a large market for
cartilage tissue engineering solutions [1]. Although engineering cartilage at first might
seem to be an easy task, the tissue’s stratified structure, unique tensile, compressive
and lubricative properties and compositional and ultrastructural complexity makes it
difficult if not impossible with current technologies to fully replicate the tissue [11-13].
Current available tissue engineering strategies for cartilage repair do not recapitulate
the cartilage microenvironment which provides a multitude of cues necessary for
cartilage homeostasis [14].

Introduction
2
1.1 The structure of articular cartilage
Articular cartilage is the tissue that covers the surface of articulating joints
providing an almost frictionless interaction and absorbing shocks from daily physical
activities. Articular cartilage is composed of extracellular matrix containing a single
type of sparsely distributed highly specialized cells (chondrocytes) and lacking
vasculature, lymphatic vessels and nerves (Figure 1.1) [13]. Chondrocytes vary in
morphology and metabolic activity depending on their location within the thickness of
the tissue. The distribution and organization of the extracellular matrix also varies
allowing the identification of four different zones from the surface of articular cartilage
to the subchondral bone: the superficial zone, transitional zone, deep zone and
calcified cartilage zone. The superficial zone is the thinnest zone comprising an
acellular thin sheet of collagen fibrils. The ellipsoid-shaped chondrocytes below this
sheet are arranged with their major axis parallel to the cartilage surface. The thin sheet
of collagen fibrils provides this layer with higher tensile strength compared to deeper
layers allowing it to resist high shear forces on the surface of the tissue. The dense
sheet of collagen also acts as a barrier preventing the leakage of cartilage molecules
out of and/or the entrance of antibodies and cytokines into the cartilage. Disruption of
the collagen layer is one of the first observed physical alterations in experimentally
induced degeneration of articular cartilage [15]. Chondrocytes of this layer synthesize
different amounts and types of matrix proteins compared to deeper layers,
particularly, they synthesize superficial zone protein (SZP) which is highly homologous
to lubricin [16], a major contributor of cartilage lubrication [17]. The transitional zone
has several times the volume of the superficial zone with a matrix composition and cell
morphology intermediate between the superficial and deep zones. Cells of the
transitional zone have a spheroidal morphology and synthesize thicker collagen fibrils
and proteoglycans at a higher concentration compared to cells of the superficial zone.
The deep zone has the highest concentration of proteoglycans and thickest collagen
fibrils. Chondrocytes in the middle layer assume a spheroidal morphology and are
often aligned in columns perpendicular to the surface of cartilage. The calcified

Chapter 1
3
cartilage zone separates the deep zone and the subchondral bone and provides a
physical barrier between cartilage and bone.
The cellular microenvironment plays a pivotal role in the function of cells [18].
In chondrocytes several factors are known to influence chondrocyte migration,
proliferation, matrix synthesis and degradation. Moreover chondrocytes of the
different layers in cartilage have a different microenvironment as shown in Figure 1.1.
Chondrocytes are surrounded by an extracellular matrix which is predominantly
composed of type II collagen and proteoglycans [13]. Chondrocytes possess cell
membrane receptors called integrins which mediate interactions with matrix proteins.
Integrin α10β1 recognizes the GFOGER sequences in collagen [19] while the receptor
CD44 mediates cell interaction with hyaluronic acid and chondroitin sulfate [20]. The
oxygen levels vary through the cartilage thickness from 7-10% on the surface down to
0.1% near the subchondral bone [21]. Mechanical stimulation also varies in type and
magnitude throughout cartilage [22]. In stem cells, the microenvironment changes
considerably throughout the stages of chondrogenesis. Initially, MSCs produce
fibronectin followed closely by an increase in type I collagen expression which
promotes their motility leading to condensation, a pre-requisite step to MSC
chondrogenesis. Differentiation into chondrocytes follows with cells decreasing
fibronectin expression and replacing type I collagen with up-regulation of type II
collagen and aggrecan [23].

Introduction
4
Figure 1. 1 | Articular cartilage structure and composition. Electronic micrographs of cells from the
various cartilage zones (left panel), full thickness articular cartilage (middle panel) and collagen and
matrix composition and structure (right panel). Reproduced with permission from [24, 25].
1.2 Cartilage disease, injury and management
Osteoarthritis is a common joint disease affecting over 200 million people
worldwide. In the United States alone, more than 40 million people are affected by
arthritis (approximately 15% of the total population) and the number is estimated to
grow to 60 million by 2020 [26]. Osteoarthritis is a low mortality disease which causes
pain and can lead to disability in advanced stages. Osteoarthritis is primarily a disease
of the aging population [27], but is occurring more commonly in the younger
population especially those engaged in high-impact sport activities. Sports-related
increase in the risk of osteoarthritis in soccer and ice hockey athletes has been
reported to be related to knee injuries and not to physical activity during sports
engagement [28].

Chapter 1
5
Articular cartilage has very limited ability to self-repair due to its avascular
nature, limited proliferative ability of mature chondrocytes and limited access to
progenitive repair cells. Furthermore, the ability of healthy chondrocytes to migrate to
injured sites is restricted by the surrounding extracellular matrix (ECM) [29]. Currently
no treatment is available to fully regenerate damaged cartilage. Treatments for
cartilage can be categorized into non-surgical and surgical treatments, with the choice
being dependent on the severity of the injury. Tissue engineering holds strong promise
in providing functional replacement for cartilage lesions. However, improvement of
the three elements of tissue engineering namely cells, biomaterials and culture
conditions are necessary to achieve ideal reconstruction of cartilage defects.
1.2.1 Non-surgical approaches (focus on chondroitin sulfate)
The pharmacological agents used for osteoarthritis treatment can be
categorized into three main groups: 1) non-steroidal anti-inflammatory agents and
analgesics (NSAIDs); 2) symptomatic slow-acting drugs for OA (SYSADOA); 3)
chondroprotective or truly disease-modifying agents known as disease-modifying OA
drugs (DMOADs) [30]. Paracetamol is a safe recommended oral analgesic but does not
always provide sufficient relief from severe pain [31, 32]. NSAIDs are commonly used
when significant inflammation occurs. However, several serious side-effects are
associated with long-term use of NSAIDs including gastric erosion, kidney injury, ulcer
disease, acute renal complications among others [33-36]. NSAIDs are useful for pain
relief but they do not have direct influence on the progression of osteoarthritis,
therefore safer disease modifying agents are required. Some ECM molecules such as
chondroitin sulfate (CS), hyaluronan and glucosamine are believed to have
chondroprotective properties [27, 37-42]. In cartilage, chondroitin sulfate is attached
to the core protein of proteoglycans which contributes to the tissue’s compressive
properties by entrapping water. Chondroitin sulfate has been shown to stimulate ECM
production, supress inflammation and inhibit cartilage degeneration [40, 43-46].
Chondroitin sulfate and glucosamine can be taken orally unlike hyaluronan which

Introduction
6
requires injection. The safety and tolerability of chondroitin sulfate were confirmed in
previous reports and it is currently available in several products such as Condrosulf®
and Matrix® [47, 48].
1.2.2 Surgical approaches
For patients who are diagnosed with full-thickness cartilage injuries and where
non-surgical pharmacological treatments fail in restoring function and pain relief, a
surgical procedure may be required. The current surgical approaches include lavage
and debridement, microfracture (marrow stimulation), osteochondral grafting and cell
based therapies (autologous chondrocyte implantation). Lavage involves washing away
small molecules, debris and inflammatory mediators and debridement is the removal
of larger cartilage, meniscal fragments and osteophytes. Lavage and debridement can
provide temporary relief but does not directly address the cartilage injury.
Microfracture or marrow stimulation is the creation of holes in the subchondral bone
which causes bleeding and the formation of a fibrin clot. Bone marrow stem cells can
then migrate to the clot and form a fibrocartilage repair tissue. The inferior mechanical
and biochemical properties of the resulting repair tissue are major disadvantages of
the method. Osteochondral grafting is usually done by taking autologous
osteochondral plugs from low-weight-bearing regions of cartilage and repairing the
chondral injuries with a mosaic of harvested plugs. The main drawbacks of this method
are donor site morbidity and low integration of the grafts with the surrounding tissue.
The gap between the graft and the surrounding cartilage is normally filled with
fibrocartilage which leads to strong alterations in load distribution. Finally autologous
chondrocytes implantation (ACI) is the process of using autologous chondrocytes
which are then expanded in the laboratory to fill the cartilage defect [49, 50]. ACI is a
promising technique for the treatment of cartilage and is described in more details in
the following sections.

Chapter 1
7
1.3 Autologous chondrocyte implantation (ACI)
Autologous chondrocyte implantation is the first approach that applies the
principle of tissue engineering to treat cartilage defects. In this thesis, we focus on
providing the proper microenvironment to enhance current culturing techniques with
the aim of addressing the areas where ACI currently falls short.
1.3.1 History and development of ACI
Autologous chondrocyte transplantation was developed in the late 1980s by
Peterson and co-workers and described in humans by Brittberg et al. in 1994 [51-53].
In this procedure a biopsy of healthy cartilage tissue (approx. 300 milligrams) is taken
from a low-weight-bearing region of the knee. The tissue is enzymatically digested in
the laboratory to isolate the cells which are then are grown in-vitro to reach sufficient
cell numbers. In the original embodiment, a periosteal flap large enough to cover the
defect is harvested from the proximal medial tibia and sutured to the surface of the
cartilage leaving a small gap for cell injection. The spaces between the sutures are
filled with fibrin glue creating a water-tight seal that prevents cell leakage and cells are
injected to fill the defect site (Figure 1.2). Although ACI presented a major
advancement in cartilage therapy, periosteal hypertrophy encouraged the research for
improved repair strategies [54, 55]. In order to address the above problems, a
membrane based on type I/III porcine collagen has been developed and is used to
replace the periosteal flap in ACI procedures [56-58]. The membrane commercialized
as (Chondrogide) has a porous surface on the side that faces the defect which allows
cell attachment and a smooth compact surface that prevents cell leakage.
Research is currently focused on the use of cells from more promising sources,
chondroinductive/conductive biomaterials, gene therapy, simplifying the surgical
procedures, mechanical stimulation and preconditioning of cells [14]. The work in this
thesis contributes to the research on the use of stem cells, biomaterials with

Introduction
8
biochemical cues and cell affinity, growth factors and in-vitro preconditioning with
mechanical stimulation.
Figure 1. 2. | ACI procedure as described in Brittberg et al. 1994. Reproduced with permission from
[52], Copyright Massachusetts Medical Society
1.3.2 The limitations of ACI
ACI relies on the growth of cells in the lab in monolayers which causes the loss
of the cartilage phenotype or what is commonly known as dedifferentiation [59].
Chondrocyte dedifferentiation is associated with morphological and gene expression
changes where cells behave more as fibroblasts [60, 61]. Dedifferentiated
chondrocytes produce type I collagen rather type II collagen and thus cells implanted
in ACI procedures often produce fibrous tissue and not hyaline cartilage [62]. The
current research focuses on developing a variety of methods to either prevent
dedifferentiation during serial expansion or to restore the cartilage phenotype of cells
prior or after the delivery to the defect site [63, 64]. Another disadvantage of ACI is

Chapter 1
9
that it relies on the use of articular chondrocytes which are limited in supply. Much of
the research now focuses on using different cell sources which can provide the same
repair properties as chondrocytes [65, 66]. Stem cells are a promising cell source and
various studies have reported the induction of chondrogenesis using TGFβ and
dexamethasone [67, 68]. Materials which are able to induce proliferation while
maintaining the cartilage phenotype of chondrocytes or inducing chondrogenesis of
stem cells would be highly desirable.
1.4 3D culture systems
Chondrocytes cultured on tissue culture plastic undergo dedifferentiation
where they assume a fibroblast like phenotype and alter their gene expression
producing proteins typical of fibroblasts [62]. Chondrocytes gene expression has been
shown to be tightly linked to its morphology where a round cell is typical of a
differentiated chondrocyte while a flattened morphology triggers chondrocyte
dedifferentiation [60, 69-71]. Culturing chondrocytes in 3D gels such as agarose [60] or
alginate [72] preserves the cartilage phenotype. Nevertheless, chondrocytes
proliferation in alginate and agarose is very limited thus it might not be the most
suitable method to culture chondrocytes when high cell yields are required. Synthetic
gels such as poly(ethylene glycol) (PEG) are attractive for 3D culturing of cells, as they
can be functionalized with specific adhesion sequences, growth factor binding sites
and protease-sensitive motifs [73-76]. Providing adhesion sequences within these
hydrogels would increase cell proliferation and may potentially maintain the cartilage
phenotype. Moreover, application of mechanical stimulation and low oxygen tension
can also help restoring the cartilage phenotype. Attempts to maintain the cartilage
phenotype in 2D cultures by cell spreading restriction or drugs have also been pursued
and are further discussed below.

Introduction
10
1.4.1 Preventing dedifferentiation in monolayer cultures
Experiments that were previously performed to achieve a cartilage phenotype
in 2D focus mainly on preventing cell spreading such as inhibiting RhoA/ROCK
signalling using Y-27632. Consequently, preventing actin polymerization by adding
cytochalasin D or stabilizing existing actin filaments with jasplakinolide helped recover
the cartilage phenotype in dedifferentiated cells [70, 71]. The addition of concanavalin
A to chondrocyte cultures also caused cell rounding and increased proteoglycan
synthesis [77]. In these experiments it was clearly demonstrated that a round
chondrocyte phenotype correlates with maintaining the cartilage phenotype. In a
collaboration with Rottmar et al. (submitted to Exp. Cell. Biol.), it was concluded that
the cartilage phenotype can be maintained in spread cells with a disrupted
cytoskeleton. A more high-tech approach involves the use of micro-patterned
substrates where the cells can be constrained to a round morphology [63, 78]. While
these approaches were shown to be beneficial, proliferation is constrained by the
patterns and thus the system does not serve the requirement of high cell numbers for
ACI. On the other hand the use of drugs might be associated with toxicity and would
require costly clinical trials before it is accepted for use in patients. An approach that
may help address these issues is to engineer substrates that allow cell proliferation
while maintaining the cartilage phenotype using safe molecules and possible external
stimuli.
The layer-by-layer (LbL) technique has been widely used to produce nanofilms
for biomedical applications [79, 80]. The LbL approach allows the build-up of thin films
simply by the alternating deposition of polyanions and polycations [81]. Since the
introduction of the LbL technique, a large body of research has been produced
describing the build-up of films of different compositions [82-89]. The majority of
these films rely on synthetic polymers with relatively high charge densities. Films such
as PSS/PAH or PLL/HA promote cell adhesion and are not cytotoxic [83, 84].
Nevertheless, polymers that diffuse out of the layers and could be taken up through
the cell membrane raise strong concerns of cytotoxicity [87]. The growth of cells has

Chapter 1
11
been shown to be dependent on the stiffness, chemistry and charge of the films [82-
86].
The preparation of films from natural polymers is limited to a few combinations
which are rarely stable [82, 88, 89]. The build-up can be achieved using ECM molecules
such as collagen which is positively charged at a pH lower than 5.5 and CS or any other
negatively charged molecule [90, 91]. Chondrocytes cultured in contact with ECM
molecules and concomitantly exposed to mechanical load are expected to have a high
chondrogenic activity optimal for cartilage tissue engineering applications (Figure 1.3).
Such systems resemble the natural environment in which chondrocytes exist and thus
are expected to be more physiologic.
Figure 1. 3. | Schematic representation of the build-up of ECM films and application of load on seeded
cells.
1.4.2 Biomimetic systems
Biomimetic systems are systems designed based on biological structures [92].
Biomimetic approaches were shown to be beneficial in cartilage tissue engineering
applications. Mimicking the natural chondrocyte round morphology in-vitro by
encapsulation in alginate and agarose leads to restoration of the cartilage phenotype
[60, 72]. Molecules that are naturally present in cartilage such as chondroitin sulfate
and hyaluronic acid were shown to have chondroprotective properties and are used
for cartilage treatment [27, 49]. Scaffolds containing these molecules or certain

Introduction
12
moieties of these molecules such as sulfate groups might have beneficial effects on
chondrocytes. Mechanical stimulation at physiological levels is known to stimulate
expression and synthesis of cartilage markers [93, 94]. Chondrocytes cultured in
hypoxic conditions were shown to synthesize more type II collagen and aggrecan [21,
95]. The fibronectin cell binding domain RGD has been shown to be involved in cell
adhesion, proliferation and differentiation of stem cells [96, 97]. However, the spread
morphology induced by this sequence may not be optimal for chondrocytes, which
form primary interactions with collagen 2 and hyaluronic acid in native cartilage [96].
Recently the GFOGER peptide was identified as the collagen sequence responsible for
chondrocyte adhesion [19, 98]. Incorporation of the GFOGER sequence in hydrogels
would provide a better cartilage mimic than the commonly used RGD sequences. The
GFOGER peptide might improve the chondrogenic performance of 3D encapsulated
chondrocytes and chondrogenesis of stem cells.
1.4.3 Mechanical stimulation
Mechanical stimulation varies throughout cartilage where cells of the
superficial layer are subject to shear, stress and compressive forces, cells of the
transitional layer and deep layer are subject to compressive forces and hydrostatic
pressure and cells of the calcified layer are subject mainly to hydrostatic pressure [22].
Mechanical loading experiments have been widely performed to obtain information
about mechanisms of cell mechanotransduction and to enhance proliferation, gene
expression, and protein synthesis in chondrocytes [94]. Mechanical signal sensing by
cells may occur through changes in the cell membrane sensed through integrins and
ion channels and their downstream signalling pathways [94, 99]. It has been shown
that in chondrocytes the application of compression at physiological levels increases
matrix protein and GAG synthesis [100]. Excessive loading, on the other hand, leads to
the production of metalloproteinases (MMPs) and aggrecanases (ADAMTS-5) that
degrade the ECM proteins [101]. Systems have been developed for the application of
tensile load, compressive load, hydrostatic pressure, shear and perfusion [102-108].

Chapter 1
13
Notably, stem cell fate can be steered through the application of phenotypic loading.
To illustrate: application of tensile loads help steer stem cells towards ligament [103],
tendon [104], or bone tissue depending on the tensile load parameters [105, 109]
while shear loads can help stem cells differentiate towards cardiac muscle [110] or
endothelial cells [107]. Finally hydrostatic pressure or compression can lead to
chondrogenic differentiation [94, 111-113]. Mechanical stimulation of chondrocytes
modulates the expression of chondrocyte marker genes such as type II collagen,
aggrecan and COMP and the synthesis of glycosaminoglycans (GAGs) [93, 106, 114,
115]. Expression of the superficial zone marker, superficial zone protein is influenced
by shear, stress and surface motion [116-118]. The mechanisms through which
chondrocytes perceive load and react to their surrounding environment are not fully
elucidated but may comprise stretch activated ion-channels and integrins [119, 120].
Understanding these mechanisms will provide basis for developing new tools in the
area of cartilage tissue engineering and possibly discovering new useful molecules in
the treatment of osteoarthritis and cartilage injuries.

Introduction
14


Chapter 2
15
2 Scope of the thesis
Cartilage disease and injury affects over 200 million people worldwide.
Osteoarthritis of cartilage is more prevalent in the aging population causing an
immense socioeconomical burden on societies. Cartilage does not heal after injury or
disease and thus requires medical intervention. Currently no disease modifying drugs
are clinically available and the surgical procedures are not yet capable of fully
recovering damaged articular cartilage. Cell based techniques such as ACI are
promising however they have limitations caused mainly by the dedifferentiation of the
transplanted cells. The use of biomimetic materials to engineer 2D and 3D substrates
for cartilage tissue engineering would potentially lead to a better outcome for ACI-like
procedures. This thesis focuses on providing the right microenvironment in
monolayers and 3D scaffolds for cartilage tissue engineering applications.
In chapter 4 we used the LbL approach to improve cell culturing conditions and
prevent dedifferentiation of in-vitro cultured chondrocytes. The layer-by-layer (LbL)
technique was used before to produce nanofilms made of ECM macromolecules.
However, most of these studies resulted in either non-stable or cell repellent films. The
build-up was also never characterized on polydimethylsiloxane (PDMS) which is a
common elastic biomaterial used in a wide range of biomedical applications. In
particular to our interest, PDMS can be used in combination with the produced natural
films to apply mechanical strain on adherent cells. We assessed the topography, build-
up, thickness and stability of type 1 collagen (Col1)/CS or Col1/heparin (HN) on PDMS
substrates using quartz crystal microbalance with dissipation (QCM-D) and atomic
force microscopy (AFM). Integrin-mediated cell adhesion was assessed by studying the
cytoskeletal organization of mammalian primary chondrocytes seeded on different end
layers and number of layers. This work provided important information on the build-up
of films made from cartilage relevant molecules on a stretchable substrate. However,

Scope
16
the chondrocytes exhibited a fibroblastic morphology on these films which lead us to
focus on 3D scaffolds in the subsequent chapters of this thesis.
In chapter 5 we focused on developing a biomimetic 3D scaffold for cartilage
tissue engineering. Inspired by the presence of sulfate groups in cartilage, we
hypothesized that sulfation of alginate which is known to restore the cartilage
phenotype would result in a more chondrogenic material. Hydroxyl groups of alginate
were converted to sulfates by incubation with sulfur trioxide-pyridine complex
(SO3/pyridine), yielding a sulfated material crosslinkable with calcium chloride.
Dedifferentiated passage 3 bovine chondrocytes were encapsulated in alginate and
alginate sulfate hydrogels for up to 35 days. Cell morphology, proliferation, expression
and synthesis of type II collagen, type I collagen and aggrecan were assessed by
quantitative real time PCR (qRT-PCR) and immunohistochemistry. The alginate sulfate
hydrogel provided a 3D microenvironment which promoted both chondrocyte
proliferation and maintenance of the chondrogenic phenotype. The results of this
study were remarkably striking especially concerning the unexpected increase in cell
proliferation emphasizing how small microenvironmental changes can completely alter
the response of cells encapsulated in a 3D scaffold.
Much of today’s research focuses on exploring new cell sources such as stem
cells for cartilage tissue engineering applications. We therefore studied in chapter 6
the possibility of inducing chondrogenesis of human mesenchymal stem cells (hMSCs)
by controlling microenvironmental cues involved in adhesion and matrix degradation
in stem cells. Interactions of human mesenchymal stem cells (hMSCs) with the triple
helical collagen mimetic, GPC(GPP)5-GFOGER-(GPP)5GPC-NH2, and the fibronectin
adhesion peptide, RGD, were studied in MMP-sensitive (degradable) or MMP-non-
sensitive (non-degradable) PEG gels formed by Michael addition chemistry. Viability,
proliferation, cytoskeletal morphology, and chondrogenic differentiation of
encapsulated hMSCs were evaluated. The type of adhesion sequence and whether it
was presented together with MMP-sensitive motifs or not had a profound influence on
the response of encapsulated hMSCs. The GFOGER peptide enhanced proliferation in

Chapter 2
17
degradable PEG gels and provided a better chondrogenic microenvironment compared
to the RGD peptide or gels with no adhesion peptide.
Finally, the microenvironmental conditions which promote expression of
cartilage zonal markers such as SZP in the superficial zone and type II collagen in the
lower cartilage zones were studied in chapter 7. Chondrocytes from 6 months old
calves were expanded in monolayer culture and the expression of SZP in alginate bead
and monolayer culture was quantified with qRT-PCR and immunostaining. The effect of
cyclic tensile strain and oxygen tension on expression of SZP and type II collagen in 2D
and 3D cultures was quantified. Bovine chondrocytes in monolayer showed a drastic
decrease in SZP expression which could be fully restored in alginate beads after 4 days
of culture. This finding indicates that although cells of the superficial zone have a more
discoidal shape than cells of the deeper layers they still require a 3D environment to
maintain their phenotype. Cyclic mechanical strain and normoxic conditions improved
SZP expression whereas Col2 was upregulated only in alginate beads under hypoxic
conditions. The results of this chapter indicate that engineering stratified articular
cartilage will require gradients both of oxygen and mechanical signals.
The thesis concludes with a summary of findings and an outlook to challenges
and future possibilities.

Scope
18

Chapter 3
19
3 Materials and methods
In this chapter, descriptions of the materials, instruments and protocols used in
this thesis are given.
3.1 Materials
Cell culture and chondrocyte isolation solutions: Phosphate buffer saline (PBS),
fetal bovine serum (FBS), cell culture media (DMEM-Glutamax), antibiotic-antimycotic
(Anti-Anti) and trypsin/EDTA were from Invitrogen AG, Basel, Switzerland. ITS+ Premix,
non-tissue culture treated 24-well plates and 40 μm cell strainers were from Becton
Dickson AG Allschwill, Switzerland. Formaldehyde, L-proline, dexamethasone, tris(2-
carboxyethyl)phosphine hydrochloride solution (TCEP), acetic acid, Spectra-Por Float-
A-Lyzer G2 black 3.5-5 kDa, hexane, cycloheximide, sodium dodecyl chloride (SDS),
sodium chloride (NaCl), pronase E from Streptomyces Griseus, bovine serum albumin
(BSA), cycloheximide, collagenase type II from Clostridium Histolyticu,
ethylenediaminetetraacetic acid (EDTA), sodium phosphate, 1,9-dimethyl-methylene
blue (DMMB), sodium formate, formic acid, ethanol, Papain from papaya latex, L-
cysteine and chondroitin 4-sulfate sodium salt from bovine trachea were purchased
from Sigma Aldrich Chemie GmbH, Buchs, Switzerland. L-Ascorbic acid phosphate
magnesium salt was obtained from Wako (IG instrumenten-Gesellschaft AG, Zurich).
Human fibroblast growth factor-2 (FGF-2) and human transforming growth factor beta
3 (TGF-β3) were from Peprotech, Rocky Hill, NJ, USA.

Materials and methods
20
Materials used in preparation of extracellular matrix multilayers: Bovine type
1 collagen (Col1) was from Becton Dickson AG Allschwill, Switzerland. Chondroitin 4-
sulfate from bovine trachea (CS) was from Fluka, Switzerland. Heparin sodium salt
(HN), hydrochloric acid (HCl), 4-(2-hydroxyethyl)piperazine-1-ethanesulfonic acid
(HEPES), N-(3-dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride (EDC) and N-
hydroxysulfosuccinimide sodium salt (NHS) were purchased from Sigma Aldrich
Chemie GmbH, Buchs, Switzerland. 150mM NaCl solutions used for preparation of
polyelectrolytes had a pH of ~6 and 150mM with 1mM HCl had a pH of ~3. All
polyelectrolyte and rinsing solutions were prepared with deionized ultra-pure water
(DIW) and filtered with a 0.2 µm sterile filter prior to use.
Materials used in preparation of hydrogels: Alginate Pronova UP LVG (low
viscosity (20-200 mPas) sodium alginate in which at a minimum 60% of the monomer
units are α-L-guluronate) was from Novamatrix, Norway. DOWEX ion exchanger,
tetrabutyl ammonium bromide, (D-(+)-glucono-δ-lactone), SO3/pyridine and anhydrous
DMF were purchased from Sigma Aldrich Chemie GmbH, Buchs, Switzerland.
Atelocollagen bovine type 1 collagen (Koken) was from Holzel Diagnostika, Koln,
Germany. CaCO3 particles of 5 µm size were obtained from PlasmaChem GmbH, Berlin.
MMP-degradable and non-degradable PEG powder without RGD-peptides (Reference
1004 and 1010) and disc casters were obtained from QGel, Lausanne, Switzerland.
Antibodies, peptides and staining materials: Immunoglobin G (IgG),
chondroitinase ABC (C2905), Triton X100, phalloidin TRITC labeled mixed isomers
(phalloidin-rhodamine) and Immunoglobin G (IgG), Fluoroshield™ with DAPI were
purchased from Sigma Aldrich Chemie GmbH, Buchs, Switzerland. Alpha 1, alpha 3 and
alpha5 integrin antibodies were obtained from Chemicon International. Type II
collagen antibody II-II6B3, type I collagen antibody M38, proteoglycan, hyaluronic acid
binding region (12/21/1-C-6) and AIIB2 beta1 integrin antibody were from
Developmental Studies Hybridoma Bank, University of Iowa. Monoclonal antibody
against native bovine lubricin (3A4) was from MD Bioproducts, Zurich, Switzerland.
Alexa 488 goat anti-mouse conjugated IgG IgM (H+L) and 4´,6-diamidino-2-

Chapter 3
21
phenylindole dilactate (DAPI) were from Invitrogen AG, Basel, Switzerland. The RGD
peptide with QGel linker was obtained from QGel, Lausanne, Switzerland. The GFOGER
peptide with the sequence GPC(GPP)5-GFOGER-(GPP)5GPC-NH2 was obtained from
professor Richard Farndale (University of Cambridge, Cambridge, UK) and preparation
of the peptide for hydrogel incorporation was followed according to an established
protocol [98]. 2mg GFOGER peptide was reduced in 10 mM acetic acid 2 mM TCEP
then heated for 2 min to 70 °C and allowed to form triple helices at 4 °C for 24 hours.
The triple helical trimer (11.1 KDa) was dialyzed against 10 mM acetic acid with a 3.5-5
KDa cutoff dialysis column to remove unfolded peptides and the TCEP.
3.2 Instruments
Quartz crystal microbalance with dissipation (QCM-D) and crystal
preparation: Assessment of the build-up of layer-by-layer films was followed using
QCM-D (Q-Sense E4, Gothenburg, Sweden). QCM crystals coated with thin PDMS
layers were cleaned for 30 minutes in 2% SDS then rinsed with DIW. The coated
crystals were then plasma treated in air for 2 minutes, mounted in the QCM-D flow cell
and 1ml of 150mM NaCl was injected. After the signal was stable adsorption of the
various layers and observation of the build-up was followed. Overtones 1, 3, 5 and 7
were monitored and the 3rd overtone was used for the assessment of films’ build-up.
QCM gold coated crystals were coated with a thin layer of PDMS using a method
previously described by Voros et al. [121]. Briefly, base silicone oil and crosslinker were
well mixed at a ratio of 10:3 (w/w) and diluted 1:100 (w/w) in hexane. The solution of
base silicon oil and crosslinker in hexane was spin coated on the gold coated QCM
crystals at 2000 rpm for 40 seconds. The crystals were then cured overnight at 70 °C to
allow polymerization of PDMS and to evaporate the hexane. Measurements of the
resonant frequency of the QCM crystals were taken before and after preparing the
thin PDMS layers and were used to calculate the thickness of the PDMS coating. The
PDMS film thickness calculated using the measured resonant frequency shift was 39.2

Materials and methods
22
±2.1 nm. The presence of PDMS coating on the QCM crystals and the films on the
PDMS were confirmed by contact angle measurements.
Atomic force microscopy (AFM): Atomic force microscopy (AFM) images were
taken using a JPK AFM instrument with CSC38 cantilevers (MikroMash).
Measurements were performed in contact mode in liquid at set points ranging from
0.1 V to 0.5 V for areas of 10 x 10 µm at a frequency fX range of 0.1 Hz to 1 Hz.
Quantitative real time polymerase chain reaction (qRT-PCR): Genes were
designed using Primer Select software and gene expression of relevant cartilage
markers was analysed using a StepOnePlus qRT-PCR machine (Applied Biosystems).
Homogenization of hydrogels was performed using a rotor stator homogenizer (TH 220
tissue homogenizer, Omni International, LabForce, Nunningen, Switzerland). Total
RNA was quantified using a spectrophotometer (Nanodrop ND-1000) and the
measured 260/280 ratio was consistently 2.0 ± 0.1 in all samples. Ribosomal protein
L13 (RPL13) was used as a housekeeping gene as it has been recently demonstrated to
be more stable than GAPDH [122]. The Livak method was used for analysis of qRT-PCR
data [123]. Investigated primers are listed in Table 3.1.
Microscopy: Images of the cells’ cytoskeletal organization and
immunohistological staining were taken with a confocal microscope (Carl Zeiss AG/LSM
510, equipped with a 40x 0.6 NA objective). Average cell area and image processing
were carried out using ImageJ 1.43 image processing program. For the GFOGER
experiments (Chapter 5) a Leica SP8 two photon microscope (Leica Microsystems,
Bensheim, Germany) was used to enable imaging deep within the opaque samples.
Macroscopic images of whole hydrogels were taken using a stereomicroscope (Leica

Chapter 3
23
WILD M650, equipped with a 6x objective). Other transmission images were acquired
using a Leica DFC420 C digital microscope camera.
Table 3. 1. Genes used in qRT-PCR analysis
Mechanical properties of the hydrogels: Compressive moduli of hydrogels
were measured using a texture analyser (TA.XTPlus, Stable Microsystems, UK). Samples
were compressed without preload at a speed of 0.01 mm/s and the force was

Materials and methods
24
measured at 0.1 mm compression, where the gels showed linear behavior. Young’s
moduli 𝐸 were calculated according to the formula:
𝜎 = 𝐸 ∙ 𝜀 (Equation 1)
where σ is the measured force per hydrogel area and ε is the strain.
Plate reader: The Synergy H1 Hybrid Reader (Biotek) was used to quantify
proliferation, DNA, RhoA activity and GAG. The plate reader allows top and bottom
UV-visible absorbance, fluorescence intensity, and luminescence detection in
microwell plates.
Mechanical strain machine and strain chambers: The STREX device (ST-140-10,
B-Bridge International) was used to apply mechanical strain to cells seeded in 2D and
3D. Silicon oil was well mixed 1:10 (w/w) with its crosslinker, then degassed PDMS was
poured in chamber molds (32 x 32 mm) and allowed to cure at 80 °C for 6 hours. For
the application of mechanical strain in 3D, STREX PDMS chambers were modified to
contain a series of support ridges to hold the gel as it is stretched. The homogeneity of
the applied strain was assessed by measuring the displacement of 10-30 μm glass
beads embedded in an alginate gel before and after strain. Assessment of
homogeneity was performed with 10% strain on 18 different areas within the strained
gel with 4 images taken in each area (n=3). Measurements of the strain values were
also determined for the other magnitudes applicable by the device.
3.3 Protocols
Chondrocyte isolation: Chondrocytes were harvested from the knees of 6
month old calves obtained from the local slaughterhouse as previously described
[124]. Cartilage shavings were minced using a sterile blade and treated with 0.2%
pronase in DMEM containing 1% antibiotic-antimycotic for 2 hours at 37°C, 7% CO2
with gentle stirring. Following the pronase digestion, the tissue was washed 3 x 2 min
with DMEM containing 1% antibiotic antimycotic then incubated 6 hours in 0.03%

Chapter 3
25
collagenase in DMEM supplemented with 1% antibiotic antimycotic at 37°C, 7% CO2
with gentle stirring. Cells were separated from the digested matrix by filtering through
a 100 μm cell strainer followed by a 40 μm cell strainer. Cells were counted and
viability was determined using an automated cell counter (Countess™ Automated Cell
Counter, Invitrogen AG, Basel, Switzerland). Cell viability in all isolations was above
90%. The isolated cells were seeded at 10,000/cm2 in DMEM supplemented with 1%
antibiotic antimycotic, 10% FBS and 50 µg/mL L-ascorbic acid. At 80-90% confluency
cells were released using trypsin/EDTA and seeded at 5,000/cm2 for the later passages.
Cell culture (Chapter 4): Passage 2 bovine chondrocytes were suspended in
DMEM supplemented with 1% Anti-Anti and 50 µM cycloheximide to prevent de novo
protein synthesis. The cells were seeded on different films of Col1/CS and Col1/HN
prepared on PDMS pieces for 4h at 37°C, 7% CO2. The films were blocked with 1% BSA
in PBS for 30 minutes prior to cell seeding. In order to determine if the cell spreading
was integrin mediated, cells were incubated with antibodies for beta1, alpha1, alpha3
and alpha5 integrins for 15 min prior to seeding on 2 bilayers of Col1/CS denoted as
(Col1/CS)2. IgG was used as control.
Cell culture (Chapter 5): Passage 3 or 4 bovine chondrocytes were mixed with
solutions of alginate, alginate sulfate or a mixture of alginate and alginate sulfate
sterile filtered through a 0.2 µm sterile Millipore filter. The final polymer concentration
was always kept at 2% (w/v) in 150 mM NaCl. Gels were cast in 30 μL volume disks
using the disc caster (Qgel, AG) and polymerized by placing the caster in a 50 mL falcon
tube containing 102 mM CaCl2 solution for 8 min. Disks were removed from the caster
with a spatula and incubated for 8 min free floating in a 60 mm petri dish containing 51
mM CaCl2 to allow equilibration of CaCl2 throughout the gel. Disks were then washed 3
x 2 min each in PBS containing 3 mM CaCl2 and 1 x 2 min in cell culture media
containing 3 mM CaCl2. Each disk was incubated in one well of non-treated 24-well
plates containing 1 mL cell culture media to which an additional 3 mM CaCl2 was
added. Medium was changed every 3 days. Cultures were carried out for 7, 21 or 35
days.

Materials and methods
26
Cell culture of human mesenchymal stem cells (hMSCs, chapter 6): Passage 2
human mesenchymal stem cells (hMSC, Lonza, Switzerland) were seeded and
expanded until passage 6 in DMEM supplemented with 10% FBS, 1% antibiotic-
antimycotic and 1 ng/mL FGF-2 which has been shown to maintain the multilineage
and chondrogenic potential of the stem [125]. Cells were released with trypsin/EDTA
and encapsulated in degradable or non-degradable PEG gels modified with 100 µM
RGD or 100 µM GFOGER peptide or non-modified according to the manufacturer’s
protocol (QGel, Lausanne, Switzerland). The cell/PEG solution was cast in discs of 30 µL
volume each. After 45 min incubation at 37 °C, discs were removed from the caster
and cultured in a non-tissue culture 24 well-plate pre-filled with 1 mL chondrogenic
media/well (DMEM 31966, 1% antibiotic antimycotic, 50 µg/mL L-ascorbic acid, 1%
ITS+ premix, 40 µg/mL L-proline, 100 ng/mL dexamethasone, 10 ng/mL TGF-β3). Non-
modified non-degradable gels cultured in medium without dexamethasone and TGF-β3
were designated as controls. Pellets were also prepared by centrifuging a 250,000 cell
suspension for 5 min at 300 g in 15 mL falcon tubes then culturing the pellets in 500 µL
chondrogenic or control media as differentiation controls. The media was changed
every 3 days.
Chondrocyte dedifferentiation assessment (Chapter 7): Cartilage tissue was
minced then placed in 350 μL RLT Plus Buffer (Qiagen AG, Zurich, Switzerland) and
homogenised using a rotor stator homogenizer (TH 220 tissue homogenizer, Omni
International, LabForce, Nunningen, Switzerland). Chondrocytes isolated from at least
3 different calves were seeded at 10,000 cells/cm2, these cells were designated as
passage 0 (P0). After 4 days of culture, part of the P0 cells seeded in a 25 cm2 tissue
culture polystyrene (TCP) flask were washed in PBS and lysed for mRNA analysis. At
80% confluence after 7±1 days, cells were trypsinized with 0.25% Tripsin/EDTA for 5
min, counted and seeded at 5,000 cells/cm2. Cells were passaged every 4±1 days or
lysed by adding 350 µL RLT plus buffer after washing once with PBS for mRNA
quantification. This was repeated till passage 4 (P4). The 260/280 ratio measured by
the spectrophotometer in all samples was consistently 2.0 ± 0.1. Total RNA was

Chapter 3
27
reverse transcribed starting with 500 ng RNA and gene expression of SZP and Col2 was
determined using qRT-PCR (StepOnePlus, Applied Biosystems). Ribosomal protein L13
(RPL13) was used as a housekeeping gene. Analysis of qRT-PCR data was performed
following the method of Livak.
Cell culture (Chapter 7): Chondrocytes at passage 3 or 4 were seeded on TCP at
5,000 cells/cm2 or embedded in alginate beads at 6x10⁶ cells/mL. To prepare alginate
beads, a cell pellet was suspended in 1.2% (w/v) alginate (Pronova UPLVG, Novamatrix)
and collected in a syringe connected to a 21-gauge needle. All alginate solutions were
sterilized by filtering through a 0.2 µm sterile Millipore filter. The alginate/cell mixture
was dispensed in a 102 mM CaCl2 mixture under continuous gentle stirring for 10min.
Beads were washed 3 x 2 min each with PBS and cultured in a 6-well plate containing
10-11 beads/well, 3 mL media/well. Cells in beads or TCP were incubated for 3-4 days
in a humidified incubator at 37°C under normoxic conditions (21% pO2) or hypoxic
conditions (1% pO2).
Application of mechanical strain (Chapter 7): For the application of strain in
2D, PDMS chambers were plasma treated for 2 min in air. The chambers were then
incubated with 0.1 mg/mL Col1 (Koken CosmoBio) in 150 mM NaCl, 1 mM HCl for 30
min to coat the chambers with a thin Col1 layer. Chambers were washed once with
150 mM NaCl and once with cell culture media followed by seeding passage 3
chondrocytes at 10,000 cells/cm2. 5% tensile strain at 1 Hz was applied for 2 hours, 24
hours post seeding for 4 days or once after 4 days of culture. Chondrocytes were
seeded on non-strained chambers and in 25 cm2 TCP flasks as controls. For application
of 3D strain, 0.75 mL of 2% alginate (w/v) + chondrocytes was injected in modified
PDMS chambers. Alginate was prepared using CaCO3 as a source for calcium ions
which results in uniform, slow-gelation of the material which can be injected [126,
127]. The gel was prepared by mixing 886 μL 2.25% alginate in 150 mM NaCl, 41 μL of
100 mg/mL CaCO3 in DIW, 73 μL of 200 mg/mL freshly prepared D-(+)-glucono-δ-
lactone (GDL) in DIW and 6 million cells. The solution was allowed to gel for 15-30 min
at 37°C. The hydrogels were covered with 5 mL of cell culture media after gelation and

Materials and methods
28
cultured for 24 hours. Mechanical strain was applied at 5.8% magnitude and 1 Hz for 2
hours, 24 hours post seeding for 4 days or once after 4 days of culture. Samples were
lysed 2 hours after the final strain was applied.
Layer-by-layer build-up assessment using QCM-D: Assessment of the build-up
of the films of Col1/CS and Col1/HN was followed using QCM-D (Q-Sense E4,
Gothenburg, Sweden). QCM crystals coated with thin PDMS layers were cleaned for 30
minutes in 2% SDS then rinsed with DIW. The coated crystals were then plasma treated
in air for 2 minutes, mounted in the QCM-D flow cell and 1ml of 150 mM NaCl was
injected. After the signal was stable, 0.5ml of Col1 dissolved at 0.1 mg/ml in 150 mM
NaCl, 1 mM HCl (pH ~3) was injected. After 30 min, crystals were rinsed with 150 mM
NaCl, 1 mM HCl, followed by injection of 1 mg/ml of CS or HN dissolved in 150 mM
NaCl (pH ~6). After 15 min incubation, a rinsing step with 150 mM NaCl was
performed. The steps were repeated for the desired number of layers. To test the
stability of the films at physiologic pH and in cell culture media, 1 ml of 10 mM HEPES,
pH 7.4, 150 mM sodium chloride (HEPES-2 buffer) was injected for 1 hour. Afterwards,
serum free media containing 1% antibiotic antimycotic was injected and the signal was
observed for 24 hours. Overtones 1, 3, 5 and 7 were monitored and the 3rd overtone
was used for the assessment of films’ build-up.
Chemical cross-linking of layer by layer films: Cross-linking was performed on
films of (Col1/CS)10 and (Col1/HN)10 prepared on PDMS pieces. 400 mM EDC and 100
mM NHS were freshly prepared in 150 mM NaCl. The substrates with the films were
crosslinked in the EDC/NHS solution for 12 h at 4 °C. [84] Films were then rinsed with
150 mM NaCl followed by deionized water and dried with a low stream of N2. Contact
angle measurements and cell adhesion assessment were then performed on the
prepared films.
Assessment of films’ topography and thickness: Silicon oil was well mixed 1:10
(w/w) with its cross linker. 3 mL of the degassed PDMS were poured in 35 mm cell
culture Petri dishes and allowed to cure at 80 °C for 6 hours. The Petri dishes were
then plasma treated in air for 2 min. The PDMS was rinsed with 150 mM NaCl and

Chapter 3
29
build-up of the films of Col1/CS or Col1/HN was initiated. Films were prepared using
the same protocol described in the QCM-D section with different number of layers and
different end layers. AFM images were taken using a JPK AFM instrument with CSC38
cantilevers (MikroMash). Fibril thickness was determined by measuring the height of
10 individual fibrils in a given image using the JPK image processing program and
calculating the average thickness and standard deviation. The thickness of the films
was measured by AFM using a previously-described method [128]. Briefly, a 2x2 µm
area of the sample was scanned several times at a high set point of approximately 5 V
and high frequency (10-20 Hz) to remove the film. A 10x10 µm area of the sample
including the scratched area was then scanned and the film thickness was determined
by measuring the average difference between 30 randomly chosen points of the
unscratched area and the scratched area using the JPK image processing program.
Cell viability: Cell viability was assessed using the Live/Dead® viability assay
from life technologies. Calcein AM is a green fluorophore that can only be activated
upon esterase cleavage by viable cells, while ethidium homodimer (EthD-1) is a red dye
that can only penetrate the membrane of a dead cell where it binds to DNA in the
nucleus. Cells in 2D or 3D were washed three times in warm PBS and subsequently
incubated in a solution containing 2 µM calcein AM and 4 µM EthD-1 for 30 minutes at
37 °C. Samples were then washed three times in warm PBS and then imaged directly
with a Zeiss LSM 510 Confocal Microscope (Carl Zeiss AG, Oberkochen, Germany) at a
minimum of three separate imaging planes per gel.
Cell morphology: Cell nuclei and the f-actin were visualized using double
labelling by DAPI and rhodamine-labelled phalloidin respectively. Cells were fixed with
5% formalin for 30 minutes at 4 °C, washed 3x with PBS, permeabilized with 0.4%
Triton X-100 for 20 minutes, and washed again. F-actin fibers were stained for 1 h
total with 0.13 µg/mL phalloidin. After 30 minutes, 2 µg/mL of DAPI was added at 1:1
(v/v) to the phalloidin solution for the remaining 30 minutes. All staining was done at
4 °C and in complete darkness. After staining, samples were washed in PBS and z-
Stacks were acquired using a two photon microscope and (SP8 Leica Microsystems,

Materials and methods
30
Bensheim, Germany). Samples grown in monolayer cultures or in translucent hydrogels
were imaged using a confocal microscope (Carl Zeiss AG/LSM 510, equipped with a 40x
0.6 NA objective).
Proliferation within hydrogels: Proliferation was assessed using a BrdU assay
Calbiochem® BrdU Cell Proliferation Assay (Calbiochem, Millipore, Switzerland). The
assay was performed according to the manufacturer’s recommendations with slight
modifications. Briefly, the gels were placed in media containing 1:200 BrdU label in a
96-well plate for 24 hours, post which the medium was carefully replaced with 270 μL
of dissolving buffer (0.055 M sodium citrate, in 0.03 M EDTA, 0.15 M NaCl, pH 6.8) and
the plate was rocked for 20 min at 1000 rpm at room temperature. For PEG gels the
dissolving solution used was trypsin. A volume of 30 µL was transferred to a new 96-
well plate and cells were centrifuged for 10 min at 1000 rpm, dried with an N2 stream
and fixed using the fixative solution supplied in the kit. Staining was then carried out
by an Anti-BrdU antibody followed by HRP conjugation and substrate addition
according to the manufacturer’s protocol. Absorbance was measured at dual
wavelengths (450-540) and the difference was blanked with the signal obtained from
cell free gels using a plate reader (Synergy H1 Hybrid Reader, Biotek).
DNA quantification in gels: Gels were collected at day 7 and day 21 for
assessment of DNA content and dissolved in 100 µL of papain lysis buffer (10 mM
EDTA, 100 mM sodium phosphate, 10 mM L-cysteine, 125 µg/mL Papain type III at pH
6.3) overnight at 60 ˚C with shaking at 1000 rpm. DNA quantification was carried out
with Quant-iTTM PicoGreen dsDNA Kit (Invitrogen). All samples were diluted 1:10 in TE
buffer supplied by the kit. DNA standard was diluted in TE Buffer to 2 μg/mL, 200
ng/mL, 20 ng/mL, 2 ng/mL and 0.2 ng/mL concentrations. Lysis buffer was diluted 1:10
and Picogreen reagent was diluted 1:200 in TE buffer. A volume of 100 µL of each
sample and DNA standard was transferred to a 96-well plate in triplicates and an equal
volume of PicoGreen solution was added in each well. Measurement was performed
using an excitation wavelength of 480 nm and emission wavelength of 520 nm with a
plate reader.

Chapter 3
31
1,9-Dimethylmethylene blue (DMMB) assay: DMMB dye was prepared
according to Estes et al. [129]. Briefly, 21 g of DMMB was dissolved in 5 mL ethanol
with 2 g sodium formate and mixed into 800 mL distilled water. The pH was adjusted
to 3.0 with formic acid followed by bringing up the solution to 1 L. Gels were collected
at day 7 and day 21 for assessment of GAG content and dissolved in 100 µL of papain
lysis buffer (10 mM EDTA, 100 mM sodium phosphate, 10 mM L-cysteine, 125 µg/mL
Papain type III at pH 6.3) overnight at 60 ˚C with shaking at 1000 rpm. Chondroitin 4-
sulfate (Sigma) was diluted in papain buffer dilutions from 0 to 35 µg/mL were used to
create the standard curve. A volume of 40 µL of each sample or standard was pipetted
into a 96-well plate. A volume of 125 µL of the DMMB dye was added to each well and
optical density was measured at 595 nm with a plate reader. All GAG quantification
was normalized to DNA content for each gel.
Gene expression: Samples in 2D were washed once in PBS followed by addition
of 350 μL of RLT plus buffer immediately to chambers or TCP. Alginate and alginate
sulfate samples were washed once with PBS and dissolved completely in 1 mL sodium
citrate dissolving buffer. Samples were centrifuged at 10,000 rpm for 2 min, re-
suspended in PBS and centrifuged at 14,500 rpm for 2 more minutes. The pellet was
then lysed in 350 μL of RLT Plus Buffer. Expression of superficial zone protein was
determined by qRT-PCR using RPL13 as a housekeeping gene. In alginate sulfate
samples, 0.5% (w/v) BSA was added to the qRT-PCR mixture to prevent interference of
the sulfate groups with the amplification reaction [130]. PEG gels and pellets were
homogenised using a rotor stator homogenizer (TH 220 tissue homogenizer, Omni
International, LabForce, Nunningen, Switzerland). A 540 μL of RNA free water and 10
µL Protease K solution (Qiagen) were added to each sample. Samples were incubated
for 10 min at 55 °C with continuous shaking at 1000 rpm. RNA was isolated using the
RNeasy Mini Kit (Qiagen) following the fibrous tissue isolation protocol given by the
manufacturer. Total RNA was quantified using a spectrophotometer (Nanodrop ND-
1000) and the measured 260/280 ratio was consistently 2.0 ± 0.1 in all samples. Total
RNA was reverse transcribed and gene expression of type II collagen (Col2), type I
collagen (Col1) and aggrecan (Agg) were determined using quantitative real time PCR

Materials and methods
32
(StepOnePlus, Applied Biosystems). Ribosomal protein L13 (RPL13) was used as a
housekeeping gene and the Livak method was used for analysis of qRT-PCR data [123].
Immunostaining: Samples were washed once with PBS, fixed with 4%
paraformaldehyde, 0.2% Triton-X100 for 30 min then washed once with PBS. Alginate
samples were stored in PBS containing 10 mM CaCl2 to prevent gel dissolution.
Samples were embedded in Optimum Cutting Temperature (O.C.T) compound and
frozen on a dry ice block for 5 min and 6 μm-thick slices were cut using a microtome
(CryoStar NX70, ThermoScientific). The slices were fixed in ethanol then washed with
10 mM CaCl2 in PBS. Antigen retrieval was performed using pronase at 1 mg/mL for 15
min at 37 °C for Col2 and Col1 and using chondroitinase at 0.02 U/mL for 40 min at
37 °C for proteoglycans. Samples were then blocked with 5% BSA in PBS for 1 h at
room temperature. Samples were incubated with primary antibodies specific against
Col2 (II-II6B3, Developmental Studies Hybridoma Bank), Col1 (M38, Developmental
Studies Hybridoma Bank) and proteoglycan, hyaluronic acid binding region (12/21/1-C-
6, Developmental Studies Hybridoma Bank) at 1:10 dilutions in 1% BSA in PBS
overnight at 4 °C. Samples were washed 3x in PBS and incubated with 1:400 Alexa-488
goat anti-mouse secondary antibody for 1 h at room temperature. Controls were
prepared following the same procedure but omitting the primary antibody. Samples
were imaged using a confocal microscope (Carl Zeiss AG/LSM 510, equipped with a 40x
0.6 NA objective).
Alcian blue staining: Frozen sections of the hydrogels were defrosted at room
temperature, washed in PBS for 5 minutes and fixed with 4% formaldehyde in PBS for
7 minutes. Samples were washed 3x 5 minutes each in PBS, hydrated with DIW and
then stained in alcian blue solution (pH 2.5) for 20 minutes. Slides were washed in
running tap water until unspecific staining was washed away, samples were rinsed in
DIW and dehydrated using ascending alcohol solutions (95% to absolute ethanol) for 3
minutes each. Sections were cleaned in xylene, air-dried and mounted with resinous
mounting medium (Mounting Medium ref. 4112, Richard-Allan Scientific). A cover slip

Chapter 3
33
was added on the top of the section and sealed with nail polish. Images were acquired
using a Leica DFC420 C digital microscope camera equipped with a 40x objective.
Statistical analysis: All quantitative data were obtained from at least 3
independent donors and expressed as the mean ± standard error. Statistical evaluation
was carried out by analysis of variance (ANOVA) and post-hoc Tukey’s tests where P
values of less than 0.05 were considered significant. Statistical analysis was performed
using OriginPro version 8.1.

Materials and methods
34

Chapter 4
35
4 Layer-by-Layer Films Made from
Extracellular Matrix Macromolecules on
Silicone Substrates
The work in this chapter has been published in R.F. Mhanna et al., Biomacromolecules 2011;
12: 609-616
In the introduction we identified the current problems with monolayer cultures
mainly represented by chondrocyte dedifferentiation. We hypothesized that the use of
molecules from ECM origin might be a tool to improve the chondrogenic performance
of serially passaged chondrocytes. Furthermore, such molecules can also be used to
improve biocompatibility of implanted prosthetic devices. In this chapter we
demonstrate the build-up of Col1/CS and Col1/HN films prepared using the layer-by
layer technique.
4.1 Current monolayer culturing techniques
The layer-by-layer (LbL) technique has been widely used to produce nanofilms
for biomedical applications [79, 80]. The LbL approach allows the build-up of thin films
simply by the alternating deposition of polyanions and polycations [81]. A wide variety
of polymeric compositions both synthetic [131] and natural [82, 88, 132] have been
studied since the first introduction of the technique by Decher and coworkers [81]. LbL
films have been used to improve cytocompatibility of poly ethylene terephthalate and
poly(l-lactic acid) [133, 134]. Films can be tuned to favour cell adhesion [83, 85] or
hinder cell adhesion through the addition of antifouling end layers such as
poly(ethylene glycol) [121, 135].

Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
36
Despite their success in cell cultures, the use of LbL engineered multilayers in
biomedical applications raised the concern of cytotoxicity of these films. Recent
studies show that polymers used to prepare films such as the traditional
polystyrenesulfonate/polyallylamine hydrochloride (PSS/PAH) multilayers can be
cytotoxic if released to the culture media [87]. Thus having systems of natural
polymers such as ECM macromolecules or other naturally occurring proteins is of high
importance. Some ECM molecules such as chondroitin sulfate (CS) and heparin (HN)
contain sulfate groups that render these polymers negatively charged at physiological
pH [91]. These can then be used in combination with type I collagen (Col1) [90] which
is protonated at a pH below its isoelectric point (5.5) [136] to form natural thin films,
for a variety of applications. CS is the most abundant GAG in plasma [137] and has
been shown to increase proteoglycan synthesis and to have roles in
mechanotransduction of articular cartilage [37, 38]. CS is available as a food
supplement and as a disease protective drug for arthritis patients [27, 39]. On the
other hand HN is a well-known anticoagulant and has been used in various biomedical
applications [91]. All these polymers have been used for generation of scaffolds for
tissue engineering especially Col1 which has been well tested in-vivo for the delivery of
chondrocytes in autologous chondrocyte implantation [29, 138-140].
Polydimethylsiloxane (PDMS) is a common elastic biomaterial used in a wide
range of biomedical applications including microfluidic devices [141], breast implants
[142], contact lenses [143], cardiac pacemakers, glaucoma drainage devices [144] and
the study of cells under static and cyclic strain [108, 145]. PDMS is naturally
hydrophobic and cell repellent. One of the most common methods used to modify
PDMS is plasma treatment which creates free hydroxyl groups on its surface rendering
PDMS hydrophilic [146]. This process is reversible as PDMS can recover hydrophobicity
within days or even hours at normal temperature and in dry environment due to the
migration of non-crosslinked monomers to the surface. Hydrophobicity recovery can
be delayed by storing PDMS in water and low temperature [147, 148]. A more efficient

Chapter 4
37
and more stable modification of the PDMS surface is by adsorption of hydrophilic
polymers on its surface [144].
In this study we assessed the build-up, stability, topography and thickness of
films prepared from (Col1/CS) and (Col1/HN). Moreover, we assessed wettability and
cell adhesion on these LbL films. The current work provides insight for the use of these
layers for biomedical applications which make use of PDMS substrates.
4.2 Build-up of Col1/CS and Col1/HN films on PDMS
The QCM-D data showed a clear build-up of the Col1/CS and Col1/HN systems
on the PDMS coated crystals (Figures 4.1 and 4.2). After Col1 adsorption the frequency
decreased about 70 Hz for all crystals with a high increase in dissipation indicative of
loose and highly hydrated layers. After the first Col1 layer, adsorption of HN or CS
caused an increase in the frequency of about 50 Hz with a decrease in the dissipation
in both systems. In the following layers, build-up of the Col1/HN system had larger
frequency drop steps than the Col1/CS (Figure 4.3).
In the Col1/CS system the adsorption of CS caused a decrease in the dissipation
and increase in frequency indicating that the film became stiffer as a negative layer
was adsorbed, a phenomenon often observed in the stratification of LbL films (Figure
4.1). The injection of CS solution (pH~6) might also initiate crosslinking of collagen
monomers induced by the local change in the film pH resulting in the formation of
fibrils. The adsorption of Col1 resulted in a decrease in the frequency and increase in
the dissipation regardless of the layer number.
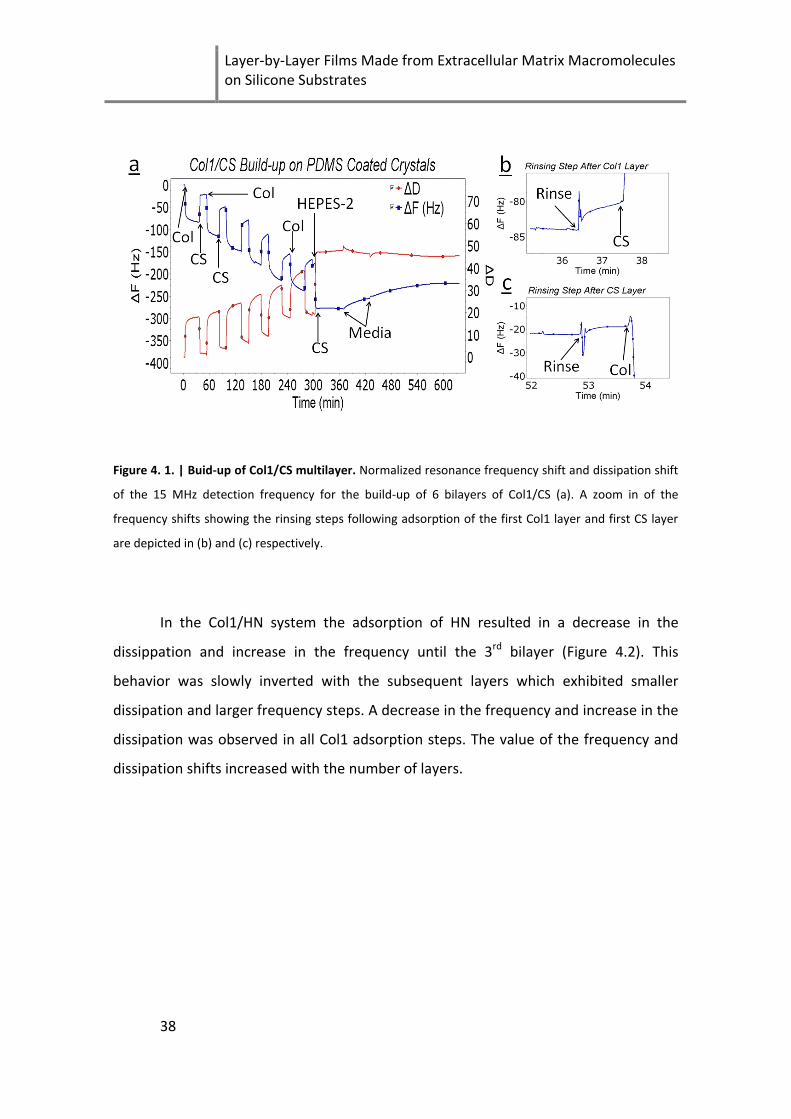
Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
38
Figure 4. 1. | Buid-up of Col1/CS multilayer. Normalized resonance frequency shift and dissipation shift
of the 15 MHz detection frequency for the build-up of 6 bilayers of Col1/CS (a). A zoom in of the
frequency shifts showing the rinsing steps following adsorption of the first Col1 layer and first CS layer
are depicted in (b) and (c) respectively.
In the Col1/HN system the adsorption of HN resulted in a decrease in the
dissippation and increase in the frequency until the 3rd bilayer (Figure 4.2). This
behavior was slowly inverted with the subsequent layers which exhibited smaller
dissipation and larger frequency steps. A decrease in the frequency and increase in the
dissipation was observed in all Col1 adsorption steps. The value of the frequency and
dissipation shifts increased with the number of layers.

Chapter 4
39
Figure 4. 2. | Buid-up of Col1/HN multilayer. Normalized resonance frequency shift and dissipation shift
of the 15 MHz detection frequency for the build-up of 6 bilayers of Col1/HN (a). A zoom in of the
frequency shifts showing the rinsing steps following adsorption of the first Col1 layer and the first HN
layer are depicted in (b) and (c) respectively.
A linear build-up was observed with the Col1/CS films (Figure 4.3), which is in
aggreement with the results reported by Zhang et al. [82] on the build-up of
Col1/hyaluronic acid (HA). On the other hand, the build-up in the Col1/HN system was
exponential, indicating a less compact structure that permits the diffusion of one or
both polymers (Figure 4.3). The exponential growth of LbL films constituting
polysacharides such as chitosan (CH) and HA have been previously observed and well
characterized [149, 150]. Diffusion of CH molecules is believed to drive the exponential
growth of these films. In the CH/HA films, isolated islets are formed during the first few
layers which coalesce to vermiculate structures as the number of deposited layers
increases. This growth behavior differs from the systems presented in this study and
other systems prepared with Col1 as the cationic polymer [82, 88], which exhibit a
fibrillar structure covering the surface from the initial layers.
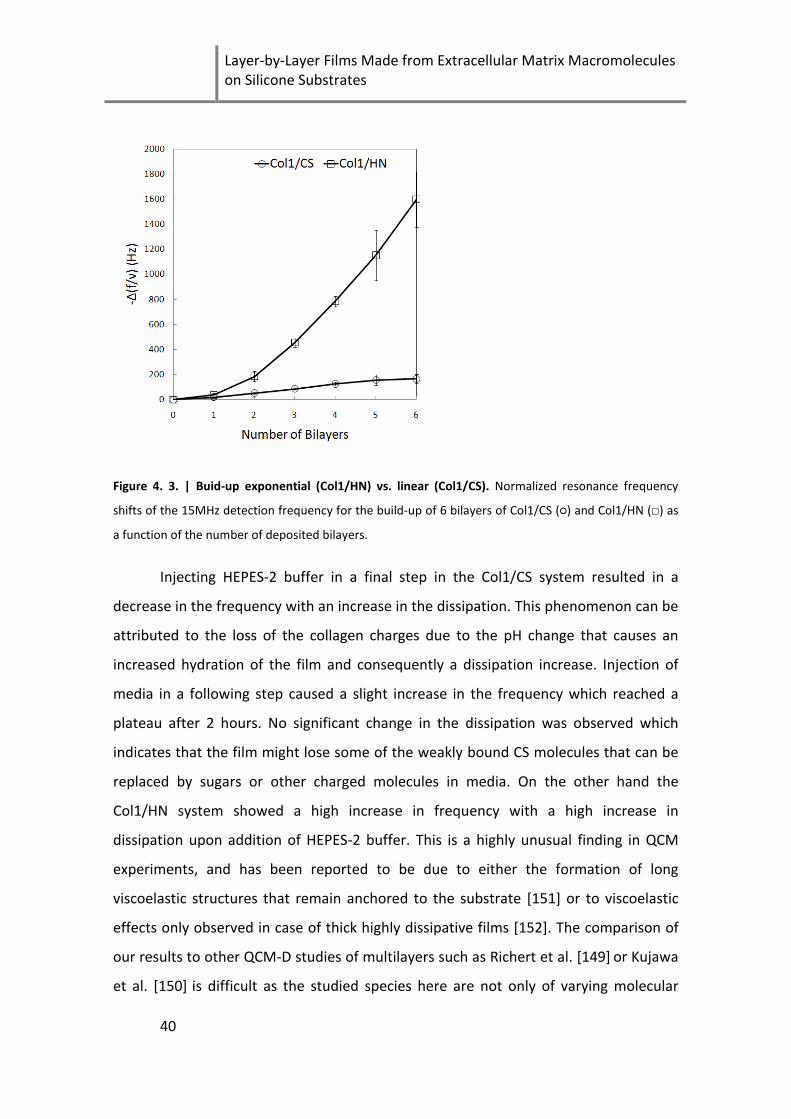
Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
40
Figure 4. 3. | Buid-up exponential (Col1/HN) vs. linear (Col1/CS). Normalized resonance frequency
shifts of the 15MHz detection frequency for the build-up of 6 bilayers of Col1/CS (○) and Col1/HN (□) as
a function of the number of deposited bilayers.
Injecting HEPES-2 buffer in a final step in the Col1/CS system resulted in a
decrease in the frequency with an increase in the dissipation. This phenomenon can be
attributed to the loss of the collagen charges due to the pH change that causes an
increased hydration of the film and consequently a dissipation increase. Injection of
media in a following step caused a slight increase in the frequency which reached a
plateau after 2 hours. No significant change in the dissipation was observed which
indicates that the film might lose some of the weakly bound CS molecules that can be
replaced by sugars or other charged molecules in media. On the other hand the
Col1/HN system showed a high increase in frequency with a high increase in
dissipation upon addition of HEPES-2 buffer. This is a highly unusual finding in QCM
experiments, and has been reported to be due to either the formation of long
viscoelastic structures that remain anchored to the substrate [151] or to viscoelastic
effects only observed in case of thick highly dissipative films [152]. The comparison of
our results to other QCM-D studies of multilayers such as Richert et al. [149] or Kujawa
et al. [150] is difficult as the studied species here are not only of varying molecular

Chapter 4
41
weight but also of different structure and charge density. In addition, in our case the
formation of fibres induced by pH change further complicates the system.
Build-up with Col1 was only possible at pH less than 5. As the isoelectric point
of Col1 is about 5.5 [136], if Col1 is rinsed with a solution of pH higher than 5, most of
the amino charged groups of collagen will be eliminated preventing electrostatic
adsorption of negatively charged CS or HN. At low pH Col1 is highly charged causing
strong repulsion between the Col1 monomers. At higher pH such as physiological pH,
the charges carried by Col1 are fewer which facilitates their self-assembly into larger
fibers [153].
4.3 Effect of substrate on film build-up
The build-up of these ECM-based films was dependent on the underlying
substrate. The initial Col1 adsorption step on the gold substrate was much larger than
on PDMS indicated by the almost 8-fold larger decrease in frequency and increase in
dissipation (Figure 4.4). After the 2nd bilayer, the frequency shifts of collagen become
similar to the shifts observed on PDMS. Furthermore, adsorption of CS on Col1 appears
to cause a further decrease in the frequency and decrease in dissipation. The decrease
in the frequency in this case can be attributed to the large amount of collagen
available for CS binding. It was clear from our experience that results obtained on gold
could not be used to estimate the build-up on PDMS. Therefore it is essential to assess
build-up of films on the actual substrates used in a given application.

Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
42
Figure 4. 4. | Build-up of Col1/CS on gold. Normalized resonance frequency shift and dissipation shift of
the 15 MHz detection frequency for the build-up of 6 bilayers of Col1/CS on gold.
4.4 Film topography
AFM images of Col1/CS and Col1/HN showed a fibrillar structure of the films
(Figure 4.5). The fibrils in the Col1/CS system increased in diameter from 10.1±1.4 nm
for (Col1/CS)2 (Figure 4.5A) up to ~19.2±3.25 nm for (Col1/CS)10 (Figure 4.5D).
Individual fibrils of about 1-2 µm in length were observed after 2 bilayers (Figures 4.5A
and B). The length of the fibrils increased with number of layers deposited and after 10
bilayers fibrils of 4-5 µm in length could be observed (Figure 4.5D). In the Col1/HN
system, films had sparse fibrils of 9.2±1.2 nm diameter for (Col1/HN)2 (Figure 4.5E and
F). A meshwork of fibrils was observed for (Col1/HN)5 and (Col1/HN)10 with no
significant change in fibrils` diameter (Figure 4.5G and H). The addition of a collagen
layer clearly increased the thickness of the fibrils in both systems which can be
observed in (Figure 4.5B and 5F).

Chapter 4
43
Figure 4. 5. | Morphology of Col1/CS and Col1/HN films. 10x10 µm AFM Images for various assemblies
of different layer numbers of Col1/CS and Col1/HN taken at room temperature in liquid (150 mM NaCl,
pH~6) in contact mode. (Col1/CS)2 (A), (Col1/CS)2Col (B), (Col1/CS)5 (C), (Col1/CS)10 (D), (Col1/HN)2 (E),
(Col1/HN)2Col (F), (Col1/HN)5 (G) and (Col1HN)10 (H).

Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
44
4.5 Film thickness and stability
The thickness of the films was measured with AFM by scratching with the tip of
the cantilever (Figure 4.6). This method ensures an accurate measurement of the film
thickness assuring no damage to the underlying PDMS substrate. Difficulties in
obtaining perfect squared scratches were due to the thick fibrils in the Col1/CS system
which would be bundled together when scanned with the cantilever for many times.
The thickness of the (Col1/CS)10 film was 15.6 ± 6.8nm (Figure 4.6A) while (Col1/HN)10
was 26.4 ± 9.4 (Figure 4.6C). In order to confirm the stability results obtained from the
QCM-D curves, scratches on (Col1/CS)10 and (Col1/HN)10 were performed before and
after incubating the films with HEPES-2 buffer and tissue culture media. The thickness
and also the topography of (Col1/CS)10 were stable for 24 h in HEPES-2 buffer and in
tissue culture media (Figure 4.6B) which is in accordance with the QCM-D data. On the
other hand films of Col1/HN maintained their fibrillar topography when moved to
HEPES-2 buffer or DMEM but the thickness of the films was reduced to 8.9 ± 3.5 nm in
DMEM (Figure 4.6D).

Chapter 4
45
Figure 4. 6. | Films’ thickness and stability. AFM image with a scratched region for (Col1/CS)10 in 150
mM NaCl, pH~ 6 as prepared (A), and after incubation in media for 24 h (B), AFM image with a
scratched region for (Col1/HN)10 in 150 mM NaCl, pH~6 as prepared (C) and after incubation in media
for 24 h (D). The scanned area was 10x10 µm and the scratch was 2x2 µm.

Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
46
4.6 Assessment of cell adhesion and integrin mediated spreading
In order to investigate cell adhesion to our synthesized substrates and not
extraneous proteins, cells were incubated with 1) cycloheximide, to avoid influences of
adhesive proteins synthesized by the cells themselves and 2) in serum free media to
prevent influences of serum proteins on cell adhesion (Figure 4.7). Under these
conditions cell spreading was superior on ECM films compared to PDMS and tissue
culture plastic (TCP) (Figure 4.8A), suggesting that normal spreading on TCP is
dependent on proteins synthesized by the cells or serum proteins. The average cell
area on the (Col1/CS)2 film was 2025 ± 1085 μm2 which was significantly greater than
the average cell area on TCP 661 ± 611 μm2 using the Student’s t test (p < 0.05). Cells
under these conditions do not adhere well to PDMS, however, the ECM film modified
PDMS allowed strong cell adhesion, where even application of 10% tensile strain to a
PDMS membrane for 24 h was possible without noticeable cell detachment. No
significant difference in cell adhesion and spreading was found between the different
films except the (Col1/HN)10 (Figure 4.7C) which exhibited a high percentage of round
cells with a significantly (p < 0.05) lower average cell area of 977 ± 781 μm2. Adhesion
to (Col1/HN)10 was possibly reduced due to the high film hydration indicated by the
large increase in dissipation as observed with QCM-D. The stiffness of the substrate
has been shown to play an important role on cell adhesion. In a study by Richert et al.
[84], chondrosarcoma cells adhered and spread only on cross-linked Poly(L-
lysine)/hyaluronan (PLL/HA) films but not on native films of the same composition.
In our work, cell spreading was limited by crosslinking, where the cells had a
more restricted spreading on the substrate, expressed more filopodia and exhibited a
ruffled morphology (Figure 4.7E and F). The uncrosslinked, native films used in this
study were not as hydrated as the non-cell adhesive (PLL/HA) films and allowed better
access to collagen binding sites, whereas crosslinking increased the stiffness of the
substrate and limited the cell's access to collagen binding sites.

Chapter 4
47
In the Col1/CS system, cell morphology was not affected by the number of
layers, a result which can be explained by the similarity in topography of the films
(Figure 4.7B, D and F). The end layer also had no significant effect on cell adhesion.
Since chondrocytes do not possess specific receptors for HN, and their adhesion is
mainly dictated by Col1, this observation indicates that Col1 must be accessible for the
cells even for CS and HN ending films. This contrasts with the study by Zhang et al. [82]
who found chondrosarcoma cultured on Col1 ending films formed pseudopod-like
attachment on the collagen fibrils, but such structures did not form on hyaluronic acid
(HA) ending films. These results suggest that although HA might form a completely
uniform film over the Col1, CS and HN layers are not continuous and allow cells to
access collagen from deeper layers.
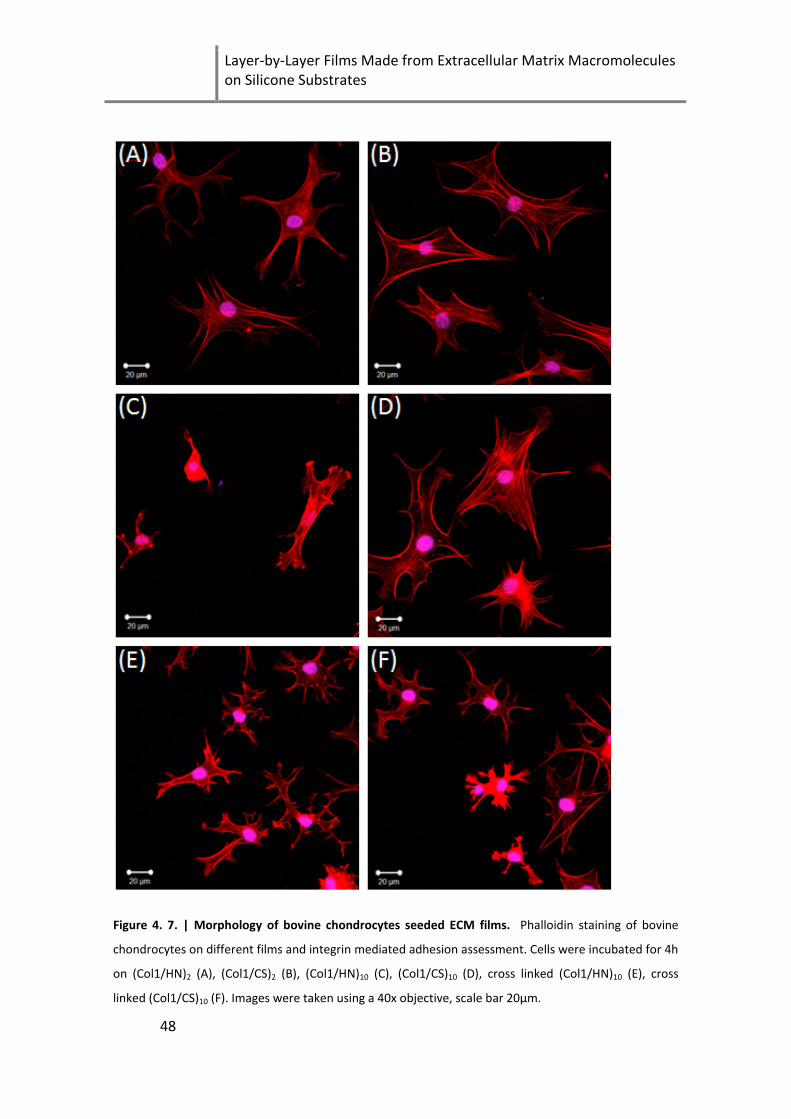
Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
48
Figure 4. 7. | Morphology of bovine chondrocytes seeded ECM films. Phalloidin staining of bovine
chondrocytes on different films and integrin mediated adhesion assessment. Cells were incubated for 4h
on (Col1/HN)2 (A), (Col1/CS)2 (B), (Col1/HN)10 (C), (Col1/CS)10 (D), cross linked (Col1/HN)10 (E), cross
linked (Col1/CS)10 (F). Images were taken using a 40x objective, scale bar 20µm.

Chapter 4
49
In order to prove that cell adhesion was integrin-mediated, cells were
incubated with antibodies to alpha1, alpha3, alpha5, beta1 integrins and IgG as
control and then cultured on films of (Col1/CS)2 (Figure 4.8). Beta1 integrin forms
dimers with several alpha subunits and controls interaction with several ECM
molecules including collagen, fibronectin and vitronectin. Alpha1 binds the monomeric
collagen fibril [154] and laminin, alpha3 binds laminin and thrombospondin, and
alpha5 binds fibronectin and fibrinogen [155]. Cell adhesion and spreading were
hindered only with the beta1 integrin antibodies (Figure 4.8C), while the other
integrins had no effect on adhesion or cell spreading. Less cell adhesion was observed
when beta1 integrin antibodies were used compared to the control and other
antibodies. Furthermore, cell area was significantly lower for cells incubated with
beta1 integrin antibodies (364 ± 142μm2 compared to IgG 1927 ± 733μm2 ) whereas
incubation with other antibodies caused no significant difference in average cell area.

Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
50
Figure 4. 8. | Integrin mediated cell spreading. Phalloidin staining of bovine chondrocytes seeded on
(Col1/CS)2 and incubated for 4h with antibodies to specific integrins. Tissue culture plastic (A), (Col1/CS)2
with IgG antibody (B), (Col1/CS)2 with beta1 integrin antibody (C), (Col1/CS)2 with alpha1 integrin
antibody (D), (Col1/CS)2 with alpha3 integrin antibody (E), (Col1/CS)2 with alpha5 integrin antibody.
Images were taken using a 40x objective, scale bar 20µm.

Chapter 4
51
4.7 Chapter summary
The goal of this study was to coat deformable PDMS substrates with specific
ECM macromolecules, so that cell/ECM interactions could take place under controlled
conditions. The thickness and morphology of films made from Col1/CS on PDMS were
stable in cell culture media. Films made from Col1/HN maintained their topography
but had a loss in the total film thickness. The substrate has a great influence on the
build-up of the films where adsorption of Col1 on gold was 8 times larger than on
PDMS. Cell adhesion was mediated mainly by beta1 integrins and cells had a
constrained spreading and ruffled morphology on crosslinked films compared to non
crosslinked. This study concerns the use of ECM molecules to coat surfaces for
studying specific ECM/cell interactions. This work is of major importance for studies of
mechanical loading of cells on PDMS. Further gene expression studies should help
reveal the role of chondroitin sulfate, heparin and other glycosaminoglycans in
cartilage mechanobiology.

Layer-by-Layer Films Made from Extracellular Matrix Macromolecules on Silicone Substrates
52

Chapter 5
53
5 Chondrocyte Culture in 3D Alginate Sulfate
Hydrogels Promotes Proliferation While
Maintaining Expression of Chondrogenic
Markers
The work in this chapter has been submitted to Biomaterials, Mhanna et al. (2013A).
In the previous chapter, we were able to prepare films made from extracellular
molecules relevant for cartilage engineering. We observed that stable films containing
the chondroprotective chondroitin sulfate molecule could be made at varying
thicknesses. However, cells cultured on these films maintained a spread fibroblast like
phenotype which motivated us to develop a 3D system that may provide similar
biological cues. To achieve this, alginate a commonly used biomaterial known to
maintain the cartilage phenotype was modified with sulfate groups and the material
was investigated for its potential use for cartilage tissue engineering.
5.1 Improving chondrogenic performance of chondrocytes in 3D
may be achieved using biomimetic materials.
Articular cartilage does not regenerate after injury or disease. Developing
strategies to restore the function of the damaged tissue is a major objective for tissue
engineers [12, 156]. Cell based approaches used for treatment of cartilage lesions
include microfracture, osteochondral grafting and autologous chondrocyte
implantation (ACI) [13, 14, 50]. Recently ACI increased in popularity and showed
promising results for treating medium and large chondral defects [49, 157, 158]. ACI

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
54
first described by Brittberg et al. [52] in 1994, is a two-step surgical procedure. In the
first stage a cartilage tissue biopsy harvested from the patient is enzymatically digested
to separate cells from the tissue and the cells are grown in a laboratory to reach
approximately 12 million cells. In the second stage, the cells are injected in the defect
and covered with a periosteal flap or membrane [57]. One problem associated with
use of these cells is the dedifferentiation which occurs during monolayer expansion
[59, 159]. Dedifferentiated chondrocytes express fibroblastic genes such as type I
collagen and produce fibrous tissue rather than hyaline cartilage in the defect site [50,
160]. Additionally, donor site morbidity from harvesting the periosteal flap and the
need for a second surgery are some of ACI’s other limiting factors. Taken together,
these limitations have opened the quest for 3D scaffolds for cartilage tissue
engineering. Three dimensional culture systems such as agarose [60] and alginate [72,
161, 162] restore the function and phenotype of chondrocytes. However, cell
proliferation is limited in alginate and agarose restricting their use in priming cells for
ACI-like procedures.
The cartilage extracellular matrix (ECM) is composed of 10-20% proteoglycans
[50]. The core proteins of proteoglycans are heavily modified by glycosaminoglycans
(GAGs) including chondroitin sulfate, keratan sulfate and dermatan sulfate. These
linear polysaccharides have sulfate and carboxylic groups which contribute to the high
fixed negative charge of the proteoglycans at physiologic pH. The GAG side chains also
give the tissue its compressive and osmotic swelling properties by entrapping water
[13] .
Sulfation of molecules has purported effects on the biological activity of
biopolymers. Chondroitin sulfate has been shown to have therapeutic benefits for hip
and knee osteoarthritis [27, 39]. Heparan sulfate has a high affinity to a plethora of
growth factors crucial for cartilage homeostasis [163, 164]. Chitosan sulfation
enhanced fibroblast adhesion and contraction of a collagen lattice compared to the
unsulfated material [165]. Freeman et al. [166] studied the binding affinity of 10
different heparin-binding factors (including FGF, IGF and VEGF) to alginate sulfate.

Chapter 5
55
They found that except FGF most of the factors bind equally well or better to sulfated
alginate compared to heparin. Reem et al. showed that scaffolds containing a mixture
of alginate and sulfated alginate caused attenuated TGFβ1 release and consequently
improved chondrogenesis of entrapped mesenchymal stem cells compared to scaffolds
lacking alginate sulfate [167]. The scaffolds used in these studies were made by
freezing alginate sulfate into a macroporous scaffold [166]. In this study we report for
the first time a sulfated alginate hydrogel which is crosslinkable with calcium.
We hypothesized that modification of alginate with sulfate groups provides a
more chondrogenic environment compared to pure alginate. To test this hypothesis
alginate was modified with sulfate groups and chondrocytes were encapsulated in
alginate and alginate sulfate at varying ratios. Cell morphology, proliferation, RhoA
GTPase activity, gene expression, and synthesis of cartilage markers were analyzed.
5.2 Preparation and characterization of sulfated alginate
Alginate 2 g (5.0 mmol) was dissolved in 400 mL water and 40 g DOWEX
Marathon C ion exchanger that was previously charged with an equal mass of
tetrabutyl ammonium bromide. The mixture was stirred overnight, filtered and
isolated by lyophilisation resulting in a total final mass of 2.7 g. Alginate tetrabutyl
ammonium salt 2 g (2.4 mmol) was suspended in 200 ml dry DMF and a 5-fold excess
SO3/pyridine per disaccharide repeating unit was added and the mixture was stirred at
room temperature for 1 h. The opaque solution was precipitated in acetone, brought
to pH=12 (ethanolic NaOH) for 10 minutes and then neutralized. The precipitate was
filtered, dissolved in water and purified by dialysis. Lyophilisation gave the pure
product of 1.8 g in mass. The process of alginate sulfate synthesis is demonstrated in
Figure 5.1. An important parameter to characterize the chemical composition of the
alginate sulfate is the degree of substitution by sulfation (DSS). It gives the average
number of sulfate groups per disaccharide repeating unit of alginate which is formed
from β-D-mannuronate and α-L-guluronate. Based on this definition DSS values may
range between 0 (unsubstituted alginate) and 4.0 (complete substitution of all free

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
56
hydroxyls by sulfates). The DSS was determined by estimation of the sulfur content
using an automatic elemental analyser (CHNS-932, Leco, Moenchengladbach,
Germany). The degree of sulfation of the material used in this study was DSS = 0.8.
Two other materials with DSs of 1.1 and 2.6 were synthesized but did not form
hydrogels with 102 mM calcium and were not further considered. IR spectra were
obtained on an FT-IR-Spectrometer FTS 175 (BIO RAD, Krefeld, Germany) applying ATR
technique. NMR spectra were recorded in D2O at 343 K on a Bruker Advance 300 MHz
spectrometer. Spectral data found for the synthesized material were consistent with
those found in the literature for alginate sulfates prepared by different procedures
[166, 168, 169]. IR (ATR): 3445, 2946, 1613, 1417, 1232, 1169, 1021, 951, 831, 796 cm-
1. 13C-NMR (D2O): 178.1-177.6 (C 6), 102.3-101.8 (C-1), 80.9-69.9 (C-2, C-3, C-4), 66.3
ppm (C-5).
1. SO3/pyridine, DMF2. ethanolic NaOH
R = H, SO3Na
N+ Br-
DOWEX
N+2
OO
OH
OHNaOOC OH
O
HO O
NaOOC
OO
OH
OH-OOC OH
O
HO O
-OOC
OO
OR
ORNaOOC OR
O
RO O
NaOOC
Figure 5. 1. | Reaction scheme for the synthesis of alginate sulfate. In a first reaction step sodium
alginate is transformed into a tetrabutyl ammonium salt to improve the solubility in the reaction
medium followed by the sulfation of free hydroxyl groups of alginate with SO3/pyridine.

Chapter 5
57
5.3 Morphology of chondrocytes encapsulated in sulfated alginate
(DSs = 0.8)
Transmission and confocal microscopy were used to assess changes in the
cytoskeletal organization. The morphology of cells encapsulated in the different ratios
of unmodified to sulfated alginate hydrogels was similar except for cells cultured in
pure alginate sulfate (2%), which assumed a more spread, fibroblast-like morphology
after 7 days in culture (Figure 5.2E). Cells encapsulated in 1.5% alginate sulfate + 0.5%
alginate (Figures 5.2D and I) exhibited minor spreading and protruding filopodia. Cells
in 2% alginate, 0.5% alginate sulfate + 1.5% alginate and 1% alginate sulfate + 1%
alginate (Figures 5.2A-C and F-H) had a typical round morphology as commonly
observed for alginate encapsulated chondrocytes. Proliferation was visually higher in
the pure alginate sulfate hydrogel (Figures 5.2E and J) and more multi-nucleated cells
could be observed in the 1.5% alginate sulfate + 0.5% alginate (Figures 5.2D and I)
hydrogels compared to hydrogels with lower alginate sulfate content.

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
58
Figure 5. 2. | Morphology of cells encapsulated in alginate and alginate sulfate. Phalloidin/DAPI
staining and transmission imaging of P3 bovine chondrocytes encapsulated in alginate (Alg), alginate
sulfate (AlgSulf) and mixtures of Alg and AlgSulf for 7 or 21 days.
Sulfated GAGs are present in the ECM of most tissues; among these are
heparin, heparan sulfate, chondroitin sulfate, keratin sulfate and dermatan sulfate [13,
170]. These molecules have been shown to have important functions such as
anticoagulant [171, 172], growth factor and cytokine affinity [163, 164],
chondroprotective [27, 39], neuroprotective [173] and antioxidant properties [174].
Lately, Ronghua et al. [169] showed that alginate sulfate has comparable anticoagulant
properties to heparin. The use of alginate sulfate for tissue engineering applications is
limited to a few studies [167, 175]. In particular to our knowledge, a pure alginate
sulfate hydrogel was not previously reported. Different sulfating agents including
sulfuric acid-carbodiimide [166], chlorosulfonic acid-formamide [169] and various

Chapter 5
59
SO3/complexes [176, 177] have been used in the past to prepare sulfated alginate and
differences in the sulfation method may explain why calcium crosslinking was not
previously possible. Recently, it was shown that SO3/pyridine is a mild sulfating agent
for GAGs such as hyaluronan allowing proper control of the degree of sulfation (DSS),
especially if a homogeneous sulfate group distribution and low DSS values are desired
[178, 179]. We therefore decided to use SO3/pyridine for the sulfation of alginate. In
our work, addition of 3 mM CaCl2 was necessary to maintain stability of the gel for the
first 21 days of culture, however CaCl2 was removed from the cultures for the last 14
days and gels maintained stability due to high cell proliferation and matrix synthesis.
5.4 Assessment of cell proliferation within the hydrogels
Chondrocyte proliferation of cells encapsulated in 2% alginate sulfate, 2%
alginate and mixtures of the two components with 2% (w/v) total concentration was
assessed using the Calbiochem BrdU proliferation kit. Proliferation of encapsulated
cells measured by this assay corroborated the visual images of Figure 5.2. The 2%
alginate sulfate hydrogels had 5-fold higher proliferation when compared to 2%
alginate (p= 0.038) and the other mixtures (0.5% alginate sulfate + 1.5% alginate
(p=0.055), 1% alginate sulfate + 1% Alg (p=0.059) and 1.5% alginate sulfate + 0.5%
alginate (p=0.11)) (Figure 5.3).
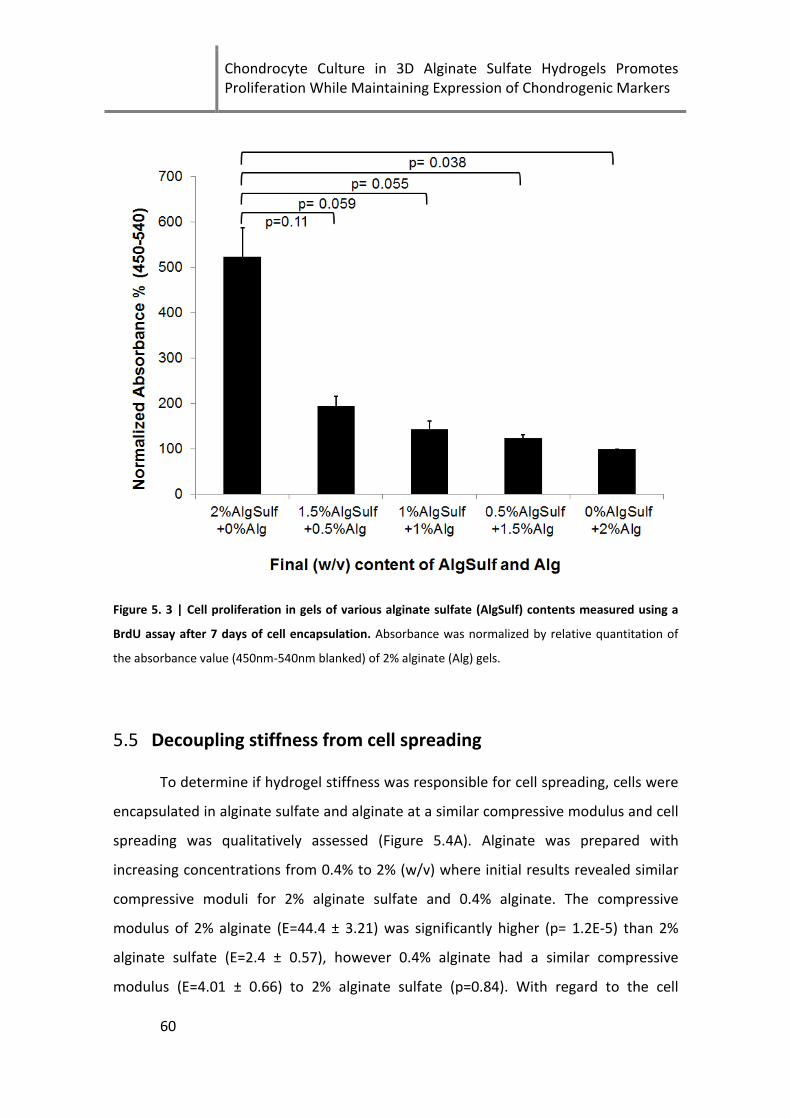
Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
60
Figure 5. 3 | Cell proliferation in gels of various alginate sulfate (AlgSulf) contents measured using a
BrdU assay after 7 days of cell encapsulation. Absorbance was normalized by relative quantitation of
the absorbance value (450nm-540nm blanked) of 2% alginate (Alg) gels.
5.5 Decoupling stiffness from cell spreading
To determine if hydrogel stiffness was responsible for cell spreading, cells were
encapsulated in alginate sulfate and alginate at a similar compressive modulus and cell
spreading was qualitatively assessed (Figure 5.4A). Alginate was prepared with
increasing concentrations from 0.4% to 2% (w/v) where initial results revealed similar
compressive moduli for 2% alginate sulfate and 0.4% alginate. The compressive
modulus of 2% alginate (E=44.4 ± 3.21) was significantly higher (p= 1.2E-5) than 2%
alginate sulfate (E=2.4 ± 0.57), however 0.4% alginate had a similar compressive
modulus (E=4.01 ± 0.66) to 2% alginate sulfate (p=0.84). With regard to the cell

Chapter 5
61
morphology, no cell spreading was observed in pure alginate hydrogels (both 0.4% and
2%) while cells showed significant spreading in alginate sulfate hydrogels (Figure 5.4B).
This finding strongly suggests that the lower stiffness of the alginate sulfate hydrogels
was not the cause of the extensive cell spreading in this material.
Figure 5. 4 | Mechanical properties of alginate sulfate (AlgSulf) hydrogels and effect on cell
proliferation. A) Average compressive modulus of alginate (Alg) gels at 2% and 0.4% (w/v) and AlgSulf
2% when subjected to unconfined compressive strain and B) Transmission images of passage 3
chondrocytes encapsulated in Alg at 2% and 0.4% (w/v) and in AlgSulf at 2% (w/v) for 7 days, scale bar
(50 µm).

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
62
5.6 RhoA and integrin signalling of chondrocytes in alginate sulfate
To explore whether cell morphology in alginate sulfate hydrogels was integrin
mediated, we tested the effect of blocking beta1 integrins on cell spreading.
Encapsulation within alginate sulfate hydrogels after incubation with the beta1
blocking antibodies showed a remarkable inhibition of cell spreading (Figure 5A). We
further investigated the effect of beta1 integrin blocking on cell proliferation. As
shown in Figure 5B, anti-beta1 integrin antibody caused a statistically significant
decrease in cell proliferation compared to untreated control (p=0.002), and to IgG
treated control (p=0.014). This evidence indicates that integrin signaling plays a role in
the physical interaction of the cells with the hydrogel and the increase in cell
proliferation. Chondrocyte proliferation has been associated with RhoA mediated
signaling pathways [180]. Therefore, we also investigated whether beta1 integrin could
act through a RhoA mediated mechanism. For this purpose, cells within 2%, 0.4%
alginate and 2% alginate sulfate maintained in culture in the presence/absence of
beta1 integrin blocking antibody or non-specific IgG were assayed for the degree of
RhoA activation. Interestingly, we found that RhoA activity was significantly higher in
2% alginate sulfate compared to pure alginate at 2% (p=0.003) and at 0.4% (p=0.026)
(Figure 5C). Moreover, beta1 integrin blocking produced a significant inhibition of
RhoA activity compared to the non-treated control (p=0.017), while IgG did not have
any significant effect (p=0.374). Those findings, taken together, indicate that beta1
integrin is responsible of cell protrusion and proliferation within alginate sulfate and
could act through the RhoA GTPase pathway.

Chapter 5
63
Figure 5. 5. | Integrin mediated cell spreading and proliferation within alginate sulfate (AlgSulf)
hydrogels and RhoA activity. A) Optical images of day 7 cultures of bovine chondrocyte encapsulated
within 2% AlgSulf gels + beta1 integrin antibody (β1) or IgG B) proliferation assay (the graph bars
represent the percentage values normalized against the untreated AlgSulf 2%) and C) OD 490 nm which
corresponds to the degree of RhoA activation in day 7 cultures within the hydrogels.

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
64
Proliferation and spreading of bovine chondrocytes on collagen based films
were shown earlier to be reduced by blocking beta1 integrins [124]. Moreover,
remodeling of three-dimensional collagen fibers and formation of aggregates by
mesenchymal stem cells was shown to be dependent on beta1 integrins [181]. We
observed here that blocking beta1 integrins prevented cell spreading and reduced
proliferation, suggesting that proliferation was integrin based and possibly through cell
synthesized collagen. Furthermore, the activity of RhoA was reduced with beta1
blocking indicating that proliferation was RhoA mediated. This is in agreement with
previous findings which report RhoA as an important regulator of chondrocyte
proliferation and differentiation.
5.7 Cyclin D1 expression is upregulated in alginate sulfate samples
To investigate if the proliferation within alginate sulfate was controlled by
genes regulating the cell cycle, expression of Cyclin D1 within alginate or alginate
sulfate hydrogels was investigated (Figure 5.6). Cyclin D1 gene expression was
upregulated 2-fold in alginate sulfate compared to alginate (p=0.74), indicating that
alginate sulfate possibly enhances proliferation by upregulation of Cyclin D1.

Chapter 5
65
Figure 5. 6. | Cyclin D1 expression. Relative gene expression of Cyclin D1 for bovine chondrocytes
encapsulated in alginate sulfate (AlgSulf) relative to alginate (Alg). RPL13 was used as a reference gene.
Beier and collaborators correlated the activity of RhoA with the increased
expression of Cyclin-D1, which is involved in regulating the cell cycle G1/S transition
[180]. Here we showed that cyclin-D1 gene expression was upregulated 2-fold in
alginate sulfate compared to alginate, thus indicating that alginate sulfate enhances
proliferation by ultimately regulating expression of cell cycle genes.
5.8 Expression of cartilage markers for dedifferentiated
chondrocytes within alginate sulfate hydrogels
The expression of relevant cartilage markers showed no significant difference in
expression between alginate sulfate and alginate indicating that the chondrocytes can
proliferate without undergoing de-differentiation which is typically associated with
expansion in monolayer culture. Interestingly, the expression of the catabolic gene
MMP13 was 11-fold lower in alginate sulfate samples after 35 days in culture (Figure

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
66
5.7). A 2-way ANOVA showed that sulfation had statistically no significant effect on
Col2/Col1 (p=0.56), Sox9/RUNX2 (p=0.48), Aggrecan (p=0.067) or MMP13 (p=0.99). On
the other hand the effect of culture time was only significant for Col2/Col1 ratio
(p=0.0087) and not significant for SOX9/RUNX2 (p=0.91), Aggrecan (p=0.16) and
MMP13 (p=0.12). Cells cultured on tissue culture plastic for 21 days exhibited a 36-fold
and 38-fold downregulation in Col2 and Aggrecan respectively compared to expression
in alginate at the same time point (data not shown).
Figure 5. 7. | Expression of relevant cartilage markers. Relative gene expression of Col2/Col1,
SOX9/RUNX2, Aggrecan and MMP13 for bovine chondrocytes encapsulated in alginate (Alg) or alginate
sulfate (AlgSulf) for 21 or 35 days. The gene expression of cells encapsulated in Alg for 21 days was used
as a reference sample and RPL13 as a reference gene.
5.9 Immunohistological staining and gross appearance of alginate
sulfate hydrogels
Encapsulation of bovine chondrocytes within the alginate sulfate hydrogels
induced an increase in cell proliferation ultimately resulting in an opaque cartilage like
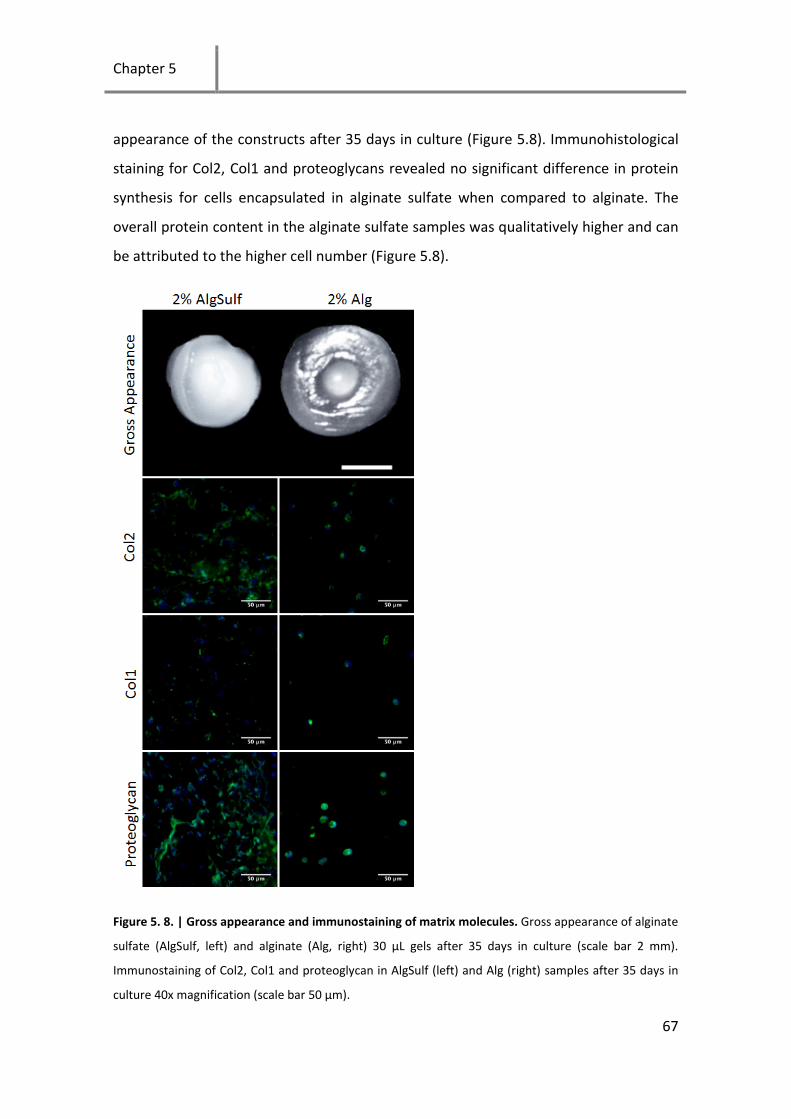
Chapter 5
67
appearance of the constructs after 35 days in culture (Figure 5.8). Immunohistological
staining for Col2, Col1 and proteoglycans revealed no significant difference in protein
synthesis for cells encapsulated in alginate sulfate when compared to alginate. The
overall protein content in the alginate sulfate samples was qualitatively higher and can
be attributed to the higher cell number (Figure 5.8).
Figure 5. 8. | Gross appearance and immunostaining of matrix molecules. Gross appearance of alginate
sulfate (AlgSulf, left) and alginate (Alg, right) 30 µL gels after 35 days in culture (scale bar 2 mm).
Immunostaining of Col2, Col1 and proteoglycan in AlgSulf (left) and Alg (right) samples after 35 days in
culture 40x magnification (scale bar 50 μm).

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
68
Alginate sulfate has been shown to have a high affinity to most heparin binding
growth factors [166]. Moreover, a scaffold containing alginate sulfate was shown to
improve chondrogenesis of encapsulated mesenchymal stem cells by gradual delivery
of transforming growth factor beta (TGFβ-1) [167]. Bovine chondrocytes were never
reported in combination with alginate sulfate and we hypothesized that the high
affinity to growth factors would result in a more chondrogenic environment for
encapsulated chondrocytes. However the focus of this study turned into the
unexpected strong proliferation and spreading of encapsulated chondrocytes and
whether chondrocytes would maintain their cartilage phenotype after long term
culturing. Chondrocytes have been traditionally reported to redifferentiate to a
cartilage phenotype after dedifferentiation when cultured in 3D hydrogels such as
agarose [60] or alginate [72, 161, 162]. However, cell proliferation in these systems is
limited and thus the regeneration capacity is low [182]. Collagen based hydrogels were
shown to induce cell proliferation at the expense of loss of the cartilage phenotype
[182, 183]. Cells encapsulated in alginate sulfate hydrogels exhibited a high degree of
proliferation while maintaining similar levels of cartilage markers and reducing
expression of MMP13. The effect of alginate sulfate on MMP13 expression may be
similar to the anti-inflammatory effect of chondroitin sulfate on MMPs through
interleukin-1 beta (IL-1β) [27, 41, 42]. However further investigation is needed to
confirm this hypothesis.
The large differences in proliferation rates between alginate and alginate
sulfate encouraged us to further investigate if the material was suitable for cartilage
tissue engineering applications. To test this possibility, dedifferentiated bovine
chondrocytes were encapsulated for up to 5 weeks within alginate and alginate sulfate
and the expression of relevant cartilage markers was analysed using qRT-PCR. Alginate
sulfation had no significant effect on the expression of Col2/Col1, SOX9/RUNX2, Agg
and MMP13. Similarly, at the protein level cells in both systems were not different.
Remarkably, alginate sulfate constructs attained a cartilage-like opaque appearance
after 5 weeks in culture.

Chapter 5
69
5.10 Chapter summary
In conclusion, alginate sulfate is a potential hydrogel for autologous
chondrocyte implantation that allows cells to synthesize their own matrix, proliferate
and maintain their cartilage phenotype. In addition, it has a proven high affinity to
important growth factors and can thus be loaded with relevant cartilage growth
factors for improved performance. Finally, the system may have strong action on the
inflammatory responses and thus may reduce complications resulting from increased
inflammation in the implant site. In conclusion, alginate sulfate may prove to be an
ideal biomaterial for cartilage tissue engineering.

Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers
70

Chapter 6
71
6 GFOGER Modified MMP-Sensitive
Polyethylene Glycol Hydrogels Induce
Chondrogenic Differentiation of Human
Mesenchymal Stem Cells
The work in this chapter has been submitted to Acta Biomaterialia Mhanna et al. (2013B)
In the previous chapter, we saw that functionalization of alginate with GAG
mimetic groups such as sulfates was sufficient to induce strong cell proliferation and
maintain the cartilage phenotype of 3D encapsulated chondrocytes. This was a
motivation to explore the effect of using a collagen mimetic peptide on the biological
responses of cells. In this chapter, the effects of the fibronectin adhesion sequence
RGD and the relatively novel collagen mimetic peptide GFOGER on chondrogenesis of
3D encapsulated human mesenchymal stem cells were investigated. Furthermore, the
effects of presenting the peptides in a degradable or non-degradable
microenvironment were analyzed.
6.1 Chondrogenic differentiation and the microenvironment
The limited ability of cartilage tissue to self-repair after injury or disease has
driven intensive efforts to engineer replacement tissue [12, 156]. Cell-based cartilage
tissue engineering holds strong promise to provide living tissue for cartilage repair [14,
49, 52]. Chondrogenic differentiation of stem cells has been widely studied using pellet
cultures [67, 184, 185], micromass cultures [186] and in hydrogels [187]. Both natural
[187-191] and synthetic [192-194] materials have been employed to induce

GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells
72
chondrogenesis of stem cells in 3D. Synthetic gels such as poly(ethylene glycol) (PEG)
are attractive for 3D culturing of cells [74], as they can be functionalized with specific
adhesion sequences [193], growth factor binding sites [195, 196] and protease-
sensitive motifs [197]. A PEG hydrogel modified with a matrix metalloproteinase
(MMP)-sensitive peptide was shown to increase chondrocyte proliferation and
upregulate mRNA levels of type II collagen and aggrecan in bovine chondrocytes [73].
The extracellular microenvironment is believed to play a key role in the
biological responses of stem cells [198, 199]. Modification of hydrogels with small
functional groups such as amines, phosphates and others were shown to induce
differentiation of stem cells into the osteogenic or adipogenic pathways [200].
Furthermore, adhesion sequences such as the RGD sequence has been shown to be
involved in adhesion [197], viability [201], proliferation [202] and differentiation of
stem cells [203, 204] however, an inhibition of chondrogenesis was earlier reported in
RGD-modified alginate hydrogels [96]. Chondrocytes form primary interactions with
collagen 2 and hyaluronic acid in the native tissue [13]. Recently Knight et al. [98]
identified the GFOGER sequence as the recognition site in type I collagen and type IV
collagen. Aggregation of chondrocytes in suspension was also shown to be mediated
by the GFOGER sequence which is recognized by α10β1 integrins [19]. Moreover, type II
collagen induced GAG deposition in chondrocytes was inhibited by GFOGER integrin
blocking peptides [205].
The GFOGER sequence has been used for modification of surfaces [206-208]
and hydrogels [205, 209] to control adhesion and biological responses of chondrocytes
and stem cells. When bovine bone marrow stromal cells were encapsulated in an
agarose gel with covalently coupled GFOGER sequences, type II collagen and aggrecan
mRNA stimulation as well as GAG deposition by chondrogenic factors was inhibited
[209]. However, human mesenchymal stem cells (hMSC) encapsulated in a soft PEG
hydrogel modified with the triple helical GFOGER peptide showed enhanced
chondrogenesis compared to the unmodified and the stiffer modified hydrogels [210].

Chapter 6
73
We hypothesized that GFOGER functionalized MMP-sensitive PEG gels would
enhance the chondrogenic differentiation of encapsulated hMSC stem cells, and that
RGD and GFOGER peptides would induce differential biological responses on
encapsulated cells. To test this hypothesis, hMSC were encapsulated in degradable
(MMP-sensitive) or non-degradable PEG hydrogels containing RGD or GFOGER
sequences. Initial hydrogel compressive modulus, cell morphology, proliferation, gene
expression and synthesis of chondrogenic markers were analysed.
6.2 Hydrogel modification did not affect mechanical properties
Degradable and non-degradable hydrogels modified with 100 µM RGD, 100 µM
GFOGER or non-modified were allowed to swell in cell culture medium in the incubator
for 24 hours after production. Compressive moduli of the equilibrated hydrogels were
measured using a texture analyzer (TA.XTPlus, Stable Microsystems, UK). This step was
essential to prove that differential biological responses from the peptide conjugations
were not due to changes in the materials’ compressive moduli.
The compressive moduli of the hydrogels were not significantly different among
the different tested conditions (Figure 6.1). A two-way ANOVA test revealed that there
was no significant influence on the mechanical properties, neither due to the
degradability of the gels (p=0.35) nor to the added adhesion peptide (p=0.32). The
interaction of degradability and peptide parameters also showed no significance
(p=0.75). We can therefore assume that cell behavior was not affected by the initial
mechanical properties of the gels. Furthermore, the average compressive modulus of
the hydrogels was 3.7±0.3 kPa which is similar in magnitude to the values stated by the
manufacturer in the materials certificate of analysis.

GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells
74
Figure 6. 1. | Compressive moduli of the hydrogels. Compression test of hydrogels containing 100 µM
RGD, 100 µM GFOGER or without adhesion peptides (No peptide). Young's moduli are shown for the
non-degradable gels (Non-deg) as well as for the degradable gels (Deg).
6.3 Cell viability
The viability of MSCs encapsulated in the PEG hydrogels was affected by gel
degradability as well as peptide incorporation (Figure 6.2). Cell viability was higher in
degradable PEG hydrogels compared to non-degradable. Cells had a higher viability in
gels modified with the adhesion sequences, though there was not a major difference
between gels containing the RGD sequence versus those containing the GFOGER
sequence.

Chapter 6
75
Figure 6. 2. | Cell viability in the hydrogels. Merged live/dead assay of stem cells encapsulated for 21
days in non-degradable (Non-Deg) and degradable (Deg) PEG gels. The image shows live (green) and
dead (red) cells simultaneously. Images were taken with a 10x objective, scale bar 100 µm.
6.4 Cell morphology
MSCs in PEG hydrogels exhibited very different actin cytoskeletal architecture
depending on the specific integrin binding domain incorporated into the PEG matrix. If
no peptide adhesion sequence was used, cells remained round in the maximum
intensity projections of the Z-stacks (Figure 2A, D and G). In RGD and GFOGER
containing PEG gels, cells spread and developed stress fibers (Figure 2B, C, E, F, H and
I). This effect was much more pronounced in the degradable gels that permitted the
cells to remodel their matrix and physically create space in the 3D network for this
elongated morphology (Figure 2E, F, H and I). Cell protrusions were observed in non-
degradable peptide containing gels, though spreading was clearly restricted. The
elongated shape of spread cells varied strikingly in RGD-PEG versus GFOGER-PEG gels.
In RGD-PEG gels, the cells tended to be smaller and contained thin star-like projections
of cytoskeleton ending at a matrix attachment site in a sharp point (Figure 2B, E and
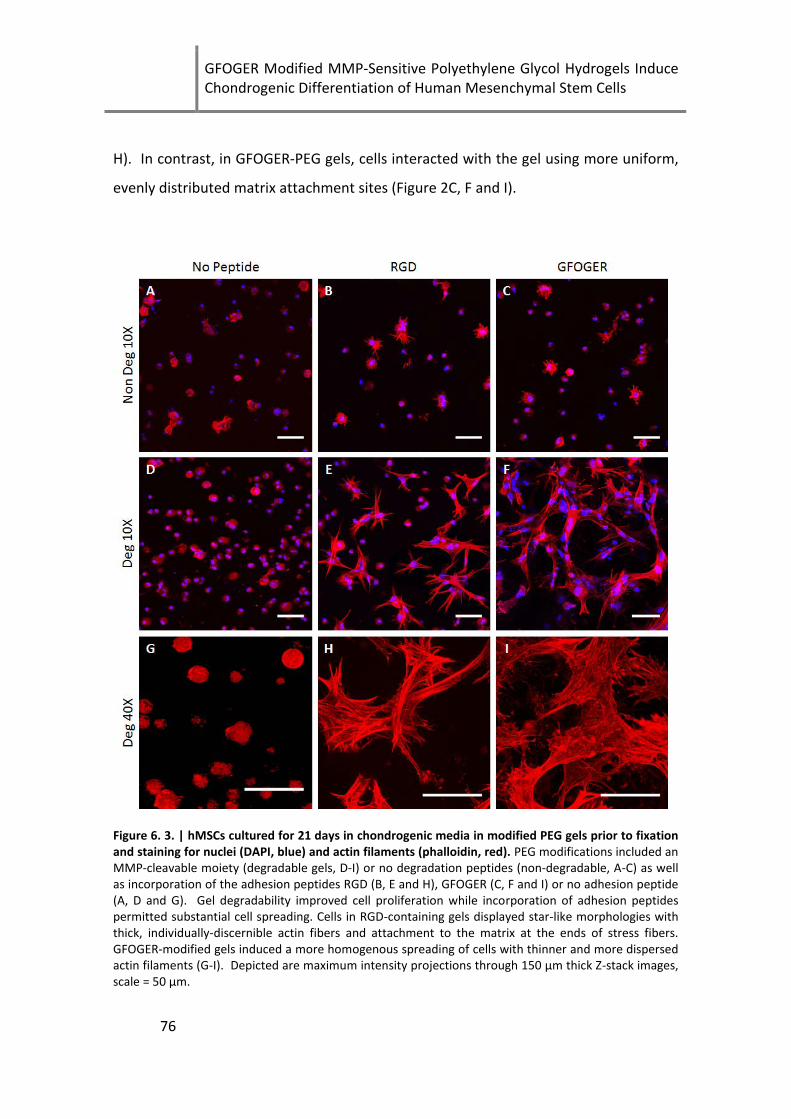
GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells
76
H). In contrast, in GFOGER-PEG gels, cells interacted with the gel using more uniform,
evenly distributed matrix attachment sites (Figure 2C, F and I).
Figure 6. 3. | hMSCs cultured for 21 days in chondrogenic media in modified PEG gels prior to fixation and staining for nuclei (DAPI, blue) and actin filaments (phalloidin, red). PEG modifications included an MMP-cleavable moiety (degradable gels, D-I) or no degradation peptides (non-degradable, A-C) as well as incorporation of the adhesion peptides RGD (B, E and H), GFOGER (C, F and I) or no adhesion peptide (A, D and G). Gel degradability improved cell proliferation while incorporation of adhesion peptides permitted substantial cell spreading. Cells in RGD-containing gels displayed star-like morphologies with thick, individually-discernible actin fibers and attachment to the matrix at the ends of stress fibers. GFOGER-modified gels induced a more homogenous spreading of cells with thinner and more dispersed actin filaments (G-I). Depicted are maximum intensity projections through 150 μm thick Z-stack images, scale = 50 μm.
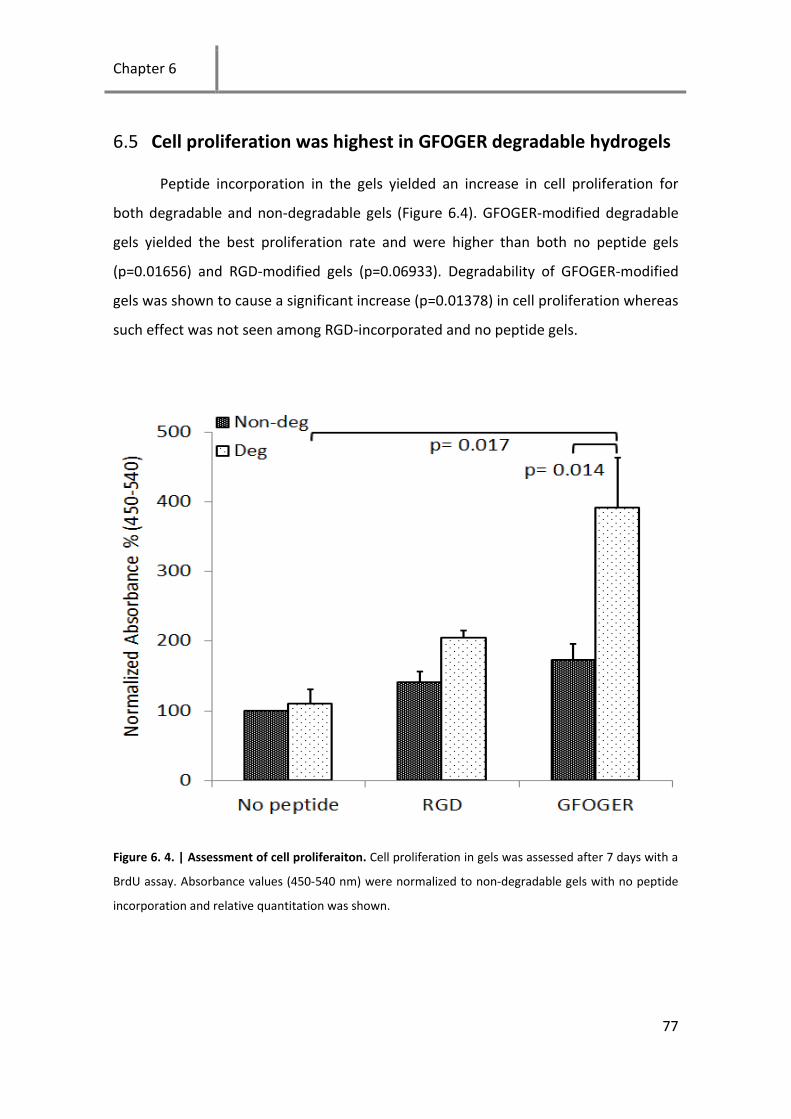
Chapter 6
77
6.5 Cell proliferation was highest in GFOGER degradable hydrogels
Peptide incorporation in the gels yielded an increase in cell proliferation for
both degradable and non-degradable gels (Figure 6.4). GFOGER-modified degradable
gels yielded the best proliferation rate and were higher than both no peptide gels
(p=0.01656) and RGD-modified gels (p=0.06933). Degradability of GFOGER-modified
gels was shown to cause a significant increase (p=0.01378) in cell proliferation whereas
such effect was not seen among RGD-incorporated and no peptide gels.
Figure 6. 4. | Assessment of cell proliferaiton. Cell proliferation in gels was assessed after 7 days with a
BrdU assay. Absorbance values (450-540 nm) were normalized to non-degradable gels with no peptide
incorporation and relative quantitation was shown.

GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells
78
6.6 Gene expression
Gene expression of type II collagen and aggrecan were highest in GFOGER-
modified degradable gels after 7 and 21 days (Figure 6.5). Gene expression of type II
collagen and aggrecan changed significantly with time (p=0.016) and (p=0.026)
respectively. After 21 days in culture only GFOGER and RGD modified degradable
hydrogels exhibited significantly higher levels of type II collagen when compared to
pellets cultured in non-chondrogenic medium (p=0.008) and (p=0.0324) respectively.
Moreover, expression of type II collagen was higher in GFOGER-modified degradable
gels compared to non-modified degradable gels (p=0.038). On the other hand, only
GFOGER-modified degradable gels had a significantly higher level of aggrecan when
compared to pellets in non-chondrogenic medium (p=0.018).

Chapter 6
79
Figure 6. 5. | Gene expression of relevant cartilage markers. Gene expression of type II collagen and
aggrecan were determined using quantitative real time PCR (StepOnePlus, Applied Biosystems). Non-
degradable hydrogels without modification (0 µM Deg) cultured in control media (control) were used as
reference sample and RPL 13 was used as a reference gene.
6.7 GAG production in peptide-modified gels
Peptide incorporation increased GAG production in degradable gels after 7 days
and 21 days and GFOGER-modified degradable hydrogels had on average the highest
GAG content (Figure 6.6). After 7 days GFOGER-modified degradable gels had an
almost significant increase in GAG/DNA (p=0.059). Non-degradable gels led to similar
production of GAG with or without peptide at both time points.

GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells
80
Figure 6. 6. | GAG/DNA quantification in the various conditions. DNA was quantified using picogreen
and GAG was quantified using the DMMB assay. GAG quantity was normalized to the DNA amount.
6.8 Histology and immunostaining
GAG synthesis was confirmed qualitatively using Alcian blue staining. GFOGER
degradable hydrogels showed the most prominent staining compared to all other
hydrogel conditions (Figure 6.7). Pellets cultured in chondrogenic medium (Figure 6.7
A) showed stronger GAG staining compared to cells cultured in control medium (Figure
6E). In hydrogels with no adhesion peptide modification, staining was limited and only
visible around the cells (Figure 6 B and F). In RGD modified gels, staining was similar to
the unmodified gels (Figure 6 C and G) whereas in GFOGER modified gels staining was
very strong around the cells in the non-degradable hydrogels with little GAGs in the
matrix (Figure 6 D) and was distributed strongly all over the sample in degradable
hydrogels (Figure 6 H). Alcian blue staining in chondrogenic pellets (Figure 6E) was
comparable to degradable hydrogels modified with the GFOGER peptide although the
cell concentration in pellets is clearly higher.
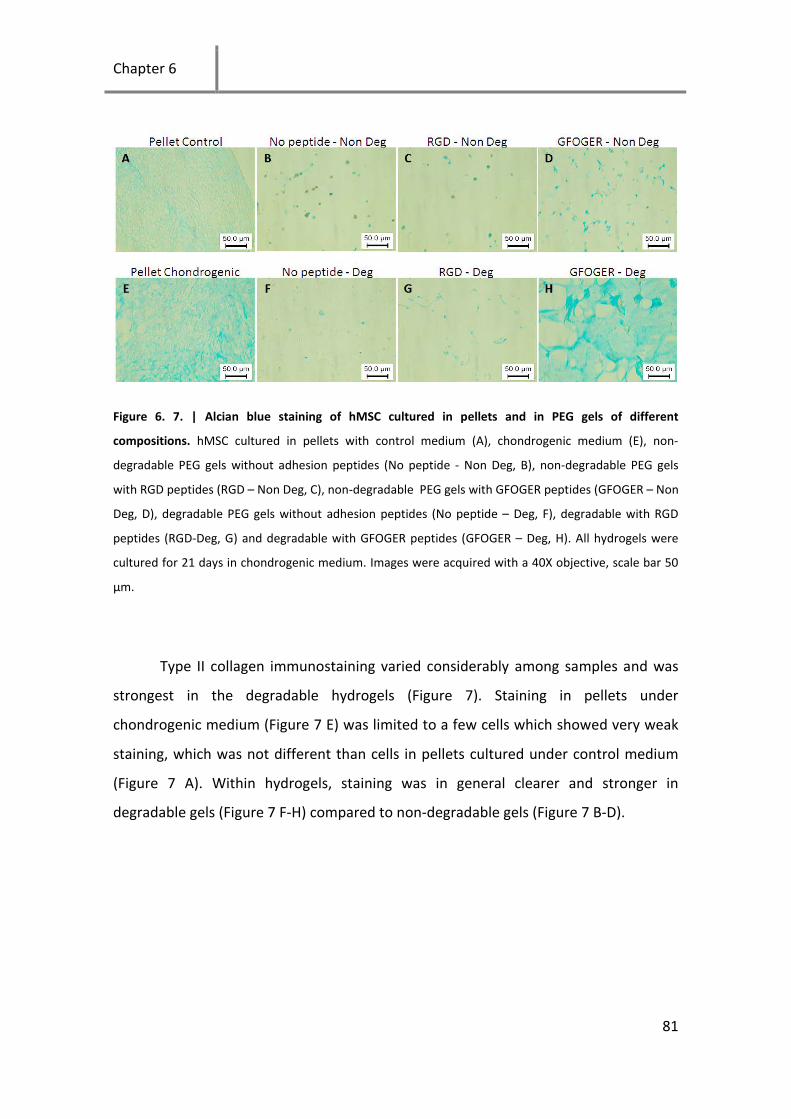
Chapter 6
81
Figure 6. 7. | Alcian blue staining of hMSC cultured in pellets and in PEG gels of different
compositions. hMSC cultured in pellets with control medium (A), chondrogenic medium (E), non-
degradable PEG gels without adhesion peptides (No peptide - Non Deg, B), non-degradable PEG gels
with RGD peptides (RGD – Non Deg, C), non-degradable PEG gels with GFOGER peptides (GFOGER – Non
Deg, D), degradable PEG gels without adhesion peptides (No peptide – Deg, F), degradable with RGD
peptides (RGD-Deg, G) and degradable with GFOGER peptides (GFOGER – Deg, H). All hydrogels were
cultured for 21 days in chondrogenic medium. Images were acquired with a 40X objective, scale bar 50
μm.
Type II collagen immunostaining varied considerably among samples and was
strongest in the degradable hydrogels (Figure 7). Staining in pellets under
chondrogenic medium (Figure 7 E) was limited to a few cells which showed very weak
staining, which was not different than cells in pellets cultured under control medium
(Figure 7 A). Within hydrogels, staining was in general clearer and stronger in
degradable gels (Figure 7 F-H) compared to non-degradable gels (Figure 7 B-D).

GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells
82
Figure 6. 8. | Type II collagen immunostaining of hMSC cultured in pellets and in PEG gels of different
compositions. hMSC cultured in pellets with control medium (A), chondrogenic medium (E), non-
degradable PEG gels without adhesion peptides (No peptide - Non Deg, B), non-degradable PEG gels
with RGD peptides (RGD – Non Deg, C), non-degradable PEG gels with GFOGER peptides (GFOGER – Non
Deg, D), degradable PEG gels without adhesion peptides (No peptide – Deg, F), degradable with RGD
peptides (RGD-Deg, G) and degradable with GFOGER peptides (GFOGER – Deg, H). All hydrogels were
cultured for 21 days in chondrogenic medium. Images were acquired with a 40X objective, scale bar 50
μm.
A major requirement for an engineered 3D biomaterial to be used for tissue
engineering applications is to maintain cell viability for a long culturing period.
Additionally, increased proliferation may be desirable for tissue formation [211].
Although the viability of fibroblasts [197] or chondrocytes [73] is not significantly
affected by the materials degradability, stem cells have been reported to have a higher
viability in degradable hydrogels [211]. Furthermore incorporation of adhesion
sequences like RGD [212], IKVAV [211] and GFOGER [210] were shown to improve
stem cells viability in PEG gels. We show here that the presence of MMP motifs alone

Chapter 6
83
did not have a significant effect on cell proliferation or cell viability. However, the
incorporation of adhesion peptides in both degradable and non-degradable hydrogels
improved cell viability and cell proliferation which is consistent with the literature
[210-212]. In general, incorporation of peptide adhesion sequences improved
proliferation however only GFOGER degradable hydrogels had a significantly higher
proliferation (p=0.017) compared to the degradable hydrogels without any adhesion
peptides. Degradability was also only a statistically significant factor in the presence of
GFOGER sequences (p=0.014).
A round cell morphology is often linked to a cartilage phenotype [60, 213].
Chondrogenesis of bone marrow stem cells has been shown to be inhibited in RGD
modified alginate [96]. Moreover, chondrogenesis in agarose was inhibited in the
presence of RGD sequences and this inhibition was blocked by disruption of the f-actin
cytoskeleton using cytochalasin D [97]. These results are in contradiction with the
findings of the current study where chondrogenic differentiation was strongest in RGD
and GFOGER modified degradable hydrogels in which cells had a clearly more spread
morphology than the other conditions. In general the trend observed in the current
study was that the increase in proliferation was associated with increase in type II
collagen. These results indicate that cell spreading and proliferation do not necessarily
inhibit the onset of chondrogenesis and that the microenvironment and growth factors
may be stronger players. Moreover, cells encapsulated in RGD modified degradable
hydrogels had a star-like morphology with strong actin fibers whereas cells in GFOGER
had a uniaxial spreading with more diffuse and less intense actin fibers which might
better support chondrogenesis.
Several studies have employed RGD sequences for improving cell adhesion
[197], viability [201] and chondrogenesis [203, 204] of stem cells in 3D PEG gels.
However, very few studies investigated the incorporation of the GFOGER peptide in 3D
hydrogels and their influence on chondrogenesis [209, 210]. In particular the effect of
hydrogel degradability on the response of stem cells to the GFOGER peptide was not

GFOGER Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells
84
reported. Furthermore, a study comparing the effect of presenting equal amounts of
the GFOGER and RGD peptides on chondrogenesis is presented here for the first time.
We observed that the presence of MMP motifs in the PEG hydrogels greatly influenced
the cell response to GFOGER and RGD peptides in terms of cell spreading and
proliferation. Furthermore, degradability was essential to induce chondrogenesis in
peptide modified hydrogels. Finally, it was observed that when degradable PEG gels
were modified with adhesion sequences chondrogenesis was better than in pellet
cultures.
6.9 Chapter summary
GFOGER modified hydrogels represent a new biomimetic option for cell growth
in a 3D hydrogel. The GFOGER peptide induces higher proliferation of stem cells and
better maintains a chondrogenic phenotype compared to RGD in terms of expression
of the chondrogenic markers type II collagen and aggrecan and alcian blue staining.
Degradability and the presence of adhesion peptides in 3D cultures are shown here to
be necessary to maintain a high viability and initiate chondrogenesis of MSCs. Taken
together, the results of the current study suggest that GFOGER peptides presented in a
degradable hydrogel provide a suitable microenvironment for proliferation and
chondrogenesis of hMSCs.

Chapter 7
85
7 Probing the Microenvironmental Conditions
for Induction of Superficial Zone Protein
Expression
The work in this chapter is in revision in Osteoarthritis and Cartilage, Mhanna et al. (2013C).
In the previous chapters we demonstrated the importance of the
microenvironment on chondrocytes and stem cells. We showed that small sulfate
groups and peptides had profound effects on cell proliferation and expression of
chondrogenic markers. In the current chapter we investigate the effect of cell culture
dimensionality, mechanical stimulation, oxygen tension and cell morphology on the
expression of superficial zone protein and type II collagen.
7.1 Chondrocyte dedifferentiation and superficial zone protein
The development of replacement tissue for healing cartilage injuries poses a
major scientific challenge. The avascular nature, limited proliferation and restricted
access to nutrients and regenerative cells prevent the spontaneous regeneration of
cartilage defects [12, 214]. Furthermore, the dense matrix that surrounds cartilage
cells (chondrocytes) inhibits their migration to the defect site [14, 156]. Articular
cartilage is organized into the deep, middle and superficial zones [13, 215]. The
superficial zone in cartilage is believed to be crucial for long term stability and function
of the tissue by protecting the deeper layers and providing lubricants that limit friction
and prevent wear [50]. Superficial zone protein (SZP) which is highly homologous to

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
86
lubricin is mainly produced by the chondrocytes of the superficial zone and is a key
molecule involved in cartilage lubrication [17, 216, 217].
Chondrocytes have been shown to dedifferentiate when cultured on two-
dimensional (2D) tissue culture plastic where they assume a fibroblastic phenotype
[59, 61, 159]. Culturing chondrocytes in alginate [72, 161, 162] (Alg) or agarose [60]
hydrogels has been shown to maintain the cartilage phenotype. However, most studies
address the re-expression of type II collagen (Col2) and aggrecan which are present
throughout cartilage tissue but are synthesized in lower amounts by superficial zone
cells compared to cells of the deeper layers [13]. Only a few studies have investigated
the effects of dedifferentiation on the superficial zone phenotype and particularly the
expression of SZP [61].
The zones of articular cartilage are important to the function of the tissue,
making an engineered cell-based cartilage replacement that recapitulates the stratified
cartilage structure desirable. In the current chapter we focus on microenvironmental
factors which regulate the expression of superficial zone protein. Synthesis of SZP has
been shown to be affected by growth factors such as bone morphogenic protein-7
(BMP-7) [218] and transforming growth factor beta (TGFβ) [219] as well as cytokines
such as interleukin-1 beta (IL-1β) [220]. Gene expression and synthesis of superficial
zone protein is also modulated by mechanical stimulation such as shear [118], surface
motion [116] and mechanical stress [117]. Finally, differentiation under hypoxic
conditions stimulated matrix production by middle/deep chondrocytes and
proteoglycan 4 by superficial zone chondrocytes [95].
We hypothesized that an in-vitro system that most closely resembles the in-vivo
cartilage superficial zone environment would allow dedifferentiated chondrocytes to
re-express markers of the superficial zone. To test this hypothesis we designed
experiments to address the effect of 3D cultures, cyclic tensile strain, oxygen tension
and cell morphology on the expression of superficial zone protein. We designed a
mechanical loading chamber for application of homogeneous 3D strain compatible
with the commercially-available STREX machine. We then cultured chondrocytes on 2D

Chapter 7
87
substrates and 3D hydrogels exhibiting varying degrees of spreading in the presence
and absence of cyclic mechanical strain and quantified the expression of SZP. We also
studied the expression of SZP at ambient oxygen levels as well as under hypoxic
conditions and compared its expression profile to that of Col2.
7.2 Both SZP and Col2 undergo dedifferentiation during serial
passaging
Serially passaged chondrocytes exhibited a massive downregulation in Col2
expression (Figure 7.1B) similar to what has been reported in previous studies. The
downregulation was significant in cartilage tissue, p0 and P1 when compared to
passage 4. The expression of SZP decreased with passaging, exhibiting a similar trend
to that reported for Col2 (Figure 7.1A). The relative quantity decreased from 78 fold in
cartilage tissue to 32 fold at P0 d4 normalized to P4 d4. The downregulation was
significant between cartilage tissue and P0 compared to passage 4, but not between
passage 2 and 4.
Figure 7. 1. | Effect of chondrocyte dedifferentiation on gene expression of SZP and type II collagen.
Chondrocytes were harvested from cartilage tissue and serially passaged by culturing on tissue culture
plastic for 4 days from passage 0 to passage 4. Analysis of SZP mRNA levels (A) and Col2 (B) showed a
decrease in expression with passaging. Gene expression was normalized to expression in P4, (n=4), *
p<0.05 and *** p<0.001.

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
88
Chondrocyte dedifferentiation is a common phenomenon observed in 2D
culture and we show here that the reduction in SZP expression along with the
precipitous drop in Col2 expression can be now considered a part of the de-
differentiation phenomenon associated with monolayer culture.
7.3 Redifferentiation of serially passaged primary chondrocytes
Chondrocytes encapsulated in alginate beads for 4 days re-expressed SZP but
not Col2 (Figure 7.2). The expression of SZP was upregulated 52.3 fold (p=0.00013) in
alginate beads compared to chondrocytes cultured on TCP (Figure 7.2A). This is
comparable to the expression in cartilage tissue indicating that the cells could almost
completely restore the expression of SZP by 3D culture. The expression of Col2 was
however downregulated 2.7 fold after 4 days in alginate culture (Figure 7.2A). The
downregulation of Col2 expression in alginate beads although statistically insignificant
(p=0.08) is in contradiction to the finding of several studies which show that
chondrocytes re-express col2 in alginate beads [72, 161, 162]. In order to confirm the
expression of SZP at the protein level, immunostaining for SZP was performed on
alginate beads and TCP samples. Strong staining for SZP was found in alginate
encapsulated cells but not in cells cultured on TCP (Figure 7.2B).

Chapter 7
89
Figure 7. 2. | Redifferentiation of serially passaged chondrocytes. Passage 3 chondrocytes were
cultured on tissue culture plastic (TCP) for 4 days or encapsulated in alginate beads (Beads) for the same
period to determine if expression of SZP and Col2 could be restored. A) qRT-PCR mRNA levels for SZP
and Col2 on TCP and within alginate beads normalized to TCP gene expression (n=5) and B)
immunostaining of SZP in cells cultured on TCP and in alginate beads, scale bar 50 μm.
Chondrocytes have been traditionally reported to redifferentiate to a more
chondrogenic phenotype when cultured in 3D hydrogels [62]. The mechanism by which
3D cultures allow redifferentiation of chondrocytes is not well understood but involves
the integrity of the cytoskeleton. The prevention of cell spreading in 2D cultures by the

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
90
ROCK kinase inhibitor Y-27632 induced a cortical actin morphology and was shown to
redifferentiate chondrocytes. Likewise, interference with actin polymerization by
cytochalasin D or jasplakinolide helped restore the chondrogenic phenotype in
dedifferentiated cells [70, 71]. The addition of concanavalin A to chondrocyte cultures
also caused cell rounding and increased proteoglycan synthesis [77]. Dedifferentiation
and redifferentiation studies have often focused on the expression of Col2, Col1 and
aggrecan while little attention has been given to the behavior of the main superficial
zone marker, SZP. We observed that SZP is downregulated with passaging similar to
Col2 and its expression can be restored within 4 days of encapsulation in alginate
beads unlike type II collagen.
7.4 Design and evaluation of a 3D tension and compression
chamber compatible with the STREX strain machine
The chamber was designed using Key Creator v7.02 (Figure 7.3A) and metal
molds were produced with a milling machine (Deckel FP 3 CC, Germany). The molds
were used for casting the 3D PDMS chamber (Figure 7.3B).

Chapter 7
91
Figure 7. 3. | Design for the 3D molds A and the final 3D chamber B.
The STREX machine allows for different magnitudes of strain to be applied to a
given chamber. For each tension/compression setting available in the STREX device,
the corresponding measured strain in the strained hydrogel was measured (Table 7.1).
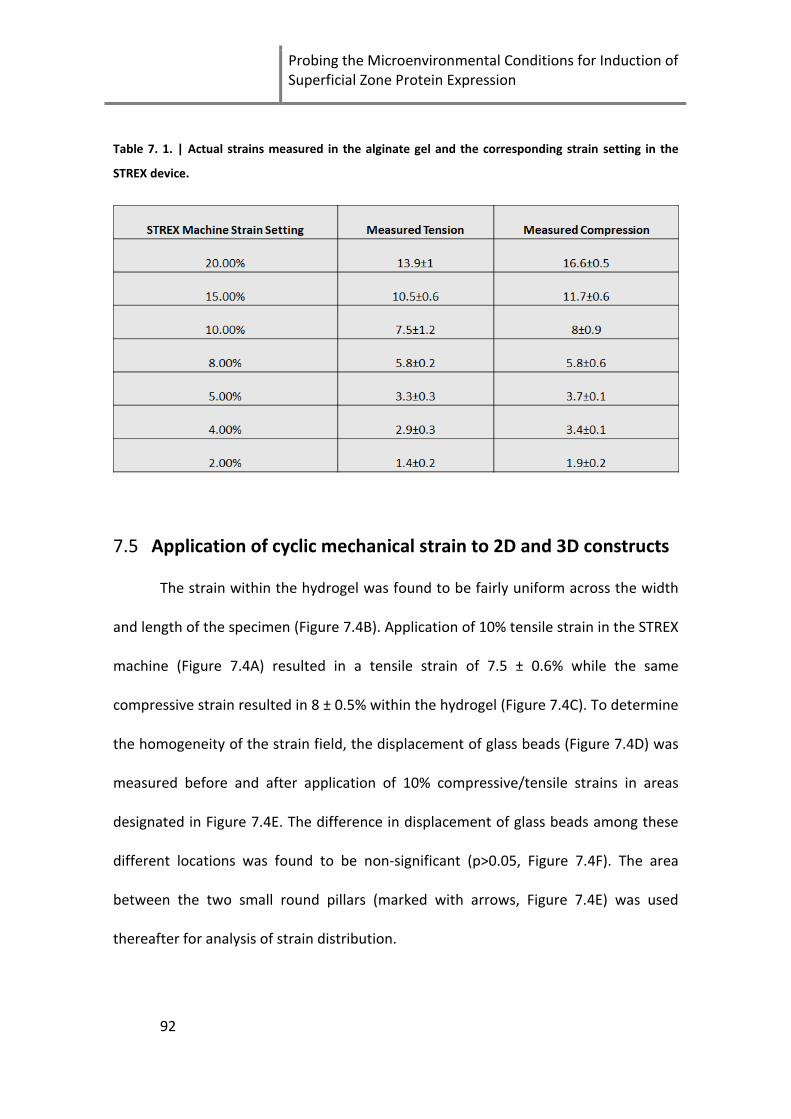
Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
92
Table 7. 1. | Actual strains measured in the alginate gel and the corresponding strain setting in the
STREX device.
7.5 Application of cyclic mechanical strain to 2D and 3D constructs
The strain within the hydrogel was found to be fairly uniform across the width
and length of the specimen (Figure 7.4B). Application of 10% tensile strain in the STREX
machine (Figure 7.4A) resulted in a tensile strain of 7.5 ± 0.6% while the same
compressive strain resulted in 8 ± 0.5% within the hydrogel (Figure 7.4C). To determine
the homogeneity of the strain field, the displacement of glass beads (Figure 7.4D) was
measured before and after application of 10% compressive/tensile strains in areas
designated in Figure 7.4E. The difference in displacement of glass beads among these
different locations was found to be non-significant (p>0.05, Figure 7.4F). The area
between the two small round pillars (marked with arrows, Figure 7.4E) was used
thereafter for analysis of strain distribution.

Chapter 7
93
Figure 7. 4. | Design for application of 3D cyclic strain. A) STREX Device. B) New chamber designed for
3D loading. Metal tubes were inserted in the holes connecting the chamber to the machine to minimize
non-uniform deformations around the holes. C) Strain measured within a hydrogel in response to 10%
compressive/tensile strain set by the instrument. D) Glass beads embedded in an alginate gel (scale bar
200 μm). E) Schematic representation of the chamber: The gel was injected in the blue area, letters (a, b
and c) mark areas where images of glass beads were taken. Three images were taken 1 mm away from
the small round pillar marked with an arrow (a), 3 images at the same level closer to the wall (b) and 3
images were taken in the center middle (c) to cover the whole gel in one plane. To assess variation in
the z-axis, images were taken at 2 different depths resulting in a total of 18 images per gel. In each
image 4 measurements of displacement were performed. F) The homogeneity was assessed by
measuring the displacement before/after compression/tension of 10-30 μm glass beads. Displacement
was found to have no significant difference between the various locations. (n=3, p>0.05).
The STREX device has been previously employed for application of mechanical
strain in 2D; however limited studies have employed the system for 3D strain [221].
These studies did not verify uniformity of strains within the strained hydrogel and in
our experience could result in non-homogeneous strains throughout the gel.

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
94
Furthermore, the previously reported chambers make it difficult if not impossible to
apply strain to alginate gels without adhesion to PDMS. The 3D pillared chamber
designed for this study was shown to allow application of homogenous strain to
alginate. The strain within the gel appeared to be lower than the strain applied to the
chamber where a 20% applied strain corresponded to approximately 14% strain in the
hydrogel. Inhomogeneity in strain was partially solved by inserting metal tubes that fit
tightly in the connection points between the chamber and the device. Additionally, the
alginate beam was cast in only the middle third of the chamber which gave a uniform
strain across the specimen width. In addition to the homogeneous 3D strain
distribution, the system allows a direct comparison between application of cyclic strain
to cells in 2D and cells in 3D. The current system is an excellent tool for application of
homogeneous 3D strain to cells embedded in a hydrogel and may be used for several
biomedical applications including but not limited to tendon, muscle and bone tissue
engineering.
7.6 Effect of mechanical strain on SZP expression
Gene expression of superficial zone protein (SZP) was upregulated in 2D and 3D
cultures by mechanical strain of 2h/day (Figure 7.5). A single application of mechanical
strain at day 4 of culture resulted in 1.5 fold upregulation in SZP while continuous
strain for 2h/day for 4 days resulted in a 2.4 fold upregulation in 3D cultures (Figure
7.5B). Cells seeded on collagen coated PDMS did not show a response to the single
strain treatment while a strain of 2h/day resulted in a 1.7 fold upregulation (Figure
7.5A). Although no statistical significant differences were obtained between group
means as calculated by two-way ANOVA (p=0.31), the data indicate a clear
upregulation trend especially with application of 2h/day strain.

Chapter 7
95
Figure 7. 5. | The effect of mechanical strain on SZP mRNA levels was measured by qRT-PCR. A)
Response of cells seeded on collagen coated PDMS to 2h strain applied 4 days post seeding or applied
every day for 2 h throughout the culture period (total of 4 times). B) Response of cells to the same strain
regime when encapsulated in alginate gels (n=4). Gene expression was normalized to unstrained
samples.
Physiologic mechanical loading has been shown to stimulate gene expression
and synthesis of chondrogenic markers [93, 99, 106, 222, 223]. The mechanism
through which chondrocytes sense mechanical load is not completely understood but
may involve stretch-activated ion channels and integrin signalling [114]. External
signals sensed by cell membrane proteins are transduced through molecular pathways
leading to the expression of certain genes [94]. To further augment the expression of
SZP, we used the commercially available STREX device to apply cyclic mechanical strain
to chondrocytes cultured in 2D and 3D [108, 221]. Tensile strains are expected to be
highest in the superficial zone and thus were expected to induce expression of
superficial zone markers [22]. We showed using this system that mechanical strain
upregulated SZP expression up to 2.4 fold in 3D and 1.7 fold in 2D. These values are
similar in magnitude to previously reported responses to mechanical stimulation [116,
117].
7.7 Effect of oxygen tension on expression of SZP
Under hypoxic conditions (1% pO2) Col 2 was upregulated while SZP was
downregulated. Cells encapsulated in alginate beads exhibited a 49.5 fold upregulation

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
96
in Col2 gene expression (p=0.014) under hypoxic conditions compared to normoxia
(Figure 7.6A). The expression of SZP on the other hand was downregulated 3 fold in
hypoxia (p=0.15). This observation is in accordance with the oxygen distribution in
cartilage tissue where the superficial zone oxygen levels are estimated to be between
7-10% (Figure 7.6B and C). In the deep and middle zones, where oxygen levels are less
than 1 %, SZP is not present and Col2 is highly abundant (Figures 7.6B and C). The
expression of SZP was also downregulated 7.7 fold in TCP cultures under hypoxia
(p=0.055), however, Col2 levels did not vary (RQ=1.1 fold, P=0.87). This indicates that
SZP is responsive to oxygen levels in 2D but Col2 is not.
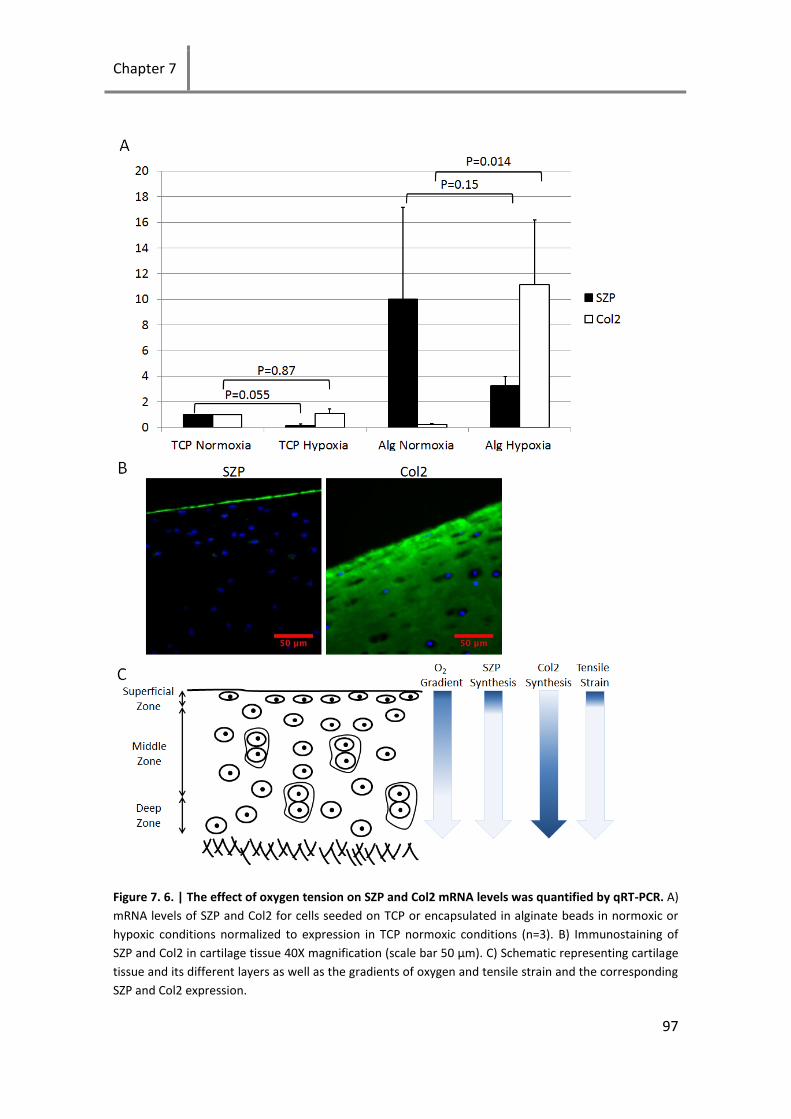
Chapter 7
97
Figure 7. 6. | The effect of oxygen tension on SZP and Col2 mRNA levels was quantified by qRT-PCR. A) mRNA levels of SZP and Col2 for cells seeded on TCP or encapsulated in alginate beads in normoxic or hypoxic conditions normalized to expression in TCP normoxic conditions (n=3). B) Immunostaining of SZP and Col2 in cartilage tissue 40X magnification (scale bar 50 μm). C) Schematic representing cartilage tissue and its different layers as well as the gradients of oxygen and tensile strain and the corresponding SZP and Col2 expression.

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
98
The avascular nature of cartilage leads to low oxygen levels compared to
vascularized tissues. Several studies have investigated the influence of oxygen tension
on chondrocyte metabolic and catabolic activity [21, 64, 95, 224-226]. These studies
showed an increased Col2 and aggrecan expression [64, 95, 224, 226] and improved
GAG deposition [225] in hypoxic compared to normoxic conditions. The effects of
oxygen tension on the expression of superficial zone markers are less investigated.
Schrobback et al. [95] found increased SZP mRNA levels in hypoxic compared to
normoxic conditions. However, this study used 5% oxygen as a hypoxic condition and
20% for normoxic conditions. The oxygen level in cartilage is believed to be between 7-
10% at the surface and near 0.1% adjacent to the subchondral bone [21, 95].
Therefore, using 5% oxygen as a hypoxia model might not induce different responses
in SZP expression. In our study 1% oxygen level was used as a hypoxic environment.
We hypothesized that simulation of the physiological cartilage conditions would direct
in-vitro cultured chondrocytes to express zonal cartilage phenotypes. This hypothesis
was confirmed by the results which showed that normoxic conditions were required
for SZP expression while hypoxic conditions induced Col2 re-expression and
downregulated SZP expression. The expression of Col2 did not increase during the 4
days of alginate bead culture and partial restoration of the expression could only be
achieved under hypoxic conditions. This is an interesting observation which indicates
that 3D culture is not sufficient for re-expression of the cartilage phenotype. Although
the common consensus is that encapsulation in alginate allows recovery of Col2
expression [72, 161, 162], some studies have shown this is not always the case [61].
7.8 Cell morphology and the expression of SZP
To address the interdependence of cell morphology and expression of SZP we
performed pilot studies to induce cell morphologies from round to spread and
quantified SZP mRNA levels. Chondrocytes cultured on the various substrates induced
a range of different morphologies, from highly spread on the 2D substrates to less

Chapter 7
99
spread in the 3D samples. Culturing chondrocytes on TCP and Col1 2D resulted in a
spread morphology typical of dedifferentiated chondrocytes. Chondrocytes exhibited a
slightly spread morphology in the Col1 hydrogel (Col1 3D) and maintained a
completely round morphology in the alginate gel (Figure 7.7).
Figure 7. 7. |Morphology of chondrocytes cultured in 2D and 3D substrates. Phase contrast (top) and
confocal images of phalloidin-rhodamine stained (bottom) passage 2 Bovine chondrocytes cultured on
TCP, Col1 coated PDMS (Col1 2D), Col1 3D and 0.8% Alg. All images were taken 3 days after cell seeding
using a 40x objective.
Gene expression of SZP was higher in 3D than 2D cultures (Figure 7.8). Cells
within alginate maintained the highest expression indicating that the spread
morphology in 3D does not necessarily lead to SZP upregulation.

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
100
Figure 7. 8. | Effect of cell morphology and dimensionality on SZP gene expression. TCP was set as the
reference sample and RPL13 was used as a housekeeping gene.
Chondrocytes of the superficial zone have a more flat morphology compared to
those of deeper layers which suggest that these cells might show a better response to
culture conditions that induce a flat morphology such as 2D cultures. It has been
reported that passage 0 cells have a higher expression of SZP in 2D compared to 3D
which is in contradiction with our results [227]. The use of passage 0 cells in the
previous report, however, may explain their results as freshly isolated cells are
weakened by the enzymatic digestion process and may be less active when embedded
in a 3D matrix. We have also observed in preliminary studies that a spread
morphology in 3D within a collagen gel resulted in lower expression of SZP compared
to alginate which induces a completely round morphology (Figures 7.7 and 7.8).
However the different responses in alginate and collagen cultures may reflect distinct
material and biological properties rather the effect of spreading. To address this issue,
we also compared SZP expression of chondrocytes in RGD modified alginate compared

Chapter 7
101
to non-modified alginate. Cells were encapsulated in alginate and RGD modified
alginate (Novatach, Novamatrix) using the CaCO3-GDL method at 0.4% gel
concentration and casted in the 3D chambers. Mechanical stimulation was applied at
day 4 for 2 h. Cell morphology was only affected on the edge of the RGD modified gel
where cells were able to spread (Figure 7.9B). This is consistent with previous reports
that show RGD-modified alginate induces cell spreading only when alginate is made
degradable through conjugation of MMP sensitive motifs [228]. The presence of RGD
sequences did not induce any effect on SZP expression nor did it induce a stronger
response to mechanical stimulation. This indicates that the presence of such adhesion
sequences do not trigger any pathways that affect SZP (Figure 7.9A).
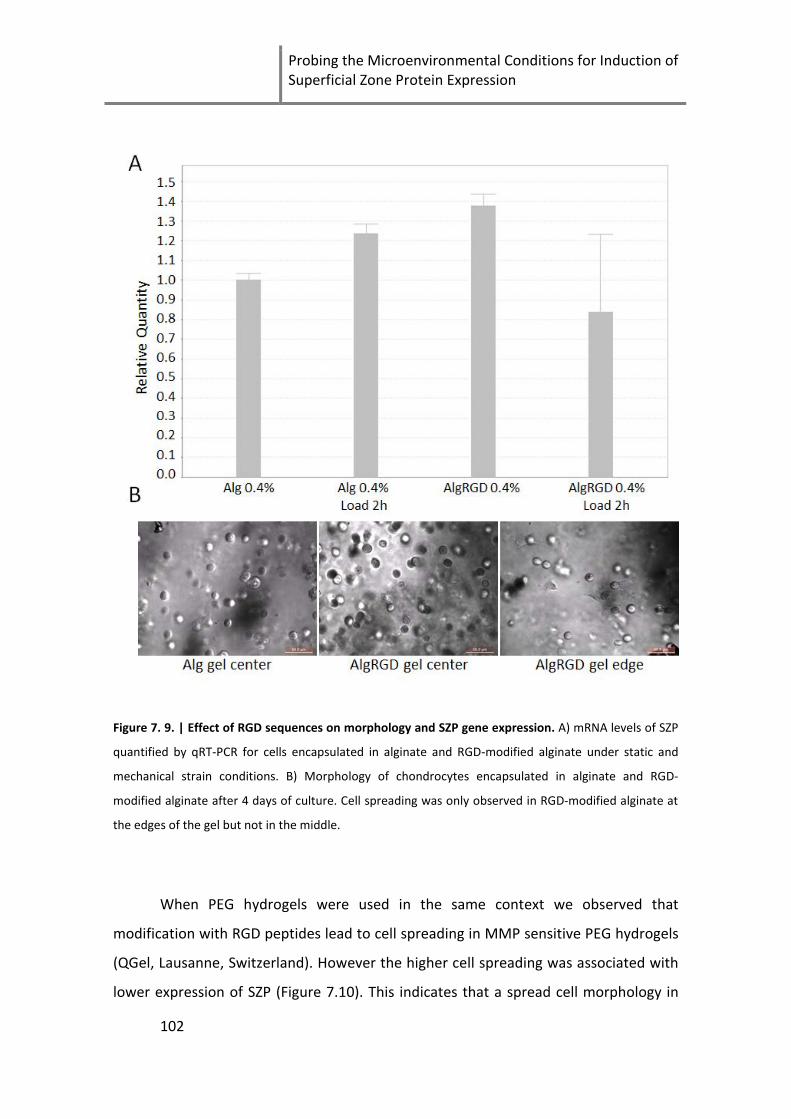
Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
102
Figure 7. 9. | Effect of RGD sequences on morphology and SZP gene expression. A) mRNA levels of SZP
quantified by qRT-PCR for cells encapsulated in alginate and RGD-modified alginate under static and
mechanical strain conditions. B) Morphology of chondrocytes encapsulated in alginate and RGD-
modified alginate after 4 days of culture. Cell spreading was only observed in RGD-modified alginate at
the edges of the gel but not in the middle.
When PEG hydrogels were used in the same context we observed that
modification with RGD peptides lead to cell spreading in MMP sensitive PEG hydrogels
(QGel, Lausanne, Switzerland). However the higher cell spreading was associated with
lower expression of SZP (Figure 7.10). This indicates that a spread cell morphology in

Chapter 7
103
3D does not alone cause an upregulation of SZP. A more in depth analysis of the
biological pathways would possibly shed light on the regulation of SZP and its relation
to cell morphology.
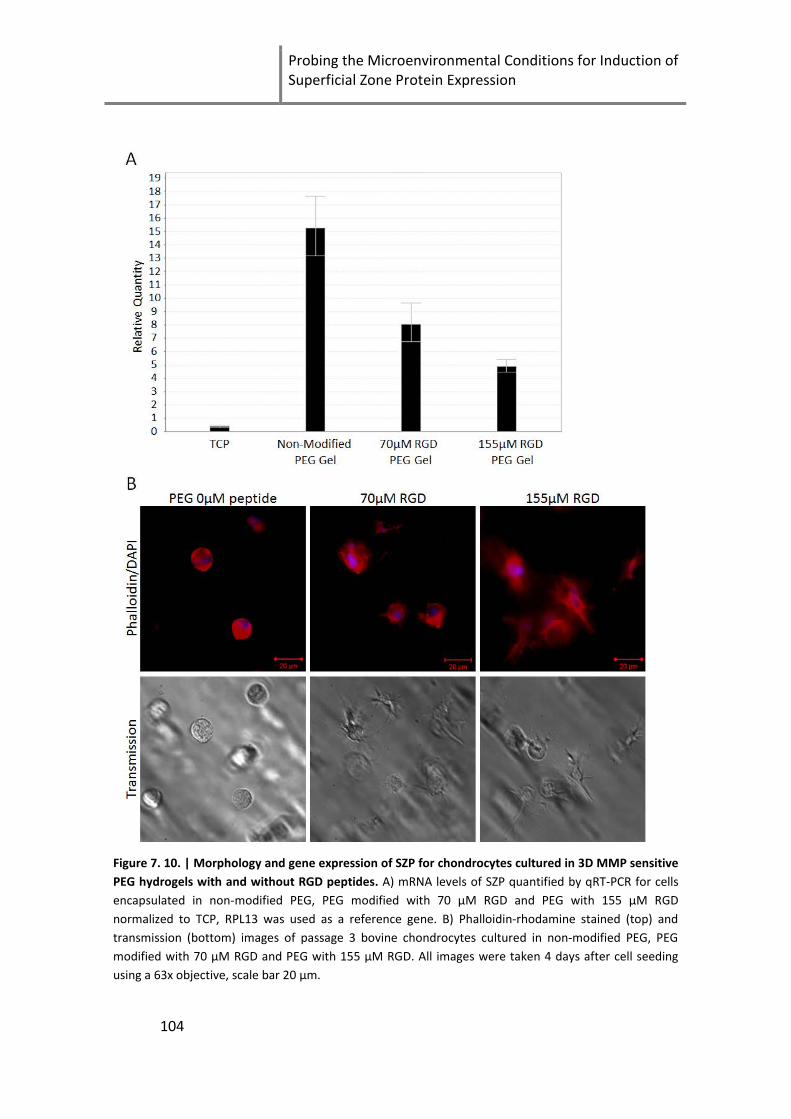
Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
104
Figure 7. 10. | Morphology and gene expression of SZP for chondrocytes cultured in 3D MMP sensitive PEG hydrogels with and without RGD peptides. A) mRNA levels of SZP quantified by qRT-PCR for cells encapsulated in non-modified PEG, PEG modified with 70 µM RGD and PEG with 155 µM RGD normalized to TCP, RPL13 was used as a reference gene. B) Phalloidin-rhodamine stained (top) and transmission (bottom) images of passage 3 bovine chondrocytes cultured in non-modified PEG, PEG modified with 70 µM RGD and PEG with 155 µM RGD. All images were taken 4 days after cell seeding using a 63x objective, scale bar 20 µm.

Chapter 7
105
7.9 Chapter summary
Cartilage lubrication is crucial for maintaining the function of articular cartilage
and preventing its degeneration. Understanding the mechanisms that regulate the
expression of SZP will allow the design of engineered tissue with optimal lubrication
properties. We have shown that SZP expression in dedifferentiated chondrocytes can
be upregulated to cartilage tissue levels when cultured in alginate at normoxic
conditions. Moreover, we designed a chamber compatible with the STREX device that
allows application of homogeneous strain in 3D to any hydrogel. The results of the
current study provide insights into the regulation of SZP and tools to control its
expression in-vitro.

Probing the Microenvironmental Conditions for Induction of Superficial Zone Protein Expression
106

Chapter 8
107
8 Conclusions and outlook
The aim of this thesis was to engineer 2D and 3D microenvironments to
improve the culturing conditions of chondrocytes and stem cells used for cartilage
tissue engineering applications such as ACI. Using the layer-by-layer technique 2D
substrates made of natural ECM macromolecules were constructed and characterized
on stretchable PDMS. The films made of type I collagen and the chondroprotective
molecule chondroitin sulfate were stable in media. However, chondrocytes seeded
those films still adopted a fibroblastic phenotype and mechanical stimulation did not
help recovering the cartilage phenotype. Furthermore, the CS containing films did not
have any effect on expression of MMPs. This might have been a consequence of using
juvenile chondrocytes as in our studies cells were obtained from 6-month old
chondrocytes. It may be that the beneficial effects of chondroitin sulfate are more
pronounced when using cells from older donors or osteoarthritic cells. The well-
characterized effects of chondroitin sulfate are on inflammatory responses and thus
future work should focus on the use of osteoarthritic cells to determine any potential
benefits of the chondroitin sulfate containing films.
The major part of this thesis focused on 3D microenvironments. 3D scaffolds
were designed to present biomimetic cartilage-like microenvironments to
encapsulated chondrocytes and stem cells. The use of the alginate sulfate material as a
hydrogel for cartilage tissue engineering was a major breakthrough in this thesis. We
showed that by simply modifying alginate with sulfate moieties, cell proliferation was
increased 5 fold and the hydrogel attained a cartilage-like appearance after only 5
weeks in culture. Most importantly, the material could preserve the cartilage
phenotype of encapsulated chondrocytes similar to unmodified alginate which is well
known to maintain the cartilage phenotype. Future work should focus on investigating
proliferation of non-passaged chondrocytes within alginate sulfate to see if de-
differentiation can be completely eliminated and to study the performance of this
novel material in animal models.

Conclusions and outlook
108
We have also seen interesting responses for stem cells encapsulated in the
GFGOER degradable hydrogels. Our studies were carried out for a maximum of 21 days
and we observed an opaque appearance of these constructs as a result of high
collagen deposition and cell proliferation. We believe that longer times are necessary
to fully establish chondrogenic differentiation of the stem cells. Therefore future work
should investigate longer culture periods within the GFOGER modified hydrogels.
Finally, we made a step towards building a stratified articular cartilage. We first
present superficial zone protein as a new marker for chondrogenesis, whose
expression drops during passaging and is fully recovered by day 4 encapsulation in
alginate under normoxic conditions. This expression was further augmented with
mechanical strain. On the other hand, type II collagen was only partially restored in
alginate beads under hypoxic conditions. We have also seen in data not presented in
this thesis that longer time periods are required to recover type II collagen expression
and potentially different mechanical stimuli such as compression or hydrostatic
pressure could be beneficial. Future work should help identify the parameters
important in constructing stratified cartilage.
In this thesis we worked on a variety of techniques to engineer 2D and 3D
microenvironments. We were successful in preparing natural ECM based films on
stretchable substrates that may be of relevance in a variety of biomedical applications.
We presented for the first time the alginate sulfate hydrogel as a material for cartilage
tissue engineering applications. We showed that the GFOGER adhesion peptide
presented in a degradable PEG hydrogel provides a better chondrogenic
microenvironment than the more commonly used RGD peptide. Finally, we
determined parameters that regulate expression of superficial zone protein as a step
towards building stratified articular cartilage. The results of this thesis are expected to
have a high impact on the tissue engineering field particularly for cartilage
applications.

Chapter 9
109
9 References
[1] Berthiaume F, Maguire TJ, Yarmush ML. Tissue Engineering and Regenerative Medicine: History, Progress, and Challenges. Annual Review of Chemical and Biomolecular Engineering. 2011;2:403-30.
[2] Langer R, Vacanti JP. Tissue Engineering. Science. 1993;260:920-6.
[3] Webster JP. Refrigerated Skin Grafts *. Annals of Surgery. 1944;120:431-49.
[4] Polge C, Smith A, Parkes A. Revival of spermatozoa after vitrification and dehydration at low temperatures. Nature. 1949;164:666.
[5] Billingham R, Medawar P. The freezing, drying and storage of mammalian skin. Journal of Experimental Biology. 1952;29:454-68.
[6] Rheinwatd JG, Green H. Seria cultivation of strains of human epidemal keratinocytes: the formation keratinizin colonies from single cell is. Cell. 1975;6:331-43.
[7] Green H, Kehinde O, Thomas J. Growth of cultured human epidermal cells into multiple epithelia suitable for grafting. Proceedings of the National Academy of Sciences. 1979;76:5665-8.
[8] Bell E, Ehrlich H, Buttle D, Nakatsuji T. Living tissue formed in vitro and accepted as skin-equivalent tissue of full thickness. Science. 1981;211:1052-4.
[9] Viola J, Lal B, Grad O. The emergence of tissue engineering as a research field. http://www.nsf.gov/pubs/2004/nsf0450/exec_summ.pdf; 2003.
[10] Eberli D, Filho LF, Atala A, Yoo JJ. Composite scaffolds for the engineering of hollow organs and tissues. Methods. 2009;47:109-15.
[11] Chen FH, Rousche KT, Tuan RS. Technology Insight: adult stem cells in cartilage regeneration and tissue engineering. Nat Clin Pract Rheum. 2006;2:373-82.
[12] Becerra J, Andrades JA, Guerado E, Zamora-Navas P, López-Puertas JM, Reddi AH. Articular cartilage: structure and regeneration. Tissue Engineering Part B: Reviews. 2010;16:617-27.
[13] Buckwalter JA, Mankin HJ, Grodzinsky AJ. Articular cartilage and osteoarthritis. INSTRUCTIONAL COURSE LECTURES-AMERICAN ACADEMY OF ORTHOPAEDIC SURGEONS. 2005;54:465.
[14] Chiang H, Jiang CC. Repair of articular cartilage defects: review and perspectives. Journal of the Formosan Medical Association. 2009;108:87-101.
[15] Guilak F, Ratcliffe A, Lane N, Rosenwasser MP, Mow VC. Mechanical and biochemical changes in the superficial zone of articular cartilage in canine experimental osteoarthritis. Journal of Orthopaedic Research. 1994;12:474-84.
[16] Jay GD, Tantravahi U, Britt DE, Barrach HJ, Cha C-J. Homology of lubricin and superficial zone protein (SZP): Products of megakaryocyte stimulating factor (MSF) gene expression by human synovial fibroblasts and articular chondrocytes localized to chromosome 1q25. Journal of Orthopaedic Research. 2001;19:677-87.

References
110
[17] Schumacher BL, Block JA, Schmid TM, Aydelotte MB, Kuettner KE. A Novel Proteoglycan Synthesized and Secreted by Chondrocytes of the Superficial Zone of Articular Cartilage. Archives of Biochemistry and Biophysics. 1994;311:144-52.
[18] Li L, Xie T. Stem cell niche: structure and function. Annu Rev Cell Dev Biol. 2005;21:605-31.
[19] Gigout A, Jolicoeur M, Nelea M, Raynal N, Farndale R, Buschmann MD. Chondrocyte Aggregation in Suspension Culture Is GFOGER-GPP- and β1 Integrin-dependent. Journal of Biological Chemistry. 2008;283:31522-30.
[20] Rolls A, Shechter R, London A, Segev Y, Jacob-Hirsch J, Amariglio N, et al. Two faces of chondroitin sulfate proteoglycan in spinal cord repair: a role in microglia/macrophage activation. PLoS medicine. 2008;5:e171.
[21] Grimshaw MJ, Mason RM. Modulation of bovine articular chondrocyte gene expression in vitro by oxygen tension. Osteoarthritis and Cartilage. 2001;9:357-64.
[22] Wong M, Carter DR. Articular cartilage functional histomorphology and mechanobiology: a research perspective. Bone. 2003;33:1-13.
[23] DeLise AM, Fischer L, Tuan RS. Cellular interactions and signaling in cartilage development. Osteoarthritis and Cartilage. 2000;8:309-34.
[24] Woo SLY, Buckwalter JA. Injury and repair of the musculoskeletal soft tissues: workshop, Savannah, Georgia, June 1987: American Academy of Orthopaedic Surgeons; 1988.
[25] Herzog W, Longino D. The role of muscles in joint degeneration and osteoarthritis. Journal of Biomechanics. 2007;40, Supplement 1:S54-S63.
[26] Schulz R, Bader A. Cartilage tissue engineering and bioreactor systems for the cultivation and stimulation of chondrocytes. Eur Biophys J. 2007;36:539-68.
[27] Kubo M, Ando K, Mimura T, Matsusue Y, Mori K. Chondroitin sulfate for the treatment of hip and knee osteoarthritis: Current status and future trends. Life Sciences. 2009;85:477-83.
[28] Thelin N, Holmberg S, Thelin A. Knee injuries account for the sports-related increased risk of knee osteoarthritis. Scandinavian Journal of Medicine & Science in Sports. 2006;16:329-33.
[29] Ochi M, Uchio Y, Kawasaki K, Wakitani S, Iwasa J. Transplantation of cartilage-like tissue made by tissue engineering in the treatment of cartilage defects of the knee. Journal of Bone & Joint Surgery, British Volume. 2002;84-B:571-8.
[30] Lequesne M, Brandt K, Bellamy N, Moskowitz R, Menkes CJ, Pelletier JP, et al. Guidelines for testing slow acting drugs in osteoarthritis. The Journal of rheumatology Supplement. 1994;41:65-71; discussion 2-3.
[31] Zhang W, Moskowitz RW, Nuki G, Abramson S, Altman RD, Arden N, et al. OARSI recommendations for the management of hip and knee osteoarthritis, Part I: Critical appraisal of existing treatment guidelines and systematic review of current research evidence. Osteoarthritis and Cartilage. 2007;15:981-1000.
[32] Zhang W, Moskowitz RW, Nuki G, Abramson S, Altman RD, Arden N, et al. OARSI recommendations for the management of hip and knee osteoarthritis, Part II: OARSI evidence-based, expert consensus guidelines. Osteoarthritis and Cartilage. 2008;16:137-62.
[33] Stürmer T, Erb A, Keller F, Günther K-P, Brenner H. Determinants of impaired renal function with use of nonsteroidal anti-inflammatory drugs: the importance of half-life and other medications. The American Journal of Medicine. 2001;111:521-7.

Chapter 9
111
[34] Winkelmayer WC, Waikar SS, Mogun H, Solomon DH. Nonselective and Cyclooxygenase-2-Selective NSAIDs and Acute Kidney Injury. The American Journal of Medicine. 2008;121:1092-8.
[35] Laine L. Nonsteroidal anti-inflammatory drug gastropathy. Gastrointestinal endoscopy clinics of North America. 1996;6:489-504.
[36] Lanza FL, Chan FKL, Quigley EMM. Guidelines for Prevention of NSAID-Related Ulcer Complications. Am J Gastroenterol. 2009;104:728-38.
[37] Wu C-H, Ko C-S, Huang J-W, Huang H-J, Chu IM. Effects of exogenous glycosaminoglycans on human chondrocytes cultivated on type II collagen scaffolds. J Mater Sci: Mater Med. 2010;21:725-9.
[38] Sharma G, Saxena RK, Mishra P. Synergistic effect of chondroitin sulfate and cyclic pressure on biochemical and morphological properties of chondrocytes from articular cartilage. Osteoarthritis and Cartilage. 2008;16:1387-94.
[39] Michel BA, Stucki G, Frey D, De Vathaire F, Vignon E, Bruehlmann P, et al. Chondroitins 4 and 6 sulfate in osteoarthritis of the knee: A randomized, controlled trial. Arthritis & Rheumatism. 2005;52:779-86.
[40] Uebelhart D, Thonar EJMA, Zhang J, Williams JM. Protective effect of exogenous chondroitin 4,6-sulfate in the acute degradation of articular cartilage in the rabbit. Osteoarthritis and Cartilage. 1998;6, Supplement A:6-13.
[41] Jomphe C, Gabriac M, Hale TM, Héroux L, Trudeau L-É, Deblois D, et al. Chondroitin Sulfate Inhibits the Nuclear Translocation of Nuclear Factor-κB in Interleukin-1β-Stimulated Chondrocytes. Basic & Clinical Pharmacology & Toxicology. 2008;102:59-65.
[42] Legendre F, Baugé C, Roche R, Saurel AS, Pujol JP. Chondroitin sulfate modulation of matrix and inflammatory gene expression in IL-1beta-stimulated chondrocytes--study in hypoxic alginate bead cultures. Osteoarthritis and cartilage / OARS, Osteoarthritis Research Society. 2008;16:105-14.
[43] Bassleer C, Henrotin Y, Franchimont P. In-vitro evaluation of drugs proposed as chondroprotective agents. International journal of tissue reactions. 1992;14:231-41.
[44] Johnson KA, Hulse DA, Hart RC, Kochevar D, Chu Q. Effects of an orally administered mixture of chondroitin sulfate, glucosamine hydrochloride and manganese ascorbate on synovial fluid chondroitin sulfate 3B3 and 7D4 epitope in a canine cruciate ligament transection model of osteoarthritis. Osteoarthritis and Cartilage. 2001;9:14-21.
[45] Lippiello L, Woodward J, Karpman R, Hammad TA. In Vivo Chondroprotection and Metabolic Synergy of Glucosamine and Chondroitin Sulfate. Clinical Orthopaedics and Related Research. 2000;381:229-40.
[46] Ronca F, Palmieri L, Panicucci P, Ronca G. Anti-inflammatory activity of chondroitin sulfate. Osteoarthritis and Cartilage. 1998;6, Supplement A:14-21.
[47] Bourgeois P, Chales G, Dehais J, Delcambre B, Kuntz J-L, Rozenberg S. Efficacy and tolerability of chondroitin sulfate 1200 mg/day vs chondroitin sulfate 3×400 mg/day vs placebo. Osteoarthritis and Cartilage. 1998;6, Supplement A:25-30.

References
112
[48] Bucsi L, Poór G. Efficacy and tolerability of oral chondroitin sulfate as a symptomatic slow-acting drug for osteoarthritis (SYSADOA) in the treatment of knee osteoarthritis. Osteoarthritis and Cartilage. 1998;6, Supplement A:31-6.
[49] Strauss EJ, Fonseca LE, Shah MR, Yorum T. Management of focal cartilage defects in the knee-Is ACI the answer? Bulletin of the NYU hospital for joint diseases. 2011;69:63.
[50] Bhosale AM, Richardson JB. Articular cartilage: structure, injuries and review of management. British Medical Bulletin. 2008;87:77-95.
[51] Grande DA, Pitman MI, Peterson L, Menche D, Klein M. The repair of experimentally produced defects in rabbit articular cartilage by autologous chondrocyte transplantation. Journal of Orthopaedic Research. 1989;7:208-18.
[52] Brittberg M, Lindahl A, Nilsson A, Ohlsson C, Isaksson O, Peterson L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. New england journal of medicine. 1994;331:889-95.
[53] Brittberg M, Peterson L, Sjouml, gren-Jansson E, Tallheden T, Lindahl A. Articular Cartilage Engineering with Autologous Chondrocyte Transplantation A Review of Recent Developments. The Journal of Bone & Joint Surgery. 2003;85:109-15.
[54] Henderson I, Gui J, Lavigne P. Autologous Chondrocyte Implantation: Natural History of Postimplantation Periosteal Hypertrophy and Effects of Repair-Site Debridement on Outcome. Arthroscopy: The Journal of Arthroscopic & Related Surgery. 2006;22:1318-24.e1.
[55] Rosenberger RE, Gomoll AH, Bryant T, Minas T. Repair of Large Chondral Defects of the Knee With Autologous Chondrocyte Implantation in Patients 45 Years or Older. The American Journal of Sports Medicine. 2008;36:2336-44.
[56] Haddo O, Mahroof S, Higgs D, David L, Pringle J, Bayliss M, et al. The use of chondrogide membrane in autologous chondrocyte implantation. The Knee. 2004;11:51-5.
[57] Behrens P, Bitter T, Kurz B, Russlies M. Matrix-associated autologous chondrocyte transplantation/implantation (MACT/MACI)—5-year follow-up. The Knee. 2006;13:194-202.
[58] Behrens P, Ehlers EM, Köchermann KU, Rohwedel J, Russlies M, Plötz W. New therapy procedure for localized cartilage defects. Encouraging results with autologous chondrocyte implantation. MMW Fortschritte der Medizin. 1999;141:49-51.
[59] Schiltz JR, Mayne R, Holtzer H. The Synthesis of Collagen and Glycosaminoglycans by Dedifferentiated Chondroblasts in Culture. Differentiation. 1973;1:97-108.
[60] Benya PD, Shaffer JD. Dedifferentiated chondrocytes reexpress the differentiated collagen phenotype when cultured in agarose gels. Cell. 1982;30:215-24.
[61] Darling EM, Athanasiou KA. Rapid phenotypic changes in passaged articular chondrocyte subpopulations. Journal of Orthopaedic Research. 2005;23:425-32.
[62] Schnabel M, Marlovits S, Eckhoff G, Fichtel I, Gotzen L, Vécsei V, et al. Dedifferentiation-associated changes in morphology and gene expression in primary human articular chondrocytes in cell culture. Osteoarthritis and Cartilage. 2002;10:62-70.
[63] Darling E, Pritchett P, Evans B, Superfine R, Zauscher S, Guilak F. Mechanical Properties and Gene Expression of Chondrocytes on Micropatterned Substrates Following Dedifferentiation in Monolayer. Cel Mol Bioeng. 2009;2:395-404.

Chapter 9
113
[64] Domm C, Schünke M, Christesen K, Kurz B. Redifferentiation of dedifferentiated bovine articular chondrocytes in alginate culture under low oxygen tension. Osteoarthritis and Cartilage. 2002;10:13-22.
[65] Candrian C, Vonwil D, Barbero A, Bonacina E, Miot S, Farhadi J, et al. Engineered cartilage generated by nasal chondrocytes is responsive to physical forces resembling joint loading. Arthritis & Rheumatism. 2008;58:197-208.
[66] Candrian C, Miot S, Wolf F, Bonacina E, Dickinson S, Wirz D, et al. Are ankle chondrocytes from damaged fragments a suitable cell source for cartilage repair? Osteoarthritis and Cartilage. 2010;18:1067-76.
[67] Mackay AM, Beck SC, Murphy JM, Barry FP, Chichester CO, Pittenger MF. Chondrogenic differentiation of cultured human mesenchymal stem cells from marrow. Tissue engineering. 1998;4:415-28.
[68] Awad HA, Halvorsen YD, Gimble JM, Guilak F. Effects of transforming growth factor beta1 and dexamethasone on the growth and chondrogenic differentiation of adipose-derived stromal cells. Tissue engineering. 2003;9:1301-12.
[69] Tew SR, Murdoch AD, Rauchenberg RP, Hardingham TE. Cellular methods in cartilage research: Primary human chondrocytes in culture and chondrogenesis in human bone marrow stem cells. Methods. 2008;45:2-9.
[70] Kumar D, Lassar AB. The transcriptional activity of Sox9 in chondrocytes is regulated by RhoA signaling and actin polymerization. Molecular and Cellular Biology. 2009;29:4262-73.
[71] Woods A, Wang G, Beier F. RhoA/ROCK signaling regulates Sox9 expression and actin organization during chondrogenesis. The Journal of biological chemistry. 2005;280:11626-34.
[72] Bonaventure J, Kadhom N, Cohen-Solal L, Ng KH, Bourguignon J, Lasselin C, et al. Reexpression of Cartilage-Specific Genes by Dedifferentiated Human Articular Chondrocytes Cultured in Alginate Beads. Experimental Cell Research. 1994;212:97-104.
[73] Park Y, Lutolf MP, Hubbell JA, Hunziker EB, Wong M. Bovine primary chondrocyte culture in synthetic matrix metalloproteinase-sensitive poly(ethylene glycol)-based hydrogels as a scaffold for cartilage repair. Tissue engineering. 2004;10:515-22.
[74] Lutolf MP, Hubbell JA. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat Biotech. 2005;23:47-55.
[75] Kraehenbuehl TP, Zammaretti P, Van der Vlies AJ, Schoenmakers RG, Lutolf MP, Jaconi ME, et al. Three-dimensional extracellular matrix-directed cardioprogenitor differentiation: Systematic modulation of a synthetic cell-responsive PEG-hydrogel. Biomaterials. 2008;29:2757-66.
[76] Lee ST, Yun JI, Jo YS, Mochizuki M, van der Vlies AJ, Kontos S, et al. Engineering integrin signaling for promoting embryonic stem cell self-renewal in a precisely defined niche. Biomaterials. 2010;31:1219-26.
[77] Yan WQ, Nakashima K, Iwamoto M, Kato Y. Stimulation by concanavalin A of cartilage-matrix proteoglycan synthesis in chondrocyte cultures. Journal of Biological Chemistry. 1990;265:10125-31.
[78] Petersen EF, Spencer RGS, McFarland EW. Microengineering neocartilage scaffolds. Biotechnology and Bioengineering. 2002;78:802-5.

References
114
[79] Boudou T, Crouzier T, Ren K, Blin G, Picart C. Multiple Functionalities of Polyelectrolyte Multilayer Films: New Biomedical Applications. Advanced Materials. 2010;22:441-67.
[80] Ai H, Jones S, Lvov Y. Biomedical applications of electrostatic layer-by-layer nano-assembly of polymers, enzymes, and nanoparticles. Cell Biochem Biophys. 2003;39:23-43.
[81] Decher G. Fuzzy Nanoassemblies: Toward Layered Polymeric Multicomposites. Science. 1997;277:1232-7.
[82] Zhang J, Senger B, Vautier D, Picart C, Schaaf P, Voegel J-C, et al. Natural polyelectrolyte films based on layer-by layer deposition of collagen and hyaluronic acid. Biomaterials. 2005;26:3353-61.
[83] Semenov OV, Malek A, Bittermann AG, Vörös J, Zisch AH. Engineered polyelectrolyte multilayer substrates for adhesion, proliferation, and differentiation of human mesenchymal stem cells. Tissue Engineering Part A. 2009;15:2977-90.
[84] Richert L, Boulmedais F, Lavalle P, Mutterer J, Ferreux E, Decher G, et al. Improvement of Stability and Cell Adhesion Properties of Polyelectrolyte Multilayer Films by Chemical Cross-Linking. Biomacromolecules. 2003;5:284-94.
[85] Thompson MT, Berg MC, Tobias IS, Rubner MF, Van Vliet KJ. Tuning compliance of nanoscale polyelectrolyte multilayers to modulate cell adhesion. Biomaterials. 2005;26:6836-45.
[86] Ai H, Lvov Y, Mills D, Jennings M, Alexander J, Jones S. Coating and selective deposition of nanofilm on silicone rubber for cell adhesion and growth. Cell Biochem Biophys. 2003;38:103-14.
[87] Chanana M, Gliozzi A, Diaspro A, Chodnevskaja I, Huewel S, Moskalenko V, et al. Interaction of Polyelectrolytes and Their Composites with Living Cells. Nano Letters. 2005;5:2605-12.
[88] Johansson JÅ, Halthur T, Herranen M, Söderberg L, Elofsson U, Hilborn J. Build-up of Collagen and Hyaluronic Acid Polyelectrolyte Multilayers. Biomacromolecules. 2005;6:1353-9.
[89] Hahn SK, Hoffman AS. Preparation and characterization of biocompatible polyelectrolyte complex multilayer of hyaluronic acid and poly-l-lysine. International Journal of Biological Macromolecules. 2005;37:227-31.
[90] Jiang F, Hörber H, Howard J, Müller DJ. Assembly of collagen into microribbons: effects of pH and electrolytes. Journal of Structural Biology. 2004;148:268-78.
[91] Volpi N, Cusmano M, Venturelli T. Qualitative and quantitative studies of heparin and chondroitin sulfates in normal human plasma. Biochimica et Biophysica Acta (BBA) - General Subjects. 1995;1243:49-58.
[92] Vincent JFV, Bogatyreva OA, Bogatyrev NR, Bowyer A, Pahl A-K. Biomimetics: its practice and theory. Journal of The Royal Society Interface. 2006;3:471-82.
[93] Giannoni P, Siegrist M, Hunziker EB, Wong M. The mechanosensitivity of cartilage oligomeric matrix protein (COMP). Biorheology. 2003;40:101-9.
[94] Wang JHC, Thampatty BP. Chapter 7 Mechanobiology of Adult and Stem Cells. In: Kwang WJ, editor. International Review of Cell and Molecular Biology: Academic Press; 2008. p. 301-46.

Chapter 9
115
[95] Schrobback K, Malda J, Crawford RW, Upton Z, Leavesley DI, Klein TJ. Effects of oxygen on zonal marker expression in human articular chondrocytes. Tissue Engineering Part A. 2012;18:920-33.
[96] Connelly JT, García AJ, Levenston ME. Inhibition of in vitro chondrogenesis in RGD-modified three-dimensional alginate gels. Biomaterials. 2007;28:1071-83.
[97] Connelly JT, García AJ, Levenston ME. Interactions between integrin ligand density and cytoskeletal integrity regulate BMSC chondrogenesis. Journal of Cellular Physiology. 2008;217:145-54.
[98] Knight CG, Morton LF, Peachey AR, Tuckwell DS, Farndale RW, Barnes MJ. The Collagen-binding A-domains of Integrins α1β1 and α2β1Recognize the Same Specific Amino Acid Sequence, GFOGER, in Native (Triple-helical) Collagens. Journal of Biological Chemistry. 2000;275:35-40.
[99] Millward-Sadler SJ, Wright MO, Davies LW, Nuki G, Salter DM. Mechanotransduction via integrins and interleukin-4 results in altered aggrecan and matrix metalloproteinase 3 gene expression in normal, but not osteoarthritic, human articular chondrocytes. Arthritis & Rheumatism. 2000;43:2091-9.
[100] Sah RLY, Kim Y-J, Doong J-YH, Grodzinsky AJ, Plass AHK, Sandy JD. Biosynthetic response of cartilage explants to dynamic compression. Journal of Orthopaedic Research. 1989;7:619-36.
[101] Lee JH, Fitzgerald JB, DiMicco MA, Grodzinsky AJ. Mechanical injury of cartilage explants causes specific time-dependent changes in chondrocyte gene expression. Arthritis & Rheumatism. 2005;52:2386-95.
[102] Scherberich A, Galli R, Jaquiery C, Farhadi J, Martin I. Three-Dimensional Perfusion Culture of Human Adipose Tissue-Derived Endothelial and Osteoblastic Progenitors Generates Osteogenic Constructs with Intrinsic Vascularization Capacity. STEM CELLS. 2007;25:1823-9.
[103] Altman GH, Horan RL, Martin I, Farhadi J, Stark PR, Volloch V, et al. Cell differentiation by mechanical stress. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2002;16:270-2.
[104] Juncosa-Melvin N, Matlin KS, Holdcraft RW, Nirmalanandhan VS, Butler DL. Mechanical stimulation increases collagen type I and collagen type III gene expression of stem cell-collagen sponge constructs for patellar tendon repair. Tissue engineering. 2007;13:1219-26.
[105] Chen Y-J, Huang C-H, Lee I-C, Lee Y-T, Chen M-H, Young T-H. Effects of Cyclic Mechanical Stretching on the mRNA Expression of Tendon/Ligament-Related and Osteoblast-Specific Genes in Human Mesenchymal Stem Cells. Connective Tissue Research. 2008;49:7-14.
[106] Davisson T, Kunig S, Chen A, Sah R, Ratcliffe A. Static and dynamic compression modulate matrix metabolism in tissue engineered cartilage. Journal of Orthopaedic Research. 2002;20:842-8.
[107] Wang H, Riha GM, Yan S, Li M, Chai H, Yang H, et al. Shear Stress Induces Endothelial Differentiation From a Murine Embryonic Mesenchymal Progenitor Cell Line. Arteriosclerosis, Thrombosis, and Vascular Biology. 2005;25:1817-23.
[108] Hirano Y, Ishiguro N, Sokabe M, Takigawa M, Naruse K. Effects of tensile and compressive strains on response of a chondrocytic cell line embedded in type I collagen gel. Journal of Biotechnology. 2008;133:245-52.

References
116
[109] Sumanasinghe RD, Bernacki SH, Loboa EG. Osteogenic differentiation of human mesenchymal stem cells in collagen matrices: effect of uniaxial cyclic tensile strain on bone morphogenetic protein (BMP-2) mRNA expression. Tissue engineering. 2006;12:3459-65.
[110] Illi B, Scopece A, Nanni S, Farsetti A, Morgante L, Biglioli P, et al. Epigenetic Histone Modification and Cardiovascular Lineage Programming in Mouse Embryonic Stem Cells Exposed to Laminar Shear Stress. Circulation Research. 2005;96:501-8.
[111] Angele P, Yoo JU, Smith C, Mansour J, Jepsen KJ, Nerlich M, et al. Cyclic hydrostatic pressure enhances the chondrogenic phenotype of human mesenchymal progenitor cells differentiated in vitro. Journal of Orthopaedic Research. 2003;21:451-7.
[112] Angele P, Schumann D, Angele M, Kinner B, Englert C, Hente R, et al. Cyclic, mechanical compression enhances chondrogenesis of mesenchymal progenitor cells in tissue engineering scaffolds. Biorheology. 2004;41:335-46.
[113] Wagner D, Lindsey D, Li K, Tummala P, Chandran S, Smith RL, et al. Hydrostatic Pressure Enhances Chondrogenic Differentiation of Human Bone Marrow Stromal Cells in Osteochondrogenic Medium. Annals of Biomedical Engineering. 2008;36:813-20.
[114] Toyoda T, Seedhom BB, Yao JQ, Kirkham J, Brookes S, Bonass WA. Hydrostatic pressure modulates proteoglycan metabolism in chondrocytes seeded in agarose. Arthritis & Rheumatism. 2003;48:2865-72.
[115] Smith RL, Rusk SF, Ellison BE, Wessells P, Tsuchiya K, Carter DR, et al. In vitro stimulation of articular chondrocyte mRNA and extracellular matrix synthesis by hydrostatic pressure. Journal of Orthopaedic Research. 1996;14:53-60.
[116] Grad S, Lee CR, Gorna K, Gogolewski S, Wimmer MA, Alini M. Surface motion upregulates superficial zone protein and hyaluronan production in chondrocyte-seeded three-dimensional scaffolds. Tissue engineering. 2005;11:249-56.
[117] Kamiya T, Tanimoto K, Tanne Y, Lin YY, Kunimatsu R, Yoshioka M, et al. Effects of mechanical stimuli on the synthesis of superficial zone protein in chondrocytes. Journal of Biomedical Materials Research Part A. 2010;92A:801-5.
[118] Nugent GE, Aneloski NM, Schmidt TA, Schumacher BL, Voegtline MS, Sah RL. Dynamic shear stimulation of bovine cartilage biosynthesis of proteoglycan 4. Arthritis & Rheumatism. 2006;54:1888-96.
[119] Ramage L, Nuki G, Salter DM. Signalling cascades in mechanotransduction: cell–matrix interactions and mechanical loading. Scandinavian Journal of Medicine & Science in Sports. 2009;19:457-69.
[120] Iqbal J, Zaidi M. Molecular regulation of mechanotransduction. Biochemical and Biophysical Research Communications. 2005;328:751-5.
[121] Lee S, Vörös J. An aqueous-based surface modification of poly (dimethylsiloxane) with poly (ethylene glycol) to prevent biofouling. Langmuir. 2005;21:11957-62.
[122] Studer D, Lischer S, Jochum W, Ehrbar M, Zenobi-Wong M, Maniura-Weber K. Ribosomal Protein L13a as a Reference Gene for Human Bone Marrow-Derived Mesenchymal Stromal Cells During Expansion, Adipo-, Chondro-, and Osteogenesis. Tissue Engineering Part C: Methods. 2012;18:761-72.

Chapter 9
117
[123] Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods. 2001;25:402-8.
[124] Mhanna RF, Vörös J, Zenobi-Wong M. Layer-by-layer films made from extracellular matrix macromolecules on silicone substrates. Biomacromolecules. 2011;12:609-16.
[125] Tsutsumi S, Shimazu A, Miyazaki K, Pan H, Koike C, Yoshida E, et al. Retention of Multilineage Differentiation Potential of Mesenchymal Cells during Proliferation in Response to FGF. Biochemical and Biophysical Research Communications. 2001;288:413-9.
[126] Kuo CK, Ma PX. Ionically crosslinked alginate hydrogels as scaffolds for tissue engineering: Part 1. Structure, gelation rate and mechanical properties. Biomaterials. 2001;22:511-21.
[127] Ingar Draget K, Østgaard K, Smidsrød O. Homogeneous alginate gels: A technical approach. Carbohydrate Polymers. 1990;14:159-78.
[128] McAloney RA, Sinyor M, Dudnik V, Goh MC. Atomic Force Microscopy Studies of Salt Effects on Polyelectrolyte Multilayer Film Morphology. Langmuir. 2001;17:6655-63.
[129] Estes BT, Diekman BO, Gimble JM, Guilak F. Isolation of adipose-derived stem cells and their induction to a chondrogenic phenotype. Nat Protocols. 2010;5:1294-311.
[130] Al-Soud WA, Ouis I-S, Li D-Q, Ljungh Å, Wadström T. Characterization of the PCR inhibitory effect of bile to optimize real-time PCR detection of Helicobacter species. FEMS Immunology & Medical Microbiology. 2005;44:177-82.
[131] Hübsch E, Ball V, Senger B, Decher G, Voegel J-C, Schaaf P. Controlling the Growth Regime of Polyelectrolyte Multilayer Films: Changing from Exponential to Linear Growth by Adjusting the Composition of Polyelectrolyte Mixtures. Langmuir. 2004;20:1980-5.
[132] Liu Y, He T, Song H, Gao C. Layer-by-layer assembly of biomacromolecules on poly(ethylene terephthalate) films and fiber fabrics to promote endothelial cell growth. Journal of Biomedical Materials Research Part A. 2007;81A:692-704.
[133] Liu Y, He T, Gao C. Surface modification of poly(ethylene terephthalate) via hydrolysis and layer-by-layer assembly of chitosan and chondroitin sulfate to construct cytocompatible layer for human endothelial cells. Colloids and Surfaces B: Biointerfaces. 2005;46:117-26.
[134] Lin Y, Wang L, Zhang P, Wang X, Chen X, Jing X, et al. Surface modification of poly(l-lactic acid) to improve its cytocompatibility via assembly of polyelectrolytes and gelatin. Acta Biomaterialia. 2006;2:155-64.
[135] Boulmedais F, Frisch B, Etienne O, Lavalle P, Picart C, Ogier J, et al. Polyelectrolyte multilayer films with pegylated polypeptides as a new type of anti-microbial protection for biomaterials. Biomaterials. 2004;25:2003-11.
[136] Luescher M, Rüegg M, Schindler P. Effect of hydration upon the thermal stability of tropocollagen and its dependence on the presence of neutral salts. Biopolymers. 1974;13:2489-503.
[137] Staprans I, Felts JM. Isolation and characterization of glycosaminoglycans in human plasma. The Journal of clinical investigation. 1985;76:1984-91.
[138] Wakitani S, Goto T, Young RG, Mansour JM, Goldberg VM, Caplan AI. Repair of large full-thickness articular cartilage defects with allograft articular chondrocytes embedded in a collagen gel. Tissue engineering. 1998;4:429-44.

References
118
[139] Varghese S, Hwang NS, Canver AC, Theprungsirikul P, Lin DW, Elisseeff J. Chondroitin sulfate based niches for chondrogenic differentiation of mesenchymal stem cells. Matrix Biology. 2008;27:12-21.
[140] Kim HK, Shim WS, Kim SE, Lee K-H, Kang E, Kim J-H, et al. Injectable In Situ–Forming pH/Thermo-Sensitive Hydrogel for Bone Tissue Engineering Tissue Engineering Part A. 2009;15:923-33.
[141] Sia SK, Whitesides GM. Microfluidic devices fabricated in Poly(dimethylsiloxane) for biological studies. ELECTROPHORESIS. 2003;24:3563-76.
[142] Bridges AJ, Conley C, Wang G, Burns DE, Vasey FB. A Clinical and Immunologic Evaluation of Women with Silicone Breast Implants and Symptoms of Rheumatic Disease. Annals of Internal Medicine. 1993;118:929-36.
[143] Okada T, Ikada Y. Modification of silicone surface by graft polymerization of acrylamide with corona discharge. Die Makromolekulare Chemie. 1991;192:1705-13.
[144] Abbasi F, Mirzadeh H, Katbab A-A. Modification of polysiloxane polymers for biomedical applications: a review. Polymer International. 2001;50:1279-87.
[145] Amma H, Naruse K, Ishiguro N, Sokabe M. Involvement of reactive oxygen species in cyclic stretch-induced NF-κB activation in human fibroblast cells. British Journal of Pharmacology. 2005;145:364-73.
[146] Lawton RA, Price CR, Runge AF, Doherty Iii WJ, Saavedra SS. Air plasma treatment of submicron thick PDMS polymer films: effect of oxidation time and storage conditions. Colloids and Surfaces A: Physicochemical and Engineering Aspects. 2005;253:213-5.
[147] Morra M, Occhiello E, Marola R, Garbassi F, Humphrey P, Johnson D. On the aging of oxygen plasma-treated polydimethylsiloxane surfaces. Journal of Colloid and Interface Science. 1990;137:11-24.
[148] Bodas D, Khan-Malek C. Hydrophilization and hydrophobic recovery of PDMS by oxygen plasma and chemical treatment—An SEM investigation. Sensors and Actuators B: Chemical. 2007;123:368-73.
[149] Richert L, Lavalle P, Payan E, Shu XZ, Prestwich GD, Stoltz J-F, et al. Layer by Layer Buildup of Polysaccharide Films: Physical Chemistry and Cellular Adhesion Aspects. Langmuir. 2003;20:448-58.
[150] Kujawa P, Moraille P, Sanchez J, Badia A, Winnik FM. Effect of Molecular Weight on the Exponential Growth and Morphology of Hyaluronan/Chitosan Multilayers: A Surface Plasmon Resonance Spectroscopy and Atomic Force Microscopy Investigation. Journal of the American Chemical Society. 2005;127:9224-34.
[151] Rossetti FF, Reviakine I, Csúcs G, Assi F, Vörös J, Textor M. Interaction of Poly(L-Lysine)-g-Poly(Ethylene Glycol) with Supported Phospholipid Bilayers. Biophysical Journal. 2004;87:1711-21.
[152] Granéli A, Edvardsson M, Höök F. DNA-Based Formation of a Supported, Three-Dimensional Lipid Vesicle Matrix Probed by QCM-D and SPR. ChemPhysChem. 2004;5:729-33.
[153] Wallace DG. The relative contribution of electrostatic interactions to stabilization of collagen fibrils. Biopolymers. 1990;29:1015-26.

Chapter 9
119
[154] Jokinen J, Dadu E, Nykvist P, Käpylä J, White DJ, Ivaska J, et al. Integrin-mediated Cell Adhesion to Type I Collagen Fibrils. Journal of Biological Chemistry. 2004;279:31956-63.
[155] Humphries JD, Byron A, Humphries MJ. Integrin ligands at a glance. Journal of Cell Science. 2006;119:3901-3.
[156] Grande DA, Southerland SS, Manji R, Pate DW, Schwartz RE, Lucas PA. Repair of articular cartilage defects using mesenchymal stem cells. Tissue engineering. 1995;1:345-53.
[157] Lim H-C, Bae J-H, Song S-H, Park Y-E, Kim S-J. Current Treatments of Isolated Articular Cartilage Lesions of the Knee Achieve Similar Outcomes. Clin Orthop Relat Res. 2012;470:2261-7.
[158] Vasiliadis H, Wasiak J, Salanti G. Autologous chondrocyte implantation for the treatment of cartilage lesions of the knee: a systematic review of randomized studies. Knee Surg Sports Traumatol Arthrosc. 2010;18:1645-55.
[159] Tallheden T, Bengtsson C, Brantsing C, Sjogren-Jansson E, Carlsson L, Peterson L, et al. Proliferation and differentiation potential of chondrocytes from osteoarthritic patients. Arthritis Research & Therapy. 2005;7:R560 - R8.
[160] Giannoni P, Cancedda R. Articular Chondrocyte Culturing for Cell-Based Cartilage Repair: Needs and Perspectives. Cells Tissues Organs. 2006;184:1-15.
[161] Hauselmann HJ, Fernandes RJ, Mok SS, Schmid TM, Block JA, Aydelotte MB, et al. Phenotypic stability of bovine articular chondrocytes after long-term culture in alginate beads. Journal of Cell Science. 1994;107:17-27.
[162] Hauselmann HJ, Masuda K, Hunziker EB, Neidhart M, Mok SS, Michel BA, et al. Adult human chondrocytes cultured in alginate form a matrix similar to native human articular cartilage. American Journal of Physiology - Cell Physiology. 1996;271:C742-C52.
[163] Kiefer MC, Stephans JC, Crawford K, Okino K, Barr PJ. Ligand-affinity cloning and structure of a cell surface heparan sulfate proteoglycan that binds basic fibroblast growth factor. Proceedings of the National Academy of Sciences. 1990;87:6985-9.
[164] Rodgers KD, San Antonio JD, Jacenko O. Heparan sulfate proteoglycans: A GAGgle of skeletal-hematopoietic regulators. Developmental Dynamics. 2008;237:2622-42.
[165] Mariappan MR, Alas EA, Williams JG, Prager MD. Chitosan and chitosan sulfate have opposing effects on collagen–fibroblast interactions. Wound Repair and Regeneration. 1999;7:400-6.
[166] Freeman I, Kedem A, Cohen S. The effect of sulfation of alginate hydrogels on the specific binding and controlled release of heparin-binding proteins. Biomaterials. 2008;29:3260-8.
[167] Re’em T, Kaminer-Israeli Y, Ruvinov E, Cohen S. Chondrogenesis of hMSC in affinity-bound TGF-beta scaffolds. Biomaterials. 2012;33:751-61.
[168] Fan L, Jiang L, Xu Y, Zhou Y, Shen Y, Xie W, et al. Synthesis and anticoagulant activity of sodium alginate sulfates. Carbohydrate Polymers. 2011;83:1797-803.
[169] Ronghua H, Yumin D, Jianhong Y. Preparation and in vitro anticoagulant activities of alginate sulfate and its quaterized derivatives. Carbohydrate Polymers. 2003;52:19-24.
[170] Knudson CB, Knudson W. Cartilage proteoglycans. Seminars in Cell & Developmental Biology. 2001;12:69-78.

References
120
[171] Murray G. Anticoagulant therapy with heparin. The American Journal of Medicine. 1947;3:468-71.
[172] Andersson LO, Barrowcliffe TW, Holmer E, Johnson EA, Sims GEC. Anticoagulant properties of heparin fractionated by affinity chromatography on matrix-bound antithrombin III and by gel filtration. Thrombosis Research. 1976;9:575-83.
[173] Galtrey CM, Fawcett JW. The role of chondroitin sulfate proteoglycans in regeneration and plasticity in the central nervous system. Brain Research Reviews. 2007;54:1-18.
[174] Campo GM, Avenoso A, Campo S, Ferlazzo AM, Micali C, Zanghı ̀L, et al. Hyaluronic acid and chondroitin-4-sulphate treatment reduces damage in carbon tetrachloride-induced acute rat liver injury. Life Sciences. 2004;74:1289-305.
[175] Freeman I, Cohen S. The influence of the sequential delivery of angiogenic factors from affinity-binding alginate scaffolds on vascularization. Biomaterials. 2009;30:2122-31.
[176] Kim KS, Lee JW, Cho SH. Anticoagulation Activities of Low Molecular Weight Sulfated Chitosan and Sulfated Sodium Alginate. Polymer Korea. 2003;27:583-8.
[177] Kasai Y, Akahira A, Kakuta S, Abudula A, Urayama K, Takigawa T. Preparation and Electrochemical Properties of Alginate Sulfate Electrolyte Membranes. Kobunshi Ronbunshu. 2008;65.
[178] Hintze V, Moeller S, Schnabelrauch M, Bierbaum S, Viola M, Worch H, et al. Modifications of Hyaluronan Influence the Interaction with Human Bone Morphogenetic Protein-4 (hBMP-4). Biomacromolecules. 2009;10:3290-7.
[179] Becher J, Möller S, Riemer T, Schiller J, Hintze V, Bierbaum S, et al. Sulfated Glycosaminoglycan Building Blocks for the Design of Artificial Extracellular Matrices. ACS Symposium Series. 2012;1107:315-28.
[180] Wang G, Woods A, Sabari S, Pagnotta L, Stanton L-A, Beier F. RhoA/ROCK Signaling Suppresses Hypertrophic Chondrocyte Differentiation. Journal of Biological Chemistry. 2004;279:13205-14.
[181] Phillips JA, Bonassar LJ. Matrix metalloproteinase activity synergizes with α2β1 integrins to enhance collagen remodeling. Experimental Cell Research. 2005;310:79-87.
[182] Yamaoka H, Asato H, Ogasawara T, Nishizawa S, Takahashi T, Nakatsuka T, et al. Cartilage tissue engineering using human auricular chondrocytes embedded in different hydrogel materials. Journal of Biomedical Materials Research Part A. 2006;78A:1-11.
[183] van Susante JLC, Buma P, van Osch GJVM, Versleyen D, van der Kraan PM, van der Berg WB, et al. Culture of chondrocytes in alginate and collagen carrier gels. Acta Orthopaedica. 1995;66:549-56.
[184] Johnstone B, Hering TM, Caplan AI, Goldberg VM, Yoo JU. In VitroChondrogenesis of Bone Marrow-Derived Mesenchymal Progenitor Cells. Experimental Cell Research. 1998;238:265-72.
[185] Sakaguchi Y, Sekiya I, Yagishita K, Muneta T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis & Rheumatism. 2005;52:2521-9.
[186] De Bari C, Dell'Accio F, Tylzanowski P, Luyten FP. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis & Rheumatism. 2001;44:1928-42.

Chapter 9
121
[187] Erickson GR, Gimble JM, Franklin DM, Rice HE, Awad H, Guilak F. Chondrogenic Potential of Adipose Tissue-Derived Stromal Cells in Vitro and in Vivo. Biochemical and Biophysical Research Communications. 2002;290:763-9.
[188] Awad HA, Quinn Wickham M, Leddy HA, Gimble JM, Guilak F. Chondrogenic differentiation of adipose-derived adult stem cells in agarose, alginate, and gelatin scaffolds. Biomaterials. 2004;25:3211-22.
[189] Ma H-L, Hung S-C, Lin S-Y, Chen Y-L, Lo W-H. Chondrogenesis of human mesenchymal stem cells encapsulated in alginate beads. Journal of Biomedical Materials Research Part A. 2003;64A:273-81.
[190] Mauck RL, Yuan X, Tuan RS. Chondrogenic differentiation and functional maturation of bovine mesenchymal stem cells in long-term agarose culture. Osteoarthritis and Cartilage. 2006;14:179-89.
[191] Wang Y, Kim U-J, Blasioli DJ, Kim H-J, Kaplan DL. In vitro cartilage tissue engineering with 3D porous aqueous-derived silk scaffolds and mesenchymal stem cells. Biomaterials. 2005;26:7082-94.
[192] Cho JH, Kim S-H, Park KD, Jung MC, Yang WI, Han SW, et al. Chondrogenic differentiation of human mesenchymal stem cells using a thermosensitive poly(N-isopropylacrylamide) and water-soluble chitosan copolymer. Biomaterials. 2004;25:5743-51.
[193] Hwang NS, Varghese S, Zhang Z, Elisseeff J. Chondrogenic differentiation of human embryonic stem cell-derived cells in arginine-glycine-aspartate-modified hydrogels. Tissue engineering. 2006;12:2695-706.
[194] Zhou G, Liu W, Cui L, Wang X, Liu T, Cao Y. Repair of porcine articular osteochondral defects in non-weightbearing areas with autologous bone marrow stromal cells. Tissue engineering. 2006;12:3209-21.
[195] Lin C-C, Anseth KS. Controlling Affinity Binding with Peptide-Functionalized Poly(ethylene glycol) Hydrogels. Advanced Functional Materials. 2009;19:2325-31.
[196] Zhu J. Bioactive modification of poly(ethylene glycol) hydrogels for tissue engineering. Biomaterials. 2010;31:4639-56.
[197] Lutolf MP, Lauer-Fields JL, Schmoekel HG, Metters AT, Weber FE, Fields GB, et al. Synthetic matrix metalloproteinase-sensitive hydrogels for the conduction of tissue regeneration: Engineering cell-invasion characteristics. Proceedings of the National Academy of Sciences. 2003;100:5413-8.
[198] Hwang NS, Varghese S, Elisseeff J. Controlled differentiation of stem cells. Advanced Drug Delivery Reviews. 2008;60:199-214.
[199] Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches. Nat Rev Immunol. 2006;6:93-106.
[200] Benoit DSW, Schwartz MP, Durney AR, Anseth KS. Small functional groups for controlled differentiation of hydrogel-encapsulated human mesenchymal stem cells. Nat Mater. 2008;7:816-23.
[201] Salinas CN, Anseth KS. The influence of the RGD peptide motif and its contextual presentation in PEG gels on human mesenchymal stem cell viability. Journal of Tissue Engineering and Regenerative Medicine. 2008;2:296-304.

References
122
[202] Lin Y-C, Brayfield CA, Gerlach JC, Peter Rubin J, Marra KG. Peptide modification of polyethersulfone surfaces to improve adipose-derived stem cell adhesion. Acta Biomaterialia. 2009;5:1416-24.
[203] Liu SQ, Tian Q, Wang L, Hedrick JL, Hui JHP, Yang YY, et al. Injectable Biodegradable Poly(ethylene glycol)/RGD Peptide Hybrid Hydrogels for in vitro Chondrogenesis of Human Mesenchymal Stem Cells. Macromolecular Rapid Communications. 2010;31:1148-54.
[204] Salinas CN, Anseth KS. The enhancement of chondrogenic differentiation of human mesenchymal stem cells by enzymatically regulated RGD functionalities. Biomaterials. 2008;29:2370-7.
[205] Chiu L-H, Chen S-C, Wu K-C, Yang C-B, Fang C-L, Lai W-FT, et al. Differential effect of ECM molecules on re-expression of cartilaginous markers in near quiescent human chondrocytes. Journal of Cellular Physiology. 2011;226:1981-8.
[206] Wojtowicz AM, Shekaran A, Oest ME, Dupont KM, Templeman KL, Hutmacher DW, et al. Coating of biomaterial scaffolds with the collagen-mimetic peptide GFOGER for bone defect repair. Biomaterials. 2010;31:2574-82.
[207] Raynor JE, Petrie TA, García AJ, Collard DM. Controlling Cell Adhesion to Titanium: Functionalization of Poly[oligo(ethylene glycol)methacrylate] Brushes with Cell-Adhesive Peptides. Advanced Materials. 2007;19:1724-8.
[208] Reyes CD, Petrie TA, Burns KL, Schwartz Z, García AJ. Biomolecular surface coating to enhance orthopaedic tissue healing and integration. Biomaterials. 2007;28:3228-35.
[209] Connelly J, Petrie T, García A, Levenston M. Fibronectin-and collagen-mimetic ligands regulate bone marrow stromal cell chondrogenesis in three-dimensional hydrogels. Eur Cell Mater. 2011;22:168-77.
[210] Liu SQ, Tian Q, Hedrick JL, Po Hui JH, Rachel Ee PL, Yang YY. Biomimetic hydrogels for chondrogenic differentiation of human mesenchymal stem cells to neocartilage. Biomaterials. 2010;31:7298-307.
[211] Jongpaiboonkit L, King WJ, Murphy WL. Screening for 3D environments that support human mesenchymal stem cell viability using hydrogel arrays. Tissue engineering Part A. 2009;15:343-53.
[212] Nuttelman CR, Tripodi MC, Anseth KS. Synthetic hydrogel niches that promote hMSC viability. Matrix Biology. 2005;24:208-18.
[213] Von Der Mark K, Gauss V, Von Der Mark H, Muller P. Relationship between cell shape and type of collagen synthesised as chondrocytes lose their cartilage phenotype in culture. Nature. 1977;267:531-2.
[214] Coates EE, Fisher JP. Phenotypic variations in chondrocyte subpopulations and their response to in vitro culture and external stimuli. Annals of Biomedical Engineering. 2010;38:3371-88.
[215] Aydelotte MB, Kuettner KE. Differences between sub-populations of cultured bovine articular chondrocytes. I. Morphology and cartilage matrix production. Connective Tissue Research. 1988;18:205-22.

Chapter 9
123
[216] Schumacher BL, Hughes CE, Kuettner KE, Caterson B, Aydelotte MB. Immunodetection and partial cDNA sequence of the proteoglycan, superficial zone protein, synthesized by cells lining synovial joints. Journal of Orthopaedic Research. 1999;17:110-20.
[217] Flannery CR, Hughes CE, Schumacher BL, Tudor D, Aydelotte MB, Kuettner KE, et al. Articular Cartilage Superficial Zone Protein (SZP) Is Homologous to Megakaryocyte Stimulating Factor Precursor and Is a Multifunctional Proteoglycan with Potential Growth-Promoting, Cytoprotective, and Lubricating Properties in Cartilage Metabolism. Biochemical and Biophysical Research Communications. 1999;254:535-41.
[218] Khalafi A, Schmid TM, Neu C, Reddi AH. Increased accumulation of superficial zone protein (SZP) in articular cartilage in response to bone morphogenetic protein-7 and growth factors. Journal of Orthopaedic Research. 2007;25:293-303.
[219] Neu CP, Khalafi A, Komvopoulos K, Schmid TM, Reddi AH. Mechanotransduction of bovine articular cartilage superficial zone protein by transforming growth factor β signaling. Arthritis & Rheumatism. 2007;56:3706-14.
[220] Lee SY, Niikura T, Reddi AH. Superficial zone protein (lubricin) in the different tissue compartments of the knee joint: modulation by transforming growth factor beta 1 and interleukin-1 beta. Tissue engineering Part A. 2008;14:1799-808.
[221] Wang JG, Miyazu M, Xiang P, Li SN, Sokabe M, Naruse K. Stretch-induced cell proliferation is mediated by FAK-MAPK pathway. Life Sciences. 2005;76:2817-25.
[222] Buschmann MD, Gluzband YA, Grodzinsky AJ, Hunziker EB. Mechanical compression modulates matrix biosynthesis in chondrocyte/agarose culture. Journal of Cell Science. 1995;108:1497-508.
[223] De Croos JNA, Dhaliwal SS, Grynpas MD, Pilliar RM, Kandel RA. Cyclic compressive mechanical stimulation induces sequential catabolic and anabolic gene changes in chondrocytes resulting in increased extracellular matrix accumulation. Matrix Biology. 2006;25:323-31.
[224] Hansen U, Schünke M, Domm C, Ioannidis N, Hassenpflug J, Gehrke T, et al. Combination of reduced oxygen tension and intermittent hydrostatic pressure: a useful tool in articular cartilage tissue engineering. Journal of Biomechanics. 2001;34:941-9.
[225] Saini S, Wick TM. Effect of low oxygen tension on tissue-engineered cartilage construct development in the concentric cylinder bioreactor. Tissue engineering. 2004;10:825-32.
[226] Wernike E, Li Z, Alini M, Grad S. Effect of reduced oxygen tension and long-term mechanical stimulation on chondrocyte-polymer constructs. Cell Tissue Res. 2008;331:473-83.
[227] Klein TJ, Schumacher BL, Blewis ME, Schmidt TA, Voegtline MS, Thonar EJ, et al. Tailoring secretion of proteoglycan 4 (PRG4) in tissue-engineered cartilage. Tissue engineering. 2006;12:1429-39.
[228] Fonseca KB, Bidarra SJ, Oliveira MJ, Granja PL, Barrias CC. Molecularly designed alginate hydrogels susceptible to local proteolysis as three-dimensional cellular microenvironments. Acta Biomaterialia. 2011;7:1674-82.

References
124

125
Curriculum Vitae
Name Rami Mhanna
Date of birth 05. 05. 1982
Nationality Lebanese
Present address Albisstrasse 96, CH-8038 Zurich, Switzerland
Education 2009 – Present PhD Candidate, Swiss Federal Institute of Technology Zurich
(ETH Zürich)
Supervised by Prof. Marcy Zenobi-Wong and Prof. Janos Vörös
Research Topics:
• The effect of the microenvironment on 3D cultured cartilage cells
• The response of cartilage cells to mechanical stimulation • Layer-by-layer films of extracellular matrix molecules • Functionalization of magnetic micro-robots for drug and gene
delivery
2007 – 2008 Masters of Biomedical Engineering, University of Melbourne
• Research Project: On-Chip Emulsions for Drug Delivery Applications, Supervised by Dr. Brigitte Stadler (Score: 89/100)
2001 – 2006 Bachelor of Engineering, Notre Dame University, Lebanon
• Major: Computer and Communication • Senior Project: Wireless Messaging System (Score: 92/100)
Employment History Jan – March 2008 Research Assistant, Department of Chemical and Biomolecular
Engineering,

126
July – Aug 2008 University of Melbourne, Nanostructured Interfaces and Materials Group, Prof. Frank Caruso
• Formation of on-chip emulsions for drug delivery applications • Drug delivery capsules using the layer-by-layer technique
Nov – Dec 2007 Research Assistant, University of Melbourne, Bio21 Institute, Dr. Sally Gras
• Studying the kinetics of collagen fibril formation
2006 – 2007 Technical IT Assistant, Full Time, Telnet, Lebanon
• Worked with a team of engineers in installation of computers, printers, projectors, servers, LANs, WANs and VOIP gateways
• Managed office work and task management for employees
1998 – 2001 Technical Electrician, Part Time, Rashaya Electric, Lebanon
• Construction of PCBs, transformers, UPSs and electrical stabilizers
Publications
Journal Publications:
• Mhanna R, Öztürk E, Vallmajo Martin Q, Millan C, Müller M and Zenobi-Wong M. Biomimetic polyethylene glycol hydrogels functionalized with GFOGER peptides induce chondrogenic differentiation of human mesenchymal stem cells. In preparation • Mhanna R, Kashyap A, Palazzolo G, and Zenobi-Wong M. Chondrocyte Culture in 3D Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers. Submitted to Biomaterials • Mhanna R, Öztürk E, Schlink P, and Zenobi-Wong M. Probing the microenvironmental conditions for induction of superficial zone protein expression. Submitted to Osteoarthritis and Cartilage • Rottmar M, Mhanna R, Vogel V, Zenobi-Wong M, and Maniura-Weber K. Interference with the contractile machinery of the fibroblastic chondrocyte cytoskeleton induces re-expression of the cartilage phenotype. Submitted to Journal of Experimental Cell Research • Sugihara K, Delai M, Mhanna R, Kusch J, Poulikakos D, Vörös J, Zambelli T, and Ferrari A. Label-free detection of cell-contractile activity with lipid nanotubes. Integrative Biology 2013;5(2):423-30 • Mhanna RF, Vörös J, Zenobi-Wong M. Layer-by-layer films made from extracellular matrix macromolecules on silicone substrates. Biomacromolecules 2011; 12: 609-616

127
Patents: • Palazzolo G, Mhanna R, Becher J, Möller S, Schnabelrauch M, and Zenobi-Wong M. Modified alginate hydrogels for tissue engineering and regenerative medicine, European Patent EP 12007934.8, November 19, 2012
Conference Contributions:
• Mhanna R, Vallmajo Martin Q, Öztürk E, Millan C, Müller M, and Wong M. Functionalized biomimetic polyethylene glycol hydrogels utilizing GFOGER peptides for cartilage tissue engineering. 3rd TERMIS World Conference, September 2012, Vienna, Austria
• Qiu F, Mhanna R, Zhang L, Ding Y, Tottori S, Sugihara K, Zenobi-Wong M, and Nelson BJ. Artificial bacterial flagella functionalized with liposomes for biomedical applications. 7th MRC Graduate Symposium, June 2012, Zurich, Switzerland • Mhanna RF, Schlink P, Vörös J, and Marcy Wong. The effect of chondrocyte morphology on their response to mechanical compression. Termis NA Conference, December 2011, Houston, USA • Sugihara K, Delai M, Stuck J, Mhanna R, Ferrari A, Vörös J, Zambelli T. The directed-assembly of lipid nanotubes from inverted-hexagonal structures. 2nd Nanotoday Conference, December 2011, Hawaii, USA • Sugihara K, Delai M, Stuck J, Mhanna R, Ferrari A, Vörös J, Zambelli T. Lipids on polyelectrolytes for biosensing and biomaterials. Biological Surfaces and Interfaces - ESF EMBO, July 2011, Sant Feliu de Guixols, Spain • Rottmar M, Mhanna RF, Wong M and Maniura K. Cell shape versus cytoskeleton integrity: probing the re-expression of the chondrogenic phenotype. Biological Surfaces and Interfaces - ESF EMBO, July 2011, Sant Feliu de Guixols, Spain • Mhanna RF, Schlink P, Vörös J, and Marcy Wong. The effect of chondrocyte morphology on their response to mechanical compression. Biological Surfaces and Interfaces - ESF EMBO, July 2011, Sant Feliu de Guixols, Spain • Mhanna RF, Vörös J, and Wong M. Build-up of layer-by-layer films of extracellular matrix molecules on PDMS. 8th International Symposium on Polyelectrolytes, November 2010, Shanghai, China • Mhanna RF, Schlink P, Vörös J, and Wong M. A new design for 3D loading of cells using controlled alginate gelation. 16th Swiss Conference on Biomaterials - SSB 2010 , May 2010, Zürich, Switzerland • Mhanna RF, Vörös J and Wong M. Layer-by-layer assemblies of extracellular matrix molecules and their effect on loaded chondrocytes. The Swiss Society for Biomedical Engineering Annual Meeting - SSBE 2009 , August 2009, Bern, Switzerland • Mhanna RF, Vörös J and Wong M. Chondrocyte response to tensile strain on layer-by-layer films made from extracellular matrix molecules. 22nd European Conference on Biomaterials - ESB 2009, August 2009, Lausanne, Switzerland

128
Research Mentoring
March – Nov 2012 Swiss Federal Institute of Technology Zurich (ETH Zürich)
Aditya Kashyap, Semester Project
Title: Effect of alginate sulfation on the cartilage phenotype
March – Nov 2012 Queralt Vallmajo Martin, Semester Project
Title: Functionalized PEG hydrogels for cartilage tissue engineering
Sept 2009 – Philippe Schlink, Master Thesis
March 2010 Title: New techniques for application of 3D strains on chondrocytes




![c-Government [Draft so much]](https://cdn.vdocument.in/doc/165x107/55627895d8b42ad1688b4879/c-government-draft-so-much.jpg)


