
Thanks to:
DOE-HGP 1987-2002
DARPA 2001-3
(* Currently unfunded* )
28-Jan-2004 DOE
Synthesis of useful replicating biosystems

DOE Synthetic Genomes: Why?Cheaper/faster "standard biology", hypothesis testing
Systems Biology: Multiple simultaneous tests
Viruses: Aid strain transfer; generate variants, new haplotypes
Anti-viral vaccines and therapeutics (including variants)
In vitro: Make products toxic in E.coli.
Microbes: Interspecific hybrids (e.g. codon usage)
Structural biology: variants
Rapid vaccine response to engineered bioterrorism.
Cell-mediated immunity + humoral.
Fix mismatch between genome analysis & synthesis

DOE Synthetic Genomes: Why?In vitroMicrobial & Human AntimutatorsArtificial ecosystems (laboratory scales)Energy aiding pathway improvementInstrustrial production: Enzymes, SingleCellProtein, Protein-
drugs Remediation: Hybrid genomes (opt. codons), combinatorial
pathway (Maxygen & Diversa). Xylose & OilPharmaceuticals: Combinatorial synthesesNano science Combinatorial syntheses, Complex nanosystems,
more general nanoassembly (in reach of polymerases and ribosome-like factories)
Health research: 10X faster results per current $ (cost/benefit)Hypothesize & test unknown gene combinations Synthetic standards (arrays, MS, quantitation, etc)Agriculture: salt, cold, drought, pest tolerant hybrid genomes

DOE Synthetic GenomesI. Charge & Intro. David & Ray A. Follow on to genome project B. DOE role in tech development in Biology & HGP C. Thumbnail of DOE missions: bioremediation, energy,
carbon sequestrationII. State of the Art of Technology GeorgeIII. New DOE technology. George A. What is new? B. Near and far future. 1&10 yr.IV. Benefits & Concerns 1&10 yr. A. GMO-gene escape (old tech) Jim B. Bioterrorism C. Ownership D. Patents/IP

DOE Synthetic GenomesIV. Benefits & Concerns: Generic, DOE, other A. GMO-gene escape (old tech) Eliminate allergenic (interspecies) pollen B. 10X greater yield for constant research $ (hypothesis
testing & systems biology) C. Pharma throughput D. Vaccines variants F. Bioterrorism (access/regulation, biohacker/vandals) DEA model for tracking chemicals tracking bioprinter cell design output (is this to hard?) G. Ownership, Patents/IP, Research sociology, agenda H. spin-offs: e.g. I/O memory nm3 vs. micron3 I. Religion life (refer to Cho et al. 1999)

Energy & CO2 Fluxes4x1013 kW of sunlight hits earth.We consume 2kW per person* 6x109 = 1010 kW.
CO2 >370 ppm = 730 x1015 g globally, increase ~3 x1015 /yr.Ocean productivity = ~100 x1015 g/yr.
Autotrophs: 1025 Prochlorococcus cells globally (108 per liter)
Undone by Cyanophages & Heterotrophs: 2x1028 SAR11 cells in the oceansPseudomonas & Caulobacter in a variety of soils & aquatic environments
http://www.gsfc.nasa.gov/gsfc/service/gallery/fact_sheets/earthsci/terra/earths_energy_balance.htmhttp://clear.eawag.ch/models/optionenE.html Morris et al. Nature 2002 Dec 19-26;420(6917):806-10. http://hosting.uaa.alaska.edu/mhines/biol468/pages/carbon.html

Synthetic Genomes: How? (examples)
How to decrease cost?Chips, error correction, polony clone/sequence
How to make new biopolymers?Altered translation, mirror proteins
How to improve energy production/conservation?Eliminate predation
How to increase safety?Eliminate DNA/RNA exchange
In vivo vs in vitro?ribosome-display selection, in vivo assembly
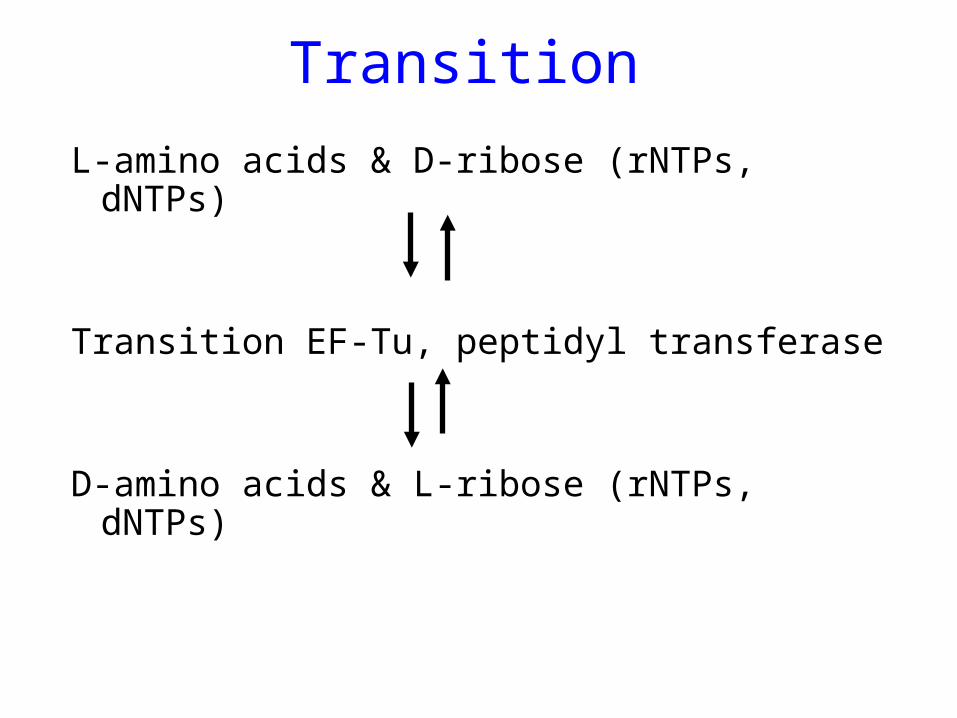
Transition
L-amino acids & D-ribose (rNTPs, dNTPs)
Transition EF-Tu, peptidyl transferase
D-amino acids & L-ribose (rNTPs, dNTPs)

Impact of mirror cells?Microbes Eliminate DNA exchange
Energy Prochlorococcus resistant to phage & predators
Remediation Engineer community resistant to predation
Pharmaceuticals Expand "natural products"
Nano science Enzyme resistant "bio"-polymers
Health Mirror humans resistant to all? viruses ?
Viruses see above
Agriculture Pest resistance

Synthetic Biology
• Test or manipulate optimality• Program minimal cells (105kbp)• Nanobiotechnology - new polymers• Manage complex systems e.g. stem cells & ocean ecology

Synthetic Genomes• Molecular Biology depends on in vitro reactions (e.g. PCR, SP6, Roche)• Utility of mirror-image & other unnatural polymers.• Combine with homologous recombination to engineer larger genomes
Toward these goals design a minimal chassis:• 100kbp genome. • All 3D structures known.• Comprehensive functional data.

Known: in vitro assembly & 3D structure of prokaryotic ribosomes
(e.g. Nomura et al.; Noller et al.)
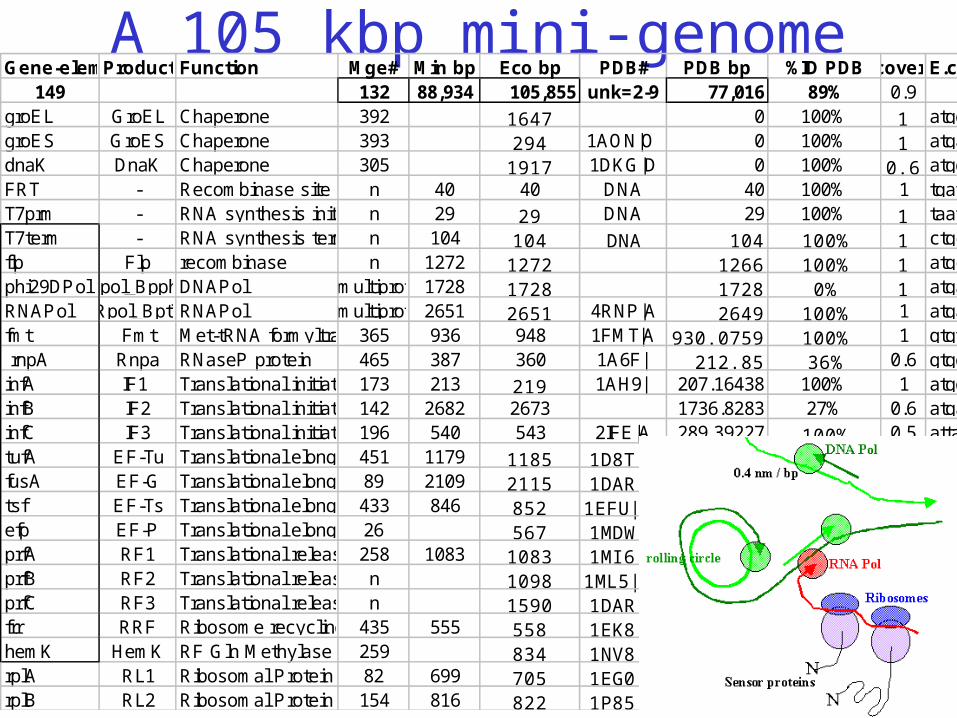
A 105 kbp mini-genomeGene-elementsProductsFunction Mge# Min bp Eco bp PDB# PDB bp %ID PDB%coveredE.coli Sequence Sequence
149 132 88,934 105,855 unk=2-9 77,016 89% 0.9groEL GroEL Chaperone 392 1647 0 100% 1 atggcagctaaagacgtaaaattcggtaacgacgctcgtgtgaaaatgctgcgcggcgtaaacgtactggcagatgcagtgaaagttaccctcggtccaaaaggccgtaacgtagttctggataaatctttcggtgcaccgaccatcaccaaagatggtgtttccgttgctcgtgaaatcgaactggaagacaagttcgaaaatatgggtgcgcagatggtgaaagaagttgcctctaaagcaaacgacgctgcaggcgacggtaccaccactgcaaccgtactggctcaggctatcatcactgaaggtctgaaagctgttgctgcgggcatgaacccgatggacctgaaacgtggtatcgacaaagcggttaccgctgcagttgaagaactgaaagcgctgtccgtaccatgctctgactctaaagcgattgctcaggttggtaccatctccgctaactccgacgaaaccgtaggtaaactgatcgctgaagcgatggacaaagtcggtaaagaaggcgttatcaccgttgaagacggtaccggtctgcaggacgaactggacgtggttgaaggtatgcagttcgaccgtggctacctgtctccttacttcatcaacaagccggaaactggcgcagtagaactggaaagcccgttcatcctgctggctgacaagaaaatctccaacatccgcgaaatgctgccggttctggaagctgttgccaaagcaggcaaaccgctgctgatcatcgctgaagatgtagaaggcgaagcgctggcaactctggttgttaacaccatgcgtggcatcgtgaaagtcgctgcggttaaagcaccgggcttcggcgatcgtcgtaaagctatgctgcaggatatcgcaaccctgactggcggtaccgtgatctctgaagagatcggtatggagctggaaaaagcaaccctggaagacctgggtcaggctaaacgtgttgtgatcaacaaagacaccaccactatcatcgatggcgtgggtgaagaagctgcaaaaattgaagagtttgatcatggctcagattgaacgctggcggcaggcctaacacatgcaagtcgaacggtaacaggaagaagcttgcttctttgctgacgagtggcggacgggtgagtaatgtctgggaaactgcctgatggagggggataactactggaaacggtagctaataccgcataacgtcgcaagaccaaagagggggaccttcgggcctcttgccatcggatgtgcccagatgggattagctagtagggroES GroES Chaperone 393 294 1AON|O 0 100% 1 atgaatattcgtccattgcatgatcgcgtgatcgtcaagcgtaaagaagttgaaactaaatctgctggcggcatcgttctgaccggctctgcagcggctaaatccacccgcggcgaagtgctggctgtcggcaatggccgtatccttgaaaatggcgaagtgaagccgctggatgtgaaagttggcgacatcgttattttcaacgatggctacggtgtgaaatctgagaagatcgacaatgaagaagtgttgatcagcgtaaagaagttgaaactaaatctgctggcggcatcgggttaagcgactaagcgtacacggtggatgccctggcagtcagaggcgatgaaggacgtgctaatctgcgataagcgtcggtaaggtgatatgaaccgttataaccggcgatttccgaatggggaaacccagtgtgtttcgacacactatcattaactgaatccataggttaatgaggcgaaccgggggaactgaaacatctaagtaccccgaggaaaagaaatcaaccgagattcccccagtagcggcgagcgadnaK DnaK Chaperone 305 1917 1DKG|D 0 100% 0.6 atgggtaaaataattggtatcgacctgggtactaccaactcttgtgtagcgattatggatggcaccactcctcgcgtgctggagaacgccgaaggcgatcgcaccacgccttctatcattgcctatacccaggatggtgaaactctagttggtcagccggctaaacgtcaggcagtgacgaacccgcaaaacactctgtttgcgattaaacgcctgattggtcgccgcttccaggacgaagaagtacagcgtgatgtttccatcatgccgttcaaaattattgctgctgataacggcgacgcatgggtcgaagttaaaggccagaaaatggcaccgccgcagatttctgctgaagtgctgaaaaaaatgaagaaaaccgctgaagattacctgggtgaaccggtaactgaagctgttatcaccgtaccggcatactttaacgatgctcagcgtcaggcaaccaaagacgcaggccgtatcgctggtctggaagtaaaacgtatcatcaacgaaccgaccgcagctgcgctggcttacggtctggacaaaggcactggcaaccgtactatcgcggtttatgacctgggtggtggtactttcgatatttctattatcgaaatcgacgaagttgacggcgaaaaaaccttcgaagttctggcaaccaacggtgatacccacctggggggtgaagacttcgacagccgtctgatcaactatctggttgaagaattcaagaaagatcagggcattgacctgcgcaacgatccgctggcaatgcagcgcctgaaagaagcggcagaaaaagcgaaaatcgaactgtcttccgctcagcagaccgacgttaacctgccatacatcactgcagacgcgaccggtccgaaacacatgaacatcaaagtgactcgtgcgaaactggaaagcctggttgaagatctggtaaaccgttccattgagccgctgaaagttgcactgcaggacgctggcctgtccgtatctgatatcgacgacgttatcctcgttggtgtgcctggcggcagtagcgcggtggtcccacctgaccccatgccgaactcagaagtgaaacgccgtagcgccgatggtagtgtggggtctccccatgcgagagtagggaactgccaggcatFRT - Recombinase site n 40 40 DNA 40 100% 1 tgatccgaagttcctattctctagaaagtataggaacttcgaagctgaccagacagtcgccgcttcgtcgtcgtcctcttcgggggagacgggcggaggggaggaaagtccgggctccatagggcagggtgccaggtaacgcctgggggggaaacccacgaccagtgcaacagagagcaaaccgccgatggcccgcgcaagcgggatcaggtaagggtgaaagggtgcggtaagagcgcaccgcgcggctggtaacagtccgtggcacggtaaactccacccggagcaaggccaaT7prm - RNA synthesis initiation siten 29 29 DNA 29 100% 1 taatacgactcactatagggagaccggccgtccccttcgtctagaggcccaggacaccgccctttcacggcggtaacaggggttcgaatcccctaggggacgccaT7term - RNA synthesis termination siten 104 104 DNA 104 100% 1 ctgctaacaaagcccgaaaggaagctgagttggctgctgccaccgctgagcaataactagcataaccccttggggcctctaaacgggtcttgaggggttttttggtgaagatttatctggtcggtggtgctgttcgggatgcattgttagggctaccggtcaaagacagagattgggtggtggtcggcagtacgccacaggagatgctcgacgcgggctaccagcaggtaggccgcgattttcctgtgtttctgcatccgcaaacgcatgaagagtatgcgctggcacgtaccgaacggaaatccggttccggttacaccggttttacttgctatgccgcaccggatgtcacgctggaaflp Flp recombinase n 1272 1272 1266 100% 1 atgccacaatttggtatattatgtaaaacaccacctaaggtgcttgttcgtcagtttgtggaaaggtttgaaagaccttcaggtgagaaaatagcattatgtgctgctgaactaacctatttatgttggatgattacacataacggaacagcaatcaagagagccacattcatgagctataatactatcataagcaattcgctgagtttcgatattgtcaataaatcactccagtttaaatacaagacgcaaaaagcaacaattctggaagcctcattaaagaaattgattcctgcttgggaatttacaattattccttactatggacaaaaacatcaatctgatatcactgatattgtaagtagtttgcaattacagttcgaatcatcggaagaagcagataagggaaatagccacagtaaaaaaatgcttaaagcacttctaagtgagggtgaaagcatctgggagatcactgagaaaatactaaattcgtttgagtatacttcgagatttacaaaaacaaaaactttataccaattcctcttcctagctactttcatcaattgtggaagattcagcgatattaagaacgttgatccgaaatcatttaaattagtccaaaataagtatctgggagtaataatccagtgtttagtgacagagacaaagacaagcgttagtaggcacatatacttctttagcgcaaggggtaggatcgatccacttgtatatttggatgaatttttgaggaattctgaaccagtcctaaaacgagtaaataggaccggcaattcttcaagcaataaacaggaataccaattattaaaagataacttagtcagatcgtacaataaagctttgaagaaaaatgcgccttattcaatctttgctataaaaaatggcccaaaatctcacattggaagacatttgatgacctcatttctttcaatgaagggcctaacggagttgactaatgttgtgggaaattggagcgataagcgtgcttctgccgtggccaggacaaatgacccccgaacaccttccaacagaacagtatgaagcgcagttagccgaaaaagtggtacgtttgcaaagtatgatggcaccgttttctgacctggttccggaagtgtttcgctcgccggtcagtcattaccggatgcgcgcggagttccgcatctggcacgatggcgatgacctgtatcacatcattttcgatcaacaaaccaaaagccgcatccgcgtggatagcttccccgccgccagtgaacttatcaacphi29DPolDpol_Bpph2DNAPol multiprot 1728 1728 1728 0% 1 atgaagcatatgccgagaaagatgtatagttgtgactttgagacaactactaaagtggaagactgtagggtatgggcgtatggttatatgaatatagaagatcacagtgagtacaaaataggtaatagcctggatgagtttatggcgtgggtgttgaaggtacaagctgatctatatttccataacctcaaatttgacggagcttttatcattaactggttggaacgtaatggttttaagtggtcggctgacggattgccaaacacatataatacgatcatatctcgcatgggacaatggtacatgattgatatatgtttaggctacaaagggaaacgtaagatacatacagtgatatatgacagcttaaagaaactaccgtttcctgttaagaagatagctaaagactttaaactaactgttcttaaaggtgatattgattaccacaaagaaagaccagtcggctataagataacacccgaagaatacgcctatattaaaaacgatattcagattattgcggaagctctgttaattcagtttaagcaaggtttagaccggatgacagcaggcagtgacagtctaaaaggtttcaaggatattataaccactaagaaattcaaaaaggtgtttcctacattgagtcttggactcgataaggaagtgagatacgcctatagaggtggttttacatggttaaatgataggttcaaagaaaaagaaatcggagaaggcatggtcttcgatgttaatagtctatatcctgcacagatgtatagtcgtctccttccatatggtgaacctatagtattcgagggtaaatacgtttgggacgaagattacccactacacatacagcatatcagatgtgagttcgaattgaaagagggctatatacccactatacagataaaaagaagtaggttttataaaggtaatgagtacctaaaaagtagcggcggggagatagccgacctctggttgtcaaatgtagacctagaattaatgaaagaacactacgattatggctaaaaaagttaattggtatgtttcttgttcacctagaagtccagaaaaaattcagcctgagttaaaagtactagcaaattttgagggaagttattggaaaggggtaaaagggtataaagcacaagaggcatttgctaaagaacttgctgctttaccacaattcttaggtactacttataaaaaagaagctgcattttctactcgagacagagtggcaccaatgaaaacttatggtttcgtatttgtagatRNAPol Rpol_Bpt7RNAPol multiprot 2651 2651 4RNP|A 2649 100% 1 atgaacacgattaacatcgctaagaacgacttctctgacatcgaactggctgctatcccgttcaacactctggctgaccattacggtgagcgtttagctcgcgaacagttggcccttgagcatgagtcttacgagatgggtgaagcacgcttccgcaagatgtttgagcgtcaacttaaagctggtgaggttgcggataacgctgccgccaagcctctcatcactaccctactccctaagatgattgcacgcatcaacgactggtttgaggaagtgaaagctaagcgcggcaagcgcccgacagccttccagttcctgcaagaaatcaagccggaagccgtagcgtacatcaccattaagaccactctggcttgcctaaccagtgctgacaatacaaccgttcaggctgtagcaagcgcaatcggtcgggccattgaggacgaggctcgcttcggtcgtatccgtgaccttgaagctaagcacttcaagaaaaacgttgaggaacaactcaacaagcgcgtagggcacgtctacaagaaagcatttatgcaagttgtcgaggctgacatgctctctaagggtctactcggtggcgaggcgtggtcttcgtggcataaggaagactctattcatgtaggagtacgctgcatcgagatgctcattgagtcaaccggaatggttagcttacaccgccaaaatgctggcgtagtaggtcaagactctgagactatcgaactcgcacctgaatacgctgaggctatcgcaacccgtgcaggtgcgctggctggcatctctccgatgttccaaccttgcgtagttcctcctaagccgtggactggcattactggtggtggctattgggctaacggtcgtcgtcctctggcgctggtgcgtactcacagtaagaaagcactgatgcgctacgaagacgtttacatgcctgaggtgtacaaagcgattaacattgcgcaaaacaccgcatggaaaatcaacaagaaagtcctagcggtcgccaacgtaatcaatgatgagtattgcgcaggtcagatcggccggaagtgccgggaactattataccgacaaggataattactatgtgctgggcagcatgggagaacgctgggccggcaggggggctgaacagctggggctgcagggcagtgtcgataaggatgtttttacccgtcttctggagggcaggctgccggacggagcggatctaagccgcatgcaggatggcagtaacaggcatcgtcccggctacgatctgaccttctccfmt Fmt Met-tRNA formyltransferase365 936 948 1FMT|A 930.0759 100% 1 gtgtcagaatcactacgtattatttttgcgggtacacctgactttgcagcgcgtcatctcgacgcgctgttgtcttctggtcataacgtcgttggcgtgttcacccagccagaccgaccggcaggacgcggtaaaaaactgatgcccagcccggttaaagttctggctgaggaaaaaggtctgcccgtttttcaacctgtttccctgcgtccacaagaaaaccagcaactggtcgccgaactgcaggctgatgttatggtcgtcgtcgcctatggtttaattctgccgaaagcagtgctggagatgccgcgtcttggctgtatcaacgttcatggttcactgctgccacgctggcgcggtgctgcaccaatccaacgctcactatgggcgggtgatgcagaaactggtgtgaccattatgcaaatggatgtcggtttagacaccggtgatatgctctataagctctcctgcccgattactgcagaagataccagtggtacgctgtacgacaagctggcagagcttggcccacaagggcttatcaccacgttgaaacaactggcagacggcacggcgaaaccagaagttcaggacgaaactcttgtcacttacgccgagaagttgagtaaagaagaagcgcgtattgactggtcactttcggcagcacagcttgaacgctgcattcgcgctttcaatccatggccaatgagctggctggaaattgaaggacagccggttaaagtctggaaagcatcggtcattgatacgcaatccatggccaatgagctggctggaaattgaaggacagccggttaaagtctggaaagcatcggtcattgatacggcaaccaacgctgcaccaggaacgatccttgaagccaacaaacaaggcattcaggttgcgactggtgatggcatcctgaacctgctctcgttacaacctgcgggatgccacaatttggtatattatgtaaaacaccacctaaggtgcttgttcgtcagtttgtggaaaggtttgaaagaccttcaggtgagaaaatagcattatgtgctgctgaactaacctatttatgttggatgattacacataacggaacagcaatcaagagagccacattcatgagctataatactatcataagcaattcgctgagtttcgatattgtcaataaatcactccagtttaaatacaagacgcaaaaa rnpA Rnpa RNaseP protein 465 387 360 1A6F| 212.85 36% 0.6 gtggttaagctcgcatttcccagggagttacgcttgttaactcccagtcaattcacattcgtcttccagcagccacaacgggctggcacgccgcaaattaccattctcggccgcctgaattcgctggggcatccccgtatcggtcttacagtcgccaagaaaaacgttcgacgcgcccatgaacgcaatcggattaaacgtctgacgcgtgaaagcttccgtctgcgccaacatgaactcccggctatggatttcgtctcggccgcctgaattcgctggggcatccccgtatcggtcttacagtcgccaagaaaaacgttcgacgcgcccatgaacgcaatcggattaaacgtctgacgcatgagtaaaggagaagaacttttcactggagttgtcccaattcttgttgaattagatggcgatgttaatgggcaaaaattctctgtcagtggagagggtgaaggtgatgcaacatacggaaaacttacccttaaatttatttgcactactgggaagctacctgttccatggccaacacttgtcactactttcgcgtatggtcttcaatgctttgcgagatacccagatcatatgaaacagcatgactttttcaaginfA IF1 Translational initiation factor 1173 213 219 1AH9| 207.16438 100% 1 atggccaaagaagacaatattgaaatgcaaggtaccgttcttgaaacgttgcctaataccatgttccgcgtagagttagaaaacggtcacgtggttactgcacacatctccggtaaaatgcgcaaaaactacatccgcatcctgacgggcgacaaagtgactgttgaactgaccccgtacgacctgagcaaaggccgcattgtcttccgtagtcgctgagtggttaagctcgcatttcccagggagttacgcttgttaactcccagtcaattcacattcgtcttccagcagccacaacgggctggcacgccgcaaattaccattctcggccgcctgaattcgctggggcatccccgtatcggtcttacagtcgccaagaaaaacgttcgacgcgcccatgaacgcaatcggattaaacgtctgacgcgtgaaagcttccgtctgcgccaacatgaactcccggctatggatttcinfB IF2 Translational initiation factor 2142 2682 2673 1736.8283 27% 0.6 atgacagatgtaacgattaaaacgctggccgcagagcgacagacctccgtggaacgcctggtacagcaatttgctgatgcaggtatccggaagtctgctgacgactctgtgtctgcacaagagaaacagactttgattgaccacctgaatcagaaaaattcaggcccggacaaattgacgctgcaacgtaaaacacgcagcacccttaacattcctggtaccggtggaaaaagcaaatcggtacaaatcgaagtccgcaagaaacgcacctttgtgaaacgcgatccgcaagaggctgaacgccttgcagcggaagagcaagcgcagcgtgaagcggaagagcaagcccgtcgtgaggcagaagaatcggctaaacgcgaggcgcaacaaaaagctgaacgtgaggccgcagaacaagctaagcgtgaagctgctgaacaagcgaaacgtgaagctgcggaaaaagacaaagtgagcaatcaacaagacgatatgactaaaaacgcccaggctgaaaaagcccgccgtgagcaggaagctgcagagctcaagcgtaaagctgaagaagaagcgcgtcgtaaactcgaagaagaagcacgtcgcgttgctgaagaagcacgtcgtatggcggaagaaaacaaatggactgataacgcggaaccgactgaagattccagcgattatcacgtcactacttctcaacatgctcgccaggcagaagacgaaagcgatcgtgaagtcgaaggcggccgtggccgtggtcgtaacgcgaaagcagcgcgtccgaagaaaggcaacaaacacgctgaatcaaaagctgatcgtgaagaagcacgcgcagcagtacgtggcggtaaaggcggaaaacgtaaaggttcttcgctgcagcaaggcttccagaagcctgctcaggccgttaaccgtgacgttgtgatcggcgaaactatcaccgttggcgaactggcgaacaagatggcggttaaaggctctcaggtcatcaaagcgatgatgaaactgggcgcaaatgagattgaagaaaaaactcgtcttaattgatggcaacagtgtggcataccgcgccttttttgccttgccacttttgcataacgacaaaggcattcatacgaatgcggtttacgggtttacgatgatgttgaacaaaattttggcggaagaacaaccgacccatttacttgtagcgtttgacgccggaaaaacgacgttccggcatgaaacgtttcaagagtataaaggcggacggcaacaaacgcccccggaainfC IF3 Translational initiation factor 3196 540 543 2IFE|A 289.39227 100% 0.5 attaaaggcggaaaacgagttcaaacggcgcgccctaaccgtatcaatggcgaaattcgcgcccaggaagttcgcttaacaggtctggaaggcgagcagcttggtattgtgagtctgagagaagctctggagaaagcagaagaagccggagtagacttagtcgagatcagccctaacgccgagccgccggtttgtcgtataatggattacggcaaattcctctatgaaaagagcaagtcttctaaggaacagaagaaaaagcaaaaagttatccaggttaaggaaattaaattccgtcctggtacagatgaaggcgactatcaggtaaaactccgcagcctgattcgctttctcgaagagggtgataaagccaaaatcacgctgcgtttccgcggtcgtgagatggcgcaccagcaaatcggtatggaagtgcttaatcgcgtgaaagacgatttgcaagaactggcagtggtcgaatccttcccaacgaagatcgaaggccgccagatgattaaattccgtcctggtacagatgaaggcgacatgaacacgattaacatcgctaagaacgacttctctgacatcgaactggctgctatcccgttcaacactctggctgaccattacggtgagcgtttagctcgcgaacagttggcccttgagcatgagtcttacgagatgggtgaagcacgcttccgcaagatgtttgagcgtcaacttaaagctggtgaggttgcggataacgctgccgccaagcctctcatcactaccctactccctaagatgattgcacgcatctufA EF-Tu Translational elongation factor Tu451 1179 1185 1D8T|A 1173.03 100% 1 gtgtctaaagaaaaatttgaacgtacaaaaccgcacgttaacgttggtactatcggccacgttgaccacggtaaaactactctgaccgctgcaatcaccaccgtactggctaaaacctacggcggtgctgctcgtgcattcgaccagatcgataacgcgccggaagaaaaagctcgtggtatcaccatcaacacttctcacgttgaatacgacaccccgacccgtcactacgcacacgtagactgcccggggcacgccgactatgttaaaaacatgatcaccggtgctgctcagatggacggcgcgatcctggtagttgctgcgactgacggcccgatgccgcagactcgtgagcacatcctgctgggtcgtcaggtaggcgttccgtacatcatcgtgttcctgaacaaatgcgacatggttgatgacgaagagctgctggaactggttgaaatggaagttcgtgaacttctgtctcagtacgacttcccgggcgacgacactccgatcgttcgtggttctgctctgaaagcgctggaaggcgacgcagagtgggaagcgaaaatcctggaactggctggcttcctggattcttatattccggaaccagagcgtgcgattgacaagccgttcctgctgccgatcgaagacgtattctccatctccggtcgtggtaccgttgttaccggtcgtgtagaacgcggtatcatcaaagttggtgaagaagttgaaatcgttggtatcaaagagactcagaagtctacctgtactggcgttgaaatgttccgcaaactgctggacgaaggccgtgctggtgagaacgtaggtgttctgctgcgtggtatcaaacgtgaagaaatcgaacgtggtcaggtactggctaagccgggcaccatcaagccgcacaccaagttcgaatctgaagtgtacattctgtccaaagatgaaggcggccgtcatactccgttcttcaaaggctaccgtccgcagttctacttccgtactactgacgtgactggtaccagtgtctaaagaaaaatttgaacgtacaaaaccgcacgttaacgttggtactatcggccacgttgaccacggtaaaactactctgaccgctgcaatcaccaccgtactggctaaaacctacggcggtgctgctcgtgcattcgaccagatcgataacgcgccggaagaaaaagctcgtggtatcaccatcaacacttctcacgttgaatacgacaccccgacccgtcactacgcacacgtagactgcccggggcacfusA EF-G Translational elongation factor G89 2109 2115 1DAR| 2085.068 60% 1 atggctcgtacaacacccatcgcacgctaccgtaacatcggtatcagtgcgcacatcgacgccggtaaaaccactactaccgaacgtattctgttctacaccggtgtaaaccataaaatcggtgaagttcatgacggcgctgcaaccatggactggatggagcaggagcaggaacgtggtattaccatcacttccgctgcgactactgcattctggtctggtatggctaagcagtatgagccgcatcgcatcaacatcatcgacaccccggggcacgttgacttcacaatcgaagtagaacgttccatgcgtgttctcgatggtgcggtaatggtttactgcgcagttggtggtgttcagccgcagtctgaaaccgtatggcgtcaggcaaacaaatataaagttccgcgcattgcgttcgttaacaaaatggaccgcatgggtgcgaacttcctgaaagttgttaaccagatcaaaacccgtctgggcgcgaacccggttccgctgcagctggcgattggtgctgaagaacatttcaccggtgttgttgacctggtgaaaatgaaagctatcaactggaacgacgctgaccagggcgtaaccttcgaatacgaagatatcccggcagacatggttgaactggctaacgaatggcaccagaacctgatcgaatccgcagctgaagcttctgaagagctgatggaaaaatacctgggtggtgaagaactgactgaagcagaaatcaaaggtgctctgcgtcagcgcgttctgaacaacgaaatcatcctggtaacctgtggttctgcgttcaagaacaaaggtgttcaggcgatgctggatgcggtaattgattacctgccatccccggttgacgtacctgcgatcaacggtatcctggacgacggtaaagacactccggctgaacgtcacgcaagtgatgacgagccgttctctgcactggcgttcaaaatcgctaccgacccgtttgttggtaacctgaccttcttccgtgtttactccggtgatggctcgtacaacacccatcgcacgctaccgtaacatcggtatcagtgcgcacatcgacgccggtaaaaccactactaccgaacgtattctgttctacaccggtgtaaaccataaaatcggtgaagttcatgacggcgctgcaaccatggactggatggagcaggagcaggaacgtggtattaccatcacttccgctgcgactactgcattctggtctggtatggctaagcagtatgagccgcatcgcatcaactsf EF-Ts Translational elongation factor Ts433 846 852 1EFU|B 840.0423 100% 1 atggctgaaattaccgcatccctggtaaaagagctgcgtgagcgtactggcgcaggcatgatggattgcaaaaaagcactgactgaagctaacggcgacatcgagctggcaatcgaaaacatgcgtaagtccggtgctattaaagcagcgaaaaaagcaggcaacgttgctgctgacggcgtgatcaaaaccaaaatcgacggcaactacggcatcattctggaagttaactgccagactgacttcgttgcaaaagacgctggtttccaggcgttcgcagacaaagttctggacgcagctgttgctggcaaaatcactgacgttgaagttctgaaagcacagttcgaagaagaacgtgttgcgctggtagcgaaaattggtgaaaacatcaacattcgccgcgttgctgcgctggaaggcgacgttctgggttcttatcagcacggtgcgcgtatcggcgttctggttgctgctaaaggcgctgacgaagagctggttaaacacatcgctatgcacgttgctgcaagcaagccagaattcatcaaaccggaagacgtatccgctgaagtggtagaaaaagaataccaggtacagctggatatcgcgatgcagtctggtaagccgaaagaaatcgcagagaaaatggttgaaggccgcatgaagaaattcaccggcgaagtttctctgaccggtcagccgttcgttatggaaccaagcaaaactgttggtcagctgctgaaagagcataacgctgaagtgactggcttcggatatcgcgatgcagtctggtaagccgaaagaaatcgcagagaaaatggttgaaggccgcatgaagaaattcaccggcgaagtatggctgaaattaccgcatccctggtaaaagagctgcgtgagcgtactggcgcaggcatgatggattgcaaaaaagcactgactgaagctaacggcgacatcgagctggcaatcgaaaacatgcgtaagtccggtgctattaaagcagcgaaaaaagcaggcaacgttgctgctgacggcgtgatcaaaaccaaaatcgacggcaactacggcatcattctggaagttaactgccagactgacttcgttgcaaaaefp EF-P Translational elongation factor P26 567 1MDW|A 0 31% 0.4 atggcaacgtactatagcaacgattttcgtgctggtcttaaaatcatgttagacggcgaaccttacgcggttgaagcgagtgaattcgtaaaaccgggtaaaggccaggcatttgctcgcgttaaactgcgtcgtctgctgaccggtactcgcgtagaaaaaaccttcaaatctactgattccgctgaaggcgctgatgttgtcgatatgaacctgacttacctgtacaacgacggtgagttctggcacttcatgaacaacgaaactttcgagcagctgtctgctgatgcaaaagcaattggtgacaacgctaaatggctgctggatcaggcagagtgtatcgtaactctgtggaatggtcagccgatctccgttactccgccgaacttcgttgaactggaaatcgttgataccgatccgggcctgaaaggtgataccgcaggtactggtggcaaaccggctaccctgtctactggcgctgtggttaaagttccgctgtttgtacaaatcggtaaatggctgctggatcaggcagagtgtatcgtaactctgtggaatggtcagccgatggcaacgtactatagcaacgattttcgtgctggtcttaaaatcatgttagacggcgaaccttacgcggttgaagcgagtgaattcgtaaaaccgggtaaaggccaggcatttgctcgcgttaaactgcgtcgtctgctgaccggtactcgcgtagaaaaaaccttcaaatctactgattccgctgaaggcgctgatgttgtcgatatgaacctgacttacctgtacaacgacggtgagttctggcacttcatgprfA RF1 Translational release factor UAA,UAG258 1083 1083 1MI6|A 1008 40% 0.9 atgaagccttctatcgttgccaaactggaagccctgcatgaacgccatgaagaagttcaggcgttgctgggtgacgcgcaaactatcgccgaccaggaacgttttcgcgcattatcacgcgaatatgcgcagttaagtgatgtttcgcgctgttttaccgactggcaacaggttcaggaagatatcgaaaccgcacagatgatgctcgatgatcctgaaatgcgtgagatggcgcaggatgaactgcgcgaagctaaagaaaaaagcgagcaactggaacagcaattacaggttctgttactgccaaaagatcctgatgacgaacgtaacgccttcctcgaagtccgagccggaaccggcggcgacgaagcggcgctgttcgcgggcgatctgttccgtatgtacagccgttatgccgaagcccgccgctggcgggtagaaatcatgagcgccagcgagggtgaacatggtggttataaagagatcatcgccaaaattagcggtgatggtgtgtatggtcgtctgaaatttgaatccggcggtcatcgcgtgcaacgtgttcctgctacggaatcgcagggtcgtattcatacttctgcttgtaccgttgcggtaatgccagaactgcctgacgcagaactgccggacatcaacccagcagatttacgcattgatactttccgctcgtcaggggcgggtggtcagcacgttaacaccaccgattcggcaattcgtattactcacttgccgaccgggattgttgttgaatgtcaggacgaacgttcacaacataaaaacaaagctaaagcactttctgttctcggtgctcgcatccacgctgctgaaatggcaaaacgccaacaggccgaagcgtctacccgtcgtaacctgctggggagtggcgatcgcagcgaccgtaaccgtacttacaacttcccgcaggggcgcgttaccgatcaccgcatcaacctgacgctctaccgcctggatgaagtgatggaaggtaagctggatatgctgattgatggccaaagaagacaatattgaaatgcaaggtaccgttcttgaaacgttgcctaataccatgttccgcgtagagttagaaaacggtcacgtggttactgcacacatctccggtaaaatgcgcaaaaactacatccgcatcctgacgggcgacaaagtgactgttgaactgaccccgtacgacctgagcaaaggccgcattgtcttccgtagtcgctgaprfB RF2 Translational release factor UAA,UAGn 1098 1ML5|Z 0 100% 1 atgtttgaaattaatccggtaaataatcgcattcaggacctcacggaacgctccgacgttcttagggggtatcttgactacgacgccaagaaagagcgtctggaagaagtaaacgccgagctggaacagccggatgtctggaacgaacccgaacgcgcacaggcgctgggtaaagagcgttcctccctcgaagccgttgtcgacaccctcgaccaaatgaaacaggggctggaagatgtttctggtctgctggaactggctgtagaagctgacgacgaagaaacctttaacgaagccgttgctgaactcgacgccctggaagaaaaactggcgcagcttgagttccgccgtatgttctctggcgaatatgacagcgccgactgctacctcgatattcaggcggggtctggcggtacggaagcacaggactgggcgagcatgcttgagcgtatgtatctgcgctgggcagaatcgcgtggtttcaaaactgaaatcatcgaagagtcggaaggtgaagtggcgggtattaaatccgtgacgatcaaaatctccggcgattacgcttacggctggctgcgtacagaaaccggcgttcaccgcctggtgcgtaaaagcccgtttgactccggcggtcgtcgccacacgtcgttcagctccgcgtttgtttatccggaagttgatgatgatattgatatcgaaatcaacccggcggatctgcgcattgacgtttatcgcacgtccggcgcgggcggtcagcacgttaaccgtaccgaatctgcggtgcgtattacccacatcccgaccgggatcgtgacccagtgccagaacgaccgttcccagcacaagaacaaagatcaggccatgaagcagatgaaagcgaagctttatgaactggagatgcagaagaaaaatgccgagaaacaggcgatggaagataacaaatccgacatcggctggggcagccagattcgttcttatgtccttgatgactcccgcattaaagatctgcgcaccggggtagaaaatgacagatgtaacgattaaaacgctggccgcagagcgacagacctccgtggaacgcctggtacagcaatttgctgatgcaggtatccggaagtctgctgacgactctgtgtctgcacaagagaaacagactttgattgaccacctgaatcagaaaaattcaggcccggacaaattgacgctgcaacgtaaaacacgcagcacccttaacattcctggtaccggtggaaaaagcaaatcggtacaaatcgaagtcprfC RF3 Translational release factor 3n 1590 1DAR| 0 28% 0.9 atgacgttgtctccttatttgcaagaggtggcgaagcgccgcacttttgccattatttctcacccggacgccggtaagactaccatcaccgagaaggtgctgctgttcggacaggccattcagaccgccggtacagtaaaaggccgtggttccaaccagcacgctaagtcggactggatggagatggaaaagcagcgtgggatctccattactacgtctgtgatgcagtttccgtatcacgattgcctggttaacctgctcgacaccccggggcacgaagacttctcggaagatacctatcgtaccctgacggcggtggactgctgcctgatggttatcgacgccgcaaaaggtgttgaagatcgtacccgtaagctgatggaagttacccgtctgcgcgacacgccgatcctcacctttatgaacaaacttgaccgtgatatccgcgacccgatggagctgctcgatgaagttgagaacgagctgaaaatcggctgtgcgccgatcacctggccgattggctgcggcaagctgtttaaaggcgtttaccacctttataaagacgaaacctatctctatcagagcggtaaaggccacaccattcaggaagtccgcattgttaaagggctgaataacccggatctcgatgctgcggttggtgaagatctggcacagcagctgcgtgacgaactggaactggtgaaaggcgcgtctaacgagttcgacaaagagctgttccttgcgggcgaaatcactccggtattcttcggtactgcgctgggtaacttcggcgtcgatcatatgttggatggcctggtggagtgggcacctgcgccgatgccgcgtcagactgatacccgtaccgtagaagcgagcgaagataaatttaccggcttcgtatttaaaattcaggccaacatggacccgaaacaccgcgaccgcgtggcgtttatgcgtgtggtgtccggtaaatatgaaaaaggcatgaaactgcgccaggtgcgcactgcgaaagatgtggtgaattaaaggcggaaaacgagttcaaacggcgcgccctaaccgtatcaatggcgaaattcgcgcccaggaagttcgcttaacaggtctggaaggcgagcagcttggtattgtgagtctgagagaagctctggagaaagcagaagaagccggagtagacttagtcgagatcagccctaacgccgagccgccggtttgtcgtataatggattacggcaaattcctctatgaaaagagcaagtcttctaaggaacagaagfrr RRF Ribosome recycling factor435 555 558 1EK8|A 552.0161 99% 1 gtgattagcgatatcagaaaagatgctgaagtacgcatggacaaatgcgtagaagcgttcaaaacccaaatcagcaaaatacgcacgggtcgtgcttctcccagcctgctggatggcattgtcgtggaatattacggcacgccgacgccgctgcgtcagctggcaagcgtaacggtagaagattcccgtacactgaaaatcaacgtgtttgatcgttcaatgtctccggccgttgaaaaagcgattatggcgtccgatcttggcctgaacccgaactctgcgggtagcgacatccgtgttccgctgccgccgctgacggaagaacgtcgtaaagatctgaccaaaatcgttcgtggtgaagcagaacaagcgcgtgttgcagtacgtaacgtgcgtcgtgacgcgaacgacaaagtgaaagcactgttgaaagataaagagatcagcgaagacgacgatcgccgttctcaggacgatgtacagaaactgactgatgctgcaatcaagaaaatgctgccgccgctgacggaagaacgtcgtaaagatctgaccaaaatcatgaagccttctatcgttgccaaactggaagccctgcatgaacgccatgaagaagttcaggcgttgctgggtgacgcgcaaactatcgccgaccaggaacgttttcgcgcattatcacgcgaatatgcgcagttaagtgatgtttcgcgctgttttaccgactggcaacaggttcaggaagatatcgaaaccgcacagatgatgctcgatgatcctgaaatgcgtgagatggcgcaggatgaactgcgcgaagcthemK HemK RF Gln Methylase 259 834 1NV8|A 0 33% 0.7 atggaatatcaacactggttacgtgaagcaataagccaacttcaggcgagcgaaagcccgcggcgtgatgctgaaatcctgctggagcatgttaccggcagagggcgtacttttattctcgcctttggtgaaacgcagctgactgacgaacaatgtcagcaacttgatgcgctactgacacgtcgtcgcgatggtgaacccattgctcatttaaccggggtgcgagaattctggtcgttgccgttatttgtttcgccagcgaccttaattccgcgcccggatacggagtgtctggtggagcaggcactggcgcggttgcctgaacaaccttgccgtattctcgatctcgggacgggtaccggggcgattgcgcttgcgctggctagcgagcgcccggactgcgaaattatcgctgtagatcgtatgcctgatgctgtctccctggcacaacgtaatgcccagcatctggcgatcaaaaatatccacattctgcaaagcgactggtttagcgcgctagccgggcagcagtttgcgatgattgtcagcaatccgccgtatattgacgagcaggaccctcatcttcaacaaggcgatgtccgctttgagccgctcactgcgctggttgcggcagacagtggaatggcagacatcgtgcatatcatcgaacagtcgcgtaacgcgctggtatccggcggctttctgcttctggaacatggctggcagcagggcgaagcggtgcgacaagcatttatcctcgcggggtatcattcttcaacaaggcgatgtccgctttgagccgctcactgcgctggttgcggcagacagtggaatggcgtgattagcgatatcagaaaagatgctgaagtacgcatggacaaatgcgtagaagcgttcaaaacccaaatcagcaaaatacgcacgggtcgtgcttctcccagcctgctggatggcattgtcgtggaatattacggcacgccgacgccgctgcgtcagctggcaagcgtaacggtagaagattcccgtacactgaaaatcaacgtgtttgatcgttcaatgtctccggccgttgaaaaagcgattatggcgtccrplA RL1 Ribosomal Protein Large subunit 82 699 705 1EG0|N 672.2298 50% 1 atggctaaactgaccaagcgcatgcgtgttatccgcgagaaagttgatgcaaccaaacagtacgacatcaacgaagctatcgcactgctgaaagagctggcgactgctaaattcgtagaaagcgtggacgtagctgttaacctcggcatcgacgctcgtaaatctgaccagaacgtacgtggtgcaactgtactgccgcacggtactggccgttccgttcgcgtagccgtatttacccaaggtgcaaacgctgaagctgctaaagctgcaggcgcagaactggtaggtatggaagatctggctgaccagatcaagaaaggcgaaatgaactttgacgttgttattgcttctccggatgcaatgcgcgttgttggccagctgggccaggttctgggtccgcgcggcctgatgccaaacccgaaagtgggtactgtaacaccgaacgttgctgaagcggttaaaaacgctaaagctggccaggttcgttaccgtaacgacaaaaacggcatcattgaagcggttaaaaacgctaaagctggccaggttcgttaccgtaacgacaaaaacggcatcatccacaccaccatcggtaaagtggactttgacgctgacaaactgaaagaaaacctggaagctctgctggttgcgctgaaaaaagcaaaaccgactcaggcgaaaggcgtgtacatcaagaaagttagcatcatggctaaactgaccaagcgcatgcgtgttatccgcgagaaagttgatgcaaccaaacagtacgacatcaacgaagctatcgcactgctgaaagagctggcgactgctaaattcgtagaaagcgtggacgtagctgttaacctcggcatcgacgctcgtaaatctgaccagaacgtacgtggtgcaactgtactgccgcacggtactggccgttccgttcgcgtagccgtatttacccaaggtgcaaacgctgaarplB RL2 Ribosomal Protein Large subunit 154 816 822 1P85|A 810.0438 100% 1 atggcagttgttaaatgtaaaccgacatctccgggtcgtcgccacgtagttaaagtggttaaccctgagctgcacaagggcaaaccttttgctccgttgctggaaaaaaacagcaaatccggtggtcgtaacaacaatggccgtatcaccactcgtcatatcggtggtggccacaagcaggcttaccgtattgttgacttcaaacgcaacaaagacggtatcccggcagttgttgaacgtcttgagtacgatccgaaccgttccgcgaacatcgcgctggttctgtacaaagacggtgaacgccgttacatcctggcccctaaaggcctgaaagctggcgaccagattcagtctggcgttgatgctgcaatcaaaccaggtaacaccctgccgatgcgcaacatcccggttggttctactgttcataacgtagaaatgaaaccaggtaaaggcggtcagctggcacgttccgctggtacttacgttcagatcgttgctcgtgatggtgcttatgtcaccctgcgtctgcgttctggtgaaatgcgtaaagtagaagcagactgccgtgcaactctgggcgaagttggcaatgctgagcatatgctgcgcgttctgggtaaagcaggtgctgcacgctggcgtggtgttcgtccgaccgttcgcggtaccgcgatgaacccggtagaccacccacatggtggtggtgaaggtcgtaactttggtaagcacccggtaactccgtggggcgttcagaccaaaggtaagaagtgcaactctgggcgaagttggcaatgctgagcatatgctgcgcgttctgggtaaatggcagttgttaaatgtaaaccgacatctccgggtcgtcgccacgtagttaaagtggttaaccctgagctgcacaagggcaaaccttttgctccgttgctggaaaaaaacagcaaatccggtggtcgtaacaacaatggccgtatcaccactcgtcatatcggtggtggccacaagcaggcttaccgtattgttgacttcaaacgcaacaaagacggtatcccggcagttgttgaacgtcttgagtacgatccg

The least well-characterized components of the mini-genome
valV GACa GUY tRNA y 77 77 rrsB 16S Small subunit Ribosomal RNA y 1418 1542 rrlB 23S Large subunit Ribosomal RNA y 2903 2904 rrfB 5S Small subunit Ribosomal RNA y 120 120rnpB 10Sb RNaseP RNA y 375 377tadA (I)TadA tRNA-Arg wobble Adenine deaminasen 504 504tilS (})TilS tRNA-Ile-Lysidine synth ? 1299 1299trmD (k)TrmD tRNA N1G37methylase anticodon445 768 768tsaA (6) min 4.6? tRNA t6A=N6-threonylcarbamoyladenosine (I,K,N,S,R)y 1000 1000miaA (+)MiaA isopentenylpyrophosphate tRNA-adenosine transferase n 951trmU (2)AsuE,MnmA,YcfB,B1133tRNA nm5U-methyltransferase(P25745) or Sulfur?? 1152iscS (2)YfhO,B2530 tRNA 2sU, Cysteine desulfurase n 1239trmE (!), MnmE,B3706, ThdFtRNA U > cmnm5U, GTPase ? 1365 1365trmC ({)MnmC, TrmC, yfcK=b2324?tRNA cmnm5U > nm5U & mnm5U? 2067gidA (!)GidA tRNA U > cmnm5U ? 1890 1890miaB MiaB tRNA i6a > s2i6A (or ms2i6A) n 1425?? (V) cmo5U = uridine 5-oxyacetic acid n 0
Forester & Church

M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
DNA Template
RNA Transcript
All 30S-Ribosomal-protein DNAs & mRNAs synthesized in vitro
Tian & Church

His-tagged ribosomal proteins synthesized in vitro
RS-2,4,5,6,9,10,12,13,15,16,17,and 21 as original constructs.
RS1 required deletion of a feedback motif in the mRNA.RS-3, 7, 8, 11, 14, 18, 19, 20 are still weakly expressed.
Note that S1, S4, S7, S8, S20, L1, L4, L10 are known to repress their own translation (and are likely titrated by rRNA).
Iteratively resynthesize all mRNAs (e.g. with less structure).
Tian & Church

Custom Oligonucleotide Chips
*Photo-Generated Acid
Electrolytic acid/base
Photolabile 5'protecting group
Ink-jet with standard monomers and acids
*http://www.xeotron.com/fw/main/default.asp
http://www.oxamer.com/
http://www.febit.com/geniom/go_DNAProc.htm
http://www.nimblegen.com/
http://www.chem.agilent.com/Scripts/PDS.asp?lPage=3071
Tian, Sheng, Gao & Church

Improving synthesis fidelity
MethodTotal
bp#
ClonesTrans-ition
Trans-version Deletion Addition Bp/error
Hyb selection, PCR 2787 9 1 0 1 0 1394Gel selection, PCR 24546 35 28 12 11 3 455
No selection, ligation+PCR 6093 25 6 6 22 4 160
No selection, PCR 9243 21 25 13 19 1 159

Why mismatch repair works for in vivo replication (& not for de novo synthesis)
Desired (or original) base pair: AT ;
Mutant bp: gc
Heteroduplexes: Ac & gT.
Mismatch repair (selecting original): At & aT
Mismatch repair (no "original"): At, aT, gc, gc
Solution: Select on majority (once or more)(1) Kinetic: + (AT) -(gc, AC , gT) (2) Equilibrium: + (AT , gc) -(Ac , gT)

Improving automated cost
oligos seq fix 5E+3 1.00E+06
Cello $1,000 $200 0 $1,200 $240,000
Smith0 $1,000 $800,000 0 $801,000 $160,200,000
Smith1 $1,000 $20 $40 $1,060 $212,000
Smith2 $1,000 $200 $24 $1,224 $244,800
Church1 $2,500 $20 $16 $2,536 $507,200
Church2 $45 $20 $16 $81 $16,129
Church0 $45 $5 0 $50 $9,929







