
The Pennsylvania State University
The Graduate School
College of Health and Human Development
VITAMIN A METABOLISM IN THE NEONATAL LUNG:
STUDIES IN A RAT MODEL
A Dissertation in
Nutrition
by
Lili Wu
2011 Lili Wu
Submitted in Partial Fulfillment
of the Requirements
for the Degree of
Doctor of Philosophy
December 2011

1
The dissertation of Lili Wu was reviewed and approved* by the following:
A. Catharine Ross
Professor of Nutritional Sciences
Occupant of Dorothy Foehr Huck Chair
Dissertation Advisor
Chair of Committee
Okhee Han
Assistant Professor of Nutritional Sciences
Katarzyna Kordas
Assistant Professor of Nutritional Sciences
Pamela J. Mitchell
Associate Professor of Biology and Molecular
Jeffery M. Dodds
Attending Veterinarian for Laboratory Animals
Gordon L. Jensen
Professor of Nutritional Sciences
Head of the Department of Nutritional Sciences
*Signatures are on file in the Graduate School

iii
ABSTRACT
Vitamin A (VA) is an essential nutrient for differentiation and maturation of the
lungs. Direct evidence has been presented that keratinizing squamous metaplasia of the
bronchopulmonary tree can be caused by VA deprivation in the lungs of VA deficient
animals and this morphologic change can be reversed by refeeding the animals with VA.
Other biochemical and molecular genetic evidence revealed that specific retinoids-
binding proteins and nuclear retinoic acid receptors are contained in the lungs and
retinoids can affect lung cells differentiation by influencing lung gene expression. In rats,
significant storage of VA in the lungs starts in late gestation just before the onset of
alveologenesis and surfactant synthesis, and then quickly is depleted during late
pregnancy and postnatal life as the lungs are still developing, suggesting a high and acute
demand of VA for postnatal lung development. However, the VA status in the lung of
human and other mammals is known to be low at birth and postnatal lungs are very
sensitive to dietary VA deprivation. VA deficiency associated with premature infants or
infants with very-low-body-weight (VLBW) can interrupt normal development and
compromise the respiratory function of the lung, thereby putting this population at high
risk to develop various respiratory diseases. Therefore, it is important to improve lung
VA status at the early time of the postnatal life.
Although supplementation with VA to neonates is an effective way to prevent VA
deficiency, it shows limited effectiveness in improving lung VA status. Previously we
have shown that retinol combined with retinoic acid (RA), a biologically active

iv
metabolite of VA (VARA), is able to increase lung retinyl ester (RE) formation
synergistically and RA redirects more of the VA given as a supplement into the neonatal
lung. Our work investigating the molecular mechanism of VARA synergy revealed that
RA is able to affect lung VA metabolism by upregulating several important retinoid
homeostatic genes: LRAT, lecithin:retinol acyltransferase, an enzyme converting retinol
to its storage form; CYP26, a cytochrome P450, an enzyme metabolizing RA to inactive
polar metabolites; and STRA6, stimulated by retinoic acid gene 6, a transmembrane
receptor for the retinol-RBP complex that mediate cellular retinol uptake. However, these
findings are based on a single dose study, and the activity of RA on gene induction
appears to be transient.
In the present study, we tested the effects of repeated supplementation with
VARA in increasing lung RE contents. We also examined whether inflammation state
and reduced RA concentration could affect the capability of RA in promoting RE
formation. At the same time, we speculated the spatial expression pattern of LRAT,
CYP26B1 and STRA6 to further understand VA metabolism in the lungs of the neonates.
We carried out several studies to 1) investigate how multiple treatments of RA during the
period of lung septation affect RE accumulation and the expression pattern of lung
retinoid homeostatic genes, or genes required for normal lung function; 2) compare a
reduced amount of RA in the VARA dose to test the potential of RA in elevating lung
RE; 3) examine how lipopolysaccharide (LPS)-induced inflammation state affects VA
homeostasis in neonatal lung; 4) determine the localization of retinoid homeostasis
proteins in the lung. The results of our studies have shown that repeated treatments of

v
VARA dramatically increase neonatal lung RE store in a cumulative and synergistic way.
Diluted RA in VARA still promotes higher RE formation in neonatal lung more than VA
alone after a single dose, but not after multiple doses. LPS-induced inflammation doesn’t
significantly impact lung RE formation promoted by RA. The localization study
suggested the expression of LRAT in lipofibroblasts, STRA6 in endothelial cells, and
CYP26B1 in bronchiolar epithelium.
Overall, these studies have shown the great ability of RA in promoting lung RE
formation, even when given in much diluted concentration. Compared with a single dose,
multiple treatments of VARA produced a cumulative effect on RE storage. The
synergistic effect of VARA was not significantly affected by inflammation. These results
together with our findings of the localization of retinoid homeostatic proteins provide a
better understanding of retinoid uptake, accumulation and metabolism in the neonatal
lung. Our findings also suggest a promising therapeutic approach in clinical use for a
rapid restoration of lung VA in preterm or VLBW infants to promote normal lung
maturation and prevent these infants from developing respiratory diseases.

vi
TABLE OF CONTENTS
LIST OF ABBREVIATIONS......................................................................................x
LIST OF FIGURES .....................................................................................................xiii
ACKNOWLEDGEMENTS.........................................................................................xvi
Chapter 1 LITERATURE REVIEW...........................................................................1
1.1 LUNG PROBLEMS IN PRETERM NEONATES .......................................1
1.2 LUNG STRUCTURE AND DEVELOPMENT ...........................................2
1.2.1 Lung structure and function..................................................................2
1.2.2 Lung development ................................................................................6
1.2.3 Lung cells .............................................................................................8
1.3 GENERAL INTRODUCTION TO VITAMIN A.........................................9
1.3.1 Functions and properties of vitamin A .................................................9
1.3.2 Vitamin A deficiency and toxicity .......................................................12
1.4 VITAMIN A METABOLISM ......................................................................13
1.4.1 Transport and metabolism of retinoids.................................................13
1.4.2 Retinoid homeostatic proteins ..............................................................19
1.5 THE REGULATORY MECHANISM OF RETINOIDS..............................22

vii
1.6 RETINOIDS IN LUNG DEVELOPMENT AND FUNCTION ...................24
1.6.1 Retinoids in lung morphogenesis .........................................................24
1.6.2 Retinoids in alveolar septation .............................................................25
1.6.3 Retinoids in lung tissue repair ..............................................................27
1.6.4 Retinoid metabolism in the lung...........................................................30
Chapter 2 MULTIPLE TREATMENT STUDY.........................................................32
2.1 ABSTRACT ..................................................................................................32
2.2 INTRODUCTION.........................................................................................33
2.3 HYPOTHESIS AND AIMS..........................................................................35
2.4 MATERIALS AND METHODS ..................................................................36
2.5 RESULTS......................................................................................................42
2.6 DISCUSSION................................................................................................52
Chapter 3 LPS-INDUCED INFLAMMATION STUDY...........................................56
3.1 ABSTRACT ..................................................................................................56
3.2 INTRODUCTION.........................................................................................57
3.3 HYPOTHESIS AND AIMS..........................................................................60
3.4 MATERIALS AND METHODS ..................................................................61
3.5 RESULTS......................................................................................................66
3.6 DISCUSSION................................................................................................77
Chapter 4 ACIDIC RETINOIDS DILUTION STUDY..............................................81

viii
4.1 ABSTRACT ..................................................................................................81
4.2 INTRODUCTION.........................................................................................82
4.3 HYPOTHESIS AND AIMS..........................................................................83
4.4 MATERIALS AND METHODS ..................................................................85
4.5 RESULTS......................................................................................................89
4.6 DISCUSSION................................................................................................95
Chapter 5 LOCALIZATION OF LUNG RETINOID HOMEOSTATIC
PROTEIN .............................................................................................................98
5.1 ABSTRACT ..................................................................................................98
5.2 INTRODUCTION.........................................................................................99
5.3 HYPOTHESIS AND AIMS..........................................................................102
5.4 MATERIALS AND METHODS ..................................................................103
5.5 RESULTS......................................................................................................106
5.6 DISCUSSION................................................................................................114
Chapter 6 DISCUSSION ............................................................................................117
6.1 VITAMIN A SUPPLEMENTATION DURING LUNG SEPTATION
PERIOD.........................................................................................................117
6.2 MOLECULAR MECHANISM OF RETINOL UPTAKE INTO THE
LUNG............................................................................................................121
6.3 THE POTENTIAL OF RA TO PROMOTE LUNG RE FORMATION ......122

ix
6.4 THE RELATIONSHIP BETWEEN INFLAMMATION AND
RETINOID METABOLISM IN NEONATAL LUNGS ..............................124
6.5 SPATIAL DISTRIBUTION OF THE RETINOID HOMEOSTATIC
PROTEINS IN NEONATAL LUNGS..........................................................125
6.6 FUTURE DIRECTIONS...............................................................................129
REFERENCES ............................................................................................................132

x
LIST OF ABBREVIATIONS
9cRA: 9-cis-retinoic acid
ADH: alcohol dehydrogenase
ADRP: adipocyte differentiation-related protein,
ALDH: aldehyde dehydrogenase
at-RA: all-trans-retinoic acid
ANOVA: analysis of variance
BPD: bronchopulmonary dysplasia
CCL2: chemokine (C-C motif) ligand 2
CRABP: cellular RA-binding protein
CRBP: cellular retinol-binding protein
CRP: c-creative protein
CYP: cytochrome P450
DAPI: 4’,6’-diamidino-2-phenylindole
ECM: extracellular matrix
h: hour(s)
HPLC: high performance liquid chromatography
IGF2R: insulin-like growth factor II receptor
IHC: immunohistochemistry

xi
IL-6: interleukin-6
ISH: in situ hybridization
IU: international unit
LPL: lipoprotein lipase
LPS: lipopolysaccharide
LRAT: lecithin:retinol acyltransferase
M6P: mannose-6-phosphate
PECAM: platelet endothelial cell adhesion molecule
RA: retinoic acid
RAE: Retinol Activity Equivalents, a term referring to vitamin A activity
RALDH: Retinaldehyde dehydrogenase
RAR: retinoic acid receptor
RARE: retinoic acid response element
RBP: retinol binding protein
RDH: retinol dehydrogenase
RDS: respiratory distress syndrome
RE: retinyl ester
rt-PCR: real-time polymerase chain reaction
RXR: retinoic X receptor

xii
RXRE: retinoid X-response elements
ROH: retinol
RXR: retinoid X receptor
SPA1: surfactant protein A1
STRA6: stimulated by retinoic acid gene 6
TTR: transthyretin
VA: vitamin A
VAD: vitamin A deficiency
VAS: vitamin A sufficiency
VEGF: vascular endothelial growth factors
VARA: vitamin A combined with retinoic acid
VLBW: very-low-body weight
VLDL: very-low-density lipoprotein

xiii
LIST OF FIGURES
Figure 1.1 Cross-section of alveolar wall and major cell types. . ................................5
Figure 1.2 Common natural and synthetic retinoids....................................................11
Figure 1.3 Schematic overview for the transport and uptake of dietrary retinoids
within the body. ....................................................................................................15
Figure 1.4 Uptake of retinoids into extrahepatic tissues..............................................17
Figure 1.5 Regulatory role of RA in retinoid metabolism. ..........................................21
Figure 1.6 Cellular retinoid metabolism and signaling pathway. ................................23
Figure 2.1 Animal experimental design.......................................................................39
Figure 2.2 Lung RE concentration after a single treatment or multiple treatments.....44
Figure 2.3 Expression level of Lung LRAT gene after treatments..............................47
Figure 2.4 Expression level of lung CYP26B1 gene after treatments. ........................48
Figure 2.5 Expression level of lung STRA6 genes after treatments............................49
Figure 2.6 Expression level of lung functional and structural genes after
treatments..............................................................................................................51
Figure 3.1 Animal experimental design.......................................................................63

xiv
Figure 3.2 Lung RE concentration after a single treatment with oil or VARA in
the presence or absence of LPS induced inflammation state................................68
Figure 3.3 Expression level of lung IL6 gene 6 h or 18 h after LPS administration. ..70
Figure 3.4 Expression level of lung CCL2 gene 6 h or 18 h after LPS
administration. ......................................................................................................71
Figure 3.5 Plasma C Reactive Proteins (CRP) level 6 h or 18 h after LPS
administration. ......................................................................................................72
Figure 3.6 Expression level of lung LRAT gene 6 h after VARA treatment. .............74
Figure 3.7 Expression level of lung CYP26B1 gene 6 h after VARA treatment.. ......75
Figure 3.8 Expression level of lung STRA6 gene 6 h after VARA treatment.............76
Figure 4.1 Animal experimental design.......................................................................87
Figure 4.2 Lung RE concentration after a single treatment with VA alone, VARA
10%, VARA 5%, VARA 2%, VARA1% and VARA0.5%..................................92
Figure 4.3 Lung RE concentration after multiple treatments with VA alone,
VARA10%, VARA2% and VARA1%.................................................................93
Figure 4.4 Lung RE concentration after multiple treatments with VA alone,
VAAm10%, VAAm2% and VAAm1%. ............................................................94
Figure 5.1 Localization of LRAT protein in 14d old rat lungs....................................109

xv
Figure 5.2 Localization of STRA6 protein in 14d old rat lungs. .................................110
Figure 5.3 Localization of CYP26B1 protein in 14d old rat lungs..............................113
Figure 6.1 Model of VA metabolism in neonatal rat lung...........................................128

xvi
ACKNOWLEDGEMENTS
First and foremost, I would like to express my sincere gratitude to my supervisor Dr. A.
Catharine Ross, whose guidance, encouragement, patience and support helped me in all
the time of research and enabled me to complete my project and writing of this thesis.
I am grateful to my committee members: Dr. Okhee Han, Dr. Katarzyna Kordas, Dr.
Pamela J. Mitchell, and Dr. Jeffery M. Dodds for sharing their expertise and providing
invaluable insights during my research and writing.
I am also thankful to my colleagues: Dr. Qiuyan Chen and Dr. Reza Zolfaghari for their
great encouragement and advice on my graduate research; Yao Zhang, Libo Tan,
Katherine Restori, Amanda Wray, Dr. Kyoko Goto, Dr. Nan-qian Li, and Madeline Stull
for their friendship and assistance in my daily work and life.
I would like to thank the Department of Nutritional Sciences, which likes a big and warm
family to students. For many years, I have got continued help from our department staff,
especially Judy Jones.
Last, but not least, I offer my gratitude and blessings to my husband, Weimin Wu and my
family for their constant support and love. Without your support, I could never have gone
this far and completed my graduate work.

1
Chapter 1
LITERATURE REVIEW
Every year, around half a million infants are born prematurely in the United States. The
human fetal lung normally is not clinically mature until after approximately 35 wk
gestation. Although many premature infants appear healthy, it has been observed that
they may suffer from interrupted lung development, underdeveloped lungs at birth, and
immature immune systems. Due to the physiological and morphological immaturity, the
lung of premature infants is functional insufficient and these infants are at increased risk
of developing various pulmonary diseases during the early postnatal life, such as
respiratory distress syndrome (RDS) and subsequent ventilatory management-caused
bronchopulmonary dysplasia (BPD). The respiratory problems are the leading cause of
the morbidity in preterm neonates. Therefore, premature infant lung development is one
of the largest issues in clinical therapy for premature infants.
1.1 LUNG PROBLEMS IN PRETERM NEONATES

2
The lungs of mammals are among of the largest organs in the body and are the major part
of the respiratory system. They are cone-shaped and have a spongy and soft texture
because of the composition of millions of alveoli, the functional unit of the lung. The
principal function of the lungs is to bring oxygen (O2) into the body, and to remove
carbon dioxide (CO2) out of the body.
In the body of mammals, the distal end of trachea divides into two bronchi which lead
into the left and right lungs, respectively. Each bronchi then starts a series of branches,
called the respiratory tree. It divides into secondary bronchi, tertiary bronchi, and after
multiple divisions, reaches to the level of bronchioles. While the process of branching
continues, bronchioles branch into the terminal bronchioles, the respiratory bronchioles,
alveolar ducts, and finally, give rise to the clusters of alveolar sacs. Each alveolar sac is
tightly wrapped by a dense network of capillaries. In human, the structure of alveoli
increases the surface of the lung to 80-140 m2, depending on body size.
According to the function and properties of each part, the lungs are divided into two
primary zones - the conducting zone and respiratory zone. The conducting zone is
composed of the trachea, bronchi, bronchioles, and terminal bronchioles. In the trachea
1.2 LUNG STRUCTURE AND DEVELOPMENT
1.2.1 Lung structure and function

3
and the upper levels of bronchi, hyaline cartilage is present in a C-ring shape around the
wall of the airway. As the bronchi divide into smaller and smaller passageways, the
amount of hyaline cartilage decreases and the cartilage ring gradually becomes irregular,
discontinued, and finally disappeared and replaced by smooth muscle in the smallest
bronchioles. As the cartilage decreases and the smooth muscle increases, the mucous
membrane also undergoes a transition from ciliated pseudostratified columnar epithelium
in the upper respiratory tract to a simple cuboidal epithelium in the terminal bronchioles.
Throughout the mucosa, goblet cells are present to provide continued mucus secretion.
The conducting zone mainly functions as passageways that deliver inspired air to the
respiratory zone and expired air to the outside environment. It also acts to warm and
humidify the inhaled air and to provide an immunological defense against intruding
organisms.
The respiratory zone is the site of gas exchange. It is made up of respiratory bronchioles,
alveolar ducts and alveoli which only have a thin wall which is primarily made up of
simple squamous epithelial cells. Each alveolar sac is surrounded by a capillary network.
The thin layer of type I epithelial cells, the basal membrane, and the thin layer of
capillary endothelial cells constitute the air-blood barrier (also named as respiratory
membrane) which permits a rapid diffusion of oxygen and carbon dioxide. The structure
of alveolar dramatically increases the surface area for contact with blood vessels. The
alveolar epithelium is simply comprised of type I alveolar cells, type II alveolar cells,
endothelial cells, macrophages and fibroblast (Fig. 1.1). Type I alveolar cells are simple
squamous cells that account for most of the surface wall, although there are just about

4
half as many as type II cells. Type I cells are large and thin, while type II alveolar cells
are small and round, and compose only about 10 % of the alveolar wall. The type II cell
is responsible for the production and secretion of surfactant, which reduces surface
tension and prevents the collapse of alveoli. When type I cell is damaged, the type II cell
is able to differentiate into a type I cell to replace it. Macrophages, derived from blood
monocytes, are also present in the alveolar space to phagocytize the invading bacteria.

5
Figure 1.1 Cross-section of alveolar wall and major cell types. This figure is from
internet resource and is modified by L. Wu.

6
In humans, the process of lung development can be subdivided into five distinct phases:
embryonic phase, pseudoglandular phase, canalicular phase, saccular phase and alveolar
phase (1).
Embryonic phase/ early branching phase
This phase of lung development takes place during weeks 4 and 5 of gestation. The lung
bud originates from endodermal epithelium and grows out from the lower pharynx. It
then subdivides into two main bronchi and begins dichotomous branching into smaller
segmental bronchi. Smooth muscle, airway cartilage, blood and lymph vessels start to
develop.
Pseudoglandular phase
In this stage (4-17 weeks of gestation), the lung resembles an exocrine gland, hence the
name. The major conducting airway continues to develop into a bronchial tree, paralleled
by the formation of a vascular bed. Under the influence of adjacent mesenchyme, the
primitive airway epithelial cells start to differentiate into ciliated cells, goblet cells and
mucous glands, while mesenchymal cells have begun to form cartilage and smooth
muscle cells (2). Type II alveolar cells appear in respiratory bronchioles and start to
produce amniotic fluid.
1.2.2 Lung development

7
Canalicular phase
In this stage (17-26 weeks of gestation), terminal airspaces expand to form primitive
alveoli. Surfactant protein is detectable by 24 weeks of gestation. During this period,
respiratory bronchioli appear, interstitial tissue decreases, the cuboidal epithelium starts
to differentiate into type I and type II alveolar cells, and the blood capillaries proliferate
around the alveoli to form air-blood barrier for limited gas exchange (3). Thus, fetus is
able to survive at around the mid-late canalicular stage (4).
Saccular phase
In the saccular stage (24-36 weeks of gestation), the distal airspaces continue to enlarge,
accompanied with a continued reduction of interstitial tissues. The air sacs are mainly
lined with flattened type I epithelial cells and rounded type II epithelial cells. The
epithelial layer becomes thinner. As a result of the structural change, the gas-exchange
function is enhanced. The primary septa between two alveolar sacculi are still thick and
contain a double capillary network. During this time, surfactant containing laminar
bodies lining in type II pneumocytes can be found. However, the lung of rodents is in this
phase at birth, which is equal to human fetal lung at 28 wks.
Alveolar phase/ septation period
The last stage, alveolar stage, begins approximately 36 week of gestation to term and
continues for at least 3 year of postnatal life. At birth, the lung is still structurally
immature, only 15-20% of alveoli in the adult lung are formed, indicating most of the
alveoli are formed within the postnatal period (5, 6). In rodents (rats and mice), this stage

8
occurs exclusively between day 4 and 14 after birth (7). The alveolar stage is mainly
characterized by repetitive subdivisions of terminal sacs into definitive alveoli by the
growth of secondary septa. In the process of subdivision, secondary septa extend from
primary alveolar wall and subdivide the terminal sacs into multiple smaller and thinner
definitive alveoli. Development of these septae occurs through deposition of new
basement membrane, outgrowth of epithelial cells and myofibroblasts at the tips of
septae, and elastin deposition (5). The epithelial cells lining the alveolar wall decrease in
number and become thinner and flatter. Accompanying the aiveolar septation is the
microvascular maturation. The capillary vessels come into close contact with the alveoli
and the basement membranes underlying the capillary endothelial cells and alveolar
epithelial cells fuse with each other, forming a very thin air-blood barrier. In contrast to
the double capillary in primary septa, the secondary septa contain only a single layer of
capillary. The continued subdivisions of alveoli greatly increases the lung surface area for
gas exchange, thus, the process is also known as alveolarization (8).
The mature lung is a heterogeneous tissue comprised of about 40 morphologically
distinct cell types (9). This cell population including cells in the epithelium, cells in the
connective tissue, blood vessels and nervous tissue, such as squamous epithelial, ciliated
columnar cells, mucus-secreting goblet cells, and clara cells lining in the upper airways,
type I and type II alveolar epithelial cells lining in alveoli; fibroblasts, myofibroblasts,
and smooth muscle cells (SMC) in the lung interstitium, endothelial cells in blood
1.2.3 Lung cells

9
vessels, macrophages and lymphocytes, migrating between circulation system and tissue,
etc. These cells serve various roles in lung development and functions.
Vitamin A (VA) is a group of compounds that play an important role in many functions.
The major functional activities of VA include 1) promoting vision (10), 2) participating
in protein synthesis and cell differentiation in epithelial tissues and skin (11, 12), 3)
supporting reproduction, development and growth (13), and 4) maintaining the integrity
of immunity (14, 15). The group of VA compounds is referred to as retinoids and is
comprised of a large number of natural and synthetic compounds (Fig. 1.2). The natural
compounds include retinol and its metabolites: RE, retinal, retinoic acid (RA) and some
water-soluble metabolites. The basic structure of a retinoid is composed of a substituted
cyclohexenyl ring, a tetraene side chain and a functional group at the end of the side
chain. Retinol (also named vitamin A) has a hydroxyl group at its terminal end. When
this group is esterified with a fatty acid, retinol becomes RE, the form in which VA is
stored. The hydroxyl group also undergoes oxidation to produce an aldehyde (retinal),
which may be further oxidized to a carboxylic acid (retinoic acid). Among these
compounds, RA is the most active metabolite of vitamin A, which interacts with RA
nuclear receptors and subsequently modulates proliferation of epithelial cells, pattern
1.3 GENERAL INTRODUCTION TO VITAMIN A
1.3.1 Functions and properties of vitamin A

10
formation in developing tissues, morphogenesis in the lung, and cellular differentiation.
In addition to these naturally occurring retinoids, a large number of artificial analogs have
been synthesized and used in studies of retinoid signaling within the cell. For example,
Am580, a stable RA analog and a selective agonist of retinoic acid receptor-α (RARα),
belongs to the retinoid family as well (16). The property of metabolism resistance of
Am580 is due to the two methyl groups at the C4 position which prevent the access of
RA metabolism enzyme, thus protect Am580 from being rapidly catabolized.

11
Figure 1.2 Common natural and synthetic retinoids. (a) All-trans-retinol; (b) All-
trans-retinoic acid; (c) Retinyl ester; (d) Retinal; (e) 9-cis-retinoic acid; (f) 13-cis-
retinoic acid; (g) Am580, (an RA analog).

12
Vitamin A deficiency is a major public health problem among young children and, to a
lesser extent, pregnant and breastfeeding women, in most developing countries. It is a
major cause of mortality and severe morbidity in children (17). It is estimated that 140–
250 million children under five years of age are affected by VA deficiency worldwide
(18). Vitamin A deficiency can be caused by inadequate intake, fat malabsorption, or
liver disorders. Deficiency can result in impaired immunity, skin keratinization,
metaplasia, poor growth and typical ocular abnormalities (e.g., xerophthalmia, night
blindness), etc. (19). VA deficiency increases the risk of infection, diarrhea and
developing respiratory disease in children (20). The traditional means of prevention are
supplementation with high-dose vitamin A, for example, infants < 6 mo can be given a
one-time dose of 50,000 IU, and those aged 6 to 12 mo can be given a one-time dose of
30,000 RAE (100,000 IU). For pregnant or breastfeeding women, therapeutic doses
should not exceed 10,000 IU /day to avoid possible damage to the fetus or infant.
On the other hand, just as a deficiency of VA affects all body systems, so does an
overabundance. The manifestations of VA toxicity include headache, vomiting, diplopia,
alopecia, dryness of the mucous membranes, bone and joint pain, liver damage and coma
(21). In very young children treated with 50,000 to 100,000 IU vitamin A, the most
frequently observed symptoms are a temporary bulging of the fontanel and vomiting (22,
23). Vitamin A also has teratogenic properties. High intakes of VA by pregnant women
1.3.2 Vitamin A deficiency and toxicity

13
may increase the incidence of teratogenic effects in the developing fetus (24), and
experiments in pregnant animals have demonstrated that excess VA given to the mothers
can result in increased birth defects in their young (25, 26) Thus, maintaining an adequate
but not excessive amount of VA in the body is vitally important.
Dietary VA is obtained mainly in the form of RE and β-carotene. When ingested, VA is
first converted to retinol in the lumen of the intestine, then re-esterified to RE in the
enterocytes, packaged into newly formed chylomicrons for delivery and stored in the
liver as RE (27). As the center of VA storage, the liver stores approximately 50-80% of
the body’s total VA. To transport VA to extrahepatic tissues, stored RE is mobilized to
produce free retinol which then binds to a liver synthesized transport protein, retinol-
binding protein (RBP). Once bound by RBP, the retinol-RBP complex is secreted into the
bloodstream for transport to peripheral tissues (28, 29). Release of retinol-RBP complex
is determined by the rate of RBP synthesis. Meanwhile, the secretion of retinol-RBP is
also highly regulated by VA status, such that VA deficiency blocks retinol-RBP secretion
(29, 30). When it circulates in the bloodstream, the retinol-RBP complex binds to a serum
carrier, transthyretin (TTR), which is believed to prevent elimination of the relatively
small RBP molecule through the kidney and thereby maintain normal levels of retinol in
1.4 VITAMIN A METABOLISM
1.4.1 Transport and metabolism of retinoids

14
the circulation (29, 31, 32) (Fig. 1.3). It has been thought that retinol enters cell through
passive transport or through a specific cell receptor. Recently, a cell surface protein,
STRA6, has been identified as an RBP receptor, which mediates cellular uptake of retinol
(33) (Fig. 1.3).
Although most of the retinoid compounds found in the circulation are in the form of
retinol bound to RBP, there are small amount of RA and RE circulating in the blood
stream and these are taken up by unknown mechanism (Fig. 1.4) (34). Dietary RA, or RA
produced by metabolism of dietary RE in the small intestine, can be absorbed via the
portal system and then circulate in the plasma bound to albumin (28, 35). A study
investigating the uptake and metabolism of all-trans-[3H]retinoic acid by human foreskin
keratinocytes suggested that the binding of RA to albumin protects RA from conversion
to polar metabolites, and controls delivery of RA from the aqueous extracellular
environment to the cell surface (36). It has been reported that RA does not require a cell
surface receptor for uptake because it is able to traverse the cellular membrane and enter
the cell efficiently (37, 38). However, studies by Kang et al. (39) revealed that a
membrane receptor, mannose-6-phosphate/insulin-like growth factor II receptor
(M6P/IGF2R) might be involved in mediating RA-response pathway and cellular activity
in cells. Moreover, the liver and other tissues do not store RA, and the pool of RA turns
over rapidly (40).
15
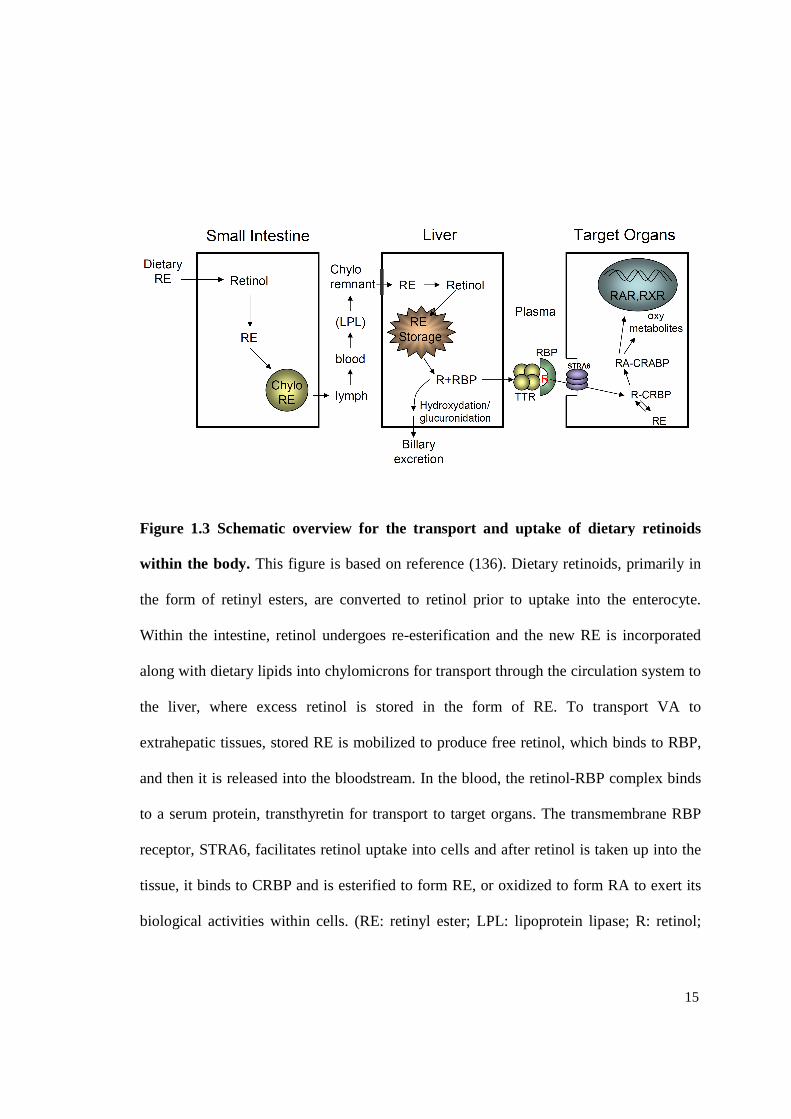
Figure 1.3 Schematic overview for the transport and uptake of dietary retinoids
within the body. This figure is based on reference (136). Dietary retinoids, primarily in
the form of retinyl esters, are converted to retinol prior to uptake into the enterocyte.
Within the intestine, retinol undergoes re-esterification and the new RE is incorporated
along with dietary lipids into chylomicrons for transport through the circulation system to
the liver, where excess retinol is stored in the form of RE. To transport VA to
extrahepatic tissues, stored RE is mobilized to produce free retinol, which binds to RBP,
and then it is released into the bloodstream. In the blood, the retinol-RBP complex binds
to a serum protein, transthyretin for transport to target organs. The transmembrane RBP
receptor, STRA6, facilitates retinol uptake into cells and after retinol is taken up into the
tissue, it binds to CRBP and is esterified to form RE, or oxidized to form RA to exert its
biological activities within cells. (RE: retinyl ester; LPL: lipoprotein lipase; R: retinol;

16
RA: retinoic acid; RBP: retinol binding protein; TTR: transthyretin; Chylo: chylomicron;
CRBP: cellular retinol-binding protein; CRABP: cellular RA-binding protein; RAR:
retinoic acid receptor; RXR: retinoid X receptor)

17
Figure 1.4 Uptake of retinoids into extrahepatic tissues. This figure is based on
reference (34). Retinoids in the circulation are present in several forms, including retinol
bound to RBP, RA bound to albumin, and RE contained in lipoproteins (primarily
chylomicrons). The transmembrane protein STRA6, which is identified as a RBP
receptor, can mediate retinol uptake into cells. The mechanisms that mediate cellular
uptake of RE and RA are not fully understood. However, a possible mechanism for RE
uptake in certain tissues has been established that RE is hydrolyzed to retinol by LPL
before it is taken up by tissues and cells.

18
It has been established that ~75% of dietary RE is taken up by the liver, while ~25% RE
is taken up by extrahepatic tissues, including adipose tissue, skeletal muscle, heart, lungs
and kidneys (41, 42). A model of mice lacking RBP indicated that although the VA
stored in the liver cannot be mobilized, RBP-deficient mice are still able to maintain a
normal phenotype (43). A further investigation on RBP-/- mice revealed that a high
concentration of RE in the circulating chylomicron/VLDL meets tissue retinoid
requirements, thus compensating for the absence of retinol-RBP (44). The physiological
importance of postprandial RE is unclear in wild-type animals, but it might be an
important factor in maintaining VA homeostasis in some specific tissues, such as the
lung, which might require a direct delivery of retinoids to accumulate VA stores (45).
There is little known about the cellular uptake of RE as well. It has been hypothesized
that lipoprotein lipase (LPL) plays a role in facilitating cellular uptake of RE, because
LPL is able to catalyze the hydrolysis of chylomicron RE (46). However, LPL-facilitated
chylomicron RE uptake is only observed in skeletal muscle, heart, and adipose tissue,
while other tissues, such as the kidneys and lungs, take up RE independent of LPL
manipulations. This suggests that more than one mechanism exists to mediate the uptake
of chylomicron RE in the extra-hepatic tissues.
Within the cytoplasm of tissue cells, retinol and its oxidized form RA bind with cellular
retinol-binding proteins (CRBP) and cellular retinoic acid-binding proteins (CRABP)
(47), which are believed to regulate the biological action of retinol and RA (48) (Fig. 1,4;
1.6). By binding with these retinoid-binding proteins, the concentration of free cellular
retinoid is limited, and the bound retinoid is directed to specific enzymes for metabolic

19
processing (49). Retinol bound to CRBP is directed to the enzyme lecithin: retinol
acyltransferase (LRAT) for esterification and thus is turned into the storage form RE (50,
51). CRBP-bound retinol is also a substrate for members of the alcohol dehydrogenase
family (ADH) (52), which catalyzes oxidization of retinol to retinal. Retinal is further
irreversibly oxidized to RA by aldehyde dehydrogenase (ALDH) and cytochrome P450
enzyme families (52, 53). CRABP has the similar function of mediating intracellular RA
concentration and directing RA to the enzymes, which metabolizes RA into polar inactive
metabolites, such as 4-oxo-RA, 4-OH-RA, 18-OH-RA and 5,18-epoxy-RA, etc (51, 54).
Thus, the biological effects of excess RA are limited.
Along the pathway of VA metabolism, two important enzymes play prominent roles in
regulating retinoid homeostasis (55). One is LRAT which catalyzes the esterification of
retinol; the other is a cytochrome P450, CYP26, which mediates oxidation of RA (Fig.
1.5). Both of these enzymes are tightly regulated by RA in a tissue-specific manner,
especially in the liver and lung (56). Also, the level of these enzymes is affected by VA
status, for example, VA deficiency down-regulates the expression level of LRAT (57)
and CYP26 (58, 59) to maintain the level of free retinol, while VA sufficiency increase
LRAT (56) and CYP26 (60) level to prevent retinol excess. This is because the level of
RA signal that regulates gene expression is determined by the status of VA. The response
1.4.2 Retinoid homeostatic proteins

20
of LRAT and CYP26 to the RA signal provides a self-regulatory mechanism that
regulates retinol homeostasis to avoid both VA deficiency and toxicity.
Another protein that may contribute to retinoid homeostasis is STRA6, a cell surface
receptor for RBP (Fig. 1.4). This transmembrane protein shows a high affinity for RBP
and mediates cellular uptake of retinol (33). Clinical cases suggest that mutations in the
STRA6 gene result in Matthew-Wood syndrome, which is characterized by multisystem
malformations that include lung hypoplasia (61). When STRA6 and LRAT are co-
expressed, the cells take up retinol more efficiently, suggesting a driving force for retinol
uptake resulting from the conversion of retinol into RE by LRAT (33, 62). Several in
vitro and in vitro studies have shown that STRA6 expression can be upregulated by RA
in several cell lines (63-65), and supplementation with retinoids is able to elevate STRA6
expression in mouse embryos or in the lungs of neonatal rats (66, 67), indicating the
regulation of STRA6 by RA. However, it is not clear whether STRA6 mediated retinol
uptake is the only mechanism facilitating retinoid uptake. As discussed before, the
chylomicron derived RE could be another important source of retinoid for some extra-
hepatic tissues.

21
Adapted from Ross, A.C. J Nutr, 1993
Figure 1.5 Regulatory role of RA in retinoid metabolism. This figure is based on
reference (68). The retinoid homeostasis is tightly regulated by RA through a gene
regulation mechanism. Expression of the LRAT and CYP26B1 mRNA are induced by
RA, or by dietary vitamin A, and are downregulated by VA depletion. STRA6 expression
may also be upregulated in response to RA for facilitating retinol uptake into the cell.

22
Vitamin A is a fat-soluble vitamin that acts like a hormone in the body (49). It functions
as a gene regulator by binding to a series of nuclear receptor proteins that belong to the
steroid/thyroid hormone receptor superfamily (69, 70) (Fig. 1.6). These retinoid receptors
are classified into two families, the RA receptors (RARs), and the retinoid X receptors
(RXRs), each containing three isoforms (α, β, and γ) (70, 71). Upon RA binding, the
RARs are activated, form homodimers of RAR/RAR, or heterodimers of RAR/RXR with
RXR, which then interacts with specific regions of DNA termed retinoid responsive
elements (RARE or RXRE) to activate or repress the transcription of downstream target
genes (70). It is believed that both all-trans and 9-cis RA bind to RAR, but all-trans RA
bind to RAR specifically, whereas 9-cis RA binds to RXR with high affinity (72). Via
this mechanism, RA is able to affect transcription of many genes. Several artificial
retinoids with specific RAR or RXR binding selectivity have been synthesized to be used
in studies of retinoid signaling within cells. For example, Am580, a retinobenzoic acid
analog of RA, which activates RARα selectively and shows resistance to metabolism (16,
73), has been used in our previous study to test the interaction of VA and acidic retinoids
on lung RE formation (67). Due to the prolonged regulating activity on retinoid
homeostatic genes, Am580 can alter neonatal lung retinoid metabolism dramatically (67).
1.5 THE REGULATORY MECHANISM OF RETINOIDS

23
Figure 1.6 Cellular retinoid metabolism and signaling pathway. This figure is based
on references (56, 68, 74). Upon entering the cell, retinol is bound by CRBP, and directed
to LRAT for esterification or directed to alcohol dehydrogenase (ADH), which catalyzes
oxidation of retinol to retinal. Retinoic acid taken into the cell or synthesized from retinal
is bound by CRABP. CRABP-RA is either delivered to CYP26 for further oxidation or
transferred to nuclear. Once in the nuclear, RA activates RAR which forms a heterodimer
with RXR, or forms a homodimer, and then interacts with specific regions of RARE or
RXRE to activate or repress the transcription of downstream target genes.

24
1.6 RETINOIDS IN LUNG DEVELOPMENT AND FUNCTION
The earliest evidence of VA’s importance in the lung can be dated back decades. In
1917, McCollum, who discovered VA, first observed that animals fed VA deficient diet
frequently suffered from prevalent bronchitis (75). Several years later, Bloch, a
pediatrician, supplemented food rich in VA to VA deficient (VAD) infants and young
children, and found the symptoms of bronchopneumonia caused by VAD could be
rescued by the VA rich diet (76). In 1933, Wolbach and Howe (77) observed that the
mucous-secreting epithelium in the trachea and the bronchi of VAD rats was replaced by
stratified squamous keratinizing epithelium, which further confirmed that the respiratory
system is a target for retinol action The morphological alteration in the airway of VAD
rats can be recovered when the dietary retinol is restored. These findings suggest that
retinoids play an important role in regulating differentiation of respiratory epithelium and
this speculation was demonstrated by a number of studies (78-81). As a biologically
active metabolite of retinoid, RA has shown stronger action on lung development than
retinol, although RA is not stored in tissues and is eliminated rapidly (53, 82).
RA signaling is detectable in the lung as early as the very beginning of organogenesis
(83). Absence of RA signaling in the early time of embryonic stage results in impaired
primary lung bud formation and branching morphogenesis (84). During the early stages
1.6.1 Retinoids in lung morphogenesis

25
of lung development, the major RA synthesizing enzyme retinaldehyde dehydrogenase 2
(RALDH2) and RA nuclear receptors (RARs, RXRs) are expressed in a specific pattern
in fetal lung. Mice with compound mutants of the RA receptors showed dramatic
abnormalities in lung phenotype (85-87). Disruption of the mouse RALDH2 gene also
generated severe defects similar to those described in vitamin A-deprivation studies (88).
In the process of airway branching, RA activity needs to be down-regulated to allow the
normal growth of distal epithelial bud. If up-regulated, the branching is inhibited and
lung structure resembles proximal airways (89, 90). When RA signaling is blocked by a
pan-RAR antagonist, the lung branching is increased (90, 91). These findings suggest the
involvement of RA in morphogenesis and in early embryonic development.
Lung septation stage occurs from later gestation until the first few postnatal years in
human. Studies in neonatal rat lung indicate that an acute and significant accumulation
and utilization of RE occurs exclusively during alveolar stage (92, 93). Concurrently,
there is an increase in active forms of retinol and RA in the lung fibroblasts, indicating an
increased demand of retinoids for normal postnatal development of the lung (94).
Associated with dramatic changes in the retinoid homeostasis is the upregulation of
cellular retinoids binding proteins (CRBP, CRABP), retinoid synthesizing enzymes
(ALDH-1, RALDH-2) and specific RA receptors (RARs) during perinatal development
(47, 94-96). After birth, the levels and binding activity of CRABP to RA rise strikingly in
1.6.2 Retinoids in alveolar septation

26
neonatal rat lung and then declines to low level until the septation process is completed
(47, 97). These findings provide evidence that RA is indeed required for alveologenesis.
It seems likely that the effects of VA on lung development, differentiation and
maintenance through the regulation of many genes that related to lung development,
including those involved in patterning, matrix proteins and certain growth factors, etc
(98), and the action of VA is through its interaction with its nuclear receptors which in
turn modulate the transcription of target genes. Studies investigating the relationship
between retinoids and lung development have shown that deletion of RA receptors RAR-
γ and RXR-α, the key nuclear receptors through which RA induces the formation of
alveoli (97, 99, 100), leads to marked failure of septation that associated with reduced
alveolar numbers and elastin production (101). In contrast to RAR-γ and RXR-α, RAR-β
is an endogenous inhibitor of septation during but not after the period of septation, since
RAR-β activation could block septation while its deletion result in early onset of
septation in mice lung (102). RA also has shown its activity in upregulating expression of
lung elastin gene in lung fibroblast (103) and administration of RA to the neonatal rats
resulted in partial recovery of the septation process and formation of alveoli (97, 99).
Inhibition of the RA synthetic enzyme, ALDH, disrupts tropoelastin mRNA and
decreases elastin levels (103).
Premature infants, associated with very low body weight (VLBW), tend to have VA
deficiency, probably because of a shortened period of transplacental VA supply resulting
from premature birth (104-106). The earlier a child is born before due date, the lower his
serum-retinol levels are (107), and this population often has a high risk to develop

27
various pulmonary diseases. In several clinical trials, VA supplements showed beneficial
effects in elevating VA status and preventing BPD in VLBW infants (108, 109).
In clinical therapy, dexamethasone, a typical synthetic glucocorticosteroid hormone, is
commonly used in the postnatal treatment of premature and VLBW infants with BPD to
prevent inflammation and stimulate the lung maturation and the production of surfactant.
Although a short-term improvement in lung function is seen in many infants, side effects
of dexamethasone have been reported in multiple organs in preterm infants (110). Animal
studies have shown that daily dexamethasone treatment to neonatal rats during the
postnatal period resulted in irreversibly impairment in saccule septation (111, 112). The
possible reasons that glucocorticosteroids inhibit the lung septation might due to its
inhibition in DNA synthesis and cell division while the process of the lung septation
needs septa formation, elongation, capillaries and fibroblasts filling (113-115). This
thought is strengthened because that serum glucocorticosteroid concentration was found
to be low during the period of septation (116), and starts to increase as septation ends (7,
117), which suggests the increased concentration of the hormone initiates the end of
septation. Interestingly, it was reported that postnatal dexamethasone treatment is able to
increase plasma VA and RBP significantly in newborn infants (118), which suggest that
dexamethasone could stimulate liver retinol mobilization and retinol-RBP release. in
response to dexamethasone.
1.6.3 Retinoids in lung tissue repair

28
In addition to dexamethasone treatment, preterm infants often receive treatments of
oxygen supplementation and mechanical ventilation support after birth since their lungs
are functional inefficient. But these treatments carry many potential complications
including airway injury, alveolar damage, and ventilator-associated inflammation because
the lung is exposed to hyperoxia. Hypoxia is known to be a major stimulator of VEGF
expression (119, 120) because it can activate the transcription factors hypoxia-inducible
factor 1 (HIF-1) and HIF-2 (121, 122), which subsequently enhance transcription of the
gene. Elevated VEGF levels in hypoxic tissues are thought to induce angiogenesis by
which more nutrients and oxygen can be delivered to the hypoxic cells (123). Therefore,
exposure of the developing lung to high levels of oxygen during the postnatal period may
downregulate VEGF expression, which in turn inhibit the formation of capillaries in the
lung (124, 125) and interferes with the process of septation (126, 127). Moreover,
hyperoxia may deprive VA storage in the lung, which is known as one of the effective
antioxidants, because oxidative stress caused by hyperoxia can result in more oxidation
of retinol.
A number of studies have demonstrated the great potential of RA in lung tissue repair. In
different neonatal rodent models, known for impaired alveologenesis, like postnatal
hyperoxia exposure or dexamethasone treatment, exogenous at-RA treatment can
partially rescue failed septation and stimulate alveolar formation in neonatal rat lung and
(97, 99, 128). In adult rat or mouse models, the similar effects of RA also have been
observed in elastase or dexamethasone-impaired lung tissue (95, 100, 129, 130). It is well
known that RA functions as a gene transcription regulator through interacting with their

29
specific nuclear receptors (69), and the VEGF genes have been reported as RA
responsive genes (131). Since VEGF family of growth factors plays an essential role in
angiogenesis (132), alveolar regeneration induced by RA may be an important novel
therapeutic approach to the treatment of respiratory diseases characterized by a reduced
gas-exchanging surface area such as BPD and emphysema.
The molecular signals that induce alveolarization are not clearly understood, but RA
signaling in lipofibroblasts appears to play a key role (133). Lipofibroblasts not only
serve as retinoid storage cells of the lung, similar to the stellate cell in the liver (134), but
also contain many components of retinoid signaling pathway including receptors and
binding proteins (94), and can produce endogenous RA (135). Furthermore,
lipofibroblasts are a major source of lung ECM proteins, such as collagen and elastin
(133), and they can synthesize elastin in response to RA (103). Collagen and elastin are
two of the most important components of the ECM that give structural support to resident
cells and contribute to the elasticity of the lung, respectively.
The interactions between ECM and cells not only regulate the development of the normal
lung, but it also plays an essential role in repair and formation of new ECM after lung
injury. Lipofibroblasts are located at the base of new septa and adjacent to type II
alveolar cells (7, 136, 137), which are involved in surfactant synthesis and secretion and
can be considered as progenitor of type I cells. Meanwhile, lipofibroblasts are progenitors
of myofibroblasts, which appear at the tips of newly formed septa and are required for
alveolar septation (137, 138). Based on these findings, lipofibroblasts might serve as a

30
proximate endogenous source of retinoid signals and a signaling center for mediating
alveolus formation during lung septation or after lung injury.
The molecular mechanism of retinol uptake by the lung tissue is still not fully
understood. A receptor-mediated cellular uptake of retinol is a possible mechanism
involved in this process. Recent discovery of a membrane receptor for RBP, STRA6,
supports this hypothesis (69, 139). Another mechanism of retinol uptake into the lung
may exist in that chylomicrons or chylomicron remnants carrying RE could be taken up
by the low-density lipoprotein or chylomicron remnant receptor (33).
It is believed that retinoids are stored as RE in lipid interstitial cells (also known as
lipofibroblasts) (140), which are characterized by their lipid droplets, located in the
proximal portion of alveolar septum, proliferate at the tip of the septum to cause the
eruption of septa into the alveolar sacs (134, 141). Abundant evidence suggest that lipid
interstitial cells play an essential role in normal lung development and injury repair via
epithelial-mesenchymal interactions (137, 142), involved in the synthesis of collagen and
elastin in extracellular matrix (133, 143-147). It also provides lipid substrate for the
synthesis of surfactant phospholipid in type II alveolar epithelial cell (133). During the
postnatal period, cellular retinoic acid binding protein (CRABP) increases in whole lung
tissue and lung lipofibroblasts of rats. Also, increases of RARβ and RARγ were observed
in lipofibroblasts from late gestational period to early postnatal period (148). Recently, it
1.6.4 Retinoid metabolism in the lung

31
was demonstrated that lipid interstitial cells are able to produce and secrete at-RA at the
sites where the secondary septum projects, thereby providing endogenous at-RA for
alveolar formation (47, 94). Based on the observations above, signaling between the
lipofibroblasts and type II cells might be crucial for normal alveologenesis.

32
Chapter 2
MULTIPLE TREATMENT STUDY
Previously we have shown that an oral dose of VA combined with acidic retinoids
synergistically increases retinol uptake and RE formation in neonatal rat lung,
concomitant with the upregulation of expression of several important retinoid
homeostatic genes: LRAT (lecithin:retinol acyltransferase), CYP26B1 (a cytochrome
P450), and STRA6 (stimulated by retinoic acid gene 6). In the present study we
compared the response to VA dose alone, or VA combined with acidic retinoids (RA or
Am580) in two timing protocols: a single early dosing (d 4) vs. multiple dosing
throughout the period of lung septation (d 4, 7, 11, 14). We also tested the influence of
dose administration on lung structural and functional genes. The results have shown that
multiple dosing resulted in a higher, cumulative increase in lung RE content for all
treatments. However, the increase of gene upregulation mediated by acidic retinoids after
the single and multiple dosing did not differ between the two treatment protocols. Lung
structural and functional genes remained nearly constant with both treatment and timing.
In conclusion, multiple treatments of VA and acidic retinoids in combination during the
septation period greatly increased neonatal lung RE content in a synergistic and
cumulative manner. Repeated but transient induction of retinoid homeostatic genes by
2.1 ABSTRACT

33
acidic retinoids at the time of each dosing may explain the observed cumulative
synergistic enhancement of RE formation.
Lung VA storage starts in late gestation and then VA is quickly depleted during the
perinatal and postnatal periods (92, 94). It is believed that the depletion of lung VA
storage is due to the high demands of retinoids for normal lung development and
maturation (149). Preterm infants who have low VA status at birth are more susceptible
to various respiratory diseases, such as BPD (106, 150, 151). Several clinical trials have
demonstrated that VA supplementation not only improves VA status but also reduce the
risk of chronic lung disease in preterm infants who often have very low birth weight at
birth (109, 152-155). In weaning rats fed a VA deprived diet, lung epithelium undergoes
striking morphological alterations, such as keratinizing metaplasia, an increased size of
rat lung airspaces, and reduced collagen and elastin in the parenchyma (149, 156, 157) .
The administration of RA to postnatal rats or mice could prevent the dexamethasone
caused low gas-exchange surface area and low alveolar number, and rescue failed
alveolar septation (97, 99). RA also enhanced alveolar septation in emphysematous rats
(100) and adult rats after pneumonectomy (158).
Given that RA is a crucial regulator of lung development, maturation and maintenance of
normal functions, studies on improving VA status in neonatal lung have been of great
interest. Shenai et al. showed that maternal VA supplementation resulted in an obvious
2.2 INTRODUCTION

34
increase in lung RE contents in the fetuses and offspring of rats (159). Previously, we
reported that providing a combination of VA and RA (10:1 molar ratio) directly to the
neonatal rats is much more effective (~4 fold) in increasing lung RE than providing the
same amounts of VA or RA separately (160, 161). Metabolic studies showed that VARA
directs more delivery of [3H]retinol, used to trace the uptake of newly absorbed retinol, to
the lung (67, 161). However, the synergistic effect of VARA was only observed in the
lungs, as RE in the liver was increased equally by VARA and an equal dose of VA only,
without RA (160, 161).
Our latest investigation on the molecular mechanism of VARA synergy suggested that
the VARA promotes RE formation in the lung by upregulating the expression of LRAT,
CYP26B1 and STRA6 mRNAs, thus, enhances uptake and storage of VA in the but
transient activity of RA on retinoid homeostatic genes (67), and this finding leads us to
the next hypotheses that repeated treatments with RA would result in much stronger and
sustained activity on lung RE formation.
In the present study, we supplemented neonatal rats with multiple doses of retinoids from
postnatal day 4 to day 14, the period of septation, to compare effects of multiple dosing
with a single dosing on RE formation. We then determined whether long-term treatment
with RA can raise expression of LRAT, CYP26B1 and STRA6 during the postnatal
period. This study also evaluated the effect of multiple treatments on the expression
pattern of several lung structural and functional genes. Overall, this study was designed
to test the effects of long-term supplementation of VARA on promoting lung RE storage
and its influences on normal lung development in postnatal life.

35
Hypothesis 1: Multiple treatments with VARA or VAAm580 throughout the postnatal
period of lung septation will significantly increase RE content in neonatal lung in a
cumulative manner. Here, we used Am580 to investigate the regulatory activity of RA
throughout the supplemented period. The repeated action from supplementation with RA
will enhance the expression of LRAT, CYP26B1 and STRA6 mRNA to higher levels
compared with the effect of a single treatment.
Hypothesis 2: Treatment with Am580 will produce a stronger induction in expression of
retinoid homeostatic genes compared with RA, thus contributing to a higher RE
formation in the neonatal lung.
Hypothesis 3: Sustained activity of acidic retinoids from multiple treatments will also
regulate the expression pattern of several lung structural and functional genes during
postnatal period.
Aim 1: To investigate how the multiple treatments of RA or Am580 during the period of
lung septation affect retinyl esters (RE) accumulation in the neonatal rat lung.
In this study, we treated neonatal rats with a single dose of VA and acidic retinoids
combination on day 4 or day14, and multiple doses through day 4 to day 14, respectively.
2.3 HYPOTHESIS AND AIMS

36
Then we determined lung total retinol contents by HPLC to determine the synergistic
effect of multiple doses on lung RE formation.
Aim 2: To determine how the retinoid homeostatic genes respond to repeated treatments
with VARA or VAAm580.
In this study, we conducted real-time polymerase chain reaction (rt-PCR) to quantify the
mRNA levels of LRAT, CYP26B1 and STRA6 genes in the lung of neonatal rats to
compare the effects of single or multiple dose of RA.
Aim 3: To determine how multiple treatments of VARA or VAAm580 will affect lung
structural and functional genes.
We examined expression levels of several lung structural and functional genes including
surfactant proteins, collagen, β-laminin, fibronection, elastin mRNA as well as vascular
endothelial growth factors (VEGF), after treatments, to test their responses to multiple
treatments of RA.
Animal procedures were approved by the Institutional Animal Use and Care Committee,
Pennsylvania State University. We conducted three studies (Fig.2.1): single early dosing
2.4 MATERIALS AND METHODS
Animals and experimental design

37
(d 4), single late dosing (d14), and multiple dosing throughout the period of lung
septation (d 4, 7, 11, 14). In each of these studies, neonatal Sprague-Dawley rats were
randomly divided into 6 groups (n=4-5/group) that received treatments of oil (control),
VA alone, RA alone, VA combined with RA (VARA), or VA combined with Am580
(VAAm). Neonatal pups were delivered and nourished by mother rats fed a VA adequate
diet. Since the pups were from several litters, sexes were evenly distributed to each
group. The average body weight of each group was close to each other. Before each
treatment, the pups were weighed and the dose was adjusted to 0.4 µl/g bodyweight.
We purchased VA, in the form of all-trans-retinyl palmitate and all-trans-RA (at-RA)
from Sigma-Aldrich (St. Louis, MO). Am580 was contributed by H. Kagechika,
University of Tokyo. Dosage selection for VA was based on the amount shown in human
newborns to reduce morbidity and mortality (50,000 IU/2.5KG) (10, 162). By converting
international unit to mass unit with the factor of 0.548 µg retinyl palmitate (RP)/IU, we
calculated the VA doses to be 10.96 µg RP (or 6 µg retinol) /g BW, scaled to body weight
of neonatal rats. The amount of RA was based on previous usage shown to induce lung
septation in neonatal rats (97): 500 µg RA ip/kg body weight, and we adjusted the
amount due to the estimation that about 80% of dose will be absorbed by oral delivery.
VA and RA were mixed at molar ratio of 10:1 and the concentration for each one was
0.05 M and 0.005 M, respectively. Am580 was prepared at the same molar concentration
2.4.2 Dose preparation

38
as RA, and mixed with VA at the same molar ratio of VARA. Canola oil was used as
placebo (control).
The volume of each dose provided to pup was 0.4 µl/g body weight and the exact volume
of dose was determined based on the pup’s body weight (20 nmol retinol and 2 nmol of
acidic retinoid, depending on treatment group, per gram of body weight). In studies I and
II, a single dose was given to pups on day 4 or day 14, respectively. In the study III,
repeated doses were given on day 4, 7, 11 and 14. Pups were killed with carbon dioxide
(CO2) 6 hours after treatment in the study I and II, and 24 hours after the last treatment in
the study III. The lung tissues were removed, trimmed and weighed. All samples were
frozen in liquid nitrogen immediately and then stored -80oC for later analysis.
2.4.3 Experimental methods and tissue collection

39
Figure 2.1 Animal experimental design. Three studies were conducted which differed
in how many doses were given and how old the neonatal rats were. In study I and II, a
single oral dose was given to pups on postnatal day 4 (Study I), and on postnatal day 14
(Study II). In study III, multiple dosing were given on day 4, 7, 11, 14. Tissues were
collected 6h after the single treatment in study I and II, and 24h after the last treatment in
study III. In each study, animals were randomly assigned to 6 groups, and received oil
(vehicle), VA, RA, VARA, Am580 and VAAm580, respectively.

40
Portions of the lung tissue were cut, weighed and extracted in chloroform:methanol, 2:1
v:v, overnight. Then, samples were processed by the Folch washing procedure (163).
After the final wash, the extract was dried down under argon; the samples were
redissolved in 2 ml hexanes. A portion of the hexanes volume was dried again and then
underwent a hydrolysis reaction by a saponification procedure. A known amount of an
internal standard, trimethylmethoxyphenyl-retinol (TMMP) was added to each sample
and the samples were dried under argon and reconstituted in 100 µl of methanol for
HPLC analysis. Portions of each sample (usually 18-22 µl) were injected onto a C-18
HPLC column and eluted with a gradient of 92.5:7.5 methanol:water at a flow rate of 1.5
ml/min for 5 min. The eluate was monitored by a Waters 960 photodiode array detector
and the areas of the peaks for TMMP and retinol were analyzed by Millenium-32
(Waters) software.
Total RNA from lung tissues from individual pups was extracted using a guanidine
extraction method and reverse transcribed into its complementary DNA (cDNA). The
diluted reaction product was used for real-time PCR (rt-PCR) analysis. Primers designed
2.4.4 Retinoid analysis
2.4.5 Gene mRNA level determination

41
to detect mRNA expression were: 5´-ATA GGA TCC TGA CCA ACA CTA CAT CCT
CTC-3´ (forward) and 5´-ATT CTC GAG TCT AAG TTT ATT GAA ACC CCA GA-3´
(reverse) for rat LRAT (NM_022280.2); 5'-TTG AGG GCT TGG AGT TGG T-3'
(forward) and 5'-AAC GTT GCC ATA CTT CTC GC-3' (reverse) for rat CYP26B1
(NM_181087); 5'-GTG CCA GTG ATT GCT GAA GA-3' (forward) and 5'-GGA GGT
GTC CTC TGG ATG AA-3' (reverse) for rat CYP26A1; 5'-CCG ATC CTG GAC AGT
TCC TA -3' (forward) and 5'-CCA CCT GGT AAG TGG CTG TT -3' (reverse) for rat
STRA6 (NM_0010029924.1); 5’-TTG TCG CTG GTA TCA AGT GC-3’(forward) and
5’-CAG CCC CTA TCA TTC CAT GT-3’ (reverse) for rat SPA1 (sftpa1)
(NM_017329.1); 5’-GAT GAT GGG GAA GCT GGT AA-3’ (forward) and 5’-ACC
ATT GGC ACC TTT AGC AC-3’(reverse) for rat collagen (Col1a1) (NM_053304.1);
5’-GAC TTG GGG CGT GTA CAG AT-3’(forward) and 5’-GCA TGA CCA TAG CAG
AAG CA-3’(reverse) for rat laminin (Lamb1) (NM_001106721.1); 5’-GAG GAA AGC
CTG GGA AAG TT-3’ (forward) and 5’-TCC ACC TCT GGC TCC ATA CT-3’
(reverse) for rat elastin (Eln) (NM_012722.1); 5’-ACC ACC CAG AAC TAC GAT GC-
3’ (forward) and 5’-TCT CCC AGG AGT CAC CAA TC-3’ (reverse) for rat fibronectin
(fn1) (NM_019143.2); 5’-GCC CAT GAA GTG AAG TT-3’ (forward) and 5’-TTT CTT
GCG CTT TCG TTT TT-3’ (reverse) for rat Vegfa (NM_031836.2). The mRNA
expression level of each sample was corrected by calculating mRNA-to-ribosomal 18S
RNA ratio. Data were normalized to the average value for the control group, set at 1.00,
prior to statistical analysis.

42
Data are presented as group means ± SEM (standard error of the mean). Group
differences were tested by one-factor ANOVA followed by Fisher’s protected least
significant difference test. The software used for statistic analysis was GraphPad Prism
(San Diego, CA). For comparison, we converted the mean mRNA value of the lung
control group to a value of 1, and the mean values of the other groups were converted
accordingly. To reduce variance of each group mean, values were transformed to log10
form before statistical analysis. Differences with p≤0.05 were considered significant.
Multiple dosing resulted in a higher, cumulative increase in lung RE contents for all
treatments. Previously we have shown that VA combined with acidic retinoids
synergistically increases retinol uptake and RE formation in neonatal rat lung. In the
present project, we designed three studies according to the timing of the dosing
throughout the period of lung septation: an early treatment study (a single dosing on day
4), a late treatment study (a single dosing on day 14) and a multiple treatment study
(multiple dosing on day 4, 7, 11, 14), to compare the level of lung RE formation
influenced by timing and numbers of times of dosing (Fig. 2.2). In each of the study,
2.4.6 Statistical analysis
2.5 RESULTS

43
neonatal rats were assigned to 6 groups and animals in each group received oral dose of
oil (control), VA alone, RA alone, VARA, Am580 alone, or VAAm respectively. In both
of the single dosing studies, VARA and VAAm promoted higher lung RE formation
compared with treatment of VA alone. The results of the early treatment studies agree
with the result of our previous study, in which VARA and VAAm580 could
synergistically increased lung RE content as early as 6 h after dosing. The treatment on
day 14 showed a similar result as the early time (day 4) administration, indicating a
consistent mechanism of VA metabolism throughout the lung septation period. Relative
to the results of the single dosing studies, multiple dosing resulted in a higher,
accumulative increase in lung RE contents for all treatments. The synergy with VARA or
VAAm was still significant in multiple dose studies. Notably, the repeated treatment of
VAAm580 exaggerated the effect of VARA in promoting RE formation, which is a 2
fold increase in lung RE. This is because of the Am580 is resistant to metabolism and its
sustaining activity on gene regulation alter neonatal lung retinoid metabolism. In
conclusion, multiple treatments of VA and acidic retinoids in combination during the
septation period greatly increased neonatal lung RE content in a synergistic and
accumulative manner.
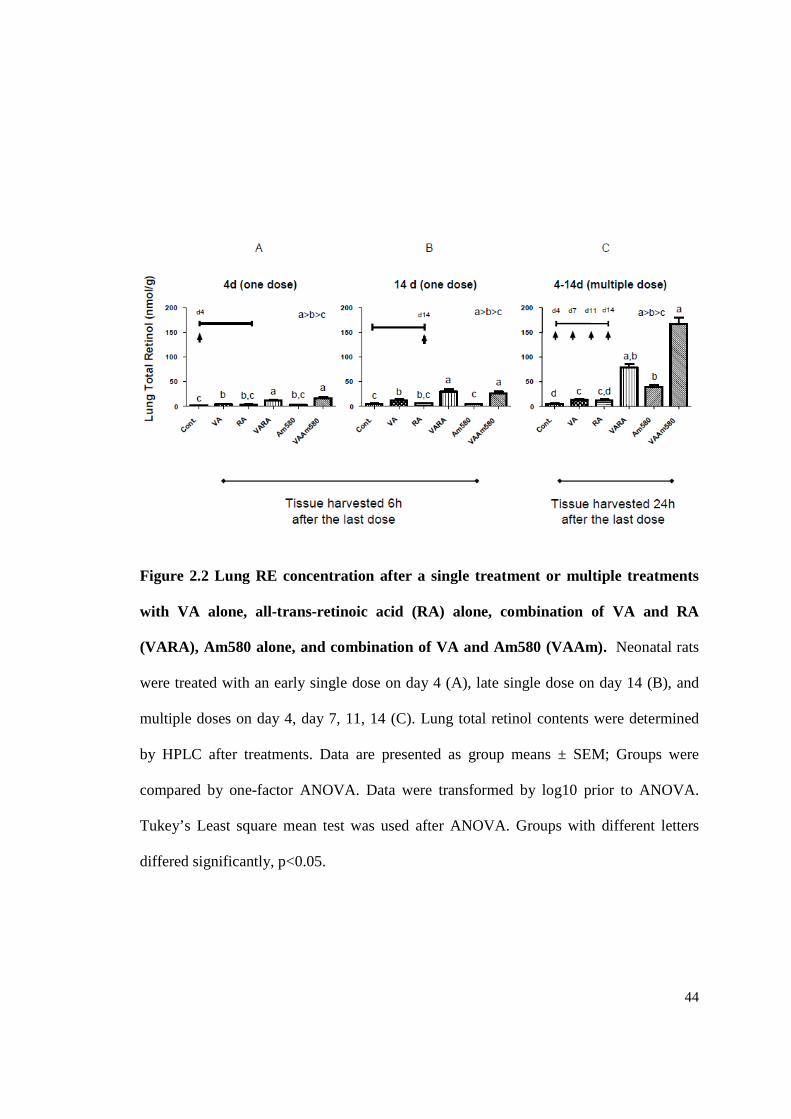
44
Figure 2.2 Lung RE concentration after a single treatment or multiple treatments
with VA alone, all-trans-retinoic acid (RA) alone, combination of VA and RA
(VARA), Am580 alone, and combination of VA and Am580 (VAAm). Neonatal rats
were treated with an early single dose on day 4 (A), late single dose on day 14 (B), and
multiple doses on day 4, day 7, 11, 14 (C). Lung total retinol contents were determined
by HPLC after treatments. Data are presented as group means ± SEM; Groups were
compared by one-factor ANOVA. Data were transformed by log10 prior to ANOVA.
Tukey’s Least square mean test was used after ANOVA. Groups with different letters
differed significantly, p<0.05.

45
Multiple treatments of VA and acidic retinoids in combination during the septation
period had no cumulative effects on the expression of lung homeostatic genes. But
VAAm can prolong the effect.
Our previous studies had shown that acidic retinoids combined with retinol could
increase retinol uptake and esterification in neonatal lung by upregulating the expression
of lung LRAT and STRA6 genes. In the present study, we examined how the long-term
administration of acidic retinoids affects the expression of retinoid homeostatic genes.
In both early and late single treatment studies, LRAT mRNA was significantly induced
by acidic retinoids at 6 h. The stable retinoid, Am580, compared with RA, had much
stronger effect of inducing LRAT mRNA compared with RA (Fig. 2.3). These results are
consistent with the results of our previous 6 h treatment study (67). However, in the
multiple treatment study, LRAT mRNA was reduced to the basal level 24 h after the last
dosing in the lungs of RA and VARA-treated neonates. By comparison with RA, Am580
dramatically increased LRAT mRNA at 6 h and maintained this level at 24 h. This result
suggests that even multiple treatments with RA still increase mRNA level transiently,
probably because RA was quickly metabolized by CYP26B1 (RA hydroxylase). In
contrast, due to its resistance to oxidative metabolism, Am580 shows its sustained
regulatory activity on LRAT gene expression even 24 h after the treatment. The extended
increase in LRAT mRNA coincided with the dramatic increase in RE content in the lung
of VAAm580 treated pups. Am580 continues to exert its regulatory activity due to its

46
resistance to oxidative metabolism, and CYP26B1 is dramatically upregulated, to a
higher extent than LRAT.
The rt-PCR results for CYP26B1 showed similarity with LRAT (Fig. 2.4). In both single
dose studies, the level of CYP26B1 mRNA was elevated 3–25-fold by all of the
treatments at 6 h, compared with control. In the multiple dose study, the mRNA level in
RA or VARA treated pups come down to basal level at 24 h, where as Am580 treatment
maintains its level at 24 h. These results indicated that CYP26B1 gene also could be
transiently induced by RA but persistently by Am580.
Then we determined STRA6 gene expression levels (Fig. 2.5). By contrast to our
pervious results, the regulation of STRA6 in the lung of 4-day old neonates by RA or
Am580 was not as strong as in the previous analysis from 7~8-day old rats, but it still
showed a similar trend of increasing in the pups treated with acidic retinoids. A
comparison of 4-day old and 2 week old rats showed that STRA6 is developmentally
regulated since the older pups, even in the vehicle group, had lower STRA6 expression
levels than younger pups.

47
Figure 2.3 Expression level of Lung LRAT gene after treatments. Neonatal rats
received an early single dose on day 4 (A), a late single dose on day 14 (B) and multiple
doses on day 4, 7, 11, 14 (C). Lung tissue from neonatal rats treated with oil, VA, RA,
VARA, Am and VAAm was processed for total RNA isolation and subjected to rt-PCR
analysis. Data was normalized to 18S rRNA and the average value for the control group
was set to 1 for each experiment. Data are presented as group means ± SEM; Groups
were compared by one-factor ANOVA. Data were transformed by log10 prior to
ANOVA. Tukey’s least square mean test was used after ANOVA. Groups with different
letters differed significantly, p<0.05.

48
Figure 2.4 Expression level of lung CYP26B1 gene after treatments. Neonatal rats
received an early single dose on day 4 (A), a late single dose on day 14 (B) and multiple
doses on day 4, 7, 11, 14 (C). Lung tissue from neonatal rats treated with oil, VA, RA,
VARA, Am and VAAm was processed for total RNA isolation and subjected to rt-PCR
analysis. Data was normalized to 18S rRNA and the average value for the control group
was set to 1 for each experiment. Data are presented as group means ± SEM; Groups
were compared by one-factor ANOVA. Data were transformed by log10 prior to
ANOVA. Tukey’s least square mean test was used after ANOVA. Groups with different
letters differed significantly, p<0.05.

49
Figure 2.5 Expression level of lung STRA6 genes after treatments. Neonatal rats
received an early single dose on day 4 (A), a late single dose on day 14 (B) and multiple
doses on day 4, 7, 11, 14 (C). Lung tissue from neonatal rats treated with oil, VA, RA,
VARA, Am and VAAm was processed for total RNA isolation and subjected to rt-PCR
analysis. Data was normalized to 18S rRNA and the average value for the control group
was set to 1 for each experiment. Data are presented as group means ± SEM; Groups
were compared by one-factor ANOVA. Data were transformed by log10 prior to
ANOVA. Tukey’s least square mean test was used after ANOVA. Groups with different
letters differed significantly, p<0.05.

50
Retinoid treatments did not perturb the expression of lung structural and functional
genes.
Lung surfactant protein A1 (SPA1), collagen, laminin, elastin, fibronectin and vascular
endothelial growth factor A (VEGF-a) are important to lung structure and function during
lung development and maturation. We next examined whether the expression of these
genes was affected by the retinoid treatments used. Primers for SPA1, collagen, laminin,
elastin, fibronectin and VEGF-a were designed and used to analyze each of these genes
necessary for normal lung functioning, determined by rt-PCR. We tested SPA1 and
collagen genes with sample from individual pup, and laminin, elastin, fibronectin,
VEGF-a genes with pooled cDNA samples for each group. The results showed that all
these genes remained nearly constant with treatment. Laminin and fibronectin genes were
developmentally down-regulated since the older pups, even in the vehicle group, had a
lower expression level compared with younger pups. These results suggest that our
treatments did not perturb the expression of these important lung structural and functional
genes.

51
Figure 2.6 Expression level of lung functional and structural genes after treatments.
Neonatal rats received an early single dose on day 4 (A), a late single dose on day 14 (B)
and multiple doses on day 4, 7, 11, 14 (C). Lung tissue from neonatal rats treated with oil,
VA, RA, VARA, Am and VAAm was processed for total RNA isolation and subjected to
rt-PCR analysis. The analyses of SPA1 and collagen genes were based on sample from
individual pups, while laminin, elastin, fibronectin and VEGF-a were based on pooled
sample of each group. Data was normalized to 18S rRNA and the average value for the
control group was set to 1 for each experiment. Data are presented as group means ±
SEM; Groups were compared by one-factor ANOVA. Data were transformed by log10
prior to ANOVA. Tukey’s Least square mean test was used after ANOVA. Groups with
different letters differed significantly, p<0.05.

52
2.6 DISCUSSION
It is known that the lung is immature at birth. It still needs a process of septation which is
dependent on the signals provided by RA. Severe VA deficiency leads to abnormal
cellular differentiation and proliferation in the epithelium of the respiratory tract (164).
Infants with extremely low birth weight usually have low VA concentrations in plasma
and tissues, and this population is more susceptible to various respiratory diseases and
infections. Therefore, strategies to improve the VA status of newborns in the postnatal
period are of great clinical interest. In our project, we used neonatal rat lung as our model
because lung development of rodents in the first two postnatal weeks parallels lung
development in the human from week 24 of gestation through the first 2 years of age
(165), therefore, it is a good model of preterm infant. Our previous studies have reported
that VA given orally to neonatal rat in combination with RA increase lung RE contents
more effectively than the same amount of VA given alone (160, 161). The administration
of RA promotes retinol uptake and esterification by upregulating several retinoid
homeostatic genes, LRAT, CYP26B1 and STRA6, and thus, redirects part of the flow of
supplemental VA in the neonate lungs, as supported by increased [3H]retinol uptake in
VARA treated neonates (67). The use of Am580, a metabolism-resistant analog of RA,
shows an extended activity in regulating retinoid homeostatic genes and further supports
the effects of RA on lung VA metabolism (67).
In the present study, we determined the effects of multiple treatments with VARA on
promoting lung RE formation during postnatal period. The dose of VA was based on a

53
previous VA supplementation study in newborn infants (10, 162). The amount of 50,000
IU VA was scaled to the body weight of neonatal rats, which is ~21 µmol of retinol per 1
kg body weight. The dose of RA was based on the amount previously used to induce lung
septation in neonatal rats, which is 1.67 µmol/kg (97). We adjusted the dose of RA to 2.1
µmol/kg to compensate for a ~20% loss during intestinal absorption. Indeed, the dose of
VARA was the exact mixture of the VA dose and the RA dose discussed above and thus
had a molar ratio of 10:1 retinol/RA. To evaluate the effect of multiple dosing, we also
conducted two single dose studies at the same time. In both of these studies, a single dose
was administrated to neonatal rats on postnatal day 4 (early treatment study) or postnatal
day 14 (late treatment study), which are the beginning and the end of the septation period,
respectively.
The early and late single treatment did not differ from each other with respect to lung RE
formation. In contrast, multiple treatments showed a higher, cumulative increase in lung
RE content for all treatments. Compared with RA, Am580 exerts a persistent and
enhanced effect on lung VA accumulation, thus, repeated treatments with VAAm580 led
to a much higher increase in RE contents than treatment with VARA (Fig.2.2). Multiple
dose of RA or Am580 given alone also increased lung RE content to a extent even
slightly higher than multiple dose of VA (Fig.2.2). Since there was no retinol mass in the
dose, the possible origin of retinol for newly formed lung RE is the liver. Thus, we
speculate that repeated administrations of RA or Am580 during postnatal period mobilize
the liver VA reserve and deliver retinol to the lung to meet the demands for lung
development, thus changing the distribution of retinol in different tissues.

54
In our single treatment studies, we determined expression of retinoid homeostatic genes
at 6h after dosing, because VARA or VAAm had a rapid effect on retinol uptake and RE
accumulation in the lungs. The gene results agree with the results of our previous 6-h
retinoid treatment study (67), in which LRAT, CYP26B1 and STRA6 were significantly
induced by RA of Am580. Although responses of these genes to the treatment with
retinoids are rapid, a 12 h study has suggested activity of RA is just transient, because
RA is quickly oxidized by induced CYP26B1 (67). To evaluate how long-term retinoids
administration affects expression of retinoid homeostatic genes, we determined gene
levels 24h after the last treatment. The data still showed the similar results to that of 12 h
study (Fig. 2.3, 2.4, 2.5). This suggests the significant increase in lung RE is the outcome
of several temporary promotions of RE formation by RA, after each dose. In contrast,
extended activity from Am580 not only maintains the expression of genes at a high level
but also exaggerates the effects observed for RA (Fig. 2.2, 2.3, 2.4, 2.5).
RA has been implicated in normal lung development and repair mechanism by promoting
expressions of several important structural and functional genes, such as surfactant
proteins (SP), collagens, elastins, etc. (157, 166-171). However, these effects usually
were observed in VA deficient animal models or cultured lung explants. In our study,
neonatal rats were delivered by mothers fed normal diet. Therefore, the VA status in
these rats before the treatments is in a normal range that is adequate to maintain a normal
lung development and functioning. We examined expression of several lung structural
and functional genes to test our hypothesis that multiple administrations of acidic
retinoids might affect their normal expression. However, our results showed that all these

55
genes remained nearly constant with all treatments and suggest that RA or Am580 did
not perturb the expression of these genes. This finding increases the feasibility of using
VARA in a clinical setting for the goal of promoting lung VA status, as no perturbing
effects were observed on important structural and functional genes.
In summary, these studies have shown that multiple treatments of VARA and VAAm
during the period of lung septation increased lung RE contents in neonatal rat lung in a
synergistic and cumulative manner. The effect is accompanied by repeated induction of
retinoid homeostatic genes, LRAT, CYP26B1 and STRA6, by RA or Am580. However,
multiple treatments had no cumulative effects on the expression of these genes. In
contrast, Am580 produced a stronger and sustained induction of LRAT and CYP26B1
mRNAs, even 24 h after the last dose, which may contribute to a higher RE formation in
the neonatal lung. The repeated administrations of acidic retinoid show no influence on
several important lung structural and functional genes. Our findings provide some clues
for a more efficient and better therapeutic strategy in clinic use that may improve lung
RE formation in neonatal lungs.

56
Chapter 3
LPS-INDUCED INFLAMMATION STUDY
Vitamin A plays an important role in postnatal lung development and maturation. Our
previous study has suggested that the principal metabolite, retinoic acid (RA),
supplemented simultaneously with VA (VARA) to neonatal rats synergistically increases
lung retinol uptake and storage by upregulating several important retinoid homeostatic
genes, LRAT (retinol esterification), CYP26B1 (RA hydroxylase) and STRA6 (retinol-
binding protein receptor). However, whether inflammation impacts expression of these
genes and thus affects retinol homeostasis in neonatal lungs is not clear. In this study, we
investigated whether LPS-induced inflammation could affect RE formation in neonatal
rat lung, promoted by VARA. 7~8 day-old neonatal rats were treated concurrently with
(6-h study) or 12 h after (18-h study) LPS administration. Lung tissues were collected 6 h
after VARA treatment. Results suggested that in both studies, LPS slightly reduced the
VARA-mediated increase in lung RE formation. Rt-PCR data indicated that in the 6-h
study, LPS attenuated the induction in LRAT by VARA, but not the CYP26B1 and
STRA6. In the 18-h study, LPS had little effects on the expression of all these genes.
Overall, these results suggest that acute inflammation modestly reduces the RA-mediated
induction in LRAT mRNA in the neonatal lung, in turn, it slightly attenuates the
3.1 ABSTRACT

57
synergistic increase in RE formation promoted by VARA. Nevertheless, VARA
increased lung RE under conditions of mild inflammation.
Vitamin A storage is low in the human lung at birth. Usually, premature delivery is one
of the major factors that contribute to severe VA deficiency and put preterm infants at
high risk for developing pulmonary diseases, such as RDS and a chronic lung disease,
BPD. Supplementing VA to the newborns directly not only improve VA status but also
produces promising results for reducing lung injury and dysfunction (109, 152, 153).
Administration of RA to neonatal rats has been shown to induce alveolar formation,
repair epithelial lesions and increase surfactant synthesis in lung (172-174).
It is believed that RA exerts its influence in development and cell differentiation through
the binding of nuclear retinoid receptors (RAR), which forms heterodimers with RXR
and subsequently interacts with one or several retinoic acid-responsive elements (RARE)
that are located in the regulatory regions of direct targets, thereby activating gene
transcription (175-177). Several important genes that are involved in retinoid homeostasis
are also regulated by RA activity. Lecithin:retinol acyltransferase, CYP26B1 (RA
hydroxylase) which catalyzes the esterification of retinol and oxidation of RA,
respectively, and STRA6 (retinol-binding receptor) which mediates retinol uptake into
tissue, are known to be tightly regulated by RA in the lung (67). Interruption of the
3.2 INTRODUCTION

58
expression of these genes might alter the balance of the retinol metabolism in the lung
tissue.
Our previous study has shown a more efficacious way in increasing VA storage in the
lung by administrating VA orally in combination with all-trans-RA (VARA) to neonatal
rats (160, 161). VA and RA were mixed in the ratio of 10:1 and the dose produces a
synergistic effect in promoting RE formation, about 4 fold as compared with same
amount of VA given alone. A metabolic study showed that the administration of RA
redirects part of the flow of supplemental [3H]retinol in the lungs in VARA-treated
neonates (67). The molecular mechanism of VARA synergy was investigated and
revealed that including RA in the dose acts as a regulator of VA homeostasis by rapidly
upregulating retinoid homeostatic genes, LRAT, CYP26B1 and STRA6 in the lung (67),
thus promoting RE formation more efficiently.
Inflammation is very often observed in preterm infants or infants with a weakened
immune system. In the process of inflammatory responses, proinflammatory cytokines
such as tumor necrosis factor-α (TNF-α), IL-1β and IL-8 are secreted by alveolar
macrophages, fibroblasts, type II pneumocytes and endothelial cells in the early
inflammatory response upon the stimulation of inflammatory agents (178). These
cytokines recruit more circulating neutrophils and macrophages to the local sites of injury
or infection and subsequently release higher amount of cytokines and chemokines to
initiate a sequence of injurious events (179). As an organ with large area exposed to the
outside, the lung is more vulnerable to be attacked by exogenous pathogens, allergens or
chemicals. The increased mRNA level and secretion of pro-inflammatory cytokines and

59
chemokines have been detected in airway of preterm infants at various stages of
developing BPD (180, 181). Risk factors contributing to the inflammatory responses in
the lung include inappropriate resuscitation, oxygen toxicity, mechanical ventilation,
pulmonary and/or systemic infection, etc. (182).
Inflammation is also known to alter the expression of many genes including genes in the
CYP family (183). Our previous in vitro study reported that suppression of RA-induced
CYP26A1 mRNA expression was observed in LPS treated THP-1 cells (184). An in vivo
study in rat liver suggested that, LPS or poly-IC induced inflammation significantly
opposes the induction of CYP26A1 and CYP26B1 expression by RA (185). It is
unknown whether inflammation (which is often associated with newborn infants)
inhibits, attenuates, or has no effect on VARA synergy in the lungs.
In the present study, we investigated effects of LPS-induced inflammation on VARA
mediated RE formation and RA induced expression of LRAT, CYP26B1 and STRA6 in
neonatal rat lungs. At the same time, we compared the lung RE and gene results of
LPS/VARA in a co-administration study and in a model of LPS pre-administration study
to test whether the time of inflammation has an effect on the VARA synergy. The results
of these studies show that LPS slightly attenuates the induction of RE formation in
neonatal lungs when VARA and LPS are administered simultaneously. Gene data
indicate that the inflammation only decreased the RA-induced increase in LRAT mRNA.
However, when LPS was administered in advance, it has little effect on RE formation and
retinoid homeostatic genes induced by RA.

60
Hypothesis 1: Inflammation markers will be dramatically increased in the lung of LPS
treated pups and inflammation will prevent the increase in RE formation in neonatal rat
lung promoted by VARA.
Hypothesis 2. The expression of retinoid homeostatic genes, LRAT, CYP26B1 and
STRA6, which respond to VARA treatment in neonatal rat lungs, will be interrupted by
LPS-induced inflammation.
Aim 1: To test how the storage of RE in the neonatal rat lung is affected by the LPS-
induced inflammation induced.
The combination of VA and its bioactive metabolite retinoic acid, VARA, has a
synergistic effect in increasing RE contents in postnatal lung tissue (161). We
investigated how LPS-induced inflammation would affect lung RE storage and whether
the inflammation would prevent the synergy of VARA. We conducted two studies, 6-h
study and 18-h study which are based on the timing of the treatments. In the 6-h study,
neonatal pups were treated with LPS and VARA simultaneously. In the 18-h study, pups
received VARA treatment 12 h after LPS administration. For both studies, the lung tissue
was collected 6 h after VARA treatments. Lung total retinol contents (RE plus retinol)
from each lung sample were quantified by HPLC. We also demonstrated the
3.3 HYPOTHESIS AND AIMS

61
inflammation state in LPS-treated rats by determining the inflammation markers, such as
IL6, CCL2 gene in the lung and C-creative protein (CRP) in plasma.
Aim 2: To determine the response of retinoid homeostatic genes related to VA uptake
and metabolism in the neonatal lungs after treatment with VARA in inflammation states.
Lung retinoid homeostatic genes LRAT, CYP26B1, and STRA6 play important roles in
VA uptake and metabolism in the lung. By analyzing the mRNA levels of these genes,
we may be able to understand how LPS-induced inflammation interfere the expression of
these genes, thus better understand VA metabolism in the neonatal lung in inflammation
state. In this study, specific genes like LRAT, CYP26B1 and STRA6 were analyzed for
their transcript levels using rt-PCR.
Animal procedures were approved by the Institutional Animals Use and Care Committee
of The Pennsylvania State University. We conducted two studies according to the time
LPS is administered relative to the time VARA was supplemented (Fig. 3.1). In a short-
term study, LPS and VARA were co-administered 6 h before collection of lung tissue. In
a long-term study, LPS was administered 18 h before and VARA was supplemented 6 h
before tissue collection. In each study, 7-8 day-old Sprague-Dawley rat pups were
3.4 MATERIALS AND METHODS
3.4.1 Animals and experimental designs

62
assigned randomly to 4 groups (n=4-5/group) and received canola oil (Control), LPS
alone, VARA, and combination of LPS and VARA. ). Neonatal pups were delivered and
nourished by mother rats fed a VA adequate diet. ). Neonatal pups were delivered and
nourished by mother rats fed a VA adequate diet.

63
Figure 3.1 Animal experimental design. Two studies were conducted according to the
time LPS is administered relative to the time VARA was supplemented. In study I (also
termed 6 hr study), LPS and VARA were administered simultaneously 6 h before tissue
collection. In study II (also termed 18 hr study), pups received VARA treatment 12 h
after LPS administration and tissues were collected 6 h later. In each study, 7-8 day-old
Sprague-Dawley rat pups were assigned randomly to 4 groups (n=4-5/group) and
received canola oil+saline (Control), LPS+oil, VARA+saline, and LPS+VARA
respectively.
3.4.2 Dose preparation

64
Vitamin A, in the form of all-trans-retinyl palmitate, and at-RA, purchased from Sigma-
Aldrich (St. Louis, MO) were mixed in a molar ratio of 10:1 and the concentration for
each one are 0.05 M and 0.005 M, respectively. The dose of the VARA combination
given orally to neonates was based on body weight (0.4 ul/g body wt). Please refer to the
chapter 2 for the detailed information. Pseudomonas aeruginosa-LPS, purchased from
List Biological Laboratory (Campbell, CA), was prepared in ta concentration of 10 µg/ml
and the dose injected intraperitoneally to neonates was 0.02 ml/g body wt (20 µg
LPS/100g body wt, less than the 50 µg/100 g body weight dose for adults). Canola oil
and saline were used as placebos for VARA and LPS, respectively.
The dose of retinoid treatment given to each pup was delivered directly to the rat pup’s
mouth by a micropipette. Pseudomonas aeruginosa-LPS from a stock concentration of
2.5mg/ml was diluted to 0.01 mg/ml (10 µg/ml) and a single dose of 0.02 ml/g body wt
LPS was injected intraperitoneally to these neonates. Neonates from both studies were
sacrificed by carbon dioxide (CO2) inhalation 6 hours after VARA treatment for both
studies and then lung tissue was collected and frozen in liquid nitrogen immediately and
then stored -80oC for later analysis.
3.4.3 Experimental methods and tissue collection

65
Plasma anti-C Reactive Proteins (CRP) was quantified by a rat CRP ELISA TEST KIT
(Life Diagnostic, Inc., West Chester, PA). The C Reactive Proteins are indicators of
inflammation in the blood. The plasma samples were diluted and incubated in the
microtiter wells for 45 minutes. The microtiter wells were subsequently washed and
incubated with horseradish peroxidase (HRP) conjugate for 30 minutes. Then the wells
were washed to remove unbound HRP-labeled antibodies and TMB reagent was added
and incubated for 20 minutes at room temperature. Color development was stopped when
the color changed to yellow. Optical density was measured spectrophotometrically at 450
nm. A standard of serially diluted control serum was included, and titers of antibody were
calculated based on this standard curve. One titer unit was defined as the dilution fold
that produced 50% of the maximal optical density for the standard sample.
Lung tissue was processed and lung total retinol was quantified in the same way as in
chapter 2. Please refer to the chapter 2 for the detailed information.
3.4.4 Plasma anti-CRP antibody enzyme-linked immunosorbent assay
3.4.5 Retinoid analysis

66
Please refer to the chapter 2 for detailed information of LRAT, CYP26B1 and STRA6
primers. Primer sets designed to detect mRNA expression were: 5’-TGT GCA ATG
GCA ATT CTG AT-3’ (forward) and 5’-TGG TCT TGG TCC TTA GCC AC-3’ for rat
IL6 (NM_012589.1); 5’-AGG TGT CCC AAA GAA GCT GT-3’ (forward) and 5’-TGC
TTG AGG TGG TTG TGG AA-3’ (reverse) for rat CCL2 (NM_031530.1). The mRNA
expression level of each sample was corrected by calculating mRNA-to-ribosomal 18S
RNA ratio. Data were normalized to the average value for the control group, set at 1.00,
prior to statistical analysis.
Please refer to the Chapter 2 for the detailed information of statistical analysis.
LPS-induced inflammation slightly reduced the VARA-mediated increase in lung
total retinol
3.4.6 Gene mRNA level determination
3.4.7 Statistical analysis
3.5 RESULTS

67
In this study, we determined lung total retinol concentration after LPS and VARA
administration. We also examined how the time of the initiation of inflammation affected
retinoid homeostasis by conducting a short-term study (6-h study) and a long-term study
(18-h study) which differed according to the time VARA was administered relative to the
time of LPS administration. In the short-term study, LPS and VARA were provided to
neonatal rats concurrently, while in the long term study, LPS was administered 12 hours
before VARA treatment. In both studies, VARA significantly increased lung RE
formation 6 h after VARA supplementation. LPS alone has no effect in changing lung RE
level. In the short-term study, LPS attenuated the increase in RE by VARA by an ~38%
reduction (Fig. 3.2A). In the long-term study, LPS also slightly but not significantly
reduced the increase due to VARA (Fig. 3.2B). In the 18 hr study, we conducted a
metabolic study by adding a tracer of [3H]retinol to the dose and investigating how much
newly absorbed retinol was taken up by lung tissue. We found that the percentage of
newly absorbed 3H in the lungs treated with VARA did not differ between the normal
state and inflammation state (data not shown). These results suggested that an acute
inflammation induced by LPS slightly affects lung RE formation promoted by VARA.

68
Figure 3.2 Lung RE concentration after a single treatment with oil or VARA in the
presence or absence of LPS induced inflammation state. In each study, 7-8 day-old
neonatal rats were randomly assigned to 4 groups, and received a single dose of
saline+oil (control), Saline+VARA, LPS+oil, LPS+VARA. In study I (6 hr study), LPS
and VARA were administered simultaneously (Fig 2A), as in study II (18 hr study), LPS
was administered 12 h before VARA supplementation (Fig 2B). Lung total retinol
contents were determined by HPLC after treatments. Data are presented as group means
± SEM; Groups were compared by one-factor ANOVA. Data were transformed by log10
prior to ANOVA. Tukey’s Least square mean test was used after ANOVA. Groups with
different letters differed significantly, p<0.05.

69
The increase in inflammation markers indicated the state of inflammation
To check whether the neonatal rats are in a state of inflammation, we determined the
expression of certain inflammation markers in the neonatal lung by rt-PCR and the level
of C-reactive protein (CRP) in the serum by ELISA. Interleukin-6 (IL-6) and hemokine
(C-C motif) ligand 2 (CCL2) are inflammatory mediators in the early inflammatory
response and in the evolution of inflammatory events. In the lung of neonatal rats, the
expression of these two genes was increased by LPS at 6 h (Fig. 3.3A, 3.4A). But 18 h
later, the increase returned to the control level (Fig. 3.3B, 3.4B). On the other hand, the
increase in plasma CRP level was only observed at 18 h, not 6 h (Fig. 3.5A, 3.5B). These
results demonstrated that neonatal rats that received LPS administration were
experiencing an inflammation state in both the short-term and the long-term studies.
However, due to the time of gene response and protein response to inflammation, the
expression level of each inflammation indicator could be very different.

70
Figure 3.3 Expression level of lung IL6 gene 6 h or 18 h after LPS administration..
Lung tissue from neonatal rats treated with saline+oil (control), Saline+VARA, LPS+oil,
LPS+VARA were collected 6h after VARA treatment and processed for total RNA
isolation and subjected to rt-PCR analysis. In study I (6 h study), LPS and VARA were
administered simultaneously (Fig 3A), as in study II (18 h study), LPS was administered
12 h before VARA supplementation (Fig 2B). Data was normalized to 18S rRNA and the
average value for the control group was set to 1 for each experiment. Data are presented
as group means ± SEM; Groups were compared by one-factor ANOVA. Data were
transformed by log10 prior to ANOVA. Tukey’s Least square mean test was used after
ANOVA. Groups with different letters differed significantly, p<0.05.

71
Figure 3.4 Expression level of lung CCL2 gene 6 h or 18 h after LPS administration.
Lung tissue from neonatal rats treated with saline+oil (control), Saline+VARA, LPS+oil,
LPS+VARA were collected 6h after VARA treatment and processed for total RNA
isolation and subjected to rt-PCR analysis. In study I (6 hr study), LPS and VARA were
administered simultaneously (Fig 3A), as in study II (18 hr study), LPS was administered
12 h before VARA supplementation (Fig 2B). Data was normalized to 18S rRNA and the
average value for the control group was set to 1 for each experiment. Data are presented
as group means ± SEM; Groups were compared by one-factor ANOVA. Data were
transformed by log10 prior to ANOVA. Tukey’s Least square mean test was used after
ANOVA. Groups with different letters differed significantly, p<0.05.

72
Figure 3.5 Plasma C Reactive Proteins (CRP) level 6 h or 18 h after LPS
administration. The plasma samples from neonatal rats treated with saline+oil (control),
Saline+VARA, LPS+oil, LPS+VARA were collected 6h after VARA treatment and
subjected to ELISA to test plasma CRP level. Groups were compared by one-factor
ANOVA. Data were transformed by log10 prior to ANOVA. Tukey’s Least square mean
test was used after ANOVA. Groups with different letters differed significantly, p<0.05.

73
LPS-induced inflammation did not interrupt the induction of STRA6, LRAT and
CYP26B1 mRNA by VARA treatment
LRAT, CYP26B1 and STRA6 are the most important genes that regulate retinoid
homeostasis in the neonatal lung. RA is able to up-regulate these genes to enhance lung
uptake and storage in the lung. We next determined the expression level of these retinoid
homeostatic genes in the lung of LPS treated rats. Gene expression was determined by
quantitative PCR.
Treatment with LPS alone has no effects on STRA6 expression, but VARA was able to
significantly increase its expression regardless of addition of LPS and what time LPS is
administered (Fig. 3.8). Similarly, VARA significantly induced LRAT and CYP26B1
expression respectively in both the 6-hour and 18-hour studies (Fig 3.6, 3.7). Only in the
6-h study, LPS slightly reduced the induction of LRAT by VARA which is
corresponding to a slight reduction of RE formation in LPS/VARA treated group. In both
studies, expression of all retinoid homeostatic genes was significantly and rapidly
increased above the control level 6 h after VARA supplementation. These results
suggested that LPS initiated inflammation has little effect in interrupting the VARA
induced gene induction in neonatal rat lungs.

74
Figure 3.6 Expression level of lung LRAT gene 6 h after VARA treatment. Lung
tissue from neonatal rats treated with saline+oil (control), saline+VARA, LPS+oil,
LPS+VARA were collected 6 h after VARA treatment and processed for total RNA
isolation and subjected to rt-PCR analysis. In study I (6 h study), LPS and VARA were
administered simultaneously (Fig 3A), as in study II (18 h study), LPS was administered
12 h before VARA supplementation (Fig 2B). Data was normalized to 18S rRNA and the
average value for the control group was set to 1 for each experiment. Data are presented
as group means ± SEM; Groups were compared by one-factor ANOVA. Data were
transformed by log10 prior to ANOVA. Tukey’s Least square mean test was used after
ANOVA. Groups with different letters differed significantly, p<0.05.

75
Figure 3.7 Expression level of lung CYP26B1 gene 6 h after VARA treatment. Lung
tissue from neonatal rats treated with saline+oil (control), Saline+VARA, LPS+oil,
LPS+VARA were collected 6h after VARA treatment and processed for total RNA
isolation and subjected to rt-PCR analysis. In study I (6 hr study), LPS and VARA were
administered simultaneously (Fig 3A), as in study II (18 hr study), LPS was administered
12 h before VARA supplementation (Fig 2B). Data was normalized to 18S rRNA and the
average value for the control group was set to 1 for each experiment. Data are presented
as group means ± SEM; Groups were compared by one-factor ANOVA. Data were
transformed by log10 prior to ANOVA. Tukey’s Least square mean test was used after
ANOVA. Groups with different letters differed significantly, p<0.05.

76
Figure 3.8 Expression level of lung STRA6 gene 6 h after VARA treatment. Lung
tissue from neonatal rats treated with saline+oil (control), Saline+VARA, LPS+oil,
LPS+VARA were collected 6h after VARA treatment and processed for total RNA
isolation and subjected to rt-PCR analysis. In study I (6 hr study), LPS and VARA were
administered simultaneously (Fig 3A), as in study II (18 hr study), LPS was administered
12 h before VARA supplementation (Fig 2B). Data was normalized to 18S rRNA and the
average value for the control group was set to 1 for each experiment. Data are presented
as group means ± SEM; Groups were compared by one-factor ANOVA. Data were
transformed by log10 prior to ANOVA. Tukey’s Least square mean test was used after
ANOVA. Groups with different letters differed significantly, p<0.05.

77
BPD is one of the lung disorders that most commonly occur in premature infants who
received prolonged mechanical ventilation and oxygen therapy to treat RDS. It is
characterized by chronic inflammation and impaired abnormal structural changes (alveoli
and vascular) in the lung. In the state of inflammation, inflammatory cells are recruited to
the airways and lung tissues to produce pro-inflammatory mediators such as TNF-α, IL-1,
IL-8, which leads to a series of injurious immune responses. As a major organ for
inflammatory responses and VA homeostasis, liver has been investigated for the
relationship between inflammation and retinoid homeostasis. It was reported that
expression of cytochrome P450 (RA hydroxylase) mRNA can be suppressed by
inflammation or infection in rat hepatocytes (183). Another study has indicated that
inflammation induced by LPS opposes the induction of CYP26A1 and CYP26B1 to RA
in the rat liver when RA and LPS were administered at the same time or LPS was
administered 4 h after RA treatment (185). These studies suggest inflammation might
affect normal VA homeostasis by interfering with expression of retinoid homeostatic
genes.
As a major extrahepatic organ that require VA’s activity for normal development and
function, the lung has potential for storing VA after VA administration (27). When RA is
supplemented simultaneously with VA, the dose produced a synergy in increasing lung
3.6 DISCUSSION

78
RE level, which is not observed in the liver (160, 161). Therefore, VA level can be
elevated in a tissue-specific manner. However, little is known about retinoid homeostasis
in the lung under inflammation condition. In our present studies, we used a model of
LPS-induced systemic inflammation to study whether inflammation has any influence on
retinoid homeostasis in the lungs. The results indicated that the influence from LPS-
induced inflammation on VA homeostasis is mild and only slightly attenuates the
increase in lung RE by VARA. LPS-induced inflammation reduced ~38% RE increase
when LPS and VARA were administered concomitantly, and reduced only ~24% when
LPS was administered 12 h before retinoids treatment. The induction in IL-6 and CCL2
mRNA (indicators of early inflammatory responses) in the lung and the increase in CRP
protein in the plasma (marker of acute inflammation) confirmed the inflammation state in
the animals. The gene data also indicated that inflammation state had little influence on
the expression of lung retinoid homeostatic genes.
In our study, we used LPS, an outer membrane component of Gram negative bacteria, to
induce inflammation in neonatal rats. Although LPS-induced inflammation only
attenuates the VARA mediated increase in RE formation modestly, it is still interesting
for us to understand the possible mechanism that causes the attenuation. LPS is able to
initiate immune responses via binding to toll-like receptor 4 (TLR4) and subsequently to
stimulate several signaling pathways that in turn activate a variety of transcription factors
including nuclear factor kappa-B (NF-κB) and AP-1, etc (186, 187). The activated
transcription factors in the cytoplasm translocate into the nucleus where they bind to
specific response elements on target genes and induce many genes encoding

79
inflammatory mediators (187). Although our experiments showed little influence from
LPS on the expression of retinoid homeostatic genes, co-administration of LPS and
VARA still reduced expression of LRAT mRNA 6 h after treatment, which perhaps
explains the reduction in VARA promoted RE formation. On the other hand, RA was
reported to be able to attenuate immune responses by opposing LPS and/or TNF-α
induced STAT expression and lowered the binding activity of STAT-1 and NF-κB to
response elements (184). Other in vitro and in vivo studies suggested that RA is able to
reduce NF-κB activity and DNA binding (86, 188). Therefore, RA’s activity in inhibiting
TLR4 downstream signaling pathways might also contribute to attenuate inflammatory
responses in neonates.
Animal and adult human studies have shown that long term respiratory inflammation
results in increased VA consumption and depletion of hepatic stores (189, 190). But in
our studies, there is no significant reduction in the lung RE and liver RE (data not shown)
observed in LPS treated group. We speculated that the short term inflammation shows
little impact on retinoid consumption. Meanwhile, the animals we used were naturally
delivered, healthy neonatal rats. They probably have a better anti-inflammatory
mechanism or immune defense than animals and humans whose health is compromised
by chronic inflammatory lung diseases. However, it would be interesting to understand
whether long-term inflammation will prevent the synergy of VARA.
However, there are limitations to this study. First, while LPS-treated newborn rat is a
common reproducible inflammation model, the systemic inflammation produced in this
model may not simulate the local inflammation like human BPD. As our study was

80
designed to evaluate the effects of postnatal lung inflammation on lung RE formation and
gene regulation, we did not observe marked decrease in lung RE content after VARA
treatment and the changes in expression of retinoid homeostatic genes. We also failed to
see significant differences in lung RE and gene levels between the 6h and 18h studies. To
better evaluate the effects of local inflammation on lung VA homeostasis, in future, we
need to establish a pneumonia model in neonatal rats which has more strong
inflammatory reaction in the lung and is more similar to the condition of BPD in preterm
infants. Second, the does of LPS administered to pups may be too low to induce the
inflammatory conditions that can compromise VARA promoted RE formation in
neonatal rat lung. To avoid this situation, an appropriate dose of LPS or other pathogen
should be determined in our future studies.
In summary, these studies provide insights into how acute inflammation during the early
postnatal period affects retinoids homeostasis in the neonatal rat lung. We demonstrated
that the LPS-induced inflammation has little influence on the induction of retinoid
homeostatic genes by RA and only slightly attenuates the synergistic increase in RE by
VARA. These results suggest that, even in a state of inflammation, VARA could still be
an effective therapeutic strategy for improving RE status in neonatal lungs.

81
Chapter 4
ACIDIC RETINOIDS DILUTION STUDY
In contrast to VA alone, the combination of VA and 10% RA (10:1 molar ratio)
administered orally is able to increase lung RE contents synergistically in neonatal rats.
However, the lowest RA concentration to cause the same promotion is not known. In this
part of this thesis, we designed two studies to test the lowest concentration of RA in
VARA dose that still is able to promote high RE formation in neonatal rat lung using a
single dosing (day 5) vs. multiple dosing schedule throughout the period of lung septation
(day 4, 7, 11, 14). In the first study, a single oral dose of oil, VA, VARA (10% RA) and
VA with a series of diluted RA (5%, 2%, 1%, 0.5% RA) was administered to 5 day-old
pups. In the second study, doses of oil, VA, VARA (10%, 2%, 1% RA) were given to
neonatal pups on each of days 4, 7, 11, and 14. We also examined the effect of
substituting Am580 (a stable analog of RA) for RA in the multiple dosing protocol by
giving oil, VA, VAAm580 (10%, 2%, 1% Am580), with the same feeding schedule as
above. Lung tissues were collected 6 hours after the last treatment and analyzed by HPLC
for lung total retinol level. The results showed that although the concentration of RA was
reduced to 1%, it still increased lung RE to the same level 10% RA does. However, this
effect is only observed in the single dosing study. The multiple doses of diluted RA (2%)
4.1 ABSTRACT

82
did not show the same effect. However, in the Am580 dilution study, which was parallel
to the multiple dosing RA study, even 1% Am580 showed the same effect of 10%
Am580 in promoting lung RE formation. We conclude that RA even in more diluted
concentration combined with the same dose of VA still promote high retinol uptake and
RE formation in neonatal lungs at a short time (6 h) compared with VA given alone. But,
this effect is not seen after multiple treatments which might be due to a quick metabolism
of a reduced amount of RA.
VA storage in the lung shows a specific pattern during the perinatal period. It rises
significantly in late gestation, starts to decline before birth and continues into the early
postnatal period (92). The utilization of VA stores strongly implies the dependence on
VA for postnatal lung development. Improving lung VA status in the early postnatal life
has been shown to be beneficial for lung development and reducing lung injury and
dysfunction (95, 173, 174, 191, 192).
VA is stored in tissues primarily in the form of retinyl esters (RE). Although a high dose
of VA administration is able to improve VA status, it has limited capability of increasing
RE concentration in the lung, because most of the supplemented VA is distributed to the
liver, the major organ for VA storage. Our previous study has suggested that a nutrient-
metabolite combination of VA and RA, VARA, mixed in a 10:1 molar ratio, shows great
potential for elevating RE level in neonatal lung (160, 161). When VARA was repeatedly
4.2 INTRODUCTION

83
administered to neonatal rats throughout the lung septation period, the increase was
augmented not only in a synergistic but also in a cumulative manner (Fig. 2.2). In a dose
dilution study, the VARA dose diluted to 25% of its original concentration still showed
same ability of increasing lung RE as that of the standard VA dose (160). The standard
dose of VA and RA used in our previous study was based on prior usage of VA in human
infants and RA in neonatal rats (160, 161). These findings reveal great potential of RA to
promote lung RE formation, and provide a more efficient method for improving lung VA
status.
In this study, we continued to test RA’s ability to induce lung RE formation during the
postnatal period. In contrast to the past dose preparation in VARA dilution study, we
only reduced the concentration of RA but used the same amount of VA in each of the
standard and diluted doses. By using the same dose, we examined the effects of a single
treatment and multiple treatments on VA stores in the lungs of neonatal rats and found
that the synergy of increasing lung RE only could be observed after a single dose, not the
multiple doses. We also compared effects of RA and Am580, a stable analog of RA, to
test our hypothesis that the absence of VARA’s synergy in multiple treatments study is
due to the quick metabolism of RA. The results of the study revealed the great potential
of RA in promoting RE formation and provided a better understanding of retinoid
metabolism in neonatal lung during the postnatal period.
4.3 HYPOTHESIS AND AIMS

84
Hypothesis 1: A single dose of RA in more diluted concentration combined with the
same (standard) amount of VA still can produce a synergistic effect in increasing retinol
uptake and RE formation in neonatal lungs, compared with the standard VA dose.
Hypothesis 2: In contrast to a VA dose alone, diluted RA combined with VA still can
promote RE formation significantly in the lung after multiple treatments throughout the
lung septation period.
Hypothesis 3: Am580 substituted for RA in the combination dose will show a stronger
capability of inducing lung RE formation compared to the same amount of RA after
multiple treatments.
Aim 1: To test whether a single dose of RA in low concentration is still able to
promote high retinol uptake and RE formation in neonatal lungs.
In this study, the amount of RA in the VARA combination was reduced to different
concentrations as the dose of VA was kept the same. We tested how 5 d-old pups respond
to diluted doses at 6 h. The lung tissue were collected and subjected to HPLC for lung
total retinol quantification. The effects of VARA dose containing different levels of RA
were evaluated.
Aim 2: To test whether multiple doses of RA in low concentrations are still able to
promote high retinol uptake and RE formation in neonatal lungs.
In this study, the doses were prepared in the same way as in aim 1, and administered to
neonatal pups repeatedly throughout postnatal lung septation period on d4, d7, d11 and

85
14, respectively. Six hours after the last dose, lung tissue was collected and lung total
retinol from each sample was quantified by HPLC. The effect of multiple treatments
with diluted RA were analyzed and compared with the single dose.
Aim 3: To test effects of multiple treatments of VA and diluted Am580 in lung RE
formation.
In this study, we substituted Am580 for RA in multiple dose study. Diluted Am580
combined with the same amount of VA were administered to neonatal rats in the same
way as in aim 2. Lung total retinol from each sample was quantified by HPLC following
the treatments. We compared the effects of diluted Am580 and diluted RA doses in
promoting lung RE formation.
Protocols for all studies were approved by the Animal Care and Use Committee of
Pennsylvania State University. Pregnant female rats (Sprague-Dawley strain) were
purchased or obtained from natural mating. Dams were fed with semipurified AIN-93G
diet containing 4 mg retinol/kg (193), which is a VA adequate diet. After birth, pups
within litters were randomly distributed to each treatment group and sex was evenly
distributed as well. We conducted two RA dilution studies: single dosing (d 5) and
4.4 MATERIALS AND METHODS
4.4.1 Animals and Experimental Design

86
multiple dosing throughout the period of lung septation (d 4, 7, 11, 14) (Fig. 4.1). In the
single dosing study, 5 day-old neonatal rats received a single oral dose of oil (control),
VA alone, VARA (10% RA) and VA combined with a series of diluted RA (5%, 2%, 1%,
0.5%). In the multiple dosing study, neonatal rats received multiple doses of oil, VA,
VARA (10%, 2%, 1% RA). We also conducted a multiple Am580 dilution study which
was parallel to the multiple RA dilution study (Fig. 4.1). Before each treatment, the pups
were weighed and the volume of each dose provided to pup was adjusted to 0.4 µl/g
bodyweight.

87
Figure 4.1 Animal experimental design. Two studies were conducted to evaluate the
effect of a single treatment and multiple treatments of diluted RA in VARA dose. In the
single dose study, the doses of VA combined with a series of diluted RA were orally
supplemented to 5 d-old pups. In the multiple dosing study, multiple doses were given
consecutively on d4, d7, d11 and d14. Pups were randomly assigned to several groups to
receive oil (vehicle), VA and VARA doses (n=5-6/group). In the multiple dose study, an
Am580 dilution study was included to be compared with the effect of VARA. The dose
of VAAm580 was prepared and administered in the same way as in multiple VARA dose
study. Tissues were collected 6h after the last treatment.

88
The standard dose was prepared in the same way as in chapter 2. Please refer to the
chapter 2 for detailed information of how the standard doses were prepared. For the RA
or Am580 diluted doses, VA and acidic retinoids (at-RA and Am580) were first prepared
as a 2x stock solution in canola oil at 0.1 mmol/g and 0.01 mmol/g, and diluted with the
same volume of oil to form 1x oral dose of VA and 1x RA/1x Am580 respectively. The
2x stock solution were then mixed 1:1 to form the dose referred to as VARA10% and
VAAm10%. Similarly, 1x RA was further diluted with oil to form 0.4x, 0.2x and 0.1x
solutions to mix with the same volume of 2x VA stock solution to form the dose referred
to as VARA5%,VARA2%, VARA1% and VARA0.5%. The combination dose of VA
and Am580 was prepared in the same way to form VAAm10%, VAAm2% and
VAAm1% solutions. Canola oil was used as placebo (control).
The volume of each dose provided to pup was 0.4 µl/g body weight and the exact volume
of dose was determined based on the pup’s body weight. In the single dose study, a single
dose was given to pups on day 5. In the multiple dose study, multiple doses were given
on day 4, 7, 11 and 14. Pups were killed with carbon dioxide (CO2) 6 hours after the last
4.4.2 Dose preparation
4.4.3 Experimental methods and tissue collection

89
in both studies. The lung tissues were removed, trimmed and weighed. All samples were
frozen in liquid nitrogen immediately and then stored -80oC for later analysis.
Lung tissue was processed and lung total retinol was quantified in the same way as in
chapter 2. Please refer to the chapter 2 for the detailed information.
Chapter 2 provides the detailed information for statistical analysis.
A single dose of RA in more diluted concentration still promotes high lung RE
formation in younger pups
Our previous study suggested that VA and RA mixed with a molar ratio of 10:1 (RA is
10% in concentration relative to VA) for a single oral treatment is able to produce a
synergistic increase in RE contents in neonatal rat lung compared with VA given alone.
However, it was not clear whether the dose of RA is appropriate or excessive for the
synergy. In the present study, we tested whether RA in more diluted concentration still
4.4.4 Retinoid Analysis
4.4.5 Statistical Analysis
4.5 RESULTS

90
promotes high lung RE formation. In the dose of VARA, the concentration of RA was
diluted to different levels while the concentration of VA was kept the same as in our
previous study. Five-day old neonatal rats received a single oral dose of oil, VA and
VARA. For the treatment of VARA, RA was added in a series of diluted concentration
10%, 5%, 2%, 1% and 0.5%. Six hours after treatment, pup lung was collected and lung
RE was analyzed by HPLC. The result showed that RA concentration as low as 1% still
increased lung RE content to the same extent as 10% RA (Fig. 4.2). The increase did not
differ between VA and VARA0.5% treatments (Fig. 4.2). These results showed the
strong potential of RA in promoting lung RE formation.
Multiple doses of diluted RA does not promote high lung RE formation in older
pups while Am580 still does
Given the potential beneficial effects and safe use of long-term treatment with low
concentration of RA to neonates, in the following study we investigated the effect of
multiple doses of VA combined with diluted RA (RA was added in concentration of
10%, 2% and 1% relative to VA) administered to neonatal pups on d4, 7, 11 and 14. The
lung tissue was collected 6 hours after the last treatment and subjected to HPLC for total
lung retinol analysis. The result showed that, the dose of VA alone increase lung RE level
~3 folds higher than the control level. When VA combined with 10% RA, the dose
increase lung RE content significantly; this is consistent to the result of our multiple-
dosing study (Fig. 4.3). However, VARA2% and VARA1% produced no further increase
in lung RE compared with VA dose alone group (Fig. 4.3). These results indicated that

91
multiple doses with diluted RA have a much reduced ability to promote lung RE
formation. This might be explained by the transient effect of RA and quick depletion of
reduced RA mass in the lung by CYP26B1.
To interpret the lost synergy with multiple treatments, we substituted Am580 for RA in
the in the combination dose. After the multiple dose treatment, lung RE was increased
significantly by all of the diluted Am580 treatments, even with 1% Am580, which still
increased the RE level higher than VA given alone (Fig. 4.3). By contrast to VARA,
VAAm580 produced a stronger and longer induction of RE formation. This could be due
to the stability of Am580 which is resistant to CYP26B1 metabolism, thereby leading to a
sustained induction on retinol uptake and RE formation in the lung.

92
Figure 4.2 Lung RE concentration after a single treatment with VA alone, VARA
10%, VARA 5%, VARA 2%, VARA1% and VARA0.5%. Neonatal rats were
treated with an early single dose on d5. Lung total retinol contents were determined by
HPLC 6h after treatment. Data are presented as group means ± SEM; Groups were
compared by one-factor ANOVA. Data were transformed by log10 prior to ANOVA.
Tukey’s least square mean test was used after ANOVA. Groups with different letters
differed significantly, P<0.05.
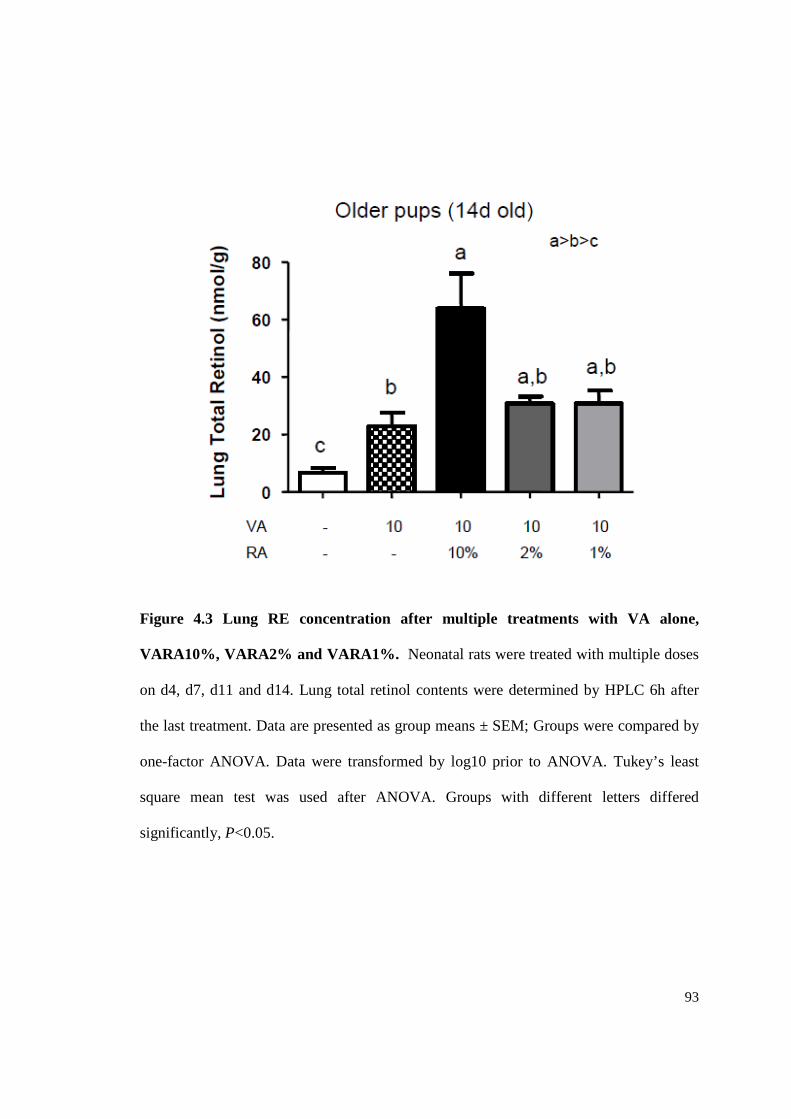
93
Figure 4.3 Lung RE concentration after multiple treatments with VA alone,
VARA10%, VARA2% and VARA1%. Neonatal rats were treated with multiple doses
on d4, d7, d11 and d14. Lung total retinol contents were determined by HPLC 6h after
the last treatment. Data are presented as group means ± SEM; Groups were compared by
one-factor ANOVA. Data were transformed by log10 prior to ANOVA. Tukey’s least
square mean test was used after ANOVA. Groups with different letters differed
significantly, P<0.05.
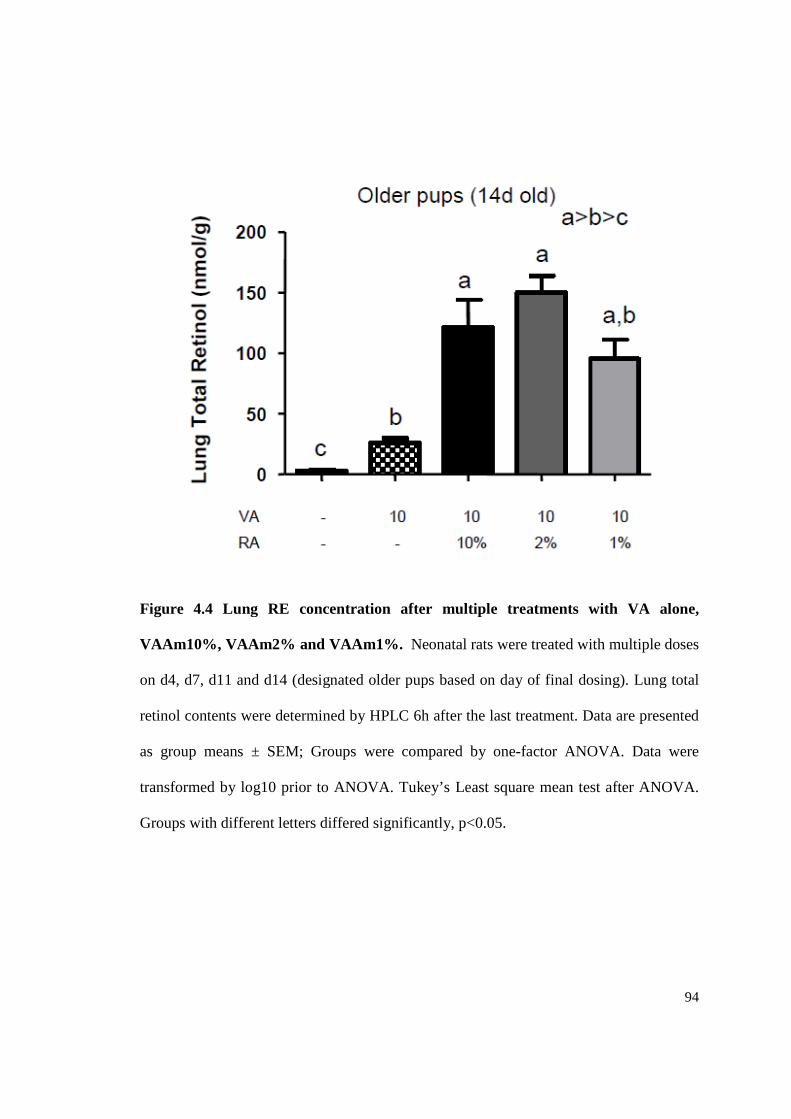
94
Figure 4.4 Lung RE concentration after multiple treatments with VA alone,
VAAm10%, VAAm2% and VAAm1%. Neonatal rats were treated with multiple doses
on d4, d7, d11 and d14 (designated older pups based on day of final dosing). Lung total
retinol contents were determined by HPLC 6h after the last treatment. Data are presented
as group means ± SEM; Groups were compared by one-factor ANOVA. Data were
transformed by log10 prior to ANOVA. Tukey’s Least square mean test after ANOVA.
Groups with different letters differed significantly, p<0.05.

95
In previous work, we showed that VARA, a combination of VA and 10% RA (we
referred to as VARA10% in the current study), increases RE content synergistically in the
lungs of neonatal rats (67, 160, 161). In this study, we examined the effects of dose with
same amount of VA but a reduced amount of RA. The amount of RA in our standard
VARA10% dose is based on the dose previously used to induce alveolar formation in
dexamethasone-treated neonatal rats during the lung septation period (99). The
mechanism by which VARA10% promotes higher RE content in the lung was
investigated and the findings suggested that 10% RA in the dose plays a role in
upregulating LRAT (retinol esterification) and STRA6 genes, which direct more of the
flow of supplemental VA into the lung tissue (67).
We started our study from a single dose study, in which a single dose of standard
VARA10% dose or VA combined with diluted RA (referred as VARA5%, 2%, 1%,
0.5%) was administered to rat pups on postnatal day 5. The increase in lung RE induced
by a single dose of VARA10% is consistent with our previous results (67). Diluted RA as
low as 1% in VARA dose still showed the effect equal to that of the VARA10% (Fig.2).
This result indicated that with the certain amount of VA, the amount of RA we used
before to produce the synergy in a short period (6 h) is saturating with respect to lung RE
formation, because a lower concentration of RA in the dose is as effective as standard
VARA dose for promoting lung RE formation.
4.6 DISCUSSION

96
Given the great potential of a single dose of VARA for elevating lung RE level at 6 h, we
expected to see the similar effect after multiple treatments with VA and diluted RA,
because in our multiple-dosing study, the lung showed great capability for RE storage
after multiple treatments with VARA10%. In the second study, we determined the effects
of the repeated doses of VARA10%, VARA2% or VARA1% throughout the lung
septation period. The standard VARA10% dose administered repeatedly to neonatal rat
has been demonstrated to be effective in increasing lung RE significantly in the multiple-
dosing study and in this study (Fig. 4.3). But VARA2% and VARA1% failed to show the
synergy that we observed in the single dose study. Both of these RA diluted doses did not
differ from an equal dose of VA only (Fig.4.3).
To interpret the discrepancy between the single- and multiple- dosing studies, we
substituted Am580 for RA in the VARA combination dose. Am580, a synthetic analog of
RA, which is known for its resistance to metabolism (73), has shown its ability to
regulate lung retinoid homeostasis and it replicates the effects of RA on lung RE
accumulation (67). In the present study, VA and Am580 were mixed in the same way
VARA was prepared (VAAm10%, VAAm2% and VAAm1%). Our results showed that
the lung RE level was elevated 3~5-fold by all VAAm doses compared with VA dose.
Based on the comparisons between single dose vs multiple dose, VARA treatments vs
VAAm treatments, we suggest several possible factors that might result in the absence of
VARA synergy in the diluted VARA formula. 1) A low amount of RA is depleted faster
than a higher amount of RA, thus losing its activity for LRAT and STRA6 induction; 2)
The lung during septation period is in high need of VA for development, therefore, the

97
temporarily improved RE content by diluted VARA is consumed rapidly; 3) The lung
responds to RA treatment more sensitively in younger rats than in older rats. To test these
hypotheses, a metabolic study to examine the distribution of supplemented retinol
combined with diluted RA, and a dose response study to test the effect of a single diluted
VARA dose at different time points during postnatal period would need to be done in the
future.
In summary, these studies were designed to test RA’s potential to induce lung RE
formation in the early time of life. The results showed that a single dose of RA in much
reduced concentration, combined with VA, still produced the same effect on RE
formation in young pups as the standard dose of RA. However, multiple treatments with
the same dose failed to increase lung RE formation synergistically, possibly due to the
transient action of RA and high demand for retinol in developing lungs. The higher
increase in RE storage caused by diluted Am580 provided evidence for our interpretation
of RA’s transient activity. Overall, we conclude that the combination of VA and diluted
RA still has great potential to induce RE formation at an early time after the treatment.
However, this effect is just transient. RA in reduced concentration is not adequate to
accumulate RE contents over time.

98
Chapter 5
LOCALIZATION OF LUNG RETINOID HOMEOSTATIC PROTEIN
STRA6, LRAT and CYP26B1 are the most important VA metabolism genes that regulate
retinoid uptake, storage and oxidation in the lung, but little is known about their protein
distribution pattern in lung tissue. Previous studies reported that the lipofibroblast is a
vitamin A-storing cell and STRA6 has been shown to be expressed in endothelial cells of
blood-organ barriers of other tissues. Due to the lack of specific antibody to CYP26B1
before, extremely limited information about this protein was available. Based on this
information, we hypothesized that LRAT is most likely expressed in lung lipofibroblast
where it can catalyze retinol esterification, while STRA6 and CYP26B1 are possibly
localized in the endothelial cells of the vascular network to mediate retinol uptake and
RA clearance. To test these hypotheses, we determined the localization of retinoid
homeostasis proteins by conducting immunofluorescence staining on lung sections from
neonatal rats. The results showed that LRAT is co-localized with adipocyte
differentiation-related protein (ADRP), the marker protein of lipofibroblast cells in lung
interstitium, STRA6 is co-localized with platelet endothelial cell adhesion molecule-1
(PECAM-1), the marker of endothelial cells, in the blood vessels and CYP26B1 is
exclusively expressed in the epithelium of bronchioles and the smooth muscle of small
5.1 ABSTRACT

99
airway and blood vessels. Overall, our results demonstrate the localization of LRAT in
lipofibroblasts and STRA6 in endothelial cells, while in addition, for the first time, we
report the localization of CYP26B1 in bronchiole epithelium. This information might
prove helpful for better understanding retinoid metabolism mechanisms in neonatal rat
lungs.
The lung is a complex tissue composed of over 40 different cell types which serve
various roles in lung development and functions. The most common lung cells include
alveolar epithelial type I, type II cells, endothelial cells, macrophage, and fibroblast, etc
(Fig. 5.1). It is known that VA plays an important role in normal lung development and
function. Vitamin A homeostasis in regulated by several important genes, LRAT,
CYP26B1 and STRA6 in the lung (67). But very limited information about the
localization and expression patterns of these proteins in the lung is available. It is
important to study this because a good understanding of the spatial and temporal
expression pattern of these proteins will provide insight on how VA is taken up, stored
and oxidized within the lung tissue.
The lung fibroblast is one type of the pulmonary mesenchymal cells that synthesizes
extracellular matrix and collagen. It plays an important role in maintaining the structural
framework for lung tissue. At the same time, there is coordinate signaling between the
epithelial cells and fibroblasts that determines normal alveolar development and mediates
5.2 INTRODUCTION

100
tissue injury repair via epithelial-mesenchymal interactions (133, 143-147). Lung
fibroblasts also provide lipid substrate for the synthesis of surfactant phospholipid in type
II alveolar epithelial cells (148). There are different populations of fibroblasts which are
classified based on the biochemical and functional characteristics of each type. The
lipofibroblast is one population that shows expression of a marker glycoprotein molecule,
Thy-1, on the cell surface (194, 195), and is characterized by abundant lipid droplets and
a high content of glycogen (137). It is located in the alveolar interstitium close to the
central region of the alveolar septum (137). Lipofibroblasts start to be present in rat lungs
around gestational day 16, and after that time the lipid content in the whole lung tissue
starts to rise significantly, especially between gestational days 17 and 19 and at birth, and
peaks during the second postnatal week (196). Compared with Thy-1 negative
fibroblasts, Thy-1 positive lipofibroblasts appear to be more abundant in the lung and to
perform a repair function after injury (194). Lipofibroblasts are also known as vitamin A-
storing cells containing retinyl esters (RE). In an animal study, feeding a VA sufficient
diet resulted in a great increase in lipid droplets size and number in lung fibroblasts (141).
Okabe et al identified that VA is contained in the isolated lung lipofibroblast and still is
present in the progeny cells after cell divisions (134). In rats, significant increase in the
lung RE storage starts from the late gestation, which is paralleled to lipid storage in the
lungs, peaks right before birth, and the depletion of this store begins and continues into
the early postnatal life (92), indicating a broad and rapid utilization of RE store for early
postnatal development of the lung. McGowan et al. have shown that the contents of
retinol and RA, the biologically active forms of vitamin A, in the lungs of rats increases
in early postnatal life (94). Concurrently, the level of cellular retinoic acid-binding

101
protein (CRABP) increases in whole lung tissue and lung lipofibroblasts (47). Also, the
increases of RARβ and RARγ were observed in lipofibroblasts from late gestational
period to early postnatal period (197, 198). It was also demonstrated that, later on,
lipofibroblasts are able to produce and secrete at-RA at the sites of secondary septum
projections, thereby providing endogenous at-RA for other lung cells (135). Overall,
these studies strongly suggest that the lipofibroblast is the cell type where expression of
the LRAT gene is localized and where it is able to generate the biological activities of
acidic retinoids in lung alveoli.
STRA6, a cell membrane protein, was recently identified as a cell-surface receptor for
RBP that mediates uptake of retinol into cells (33). Both in situ hybridization (ISH) and
immunohistochemistry (IHC) studies suggested strong expression of STRA6 could be
detected in blood-organ barriers which are made of epithelial or endothelial cells, for
example, the choroid plexus and the brain microvasculature, RPE cells in the eye, Sertoli
cells in the testis and kidney, spleen, etc. (63). These barriers play a role in facilitating
uptake of many nutrients, including retinol. Although the expression of STRA6 in the
lung is quite low, our previous studies showed that oral supplementation with RA could
increase the level of STRA6 mRNA in neonatal rat lungs (67). Based on its function and
localization in other organs, STRA6 is most probably expressed in the lung vascular
system, especially the small arterioles and the capillary network which forms the air-
blood barrier with the alveolar epithelium.
Due to the lack of proven specific antibodies against CYP26 proteins before, limited
information about CYP26 protein distribution in any organs has been demonstrated. As

102
the major function of CYP26 is to control cellular exposure to RA by inactivating RA in
cells, it is possible that CYP26B1 protein is expressed in endothelial cells of capillary in
the lung, where it might mediate the excretion of polar metabolites of RA into blood
stream. It is also possible that CYP26B1 is expressed in the lung epithelial cells where
RA activity is highly required.
We hypothesized that LRAT is expressed in the lipofibroblastic cells–a lung retinol
storing cell (135), CYP26B1 and STRA6 are expressed in the lung vascular endothelial
cells, where they might play a role in controlling the uptake of retinol and excretion of
RA. In our previous studies, these genes could be highly induced in the neonatal rat lung
by a short-term treatment (6h) with RA. However, this action is transient because RA is
quickly metabolized by the product of the induced CYP26 gene. Based on previous
findings, we speculate that multiple treatments with RA over a period of time (throughout
the postnatal period) might maintain a higher expression level of retinoid homeostatic
proteins compared with a 6-h single dose treatment.
Aim: To determine the spatiotemporal expression pattern of the retinoid
homeostatic proteins LRAT, CYP26B1 and STRA6 in neonatal rat lung.
We tested this aim by giving multiple treatments of VARA to neonatal rats throughout
the stage of lung alveolarization, and conducting immunofluorescence staining with lung
5.3 HYPOTHESIS AND AIMS

103
sections from treated rats. Aim 1a): We localized spatial expression of the LRAT,
STRA6 and CYP26B1 proteins in the lungs. Aim 2b): We compared the change in
protein expression in the absence or presence of VARA treatment throughout the
postnatal period.
The neonatal rats used in this study were under a protocol approved by the Institutional
Animal Use and Care Committee of the Pennsylvania State University. Neonatal pups
were delivered and nourished by mother rats fed a VA adequate diet. Throughout the
period of lung septation (days 4-14), the pups received 4 oral doses of VARA, one each
on day 4, 7, 11 and 14. Before each treatment, the pups were weighed and the dose was
adjusted to 0.4 µl/g bodyweight.
To inflate the lungs, the trachea was cannulated with a plastic needle. Optical cutting
temperature (OCT) was diluted 3:1 and used for inflation of the lung (1-2 ml) through the
5.4 MATERIALS AND METHODS
5.4.1 Animals
5.4.2 Preparation of lung tissue

104
needle until the lungs filled the pleural cavity. The trachea was then tied and the lungs
were dissected out and quickly minced into small pieces, subsequently. The freshly
dissected lung tissue was placed into pre-labeled plastic freezing molds and covered with
OCT. Filled molds were snap-frozen on dry ice and frozen blocks were stored at -80oC
until ready for sectioning.
Frozen tissue blocks were transferred to a cryotome cryostat (Shandon SME Cryotome
Cryostat) and the temperature of the frozen tissue block was equilibrated to the
temperature of the cryotome cryostat. Then the frozen lung tissue was cut into 6-µm
sections and mounted onto superfrost plus (Fisher) slides. Sections were dried overnight
at room temperature and then stored at -80oC for later use. Frozen tissue sections were
warmed and air dried at room temperature, fixed in pre-cooled acetone (-20oC) for 10
min and then placed at room temperature for > 20 min to allow the acetone to evaporate.
They were then washed in 10 mM phosphate buffered saline (PBS) at a neutral pH for 2
changes, 5 min each, and incubated in 0.3% H2O2 solution in PBS at room temperature
for 10 min to block endogenous peroxidase activity. Afterwards, sections were washed in
PBS again 2x5 min, and blocked with blocking buffer (10% fetal bovine serum in PBS)
in a humidified chamber at room temperature for 1 h. After blocking, sections were
incubated with the mixture of two primary antibodies in a humidified chamber overnight
at 4°C. On the second day, the sections were washed and incubated in the mixture of two
5.4.3 Preparation of tissue sections

105
fluorescent-conjugated secondary antibodies in 1% BSA in PBS in the dark for 30 min at
room temperature. After washing in PBS, a drop of anti-fading fluorescent mounting
medium (VECTASHIELD HardSet Mounting Medium with DAPI) was dispensed onto
the section to mount the tissue, and they were then cover slipped. Sections were viewed
using an Olympus Fluoview 1000 Confocal Laser Scanning Microscope (Olympus
America Inc.; Lehigh Valley, PA). Images were analyzed using Fluoview software.
The following antibodies were used as primary reagents for the detection of LRAT,
STRA6, CYP26B1 and PECAM-1 and ADRP as the marker proteins of endothelial cells
and lipofibroblasts, respectively.
Anti-STRA6, polyclonal antibody made in goat; anti rat, mouse, was obtained from
Sigma-Aldrich (St Louis, MO).
Anti-LRAT, monoclonal antibody made in mouse; anti human, was obtained from
Sigma-Aldrich (St Louis, MO).
Anti-CYP26B1, polyclonal antibody made in rabbit; anti human, was obtained from
Thermo Scientific Inc. (Rockford, IL).
Anti-PECAM-1, monoclonal antibody made in mouse; anti rat, was obtained from Santa
Cruz Biotechnology (Santa Cruz, CA).
5.4.4 Antibodies for section immunostaining

106
Anti-ADRP, polyclonal antibody made in goat; anti mouse, provided by the laboratory of
Dr. Constantine Londos, NIH.
Secondary fluorescent conjugated antibodies included:
Alexa Fluro 488-conjugated anti-goat IgG
Alexa Fluro 488-conjugated anti-mouse IgG
Alexa Fluro 568-conjugated anti-mouse IgG
Alexa Fluro 594-conjugated anti-rabbit IgG
These antibodies were obtained from Molecular Probes, Inc. (Eugene, OR).
We investigated the distribution of retinoid homeostatic proteins by performing
fluorescent immunostaining of lung sections from 14-day old neonatal Sprague-Dawley
rats and using confocal laser scanning microscopy to analyze the results. To test the
hypotheses that LRAT is exclusively or nearly exclusively expressed in lung
lipofibroblasts and CYP26B1 and STRA6 are expressed mainly in lung vascular
networks, we stained ADRP, a marker protein of lipofibroblasts, and PECAM1, a marker
protein of endothelial cells, simultaneously with their target proteins respectively.
5.5 RESULTS

107
Staining with respective 2nd antibody did not show any immunoreactivity (Fig 5.2.P, Q,
Fig 5.3M, N, Fig. 5.4Q, R).
Confocal laser microscopy revealed that in the lungs of 14-day old rats moderate
immunoreactivity of LRAT (red) was present in the interstitial region. The
immunostaining of ADRP, a signature lipofibroblast marker protein (green), overlapped
with LRAT, indicating the localization of LRAT in lung lipofibroblast cells (Fig. 5.2.E,
J). The immunoreactivity of LRAT was not detected in alveolar areas, bronchioles or
blood vessels (Fig. 5.2.O).
PECAM-1, a marker protein of endothelial cells, was clearly detected (red) in the
endothelium of small blood vessels, capillaries that surround connective tissues and
alveolar sacs, indeed everywhere that might be in contact with the vascular network (Fig.
5.3.B, F, J). The detected immunoflorescence of STRA6 (green) in the lung suggested
that this protein is expressed in the same area where PECAM-1 is expressed (Fig. 5.3.C,
G, K). The merged confocal images showed the co-localization of the STRA6 and
PECAM-1 expression, indicating the expression of STRA6 in the endothelial cells of
capillaries and blood vessels (Fig. 5.3.D, H, L).
Similar to the result for STRA6 localization, PECAM-1 staining (green) was easily
detected almost everywhere that is in contact with the vascular system. However, the
merged images showed a completely non-overlapping of CYP26B1 (red) and PECAM-1
immunostaining in the lung. In contrast to PECAM-1 staining, CYP26B1 was primarily
detected in the epithelium of bronchioles, terminal bronchioles and respiratory

108
bronchioles (Fig. 5.4.C, G, K). Less intense staining could be detected in the smooth
muscle of bronchioles, terminal bronchioles walls, as well as in the smooth muscle in the
media of small arteries and arterioles (Fig. 5.4.C, G). However, there is no detectable
staining of CYP26B1 in alveolar sacs and lung interstitium (Fig 5.4.O). Merged images
show non-colocalization of the two proteins.

109
Figure 5.1 Localization of LRAT protein in 14d old rat lungs. Confocal laser scanning
microscopy of immunoreactivity for LRAT (red; C, H, M) on 14 day old rat lungs in
combination with ADRP (green; B, G, L) and DIC display (A, F, K). Merged images (E,
J, O) indicate that ADRP and LRAT proteins are co-expressed and slightly detected in
the interstitium of the lung (black arrows in E and J). But the immunoreactivity of both
proteins is absent in most alveolar area, air-ways and blood vessels (O). Figure P and Q
show the negative control of the secondary antibody and DAPI for nuclear staining.
(as=alveolar sac; B=bronchiole; V=vessel)

110
Figure 5.2 Localization of STRA6 protein in 14d old rat lungs. Confocal laser
scanning microscopy of immunoreactivity for STRA6 (green; C, G, K) on 14-day old rat
lungs in combination with PECAM-1 (green; B, F, J) and DIC display (A, E, I). In the
lung, PECAM-1 is present in whole vascular network, especially the endothelium of
small blood vessels (F, J). Merged images indicate that STRA6 co-localize with PECAM-

111
1 in the vascular network. Figure M and N show the negative control of the secondary
antibody and DAPI for nuclear staining. (as=alveolar sac; B=bronchiole; TB=terminal
bronchiole; RB=respiratory bronchiole; V=vessel)
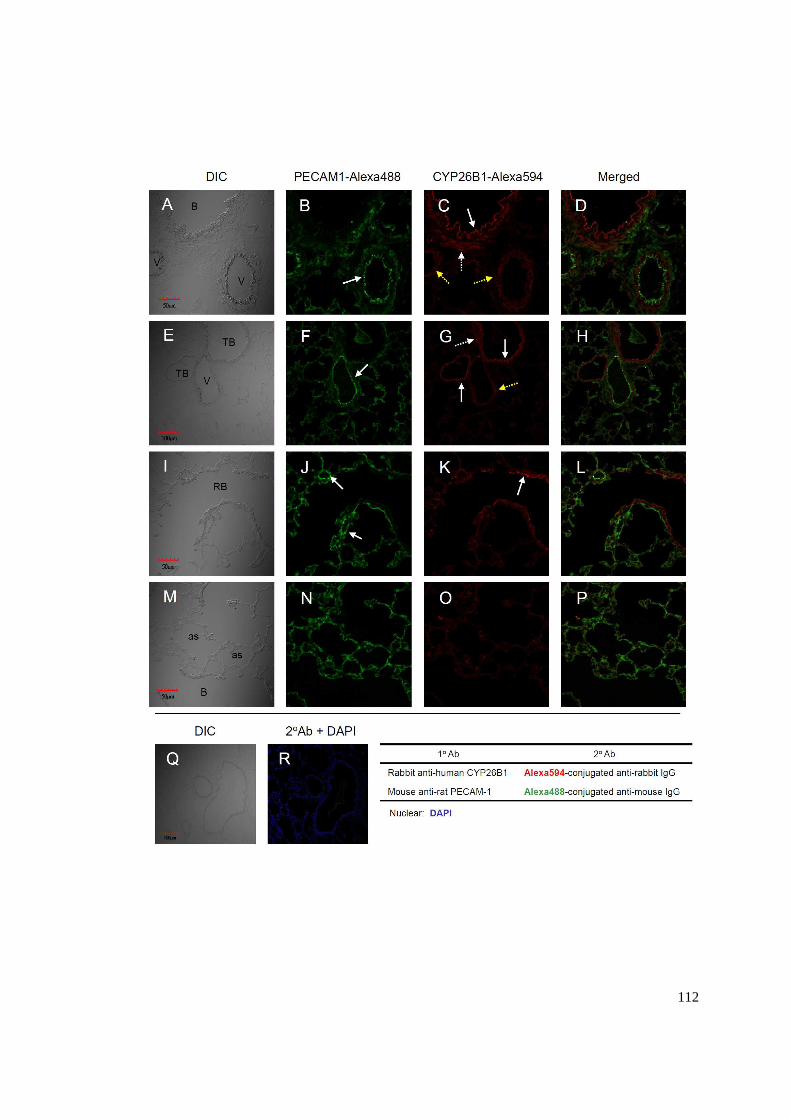
112

113
Figure 5.3 Localization of CYP26B1 protein in 14d old rat lungs. Confocal laser
scanning microscopy of immunoreactivity for CYP26B1 (red; C, G, K, O) on 14-day old
rat lungs in combination with PECAM-1 (green; B, F, J, N) and DIC display (A, E, I, M).
In the lung, PECAM-1 is present in whole vascular network, especially the endothelium
of small blood vessels (white solid arrows in B, F and J). CYP26B1 is primarily
expressed in the epithelium of bronchioles, terminal bronchioles and respiratory
bronchioles (white arrows in C, G, K). Moderate expression of CYP26B1 could be
detected in the smooth muscle of bronchiole and terminal bronchiole walls (white dashed
arrows in C and G), as well as the smooth muscle in the media of small arteries and
arterioles (yellow dashed arrows in C and G). Merged images show that CYP26B1 is not
overlapped with PECAM-1, indicating no expression of CYP26B1 in vascular network
(D, H, L). CYP26B1 immunoreactivity is also absent in alveolar area (O, P). Figure Q
and R show the negative control of the secondary antibody and DAPI for nuclear
staining. (as=alveolar sac; B=bronchiole; TB=terminal bronchiole; RB=respiratory
bronchiole; V=vessel)

114
Over the past years, significant progress has been made in elucidating the mechanism of
retinoid metabolism in various tissues, and improving the understanding of the roles that
retinoid homeostatic enzymes play in the lung during development. However, a clear
profile of the spatial and temporal expression pattern of these lung retinoid homeostatic
proteins has not been obtained yet. In our study, we used 14 d-old neonatal rats, because
the lungs have just experienced a rapid process of alveolization. During this stage, RA
action is required for normal alveolar development. Therefore, VA homeostasis is
important to maintain. Where is VA taken up into the lung tissue? Where is VA locally
stored? And where is RA cleared? Using immunofluorecence staining, our present
experiments report the localization of the retinoid homeostatic proteins, LRAT,
CYP26B1 and STRA6, in neonatal rat lung. The presence of LRAT in interstitial tissue
corresponds to the presence of ADRP, a marker protein of lipofibroblast, suggest the
possible localization of LRAT in lipofibroblasts. STRA6 expressed in the vascular
endothelium could ideally play a role in taking up VA from the blood stream. Our results
also clearly indicate the expression of CYP26B1 in the epithelium of bronchioles and
smooth muscle of small airways and blood vessels.
5.6 DISCUSSION

115
It is well known that CYP26 functions to prevent the action of RA and deplete RA in
tissue. But, significant gaps exist in understanding the spatio-temporal expression of the
CYP26 enzymes in lung tissue. The primary oxidized metabolites of at-RA are polar
metabolites that could be eliminated through the blood stream, thus we hypothesized that
CYP26B1 is localized in the endothelial cells of the vascular network, which is believed
to be an ideal place for a ready clearance of RA. However, in our experiment results, the
immunostaining of CYP26B1 protein suggests its presence in the epithelial cells of
bronchioles, not the endothelial cells. The possibilities to explain this are that, first,
CYP26B1 protein is produced locally in epithelial cells to prevent RA action in this
tissue, and excess RA is trafficked to epithelial cells for metabolism, or second, that
CYP26B1 produced in other cell types is transferred to epithelial cells to clear excess RA
here. But we are not sure why CYP26B1 is only expressed in epithelium of bronchioles.
One possibility is that the basal level of CYP26B1 is too low to be detected in other
areas, like alveolar walls, or even vascular endothelial cells, and a longer time might be
required for detectable expression of CYP26B1 after RA or Am580 administration.
Furture studies with longer time of treatment and gene localization study (e.g. ISH) are
needed to further determine where the CYP26B1 gene is expressed and the enzyme is
localized.
Our previous study reported that the mRNAs of retinoid homeostatic genes in the
neonatal rat lungs could be quickly increased by oral treatment of VARA as early as 6h
(67). However, we did not observe any changes in protein level between oil (control) and
VARA treated rat lung samples (data not shown). This could be due to the late protein

116
expression after gene induction. Although the signals of CYP26B1, LRAT and CYP26B1
proteins could be detected in the lung and marker proteins provide evidence and
information of where they are localized, it is still very important to see the changes in
protein level that are regulated by RA, if they exist. To solidify our findings on the
localization of lung retinoid homeostatic proteins and their responses to RA regulation,
protein levels in lung tissue with a longer time of treatment should be examined.
In conclusion, based on immunofluorescence staining, we have described a possible
distribution of retinoid homeostatic proteins in neonatal rat lungs. The dietary VA
transported to the lungs is taken up by STRA6 which is expressed in endothelial cells,
and then VA is esterified and stored in lipofibroblast cells by LRAT which is produced
locally. The excess RA is metabolized by CYP26B1 in epithelial cells of bronchioles.
This documentation provides some new clues for a better understanding of overall VA
homeostasis in the neonatal lung.

117
Chapter 6
DISCUSSION
VA is essential for the growth and development of the lung. During the late gestation
(day 14 – 18), lung RE concentration increases significantly and then starts to deplete
before birth and throughout the postnatal period (92). And this depletion is accompanied
by a morphological maturation of the lungs (93), indicating an increased demand for
retinoids for the process of postnatal lung development.
Given that VA has shown its essential role in lung development, supplementation of VA
during late fetal life and early postnatal life to maintain adequate lung VA storage is of
great biological advantage. Before birth, the fetus is exclusively dependent on the
maternal supply of VA. When a large dose of retinyl palmitate (50,000 I.U.) was given to
pregnant rat on day 16, the levels of RE in the lungs of fetus and neonatal rats were
significantly elevated (159). Although this supplementation also increased the RE
concentration in the liver of the offspring, this increase was smaller than the increase in
the lung, suggesting a more efficient mechanism of VA uptake in the fetal lung during the
late gestation. Prematurity in infants is usually associated with severe VA deficiency in
the lungs, which might be attributed to the shortage of maternal VA supply during late
6.1 VITAMIN A SUPPLEMENTATION DURING LUNG SEPTATION PERIOD

118
pregnancy and the relatively immaturity of the liver for RBP synthesis. Animal and
human studies have shown that VA supplementation of lactating mothers during the
postnatal period can improve VA status in their offspring (199-201). Direct oral VA
supplementation of the offspring also can increase the lung RE level. However, the
uptake of retinol by the lung is not as sufficient as by the liver (160, 161). This is because
the liver, as the major organ for VA storage, stores most of excess dietary VA. Other
studies in adult rats treated with RA, the principal active metabolite of retinol, have
shown that the storage of VA in the lung can be increased by RA treatment as well (202,
203). Since RA can not be reversibly reduced to produce retinol, the increased amount of
RE in the lungs maybe originate from the liver RE store, and RA may play a regulatory
role facilitating retinol uptake and RE formation in the lung.
We previously tested the effects of VA and RA combination (10:1 molar mixture of VA
and RA) in promoting lung RE formation and the results showed that the administration
of this nutrient-metabolite combination for 3 consecutive days (p5-7) can synergistically
increase lung RE, at least 4-fold more than the same amount of VA dose given alone
(160). The VA dose used in our studies is based on the dose (50,000 IU) of retinol used
in human studies that has been shown to reduce mortality of newborns in the first year of
life (10, 162). The dosage we selected for RA is based on previous usage shown to induce
lung septation in neonatal rats (97). In a metabolic study, we found that the
administration of RA direct more of the orally supplemented [3H]retinol into the lung
(161).

119
Following these findings, we investigated the molecular mechanism by which VARA
promotes lung RE formation. We examined the expression of lung retinoid homeostatic
genes, LRAT, CYP26B1 and STRA6, as they can control the balance of the retinoid
metabolism by mediating cellular retinol uptake and converting retinol to its storage for,
or by oxidizing RA to inactive polar metabolites. We found that all of these three gene
were upregulated by RA administration at 6 h (67) and concluded that the redirection of
VA flow into the lung by VARA is a result of the upregulation of LRAT and STRA6
genes. The other evidence that demonstrated the cooperation of LRAT and STRA6 in
mediating retinol uptake and storage is from the study of retina and retinal pgment
epithelial cells, in which STRA6 and LRAT are co-expressed. These cells take up retinol
more efficiently, suggesting a driving force for retinol uptake resulting from the
conversion of retinol into RE by LRAT (33). However, by 12 h after dose administration,
the regulatory activity of RA is no longer evident, and this might be attributed to the
simultaneous induction of CYP26B1 by RA at 6 h which prevent RA’s action by
oxidizing it (67).
The experiments described above were designed to examine the response to VARA at
early postnatal life (p7~p8) and over relative short times (6 h after treatment). A kinetic
study checked the consumption of lung RE store in the pups treated with VARA on days
5, 6, and 7, showed that although RE declined between days 8 and 16, the VARA-derived
RE store was retained during the period of postnatal lung septation (161). In the present
studies, we determined the effect of multiple treatments with VARA throughout the
septation period on promoting RE formation in neonatal rat lung and found that multiple

120
treatments increased lung RE contents in a synergistic and cumulative manner (Fig. 2.2).
In this study, we included the usage of Am580, a stable analog of RA, to compare with
RA. From the results, VARA and VAAm did not differ from each other with respect to
lung RE formation after a single dose, but VAAm produced a stronger effect than VARA
in increasing RE after multiple doses (Fig. 2.2). The data of gene expression provides a
good explanation for the difference between VARA and VAAm treatments that the
constant induction on lung retinoid homeostatic genes by Am580, which can persist even
24 h after the dosing leads to an exaggerated effects of VARA on lung RE promotion
(Fig. 2.3, 2.4).
Retinoids have been implicated in lung development by promoting alveolar septation,
angiogenesis, and surfactant synthesis (172). RA has also shown promising effects in
lung repair by regulating genes involved in lung functioning, cell proliferation,
differentiation, cell-cell communication, and cell-matrix interactions (204). Products of
such genes like surfactant proteins, collagen, fibronectin, tropoelastin, β-laminin, as well
as VEGF are all required for normal lung structure and function (204, 205). Given that
RA has shown its regulatory activity on these important lung functional and structural
genes, we assessed the impact of VARA treatments on these genes. The results of this
experiment revealed that the expression of lung functional and structural genes was not
affected by either a single dose or multiple doses of VARA (Fig. 2.5). It is not surprised
to see this result because we used a model of full-term healthy rat pups which were
delivered and nourished by mother rats fed a VA adequate diet. The lungs of these pups
were undergoing or had completed the process of development by the time the gene

121
expression level was analyzed. This result suggests that under physiological condition,
our VARA treatments did not perturb the expression of the important lung structural and
functional genes.
In summary, VARA is more effective than VA alone or RA alone in increasing RE
content in neonatal lungs, and it has little effect on lung structural and functional genes.
These properties make VARA a promising therapeutic option in clinic medicine for
increasing RE formation.
In the fasting state, >95% of retinoid in the circulation is found as retinol bound to RBP,
while the rest is comprised of a small portion of RA bound to albumin and RE contained
in chylomirons. In the fed state, the proportion of RE in the circulation is significantly
increased (206). The precise mechanism of how retinoids are taken up by lung tissue is
not defined yet. But this process possibly involves the direct uptake of retinoids in all
these three forms.
The uptake of retinol bound to RBP mediated by a cell-surface receptor for RBP has been
hypothesized for many years until recently Kawaguchi et al. provided solid evidence
which demonstrated the existence of a RBP receptor, STRA6. STRA6 is a
transmembrane protein, which shows high affinity to RBP and facilitates uptake of
retinol in most of tissues (33). In vitro and in vivo studies have shown that the STRA6
6.2 MOLECULAR MECHANISM OF RETINOL UPTAKE INTO THE LUNG

122
gene can be upregulated by RA (63-67). However, in the multiple treatment study, the
expression level of lung STRA6 is not evidently increased by either RA or Am580 (Fig.
2.5). This result suggested that although STRA6 is involved in the process of retinol
uptake, it may functions at a basal level to transport retinol into lung cells, no matter
whether the RA action is present. In contrast, LRAT gene expression is rapidly and
significantly induced by RA (Fig. 2.2), suggesting that LRAT is the more important
contributing factor that leads to RE formation in lung cells.
RE or retinol present in the circulating chylomicrons may be another important source for
lung RE store after meal or oral VA supplementation. One of our previous studies
suggested that although supplementation with VA or VARA at early postnatal age can
elevate RE markedly in the lungs of neonatal rats, and the plasma retinol was maintained
at the same level by either VA-deficient or VA-adequate diet after weaning, these rats are
still dependent on the VA-adequate diet to prevent the drop of lung RE store during the
postweaning growth period (45). These results imply that chylomicron-associated RE or
retinol served as a secondary source of VA for lung RE store, in addition to that provided
by retinol bound to RBP. Previous study had shown that LPL is able to hydrolyze
chylomicron-associated RE to retinol and facilitate retinoid uptake by adipocytes (46).
However, the direct evidence for the uptake of chylomicron-associated RE in the lung is
still lacking, and whether this process is regulated by RA is not clear.
6.3 THE POTENTIAL OF RA TO PROMOTE LUNG RE FORMATION

123
The concentration of RA in the VARA dose used for our animal experiments is 5 mM,
and the dose administered to neonatal pups is 2 nmol per gram of body weight. Both the
single dose and the multiple doses of VARA can increase lung RE content significantly
in neonatal lungs (Fig. 2.2). Is the concentration of RA over-saturated (more than
necessary) for producing the same effect? In the RA dilution study, we tested the
potential of RA to induce lung RE formation by diluting its concentration. We found that
the RA reduced down to 0.5 mM can still synergize with the same (standard) amount of
VA to increase lung RE in younger pups (5-day old) after a single dose. Interestingly, this
effect is not observed in older pups (14-day old), which received multiple doses (Fig.4.3).
Our previous 12 h experiment had suggested that the activity of RA is just transient
because it is quickly oxidized. Therefore, we speculated that the loss of synergy in the
multiple dose study is due to the quicker elimination of the small amount of RA by lung
or other tissues. This hypothesis was supported by a parallel multiple dose experiment in
which RA was substituted by Am580. The synergistic effect of VAAm was at least
partially retained with the 1% concentration of Am580 (Fig. 4.4), indicating a prolonged
regulatory activity of Am580.
Nevertheless, some other possible mechanisms that leading to the discrepancy between
the single dose study and the multiple dose study may also exist. For example, the lung
tissue may respond to RA or Am580 regulation with different sensitivity at different
ages, or there may be a high rate of RA clearance in the lungs of older rat pups. Whether
the VARA synergy in neonatal lung is time-dependent should be tested in the future.

124
It is well known that VA deficiency increases the risk of infectious diseases in young
children, which can lead to increased morbidity and mortality (207, 208). VA
supplementation has been shown to effectively reduce the severity of infectious diseases,
morbidity and mortality (209-212). Although it is already well established that retinoids
help maintain integrity of the immune system (213) and have shown anti-inflammatory
activity (214), little is known about how inflammation affects retinoid metabolism.
It has been reported that inflammation is able to alter the expression of several
cytochrome P450 genes in the liver (183). In a previous study in which adult rats were
treated with LPS and RA simultaneously, we found that LPS can abrogate the RA-
induced increase in CYP26A1 and CYP26B1 mRNA in the liver (185). Similarly, we
tested the effects of inflammation on retinoid homeostasis in neonatal lungs. In the LPS
study, we used a model of LPS-induced inflammation in neonatal rats to determine
whether the inflammation can prevent RE promotion by VARA, and opposes the
response of LRAT, CYP26B1 and STRA6 to RA in the lung, in which CYP26B1 mRNA
is more abundant. The results of our experiments suggested that the acute inflammation
just modestly decreased but did not prevent VARA-promoted RE formation in the lung,
and did not significantly alter the retinoid homeostatic genes expression pattern.
6.4 THE RELATIONSHIP BETWEEN INFLAMMATION AND RETINOID
METABOLISM IN NEONATAL LUNGS

125
Inflammation can reduce the production of RBP in the rat liver as well as the RBP-retinol
complex concentration in the plasma (215), which might contribute to the slight reduction
in the VARA-promoted RE formation. Despite this, the synergistic effect of VARA was
still very obvious in the inflammatory state. Our results also demonstrated that LPS
administration alone has little effects on the basal level of RE, and the expression of
LRAT, CYP26B1 and STRA6 genes in the lung, suggesting effective maintenance of
retinoid homeostasis under the inflammatory stimuli.
Alveologenesis is associated with dramatic changes not only in lung morphological
maturation but also in the metabolism of endogenous retinoids from storage forms, such
as RE, to oxidized metabolites (93, 94, 216). Important components involved in the
retinoid metabolism pathway include STRA6, a RBP receptor which mediate cellular
retinol uptake, LRAT, an enzyme which convert retinol to RE, retinoid binding proteins
(CRBP, CRABP), cellular proteins which regulate the biological action of retinol and
RA, retinoid oxidizing enzymes (RDH, RALDH and cytochrome P450), and RA
receptors (RARs, RXRs), nuclear proteins which act as transcription factors to regulate
downstream gene expression, etc. Previous studies have shown the distribution of CRBP-
I and CRABP-I proteins in lipofibroblasts in postnatal lungs and their upregulation
during alveolarization (94, 217), suggesting the appearance or production of retinol and
6.5 SPATIAL DISTRIBUTION OF THE RETINOID HOMEOSTATIC
PROTEINS IN NEONATAL LUNGS

126
RA in lung lipofibroblast cells. RAR expression also has been reported to be localized in
bronchial epithelium, bronchial and vascular smooth muscle, pleura, and scattered cells
within the alveolar regions (217), implying RA’s activity at these sites.
In the localization study, we chose to determine the spatial expression of LRAT,
CYP26B1 and STRA6 in postnatal lung. Our confocal microscopy images revealed that
STRA6 is specifically expressed in the endothelial cells, and LRAT is expressed in the
lipofibroblast (Fig. 5.2, 5.3). Interestingly, the distribution of CYP26B1 in bronchial
epithelium and smooth muscle of bronchioles and vessel is similar to the distribution of
RAR. It is well known that VA deficiency can result in morphological alteration in the
bronchial epithelium, a tissue known to be very retinoid sensitive (11, 77) and this effect
suggests that RA signaling is required for maintaining the integrity of airway epithelium.
Contradictorily, the CYP26B1 was identified strongly at the same site, which may mean
there is a prevention of RA’s action through RA nuclear receptors. It is not clear what
role CYP26B1 plays in this tissue, possibly, a mechanism with subtle controlling on
cellular concentration and biological activity of RA may exist.
The lipofibroblast cells in postnatal rat lung are primarily localized in the alveolar
interstitium. They are found adjacent to septa junctions when septa are being formed
(137). This cell type might play a role as a retinoid signaling center not only due to its
localization, but also because many components of the retinoid signaling pathway
including receptors and binding proteins are contained in these cells (94). The formation
of septa occurs in the rat mainly between postnatal day 4 and 14 (114). In our
experiments, we used the samples from 14-day old pups. The staining of a lipofibroblast

127
marker protein, ADRP, and LRAT protein is not evident in alveolar wall. This is might
be due to low formation of septa at the end of alveolar stage.
Based on our findings and information from previous studies, we propose a possible
model of retinoid metabolism in the lung (Fig. 6.1): plasma retinol bound to RBP is taken
up by lung tissue through a membrane receptor, STRA6, which is expressed in the
endothelial cells of capillaries. Subsequently retinol is transported to lipofibroblasts, at
where excess retinol will be converted to RE by LRAT for storage, or metabolized to
produce endogenous RA. The locally synthesized RA thereafter is secreted to
neighboring cells, including type II alveolar cells, endothelial cells and myofibroblasts, in
a paracrine manner to initiate or coordinate the alveolar septal eruption during
alveologenesis. In bronchial epithelium, where CYP26B1 is predominantly expressed,
the excess RA will be cleared to maintain the balance of retinoid metabolism pathway.
In conclusion, this experiment determined the spatial distribution of the retinoid
homeostatic proteins in the lung. Together with the information from previous studies,
our findings further demonstrate the key role of lipofibroblast as a retinoid signaling
center during alveologenesis, and provide better understanding of retinol uptake and
metabolism in neonatal lung tissue.
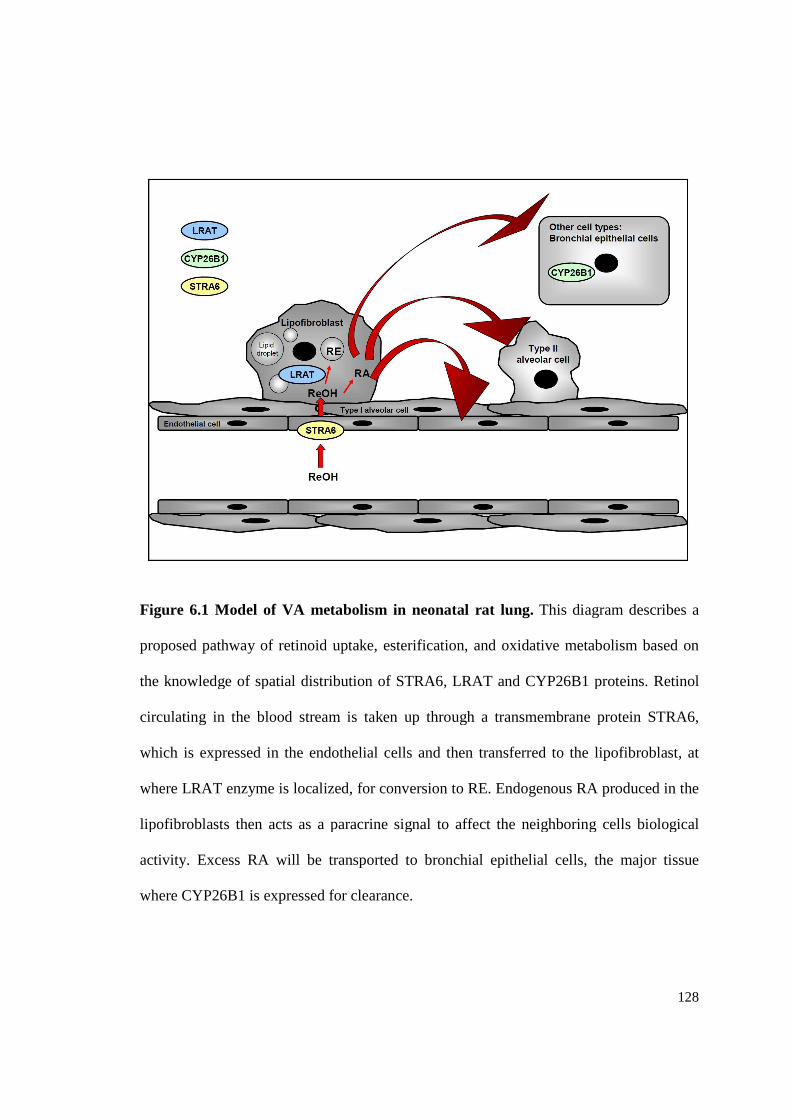
128
Figure 6.1 Model of VA metabolism in neonatal rat lung. This diagram describes a
proposed pathway of retinoid uptake, esterification, and oxidative metabolism based on
the knowledge of spatial distribution of STRA6, LRAT and CYP26B1 proteins. Retinol
circulating in the blood stream is taken up through a transmembrane protein STRA6,
which is expressed in the endothelial cells and then transferred to the lipofibroblast, at
where LRAT enzyme is localized, for conversion to RE. Endogenous RA produced in the
lipofibroblasts then acts as a paracrine signal to affect the neighboring cells biological
activity. Excess RA will be transported to bronchial epithelial cells, the major tissue
where CYP26B1 is expressed for clearance.

129
It is known that septation occurs mainly between the postnatal day 4 and day 14th
postnatal days in rats (7). Our present studies have determined the localization of retinoid
homeostatic proteins in the lungs of neonatal rat at the age of postnatal day 14, a time
point by which major septation process is completed. To better understand the regulation
of retinoid homeostasis in neonatal rat lungs, we need to investigate the temporal
expression of the retinoid homeostatic proteins during the postnatal septation period. We
believe that retinoid homeostatic proteins, similar to CRBP, CRABP and RARs, which
are tightly regulated in the postnatal lung (216), will be temporally and spatially
associated with alveolar formation, because lung VA status and metabolism undergoes a
significant change during this period.
In the RA dilution study, we observed that diluted RA combined with the same dose of
VA showed different effects in promoting lung RE formation in a single dose study and
multiple dose study. To interpret this phenomenon, we speculate that a response to RA
administration with a high sensitivity only exists at the very beginning of the postnatal
life, maybe even earlier than birth, because a high demand of retinoids is required for the
process of alveologenesis. To test this speculation, we need to examine the effects of a
single dose of VA combined with diluted RA administered at different postnatal ages. If
it is the case, supplementing with VA at an earlier postnatal time may be more
6.6 FUTURE DIRECTIONS

130
meaningful than at a later time, and we should then adjust the treatment strategy
according to a best timing to approach the more efficient method of increasing lung RE
level.
In the LPS study, we examined how inflammation affects retinoids metabolism in
neonatal lungs. However, there are many other factors that might impact lung retinoid
metabolism during postnatal stage, such as mechanical ventilation, dexamethasone
treatment, hyperoxia, pulmonary infections, etc, which are often associated with
neonates, especially newborns with very low body weight. In the future, it will also be of
interest to test the synergistic effect of VARA in promoting lung RE formation under
these conditions.
Since the present study has reported that RA combined with PIC promoted TT-induced
vaccine response in both adult and neonatal mice, the following question will be how to
apply to human vaccination.
The recent data in our lab reported that oral administration of retinyl palmitate (RP)
combined with a small amount of RA (1:10 molar of RP) synergistically increased lung
VA concentration about 5 to 10-fold in rat pups, and this potency was maintained even
though the dose of RP+RA was reduced to half or one fourth (Ross et al, unpublished
data). These data suggest a novel supplementation that VA combined with a small
amount of RA can not only reduce the dosage of VA but also increase the capacity of
improving VA status.

131
More importantly, the combination of VA and RA may be more effective than VA alone
to stimulate immune system in VA-sufficient population. Therefore, in the future study,
we will test the immunoregulatory effect of VA+RA in adult and neonatal mice.

132
REFERENCES
1. Joshi S, Kotecha S. 2007. Lung growth and development. Early Hum Dev 83: 789-94
2. Jeffrey PK. 1998. The development of large and small airways. Am J Respir Crit Care Med 157: S174-80
3. Hislop AA. 2002. Airway and blood vessel interaction during lung development. J Anat 201: 325-34
4. DiFiore JW, Wilson JM. 1994. Lung development. Semin Pediatr Surg 3: 221-32
5. Thurlbeck WM. 1982. Postnatal human lung growth. Thorax 37: 564-71
6. Zeltner TB, Burri PH. 1987. The postnatal development and growth of the human lung. II. Morphology. Respir Physiol 67: 269-82
7. Burri PH. 1974. The postnatal growth of the rat lung. 3. Morphology. Anat Rec 180: 77-98
8. Burri PH. 1984. Fetal and postnatal development of the lung. Annu Rev Physiol 46: 617-28
9. Williams MC. 1977. Development of the alveolar structure of the fetal rat in late gestation. Fed Proc 36: 2653-9
10. Humphrey JH, Agoestina T, Wu L, Usman A, Nurachim M, Subardja D, Hidayat S, Tielsch J, West KP, Jr., Sommer A. 1996. Impact of neonatal vitamin A supplementation on infant morbidity and mortality. J Pediatr 128: 489-96
11. Wolbach SB, Howe PR. 1925. Tissue Changes Following Deprivation of Fat-Soluble a Vitamin. J Exp Med 42: 753-77

133
12. Strickland S, Mahdavi V. 1978. The induction of differentiation in teratocarcinoma stem cells by retinoic acid. Cell 15: 393-403
13. Wilson JG, Roth CB, Warkany J. 1953. An analysis of the syndrome of malformations induced by maternal vitamin A deficiency. Effects of restoration of vitamin A at various times during gestation. Am J Anat 92: 189-217
14. Scrimshaw NS, Taylor CE, Gordon JE. 1968. Interactions of nutrition and infection. Geneva. World Health Organization
15. Hughes DA. 1999. Effects of carotenoids on human immune function. Proc Nutr Soc 58: 713-8
16. Kagechika H. 2002. Novel synthetic retinoids and separation of the pleiotropic retinoidal activities. Curr Med Chem 9: 591-608
17. Sommer A, Tarwotjo I, Hussaini G, Susanto D. 1983. Increased mortality in children with mild vitamin A deficiency. Lancet 2: 585-8
18. Goodman T, Dalmiya N, de Benoist B, Schultink W. 2000. Polio as a platform: using national immunization days to deliver vitamin A supplements. Bull World Health Organ 78: 305-14
19. Underwood B. 1994. Vitamin A in human nutrition: public health considerations. In The retinoids: biology, chemistry, and medicine., ed. RA Sporn MB, Goodman DS, pp. 211-27. New York: Raven Press
20. Sommer A, Katz J, Tarwotjo I. 1984. Increased risk of respiratory disease and diarrhea in children with preexisting mild vitamin A deficiency. Am J Clin Nutr 40: 1090-5
21. Stofft E, Biesalski HK, Zschaebitz A, Weiser H. 1992. Morphological changes in the tracheal epithelium of guinea pigs in conditions of "marginal" vitamin A deficiency. A light, scanning- and transmission-electron microscopic study under special breeding conditions appropriate to early vitamin A deficiency. Int J Vitam Nutr Res 62: 134-42

134
22. de Francisco A, Chakraborty J, Chowdhury HR, Yunus M, Baqui AH, Siddique AK, Sack RB. 1993. Acute toxicity of vitamin A given with vaccines in infancy. Lancet 342: 526-7
23. West KP, Jr., Khatry SK, LeClerq SC, Adhikari R, See L, Katz J, Shrestha SR, Pradhan EK, Pokhrel RP, Sommer A. 1992. Tolerance of young infants to a single, large dose of vitamin A: a randomized community trial in Nepal. Bull World Health Organ 70: 733-9
24. Werler MM, Lammer EJ, Mitchell AA, Brent RL, Hendrickx AG, Holmes LB, Miller RK, Watkins M, Moore C, Mulinare J. 1996. Teratogenicity of High Vitamin A Intake. N Engl J Med 334: 1195-7
25. Geelen JA. 1979. Hypervitaminosis A induced teratogenesis. CRC Crit Rev Toxicol 6: 351-75
26. Pinnock CB, Alderman CP. 1992. The potential for teratogenicity of vitamin A and its congeners. Med J Aust 157: 804-9
27. Nagy NE, Holven KB, Roos N, Senoo H, Kojima N, Norum KR, Blomhoff R. 1997. Storage of vitamin A in extrahepatic stellate cells in normal rats. J Lipid Res 38: 645-58
28. Smith JE, Milch PO, Muto Y, Goodman DS. 1973. The plasma transport and metabolism of retinoic acid in the rat. Biochem J 132: 821-7
29. Smith JE, Goodman DS. 1979. Retinol-binding protein and the regulation of vitamin A transport. Fed Proc 38: 2504-9
30. Soprano DR, Blaner WS. 1994. Plasma Retinol-Binding Protein. In The Retinoids, Biology, Chemistry and Medicine, 2nd ed.; Sporn, M.B., Roberts, A.B., Goodman, D.S., Eds.;. Raven Press: New York, NY, USA, 1994; PP. 257-282: Raven Press. 257-82 pp.
31. Episkopou V, Maeda S, Nishiguchi S, Shimada K, Gaitanaris GA, Gottesman ME, Robertson EJ. 1993. Disruption of the transthyretin gene results in mice with depressed levels of plasma retinol and thyroid hormone. Proc Natl Acad Sci U S A 90: 2375-9

135
32. van Bennekum AM, Wei S, Gamble MV, Vogel S, Piantedosi R, Gottesman M, Episkopou V, Blaner WS. 2001. Biochemical basis for depressed serum retinol levels in transthyretin-deficient mice. J Biol Chem 276: 1107-13
33. Kawaguchi R, Yu J, Honda J, Hu J, Whitelegge J, Ping P, Wiita P, Bok D, Sun H. 2007. A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 315: 820-5
34. D'Ambrosio DN, Clugston RD, Blaner WS. 2011. Vitamin A Metabolism: An Update. 3: 63-103
35. Blomhoff R, Green MH, Berg T, Norum KR. 1990. Transport and storage of vitamin A. Science 250: 399-404
36. Hodam JR, Creek KE. 1996. Uptake and metabolism of [3H]retinoic acid delivered to human foreskin keratinocytes either bound to serum albumin or added directly to the culture medium. Biochim Biophys Acta 1311: 102-10
37. Noy N. 1992. The ionization behavior of retinoic acid in lipid bilayers and in membranes. Biochim Biophys Acta 1106: 159-64
38. Noy N. 1992. The ionization behavior of retinoic acid in aqueous environments and bound to serum albumin. Biochim Biophys Acta 1106: 151-8
39. Kang JX, Bell J, Leaf A, Beard RL, Chandraratna RA. 1998. Retinoic acid alters the intracellular trafficking of the mannose-6-phosphate/insulin-like growth factor II receptor and lysosomal enzymes. Proc Natl Acad Sci U S A 95: 13687-91
40. Rigas JR, Francis PA, Muindi JR, Kris MG, Huselton C, DeGrazia F, Orazem JP, Young CW, Warrell RP, Jr. 1993. Constitutive variability in the pharmacokinetics of the natural retinoid, all-trans-retinoic acid, and its modulation by ketoconazole. J Natl Cancer Inst 85: 1921-6
41. Goodman DW, Huang HS, Shiratori T. 1965. Tissue Distribution and Metabolism of Newly Absorbed Vitamin a in the Rat. J Lipid Res 6: 390-6

136
42. van Bennekum AM, Kako Y, Weinstock PH, Harrison EH, Deckelbaum RJ, Goldberg IJ, Blaner WS. 1999. Lipoprotein lipase expression level influences tissue clearance of chylomicron retinyl ester. J Lipid Res 40: 565-74
43. Quadro L, Blaner WS, Salchow DJ, Vogel S, Piantedosi R, Gouras P, Freeman S, Cosma MP, Colantuoni V, Gottesman ME. 1999. Impaired retinal function and vitamin A availability in mice lacking retinol-binding protein. Embo J 18: 4633-44
44. Quadro L, Hamberger L, Gottesman ME, Colantuoni V, Ramakrishnan R, Blaner WS. 2004. Transplacental delivery of retinoid: the role of retinol-binding protein and lipoprotein retinyl ester. Am J Physiol Endocrinol Metab 286: E844-51
45. Ross AC, Li NQ. 2007. Lung retinyl ester is low in young adult rats fed a vitamin A deficient diet after weaning, despite neonatal vitamin A supplementation and maintenance of normal plasma retinol. J Nutr 137: 2213-8
46. Blaner WS, Obunike JC, Kurlandsky SB, al-Haideri M, Piantedosi R, Deckelbaum RJ, Goldberg IJ. 1994. Lipoprotein lipase hydrolysis of retinyl ester. Possible implications for retinoid uptake by cells. J Biol Chem 269: 16559-65
47. Ong DE, Chytil F. 1976. Changes in levels of cellular retinol- and retinoic-acid-binding proteins of liver and lung during perinatal development of rat. Proc Natl Acad Sci U S A 73: 3976-8
48. Napoli JL. 1999. Interactions of retinoid binding proteins and enzymes in retinoid metabolism. Biochim Biophys Acta 1440: 139-62
49. Ross AC, Ternus ME. 1993. Vitamin A as a hormone: recent advances in understanding the actions of retinol, retinoic acid, and beta carotene. J Am Diet Assoc 93: 1285-90; quiz 91-2
50. Yost RW, Harrison EH, Ross AC. 1988. Esterification by rat liver microsomes of retinol bound to cellular retinol-binding protein. J Biol Chem 263: 18693-701
51. Napoli JL. 1996. Retinoic acid biosynthesis and metabolism. Faseb J 10: 993-1001

137
52. Duester G. 1996. Involvement of alcohol dehydrogenase, short-chain dehydrogenase/reductase, aldehyde dehydrogenase, and cytochrome P450 in the control of retinoid signaling by activation of retinoic acid synthesis. Biochemistry 35: 12221-7
53. Zile M, DeLuca HF. 1968. Retinoic acid: some aspects of growth-promoting activity in the albino rat. J Nutr 94: 302-8
54. Fiorella PD, Napoli JL. 1991. Expression of cellular retinoic acid binding protein (CRABP) in Escherichia coli. Characterization and evidence that holo-CRABP is a substrate in retinoic acid metabolism. J Biol Chem 266: 16572-9
55. Ross AC, Zolfaghari R, Weisz J. 2001. Vitamin A: recent advances in the biotransformation, transport, and metabolism of retinoids. Curr Opin Gastroenterol 17: 184-92
56. Ross AC. 2003. Retinoid production and catabolism: role of diet in regulating retinol esterification and retinoic Acid oxidation. J Nutr 133: 291S-6S
57. Randolph RK, Ross AC. 1991. Vitamin A status regulates hepatic lecithin: retinol acyltransferase activity in rats. J Biol Chem 266: 16453-7
58. Wang Y, Zolfaghari R, Ross AC. 2002. Cloning of rat cytochrome P450RAI (CYP26) cDNA and regulation of its gene expression by all-trans-retinoic acid in vivo. Arch Biochem Biophys 401: 235-43
59. Yamamoto Y, Zolfaghari R, Ross AC. 2000. Regulation of CYP26 (cytochrome P450RAI) mRNA expression and retinoic acid metabolism by retinoids and dietary vitamin A in liver of mice and rats. Faseb J 14: 2119-27
60. Petkovich PM. 2001. Retinoic acid metabolism. J Am Acad Dermatol 45: S136-42
61. Golzio C, Martinovic-Bouriel J, Thomas S, Mougou-Zrelli S, Grattagliano-Bessieres B, Bonniere M, Delahaye S, Munnich A, Encha-Razavi F, Lyonnet S, Vekemans M, Attie-Bitach T, Etchevers HC. 2007. Matthew-Wood syndrome is caused by truncating mutations in the retinol-binding protein receptor gene STRA6. Am J Hum Genet 80: 1179-87

138
62. Isken A, Golczak M, Oberhauser V, Hunzelmann S, Driever W, Imanishi Y, Palczewski K, von Lintig J. 2008. RBP4 disrupts vitamin A uptake homeostasis in a STRA6-deficient animal model for Matthew-Wood syndrome. Cell Metab 7: 258-68
63. Bouillet P, Sapin V, Chazaud C, Messaddeq N, Decimo D, Dolle P, Chambon P. 1997. Developmental expression pattern of Stra6, a retinoic acid-responsive gene encoding a new type of membrane protein. Mech Dev 63: 173-86
64. Cai K, Gudas LJ. 2009. Retinoic acid receptors and GATA transcription factors activate the transcription of the human lecithin:retinol acyltransferase gene. Int J Biochem Cell Biol 41: 546-53
65. Tice DA, Szeto W, Soloviev I, Rubinfeld B, Fong SE, Dugger DL, Winer J, Williams PM, Wieand D, Smith V, Schwall RH, Pennica D, Polakis P. 2002. Synergistic induction of tumor antigens by Wnt-1 signaling and retinoic acid revealed by gene expression profiling. J Biol Chem 277: 14329-35
66. Kim YK, Wassef L, Hamberger L, Piantedosi R, Palczewski K, Blaner WS, Quadro L. 2008. Retinyl ester formation by lecithin: retinol acyltransferase is a key regulator of retinoid homeostasis in mouse embryogenesis. J Biol Chem 283: 5611-21
67. Wu L, Ross AC. 2010. Acidic retinoids synergize with vitamin A to enhance retinol uptake and STRA6, LRAT, and CYP26B1 expression in neonatal lung. J Lipid Res 51: 378-87
68. Ross AC. 1993. Overview of retinoid metabolism. J Nutr 123: 346-50
69. Chambon P. 1996. A decade of molecular biology of retinoic acid receptors. Faseb J 10: 940-54
70. De Luca LM. 1991. Retinoids and their receptors in differentiation, embryogenesis, and neoplasia. Faseb J 5: 2924-33
71. Petkovich M, Brand NJ, Krust A, Chambon P. 1987. A human retinoic acid receptor which belongs to the family of nuclear receptors. Nature 330: 444-50

139
72. Heyman RA, Mangelsdorf DJ, Dyck JA, Stein RB, Eichele G, Evans RM, Thaller C. 1992. 9-cis retinoic acid is a high affinity ligand for the retinoid X receptor. Cell 68: 397-406
73. Hashimoto Y, Kagechika H, Shudo K. 1990. Expression of retinoic acid receptor genes and the ligand-binding selectivity of retinoic acid receptors (RAR's). Biochem Biophys Res Commun 166: 1300-7
74. Mangelsdorf DJ. 1994. Vitamin A receptors. Nutr Rev 52: S32-44
75. McCollum EV. 1917. The supplementary dietary realtionships among our natural foodstuffs. J. Am. Med. Assoc 68: 1379-86
76. Bloch CE. 1921. Clinical investigation of Xerophthalmia and Dystrophy in Infants and Young Children (Xerophthalmia et Dystrophia Alipogenetica). J Hyg (Lond) 19: 283-304 5
77. Wolbach SB, Howe PR. 1933. Epithelial Repair in Recovery from Vitamin a Deficiency : an Experimental Study. J Exp Med 57: 511-26
78. Emura M, Mohr U, Riebe M, Aufderheide M, Dungworth DL. 1988. Regulation of growth and differentiation by vitamin A in a cloned fetal lung epithelial cell line cultured on collagen gel in hormone-supplemented medium. In Vitro Cell Dev Biol 24: 639-48
79. Ochiai A, Emura M, Mohr U, Tahara E, Dungworth DL. 1991. Induction and characterization of secretory differentiation in human fetal bronchial epithelial cell line (HFBE) cultured on collagen gel in growth hormone and vitamin A-supplemented medium. Exp Pathol 41: 157-68
80. Jetten AM, Brody AR, Deas MA, Hook GE, Rearick JI, Thacher SM. 1987. Retinoic acid and substratum regulate the differentiation of rabbit tracheal epithelial cells into squamous and secretory phenotype. Morphological and biochemical characterization. Lab Invest 56: 654-64
81. Wu R, Wu MM. 1986. Effects of retinoids on human bronchial epithelial cells: differential regulation of hyaluronate synthesis and keratin protein synthesis. J Cell Physiol 127: 73-82

140
82. Haq R, Pfahl M, Chytil F. 1991. Retinoic acid affects the expression of nuclear retinoic acid receptors in tissues of retinol-deficient rats. Proc Natl Acad Sci U S A 88: 8272-6
83. Malpel S, Mendelsohn C, Cardoso WV. 2000. Regulation of retinoic acid signaling during lung morphogenesis. Development 127: 3057-67
84. Mollard R, Ghyselinck NB, Wendling O, Chambon P, Mark M. 2000. Stage-dependent responses of the developing lung to retinoic acid signaling. Int J Dev Biol 44: 457-62
85. Mendelsohn C, Lohnes D, Decimo D, Lufkin T, LeMeur M, Chambon P, Mark M. 1994. Function of the retinoic acid receptors (RARs) during development (II). Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 120: 2749-71
86. Luo J, Sucov HM, Bader JA, Evans RM, Giguere V. 1996. Compound mutants for retinoic acid receptor (RAR) beta and RAR alpha 1 reveal developmental functions for multiple RAR beta isoforms. Mech Dev 55: 33-44
87. Kastner P, Mark M, Ghyselinck N, Krezel W, Dupe V, Grondona JM, Chambon P. 1997. Genetic evidence that the retinoid signal is transduced by heterodimeric RXR/RAR functional units during mouse development. Development 124: 313-26
88. Niederreither K, Subbarayan V, Dolle P, Chambon P. 1999. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat Genet 21: 444-8
89. Cardoso WV, Williams MC, Mitsialis SA, Joyce-Brady M, Rishi AK, Brody JS. 1995. Retinoic acid induces changes in the pattern of airway branching and alters epithelial cell differentiation in the developing lung in vitro. Am J Respir Cell Mol Biol 12: 464-76
90. Chazaud C, Dolle P, Rossant J, Mollard R. 2003. Retinoic acid signaling regulates murine bronchial tubule formation. Mech Dev 120: 691-700
91. Wongtrakool C, Malpel S, Gorenstein J, Sedita J, Ramirez MI, Underhill TM, Cardoso WV. 2003. Down-regulation of retinoic acid receptor alpha signaling is

141
required for sacculation and type I cell formation in the developing lung. J Biol Chem 278: 46911-8
92. Shenai JP, Chytil F. 1990. Vitamin A storage in lungs during perinatal development in the rat. Biol Neonate 57: 126-32
93. Geevarghese SK, Chytil F. 1994. Depletion of retinyl esters in the lungs coincides with lung prenatal morphological maturation. Biochem Biophys Res Commun 200: 529-35
94. McGowan SE, Harvey CS, Jackson SK. 1995. Retinoids, retinoic acid receptors, and cytoplasmic retinoid binding proteins in perinatal rat lung fibroblasts. Am J Physiol 269: L463-72
95. Hind M, Maden M. 2004. Retinoic acid induces alveolar regeneration in the adult mouse lung. Eur Respir J 23: 20-7
96. Whitney D, Massaro GD, Massaro D, Clerch LB. 1999. Gene expression of cellular retinoid-binding proteins: modulation by retinoic acid and dexamethasone in postnatal rat lung. Pediatr Res 45: 2-7
97. Massaro GD, Massaro D. 1996. Postnatal treatment with retinoic acid increases the number of pulmonary alveoli in rats. Am J Physiol 270: L305-10
98. Chytil F. 1992. The lungs and vitamin A. Am J Physiol 262: L517-27
99. Massaro GD, Massaro D. 2000. Retinoic acid treatment partially rescues failed septation in rats and in mice. Am J Physiol Lung Cell Mol Physiol 278: L955-60
100. Massaro GD, Massaro D. 1997. Retinoic acid treatment abrogates elastase-induced pulmonary emphysema in rats. Nat Med 3: 675-7
101. McGowan S, Jackson SK, Jenkins-Moore M, Dai HH, Chambon P, Snyder JM. 2000. Mice bearing deletions of retinoic acid receptors demonstrate reduced lung elastin and alveolar numbers. Am J Respir Cell Mol Biol 23: 162-7

142
102. Massaro GD, Massaro D, Chan WY, Clerch LB, Ghyselinck N, Chambon P, Chandraratna RA. 2000. Retinoic acid receptor-beta: an endogenous inhibitor of the perinatal formation of pulmonary alveoli. Physiol Genomics 4: 51-7
103. McGowan SE, Doro MM, Jackson SK. 1997. Endogenous retinoids increase perinatal elastin gene expression in rat lung fibroblasts and fetal explants. Am J Physiol 273: L410-6
104. Bhatia J, Ziegler EE. 1983. Retinol-binding protein and prealbumin in cord blood of term and preterm infants. Early Hum Dev 8: 129-33
105. Brandt RB, Mueller DG, Schroeder JR, Guyer KE, Kirkpatrick BV, Hutcher NE, Ehrlich FE. 1978. Serum vitamin A in premature and term neonates. J Pediatr 92: 101-4
106. Shenai JP, Chytil F, Jhaveri A, Stahlman MT. 1981. Plasma vitamin A and retinol-binding protein in premature and term neonates. J Pediatr 99: 302-5
107. Mupanemunda RH, Lee DS, Fraher LJ, Koura IR, Chance GW. 1994. Postnatal changes in serum retinol status in very low birth weight infants. Early Hum Dev 38: 45-54
108. Bancalari E, Claure N, Sosenko IR. 2003. Bronchopulmonary dysplasia: changes in pathogenesis, epidemiology and definition. Semin Neonatol 8: 63-71
109. Shenai JP, Kennedy KA, Chytil F, Stahlman MT. 1987. Clinical trial of vitamin A supplementation in infants susceptible to bronchopulmonary dysplasia. J Pediatr 111: 269-77
110. Ng PC. 1993. The effectiveness and side effects of dexamethasone in preterm infants with bronchopulmonary dysplasia. Arch Dis Child 68: 330-6
111. Massaro D, Massaro GD. 1986. Dexamethasone accelerates postnatal alveolar wall thinning and alters wall composition. Am J Physiol 251: R218-24
112. Blanco LN, Massaro GD, Massaro D. 1989. Alveolar dimensions and number: developmental and hormonal regulation. Am J Physiol 257: L240-7

143
113. Loeb JN. 1976. Corticosteroids and growth. N Engl J Med 295: 547-52
114. Massaro D, Teich N, Maxwell S, Massaro GD, Whitney P. 1985. Postnatal development of alveoli. Regulation and evidence for a critical period in rats. J Clin Invest 76: 1297-305
115. Morishige WK, Joun NS. 1982. Influence of glucocorticoids on postnatal lung development in the rat: possible modulation by thyroid hormone. Endocrinology 111: 1587-94
116. Henning SJ. 1978. Plasma concentrations of total and free corticosterone during development in the rat. Am J Physiol 235: E451-6
117. Burri PH, Dbaly J, Weibel ER. 1974. The postnatal growth of the rat lung. I. Morphometry. Anat Rec 178: 711-30
118. Georgieff MK, Mammel MC, Mills MM, Gunter EW, Johnson DE, Thompson TR. 1989. Effect of postnatal steroid administration on serum vitamin A concentrations in newborn infants with respiratory compromise. J Pediatr 114: 301-4
119. Ikeda E, Achen MG, Breier G, Risau W. 1995. Hypoxia-induced transcriptional activation and increased mRNA stability of vascular endothelial growth factor in C6 glioma cells. J Biol Chem 270: 19761-6
120. Levy AP, Levy NS, Wegner S, Goldberg MA. 1995. Transcriptional regulation of the rat vascular endothelial growth factor gene by hypoxia. J Biol Chem 270: 13333-40
121. Forsythe JA, Jiang BH, Iyer NV, Agani F, Leung SW, Koos RD, Semenza GL. 1996. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol Cell Biol 16: 4604-13
122. Ema M, Taya S, Yokotani N, Sogawa K, Matsuda Y, Fujii-Kuriyama Y. 1997. A novel bHLH-PAS factor with close sequence similarity to hypoxia-inducible factor 1alpha regulates the VEGF expression and is potentially involved in lung and vascular development. Proc Natl Acad Sci U S A 94: 4273-8

144
123. Shweiki D, Itin A, Soffer D, Keshet E. 1992. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature 359: 843-5
124. Maniscalco WM, Watkins RH, D'Angio CT, Ryan RM. 1997. Hyperoxic injury decreases alveolar epithelial cell expression of vascular endothelial growth factor (VEGF) in neonatal rabbit lung. Am J Respir Cell Mol Biol 16: 557-67
125. Hosford GE, Olson DM. 2003. Effects of hyperoxia on VEGF, its receptors, and HIF-2alpha in the newborn rat lung. Am J Physiol Lung Cell Mol Physiol 285: L161-8
126. Bartlett D, Jr. 1970. Postnatal growth of the mammalian lung: influence of low and high oxygen tensions. Respir Physiol 9: 58-64
127. Bucher JR, Roberts RJ. 1981. The development of the newborn rat lung in hyperoxia: a dose-response study of lung growth, maturation, and changes in antioxidant enzyme activities. Pediatr Res 15: 999-1008
128. Veness-Meehan KA, Pierce RA, Moats-Staats BM, Stiles AD. 2002. Retinoic acid attenuates O2-induced inhibition of lung septation. Am J Physiol Lung Cell Mol Physiol 283: L971-80
129. Stinchcombe SV, Maden M. 2008. Retinoic acid induced alveolar regeneration: critical differences in strain sensitivity. Am J Respir Cell Mol Biol 38: 185-91
130. Maden M. 2006. Retinoids have differing efficacies on alveolar regeneration in a dexamethasone-treated mouse. Am J Respir Cell Mol Biol 35: 260-7
131. Akiyama H, Tanaka T, Maeno T, Kanai H, Kimura Y, Kishi S, Kurabayashi M. 2002. Induction of VEGF gene expression by retinoic acid through Sp1-binding sites in retinoblastoma Y79 cells. Invest Ophthalmol Vis Sci 43: 1367-74
132. Chinoy MR, Graybill MM, Miller SA, Lang CM, Kauffman GL. 2002. Angiopoietin-1 and VEGF in vascular development and angiogenesis in hypoplastic lungs. Am J Physiol Lung Cell Mol Physiol 283: L60-6
133. McGowan SE, Torday JS. 1997. The pulmonary lipofibroblast (lipid interstitial cell) and its contributions to alveolar development. Annu Rev Physiol 59: 43-62

145
134. Okabe T, Yorifuji H, Yamada E, Takaku F. 1984. Isolation and characterization of vitamin-A-storing lung cells. Exp Cell Res 154: 125-35
135. Dirami G, Massaro GD, Clerch LB, Ryan US, Reczek PR, Massaro D. 2004. Lung retinol storing cells synthesize and secrete retinoic acid, an inducer of alveolus formation. Am J Physiol Lung Cell Mol Physiol 286: L249-56
136. Sannes PL. 1991. Structural and functional relationships between type II pneumocytes and components of extracellular matrices. Exp Lung Res 17: 639-59
137. Vaccaro C, Brody JS. 1978. Ultrastructure of developing alveoli. I. The role of the interstitial fibroblast. Anat Rec 192: 467-79
138. Lindahl P, Karlsson L, Hellstrom M, Gebre-Medhin S, Willetts K, Heath JK, Betsholtz C. 1997. Alveogenesis failure in PDGF-A-deficient mice is coupled to lack of distal spreading of alveolar smooth muscle cell progenitors during lung development. Development 124: 3943-53
139. Ghyselinck NB, Dupe V, Dierich A, Messaddeq N, Garnier JM, Rochette-Egly C, Chambon P, Mark M. 1997. Role of the retinoic acid receptor beta (RARbeta) during mouse development. Int J Dev Biol 41: 425-47
140. Blomhoff R, Skrede B, Norum KR. 1990. Uptake of chylomicron remnant retinyl ester via the low density lipoprotein receptor: implications for the role of vitamin A as a possible preventive for some forms of cancer. J Intern Med 228: 207-10
141. Spit BJ. 1983. Induction of lipid droplets in fibroblasts of the hamster lung by a diet high in vitamin A. Exp Lung Res 4: 247-57
142. Kaplan NB, Grant MM, Brody JS. 1985. The lipid interstitial cell of the pulmonary alveolus. Age and species differences. Am Rev Respir Dis 132: 1307-12
143. Torday JS, Torres E, Rehan VK. 2003. The role of fibroblast transdifferentiation in lung epithelial cell proliferation, differentiation, and repair in vitro. Pediatr Pathol Mol Med 22: 189-207

146
144. Adamson IY, Young L, King GM. 1991. Reciprocal epithelial: fibroblast interactions in the control of fetal and adult rat lung cells in culture. Exp Lung Res 17: 821-35
145. Shannon JM, Pan T, Nielsen LD, Edeen KE, Mason RJ. 2001. Lung fibroblasts improve differentiation of rat type II cells in primary culture. Am J Respir Cell Mol Biol 24: 235-44
146. O'Reilly MA, Stripp BR, Pryhuber GS. 1997. Epithelial-mesenchymal interactions in the alteration of gene expression and morphology following lung injury. Microsc Res Tech 38: 473-9
147. Shannon JM, Hyatt BA. 2004. Epithelial-mesenchymal interactions in the developing lung. Annu Rev Physiol 66: 625-45
148. Torday J, Hua J, Slavin R. 1995. Metabolism and fate of neutral lipids of fetal lung fibroblast origin. Biochim Biophys Acta 1254: 198-206
149. Chytil F. 1996. Retinoids in lung development. Faseb J 10: 986-92
150. Chan V, Greenough A, Cheeseman P, Gamsu HR. 1993. Vitamin A status in preterm and term infants at birth. J Perinat Med 21: 59-62
151. Mactier H, Weaver LT. 2005. Vitamin A and preterm infants: what we know, what we don't know, and what we need to know. Arch Dis Child Fetal Neonatal Ed 90: F103-8
152. Tyson JE, Wright LL, Oh W, Kennedy KA, Mele L, Ehrenkranz RA, Stoll BJ, Lemons JA, Stevenson DK, Bauer CR, Korones SB, Fanaroff AA. 1999. Vitamin A supplementation for extremely-low-birth-weight infants. National Institute of Child Health and Human Development Neonatal Research Network. N Engl J Med 340: 1962-8
153. Ambalavanan N, Tyson JE, Kennedy KA, Hansen NI, Vohr BR, Wright LL, Carlo WA. 2005. Vitamin A supplementation for extremely low birth weight infants: outcome at 18 to 22 months. Pediatrics 115: e249-54

147
154. Shenai JP, Rush MG, Stahlman MT, Chytil F. 1992. Vitamin A supplementation and bronchopulmonary dysplasia--revisited. J Pediatr 121: 399-401
155. Darlow BA, Graham PJ. 2002. Vitamin A supplementation for preventing morbidity and mortality in very low birthweight infants. Cochrane Database Syst Rev: CD000501
156. Baybutt RC, Hu L, Molteni A. 2000. Vitamin A deficiency injures lung and liver parenchyma and impairs function of rat type II pneumocytes. J Nutr 130: 1159-65
157. McGowan SE, Holmes AJ. 2007. Vitamin A deficiency alters pulmonary parenchymal collagen and tissue mechanics. Respir Physiol Neurobiol 156: 312-9
158. Kaza AK, Kron IL, Kern JA, Long SM, Fiser SM, Nguyen RP, Tribble CG, Laubach VE. 2001. Retinoic acid enhances lung growth after pneumonectomy. Ann Thorac Surg 71: 1645-50
159. Shenai JP, Chytil F. 1990. Effect of maternal vitamin-A administration on fetal lung vitamin-A stores in the perinatal rat. Biol Neonate 58: 318-25
160. Ross AC, Li NQ, Wu L. 2006. The components of VARA, a nutrient-metabolite combination of vitamin A and retinoic acid, act efficiently together and separately to increase retinyl esters in the lungs of neonatal rats. J Nutr 136: 2803-7
161. Ross AC, Ambalavanan N, Zolfaghari R, Li NQ. 2006. Vitamin A combined with retinoic acid increases retinol uptake and lung retinyl ester formation in a synergistic manner in neonatal rats. J Lipid Res 47: 1844-51
162. Rahmathullah L, Tielsch JM, Thulasiraj RD, Katz J, Coles C, Devi S, John R, Prakash K, Sadanand AV, Edwin N, Kamaraj C. 2003. Impact of supplementing newborn infants with vitamin A on early infant mortality: community based randomised trial in southern India. Bmj 327: 254
163. Folch J LM, and Sloane Stanley GH. 1957. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem 226: 497-509
164. Takatsuka J, Takahashi N, de Luca LM. 1996. Retinoic acid metabolism and inhibition of cell proliferation: an unexpected liaison. Cancer Res 56: 675-8

148
165. Thurlbeck WM. 1975. Postnatal growth and development of the lung. Am Rev Respir Dis 111: 803-44
166. Bogue CW, Jacobs HC, Dynia DW, Wilson CM, Gross I. 1996. Retinoic acid increases surfactant protein mRNA in fetal rat lung in culture. Am J Physiol 271: L862-8
167. Chailley-Heu B, Chelly N, Lelievre-Pegorier M, Barlier-Mur AM, Merlet-Benichou C, Bourbon JR. 1999. Mild vitamin A deficiency delays fetal lung maturation in the rat. Am J Respir Cell Mol Biol 21: 89-96
168. Liu B, Harvey CS, McGowan SE. 1993. Retinoic acid increases elastin in neonatal rat lung fibroblast cultures. Am J Physiol 265: L430-7
169. McGowan SE, Jackson SK, Olson PJ, Parekh T, Gold LI. 1997. Exogenous and endogenous transforming growth factors-beta influence elastin gene expression in cultured lung fibroblasts. Am J Respir Cell Mol Biol 17: 25-35
170. Veness-Meehan KA, Bottone FG, Jr., Stiles AD. 2000. Effects of retinoic acid on airspace development and lung collagen in hyperoxia-exposed newborn rats. Pediatr Res 48: 434-44
171. Esteban-Pretel G, Marin MP, Renau-Piqueras J, Barber T, Timoneda J. 2010. Vitamin A deficiency alters rat lung alveolar basement membrane: reversibility by retinoic acid. J Nutr Biochem 21: 227-36
172. Fraslon C, Bourbon JR. 1994. Retinoids control surfactant phospholipid biosynthesis in fetal rat lung. Am J Physiol 266: L705-12
173. Maden M, Hind M. 2004. Retinoic acid in alveolar development, maintenance and regeneration. Philos Trans R Soc Lond B Biol Sci 359: 799-808
174. Belloni PN, Garvin L, Mao CP, Bailey-Healy I, Leaffer D. 2000. Effects of all-trans-retinoic acid in promoting alveolar repair. Chest 117: 235S-41S
175. Ross AC. 1999. Vitamin A and retinoids. In: Modern Nutrtion in Health and Disease (Shils, M.E., Olson, J.A., Shike, M. & Ross, A.C., eds.) 9th ed.: pp. 305-32

149
176. Altucci L, Gronemeyer H. 2001. Nuclear receptors in cell life and death. Trends Endocrinol Metab 12: 460-8
177. Bastien J, Rochette-Egly C. 2004. Nuclear retinoid receptors and the transcription of retinoid-target genes. Gene 328: 1-16
178. Speer CP. 2006. Inflammation and bronchopulmonary dysplasia: a continuing story. Semin Fetal Neonatal Med 11: 354-62
179. Speer CP. 2003. Inflammation and bronchopulmonary dysplasia. Semin Neonatol 8: 29-38
180. Groneck P, Gotze-Speer B, Oppermann M, Eiffert H, Speer CP. 1994. Association of pulmonary inflammation and increased microvascular permeability during the development of bronchopulmonary dysplasia: a sequential analysis of inflammatory mediators in respiratory fluids of high-risk preterm neonates. Pediatrics 93: 712-8
181. Groneck P, Schmale J, Soditt V, Stutzer H, Gotze-Speer B, Speer CP. 2001. Bronchoalveolar inflammation following airway infection in preterm infants with chronic lung disease. Pediatr Pulmonol 31: 331-8
182. Jobe AH, Ikegami M. 1998. Mechanisms initiating lung injury in the preterm. Early Hum Dev 53: 81-94
183. Morgan ET, Li-Masters T, Cheng PY. 2002. Mechanisms of cytochrome P450 regulation by inflammatory mediators. Toxicology 181-182: 207-10
184. Hoo AF, Dezateux C, Henschen M, Costeloe K, Stocks J. 2002. Development of airway function in infancy after preterm delivery. J Pediatr 141: 652-8
185. Zolfaghari R, Cifelli CJ, Lieu SO, Chen Q, Li NQ, Ross AC. 2007. Lipopolysaccharide opposes the induction of CYP26A1 and CYP26B1 gene expression by retinoic acid in the rat liver in vivo. Am J Physiol Gastrointest Liver Physiol 292: G1029-36
186. Janssens S, Beyaert R. 2003. Role of Toll-like receptors in pathogen recognition. Clin Microbiol Rev 16: 637-46

150
187. Guha M, Mackman N. 2001. LPS induction of gene expression in human monocytes. Cell Signal 13: 85-94
188. Rockel JS, Kudirka JC, Guzi AJ, Bernier SM. 2008. Regulation of Sox9 activity by crosstalk with nuclear factor-kappaB and retinoic acid receptors. Arthritis Res Ther 10: R3
189. Shoseyov D, Bibi H, Biesalski H, Reifen R. 2002. Repeated allergen challenge in rats increases vitamin A consumption. Chest 122: 1407-11
190. Paiva SA, Godoy I, Vannucchi H, Favaro RM, Geraldo RR, Campana AO. 1996. Assessment of vitamin A status in chronic obstructive pulmonary disease patients and healthy smokers. Am J Clin Nutr 64: 928-34
191. Massaro D, Massaro GD. 2002. Invited Review: pulmonary alveoli: formation, the "call for oxygen," and other regulators. Am J Physiol Lung Cell Mol Physiol 282: L345-58
192. Massaro D, Massaro GD. 2006. Toward therapeutic pulmonary alveolar regeneration in humans. Proc Am Thorac Soc 3: 709-12
193. Reeves PG, Nielsen FH, Fahey GC, Jr. 1993. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J Nutr 123: 1939-51
194. Fries KM, Blieden T, Looney RJ, Sempowski GD, Silvera MR, Willis RA, Phipps RP. 1994. Evidence of fibroblast heterogeneity and the role of fibroblast subpopulations in fibrosis. Clin Immunol Immunopathol 72: 283-92
195. Penney DP, Keng PC, Derdak S, Phipps RP. 1992. Morphologic and functional characteristics of subpopulations of murine lung fibroblasts grown in vitro. Anat Rec 232: 432-43
196. Tordet C, Marin L, Dameron F. 1981. Pulmonary di-and-triacylglycerols during the perinatal development of the rat. Experientia 37: 333-4
197. Grummer MA, Thet LA, Zachman RD. 1994. Expression of retinoic acid receptor genes in fetal and newborn rat lung. Pediatr Pulmonol 17: 234-8

151
198. Grummer MA, Zachman RD. 1995. Postnatal rat lung retinoic acid receptor (RAR) mRNA expression and effects of dexamethasone on RAR beta mRNA. Pediatr Pulmonol 20: 234-40
199. Tanumihardjo SA, Muherdiyantiningsih, Permaesih D, Komala, Muhilal, Karyadi D, Olson JA. 1996. Daily supplements of vitamin A (8.4 mumol, 8000 IU) improve the vitamin A status of lactating Indonesian women. Am J Clin Nutr 63: 32-5
200. Valentine AR, Tanumihardjo SA. 2005. One-time vitamin A supplementation of lactating sows enhances hepatic retinol in their offspring independent of dose size. Am J Clin Nutr 81: 427-33
201. Davila ME, Norris L, Cleary MP, Ross AC. 1985. Vitamin A during lactation: relationship of maternal diet to milk vitamin A content and to the vitamin A status of lactating rats and their pups. J Nutr 115: 1033-41
202. Barua AB, McGowan SE, Ivanoff KD, Goswami BC, Olson JA. 2004. Elevation of retinyl ester level in the lungs of rats following repeated intraperitoneal injections of retinoic acid or retinoyl glucuronide. Pulm Pharmacol Ther 17: 113-9
203. Cifelli CJ, Green JB, Green MH. 2005. Dietary retinoic acid alters vitamin A kinetics in both the whole body and in specific organs of rats with low vitamin A status. J Nutr 135: 746-52
204. Schuger L, Varani J, Mitra R, Jr., Gilbride K. 1993. Retinoic acid stimulates mouse lung development by a mechanism involving epithelial-mesenchymal interaction and regulation of epidermal growth factor receptors. Dev Biol 159: 462-73
205. McGowan SE. 1992. Extracellular matrix and the regulation of lung development and repair. Faseb J 6: 2895-904
206. Vogel SG, M.V.; Blaner, W.S. 1999. Retinoid uptake, metabolism and transport. In Handbook of Experimental Pharmacology, The Retinoids;, pp. 31-96. Heidelberg, Germany: Springer Verlag
207. Reifen R. 2002. Vitamin A as an anti-inflammatory agent. Proc Nutr Soc 61: 397-400

152
208. Semba RD. 1999. Vitamin A and immunity to viral, bacterial and protozoan infections. Proc Nutr Soc 58: 719-27
209. Hussey GD, Klein M. 1990. A randomized, controlled trial of vitamin A in children with severe measles. N Engl J Med 323: 160-4
210. Villamor E, Fawzi WW. 2000. Vitamin A supplementation: implications for morbidity and mortality in children. J Infect Dis 182 Suppl 1: S122-33
211. Grubesic RB, Selwyn BJ. 2003. Vitamin A supplementation and health outcomes for children in Nepal. J Nurs Scholarsh 35: 15-20
212. Semba RD, Ndugwa C, Perry RT, Clark TD, Jackson JB, Melikian G, Tielsch J, Mmiro F. 2005. Effect of periodic vitamin A supplementation on mortality and morbidity of human immunodeficiency virus-infected children in Uganda: A controlled clinical trial. Nutrition 21: 25-31
213. Ross AC. 2000. Vitamin A, retinoids and immune responses. In Vitamin A and retinoids: an update of biological aspects and clinical application, ed. MA Livrea, pp. 83-96. Switzerland.: Birkhäuser Verlag Basel
214. Kuenzli S, Tran C, Saurat JH. 2004. Retinoid receptors in inflammatory responses: a potential target for pharmacology. Curr Drug Targets Inflamm Allergy 3: 355-60
215. Rosales FJ, Ritter SJ, Zolfaghari R, Smith JE, Ross AC. 1996. Effects of acute inflammation on plasma retinol, retinol-binding protein, and its mRNA in the liver and kidneys of vitamin A-sufficient rats. J Lipid Res 37: 962-71
216. Hind M, Corcoran J, Maden M. 2002. Alveolar proliferation, retinoid synthesizing enzymes, and endogenous retinoids in the postnatal mouse lung. Different roles for Aldh-1 and Raldh-2. Am J Respir Cell Mol Biol 26: 67-73
217. Hind M, Corcoran J, Maden M. 2002. Temporal/spatial expression of retinoid binding proteins and RAR isoforms in the postnatal lung. Am J Physiol Lung Cell Mol Physiol 282: L468-76

Vita
Lili Wu
EDUCATION 2008- 2011 Ph.D., Major: Nutritional Sciences The Pennsylvania State University, University Park, PA 2005-2008 M.S., Major: Nutritional Sciences The Pennsylvania State University, University Park, PA 1996-2001 B.S., Major: Clinical Medicine Anhui Medical University, Hefei, China PROFESSIONAL EXPERENCE Graduate Research Assistant Jan 2005 – Dec 2010 Department of Nutritional Sciences, The Pennsylvania State University, University Park Investigated Vitamin A metabolism in neonatal rat lungs PUBLICATIONS Wu L and Ross AC, Multiple treatments of acidic retinoids during postnatal period accumulate retinol storage in the lung of neonatal rats. (In preparation) Wu L and Ross AC, LPS-induced inflammation modestly affects retinoic acid promoted retinol storage in neonatal lungs. (In preparation) Wu L and Ross AC (2010) Acidic retinoids synergize with vitamin A to enhance retinol uptake and STRA6, LRAT, and CYP26B1 expression in neonatal lung. J Lipid Res 51: 378-387. Wu L, (2008) Master thesis: Investigation on the mechanism of vitamin A uptake, accumulation and metabolism in the lungs of the neonatal rat. Ross AC, Li NQ, and Wu L (2006) The components of VARA, a nutrient-metabolite combination of vitamin A and retinoic acid, act efficiently together and separately to increase retinyl esters in the lungs of neonatal rats. J Nutr 136: 2803-2807. TEACHING EXPERENCE Teaching Assistant 2009 NUTR 445 (Nutrient Metabolism I) PROFESSIONAL MEMBERSHIP American Society for Nutritional Sciences (ASNS)







