early events in photodynamic therapy: chemical and physical changes in a popc:cholesterol bilayer...
TRANSCRIPT

Early Events in Photodynamic Therapy: Chemical and Physical Changesin a POPC:Cholesterol Bilayer due to Hematoporphyrin IX-mediatedPhotosensitization
Antonio Santos1, Antonio M. Rodrigues1, Abılio J. F. N. Sobral1, Paula V. Monsanto2,Winchil L. C. Vaz1 and Maria Joao Moreno*1
1Departamento de Quımica da FCTUC, Coimbra, Portugal2Forensic Toxicology Service, National Institute of Legal Medicine—Center Branch, Coimbra, Portugal
Received 22 March 2009, accepted 8 June 2009, DOI: 10.1111 ⁄ j.1751-1097.2009.00606.x
ABSTRACT
We studied the interaction of hematoporphyrin IX (HpIX) with
bilayers of 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine
(POPC) containing cholesterol at a molar fraction between 0
and 0.5. The membrane-associated fraction of HpIX decreases
significantly over a period of hours, for porphyrin concentrations
in the aqueous phase above 50 nMM. This was attributed to self-
aggregation of HpIX and was well described by a dimerization
process. A model was developed to correct for aggregation and
obtain the true partition coefficient which is dependent on the
molar fraction of cholesterol with a maximum at 20 mol%. The
chemical and physical effects on the lipid bilayer upon irradiation
of HpIX were studied for lipid bilayers with POPC:Chol 1:1.
Exposure of these bilayers to visible light in the presence of
HpIX leads to several cholesterol oxidation products that were
identified using GC-MS. A dramatic increase in the membrane
leakiness was also observed, even for short irradiation times and
small light intensities, as evaluated from the rate of pH
equilibration and dithionite permeability. The relevance of these
results for the mechanism of photodynamic therapy is discussed.
INTRODUCTION
Photodynamic therapy (PDT) uses the combined action oflight, oxygen and sensitizers in selective destruction of tissue.
In the last three decades this therapeutic approach has beenexploited for the treatment of neoplastic diseases (1–3) andmore recently in disinfection (4) and the treatment of
macular degeneration (5). PDT has also been used in thetreatment of skin diseases (4) dating back to its early use inthe treatment of psoriasis (1,3). Recent advances in PDT
focus on the development of more efficient photosensitizers,with improved tissue selectivity (6) and absorption in the red(7) and also on the combined approach of different therapies(6,8) and multifunctionality with the use of nanoplatforms
(9,10).Several aspects of the mechanism of PDT are well
established with the initial production of reactive oxygen
species and the resulting oxidation of cell components as the
first event (11,12). Oxidative stress in PDT may be morelocalized in the lipid bilayer of cell membranes or in theaqueous compartments of the cell depending on the polarity
and charge of the photosensitizer (2,11,12). However, due tothe higher concentration of oxygen in the nonpolar envi-ronment of lipid bilayers (13), the oxidation of bilayer
components is always an outcome of PDT, polyunsaturatedfatty acids and cholesterol being the most susceptible lipids(14,15). The dependence of the outcome of PDT on the totalfluence and fluence rate used is now well recognized with
low doses resulting mainly in apoptosis while high doseslead to necrosis (16–19). In this study we characterize thechemical and physical effects of irradiation at different
fluence rates and a possible relation with the different modesof cell death is proposed.
Another important aspect of the mechanism of PDT is the
toxicity of the oxidized products formed and the biologicalresponse that they generate. It is now accepted that theoxidized forms of cholesterol generated during PDT are very
cytotoxic products, affect several cell pathways and induceapoptosis (20–24). Some insights into the relation betweenthose cellular outcomes and the change in the properties of thelipid bilayers will be given.
In this work we used hematoporphyrin IX (HpIX) togenerate singlet oxygen. This was one of the first photo-sensitizers studied and used in PDT clinical applications (3).
Currently, in therapeutic studies, it has been substituted bymore recent sensitizers with improved light absorption prop-erties as well as tissue selectivity (1,3). It is however a classical
photosensitizer, used in in vitro model systems in which lightpenetration is not an issue (25–27).
MATERIALS AND METHODS
1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC), 1,2-di-myristoyl-sn-glycero-3-phospho-ethanolamine-NBD (NBD-DMPE)and cholesterol were from Avanti Polar Lipids, Inc. (Alabaster,AL), anthracene-9,10-dipropionic acid (ADPA) was from MolecularProbes (Eugene, OR) and diphenyl anthracene (DPA) from Koch-Light Laboratories. All other reagents and solvents were ofanalytical grade, or higher purity, from Sigma-Aldrich QuımicaS.A. (Sintra, Portugal). Hematoporphyrin was purified by chroma-tography on cellulose (cellulose for column chromatography, acidwashed, powder [Fluka 22184]) using tetrahydrofuran as eluent.
*Corresponding author email: [email protected] (Maria Joao Moreno)� 2009TheAuthors. JournalCompilation.TheAmericanSociety ofPhotobiology 0031-8655/09
Photochemistry and Photobiology, 2009, 85: 1409–1417
1409

Cholesterol concentrations were determined using the Lieberman-Burchard method and phospholipid concentrations were determinedusing a modified version of the Bartlett phosphate assay as describedpreviously (28). The fluorescence measurements were performed on aCary Eclipse Fluorescence spectrophotometer (Varian) equipped witha thermostated multicell holder accessory and UV–Vis absorption wasperformed on a Unicam UV530 spectrophotometer (Cambridge, UK).The oxidized cholesterol derivatives were analyzed using GC-MS6890 ⁄HP 5973 from Hewlett-Packard (and ⁄or GC-2010 ⁄GCMS-QP2010 from Shimadzu) with the column HP-5MS from Agilent (orDB-5MS from J&W Scientific for the Shimadzu equipment), atemperature of 250�C at the injector and ionization source and atemperature gradient at the column increasing from 120 to 240�C at5 min and to 270�C at 30 min. Peroxides were analyzed using theperoxide sticks Quantofix (Aldrich Cat Z24,925-4).
For the measurement of the partition coefficient of HpIX betweenthe aqueous phase and the lipid bilayers, HpIX was first dissolved inglacial acetic acid at a concentration of 4.4 · 10)5
MM and then dilutedwith sodium phosphate buffer 50 mMM (pH = 7.4 with 0.15 MM NaCl) tothe required final concentration. Large unilamellar vesicles (LUV)were prepared by evaporation of an azeotropic mixture of chloroformand methanol (87:13, vol:vol) containing the required lipids and probesat the specified molar ratio followed by hydration of the film withphosphate buffer and extrusion (Extrusor from Lipex Biomembranes,Vancouver, BC, Canada) through 100 nm pore size filters (Nucleo-pore; Whatman, Springfield Hill, UK) as described previously (28).
Irradiation of the samples was performed with a 150 W Xenonlamp filtered for the UV and IR radiation with a cutoff filter at 380 nmand a 10 cm water filter, respectively. Depending on the irradiationwavelengths and fluence desired, additional cutoff and ⁄or band-passfilters were used as indicated in the respective experiment. The lightwas focused on a liquid light guide (Newport Stratford, Inc.) and thesample was illuminated from above. The intensity of the incident beamwas determined using potassium ferrioxalate actinometry. The oxygenconcentration in the samples was kept constant by bubbling airpreviously saturated with water. The measurement of the singletoxygen concentration was performed by irradiation of a LUVsuspension 0.5 mMM POPC:Chol 1:1 containing 5 lMM HpIX with anirradiation intensity of 5.3 · 10)5 Einstein dm)3 s)1 at 506 nm. Forthe experiments on the measurement of the singlet oxygen concentra-tion in the aqueous phase, ADPA was added to the LUV aqueoussuspension prior to irradiation at a final concentration of 10 lMM whileDPA was included in the preparation of the LUV, at a molar ratio ofprobe to total lipid of 1:100, for the measurement of singlet oxygen inthe lipid bilayer.
For the experiments on the rate of pH equilibration across the lipidbilayer, a lipid film of POPC:Chol 1:1, with or without HpIX (at aHpIX:lipid molar ratio of 1:500), was hydrated with 1 mMM phosphatebuffer, pH = 5.6 containing 0.15 MM NaCl and 50 lMM carboxyfluores-cein (CBF). The LUVs were prepared by extrusion, as mentionedabove, and were passed through a Sephadex G-75 (Pharmacia) sizeexclusion column to remove the CBF in the aqueous phase outside theLUVs. The sample was irradiated at 20�C for the desired period andlight intensity, was then equilibrated at 37�C and the pH outside theLUVs was changed to 6.6 by dilution with 1 mMM phosphate buffer atpH = 8 (also containing 0.15 MM NaCl). The equilibration of the pH inthe aqueous solution inside the LUVs was followed via the change inthe CBF fluorescence.
Permeation of dithionite through lipid bilayers was measured forLUVs with the lipid composition POPC:Chol 1:1 containing thefluorescent probe, NBD-DMPE, at a molar ratio relative to the totallipid of 1:5000, without HpIX or with 0.2 mol% of this sensitizer. Thesamples were irradiated at 20�C as described above, then equilibratedat the required temperature, and 20 lL of a solution of dithionite 1 MM
dissolved in Tris buffer 1 MM pH = 10 was added to 2 mL of thesample. The decrease in NBD-DMPE due to reduction by dithionitewas followed via NBD fluorescence as described previously (28)allowing the recovery of the rate of dithionite permeability. In brief,the fraction of NBD-DMPE in the outer monolayer of the LUVsreacts within seconds with the dithionite added to the aqueous solutionoutside the LUVs leading to a fast reduction of about 50% in theNBD-DMPE fluorescence. Dithionite slowly permeates the lipidbilayer toward internal aqueous compartment of the LUVs where itreacts with NBD-DMPE in the inner monolayer leading to the
decrease of the fluorescence intensity toward zero and allowing thecalculation of dithionite permeability coefficient (28). The NBD-DMPE initially in the inner monolayer may also translocate intothe outer monolayer (where it reacts promptly with dithionite) andthe faster process (NBD-DMPE translocation or dithionite perme-ability) may be quantitatively characterized from the fluorescencedecrease profile. The two processes have been characterized indepen-dently for lipid bilayers prepared from pure POPC and POPC:Cholmixtures, the permeation of dithionite being faster then NBD-DMPEtranslocation (28).
RESULTS AND DISCUSSION
Partition to the lipid bilayer
One difficulty in performing quantitative studies with HpIX isits limited solubility in aqueous solutions. In most studies this
difficulty has been overcome via the use of organic solvents toprepare the stock solution. To avoid interference of the residualorganic solvent in the results obtained (29), we used glacial
acetic acid as the primary solvent. This solution was found to bestable for several months, without evidence for aggregation ordegradation, at a concentration of HpIX up to 1 · 10)4
MM.Different aqueous solutions of HpIX were prepared from this
stock solution through dilutions with a strong pH buffer(sodium phosphate buffer 5 · 10)2
MM, at pH = 7.4, containing0.15 MM NaCl) and used within 30 min. The pH of the final
solution was always within the range 7.3–7.4.The equilibrium partition of HpIX between the aqueous
phase and lipid bilayers was measured at 25 and 37�C for LUV
prepared from POPC and different molar fractions of choles-terol up to 0.5. The partition was followed through the redshiftand quantum yield increase in HpIX fluorescence in the lipid
bilayer compared to its fluorescence in the aqueous phase(Fig. 1A).
The interaction between a small molecule and a lipid bilayermay be treated as a partition between the aqueous phase and
the lipid phase, Scheme 1, where HpIXW and HpIXL representHpIX in the aqueous phase or associated with the lipid bilayer,respectively.
A B
Figure 1. Titration of an aqueous solution of hematoporphyrin IX(HpIX), at a concentration of 2 · 10)7
MM, with large unilamellarvesicles prepared from 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocho-line (POPC) at 37�C. (A) Fluorescence emission spectrum of HpIX inthe presence of different lipid concentrations. (B) Change in thefluorescence intensity at 630 nm (h) as a function of the lipidconcentration, the best fit of Eq. (3) with KP = 1.5 · 105 is alsoshown (—).
1410 Antonio Santos et al.

HpIXW! KP
HpIXL
Scheme 1. Equilibrium partition of HPIX from the aqueous phase andthe lipid bilayer.
The equilibrium constant in Scheme 1, KP, is the partitioncoefficient and is defined by Eq. (1) where the concentrationsof HpIX are defined with respect to the volume of therespective phase.
KP ¼½HpIXL�L½HpIXW�W
ð1Þ
From Eq. (1) the concentration of HpIX in each phase maybe calculated with respect to the total volume of the solution,
ð½HpIXX�TÞ, which is given by Eq. (2) when the volume of theaqueous phase is considered equal to the total volume of thesolution.
HpIXW½ �T¼HpIXT½ �T
1þ KPVL½L�T
HpIXL½ �T ¼HpIXT½ �TKPVL½L�T1þ KPVL½L�T
ð2Þ
To obtain Eq. (2) from Eq. (1), the total volume of the lipidphase was expressed as the molar volume of the lipid, VL,
multiplied by the concentration of lipid with respect to thetotal volume of the solution, [L]T.
The partition coefficient is calculated from the dependence
of the total fluorescence intensity of HpIX at 630 nm, I630F , onthe lipid concentration according to Eq. (3):
I 630F ¼ U630W ½HpIXW�T þ U630
L ½HpIXL�T ð3Þ
The contribution of the fluorescence from each HpIXpopulation to the total fluorescence observed is given by their
relative quantum yields at 630 nm, U630X . The molar volume of
the lipid, VL, was calculated from data in the literature (30–32), and the values of 0.8 and 0.64 dm3 mol)1 were considered
for bilayers prepared from pure POPC or POPC:Chol 1:1,respectively. The molar volume of the bilayers with interme-diate compositions was calculated assuming a linear depen-dence on the fraction of cholesterol. The total concentration of
lipid, L½ �T, takes into account both POPC and cholesterol. Theresults obtained in a typical titration with POPC at 37�C areshown in Fig. 1B, the average of several experiments and the
data for other lipid compositions are presented in Table 1.The rate constants for the interaction of HpIX with POPC
lipid bilayers have been reported (33), and the calculated time
required to attain equilibrium, after addition of HpIX to theLUVs, is less than 1 min for all the lipid concentrations used.The time required to prepare and analyze all the solutions in agiven titration curve was about 30 min and the partition
coefficient recovered after this incubation time is reported inFig. 1. The fluorescence of the different solutions was thenfollowed over time. A strong decrease in the intensity of the
fluorescence emission at 630 nm accompanied by a smallincrease at 580 nm was observed when the concentration oflipid was small (Fig. 2A). This decrease in the total fluorescence
was reversible (the fluorescence increased to the expected valuewhen high concentrations of POPC were added to the solutions
after a long incubation) and its temporal behavior was notcompatible with a first order process. Additionally, the fluores-cence decrease was dependent upon the concentration of HpIXused, being less pronounced for lower concentrations. This
suggested aggregation ofHpIXas the process responsible for thefluorescence decrease. Aggregation of porphyrins is well docu-mented in the literature (34) and for the small concentrations
used a dimerization is usually assumed. The experimental datawere analyzed with a model that included aggregation of HpIXin the aqueous phase to form aggregates with nHpIXmolecules
(HpIXn), Scheme 2, and partition of the monomers from theaqueous phase into the POPC bilayer, Scheme 1. The formationof aggregates decreases the total concentration of HpIX
available for interacting with the lipid bilayer, which is nowgiven by ½HpIXT� � n½HpIXn�, and in this case the titration ofHpIX with lipid bilayers is described by Eq. (4):
I630F ¼ U630W
ð½HpIXT�T � n½HpIXn�TÞ1þ KPVL½L�T
� �
þ U630L
ð½HpIXT�T � n½HpIXn�TÞKPVL½L�T1þ KPVL½L�T
� � ð4Þ
Table 1. Equilibrium partition coefficients for partitioning of HpIXbetween LUV made from binary mixtures of POPC and cholesteroland the aqueous phase at 25�C.
Molar fraction ofcholesterol KP · 10)5*
0 2.0 ± 0.30.1 2.5 ± 0.30.2 3.0 ± 0.30.3 2.0 ± 0.30.4 1.6 ± 0.30.5 1.2 ± 0.2
HpIX=hematoporphyrin IX; LUV=large unilamellar vesicle; POPC=1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine. *Values shown arethe average and standard deviation of two to five independent experi-ments.
A B
Figure 2. Time dependence of hematoporphyrin IX fluorescence at630 nm in the presence of different concentrations of POPC at 37�C.(A) Decrease in the fluorescence intensity with time after preparationof the solutions for a 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocho-line (POPC) concentration of 0 (h), 5 (s), 15 (,), 30 (v) and 100 (+)lMM. (B) Shape of the titration curve for an incubation time of <30 min(h), 6 h (4), 24 h (e) and 48 h (x). The lines are the best fit of thetitration Eqs. (4) and (5) using the same value of KP = 1.4 · 105,k1 = 3 · 102 MM
)1 s)1, k-1 = 3 · 10)6 s)1 and n = 2 for all curves.
Photochemistry and Photobiology, 2009, 85 1411

The concentration of aggregates at a given time may beobtained from the numerical integration of the differentialequation derived from Scheme 2, Eq. (5):
d½HpIXn�dt
¼ k1½HpIXW�n � k�1½HpIXn� ð5Þ
The best fit of the model, described by Eqs. (4) and (5) withn = 2, is shown in Fig. 2 for POPC at 37�C. Depending on thesystem and replicate, the minimum of the v2 for the best fit ofthe model to the fluorescence decrease was obtained for n
between 1.5 and 3 suggesting that aggregation is not a puredimerization as noted by other authors (35). However, the v2
surface was very shallow and the increase when fixing n = 2
was always smaller than 15%. The effect on the recoveredpartition coefficient was even less significant, always less than10%, being smaller than the standard deviation obtained for
different replicates. Therefore, to increase consistency andbecause dimerization is the dominant process in the concen-tration range used (35), we applied n = 2 for all systems in therecovery of the partition coefficient.
nHpIXW! k1k�1
HpIXn
Scheme 2. Aggregation of HPIX in the aqueous phase.
From the data obtained at different incubation times, thepartition coefficient may be recovered and this is shown in
Fig. 2B. If the experimental results are adjusted with Eq. (3),ignoring aggregation of HpIX, the apparent partition coeffi-cient obtained decreases with increasing time of incubation,
being an order of magnitude smaller after 24 h of incubation.Using Eqs. (4) and (5), the true partition coefficient is obtainedwhich is not dependent on the incubation time and is given inTable 1.
HpIX was titrated with liposomes containing POPC anddifferent molar fractions of cholesterol, at 25�C, and theoverall results obtained are shown in Table 1. The partition
was found to depend strongly on the cholesterol content ofthe membrane with a maximum at 0.2 molar fraction. Thepublished phase diagram for POPC:Chol mixtures shows a
region of coexistence of liquid disordered and liquid orderedphases between 0.1 and 0.4 molar fraction of cholesterol at25�C (36), and our data are consistent with a preferential
partition of HpIX to the phase boundaries. Evidence for aparticularity at 0.1–0.2 molar fraction of cholesterol wasalso found using other methods (37). We recently proposeda model for the microscopic structure of POPC bilayers
enriched in cholesterol which questions the existence of twocoexisting macroscopic phases and proposes a two-dimen-sional micelle-like microheterogeneous structure (38).
According to that model, the increased partition of HpIXtoward POPC bilayers with intermediate molar fractions ofcholesterol may reflect a stronger affinity for the interfaces
between those micelle-like structures.The value obtained for the partition coefficient in POPC
bilayers compares very well with that obtained for lipidbilayers prepared from DOPC (33). The effect of cholesterol
is very similar to that observed in bilayers prepared fromDMPC and different molar fractions of cholesterol (37)where an increase in the affinity was also observed in the
Figure 3. Chromatogram obtained for the POPC:Chol 1:1 lipid bilayer photosensitized by hematoporphyrin IX at a molar ratio of 1:100 lipidmolecules by irradiation with UV–visible light (k > 345 nm), I0 = 5 · 10)3 Einstein dm)3 s)1, for 2 h. The oxysterols formed were identified bytheir MS spectrum and some relevant species are shown.
1412 Antonio Santos et al.

region of 10–20% cholesterol. The titration of HpIX withpure POPC bilayers, and with POPC bilayers containing a0.2 molar fraction of cholesterol, was also carried out at37�C. The values obtained for the partition coefficients were
very similar to those obtained at 25�C being slightly smallerbut within the experimental error. This small variation wasexpected because the partition of amphiphilic molecules
into neutral lipid bilayers is essentially driven by thehydrophobic effect which is mainly of an entropic nature(38–40).
Chemical changes in a POPC:Chol 1:1 bilayer due to
HpIX-mediated photosensitization
When the singlet excited state of hematoporphyrin is popu-
lated by absorption of radiation in the UV–VIS region, there isan efficient intersystem crossing with the formation of thetriplet excited state of hematoporphyrin. In the presence ofmolecular oxygen energy transfer occurs between those two
molecules in the triplet state with the formation of hemato-porphyrin in the ground state (singlet) and molecular oxygenin the singlet excited state (1O�2). In a nonpolar environment
like the lipid bilayer, the production of singlet oxygen is anefficient process (26,27,41) and this is the main route for thedamaging effects observed on cells due to photosensitization
with porphyrin derivatives (11). The singlet oxygen molecule isa very potent oxidant and reacts with unsaturated moleculesthat are in the vicinity of its production site. Due to its lowsolubility in aqueous solutions, HpIX localizes mainly in the
lipid bilayers and oxidized lipids are most probably among theinitial products of photosensitization. From the different lipidscommonly found in biological membranes, polyunsaturated
lipids and cholesterol are the major targets of oxidation bysinglet oxygen (14,15). Following cholesterol oxidation prod-ucts is particularly convenient because it may give insights into
the mechanism of photosensitization-induced cell damage(14,42,43).
We analyzed the oxidation products formed after irradia-
tion of LUVs prepared from a mixture of POPC:Chol (1:1) at amolar ratio of 1 molecule of HpIX to 100 lipid molecules. Thesolution was irradiated, k > 345 nm and I0 = 1.3 · 10)3 -Einstein dm)3 s)1, for 0.5, 1 or 2 h and its chloroform extract
was analyzed using GC-MS. The results obtained for 2 hirradiation are presented in Fig. 3. The chloroform extractswere also analyzed for peroxides, immediately after prepara-
tion, and a concentration of up to 5 mg L)1 in the irradiatedsamples was observed. These peroxides are thermally unstable,decomposing to more stable products, and they are not
observed in the GC-MS results.Several forms of oxidized cholesterol were detected
including cholesta-3,5-diene-7-one, cholest-4-en-3one, choles-tene-3,6-dione and 7-ketocholesterol. Up to 25% of the
cholesterol was oxidized as judged from the relative areas ofall cholesterol derivatives observed. To evaluate the rele-vance of the total fluence and fluence rate used in these
experiments compared to in vivo PDT the light intensitymust be converted into the more common units of W cm)2.For this one needs to consider the wavelength of light and
the size and geometry of the irradiated sample (which in thiswork was top illumination of a cell with 1 cm2 area and3 cm height), Eq. (6);
I0jWcm�2j ¼Zkmax
kmin
I0ðkÞ dk
¼Zkmax
kmin
NAjmol�1j hjJsj cjms�1jI0ðkÞjEinstein dm�3s�1jVjdm3jkjmj Sjcm2j ;
ð6Þ
where NA is the Avogadro number, h is the Planck constant, c
is the speed of light, k is the wavelength, V and S are thevolume and surface of the irradiated sample, respectively, andI0(k) is the intensity of the incident light at a given wavelength,
in the units indicated inside the parenthesis. Taking intoaccount the spectral distribution of the Xenon lamp used andconsidering only the wavelengths of the maximum absorptivityof HpIX (kmin = 380 nm, kmax = 420 nm), the intensity of
light used (I0 = 1.3 · 10)3 Einstein dm)3 s)1), correspondingto I0 = 0.15 W cm)2, is comparable to the high fluence ratesused in in vivo photodynamic experiments (2,16). HpIX also
absorbs light at other wavelengths and the intensity of the totallight absorbed, at the beginning of the experiment, wasIabs = 5 · 10)4 Einstein dm)3 s)1 or 0.4 W cm)2. The light
absorbed by HpIX decreased as the irradiation proceeded dueto auto-oxidation of the porphyrin moiety. After 1 h ofirradiation the concentration of HpIX was reduced to about
half the initial concentration resulting in a reduction in 30% inthe light absorbed, in reasonable agreement with previouslyfound quantum yields of HpIX photobleaching (44).
Oxidation of cholesterol yields different products depending
on the oxidants involved (free radicals or singlet oxygen) andthis has been used to distinguish between Type I and Type IIphotosensitization (42,43). The observation of several products
of cholesterol oxidation at the 5 and 6 carbons is indicative ofoxidation via singlet oxygen. Cholesterol oxidized at carbon 7is indicative of free radical oxidation but may also result from
decomposition of cholesterol oxidized in position 5 (43,45). Toevaluate the regioselectivity effects in the oxidation of choles-terol, we also analyzed the products formed upon irradiationof HpIX in the presence of cholesterol in homogeneous media.
The products formed, and more particularly the ratio betweenthem, were very different from those found upon oxidation inthe lipid bilayer indicating that the local environment is very
important in the oxidation products formed. For irradiation ofHpIX in acetonitrile, the most abundant cholesterol oxidationproduct obtained was cholesta-4,6-dien-3-one which was a
minor product found for the irradiation of cholesterol in aPOPC:chol lipid bilayer. These results clearly indicate that thesystems used to characterize the effects of PDT in live cells
should resemble the biological environment as much aspossible.
The amount of oxidation products found is indicative of anefficient reaction between the singlet oxygen formed by
irradiation of HpIX and cholesterol. To better evaluate theefficiency of this process we measured the amount of singletoxygen that reaches the aqueous phase via its reaction with
ADPA (10). From the rate of the decrease in the concentrationof ADPA the concentration of 1O2 in the aqueous phase maybe calculated (knowing the intensity of light absorbed, the rate
of decay of 1O2 in the different environments due to quenchingby the solvent, the relative concentration of 1O2 in the different
Photochemistry and Photobiology, 2009, 85 1413

phases and the rate of reaction with ADPA) and from this theapparent quantum yield of 1O2 production may be obtained(10). Considering a partition coefficient for 1O2 between theaqueous phase and the lipid bilayer of 3 (13), we obtain an
apparent quantum yield of 1O2 production by HpIX in thePOPC:Chol 1:1 bilayer of 0.01. This value is much smallerthan the reported quantum yield of 0.7 for HpIX in lipid
bilayers (26) and the difference reflects the very efficientreaction of 1O2 with the bilayer components resulting in areduction in its steady-state concentration. We evaluated the
concentration of 1O2 in the bilayer phase using its reactionwith DPA and the method reported in the literature (26).Surprisingly we found a strong interaction between HpIX and
DPA with a reduction in the fluorescence from DPA and didnot continue those experiments. A decrease in the fluorescenceof DPA was expected due to energy transfer to HpIX as theemission spectrum of DPA overlaps significantly with the
absorption of HpIX. However, the fluorescence reduction wasmuch stronger than that predicted for a random distribution ofboth solutes in the lipid bilayer. This behavior may be due to
the microheterogeneous structure of the POPC:Chol lipidbilayer (38) that can lead to the accumulation of both solutesat a given location in the bilayer promoting the interaction
between them.
Physical changes in a POPC:Chol 1:1 bilayer due to
HpIX-mediated photosensitization
The products of cholesterol oxidation are more polar, and
have a different spatial distribution of the polar groupscompared with cholesterol. This is expected to have profoundeffects on the fluidity and elasticity (23,46) and dipole potential
(47–49) as well as changes in the phase behavior of mixtures ofphospholipids and sterol (50,51). The solubilization of choles-terol enriched lipid bilayers by Apo A1, to form high-density
lipoproteins, is also strongly inhibited when cholesterol issubstituted by an oxidized derivative (23) possibly via some of
the above mentioned changes in the physical properties of thelipid bilayer.
A fundamental property of biological membranes is the lowpermeation of polar molecules ⁄ ions across them which enables
control of the chemical composition of aqueous compartmentsin the cell. This property is based on the nonpolar nature of thelipid bilayer interior and on the high cohesion between the well-
packed lipid hydrocarbons. Cholesterol plays an essential role inthis respect (52) due to its flat and rigid structure with a singlepolar group anchoring the molecule in the polar interface of the
bilayer and the flat, rigid and nonpolar fused rings in the centerof each monolayer of the lipid bilayer. The permeation rate ofpolar solutes through the lipid bilayer is strongly reduced for
bilayers containing cholesterol (28,52–54). The higher hydro-philic ⁄ hydrophobic ratio of oxidized cholesterol, and in partic-ular the different position of the polar groups (20,23), increasesthe polarity of the lipid bilayer center and impairs its function in
the reduction of membrane leakiness.We assessed the effects of oxidation of a lipid bilayer
prepared from POPC:Chol 1:1, by HpIX-mediated photosen-
sitization, on the rates of permeation of small ions; H+ ⁄OH)
and dithionite. In those experiments a band-pass filter at525 nm was used which significantly reduced the intensity of
the incident light compared to that used in the previous sectionfor the evaluation of the chemical effects. The intensity of theincident light was 6.4 · 10)5 Einstein dm)3 s)1 correspondingto 44 mW cm)2. The total concentration of HpIX was also
smaller (1:500 HpIX:lipid compared with 1:100) correspondingto an intensity of light absorbed equal to 7.6 · 10)7 Ein-stein dm)3 s)1 or 0.5 mW cm)2. This fluence rate is 2 orders of
magnitude smaller than that used in the previous section andmuch smaller than the one used in in vivo photodynamic cellkilling. We are therefore evaluating the physical effects of very
small oxidation injuries.The results obtained for the rate of equilibration of a pH
gradient are shown in Fig. 4. The experimental data was the
fluorescence intensity at 520 nm from CBF, encapsulated inthe internal aqueous media of the LUVs, when excited at 439and 493 nm. The ratio between the fluorescence intensity forexcitation at both wavelengths reflects the fraction of CBF in
two different ionization states and is therefore related to thepH of the aqueous solution (Fig. 4, plot A). The time profile ofthe ratio between the fluorescence intensities for excitation at
both wavelengths may be converted into the time course forvariation of the pH inside the LUVs and this is shown in plot Bfor different irradiation times.
Carboxyfluorescein fluorescence has been widely used as apH sensor in the range of pH from 5 to 9 due to the strongvariation of its fluorescence when excited at around 500 nm(55). The advantages of using the ratio of the fluorescence
intensity, when excited at around 500 and 440 nm, (56) includethe fact that the signal becomes independent of the totalconcentration of the sensor (including insensitivity to photo-
bleaching). We therefore used this pH sensor in the ratiometricmode and the calibration of the sensor is shown inFig. 4, plotA.
The LUVs were prepared at pH = 5.6 with CBF in the
internal aqueous compartment at a concentration of 50 lM.M.
The external CBF was removed by size exclusion chromatog-raphy, and the pH in the outer aqueous compartment was
rapidly changed to 6.6 by the addition of a solution atpH = 8. The equilibration of the pH across the lipid bilayer
A B
Figure 4. (A) Ratio of the fluorescence from carboxyfluorescein at520 nm when excited with radiation with wavelengths of 493 and439 nm, as a function of the pH of the aqueous solution. The line is thebest fit of a titration curve with a pKa = 6.8. (B) Change in the pHinside large unilamellar vesicles (LUVs) of POPC:Chol 1:1 afterchanging the pH in the aqueous solution outside the LUVs from 5.6 to6.6. The lipid bilayer contained hematoporphyrin IX (HpIX) at amolar ratio of 1:500 relative to total lipid and was irradiated withvisible light (525 nm, Iabs = 0.5 mW cm)2) for 0 (s), 15 (4) or30 min (e). The profile of pH equilibration in the absence of HpIX isalso shown (h).
1414 Antonio Santos et al.
was followed over time and the results are shown in Fig. 4,plot B. A fast partial pH equilibration is observed in the first
minutes followed by a slow equilibration. The quantitativeinterpretation of the pH equilibration profile is a complicatedtask (57) that has not yet been fully solved and therefore we
will interpret the results obtained solely on a qualitative basis.The introduction of HpIX in the lipid bilayer increased slightlythe rate of pH equilibration in agreement with an increase
in the membrane polarity and with the weak acid properties ofHpIX. Irradiation of the lipid bilayer containing HpIXstrongly increased the rate of pH equilibration with 75%approach to equilibrium attained after 1 min for the sample
irradiated for 30 min as compared with 48% for the lipidbilayer containing HpIX but without irradiation, and 43% forthe lipid bilayer in the absence of HpIX. Irradiation in the
absence of HpIX did not change the profile of pH equilibra-tion (results not shown).
To quantitatively assess the changes in the leakiness of the
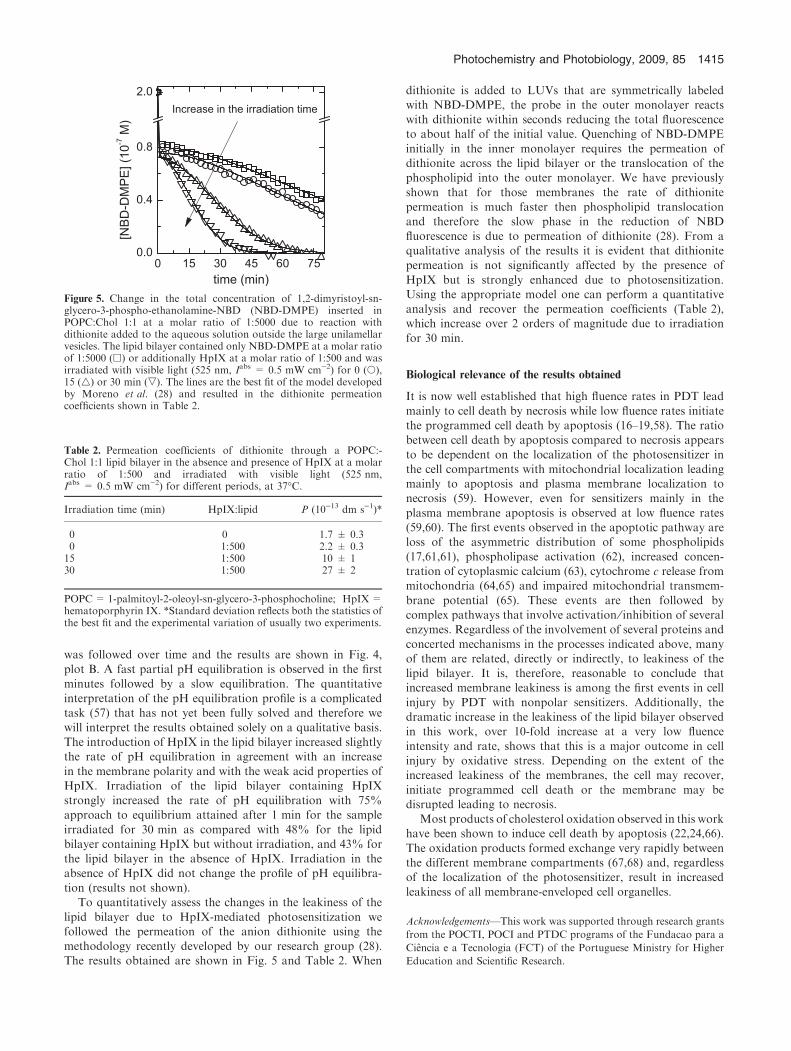
lipid bilayer due to HpIX-mediated photosensitization wefollowed the permeation of the anion dithionite using themethodology recently developed by our research group (28).The results obtained are shown in Fig. 5 and Table 2. When
dithionite is added to LUVs that are symmetrically labeledwith NBD-DMPE, the probe in the outer monolayer reactswith dithionite within seconds reducing the total fluorescenceto about half of the initial value. Quenching of NBD-DMPE
initially in the inner monolayer requires the permeation ofdithionite across the lipid bilayer or the translocation of thephospholipid into the outer monolayer. We have previously
shown that for those membranes the rate of dithionitepermeation is much faster then phospholipid translocationand therefore the slow phase in the reduction of NBD
fluorescence is due to permeation of dithionite (28). From aqualitative analysis of the results it is evident that dithionitepermeation is not significantly affected by the presence of
HpIX but is strongly enhanced due to photosensitization.Using the appropriate model one can perform a quantitativeanalysis and recover the permeation coefficients (Table 2),which increase over 2 orders of magnitude due to irradiation
for 30 min.
Biological relevance of the results obtained
It is now well established that high fluence rates in PDT lead
mainly to cell death by necrosis while low fluence rates initiatethe programmed cell death by apoptosis (16–19,58). The ratiobetween cell death by apoptosis compared to necrosis appears
to be dependent on the localization of the photosensitizer inthe cell compartments with mitochondrial localization leadingmainly to apoptosis and plasma membrane localization tonecrosis (59). However, even for sensitizers mainly in the
plasma membrane apoptosis is observed at low fluence rates(59,60). The first events observed in the apoptotic pathway areloss of the asymmetric distribution of some phospholipids
(17,61,61), phospholipase activation (62), increased concen-tration of cytoplasmic calcium (63), cytochrome c release frommitochondria (64,65) and impaired mitochondrial transmem-
brane potential (65). These events are then followed bycomplex pathways that involve activation ⁄ inhibition of severalenzymes. Regardless of the involvement of several proteins and
concerted mechanisms in the processes indicated above, manyof them are related, directly or indirectly, to leakiness of thelipid bilayer. It is, therefore, reasonable to conclude thatincreased membrane leakiness is among the first events in cell
injury by PDT with nonpolar sensitizers. Additionally, thedramatic increase in the leakiness of the lipid bilayer observedin this work, over 10-fold increase at a very low fluence
intensity and rate, shows that this is a major outcome in cellinjury by oxidative stress. Depending on the extent of theincreased leakiness of the membranes, the cell may recover,
initiate programmed cell death or the membrane may bedisrupted leading to necrosis.
Most products of cholesterol oxidation observed in this workhave been shown to induce cell death by apoptosis (22,24,66).
The oxidation products formed exchange very rapidly betweenthe different membrane compartments (67,68) and, regardlessof the localization of the photosensitizer, result in increased
leakiness of all membrane-enveloped cell organelles.
Acknowledgements—This work was supported through research grants
from the POCTI, POCI and PTDC programs of the Fundacao para a
Ciencia e a Tecnologia (FCT) of the Portuguese Ministry for Higher
Education and Scientific Research.
Table 2. Permeation coefficients of dithionite through a POPC:-Chol 1:1 lipid bilayer in the absence and presence of HpIX at a molarratio of 1:500 and irradiated with visible light (525 nm,Iabs = 0.5 mW cm)2) for different periods, at 37�C.
Irradiation time (min) HpIX:lipid P (10)13 dm s)1)*
0 0 1.7 ± 0.30 1:500 2.2 ± 0.315 1:500 10 ± 130 1:500 27 ± 2
POPC= 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine; HpIX=hematoporphyrin IX. *Standard deviation reflects both the statistics ofthe best fit and the experimental variation of usually two experiments.
Figure 5. Change in the total concentration of 1,2-dimyristoyl-sn-glycero-3-phospho-ethanolamine-NBD (NBD-DMPE) inserted inPOPC:Chol 1:1 at a molar ratio of 1:5000 due to reaction withdithionite added to the aqueous solution outside the large unilamellarvesicles. The lipid bilayer contained only NBD-DMPE at a molar ratioof 1:5000 (h) or additionally HpIX at a molar ratio of 1:500 and wasirradiated with visible light (525 nm, Iabs = 0.5 mW cm)2) for 0 (s),15 (4) or 30 min (,). The lines are the best fit of the model developedby Moreno et al. (28) and resulted in the dithionite permeationcoefficients shown in Table 2.
Photochemistry and Photobiology, 2009, 85 1415

REFERENCES1. Dolmans, D. E. J. G., D. Fukumura and R. K. Jain (2003)
Photodynamic therapy for cancer. Nat. Rev. Cancer 3, 380–387.2. Dougherty, T. J. (1993) Photodynamic therapy. Photochem.
Photobiol. 58, 895–900.3. Ackroyd, R., C. Kelty, N. Brown and M. Reed (2001) The history
of photodetection and photodynamic therapy. Photochem.Photobiol. 74, 656–669.
4. Hamblin, M. R. and T. Hasan (2004) Photodynamic therapy: Anew antimicrobial approach to infectious disease? Photochem.Photobiol. Sci. 3, 436–450.
5. Dougherty, T. J. (2002) An update on photodynamic therapyapplications. J. Clin. Laser Med. Surg. 20, 3–7.
6. Verma, S., G. M. Watt, Z. Mal and T. Hasan (2007) Strategies forenhanced photodynamic therapy effects. Photochem. Photobiol.83, 996–1005.
7. Sobral, A. J. F. N., S. Eleouet, N. Rousset, A. M. D. Gonsalves,O. Le Meur, L. Bourre and T. Patrice (2002) New sulfonamideand sulfonic ester porphyrins as sensitizers for photodynamictherapy. J. Porphyrins Phthalocyanines 6, 456–462.
8. Zuluaga, M. F. and N. Lange (2008) Combination of photo-dynamic therapy with anti-cancer agents. Curr. Med. Chem. 15,1655–1673.
9. Koo, Y. E. L., G. R. Reddy, M. Bhojani, R. Schneider, M. A.Philbert, A. Rehemtulla, B. D. Ross and R. Kopelman (2006)Brain cancer diagnosis and therapy with nanoplatforms. Adv.Drug Deliv. Rev. 58, 1556–1577.
10. Moreno, M. J., E. Monson, R. G. Reddy, A. Rehemtulla, B. D.Ross, M. Philbert, R. J. Schneider and R. Kopelman (2003)Production of singlet oxygen by Ru(dpp(SO3)2)3 incorporatedin polyacrylamide PEBBLES. Sensor. Actuator. B-Chem. 90,82–89.
11. MacDonald, I. J. and T. J. Dougherty (2001) Basic principles ofphotodynamic therapy. J. Porphyrins Phthalocyanines 5, 105–129.
12. Henderson, B. W. and T. J. Dougherty (1992) How does photo-dynamic therapy work? Photochem. Photobiol. 55, 145–157.
13. Subczynski, W. K. and J. S. Hyde (1983) Concentration of oxygenin lipid bilayers using a spin-label method. Biophys. J. 41, 283–286.
14. Ochsner, M. (1997) Photophysical and photobiological processesin the photodynamic therapy of tumours. J. Photochem. Photobiol.B, Biol. 39, 1–18.
15. Degoeij, A. F. P. M. and J. VanSteveninck (1976) Photodynamiceffects of protoporphyrin on cholesterol and unsaturated fatty-acids in erythrocyte-membranes in protoporphyria and in normalred blood-cells. Clin. Chim. Acta 68, 115–122.
16. Henderson, B. W., S. O. Gollnick, J. W. Snyder, T. M. Busch,P. C. Kousis, R. T. Cheney and J. Morgan (2004) Choice ofoxygen-conserving treatment regimen determines the inflamma-tory response and outcome of photodynamic therapy of tumors.Cancer Res. 64, 2120–2126.
17. Di Stefano, A., A. Ettorre, S. Sbrana, C. Giovani and P. Neri(2001) Purpurin-18 in combination with light leads to apoptosis ornecrosis in HL60 leukemia cells. Photochem. Photobiol. 73, 290–296.
18. Wyld, L., M. W. R. Reed and N. J. Brown (2001) Differential celldeath response to photodynamic therapy is dependent on dose andcell type. Br. J. Cancer 84, 1384–1386.
19. Kessel, D. and R. D. Poretz (2000) Sites of photodamage inducedby photodynamic therapy with a chlorin e6 triacetoxymethyl ester(CAME). Photochem. Photobiol. 71, 94–96.
20. Massey, J. B. and H. J. Pownall (2006) Structures of biologicallyactive oxysterols determine their differential effects on phospho-lipid membranes. Biochemistry 45, 10747–10758.
21. Wielkoszynski, T., K. Gawron, J. Strzelczyk, P. Bodzek, M.Zalewska-Ziob, G. Trapp, M. Srebniak and A. Wiczkowski(2006) Cellular toxicity of oxycholesterols. Bioessays 28, 387–398.
22. Berthier, A., S. Lemaire-Ewing, C. Prunet, T. Montange,A. Vejux, J. P. Pais de Barros, S. Monier, P. Gambert, G. Lizardand D. Neel (2005) 7-Ketocholesterol-induced apoptosis. FEBS J.272, 3093–3104.
23. Massey, J. B. and H. J. Pownall (2005) The polar nature of 7-ketocholesterol determines its location within membrane domains
and the kinetics of membrane microsolubilization by apolipo-protein A-I. Biochemistry 44, 10423–10433.
24. Seo, D. W., H. S. Choi, S. P. Lee and R. Kuver (2004) Oxysterolsfrom human bile induce apoptosis of canine gallbladder epithelialcells in monolayer culture. Am. J. Physiol. Gastrointest. LiverPhysiol. 287, G1247–G1256.
25. Calzavara-Pinton, P. G., M. Venturini and R. Sala (2005) Acomprehensive overview of photodynamic therapy in the treat-ment of superficial fungal infections of the skin. J. Photochem.Photobiol. B, Biol. 78, 1–6.
26. Bronshtein, I., M. Afri, H. Weitman, A. A. Frimer, K. M. Smithand B. Ehrenberg (2004) Porphyrin depth in lipid bilayers asdetermined by iodide and parallax fluorescence quenching meth-ods and its effect on photosensitizing efficiency. Biophys. J. 87,1155–1164.
27. Lavi, A., H. Weitman, R. T. Holmes, K. M. Smith andB. Ehrenberg (2002) The depth of porphyrin in a membrane andthe membrane’s physical properties affect the photosensitizingefficiency. Biophys. J. 82, 2101–2110.
28. Moreno, M. J., L. M. B. B. Estronca and W. L. C. Vaz (2006)Translocation of phospholipids and dithionite permeability in li-quid-ordered and liquid-disordered membranes. Biophys. J. 91,873–881.
29. Kuzelova, K. and D. Brault (1994) Kinetic and equilibrium studiesof porphyrin interactions with unilamellar lipidic vesicles.Biochemistry 33, 9447–9459.
30. Wiener, M. C. and S. H. White (1992) Structure of a fluid diol-eoylphosphatidylcholine bilayer determined by joint refinement ofX-ray and neutron-diffraction data. 3. Complete structure.Biophys. J. 61, 434–447.
31. Wiener, M. C. and S. H. White (1992) Structure of a fluid diol-eoylphosphatidylcholine bilayer determined by joint refinement ofX-ray and neutron-diffraction data. 2. Distribution and packingof terminal methyl-groups. Biophys. J. 61, 428–433.
32. Smaby, J. M., M. M. Momsen, H. L. Brockman and R. E. Brown(1997) Phosphatidylcholine acyl unsaturation modulates thedecrease in interfacial elasticity induced by cholesterol. Biophys.J. 73, 1492–1505.
33. Maman, N. and D. Brault (1998) Kinetics of the interactions of adicarboxylic porphyrin with unilamellar lipidic vesicles: Interplaybetween bilayer thickness and pH in rate control. Biochim. Bio-phys. Acta Biomembr. 1414, 31–42.
34. Margalit, R. and M. Rotenberg (1984) Thermodynamics of por-phyrindimerization in aqueous-solutions.Biochem. J.219, 445–450.
35. Brown, S. B., M. Shillcock and P. Jones (1976) Equilibrium andkinetic studies of aggregation of porphyrins in aqueous-solution.Biochem. J. 153, 279–285.
36. Mateo, C. R., A. U. Acuna and J. C. Brochon (1995) Liquid-crystalline phases of cholesterol lipid bilayers as revealed by thefluorescence of trans-parinaric acid. Biophys. J. 68, 978–987.
37. Kuzelova, K. and D. Brault (1995) Interactions of dicarbox-ylic porphyrins with unilamellar lipidic vesicles—drastic effectsof pH and cholesterol on kinetics. Biochemistry 34, 11245–11255.
38. Estronca, L. M. B. B., M. J. Moreno and W. L. C. Vaz (2007)Kinetics and thermodynamics of the association of dehydroer-gosterol with lipid bilayer membranes. Biophys. J. 93, 4244–4253.
39. Sampaio, J. L., M. J. Moreno and W. L. C. Vaz (2005) Kineticsand thermodynamics of association of a fluorescent lysophos-pholipid derivative with lipid bilayers in liquid-ordered and liquid-disordered phases. Biophys. J. 88, 4064–4071.
40. Abreu, M. S. C., M. J. Moreno and W. L. C. Vaz (2004) Kineticsand thermodynamics of association of a phospholipid derivativewith lipid bilayers in liquid-disordered and liquid-ordered phases.Biophys. J. 87, 353–365.
41. Madden, K. P. The Radiation Chemistry Data Center of theNotre Dame Radiation Laboratory. Available at: http://www.rcdc.nd.edu/index.html. Accessed on 26 April 2009.
42. Girotti, A. W. and W. Korytowski (2000) Cholesterol as a singletoxygen detector in biological systems. Methods Enzymol. 319, 85–100.
43. Kulig, M. J. and L. L. Smith (1973) Sterol metabolism. 25. Cho-lesterol oxidation by singlet molecular-oxygen. J. Org. Chem. 38,3639–3642.
1416 Antonio Santos et al.

44. Spikes, J. D. (1992) Quantum yields and kinetics of the photo-bleaching of hematoporphyrin, photofrin-II, tetra(4-sulfonatophe-nyl)-porphine and uroporphyrin. Photochem. Photobiol. 55, 797–808.
45. Nakano, M., K. Sugioka, T. Nakamura and T. Oki (1980)Interaction between an organic hydroperoxide and an unsaturatedphospholipid and alpha-tocopherol in model membranes.Biochim. Biophys. Acta 619, 274–286.
46. Li, X. M., M. M. Momsen, H. L. Brockman and R. E. Brown(2003) Sterol structure and sphingomyelin acyl chain lengthmodulate lateral packing elasticity and detergent solubility inmodel membranes. Biophys. J. 85, 3788–3801.
47. Asawakarn, T., J. Cladera and P. O’Shea (2001) Effects of themembrane dipole potential on the interaction of saquinavir withphospholipid membranes and plasma membrane receptors ofCaco-2 cells. J. Biol. Chem. 276, 38457–38463.
48. Gross, E., R. S. Bedlack and L. M. Loew (1994) Dual-wavelengthratiometric fluorescence measurement of the membrane dipolepotential. Biophys. J. 67, 208–216.
49. Alakoskela, J. M. I., T. Soderlund, J. M. Holopainen and P. K. J.Kinnunen (2004) Dipole potential and head-group spacing aredeterminants for the membrane partitioning of pregnanolone.Mol. Pharmacol. 66, 161–168.
50. Beattie, M. E., S. L. Veatch, B. L. Stottrup and S. L. Keller (2005)Sterol structure determines miscibility versus melting transitions inlipid vesicles. Biophys. J. 89, 1760–1768.
51. Wang, J. W., Megha and E. London (2004) Relationship betweensterol ⁄ steroid structure and participation in ordered lipid domains(lipid rafts): Implications for lipid raft structure and function.Biochemistry 43, 1010–1018.
52. Szabo, G. (1974) Dual mechanism for action of cholesterol onmembrane-permeability. Nature 252, 47–49.
53. Degier, J., J. G. Mandersl and L. L. Vandeene (1968) Lipidcomposition and permeability of liposomes. Biochim. Biophys.Acta 150, 666–675.
54. Marsh, D. (2001) Polarity and permeation profiles in lipid mem-branes. Proc. Natl Acad. Sci. USA 98, 7777–7782.
55. Rink, T. J., R. Y. Tsien and T. Pozzan (1982) Cytoplasmic pH andfree Mg-2+ in lymphocytes. J. Cell Biol. 95, 189–196.
56. Selvaggio, A. M., J. H. Schwartz, H. H. Bengele and E. A.Alexander (1986) Kinetics of the Na+-H+ antiporter as assessed
by the change in intracellular pH in Mdck-Cells. Am. J. Physiol.251, C558–C562.
57. Deamer, D. W. and J. W. Nichols (1983) Proton hydroxidepermeability of liposomes. Proc. Natl Acad. Sci. USA 80, 165–168.
58. Fabris, C., G. Valduga, G. Miotto, L. Borsetto, G. Jori,S. Garbisa and E. Reddi (2001) Photosensitization with zinc (II)phthalocyanine as a switch in the decision between apoptosis andnecrosis. Cancer Res. 61, 7495–7500.
59. Moor, A. C. E. (2000) Signaling pathways in cell death and sur-vival after photodynamic therapy. J. Photochem. Photobiol.B, Biol. 57, 1–13.
60. Lin, C. P., M. C. Lynch and I. E. Kochevar (2000) Reactiveoxidizing species produced near the plasma membrane induceapoptosis in bovine aorta endothelial cells. Exp. Cell Res. 259,351–359.
61. Ruck, A., K. Heckelsmiller, R. Kaufmann, N. Grossman, E.Haseroth and N. Akgun (2000) Light-induced apoptosis involves adefined sequence of cytoplasmic and nuclear calcium release inAlPcS4-photosensitized rat bladder RR 1022 epithelial cells.Photochem. Photobiol. 72, 210–216.
62. Ahmad, N. and H. Mukhtar (2000) Mechanism of photodynamictherapy-induced cell death. Methods Enzymol. 319, 342–358.
63. Panini, S. R. and M. S. Sinensky (2001) Mechanisms of oxysterol-induced apoptosis. Curr. Opin. Lipidol. 12, 529–533.
64. Chiu, S. M. and N. L. Oleinick (2001) Dissociation of mito-chondrial depolarization from cytochrome c release duringapoptosis induced by photodynamic therapy. Br. J. Cancer 84,1099–1106.
65. Oleinick, N. L., R. L. Morris and T. Belichenko (2002) The role ofapoptosis in response to photodynamic therapy: What, where,why, and how. Photochem. Photobiol. Sci. 1, 1–21.
66. Schroepfer, G. J. (2000) Oxysterols: Modulators of cholesterolmetabolism and other processes. Physiol. Rev. 80, 361–554.
67. Girotti, A. W. and T. Kriska (2004) Role of lipid hydroperoxidesin photo-oxidative stress signaling. Antioxid. Redox Signal. 6, 301–310.
68. Vila, A., W. Korytowski and A. W. Girotti (2001) Spontaneousintermembrane transfer of various cholesterol-derived hydroper-oxide species: Kinetic studies with model membranes and cells.Biochemistry 40, 14715–14726.
Photochemistry and Photobiology, 2009, 85 1417