ecophysiology of the tree fern species dicksonia ......ecophysiology of the tree fern species...
TRANSCRIPT

Ecophysiology of the tree fern species
Dicksonia antarctica Labill
and Cyathea australis (R. Br.) Domin
Liubov Vladimirovna Volkova
Submitted in total fulfilment of the requirements
of the degree of Doctor of Philosophy
October 2009
Department of Forest and Ecosystem Science
Melbourne School of Land and Environment
The University of Melbourne
Produced on archival quality paper

-i-
Abstract
Predictions of global warming and associated climate change indicate widespread in-
creases in light intensities, temperatures, and the frequency and severity of droughts in
south-eastern Australia. Understanding the ability of plants to respond and acclimate to
these events is essential to predict species survival and potential impacts on biodiver-
sity.
This study focuses on two tree fern species – Dicksonia antarctica and Cyathea aus-
tralis – two iconic understorey species of south-east Australian forests. These tree ferns
belong to different families and are of contrasting origins, yet often grow together in
south-eastern Australia, typically in shade, often along waterways. Their ecological im-
portance is evident in the high epiphytic diversity on their trunks (ferns, mosses, bryo-
phytes, liverwort etc), and the provision of nursery sites for many tree and shrub spe-
cies. Both species are decreased by timber harvesting practices such as clearcut logging,
with deaths continuing for up to five years in the post-harvest environment. Understand-
ing the relative roles of changing light, water, and temperature in these ongoing declines
is essential to conserving both tree fern populations and their dependent biota.
The Thesis encompasses three controlled experiments and a field study. In the con-
trolled experiments, the tree ferns were acclimated to contrasting growth light environ-
ments (shade or moderate light) and then exposed to an environmental stress (i.e. light,
heat, water deficit). The field study examined relationships between environmental
variables (i.e. light, temperature, plant water status) and photosynthetic capacity pa-
rameters of the tree ferns in their natural environment. Stress responses and acclimation
potential of photosynthetic traits, water relation parameters, and frond traits of the tree
ferns were studied using infra-red gas analysis, pigment determination techniques, and
stable isotope methods.
It was hypothesised that, consistent with their contrasting origins and micro-site prefer-
ences, the two tree fern species would possess different physiological characteristics

-ii-
and therefore respond differently to environmental stresses. It was also hypothesised
that plants grown under contrasting light environments would have different reactions to
and recoveries from environmental stresses.
Overall, plants were able to sustain and recover from high light stress, while interactive
effects of high light and heat were most detrimental to tree fern performance. Both spe-
cies were susceptible to water stress, either alone or in combination with high light. The
hypothesised different responses of the two species (associated with their different ori-
gins) were not confirmed, and reaction to and recovery from stress was mainly unaf-
fected by growth light environment. Both species had low acclimation potential to any
of the applied environmental stresses. Overall, findings from this study indicate that
combined effects of high light and heat most likely cause ongoing decline of tree ferns
in post-harvest environments, and that the distribution of tree ferns will most likely con-
tract under future climate scenarios of higher light, increased temperatures, and de-
creased water availability.

-iii-
Declaration
This is to certify that:
the thesis comprises only my original work towards the PhD except where indicated
in the Preface,
due acknowledgement has been made in the text to all other material used,
the thesis is less than 100,000 words in length, exclusive of tables, maps, bibliogra-
phies and appendices
Liubov Volkova

-iv-
Preface
The climate chamber experiment, Chapter 3, was undertaken in Champenoux, France
using facilities of the Institut National Reserche Agronomique (INRA). I planned, con-
ducted the research, evaluated and presented the data. The results of the study were pre-
sented at the International Eco-Fizz conference, 2007 (a poster) and published in the
scientific journal Functional Plant Biology (Volkova L, Tausz M, Bennett LT, Dreyer
E, 2009. „Interactive effects of high irradiance and moderate heat on photosynthesis,
pigments, and tocopherol in the tree-fern Dicksonia antarctica’). M. Tausz and L.T.
Bennett are the supervisors of my PhD work. Erwin Dreyer, the fourth co-author of the
publication was a hosting party in INRA and supervised my activities. Professor Dreyer
is also an honorary staff member of the Department of Forest and Ecosystem Science at
The University of Melbourne.
Chapter 4 (high light and water stress experiment) has been submitted for publication in
co-authorship with my supervisors and Dr. Andrew Merchant (the University of Syd-
ney). I declare that the execution of the experiment, data evaluation and presentation
were solely my own work, and that A. Merchant gave useful tips on the experimental
design and helped to organise the isotopic analysis of my samples.
Chapters 2 and 5 are written in co-authorships with my supervisors M. Tausz and L.T.
Bennett who helped with usual supervisory roles.

-v-
Acknowledgements
Personal financial support for this study was provided by a Melbourne Research Schol-
arship. Expenses related to research activities (i.e. field study and construction of the
controlled experiments) were partly covered by a research agreement with the Victorian
Department of Sustainability and Environment (TA30874).
I am personally grateful to my University supervisors, Ass. Prof. Michael Tausz and Dr
Lauren Bennett, for their patience, ongoing support and encouragement during my
study. I admire Michael for his ability to think globally and to give me confidence that
everything is possible. I admire Ren for her strong personality, always prompt re-
sponses; ability to carefully examine every detail; her great friendship and care when I
needed it. She was (and is) the Woman, who made me deeply respect women in science.
I am indebted to Erwin Dreyer (INRA, France) for his great deal of support and advice
during my candidature; his personal friendship is very precious to me. I would like to
thank Andrew Merchant for his advice and support throughout my study. My thanks to
Chris Western for his patience, always good advice and for being my personal Counsel-
lor at difficult times.
I acknowledge staff and students at Creswick campus for their support. Thanks particu-
larly to Thomas Wright for his friendship and ongoing help, and Raymond Dempsey for
his help in the field. Thanks also to Matt Lee and Najib Ahmady for always providing
reliable and timely results, and my thanks to all others.
I am grateful to my family: husband, Fedor Torgovnikov, for his patience, support and
help during my study. His ability to fix equipment and build constructions for my ex-
periments was priceless. His patience with my often bad moods due to problems with
experiments and understanding my difficulties made me able to finish this study. I thank
my daughter, Katerina Torgovnikova, for her help with watering and re-potting plants
and her patience with “always busy mum”. I am thankful to my parents-in-law, and
most of all, I want to thank my mum, Svetlana Volkova, for teaching me to never give
up and always reach my targets.

-vi-
Table of Contents
Abstract ...................................................................................................................... i
Declaration .............................................................................................................. iii
Preface ..................................................................................................................... iv
Acknowledgements .................................................................................................... v
Table of Contents ..................................................................................................... vi
List of Figures .......................................................................................................... xi
Chapter 1. Introduction .......................................................................................... 1
1.1. Environmental stresses: light, temperature and water deficit ...................... 1
1.2. Fundamental effects of high irradiance in interaction with high
temperature or drought on plants ............................................................................. 1
1.3. Two tree ferns of contrasting origin ............................................................. 3
1.4. The tree ferns in mountain ash forests of south-eastern Australia ............... 5
1.5. Current knowledge of tree fern ecophysiology ............................................. 6
1.6. Thesis aims and outline ................................................................................. 8
Chapter 2. Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica, and Cyathea australis, acclimated to different light
intensities. ............................................................................................................ 11
(i) Abstract ....................................................................................................... 11
2.1. Introduction ................................................................................................ 11
2.2. Materials and methods ................................................................................ 13
2.2.1. Plant material ......................................................................................... 13
2.2.2. Experimental design ............................................................................... 14
2.2.3. Light environment of measured frond ..................................................... 15
2.2.4. Maximal quantum yield of photochemistry (Fv/Fm) ................................ 16
2.2.5. Gas exchange measurements .................................................................. 16
2.2.6. Plant water status ................................................................................... 17
2.2.7. Frond traits ............................................................................................. 17
2.2.8. Artificial sunfleck experiment ................................................................. 18
2.2.9. Statistical analyses .................................................................................. 19

-vii-
2.3. Results ......................................................................................................... 19
2.3.1. Light environment of the tree ferns ......................................................... 19
Maximum quantum yield of photochemistry (Fv/Fm) .......................................... 20
2.3.2. Photosynthetic capacity parameters ....................................................... 20
2.3.3. Plant water status ................................................................................... 26
2.3.4. Frond traits ............................................................................................. 26
2.3.5. Artificial sunfleck experiment ................................................................. 29
2.4. Discussion ................................................................................................... 31
2.4.1. Species overview ..................................................................................... 31
2.4.2. Acclimation to growth light environment ............................................... 31
2.4.3. High light stress ...................................................................................... 33
2.4.4. Acclimation to new light environment .................................................... 33
2.5. Summary ..................................................................................................... 34
Chapter 3. Interactive effects of high irradiance and moderate heat on
photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. .. 35
(ii) Abstract ....................................................................................................... 35
3.1. Introduction ................................................................................................ 35
3.2. Material and methods ................................................................................. 39
3.2.1. Plant material ......................................................................................... 39
3.2.2. Climate chamber conditions and experimental design ........................... 39
3.2.3. Frond temperature (Tfrond) ...................................................................... 40
3.2.4. Maximal quantum yield of photochemistry (Fv/Fm) ................................ 40
3.2.5. Gas exchange measurements .................................................................. 41
3.2.6. Frond nitrogen and chlorophyll content ................................................. 42
3.2.7. Critical temperature (Tc) ........................................................................ 42
3.2.8. Total tissue osmolality ............................................................................ 43
3.2.9. Pigments and tocopherol determination ................................................. 43
3.2.10. Statistical analysis .............................................................................. 44
3.3. Results ......................................................................................................... 45
3.3.1. Frond temperature (Tfrond) ...................................................................... 45

-viii-
3.3.2. Maximum quantum yield of PS II (Fv/Fm) and photosynthetic capacity
parameters .......................................................................................................... 46
3.3.3. Critical temperature (Tc) ........................................................................ 52
3.3.5. Carotenoids and α-tocopherol ................................................................ 53
3.3.6. Correlations between Tc and biochemical parameters ........................... 57
3.4. Discussion ................................................................................................... 58
3.4.1. Effect of high irradiance, high temperature and their interaction on
photosynthetic capacity parameters of D. antarctica ......................................... 58
3.4.2. Membrane stability of D. antarctica measured via critical temperature 60
3.4.3. Xanthophyll cycle carotenoids, pigments and α-tocopherol ................... 61
3.5. Summary ..................................................................................................... 63
Chapter 4. Interactive effects of high light and water deficit on the tree fern
species Dicksonia antarctica and Cyathea australis .............................................. 65
(iii) Abstract ................................................................................................... 65
4.1. Introduction ................................................................................................ 65
4.2. Materials and methods ................................................................................ 68
4.2.1. Plant material ......................................................................................... 68
4.2.2. Experimental design ............................................................................... 68
4.2.3. Maximum quantum yield of PSII (Fv/Fm) ................................................ 70
4.2.4. Photosynthetic capacity .......................................................................... 70
4.2.5. Frond water relations ............................................................................. 72
4.2.6. Stable isotope analysis ............................................................................ 72
4.2.7. Relative extractable soil water, REW ..................................................... 72
4.2.8. Statistical analysis .................................................................................. 73
4.3. Results ......................................................................................................... 74
4.3.1. Maximum quantum yield of PS II (Fv/Fm) ............................................... 74
4.3.2. Photosynthetic capacity .......................................................................... 74
4.3.3. Frond survival ......................................................................................... 79
4.3.4. Time course of stomatal conductance during 5 days without water ....... 79
4.3.5. Frond water relations ............................................................................. 81

-ix-
4.3.6. Intrinsic water use efficiency (calculated as Amax/gs, WUEi) and stable
carbon isotope composition (δ13
C) ..................................................................... 83
4.4. Discussion ................................................................................................... 85
4.4.1. Pre-treatment period – species differences and effect of light ............... 85
4.4.2. Water deficit and light interactions ........................................................ 86
4.4.3. Rewatering period ................................................................................... 88
4.5. Summary ..................................................................................................... 89
Chapter 5. Seasonal variations in photosynthesis of the tree ferns Dicksonia
antarctica and Cyathea australis in wet sclerophyll forests of Australia .............. 91
(iv) Abstract ................................................................................................... 91
5.1. Introduction ................................................................................................ 91
5.2. Materials and methods ................................................................................ 94
5.2.1. Study site and sampling design ............................................................... 94
5.2.2. Tree fern measurement schedule ............................................................ 95
5.2.3. Mean irradiance on measured fronds ..................................................... 96
5.2.4. Maximal quantum yield of photochemistry (Fv/Fm) ................................ 96
5.2.5. Gas exchange measurements .................................................................. 96
5.2.6. Frond water potential ............................................................................. 97
5.2.7. Frond traits ............................................................................................. 97
5.2.8. Statistical analysis .................................................................................. 98
5.3. Results ......................................................................................................... 99
5.3.1. Relationships between photosynthesis, growth irradiance and
temperature ......................................................................................................... 99
5.3.2. Water status parameters ....................................................................... 103
5.3.3. Diurnal measurements .......................................................................... 103
5.3.4. Stomatal density .................................................................................... 106
5.4. Discussion ................................................................................................. 106
5.4.1. Comparisons between the two tree fern species ................................... 106
5.4.2. Light as a limiting factor to tree fern photosynthetic performance ...... 107
5.4.3. Temperature as a limiting factor to tree fern photosynthetic performance
.............................................................................................................. 109

-x-
5.4.4. Effects of plant water status and water relation parameters on tree fern
photosynthetic performance .............................................................................. 110
5.4.5. Stomatal density .................................................................................... 111
5.5. Summary ................................................................................................... 113
Chapter 6. Ecophysiology of two tree fern species and implications for their
future management. General discussion and conclusions .................................... 115
6.1. Species overview ....................................................................................... 115
6.2. Overview of light, temperature, and water availability as stresses on tree
fern physiology ...................................................................................................... 117
6.3. Practical implications and future directions ............................................ 119
REFERENCES ...................................................................................................... 123

-xi-
List of Figures
Figure 2.1 Photosynthetic capacity parameters of the tree ferns D. antarctica and C.
australis grown under variable light and shade during the before exposure period, and
then exposed to high light, and measured after two weeks (short-term exposure) and
three months (long-term exposure). ...........................................................................
Figure 2.2 Mesophyll capacity parameters of the tree ferns D. antarctica and C.
australis grown under variable light and shade during the before exposure period, and
then exposed to high light, and measured after two weeks (short-term exposure) and tree
months (long-term exposure) ................................................................................. 24
Figure 3.1. Time course of maximum quantum efficiency of PSII and chlorophyll
content of high irradiance and shaded D. antarctica during three successive temperature
treatments ............................................................................................................... 47
Figure 3.2 Stomatal conductance versus light-saturated rate of net photosynthesis for
high irradiance and shaded D. antarctica .............................................................. 51
Figure 3.3 Time course of critical temperature Tc D. antarctica across the experiment
............................................................................................................................... 52
Figure 3.4 α -Tocopherol content of high irradiance and shaded D. antarctica under
three temperature ................................................................................................... 56
Figure 3.5 Critical temperature versus xanthophyll zeaxanthin of high irradiance and
shaded D. antarctica during three temperature treatments .................................... 57
Figure 4.1 Weather conditions during the experiment .......................................... 73
Figure 4.2 Light saturated net photosynthesis and stomatal conductance of water deficit
and control D. antarctica and C. australis under high and moderate light in three
successive experimental periods (pre-treatment, water deficit and rewatering) .... 78
Figure 4.3. Time course of stomatal conductance of water deficit D. antarctica and C.
australis grown under high and moderate light with decreasing relative extractable soil
water and increasing number of days without water ............................................. 80

-xii-
Figure 4.4 Stable isotope composition and intrinsic water use efficiency and of the tree
ferns D. antarctica and C. australis under high and moderate light in three successive
experimental periods .............................................................................................. 84
Figure 5.1 Location of the tree ferns at the study area .......................................... 95
Figure 5.2 Relationships between photosynthetic capacity parameters and frond traits of
the tree ferns D. antarctica and C. australis and environmental variables ......... 101
Figure 5.3 Climate conditions during diurnal course measurements in summer and
winter ................................................................................................................... 104
Figure 5.4 Relationships between photosynthesis, stomatal conductance and water
pressure deficit based on leaf temperature ........................................................... 105
Figure 5.5 Light response curves of the tree ferns D. antarctica and C. australis in
summer and winter ............................................................................................... 106
Figure 5.6 Stomatal density of the tree ferns D. antarctica and C. australis from light-
exposed and shaded habitats. ............................................................................... 112

-xiii-
List of Tables
Table 2.1 Relative irradiance, Isum (i.e. the fraction of penetrating irradiance in the
photosynthetically active spectral region) of the tree ferns growing under variable light
and shade during the before exposure period, and then exposed to high light, and
measured after two weeks (short-term exposure) and three months (long-term exposure)
.................................................................................................................................... 20
Table 2.2 Photosynthetic capacity parameters of the of the tree ferns D. antarctica and
C. australis grown under variable light and shade during the before exposure period,
and then exposed to high light, and measured after two weeks (short-term exposure) and
three months (long-term exposure) ........................................................................ 23
Table 2.3 Predawn frond water potentials and frond traits of the tree ferns D. antarctica
and C. australis grown under variable light and shade during the before exposure
period, and then exposed to high light, and measured after two weeks (short-term
exposure) and three months (long-term exposure) ................................................ 27
Table 2.4 Dynamic responses of photosynthesis to an artificial sunfleck ............ 30
Table 3.1 Temperature of D. antarctica fronds exposed to high irradiance and under
shade ...................................................................................................................... 60
Table 3.2 Photosynthesis and frond traits of D. antarctica exposed to high irradiance
and under shade before and during three successive temperature treatments ....... 49
Table 3.3 Pigment content and osmolality of D. antarctica fronds exposed to high
irradiance and under shade during three successive temperature treatments ........ 54
Table 4.1 Chlorophyll fluorescence and photosynthetic capacity variables of water
deficit and control tree ferns (D. antarctica and C. australis) grown under high and
moderate light during three successive experimental periods ............................... 76
Table 4.2 Frond water relations of water deficit and control tree ferns (D. antarctica
and C. australis) grown under high and moderate light during three successive
experimental periods ............................................................................................ 100

-xiv-
Table 5.1 Significance of the effect of fixed factors (species and season) and of
covariates (ANCOVA) on photosynthetic capacity parameters and frond traits of the
tree ferns D. antarctica and C. australis .............................................................. 100
Table 5.2 Photosynthetic capacity and water relation parameters of the tree ferns D.
antarctica and C. australis in summer and winter............................................... 102

-1-
Chapter 1. Introduction
1.1. Environmental stresses: light, temperature and water deficit
Predictions of global warming and associated climate change indicate widespread in-
creases in light intensities, temperatures, and the frequency and severity of droughts in
south-eastern Australia (Hennessy et al. 2007). Understanding likely responses of plants
to future threats is critical to land management, and some of the main challenges for
conservational biology will be to anticipate environmental change and to adjust man-
agement approaches accordingly (Rossetto 2008). To achieve this aim, it is crucial to
understand the mechanisms of plants to cope with such changes in environmental fac-
tors, and to understand the limitations of their coping capacity.
1.2. Fundamental effects of high irradiance in interaction with high temperature
or drought on plants
Exposure of plants to high levels of irradiance often leads to photoinhibition and photo-
oxidative stress. Photoinhibition is a decline in the quantum yield of photosynthesis.
The primary sites of light damage are associated with components located in the thyla-
koid membranes of chloroplasts (Havaux et al. 1996). Primary damage occurs within
the reaction centre of photosystem II (PSII), with associated photoinhibition effects
such as decreases in photosynthetic yield, bulk pigment loss with photo-oxidation, loss
of enzyme activity (including Rubisco), and, eventually, even cell death (Long et al.
1994). Photo-oxidative stress is caused by the toxic effects of reactive oxygen species
(ROS) produced in the photosynthetic apparatus under high irradiance (Niyogi 2000).
Plants have developed a number of adaptive mechanisms that allow the photochemical
apparatus to cope with rapid changes in light. For instance, when leaves are exposed to
strong light that is saturating for photosynthesis, the xanthophyll zeaxanthin is rapidly
and reversibly formed by violaxanthin de-opoxidation in bright light via the intermedi-
ate antheraxanthin (e.g. Demmig-Adams and Adams 2006).

-2-
Photoinhibition alone is rarely responsible for plant mortality and the plant may recover
and become fully acclimated (Lovelock et al. 1994). However, when, in addition to high
irradiance, leaves are exposed to other environmental stress factors such as high tem-
perature or drought, there can be sustained reductions in the efficiency of photosynthetic
energy conversion and inhibition of repairs to photodamaged PSII (Murata et al. 2007).
Interactive effects of high light and increased temperature are widely discussed in plant
physiology literature (e.g. Havaux et al. 1991, Kirchgeßner et al. 2003, Dieleman and
Meinen 2007). While some authors suggest that these effects are detrimental for plants
because photosynthesis is particularly sensitive to inhibition by heat stress due to labile
components in the photosynthetic apparatus (Salvucci and Crafts-Brandner 2004), oth-
ers insist that high light alleviates negative effects of high temperatures on plants (Ha-
vaux et al. 1991).
Water supply is among the most important factors limiting plant species distribution
(Howard 1973). The primary effects of water stress on photosynthesis have been com-
prehensively discussed (e.g. Flexas et al. 1998), with stomatal conductance among the
earliest responses that protect plants from extreme water loss. Decreases in intercellular
CO2 concentrations (Ci) after stomatal closure during water stress may induce down-
regulation of photosynthetic apparatus to match available carbon substrate and de-
creased growth (Chaves et al. 2003). A number of drought effects are mediated by an
excess of absorbed light energy in the photosynthetic apparatus, leading to an imbalance
between electron transport and electron consumption and causing photoinhibition and
photo-oxidative stress (Flexas et al. 1999). Hence, an interactive effect of high light and
drought can be fatal to plants (Levitt 1980, Lovelock et al. 1994).
Effects of high light, heat and water deficit on plant performance have been extensively
studied over recent decades, yet these studies have been mainly focused on productive,
overstorey tree species (e.g. oak, Eucalyptus species, Acacia species etc). Understorey
species, with low commercial value, have received much less attention, in spite of their

-3-
specific situation, growing under relatively low irradiance, but experiencing occasional
exposure to high irradiance through sunflecks or removal of overstorey (e.g. Pearcy
1988, Durand and Goldstein 2001). Hence, results on overstorey trees cannot easily be
generalised for these species. Growing concern about biodiversity protection in produc-
tive forests and increasing commitment to sustainable forest management is contribut-
ing to rising interest in the understorey component of forests.
1.3. Two tree ferns of contrasting origin
Ferns, or pteridophytes, are the largest and most complex group of flowerless plants that
reproduce by spores developed in sporangia on the underside of leaves or fronds (Large
and Braggins 2004). Some ferns have adopted the tree growth form and are thus called
tree ferns. Most tree ferns belong to the families Dicksoniaceae and Cyatheaceae (Large
and Braggins 2004). Members of the family Cyatheaceae are the most widespread tree
ferns, with many species showing high degrees of local endemism. Centres of diversity
include the Great Antilles, Central America, the Andes, Madagascar, Malesia (i.e. in-
cluding Indonesia, Philippines and New Guinea). The family Dicksoniaceae has high
diversity in Indonesia and New Guinea, with some species found in isolated pockets in-
cluding St Helena Island and the Fernandez Islands off the coast of Chile (Large and
Braggins 2004).
The tree ferns Dicksonia antarctica Labill. and Cyathea australis (R. Br.) Domin are
iconic and ecologically important understorey plants of Australian forests. Observations
by Ashton (2000) indicated that trunks of tree ferns were favourable sites for the estab-
lishment of most woody species in wet sclerophyll forest dominated by Eucalyptus reg-
nans F. Muell. Tree ferns, particularly D. antarctica, formed an impressive understorey
and were associated with numerous species of ground and epiphytic ferns. Studies by
Lindenmayer et al. (1994) found abundance of the mountain brushtail possum Tricho-
surus caninus Ogilby increased with numbers of C. australis and D. antarctica. The
dead fronds of C. australis were favorite sites for Exoneura bicolor bees (Blows and
Schwarz 1991). Crimson Rosella (Platycercus elegans) birds value sori of D. antarctica

-4-
as an energy rich food, and sori account for 20-30% of the birds‟ diet in autumn and
winter (Magrath and Lill 1983). Both tree fern species are also popular horticulture
commodities for domestic and international markets; for example, in 2003/2004 more
than 50,000 trunks of D. antarctica were exported from Tasmania (Davies 2005).
Both D. antarctica and C. australis are widespread in the temperate zones of Australia
(McCarthy 1998). The most significant habitats for tree ferns are rainforest (cool and
warm temperate) and wet sclerophyll forests, particularly in the deepest, least disturbed
sheltered gullies (Department of Natural Resources and Environment 2002). D. antarc-
tica is common in wet forest and often dominates moist, shady gullies, where it fre-
quently grows in extensive stands. C. australis‟ s habitat ranges from dark gullies to dry
forest fringes and creek banks in quite open areas (McCarthy 1998). Observations indi-
cate that the two tree fern species have overlapping but divergent micro-site prefer-
ences. For example, a study in south-east Australian wet sclerophyll forest found that
tree ferns were more likely to be C. australis than D. antarctica with increasing distance
from a stream (Dignan and Bren 2003).
D. antarctica and C. australis belong to contrasting floristic elements of the Australian
vegetation. While D. antarctica is believed to be endemic to Australia and derived di-
rectly from the original Gondwanan flora, C. australis is considered to be an intrusive
species of the Indo-Malayan flora (Barlow 1994). These different origins combined
with indications of different micro-site preferences suggest the two tree ferns would
have different physiological adaptations to environmental stresses.
During their lifetime, tree ferns can be periodically exposed to the harsh conditions of
post-wildfire environments, which are characterised by increased light intensities and
leaf temperatures, and consequently increased evapotranspiration and water loss. Effects
of these conditions on tree fern physiology have not been studied, but are indicated by
poor survival and ongoing decline of both D. antarctica and C. australis after clearcut
logging in mountain ash (E. regnans) forest (Ough and Murphy 2004). It was found that
only 11-17 % of D. antarctica and C. australis survived one year after clearcut logging,

-5-
and of those remaining, up to 40% of D. antarctica and 65% of C. australis were not
expected to survive another five years (Ough and Murphy 2004). In contrast, much
higher rates of regeneration and survival of the tree ferns were recorded after wildfires
(Ought 2001).
1.4. The tree ferns in mountain ash forests of south-eastern Australia
Mountain ash forest of south-eastern Australia is a unique wet sclerophyll ecosystem
that typically forms an interface between two broad vegetation types, rainforest and dry
sclerophyll forest (Campbell and Clarke 2006). These forests are highly prized as water
catchments for the Melbourne region, and for flora and fauna conservation and recrea-
tion purposes (Attiwill and Fewings 2001). E. regnans, itself, is a valuable timber spe-
cies, and about 40% of these forests are available for timber harvesting (Bennett and
Adams 2004).
The dominant harvesting practice in mountain ash forests includes clearcut, slash burn-
ing of debris and remaining vegetation, and seeding with E. regnans seeds (Bennett and
Adams 2004). Such harvesting practices cause major disturbance, including physical
damage to resprouting plants, changes in soil physical and chemical properties, distur-
bance to soil stored plant propagules, and sudden exposure of understorey plants to full
sunlight (Ough and Murphy 1996).
Reasons for steady declines in tree fern numbers in post harvest environments remain
uncertain. Soil disturbance was suggested as a likely major contributor to poor regenera-
tion of tree ferns a decade after clear-felling compared with wildfire regeneration (Ough
2001). Greater survival of tree ferns was recorded in understorey islands (i.e. areas
within a coupe where trees can be felled but disturbance to understorey species and soil
is minimized) than in logged coupes, but mortality also occurred in understorey islands
across all size classes of tree ferns (Ough and Murphy 1998). Apart from soil distur-
bance, there are other obvious differences between post-wildfire and post-harvesting
environments: a fire-killed forest provides much more shade and many more micro-

-6-
habitats than the relatively uniform ash-bed created by high intensity regeneration burns
after logging (Hickey 1994). Logging also results in sharp edges in the boundary zone,
increasing light penetration up to 100% (Dignan and Bren 2003), which can create high
light stress for vegetation remaining in buffer zones, including understorey islands.
High light stress gives rise to two other stress factors – heat and drought. Heat, because
direct irradiance will also increase leaf temperatures, and drought, because greater leaf
temperatures will lead to a greater evaporative demand. As discussed above, interactive
effects of these three stresses can be fatal for a plant (Levitt 1980). Thus, it is possible
that sudden changes in light intensity, water availability and temperature contribute to
D. antarctica and C. australis mortality in the post-harvest environment.
1.5. Current knowledge of tree fern ecophysiology
Little is known about the physiology of tree ferns. It is obvious that tree ferns tolerate a
broad range of environmental conditions throughout their life cycle. Periodically dis-
turbed by wildfires, they have evolved under a regime of variable light levels from high
(immediately after fires) to low or moderate after canopy re-establishment (Hunt et al.
2002).
Certainly, other studies indicate potential for fern acclimation to different light regimes.
For example, New Zealand ferns from contrasting habitats displayed contrasting charac-
teristics in terms of photosynthetic light compensation point, which were tightly corre-
lated with specific frond area (Bannister and Wildish 1982). Frond characteristics (frond
surface area, epidermis thickness, palisade/ spongy mesophyll ratio, blade size, petiole
length) of a South American Cyathea species were also correlated with the irradiance
regime at its local micro-habitat (Arens 1997). During the course of forest ecosystem
dynamics including gap formation, bushfires, or forest harvesting, tree ferns may be
suddenly exposed to full sunlight. Studies in Hawaii indicated limited capacity of shade-
acclimated tree ferns to quickly and efficiently adjust to sudden increase in irradiance

-7-
due to gap formation (Durand and Goldstein 2001). However, further studies of high
light stress on tree ferns and their rate of recovery are currently lacking.
Effects of high temperature, either alone or with high light, on the physiological per-
formance of tree ferns have also been poorly studied. Tingey et al. (1987) found that
photosynthesis of D. antarctica was particularly susceptible to inhibition with increas-
ing temperature and high light; and Nobel et al. (1984) also mentioned negative effects
of high temperature on gas exchange of ferns. Moreover, there are indications that tree
ferns are very susceptible to temperature increases due to their reticulated vascular sys-
tem (White and Weidlich 1995), which might not be as efficient in delivering water to
fronds as the vascular system of angiosperms (Brodribb et al. 2005). However, more
detailed studies, examining acclimation potential of photosynthetic apparatus of tree
ferns to temperature increases and its reversibility are lacking, despite the obvious im-
portance of this knowledge to predicting species‟ acclimation potential and survival in
the future.
Adequate water supply as an important element for tree ferns can be suggested from
their distributional patterns in the forests (mostly along waterways), and is also men-
tioned in the horticultural literature (e.g. Jones and Clemesha 1993, Large and Braggins
2004). Observational studies indicate that tree ferns can sustain periods of drought if
they are protected by canopy. Ashton (2000) observed that despite infrequent but severe
drought events, tree fern numbers increased by 80% in the lower strata of wet sclero-
phyll forests over 48 years. Hunt et al. (2002) also reported that D. antarctica can main-
tain favourable water relations during short periods of drought if its habitat is limited to
sheltered sites. However, these field observations involve potentially confounding ef-
fects of shade, temperature and (soil and air) humidity, because more shaded sites are
also cooler and moister. Thus, it often remains unresolved whether alleviation of
drought stress is a direct effect of lower irradiance – e. g. shading ameliorates drought-
related photoinhibition and photo-oxidative stress – or an indirect effect of greater water
availability and less evaporative demand in shade.

-8-
1.6. Thesis aims and outline
The principal research objectives of this thesis are the characterisation of physiological
responses of D. antarctica and C. australis to varying light conditions, temperature re-
gimes and water availability. There have been no prior studies of the comparative
physiology of D. antarctica and C. australis, and there has been little previous examina-
tion of the interactive effects of light, temperature and water deficit on tree fern physi-
ology.
In my first experimental study (Chapter 2) I examine effects of high light on photosyn-
thetic capacity parameters of D. antarctica and C. australis in a controlled glasshouse
experiment.
In my second experimental Chapter (Chapter 3), I report effects of high light and light
by temperature interactions on photosynthetic performance of D. antarctica in a con-
trolled climate chamber experiment. This experiment was based in France, which meant
that C. australis could not be included because a European source of this species could
not be found.
In my third experimental Chapter (Chapter 4), I examine effects of water deficit either
alone or in interaction with high light on the photosynthetic capacity of D. antarctica
and C. australis in a semi-controlled, open-air experiment.
In my fourth and final experimental Chapter (Chapter 5) I examine the ecophysiology of
both tree fern species under field conditions in the buffer zones surrounding a clearcut
mountain ash forest of central Victoria, Australia. Here, I examine relationships of
growth irradiance, leaf/air temperatures, plant water status with photosynthesis, frond
traits, and water relation parameters of mature tree ferns over two consecutive years.
In my final Chapter 6, I provide an overall discussion of the results and indicate possi-
ble implications of my findings.

-9-
Each experimental chapter was written as a stand-alone paper for journal submission.
Thus, some repetitions of citations and of text from this Introductory Chapter were in-
evitable.

-10-

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-11-
Chapter 2. Effects of sudden exposure to high light on two tree fern
species Dicksonia antarctica, and Cyathea australis, acclimated to
different light intensities.
(i) Abstract
We examined the responses of two tree fern species (Dicksonia antarctica and Cyathea
australis) growing under shade or variable light (intermittent shade) to sudden exposure
to high light. Steady-state gas exchange as well as dynamic responses of plants to artifi-
cial sunflecks indicated that difference in growth light environment had very little effect
on the tree ferns‟ capacity to utilise and acclimate to prevailing light conditions. Two
weeks of exposure to high light (short-term acclimation) led to decreases in all photo-
synthetic parameters and more negative predawn frond water potentials, mostly irre-
spective of previous growth light environment. After three months in high light (long-
term acclimation), D. antarctica fully recovered while C. australis previously grown
under variable light recovered only partially, suggesting high light stress effects under
the variable light environment for this species.
2.1. Introduction
The light environment in the understorey of closed forests is often characterized as a
low level of diffuse light punctuated by intense sunflecks resulting from direct-beam
solar radiation through holes in the canopy (Pearcy 1988). However, through forest eco-
system dynamics including gap formation and bushfires, or anthropogenic management
such as forest harvesting, understorey species can suddenly be exposed to prolonged full
sunlight, a stress factor that can contribute to decline in their photosynthetic perform-
ance (Levitt 1980). Rapid physiological adjustment to unfavourable levels of irradiance
(i.e. acclimation, Lambers et al. 2008) is then required for understorey species survival.

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-12-
The tree ferns Dicksonia antarctica (Labill.) and Cyathea australis (R.Br.) Domin are
characteristic and ecologically important understorey plants of south-eastern Australia
(Large and Braggins 2004). Even though both species prefer high rainfall wet sclero-
phyll forests (Jones and Clemesha 1993), they have different micro-site preferences: D.
antarctica is common in wet, shady gullies, whereas the often co-occurring species C.
australis seems to preferentially grow within the forest or even along forest margins
(McCarthy 1998). An observational study confirmed that the greater the distance to the
stream the more likely it‟s to encounter C. australis rather than D. antarctica (Dignan
and Bren 2003), suggesting a greater dependence of D. antarctica on water availability
and shade protection. D. antarctica and C. australis belong to contrasting floristic ele-
ments of the Australian vegetation (Gondwanan vs. Intrusive, Tropical; Barlow 1994),
which suggests different physiological adaptation potential and supports their distribu-
tion patterns within forests.
Periodically disturbed by wildfires, D. antarctica and C. australis have evolved under a
regime of variable light levels from high (immediately after fires) to low or moderate
after canopy re-establishment (Hunt et al. 2002). Evidence of plasticity in frond mor-
phology and anatomy in response to different levels of irradiance was found in a study
on South American Cyathea species (Arens 1997). Hunt et al. (2002) also suggested
that during the period of regeneration of woody species following fire, D. antarctica
may experience prolonged periods of exposure to high light intensities and dry atmos-
pheric conditions. Potential of D. antarctica and C. australis to tolerate a broad range of
light conditions is also indicated in horticultural publications (Jones and Clemesha
1993; Large and Braggins 2004).
Despite their apparent longer term acclimation potential to variable light conditions, tree
ferns seem particularly vulnerable after the formation of large gaps. Studies in Hawaii
found that shade-adapted tree ferns were damaged in disturbed areas and forest gaps,
because they are unable to adjust quickly or efficiently to high light environments (Du-
rand and Goldstein 2001). Moreover, Ough and Murphy (2004) found that only about
11 – 17 % of D. antarctica and C. australis survived one year after clearcut logging in

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-13-
the mountain ash (Eucalyptus regnans F. Muell) forests of the Victorian Central High-
lands. Of those remaining, up to 40 % of D. antarctica and 65 % of C. australis would
not survive another five years (Ough and Murphy 2004).
The objectives of this study were to elucidate acclimation potential and vulnerability of
D. antarctica and C. australis to sudden increases in light. The two species were grown
under either full or intermittent shade („variable light‟) in a glasshouse and, after pro-
longed acclimation, were suddenly exposed to high irradiance. We measured steady-
state and dynamic (i.e. sunflecks) photosynthetic responses of the tree ferns acclimated
to each light environment in order to test the following hypotheses:
Responses to light would be different between the two species, with D. antarctica
performing better in full shade and being more prone to high light-induced dam-
age;
Sudden exposure to high light would cause limitations in gas exchange (e.g.
photoinhibition) in both species in the short term (two weeks), with those accli-
mated to full shade most strongly affected;
Both tree fern species would have limited capacity to acclimate to high light even
in the longer term (three months).
2.2. Materials and methods
2.2.1. Plant material
Ten sporophytes of D. antarctica and ten of C. australis (Fern Acres nursery, King
Lake West, Australia) were transplanted into 25-l pots. Potting mix contained (% vol-
ume) composted pine bark (30), gravel (45), coarse fern mulch (5), composted mulch
(14.5), fine fern mulch (5), „Dynamic lifter‟ (0.16: Yates, Padstow, NSW, Australia),
and two types of slow-release fertiliser (0.17 each; Osmocote, Baulkham Hills, NSW,
Australia). Before the experiment, tree ferns were propagated from spores and grown in
an open-air nursery under a dense canopy that provided ca 70% shade. All plants were
about six months old and 20-25 cm tall at the start of the experiment.

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-14-
2.2.2. Experimental design
The experiment ran from December 2005 to July 2006 in a glasshouse at the site of the
University of Melbourne‟s Creswick campus, in south-eastern Australia (143º53‟E,
37º25‟S; elevation 392 m above sea level). After the first two weeks, old and fully de-
veloped fronds were cut off and only new fronds, which developed under a designated
growth light environment, were measured during the experiment.
Plants were randomly assigned to two growth light environments in a fully randomised
block design with five replicates (i.e. two species within two treatments per each of five
blocks). The two growth light environments – „shade‟ and „variable light‟ – were ap-
plied using wavelength neutral shadecloth. Shade allowed ca 20% uniform light pene-
tration; whereas the variable light simulated sunflecks – the shadecloth was cut into 12
cm-wide stripes, and these were alternated with uncovered gaps of the same width (the
12 cm width was based on the 20 min movement of the sun at its zenith). Under direct
sun in the glasshouse, the maximum recorded photosynthetic photon flux density
(PPFD) was 1900 µmol photons m-2
s-1
at plant height (PAR range, 400-700 nm, meas-
ured with a Li-Cor quantum sensor).
Plants were watered twice per day to maintain soils at field capacity throughout the ex-
periment. Relative humidity and air temperature in the glasshouse were maintained us-
ing a Humidex I greenhouse climate control system (Nelan Industries Pty. Ltd., Mel-
bourne, Australia). Mean conditions throughout the experiment were: 8ºC minimum
temperature, 24ºC maximum temperature, and > 60 % relative humidity.
Plants were measured at the end of three periods:
1) „Before exposure‟ (early April 2006): measurement of new fully-developed
fronds after four months of growth under the designated light environment
(shade or variable light);

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-15-
2) „Short-term exposure‟ (late April 2006): measurements of the same cohort of
fronds after two weeks of shade removal (indication of short-term acclimation
potential);
3) „Long-term exposure‟ (late July 2006): measurements of a new fully developed
cohort of fronds after three months of shade removal (indication of long-term
acclimation potential).
In addition, an artificial sunfleck experiment was conducted for two days in late March
2006 (i.e. before exposure and just after full expansion of new fronds). See below for
details.
Chlorophyll a fluorescence, predawn water potential, and gas exchange parameters
were measured at the end of each of the three experimental periods. Samples for nitro-
gen and chlorophyll were collected at the same time. All measurements were made on
the mid-third of the youngest fully expanded fronds.
2.2.3. Light environment of measured frond
The growth light environment for each plant was calculated from hemispherical photo-
graphs. These were taken at the level of each measured frond using a fish-eye lens
(Nikon, F- 601, Japan). Black and white negatives were scanned and evaluated using
Winphot software (ter Steege 1996). Relative irradiance at the measurement location
(Isum) was calculated according to Niinemets et al. (1998):
Isum = pdif Idif + (1- pdif) Idir Eqn. (1)
Where pdif is the ratio of diffuse irradiance to total irradiance in the photosynthetically
active spectral region (400-700nm) above the plant; and Idif and Idir are the factors of
diffuse and direct radiation that will penetrate to the measured location relative to the

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-16-
total irradiance above the plant (ter Steege 1996). All parameters were calculated for
each day of the experiment, taking into account amount of sunshine hours in Victoria
during each month of the experiment (data from the Australian Bureau of Meteorology,
http://www.bom.gov.au/jsp/ncc/climate _averages/sunshine-hours/index.jsp, verified 1
September 2009).
2.2.4. Maximal quantum yield of photochemistry (Fv/Fm)
Predawn chlorophyll a fluorescence was measured on overnight dark-adapted leaves
with a pulse modulated fluorometer (OS-30p, Opti-Sciences, Hudson, USA). Ground
fluorescence (Fo) was obtained with a low intensity modulated light (600 Hz, 650nm,
photosynthetic photon flux density PPFD <1 µmol m-2
s-1
). Maximum fluorescence (Fm)
was induced by a saturating flash. Maximum efficiency of PSII was estimated as Fv/Fm
= (Fm - Fo)/Fm, after Maxwell and Johnson (2000).
2.2.5. Gas exchange measurements
Gas exchange parameters were measured using a Li-Cor 6400 gas exchange system,
equipped with a 2x3 cm broadleaf chamber (Li-Cor, Lincoln, Nebraska, USA).
A light response curve was generated for each plant at CO2 concentration of 400 µmol
mol-1
, block temperature 25ºC, air flow rate 400 µmol air s-1
, and relative humidity
>60%. PPFD was increased stepwise from 0 to 2000 µmol m-2
s-1
. Fronds were induced
in the dark for approximately 10 min and the rate of dark respiration was recorded when
stability was reached. PPFD was then increased in 11 successive steps to 2000 µmol m-
2s
-1 with two measurements per PPFD level. Measurements were recorded once rates of
gas exchange were stable. Apparent quantum yield (ф) and maximum photosynthetic
rate Amax were calculated according to Lambers et al. (2008).
An A-Ci curve was generated according to Long and Bernacchi (2003) with some modi-
fications: PPFD 1000 µmol m-2
s-1
, block temperature 25ºC, air flow rate 400 µmol air s-

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-17-
1, and relative humidity > 60%. Reference CO2 concentration was increased from 75 to
2200 µmol mol-1
in 13 successive steps with two measurements per CO2 concentration.
Measurements were recorded once gas exchange parameters were stabilised, which on
average took at least 5 min. After finishing the A-Ci curve, illumination in the leaf
chamber was turned off, CO2 concentration was decreased to ambient and respiration
due to oxidative phosphorylation was recorded after 5 min in the dark.
Using the Farquhar model (Farquhar et al. 1980), maximum carboxylation rate (Vcmax),
and maximum electron transport rate (Jmax) were evaluated by fitting A- Ci curves to
non-rectangular hyperbolas (as described in Dreyer et al. 2001 and Montpied et al.
2009). Triose phosphate use (TPU) limitation was not included in the model, and corre-
sponding points with decreased Amax at elevated Ci were disregarded (Long and Bernac-
chi 2003). The set of primary parameters of Rubisco kinetic properties used here
(Kc=327µmol mol-1
, Ko=282600 µmol mol-1
, Γ*=43.7 µmol mol-1
) are from von
Caemmerer et al. (1994).
The frond area enclosed in the chamber for light response and A- Ci curves was marked,
detached, scanned and calculated using imaging software (UTHSCSA Image Tool Ver-
sion 3, University of Texas, USA). All gas exchange measurements were recalculated
on a frond-area basis.
2.2.6. Plant water status
Predawn frond water potential (Ψpredawn) of each plant was measured at the end of each
period using a pressure chamber (PMS Corvallis, OR, USA).
2.2.7. Frond traits
Specific leaf area (SLA), needed for calculation of nitrogen and chlorophylls on a
frond-area basis, was calculated as the ratio of frond area over frond dry weight (m2 kg
-1
dry weight). Fresh frond samples were collected, the frond area scanned and calculated

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-18-
using Scion Image software (Scion Corporation 2000-2001, USA), and frond material
then dried at 60ºC for 48 h for dry weight.
Frond samples were analysed for total nitrogen and carbon content using an elemental
analyser (LECO CHN-1000, Michigan, USA). Frond samples were dried as described
above and ground to a fine powder. Photosynthetic nitrogen use efficiency (PNUE) was
calculated as Amax divided by frond nitrogen content (on a frond-area basis).
For measurements of frond chlorophyll content, four frond discs (each diameter 3.75
mm) were collected, immediately immersed in liquid nitrogen, and stored at -80ºC until
extraction. Chlorophyll a and b were extracted using 1.8 ml of 100% dimethyl sulphox-
ide (DMSO). Extracts were heated for 30 min at 65ºC in a dry block heater Termoline
L+M (Northgate, Queensland, Australia). The supernatant was then transferred to a
spectrophotometer Carry 300 (Varian, The Netherlands). A blank of pure DMSO was
used to calibrate the spectrophotometer at zero absorbance. Chlorophyll a, b and total
concentrations were calculated according to Wellburn (1994).
2.2.8. Artificial sunfleck experiment
Predawn Fv/Fm was recorded for each dark-adapted plant. A frond was then enclosed in
the Li-Cor chamber at PAR 20 µmol m-2
s-1
and photosynthesis rate (A20) and gs20 were
recorded once the readings were stable (after at least 5 min). PPFD was then increased
to 2000 µmol m-2
s-1
in one step, and gas exchange parameters recorded every 10 sec-
onds for 20 min. Fv/Fm immediately and 30 min after the sunfleck were recorded using a
pulse modulated fluorometer (as above). Experimental conditions were: chamber rela-
tive humidity 75-80%, reference CO2 concentration 400 µmol m-2
s-1
and block tempera-
ture 25±1ºC.
The following parameters were calculated to characterise the dynamic response of net
photosynthesis to a sudden increase in PAR from 20 to 2000 µmol m-2
s-1
: maximal pho-

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-19-
tosynthesis rate (Amax_ind, calculated from the induction curve) and time to reach 63% of
change in photosynthesis (t63%), nomenclature after Tausz et al. (2005).
2.2.9. Statistical analyses
Repeated-measures models of SPSS 15 (SPSS Inc. Chicago, USA) were used for statis-
tical analyses, with light and species as the between-subject factors and period as the
within-subject factor (all fixed). Effects of period (before exposure; short-term expo-
sure; long-term exposure), growth light environment (variable light; shade) and species
(D. antarctica; C. australis), and period by light by species interactions on each de-
pendent variable were tested. Data for statistical analyses were the values per individual
plant at the end of each period. Significant differences between periods were examined
by using the repeated contrast function (SPSS 15).
A two-way general linear model (SPSS 15) with growth light environment and species
as fixed factors was used to analyse the artificial sunfleck data.
Each dependent variable was checked for normality using the Shapiro-Wilk test and log
transformed if assumptions of normality were not satisfied. Data were checked for ho-
mogeneity of variance using Cochrane‟s test, and it was ensured by visual examination
of scatter plots that means and variances were not correlated across experimental
groups.
2.3. Results
2.3.1. Light environment of the tree ferns
Relative irradiance, Isum, did not differ between species within growth light environment
(P=0.4, data not shown), confirming randomised block design for the two species. Yet,
Isum in the variable light was 2.5 times greater than Isum of shade (Table 2.1). Shade re-
moval increased growth light intensity almost two-fold for variable light plants, and

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-20-
more than four-fold for shaded plants (Table 2.1). These relatively high levels of irradi-
ance remained until the end of the experiment.
Table 2.1. Relative irradiance, Isum (i.e. the fraction of penetrating irradiance in the pho-
tosynthetically active spectral region) of the tree ferns growing under variable light and
shade during the before exposure period, and then exposed to high light, and measured
after two weeks (short-term exposure) and three months (long-term exposure). Values
are means (n = 5) ± s.e. Affect abbreviations: P, Period; L, Growth light environment.
Differences in Isum between growth light environments for each period were determined
using one-way ANOVAs‟. Significance level:***P<0.001.
Isum Growth light environment Effects
Variable light Shade L S
Before exposure 0.344±0.02 0.134±0.01 *** n.s. (0.4)
Short-term exposure 0.656±0.03 0.575±0.04 n.s. (0.2) n.s. (0.1)
Long-term exposure 0.617±0.04 0.676±0.05 n.s. (0.9) n.s. (0.4)
Maximum quantum yield of photochemistry (Fv/Fm)
Maximum quantum yield of photochemistry (Fv/Fm) was similar between species and
tended to be lower under variable light than shade in the before exposure period (mean
across species of 0.76 versus 0.82; Table 2.2). Short-term (two week) exposure to high
light led to significant decreases in Fv/Fm of both species irrespective of the growth light
environment (to ca 0.70; Table 2.2). After three months of exposure to high light, Fv/Fm
partially recovered in previously shaded plants but remained low in plants previously
grown under variable light (Period x Light, P<0.001, Table 2.2).
2.3.2. Photosynthetic capacity parameters
Light-saturated rate of net photosynthesis (Amax) and stomatal conductance (gs) at Amax
were similar across species and growth light environments in the before exposure period

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-21-
(Figs. 2.1 a, 2.1 b). Both parameters decreased after two weeks of exposure to high light
in both species irrespective of the growth light environment, and both parameters recov-
ered after three months of high light exposure, with the exception of Amax in C. aus-
tralis, which remained low (Figs. 2.1 a, 2.1 b).
Respiration rate in the dark (Rd) was not significantly affected by the growth light envi-
ronment, species or sudden exposure to high light (Fig. 2.1 c). However, three months
after exposure, Rd increased significantly in C. australis previously grown under vari-
able light (Fig. 2.1 c).
Apparent maximum quantum yield (ф) was significantly greater in shade than in vari-
able light plants in the before exposure period, then decreased in previously shaded
plants but increased in variable light plants after two weeks of exposure to high light
(Table 2.2). After three months of exposure to high light, ф increased to near or greater
than the before exposure values in all but previously shade-grown C. australis plants
(Table 2.2). Effect of species on ф was insignificant in all periods.
Light compensation point (LCP) was similar across species and growth light environ-
ments in the before exposure period (Table 2.2). Two weeks of exposure to high light
led to significant increases of LCP in both species with greater increased in previously
shaded than in variable light plants. LCP continued to increase after three months of ex-
posure to high light in all but previously shade-grown D. antarctica (Table 2.2).
The maximal carboxylation rate (Vcmax) as well as the maximal light driven electron flux
(Jmax) did not differ between growth light environments and species in the before expo-
sure period (Figs. 2.2 a, 2.2 b). Two weeks of exposure to high light led to significant
decreases in Vcmax and Jmax, irrespective of species and growth light environments. Vcmax
remained low even after three months of exposure in all plants and only marginally re-
covered in D. antarctica previously grown in variable light (P=0.06). In contrast, recov-
ery of Jmax was observed in all plants with exception for C. australis previously grown
in variable light (Fig. 2.2 b).

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-22-
Photosynthetic nitrogen use efficiency (PNUE) was not affected by the growth light en-
vironment but was significantly higher in C. australis in the before exposure period
(Table 2.2). Short-term (two week) exposure to high light did not affect PNUE of vari-
able light plants contrasting with a decrease in shade-grown plants. PNUE significantly
increased after three months of exposure in all plants, with greater rises in plants previ-
ously grown in variable light (Light x Period, P=0.02; Table 2.2).

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyat-
heaceae) acclimated to different light intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-23-
Table 2.2. Photosynthetic capacity parameters of the of the tree ferns D. antarctica and C. australis grown under variable light and shade during
the before exposure period, and then exposed to high light, and measured after two weeks (short-term exposure) and three months (long-term
exposure).
Values are means (n = 5) ± s.e. of: Fv/Fm, maximal quantum yield of photochemistry; ф, the apparent maximum quantum yield; LCP, light com-
pensation point; PNUE, photosynthetic nitrogen use efficiency (Amax/nitrogen content). Effect abbreviations: P, Period; L, Growth light envi-
ronment; S, species. Significance levels:*P≤0.05; **P<0.01; ***P<0.001; x, interactions. Only significant effects and interactions are presented.
Parameters Species Growth light environment
Effects Variable light Shade
Before ex-
posure
Short-term
exposure
Long-term
exposure
Before ex-
posure
Short-term
exposure
Long-term ex-
posure
Fv/Fm D. antarctica 0.78±0.02 0.72±0.03 0.69±0.02 0.83±0.01 0.68±0.02 0.71±0.02 P ***
L x P ** C. australis 0.75±0.02 0.68±0.03 0.68±0.01 0.82±0.01 0.67±0.02 0.73±0.01
ф (mol CO2
mol-1
quanta)
D. antarctica 0.068±0.005 0.079±0.009 0.096±0.031 0.085±0.008 0.059±0.008 0.047±0.002 L*; L x P *
C. australis 0.069±0.009 0.112±0.028 0.106±0.008 0.081±0.012 0.065±0.013 0.080±0.012
LCP (µmol m-
2s
-1)
D. antarctica 9±0 12±3 15±4 12±2 26±6 14±3 P***
C. australis 11±2 12±1 17±3 7±2 15±3 19±5
PNUE (µmol
CO2 molN-1
s-
1)
D. antarctica 25±4 30±1 59±11 33±3 25±3 48±4 P ***; S***
L x P *
L x S *
C. australis 32±5 33±3 70±5 66±13 23±4 56±8

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-24-
Am
ax (
mol C
O2 m
-2s-1
)
0
2
4
6
8
Shadegs a
t A
max (
mm
ol H
2O
m-2
s-1
)
0
20
40
60
80
100
120
140
Variable light
Before_exposure
Short_term
Long_term
Rd (
mol C
O2 m
-2s-1
)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Period
Before_exposure
Short_term
Long_term
P-value of effects
P ***
P-value of effects
P ***P x L x S *
P-value of effects
P x L x S*
a
b
c
Fig. 2.1. a) Stomatal conductance under saturating irradiance; b) maximum photosyn-
thetic rate and c) mitochondrial respiration in the dark of the tree ferns D. antarctica
( ) and C. australis ( ) grown under variable light and shade during the before
exposure period, and then exposed to high light, and measured after two weeks (short-
term exposure) and three months (long-term exposure). Values are means (n=5) ± s.e. Ef-
fect abbreviations: S, Species; L, Growth light environment; P, Period. Significance lev-
els:*P≤0.05; ***P<0.001; x, interactions. Only significant effects and interactions are
presented.

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-25-
Variable light
Before_exposure
Short_term
Long_term
ShadeV
cm
ax (
mo
l C
O2 m
-2s-1
)
0
10
20
30
40
Period
Before_exposure
Short_term
Long_term
Jm
ax (
mo
l C
O2 m
-2s-1
)
0
20
40
60
80
100
P-value of effects
P*
P-value of effects
P**S**P x S *
a
b
Fig. 2.2. a) Maximal carboxylation rate of Rubisco and b) maximal light driven electron
flux of the tree ferns D. antarctica ( ) and C. australis ( ) grown under vari-
able light and shade during the before exposure period, and then exposed to high light,
and measured after two weeks (short-term exposure) and tree months (long-term expo-
sure). Values are means (n=5) ± s.e. Effect abbreviations: S, Species; P, Period. Signifi-
cance levels:*P≤0.05; **P<0.01; x, interactions. Only significant effects and interactions
are presented.

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-26-
2.3.3. Plant water status
Predawn frond water potentials (Ψ predawn) were similar across growth light environ-
ments and species in the before exposure period (-0.3 to -0.4 MPa; Table 2.3). Two
weeks of exposure to high light led to small although significant decreases in Ψ predawn in
both species and both growth light environments (-0.6 to -1.1 MPa), with full recovery
after three months in high light (Table 2.3).
2.3.4. Frond traits
Nitrogen (NA) and carbon (CA) content per frond area were significantly greater in
plants grown under variable light than in shade, and in D. antarctica than in C. australis
(Table 2.3). Two weeks of exposure to high light stimulated increases in CA in shade-
grown plants and decreases in NA in variable light plants of both species. After three
months in high light, NA remained relatively unchanged in shade-grown plants, but de-
creased significantly in plants previously grown in variable light. In contras, CA was
comparable with pre-exposure levels in variable light plants while continued to increase
in previously shade-grown plants (Table 2.3).
Total chlorophyll (a + b) and chlorophyll a content per frond area were significantly
greater in shade-grown than in variable light plants, and in D. antarctica than in C. aus-
tralis (Table 2.3). Sudden exposure to high light had no immediate effect on total chlo-
rophyll, which decreased only after three months of high light exposure (Table 2.3).
Chlorophyll a/b ratio was significantly greater in D. antarctica for much of the experi-
ment, whereas growth light environment and short-term (two week) exposure to high
light had no effect on chlorophyll a/b ratio (Table 2.3).

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species Dicksonia antarctica (Dicksoniaceae) and Cyathea australis
(Cyatheaceae) acclimated to different light intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-27-
Table 2.3. Predawn frond water potentials and frond traits of the tree ferns D. antarctica and C. australis grown under variable light and
shade during the before exposure period, and then exposed to high light, and measured after two weeks (short-term exposure) and three
months (long-term exposure). Values are means (n = 5) ± s.e. of: Ψ predawn, predawn frond water potential; NA, nitrogen content on a frond
area basis; CA, carbon content on a frond area basis; Chl total, total chlorophyll content on a frond area basis; Chl a, chlorophyll a content
on a frond area basis; Chl a/b, chlorophyll a/b ratio. Effect abbreviations: P, Period; L, Growth light environment; S, Species. Significance
levels:*P≤0.05; **P<0.01; ***P<0.001; n.s., non significant; x, interactions. Only significant interactions are presented.
Parameters Species Variable light Shade Effects
Before
exposure
Short-term
exposure
Long-term
exposure
Before
exposure
Short-term
exposure
Long-term
exposure
Ψ predawn
(MPa)
D. antarctica -0.4±0.1 -1.1±0.2 -0.5±0.1 -0.3±0.1 -0.9±0.2 -0.4±0.0 P ***
C. australis -0.3±0.1 -0.7±0.1 -0.5±0.1 -0.4±0.1 -0.6±0.1 -0.3±0.2
NA (g m-2
) D. antarctica 4.0±0.3 3.2±0.1 2.0±0.3 2.7±0.1 2.6±0.1 2.1±0.2 P ***; L ***; S ***;
L x P ** C. australis 3.4±0.5 1.9±0.2 1.5±0.1 1.8±0.2 1.8±0.1 1.5±0.1
CA (g m-2
) D. antarctica 74±5 71±4 72±9 48±5 58±5 75±5 P *; L ***; S *
C. australis 62±8 64±6 69±1 39±5 45±5 57±6
Chl total
(µmol m-2
)
D. antarctica 592±71 581±80 465±83 605±65 740±86 493±34 P *; L *; S **
C. australis 433±78 352±52 367±66 567±65 554±103 427±60

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species Dicksonia antarctica (Dicksoniaceae) and Cyathea australis
(Cyatheaceae) acclimated to different light intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-28-
Parameters Species Variable light Shade Effects
Before
exposure
Short-term
exposure
Long-term
exposure
Before
exposure
Short-term
exposure
Long-term
exposure
Chl a (µmol
m-2
)
D. antarctica 454±51 442±64 337±63 454±46 538±60 394±24 P*; L*; S***
C. australis 321±57 264±40 267±55 422±48 400±72 312±45
Chl a/b D. antarctica 3.3±0.2 3.4±0.2 1.3±0.2 3.1±0.1 2.6±0.1 1.5±0.1 S***; L x P *
C. australis 2.6±0.3 3.0±0.3 2.4±0.2 2.9±0.1 2.7±0.2 2.7±0.3

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-29-
2.3.5. Artificial sunfleck experiment
Predawn Fv/Fm was significantly lower in variable light than shade-grown plants (never-
theless near 0.80), with the lowest Fv/Fm recorded for C. australis (0.78). Immediately
after a sunfleck, Fv/Fm decreased to near 0.50 for both species irrespective of the growth
light environment, but partly recovered within 30 min, although recovery of variable
light C. australis was least (0.70; Table 2.4). Photosynthesis rate in low light (A20) as
well as maximal photosynthesis rate calculated from the induction curve Amax_ind did not
differ between species and growth light environments (Table 2.4). Increase in stomatal
conductance (∆gs) in response to a sunfleck was insignificant for both species irrespec-
tive of the growth light environment (Table 2.4). Time to reach 63% of change in pho-
tosynthesis (t63%) did not differ significantly between species and growth light environ-
ments (Table 2.4).

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-30-
Table 2.4. Dynamic responses of photosynthesis to an artificial sunfleck. Values are
means (n = 5) ± s.e. of: Fv/Fm _pr, Fv/Fm predawn; Fv/Fm _ immed, Fv/Fm immediately after
the sunfleck; Fv/Fm _30 min, Fv/Fm 30 min after sunfleck; A20, maximal photosynthesis
rate at PAR 20 µmol m-2
s-1
; Amax _ind, maximal photosynthesis rate calculated from the
induction curve; ∆ gs (gsmax–gs20), difference between stomatal conductance at the end of
the experiment (gsmax) and at PAR 20 µmol m-2
s-1
(gs20); t 63%, time to reach 63% of
change in photosynthesis. Effect abbreviations: S, Species; L, Growth light environ-
ment. Significance levels:*P≤0.05; n.s.,-non significant. Only significant interactions
are presented.
Parameter Species Light regime P-value of effect
Variable Shade
Fv/Fm _pr D. antarctica 0.81±0.01 0.81±0.01 L *, L x S *
C. australis 0.78±0.01 0.82±0.00
Fv/Fm _ immed D. antarctica 0.53±0.04 0.49±0.03 n.s.
C. australis 0.53±0.04 0.53±0.03
Fv/Fm _30 min D. antarctica 0.76±0.01 0.71±0.02 L x S *
C. australis 0.70±0.02 0.72±0.01
A20
(µmol m-2
s-1
)
D. antarctica 0.51±0.12 0.15±0.18 n.s.
C. australis 0.42±0.11 0.54±0.13
Amax _ind
(µmol m-2
s-1
)
D. antarctica 6.25±0.29 3.94±0.68 n.s.
C. australis 5.46±0.89 5.86±0.59
∆ gs
(mmol m-2
s-1
)
D. antarctica 26±7 51±13 n.s.
C. australis 35±16 29±26
t 63% (s) D. antarctica 27±4 82±25 n.s.
C. australis 49±18 22±4

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-31-
2.4. Discussion
2.4.1. Species overview
D. antarctica and C. australis had comparable maximum photosynthetic rates, Amax and
mesophyll capacity parameters (i.e. Vcmax and Jmax). Photosynthetic rates were within the
range previously reported in the literature for D. antarctica (Amax of 6 – 11 µmol m-2
s-1
(Hunt et al. 2002; Volkova et. al 2009), while Vcmax and Jmax were within the lowest range
of values, measured under similar light intensities and leaf temperatures, among a large
number of species reviewed by Wullschleger (1993).
N content per frond area was significantly greater in D. antarctica than in C. australis.
Values of up to 4 g m-2
in D. antarctica are beyond the range previously recorded for tree
fern species (e.g. Cibotium menziesii (Hook) max 2.1 g m-2
, Durand and Goldstein 2001).
D. antarctica also had significantly greater chlorophyll a/b ratio, due to its greater chloro-
phyll a content, which is a characteristic of sun acclimated plants (Arens 1997). This re-
sult was at odds with this species‟ putative preference for well-shaded microhabitats
(Dignan and Bren 2003). However, potential confounding effects of tree fern age should
be considered given that our study involved young sporophytes rather than field-grown
adults (Herbinger et al. 2007). Although D. antarctica seems to show attributes of a more
light-adapted plant at the leaf scale, differences at the whole-tree scale, such as less hy-
draulic conductance (typical for trees with small rooting volumes), may contribute to re-
stricting this species to wetter and shadier sites (see, for instance, McDowell et al. 2008).
In addition, results of a field study on mature tree ferns indicated more similar values in
chlorophyll a/b ratios between the two species (Volkova et al, in preparation).
2.4.2. Acclimation to growth light environment
We observed plasticity of frond traits in response to the growth light environment in both
species (except for frond shape, which did not differ between the two growth light envi-
ronments). More carbon per frond area, CA, in plants grown under variable light, indicated
greater carbon gain under increased light availability (Oikawa et al. 2006). Shade-grown
plants had more total chlorophyll (a + b) per frond area, indicating an enhanced invest-

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-32-
ment of resources to improve light harvesting in low irradiance (Niinemets et al. 1998).
Although shade leaves commonly have lower chlorophyll a/b ratio, the growth light envi-
ronment did not affect chlorophyll a/b ratio in our tree ferns, possibly because plasticity
of this parameter is limited in ferns. Due to a lack of data on acclimation of chlorophyll
a/b ratio in other tree fern species, we compared our tree ferns with shade adapted
Trichomanes ferns, where the chlorophyll a/b ratio did not change after plants were trans-
ferred to high light (Johnson et al. 2000). We believe that such a comparison with non-
arboreal ferns is appropriate as according to Large and Braggins (2004) „tree fern‟ is an
arbitrary term, applied to any ferns with large erect rhizomes.
In terms of photosynthetic capacity parameters both species displayed limited acclimation
potential to the growth light environment (i.e. effect of light was significant only for the
maximum apparent quantum yield, ф), especially given that all measurements were made
on fronds that developed under those environments. Greater ф of shade-acclimated plants
probably allowed plants to photosynthesise more efficiently under low light (Seidlova et
al. 2009).
Acclimation of plants to variable light did not lead to more efficient use of sunflecks –
none of the measured parameters during sunflecks differed significantly from shaded
plants. These results are in agreement with findings for the fern Polypodium virginianum
L. by Gildner and Larson (1992) and confirm our suggestion of low acclimation potential
of the tree fern photosynthetic apparatus to changing light conditions.
Significant decreases in Fv/Fm after the sunfleck treatment indicated engagement of non-
photochemical quenching, which was in agreement with other studies (e.g. Watling et al.
1997). There was no significant stomatal response to the artificial sunfleck, suggesting
that stomata were not controlling dynamic photosynthetic response in the tree ferns, as
was previously found in a co-occurring tree species Nothofagus cunninghamii Oerst.
(Tausz et al. 2005).

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-33-
2.4.3. High light stress
Irrespective of previous growth light environment, sudden exposure to high light resulted
in significant decreases in a number of parameters, namely Fv/Fm, Amax, gs, Vcmax, Jmax and
Ψpredawn in both species. Decreases in photosynthetic capacity parameters (i.e. Amax, Vcmax,
Jmax) are consistent with effects of high light stress (Larcher 2003). The apparent quantum
yield of photosynthesis, ф, was the only parameter more strongly affected in shade-
acclimated plants than in those grown under variable light, probably indicating that excess
irradiance reduced the quantum yield of photosynthesis via inactivation or down-
regulation of PSII (Montgomery et al. 2008) – greater decrease in Fv/Fm in shade-grown
plants is in agreement with this statement.
Significant decreases in Ψ predawn were not associated with changes in water availability as
plants were watered to field capacity of the potting mix at all times and may point towards
the effect of increased transpiration due to increase in leaf temperature trigged by sudden
exposure to high light (Levitt 1980). Decreases in Ψ predawn in our plants (to -1.1 MPa)
possibly indicated hydraulic stress. Ferns are known to have dichotomous branching veins
which are not very efficient in delivering water to fronds. Furthermore, vein density is
very low in these species (Brodribb et al. 2007).
2.4.4. Acclimation to new light environment
Non-recovery of Fv/Fm after three months of exposure indicated mild, albeit significant,
photoinhibition in all plants, consistent with findings of Guo et al. (2006), who observed
non-recovery in Fv/Fm of tropical rainforest sub-canopy species after transfer to high light.
However, mild photoinhibition would have very little effect on Amax measured under satu-
rated light (Zhu et al. 2004) and therefore would not contribute markedly to longer-term
persistence of the tree ferns under high light conditions.
Photosynthesis (Amax) of D. antarctica but not C. australis recovered after three months
exposure to high light. This result is consistent with field observations by Ough and Mur-

This Chapter is published
Volkova L, Bennett LT and Tausz M “Effects of sudden exposure to high light on two tree fern species
Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) acclimated to different light
intensities, Australian Journal of Botany, v.57, issue 7, 2009 In press
-34-
phy (2004) of greater decline in C. australis than D. antarctica within five years of clear-
cut logging.
While Vcmax remained low even after three months of exposure to high light in all plants,
full recovery (and even further increase of Jmax in D. antarctica) suggest that electron
transport and RuBP regeneration capacity, rather than Rubisco activity, were limiting fac-
tors to tree fern photosynthesis, as suggested by Wise et al. (2004).
Increases in Rd with prolonged exposure to high light of C. australis pre-acclimated to
variable light suggests greater energy loss by this species (Seidlova et al. 2009) and may
contribute to the lack of recovery of Amax. These results are consistent with our earlier
suggestion and findings by Ough and Murphy (2004) that C. australis appeared to be
more vulnerable to high light stress.
2.5. Summary
Regardless of putative differences in origin and observed differences in micro-site prefer-
ences, both tree fern species had comparable photosynthetic capacity parameters. Steady-
state gas exchange as well as dynamic responses of plants to an artificial sunfleck indi-
cated that difference in the growth light environment had very little effect on the tree
ferns‟ capacity to utilise and acclimate to prevailing light conditions. Sudden exposure to
high light led to decreases in all photosynthetic parameters and more negative predawn
frond water potentials, indicating high light stress responses that were mostly irrespective
of the growth light environment. Despite its field preference for shadier and wetter sites,
D. antarctica showed greater acclimation capacity to sustained high light stress than C.
australis under glasshouse conditions. After three months in high light, D. antarctica fully
recovered while C. australis previously grown under variable light recovered only par-
tially, indicating limited capacity of these plants to acclimate to high light, or perhaps
suggesting some previous light stress under the variable light environment. Results of this
study can be used in planning of forest management practises for better protection of bio-
diversity.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-35-
Chapter 3. Interactive effects of high irradiance and moderate heat on
photosynthesis, pigments, and tocopherol in the tree fern Dicksonia
antarctica.
(ii) Abstract
Effects of high irradiance and moderate heat on photosynthesis of the tree fern Dick-
sonia antarctica were examined in a climate chamber under two contrasting irradiance
regimes (900 and 170 µmol photons m-2
s-1
) and three sequential temperature treatments
(15ºC; 35ºC; back to 15ºC). High irradiance led to decline in predawn quantum yield of
photochemistry, Fv/Fm (0.73), maximal Rubisco activity (Vcmax; from 37 to 29 µmol m-2
s-1
), and electron transport capacity (Jmax; from 115 to 67 µmol m-2
s-1
). Temperature
increase to 35ºC resulted in further decreases in Fv/Fm (0.45) and in chlorophyll bleach-
ing of high irradiance plants, while Vcmax and Jmax were not affected. Critical tempera-
ture for thylakoid stability (Tc) of D. antarctica was comparable with other higher plants
(ca 47ºC), and increases of Tc with air temperature were greater in high irradiance
plants. Increased Tc was not associated with accumulation of osmotica or zeaxanthin
formation. High irradiance increased the xanthophyll cycle pigment pool (V+A+Z, 91
vs. 48 mmol mol-1
chlorophyll-1
), de-epoxidation state (56% vs. 4%), and α-tocopherol.
Temperature increase to 35ºC had no effect on V+A+Z and de-epoxidation state in both
light regimes, while lutein, β-carotene and α-tocopherols increased, potentially contrib-
uting to increased membrane stability under high irradiance.
3.1. Introduction
While understorey species of evergreen forests often experience high intensity sun-
flecks, they are not usually exposed to prolonged periods of high irradiance (Lovelock
et al. 1998; Tausz et al. 2005). A protective canopy usually creates a favourable micro-
climate with more moderate temperature fluctuations and greater air humidity than in
the above-canopy atmosphere. However, during the course of forest ecosystem dynam-

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-36-
ics including gap formation, bushfires, or anthropogenic management such as forest
harvesting, understorey species may be suddenly exposed to full sunlight and high tem-
peratures, stress factors that can contribute to temporary decline of these species. Ac-
cording to climate-change projections, these factors are likely to become even more sig-
nificant in the future, as temperatures are predicted to increase, and disturbances in for-
est canopies may become more frequent (Hennessy et al. 2007).
In the short term, exposure of shade-acclimated plants to high levels of irradiance often
leads to photoinhibition and photo-oxidative stress. Photoinhibition alone is rarely re-
sponsible for plant mortality and the plant may recover and become fully acclimated.
Photo-oxidative stress is caused by the toxic effects of reactive oxygen species (ROS)
produced in the photosynthetic apparatus under high irradiance when carbon assimila-
tion is light-saturated (Niyogi 2000). Many plants can, to a certain extent, acclimate to
increased irradiance through enhanced dissipation of absorbed light energy in the thyla-
koids, a process related to the conversion of the light harvesting xanthophyll violaxan-
thin to the energy quenching zeaxanthin (Demmig-Adams and Adams 2006). Protection
against high irradiance can also involve the accumulation of tocopherol (Munné-Bosch
2005), an antioxidant that scavenges toxic ROS and contributes to thylakoid membrane
stability.
When, in addition to high irradiance, leaves are exposed to other environmental stress
factors such as high temperature, there can be sustained reductions in the efficiency of
photosynthetic energy conversion and inhibition of repairs to photodamaged photosys-
tem II (PSII; Murata et al. 2007). Photosynthesis is particularly sensitive to inhibition
by heat stress due to labile components in the photosynthetic apparatus (Salvucci and
Crafts-Brandner 2004). The thylakoid membrane is one of the main temperature stress
targets and changes during acclimation occur at that level (Ducruet et al. 2007). The de-
gree of thermostability of the thylakoids can be estimated by the critical temperature Tc
– the temperature threshold above which irreversible damage occurs to PSII (Schreiber
and Berry 1977). Tc changes with growing conditions, reflecting thermal acclimation of
the photosynthetic apparatus (Ducruet et al. 2007). Chlorophyll fluorescence yield is a

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-37-
sensitive indicator of the state of thylakoids, and can be used to assess Tc in plants as the
temperature threshold above which ground level fluorescence (F0) increases (e.g. Froux
et al. 2004). The mechanisms underlying acclimatory changes in Tc are still poorly un-
derstood, although some results point towards stabilising effects of protective com-
pounds on thylakoids. For example, the xanthophyll zeaxanthin (Havaux and Gruszecki
1993; Havaux and Tardy 1996), as well as increased soluble sugar concentration (Hüve
et al. 2006), are believed to have a stabilising effect and shift Tc towards higher tem-
peratures.
Our model understorey species, the tree fern Dicksonia antarctica (Labill., Dick-
soniaceae), is known to decline after clearcut logging in Victoria, Australia (Ough and
Murphy 2004). These tree ferns are iconic and ecologically significant understorey spe-
cies in many humid forest types in the Southern Hemisphere, including Australian tem-
perate rain forests and wet sclerophyll (eucalypt) forests (Large and Braggins 2004).
They support a large epiphytic diversity on their trunks and provide nursery sites for
many tree and shrub species as well as nesting and feeding sites for marsupials, insects
and birds (Lindenmayer et al. 1994; Roberts et al. 2005). Decline in D. antarctica num-
bers is expected to negatively impact on many dependent species and thus maintenance
of tree ferns is often an objective of forest management plans (Department of Natural
Resources and Environment 2002).
The reasons for poor survival and ongoing decline of D. antarctica after logging remain
uncertain (Ough and Murphy 2004), but exposure to high irradiance, combined with in-
creased air and frond temperatures, could be contributing factors. Periodically disturbed
by wildfires in their natural habitat, D. antarctica are exposed to a broad range of irradi-
ance during their lifetime (Hunt et al. 2002), which suggests that this species is able to
at least partly acclimate to different levels of irradiance. Certainly, other studies indicate
potential for fern acclimation to different light regimes. For example, New Zealand
ferns from contrasting habitats displayed contrasting characteristics in terms of photo-
synthetic light compensation point, which were tightly correlated with specific frond
area (Bannister and Wildish 1982). Frond characteristics (frond surface area, epidermis

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-38-
thickness, palisade/spongy mesophyll ratio, blade size, petiole length) of a South
American Cyathea species (another important tree fern genus) were also correlated with
its local irradiance (Arens 1997). However, other studies suggest limited capacity of
shade-acclimated tree ferns to efficiently adjust to increased irradiance (Durand and
Goldstein 2001). To our knowledge, only a few studies have examined effects of high
temperature, either alone or with high irradiance on the physiological performance of
tree ferns: Tingey et al. (1987) found that photosynthesis of D. antarctica was particu-
larly susceptible to inhibition with increasing temperature and high light; and Nobel et
al. (1984) also mentioned negative effects of high temperature on gas exchange of ferns.
D. antarctica’ s natural distribution in Australia is limited to the temperate zone
(McCarthy 1998), characterised by cool to warm conditions (Köppen classification Aus-
tralian Bureau of Meteorology, http://www.bom.gov.au/iwk/climate_zones, verified 13
August 2009), perhaps indicating that the species has limited potential for acclimation
to temperature increases (such as after clearcut logging, but potentially also due to cli-
mate change), making it susceptible to ongoing decline.
In this study, we investigated the responses of D. antarctica to high irradiance, moder-
ately high temperature (+35ºC), and a combination of both under fully controlled cli-
mate chamber conditions. Measuring Tc together with a number of variables related to
photosynthesis, chlorophyll fluorescence, and chloroplast pigments, we addressed the
following specific questions:
Are photosynthetic parameters of D. antarctica adversely affected by a) high ir-
radiance; b) high temperatures; and c) their interactions?
Does membrane stability (measured via critical temperature, Tc,) increase in D.
antarctica fronds with increased temperature (indicative of an acclimation to high
temperature), and if yes, are Tc changes associated with accumulation of osmotica
or zeaxanthin formation?
Do other potentially protective thylakoid compounds, such as carotenoids and to-
copherol, change in relation to high irradiance and high temperature?
Are effects of high temperature on the above parameters reversible?

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-39-
3.2. Material and methods
3.2.1. Plant material
One-year-old sporophytes of Dicksonia antarctica Labill (HSK Gardening and Leisure
Avon Dassett, UK) were transplanted into 10-l pots, containing a mixture of sand and
peat (50/50 v/v) and 40g slow release fertiliser (Nutricote 100, Chisso-Asahi Fertilizer
Co. Ltd, Tokyo, Japan, N/P/K, %, 13/13/13) per pot. The plants were grown under uni-
form sunlit conditions in a naturally illuminated glasshouse at INRA, Champenoux,
France (48º44‟N, 6º14‟E) for two months in spring 2007. At the end of this period, the
plants were transferred to a climate chamber (Chambre Phytotronique STRADER, An-
gers, France) at Champenoux.
3.2.2. Climate chamber conditions and experimental design
Irradiance in the climate chamber was provided by two types of 400 W lamps (HQI
Philips (mercury halide) and SONT Philips (sodium halide) Koninklijke Philips Elec-
tronics N.V., Eindhoven, The Netherlands) and resulted in a photosynthetic photon flux
density (PPFD) of 900 µmol photons m-2
s-1
at plant height (PAR range, 400-700 nm,
measured with a Li-Cor quantum sensor). Relative humidity was 70-80%, air tempera-
ture was controlled to ± 0.5ºC (see temperature treatments below), and photoperiod was
16 h day-1
.
The tree ferns were randomly assigned to two experimental groups (n = 7 in each). One
group was shielded by a wavelength-neutral shade mesh (17% light transmission,
PPFD: 170 µmol photons m-2
s-1
) – „shade‟, the other exposed to full light (900 µmol
photons m-2
s-1
) – „high irradiance‟. Such levels are representative of irradiance condi-
tions in the open on overcast winter and clear summer days respectively, at typical D.
antarctica field sites in mountain ash forests of Victoria, Australia. Plants were kept
well-watered at all times and were rotated daily at random within their designated ir-
radiance regime.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-40-
A sequence of temperature treatments was applied as follows:
(1) 10 days at 15ºC day and night;
(1a) 3 days at 25°C day and night (to avoid heat shock);
(2) 12 days at 35°C/25°C day/night (typical hot summer days for D. antarctica in
the field);
(3) 10 days at 15°C day and night (to check reversibility of temperature effects).
Chlorophyll a fluorescence and critical frond temperature were measured for each indi-
vidual every 1 to 2 days throughout the experiment. Photosynthesis was recorded from
net CO2 uptake (A) versus intercellular CO2 concentration (A-Ci curves) on the first day
of the experiment and at the end of each temperature treatment. Samples for nitrogen
(N) content, xanthophyll analyses, and osmolality were taken at the end of each of the
three temperature treatments. All measurements were made on the mid-third of the
youngest fully-expanded fronds that were of healthy appearance (i.e. not discoloured).
Total chlorophyll content was measured for each individual every 1 to 2 days through-
out the experiment; measurements were made over the entire plant irrespective of frond
condition.
3.2.3. Frond temperature (Tfrond)
Frond temperature (as 10 random points across the entire plant) was measured twice
during the 35ºC temperature treatment (beginning/end) at predawn and midday, and
once at the end of the experiment (during the second 15ºC temperature treatment), using
an IR laser thermometer (Raynger PM, Raytech Inc, Santa Cruz, CA, USA).
3.2.4. Maximal quantum yield of photochemistry (Fv/Fm)
Fv/Fm was derived from chlorophyll a fluorescence measured on dark-acclimated fronds
(at the end of the „night‟ period) with a modulated fluorometer (PAM 2000, Heinz Walz

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-41-
GmbH, Effeltrich, Germany). Maximum quantum yield of PSII was estimated as Fv/Fm
= (Fm–F0)/Fm, after Maxwell and Johnson (2000).
3.2.5. Gas exchange measurements
Gas exchange parameters were measured using a Li-Cor 6400 gas exchange system,
equipped with a 2x3 cm broadleaf chamber (Li-Cor, Lincoln, Nebraska, USA). All gas
exchange measurements were conducted at the reference frond temperature of 25ºC. For
each plant, an A-Ci curve was generated at PPFD 1000 µmol m-2
s-1
, frond temperature
25ºC, air flow rate 400 µmol air s-1
, and relative humidity (RH)>60%. Photosynthesis
was induced at a CO2 mole fraction of 50 µmol mol-1
for 15-20 min prior to measure-
ments to ensure maximal stomatal opening and maximal activity of Calvin cycle en-
zymes. CO2 mole fraction was then increased in 13 successive steps to 2200 µmol mol-1
with two measurements at each step. After finishing the A-Ci curve, illumination in the
leaf chamber was turned off, CO2 mole fraction was decreased to 400 µmol mol-1
and
respiration rate was recorded after 5 min in the dark. The frond area enclosed in the Li-
Cor chamber was marked, photographed and calculated using imaging software
(UTHSCSA Image Tool Version 3, University of Texas, USA). Values for Amax and gs
at ambient CO2 (400 µmol mol-1
) were derived from these curves.
Using a biochemical photosynthetic model Farquhar (Farqhuar et al. 1980), apparent
(i.e. assuming that mesophyll conductance to CO2, gi, is infinite) maximum carboxyla-
tion rate (apparent Vcmax), and the maximum apparent rate of electron transport (appar-
ent Jmax) were estimated by fitting the A-Ci curves to the model as described in Mont-
pied et al. (2009). Triose phosphate use (TPU) limitation was not included in the model;
points with decreasing A at high CO2 mole fractions were disregarded. A set of primary
parameters of Rubisco kinetic properties used herein, Kc= 327 µmol mol-1
, Ko = 282600
µmol mol-1
, Γ* = 43.7 µmol mol-1
, were taken from von Caemmerer et al. (1994).
Then, gi was estimated with the curve fitting approach introduced by Ethier and
Livingston (2004) and described by Montpied et al. (2009) and real Vcmax and Jmax were

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-42-
computed based on the A-Cc (chloroplastic CO2 mole fraction); therefore, only corrected
Vcmax and Jmax (i.e., under the hypothesis of finite gi) are given and discussed in this
study.
3.2.6. Frond nitrogen and chlorophyll content
Frond samples were analysed for nitrogen (N) content using an elemental analyser
(NCS 2500, CE instrument Thermo Quest, Milano, Italy). Samples were dried at 60ºC
for 48 h (to determine dry weight) and then ground to a fine powder. Frond area of fresh
samples was scanned and calculated using Scion Image software (Scion Corporation
2000-2001, USA), and these data used to calculate N content on a frond area basis.
Frond chlorophyll content was estimated from transmittance values measured with the
Minolta SPAD-502 Chlorophyll meter (Minolta, Illinois, USA; hereafter „SPAD‟). Val-
ues were the mean of two to three separate pinnules per plant (randomly selected irre-
spective of frond colour).
Total chlorophyll (a + b) was also measured by HPLC (see Pigments and tocopherol
determination).
3.2.7. Critical temperature (Tc)
Critical temperature was estimated in vivo from the sharp rise of basal chlorophyll a
fluorescence under increasing temperature (Schreiber and Berry 1977). Disks of tree
fern pinnules were placed into a temperature-controlled aluminium body, with the fibre-
optics of the fluorometer (PAM 2000, Walz, Effelrich, Germany) pointing at the sam-
ple. Ground fluorescence (F0) was induced with a red diode at low PPFD of about 1
µmol m-2
s-1
. Temperature of the aluminium body was increased gradually (1°C min-1
)
from 20°C to 60°C. F0 was continuously recorded and critical temperature (Tc) was es-
timated graphically at the beginning of the heat-induced fluorescence rise (Froux et al.
2004).

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-43-
3.2.8. Total tissue osmolality
Total tissue osmolality was measured using freeze-point depression from hot water ex-
tracts of dried frond tissue (Callister et al. 2006). Approximately 60 mg of dried ground
frond tissue (as prepared for N analysis) was placed in a 2-ml polypropylene vial to
which 1.6 ml of deionised water was added. The samples were placed in a water-bath at
90ºC for 60 min. The samples were left to cool to room temperature, centrifuged at
10,000 x g for 2 min and 1 ml of the supernatant was transferred to a 1.7-ml polypro-
pylene vial. Osmolality of the solution was measured using an OSMOMAT 030 cryos-
copic osmometer (Gonotec, Berlin, Germany).
3.2.9. Pigments and tocopherol determination
Frond discs (3.75 mm diameter) were collected at midday at the end of each of the three
temperature treatments and immediately frozen in liquid nitrogen. Samples were freeze-
dried, sealed with silicagel in airtight plastic bags, and kept at -20ºC until analysis.
Four discs per plant were ground in a Matrix Mill (Retsch MM301, Germany) at the
temperature of liquid nitrogen. To avoid the presence of traces of acid in the acetone
used for the extraction, 0.5 g l-1
of calcium carbonate were added to samples prior to
grinding (García-Plazaola and Becerril 1999). The resulting powder was extracted with
0.5 ml of ice-cold acetone, homogenised and centrifuged at 4ºC for 1 min at 15000 x g.
The pellets were re-extracted as described above to a combined sample volume of 1 ml.
Extracts were stored in sealed vials at -20ºC until analysis. Prior to injection, samples
were centrifuged at 4ºC for 20 min at 15000 x g and the clean supernatant was trans-
ferred into HPLC vials.
HPLC separation of chloroplast pigments and tocopherols was according to the methods
given in Tausz et al. (2003). Chromatographic conditions were:

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-44-
Pigments: 25 x 4.6 mm Spherisorb ODS 25 µm column. Gradient: solvent A: ace-
tonitrile: methanol: water = 100:5:10 (v/v/v), solvent B: ethylacetate: acetone = 1:2
(v/v), 10% B to 70% B in 17 min, hold at 70% B for 5 min, return to 10% B in 5
min. Flow rate 1 ml min-1
. The injection volume was 20 µl, photometric detection at
440 nm.
α-Tocopherol: 25 x 4.6 mm Spherisorb ODS 25 µm column. Solvent 100% metha-
nol isocratic. Flow rate 1 ml min-1
. Injection volume was 20 µl. Fluorescence detec-
tion excitation 295 nm, emission 325 nm.
Acetone, acetonitrile, methanol and ethyl acetate were of HPLC grade and water was
deionized. A standard of α-tocopherol was purchased from Sigma (Sigma-Aldrich, Cas-
tlehill, NSW, Australia); standards for carotenoids and chlorophyll a and b were pre-
pared as follows: several generic extracts were prepared in 100% acetone and measured
at three wavelengths in the spectrophotometer (Varian UV/V 300, USA). Using the
equations of Lichtenthaler (1987), chlorophyll a, b and total carotenoid concentrations
were calculated at a spectrophotometer resolution range of 1 – 4 nm. The same extracts
were then re-run in the HPLC and conversion factors for chlorophyll a, b and total caro-
tenoids were calculated, disregarding the minor differences in carotenoid absorption co-
efficients at the wavelength in question.
3.2.10. Statistical analysis
Repeated measures models of SPSS 15 (SPSS Inc. Chicago, USA) were used for statis-
tical analysis, with irradiance as the between-subject factor and temperature as the
within-subject factor (both fixed). Effects of irradiance (high irradiance, shade), and 3
levels of temperature (15ºC, 35°C, back to 15ºC), and irradiance by temperature interac-
tions on each dependent variable were analysed. Data for statistical analyses were the
values per individual plant at the end of each temperature treatment. Photosynthetic pa-
rameters, measured before the start of the experiment were not used in the model.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-45-
3.3. Results
3.3.1. Frond temperature (Tfrond)
Tfrond did not differ among plants at the beginning of the experiment (data not shown).
After the temperature increased to 35ºC, Tfrond was similar in shaded and high irradiance
plants at predawn (below 25°C) but differed on average by 1.5ºC at midday (around
35°C vs. 33.5°C, Table 3.1). By the end of the treatment at 35ºC, Tfrond of shaded plants
was on average 0.7ºC cooler at predawn, and 3.3ºC cooler at midday (Table 3.1).
Table 3.1. Temperature of Dicksonia antarctica fronds (Tfrond) exposed to high irradi-
ance and under shade. Values are means ± s.e. (n = 7 plants, ten measurements per
plant) at predawn and midday on the first (1) and last (12) days of two temperature
treatments (35ºC and back to 15ºC). P values indicate significance of difference be-
tween high irradiance and shaded plants within temperature treatments (Student‟s t-test)
35ºC
P
Back to 15ºC
P High
irradi-
ance
Shaded
Differ-
ence
High
irradi-
ance
Shaded Differ-
ence
Pre-
dawn
Day 1 24.3±0.1 24.2±0.1 0.1 0.30 15.3±0.0 16.0±0.0 -0.7 <0.001
Day 12 23.0±0.2 22.3±0.2 0.7 0.02
Midday
Day 1 34.9±0.6 33.4±0.3 1.5 0.02 19.7±0.4 17.2±0.4 2.5 <0.001
Day 12 34.6±0.2 31.3±0.5 3.3 0.002

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-46-
3.3.2. Maximum quantum yield of PS II (Fv/Fm) and photosynthetic capacity
parameters
Predawn Fv/Fm remained close to the optimum value of 0.83 in shaded plants across all
temperature treatments. In contrast, Fv/Fm declined after the first day of exposure to high
irradiance (Fig. 3.1 a). The 35ºC treatment resulted in further decreases of Fv/Fm (Fig.
3.1 a). After return to 15°C, a partial recovery of Fv/Fm was detected.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-47-
Fig. 3.1. a) Time course of maximum quantum efficiency of PSII and b) chlorophyll
content of high irradiance (open symbols) and shaded (closed symbols)D. antarctica
during three successive temperature treatments(delineated by dotted lines). Values are
means of n = 7 (± s.e.). P-values indicate significance of effects of irradiance (I), tem-
perature (T), and irradiance by temperature interaction (I x T).

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-48-
Photosynthetic parameters were comparable among all plants at the start of the experi-
ment (Table 3.2, „before‟). Ten days of high irradiance (at 15ºC) resulted in decreases in
both light-saturated rate of net photosynthesis at a reference temperature of 25°C,
Amax25
, and corresponding stomatal conductance gs (Table 3.2; Fig. 3.2). Increasing the
temperature to 35ºC induced stomatal opening in all plants (Table 3.2). However, while
Amax of all shaded plants also increased (Table 3.2. Fig. 3.2), Amax was less responsive in
high irradiance plants (Fig. 3.2). Nonetheless, all changes in gs and Amax were fully re-
versible upon return to 15ºC (Table 3.2).
When measured at the reference temperature of 25°C, maximal carboxylation rate,
Vcmax, and maximal light-driven electron flux Jmax decreased in response to increased
irradiance. Both were insensitive to temperature treatments regardless of irradiance re-
gime (Table 3.2). Mesophyll conductance to CO2, gi, was highly variable, probably ow-
ing to the low accuracy of the fitting procedure used. As a result, no significant effects
of irradiance and temperature could be detected (Table 3.2).
Nitrogen content per frond area (g m-2
) was not affected by either irradiance or tempera-
ture (Table 3.2). Photosynthetic nitrogen use efficiency (PNUE), calculated as Vcmax/N,
was lower under high irradiance than in shade and remained such until the end of the
experiment. Changes in temperature did not affect PNUE (Table 3.2).
Total chlorophyll content (in SPAD units) decreased at 35ºC under high irradiance and
remained low thereafter (Fig. 3.1 b). A similar albeit not significant effect was detected
for chlorophyll a + b in HPLC extracts (Table 3.2). It should be noted that visually
damaged fronds were avoided for HPLC analyses, while SPAD measurements were
made across entire fronds. Chlorophyll a/b ratios were similar between irradiance re-
gimes during the 15ºC temperature treatment. With temperature increase to 35ºC, chlo-
rophyll a/b ratios decreased in high irradiance plants in contrast to a significant increase
in shaded plants (Table 3.2). With temperature return to 15ºC, chlorophyll a/b ratios of
high irradiance plants recovered to the initial values.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate heat on photosynthesis, pigments, and tocopherol in the tree fern
Dicksonia antarctica. Functional Plant Biology In press.
-49-
Table 3.2. Photosynthesis and frond traits of D. antarctica exposed to high irradiance and under shade before and during three successive
temperature treatments. Values are means ± s.e. (n = 7) of Amax25
, light-saturated net CO2 assimilation rate at 25°C; gs, stomatal conduc-
tance under saturating irradiance at 25°C; NA, nitrogen content on a frond area basis; PNUE, photosynthetic nitrogen use efficiency
(Vcmax25
/NA) at 25°C; Chl total, total chlorophyll content in SPAD units and on a frond area basis; Chl a/b, chlorophyll a/b ratio; Vcmax25
,
maximal carboxylation rate of Rubisco at 25°C; Jmax25
, maximal light driven electron flux at 25°C; gi, mesophyll conductance to CO2
measured at 25°C; Effect abbreviations: I, Irradiance; T, temperature; I x T, irradiance by temperature interaction; Significance lev-
els:*P≤0.05; **P<0.01; ***P<0.001; n.s., non significant; n.d., no data
Parameter
Irradiance regime
Temperature treatments Significance of
effects (P)
Before 15ºC 35ºC Back to 15ºCº
Amax25
(µmol CO2 m-2
s-1
) High irradiance 6.0±0.5 3.7±0.5 5.1±0.6 3.9±0.4
I***; T* Shade 6.4±0.4 6.4±0.8 7.6±0.3 5.6±0.5
gs at Amax25
(mmol H2O m-2
s-1
)
High irradiance 82±11 57±6 108±7 50±6 I*; T***
Shade 80±7 80±14 152±12 70±5
Vcmax25
(µmol CO2 m-2
s-1
) High irradiance 37.0±2.0 29.3±2.7 23.2±2.8 29.1±3.3 I***
Shade 36.0±1.0 37.1±2.9 40.6±3.8 34.5±4.2
Jmax 25
(µmol CO2 m-2
s-1
) High irradiance 115±6 68±9 50±7 85±11 I**
Shade 105±6 85±10 86±12 98±10

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate heat on photosynthesis, pigments, and tocopherol in the tree fern
Dicksonia antarctica. Functional Plant Biology In press.
-50-
Parameter
Irradiance regime
Temperature treatments Significance of
effects (P)
Before 15ºC 35ºC Back to 15ºCº
gi (mmol CO2 m-2
s-1
)
High irradiance 155±64 141±53 143±115 115±25 n.s.
Shade 115±35 222±63 294±195 259±112
NA (g m-2
)
High irradiance n.d. 16.9±1.0 15.9±0.6 17.4±0.7
n.s. Shade n.d. 16.5±0.7 16.0±0.8 15.5±2.1
PNUE (Vcmax25
/N),(µmol
mol-1
N-1
m-2
)
High irradiance n.d. 204±21 201±24 175±13 I**
Shade n.d. 278±18 311±30 300±41
Chl total (SPAD units) High irradiance n.d. 49.1±1.7 39.3±2.3 34.1±2.1
I**; T***;
I x T*** Shade n.d. 48.9±0.4 47.1±0.5 47.3±1.0
Chl total (a + b)
(µmol m-2
)
High irradiance n.d. 782±142 721±111 675±90 n.s.
Shade n.d. 810±63 914±127 843±81
Chl a/b
High irradiance n.d. 2.36±0.05 2.29±0.05 2.41±0.07 I**; T**;
I x T*** Shade n.d. 2.46±0.05 2.71±0.05 2.66±0.07

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate heat on photosynthesis, pigments, and tocopherol in the tree fern
Dicksonia antarctica. Functional Plant Biology In press.
-51-
Fig. 3.2. Stomatal conductance, gs versus light-saturated rate of net photosynthesis Amax for high irradiance (on the right) and shaded (on
the left) D. antarctica measured at the standardised temperature of 25ºC during three temperature treatments: 15ºC (open circle), 35ºC
(closed diamond) and back to 15ºC (open triangle). Values are means of n = 6 (high irradiance) and n = 7 (shaded).

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-52-
3.3.3. Critical temperature (Tc)
During the 15°C treatment, critical temperature for photochemistry (Tc) was similar un-
der shade and high irradiance (means of 47.5ºC and 47.2ºC respectively; Fig. 3.3). In-
crease in temperature resulted in significant rises of Tc that were greatest under high ir-
radiance. Tc started to decrease after return to 15°C, although pre-treatment values were
not reached by the end of the experiment (Fig. 3.3).
Fig. 3.3. Time course of critical temperature, Tc of high irradiance (open symbols) and
shaded (closed symbols) D. antarctica across the experiment. Values are means of n =
7 (± s.e.). P-values indicate significance of effects of irradiance (I), temperature (T), and
irradiance by temperature interaction (I x T).

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-53-
3.3.4. Total tissue osmolality
Total tissue osmolality was not affected by irradiance but decreased significantly during
the 35ºC treatment (Table 3.3). It increased to close to the original values after return to
15°C.
3.3.5. Carotenoids and α-tocopherol
Neoxanthin and lutein contents (per mol total chlorophyll) were significantly greater
under high irradiance, whereas α- and β-carotene contents remained comparable to
shaded plants (Table 3.3). Temperature increase to 35ºC led to significant increase in β-
carotene and lutein in high irradiance plants. These pigments tended to remain high on
return to 15ºC, although α-carotene significantly decreased under high irradiance (Table
3.3).
The xanthophyll cycle pigment pool (i.e., Violaxanthin, Antheraxanthin, and Zeaxan-
thin, V+A+Z) was significantly greater under high irradiance for the whole experiment.
Changes in temperature did not affect V+A+Z under either irradiance regime (Table
3.3). The de-epoxidation state of xanthophylls (expressed as (0.5A+Z)/(V+A+Z)) was
greater under high irradiance than shade (on average 44% vs. 3%; Table 3.3). It de-
creased by ca 32% under high irradiance upon return to 15°C, but was insensitive to
temperature under shade. Small amounts of lutein-epoxide were detected (Table 3.3),
but they were not affected by light, temperature and their interactions.
Content of α-tocopherol was significantly greater under high irradiance than shade (Fig.
3.4). Whereas α-tocopherol was invariant to temperature changes in shaded plants, α-
tocopherol increased in high irradiance plants with temperature increase to 35ºC, and
remained high after the temperature returned to 15ºC; due to the lack of a high light-low
temperature control, we cannot clearly separate the high irradiance and high temperature
effects on these changes.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate heat on photosynthesis, pigments, and tocopherol in the tree fern
Dicksonia antarctica. Functional Plant Biology In press.
-54-
Table 3.3. Pigment content and osmolality of D. antarctica fronds exposed to high irradiance and under shade during three successive
temperature treatments. Values are means ± s.e. (n = 7) of: osmolality; carotenoids (on a chlorophyll basis): lutein, neoxanthin and xantho-
phyll pool (i.e. Violanxanthin, Antheraxanthin, Zeaxanthin, V+A+Z), α- and β-carotene, lutein-epoxide and the de-epoxidation state of
violaxanthin = (0.5A+Z)/(V+A+Z). Effect abbreviations: I, Irradiance; T, temperature; I x T, irradiance by temperature interaction; Signifi-
cance levels:*P≤0.05; **P<0.01; ***P<0.001; n.s. –not significant
Irradiance regime
Temperature treatments Significance of effects
(P) 15ºC 35ºC Back to 15ºC
Osmolality (mosmol g d.w.-1
) High irradiance 1.56±0.06 1.33±0.04 1.45±0.05
T*** Shade 1.55±0.02 1.40±0.03 1.51±0.01
V+A+Z (mmol mol-1
chl-1
) High irradiance 91.9±9.2 118±15.0 113.7±15.9
I*** Shade 47.8±1.7 45.8±1.2 45.5±1.2
De-epoxidation (%) High irradiance 56.6±3.4 53.1±7.7 21.4±4.7
I***; T***; I x T*** Shade 4.4±1.1 2.3±0.8 1.6±0.6
Lutein (mmol mol-1
chl-1
) High irradiance 263±9 339±25 388±29
I***; T**; I x T** Shade 185±4 186±3 198±6

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate heat on photosynthesis, pigments, and tocopherol in the tree fern
Dicksonia antarctica. Functional Plant Biology In press.
-55-
Irradiance regime
Temperature treatments Significance of effects
(P) 15ºC 35ºC Back to 15ºC
Neoxanthin (mmol mol-1
chl-1
) High irradiance 61.3±2.6 65.9±3.8 69.3±3.6
I*** Shade 45.2±1.9 46.2±1.1 48.4±2.0
α-carotene (mmol mol-1
chl-1
) High irradiance 11.1±2.3 24.2±3.4 5.8±1.5
I*; T***; I x T** Shade 16.1±2.6 24.1±2.4 20.9±1.8
β-carotene (mmol mol-1
chl-1
) High irradiance 62.2±7.8 94.6±8.7 89.3±7.0
I***; T** Shade 44.7±5.7 59.4±5.3 59.3±3.3
Lutein-epoxide
(mmol mol-1
chl-1
)
High irradiance 5.2±1.7 6.3±1.4 6.2±2.2
n.s.
Shade 3.4±0.4 3.9±0.4 4.3±0.3

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-56-
Fig. 3.4 α - Tocopherol content of high irradiance (open bars) and shaded (closed bars)
D. antarctica under three temperature treatments. Values are means of n = 7 (± s.e.). P-
values indicate significance of effects of irradiance (I), temperature (T), and irradiance
by temperature interaction (I x T).

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-57-
3.3.6. Correlations between Tc and biochemical parameters
No significant correlation was detected between Tc and osmolality, tocopherol, and α-
and β-carotenes (correlations not shown). Correlations between Tc and zeaxanthin, pre-
sented as Z/(Z+V) (after Havaux and Gruszecki 1993) were also not significant, either
for each irradiance treatment separately or for the combined data (Fig. 3.5).
Fig. 3.5. Critical temperature, Tc versus xanthophyll zeaxanthin (expressed as Z/(Z+V);
after Havaux and Gruszecki, 1993) of high irradiance (open symbols) and shaded
(closed symbols) D. antarctica during three temperature treatments: 15ºC (circle), 35ºC
(diamond) and back to 15ºC (triangle). Each point represents an individual measure-
ment. Relationships were also non-significant when data were separated by irradiance
treatment. Z, zeaxanthin; V, violaxanthin.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-58-
3.4. Discussion
3.4.1. Effect of high irradiance, high temperature and their interaction on pho-
tosynthetic capacity parameters of D. antarctica
Photosynthetic capacity of D. antarctica in this study was within ranges reported in the
literature. Maximum light saturated rates of net photosynthesis (Amax25
) were compara-
ble with values reported for the same species: 6 – 10.8 µmol m-2
s-1
(Nobel et al. 1984)
and 8.3 µmol m-2
s-1
(Hunt et al. 2002). Maximum carboxylation rates, Vcmax25
(the
maximal in vivo Rubisco activity), and the maximum rate of electron transport, Jmax25
, at
the reference temperature of 25°C were comparable to some shade tolerant tree species,
such as silver fir (Abies alba Mill, Robakowski et al. 2002), and mesophyll conduc-
tance, gi, corresponded to typical values of evergreen trees among the species reviewed
by Ethier and Livingston (2004).
Photosynthetic capacity of D. antarctica was adversely affected by high irradiance.
Jmax25
decreased with respect to shaded plants, and this decrease was paralleled by a de-
crease of maximum quantum yield of PSII, Fv/Fm, indicating a moderate but chronic
photoinhibition (Table 3.2; Tallon and Quiles 2007). High irradiance also led to de-
creases in Amax25
, due to both reduced stomatal conductance gs and decreased photosyn-
thetic capacity, i.e., Rubisco activity and Jmax25
. A number of processes may result in
deactivation of Rubisco – for example, remobilisation and export of nitrogen from the
leaves, interruption in the electron transport chain, or the presence of reactive oxygen
species (ROS). In our case, deactivation of Rubisco in high irradiance plants was not
related to a remobilisation of frond nitrogen – the N content per frond area was not af-
fected by irradiance. This can be seen also through the decline of photosynthetic nitro-
gen use efficiency (PNUE). However, high irradiance alone did not affect total chloro-
phyll content and chlorophyll a/b ratios, indicating that photoprotective mechanisms
were efficient enough to avoid chlorophyll degradation.
High temperature had no negative effects on photosynthetic capacity of D. antarctica
under shade. Increasing temperature to 35ºC even stimulated Amax25
. Increases in Amax25

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-59-
were in this case solely due to increases in gs, as we found no effects of temperature on
photosynthetic capacity (Vcmax25
, Jmax25
). The increase in chlorophyll a/b ratio in shaded
plants with increasing temperature perhaps indicates temperature-stimulated resynthesis
of photosynthetic reaction centres relative to light-harvesting antenna complexes, which
commonly coincides with other changes in pigment composition, e.g. lutein, β-carotene
(Haldimann 1999).
The interactive effect of high irradiance and high temperature led to severe photoinhibi-
tion in agreement with earlier findings (Berry and Björkman 1980). Temperature in-
crease stimulated gs yet without commensurate increases in Amax25
, indicating that meta-
bolic limitations (e. g. Rubisco activity) governed Amax25
(consistent with the findings of
e.g. Law and Crafts-Brandner 1999 on Rubisco activation). Many studies have shown a
negative effect of moderate heat and photoinhibition on the activation of Rubisco medi-
ated by an activase (for details see Salvucci and Crafts-Brandner 2004). In our study,
we did not measure the activity of Rubisco-activase, but found no increase in photosyn-
thetic capacity to increased temperatures. Prolonged photoinhibition and heat interac-
tion resulted in decreases in chlorophylls as in numerous other studies (e.g. Lambers et
al. 2008). Decreases in chlorophyll a/b ratio indicated that under prolonged light stress,
chlorophylls were destabilised with chlorophyll a being more sensitive than chlorophyll
b (Yamamoto et al. 2008).
A return of temperature to 15ºC induced stomatal closure thus reversing Amax25
to the
initial values at 15°C, as found by Ghouil et al. (2003). The only partial recovery of
Fv/Fm in high irradiance plants demonstrated detrimental interactive effects of high ir-
radiance and temperature on Fv/Fm, suggesting that degradation processes (i.e. bleaching
of chlorophylls or photo-degradation of thylakoid complexes) did not allow rapid re-
covery of Fv/Fm (Ottander et al. 1995). The fact that Vcmax and Jmax remained non-
responsive possibly underlined the high sensitivity of Rubisco to high irradiance in D.
antarctica.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-60-
In summary, photosynthetic capacity and photosynthetic nitrogen use efficiency were
rapidly affected by exposure to high irradiance under 15°C, while chlorophylls and pre-
dawn Fv/Fm declined further under the combination of high irradiance and high tempera-
ture.
3.4.2. Membrane stability of D. antarctica measured via critical temperature
Critical temperature, Tc, recorded in D. antarctica was ca 47ºC, comparable with a
range of overstorey species, such as Quercus petraea (46.7ºC; Dreyer et al. 2001).
Comparable data for other tree fern species are currently lacking.
An increase in air temperature induced an increase in Tc, as found by many authors (e.g.
Dreyer et al. 2001). This increase in Tc was larger under high irradiance than shade, co-
inciding with a 3.3°C higher midday frond temperature under high irradiance. Previ-
ously published data suggest that an increase in Tc by 1ºC (as found in our study) re-
quires an increase in ambient temperatures of about 10ºC (Froux et al. 2004). We there-
fore believe that the larger rise in Tc in high irradiance plants was not solely caused by
the difference in frond temperature, but directly related to high irradiance effects, which
resulted in enhanced thermostability of thylakoid membranes.
Contrary to findings by Hüve et al. (2006), increased thermostability of the thylakoid
membranes was not associated with the accumulation of osmotically active substances.
In our study, osmotically active solutes even decreased when temperature increased. It
may be argued that the hot water extract method results in artefacts, because some cell
wall or other material can be brought into solution as a result of the grinding and extrac-
tion procedures (Callister et al. 2006). Yet, this method is widely used (e.g. Merchant et
al. 2006) and moreover, a tight correlation was found among methods even though the
absolute results were different (Callister et al. 2006). Our observation therefore seems
reliable.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-61-
With subsequent temperature decrease to 15ºC, we found a tendency for Tc to return to
the initial values, even though this return was not complete. The observed lag confirms
the hysteresis found by Froux et al. (2004), when the increase in Tc with increasing
temperature is faster than relaxation from this effect after temperature decrease.
3.4.3. Xanthophyll cycle carotenoids, pigments and α-tocopherol
Exposure of D. antarctica to high irradiance alone resulted in almost two-fold increase
in xanthophyll cycle carotenoids (V+A+Z), both on a chlorophyll as well as on a frond
area basis. A high de-epoxidation state was also recorded (50% vs. 2-4% in the shade).
Values for V+A+Z were within the range of values presented for other species (Thayer
and Björkman 1990). The maximum de-epoxidation state of the xanthophyll cycle was
lower than maximum values measured in epiphytic ferns (Tausz et al. 2001). An in-
crease was also recorded in pools of other carotenoids such as neoxanthin and lutein in
response to high irradiance. Values were within the range reported for other ferns, e.g. a
range of epiphytic species (Tausz et al. 2001) or the tree fern Cyathea microdonta (Ma-
tsubara et al. 2009). These carotenoids (e.g. neoxanthin) may preserve PSII from photo-
inactivation and protect membrane lipids from photo-oxidation by ROS (North et al.
2007). The observed increase in lutein (located primarily in both the proximal and distal
light harvesting centres of PSI and PSII) is probably associated with the acclimation of
antennae to increasing irradiance (Senger et al. 1993). As antenna size is usually re-
duced in response to increasing irradiance, it is also likely that an increasing fraction of
these carotenoids is not bound to antenna proteins. High irradiance also induced in-
creases in α–tocopherols, known for their protective function in thylakoids, consistent
with findings elsewhere (e.g. García-Plazaola and Becerril 1999; Munné-Bosch 2005).
Increased temperature stimulated an increase in Tc in shaded plants but without simulta-
neous accumulation of zeaxanthin, which is contrary to observations by Havaux and
Gruszecki (1993) and Havaux and Tardy (1996). Irrespective of an increased Tc, high
temperature had no effect on concentration of α-tocopherol in shaded plants. According
to our results, it is therefore unlikely that increased membrane stability as measured by

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-62-
Tc is directly and generally dependent on zeaxanthin or α-tocopherol. There was some
coincidence of increased Tc with increases in α- and β-carotenes in shaded plants, which
may have reflected changes in carotenoid synthesis rates. Carotenes may also contribute
to an overall increase in membrane stability, as proposed by Tausz et al. (2001).
Interactions between irradiance and temperature had no additional effect on the xantho-
phyll cycle pool and de-epoxidation state of high irradiance plants. In contrast to
V+A+Z, the combination of high irradiance with high temperature led to further in-
creases in the amount of lutein and β-carotene, which remained greater until the end of
experiment. It is not clear from our experiment whether sustained concentrations of ca-
rotenoids indicated their role as a last resort in membrane photoprotection under ex-
treme stress, or simply reflected their superior stability under such conditions. In con-
trast to shaded plants, temperature increase stimulated an almost two-fold increase in α-
tocopherol in high irradiance plants. Although these results for high irradiance plants
appear to support findings of Llusià et al. (2005) – who suggested that increased toler-
ance to high temperatures might be at least partly due to an increase in α-tocopherol –
our results for shaded plants indicate that changes in Tc can occur independently of
changes in α-tocopherol.
Temperature return to 15°C did not affect total V+A+Z pool of high irradiance plants,
but the de-epoxidation state decreased by 30% to remain significantly higher than in
shaded plants. De-epoxidation of the xanthophyll cycle is driven by an acidic thylakoid
lumen, which can be the consequence of an imbalance between electron transport and
electron consumption (Demmig-Adams and Adams 2006). It seems that heat related
changes in the photosynthetic apparatus lead to a relaxation of the pH gradient upon
temperature decrease, despite the continuation of high irradiance. This may be related to
a sustained decrease in light use efficiency as suggested by persistently low Fv/Fm val-
ues, and other, as yet unexplained, changes in the photosynthetic membrane. Such fur-
ther changes were also expressed in the observed change in Tc, possibly in combination
with an increased electron consumption rate upon relaxation of the high temperature.
Temperature decrease did not affect concentration of carotenoids except for α-carotene.

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-63-
Significant decreases in α-carotene in high irradiance plants can be explained in terms
of its ease of oxidation to lutein under conditions of oxidative stress (Senger et al.
1993), or its conversion to β-carotene to increase scavenging of free radicals in core
complexes under conditions of stress (Kirchgeßner et al. 2003).
In summary, increased thylakoid stability in D. antarctica observed during our experi-
ment could not be explained by any of the measured changes in pigments or α-
tocopherol, although they may all play partial roles. Alternative or additional explana-
tions may involve presence of certain heat shock proteins or changes in the composition
of membrane lipids, as suggested by Sinsawat et al. (2004). Discrepancies with earlier
literature may be related to the fact that pigment changes are fast responses to tempera-
ture increases (in the order of one day; as e. g. in Havaux and Tardy 1996), while we
investigated longer term acclimation (12 days) to high temperature. Longer term accli-
mation of plants to rising temperatures is also related to the appearance of polar lipids
with saturated fatty acids causing a decrease in membrane fluidity (Zsófi et al. 2009). In
parallel with an increased threshold temperature for thermal inactivation of PSII (Down-
ton et al. 1984) this can increase thylakoid stability. The difference between the rate of
acclimation and de-acclimation supports this hypothesis: the fast rise may be initially
due to rapid changes in pigments that are later completed by slower changes in lipid
composition. The reversal of these changes may be slower, which would be the cause
for a slow return to initial levels of stability. However, our data do not allow confirma-
tion of this speculation.
3.5. Summary
High irradiance caused chronic photoinhibition (measured as sustained decrease in
maximum PSII quantum efficiency), and decreases in all photosynthetic capacity pa-
rameters of D. antarctica. Whilst we observed some acclimation in terms of increases in
protective carotenoids, which may have sufficed to avoid chlorophyll degradation, simi-
lar or even decreasing chlorophyll a/b ratios indicated limited short-term acclimation
potential of D. antarctica fronds to high irradiance. Temperature alone appeared to have

This Chapter is published
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance and moderate
heat on photosynthesis, pigments, and tocopherol in the tree fern Dicksonia antarctica. Functional Plant
Biology In press.
-64-
no negative effect on photosynthesis of D. antarctica, possibly suggesting that this spe-
cies can thrive under moderately high temperature as long as it is shaded. However,
there was ample evidence of severe damage to the photosynthetic apparatus when high
irradiance was combined with moderately high temperatures. Therefore we can specu-
late that future scenarios predicting higher temperatures and more frequent disturbances
may decrease the competitiveness of D. antarctica.

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-65-
Chapter 4. Interactive effects of high light and water deficit on the
tree fern species Dicksonia antarctica and Cyathea australis
(iii) Abstract
We examined the responses of two tree fern species (Dicksonia antarctica and Cyathea
australis) growing under moderate and high light regimes to short-term water deficit
followed by rewatering. Under adequate water supply, morphological and photosyn-
thetic characteristics differed between species. D. antarctica, although putatively the
more shade and less drought adapted species, had greater chlorophyll a/b ratio, and
greater water use efficiency and less negative δ13
C. Both species were susceptible to
water deficit regardless of the light regime showing significant decreases in photosyn-
thetic parameters (Amax, Vcmax, Jmax) and stomatal conductance (gs) in conjunction with
decreased relative frond water content (RWC) and predawn frond water potential (Ψ
predawn). Stomatal conductance under moderate light responded later and at lower soil
water content. More shaded D. antarctica seemed to be most vulnerable to drought as
evidenced by greatest decreases in Ψ predawn, and lowest stomatal conductance and pho-
tosynthetic rates. Both tree fern species were able to recover after short but severe water
stress.
4.1. Introduction
Drought is considered to be one of the most important factors limiting plant perform-
ance (e. g. Ribas-Carbo et al. 2005). Shade grown plants are potentially less tolerant to
reduced soil moisture than light grown plants because survival in light limiting envi-
ronments generally requires a large leaf area, which can only be supported under moist
conditions (Smith and Huston 1989). This is supported by a number of studies e. g. Val-
ladares and Pearcy (2002), Valladares and Niinemets (2008), which have found that
shade and drought tolerance cause conflicting requirements for biomass investment.

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-66-
Shade tolerance favours foliage for efficient light capture, whereas drought tolerance
requires predominant investment in roots for efficient water uptake.
During periods of insufficient water supply, plants may be exposed to light conditions
in excess of their ability to use in photosynthetic fixation. The resulting imbalance be-
tween electron transport and consumption leads to photoinhibition and photo-oxidative
stress (Flexas et al. 1999). Thus, an interactive effect of high light and water deficit can
be more detrimental than water deficit alone (Levitt 1980, Lovelock et al. 1994). Val-
ladares and Pearcy (2002) observed that the capacity to withstand severe drought was
not enhanced in the shade but was decreased due to increased below-ground competi-
tion for water with established trees. Furthermore, photoinhibition becomes relatively
more important for carbon gain in shade than in sun due to the relatively more important
effect of low photochemical efficiency under low light following sunflecks (Valladares
and Pearcy 2002).
The tree ferns Dicksonia antarctica (Labill.) and Cyathea australis (R.Br.) Domin are
ecologically important (Lindenmayer et al. 1994, Roberts et al. 2005) understorey spe-
cies of south-eastern Australian forest ecosystems, including wet sclerophyll forests and
cool temperate rainforests characterised by mild to warm summers with occasionally
short periods of droughts and high temperatures (Australian Bureau of Meteorology,
2006). D. antarctica and C. australis, are believed to have different micro-habitat pref-
erences as D. antarctica typically dominates wet, shady gullies while C. australis is
more common along forest margins (McCarthy 1998). An observational study con-
firmed that the greater the distance to the stream the more likely it‟s to encounter C.
australis than D. antarctica (Dignan and Bren 2003). D. antarctica and C. australis
belong to contrasting floristic elements of the Australian vegetation (Gondwanan vs.
Tropical, Groves 1994) thus physiological adaptation to micro-habitats (if they exist)
are likely to have arisen during the contrasting phytogeographical history of these spe-
cies.

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-67-
During their lifetime, tree ferns can be periodically exposed to harsh conditions of post-
wildfire environments, characterised by increased light intensities and leaf temperatures,
and consequently increased evapotranspiration and water loss. Direct effects of these
conditions on tree fern physiology have not been studied, but are indicated by poor sur-
vival and ongoing decline of both D. antarctica and C. australis after clearcut logging
(Ough 2001). Understanding negative effects of excess light and water deficit on tree
fern persistence is of increasing importance given predictions of more frequent drought
and fire events in climate change scenarios relevant to south-eastern Australia
(Hennessy et al. 2007).
Both D. antarctica and C. australis are able to tolerate short periods of drought if some
shade is available. Despite infrequent but severe drought events, tree fern numbers in-
creased by 80% in the lower strata of wet sclerophyll forests over 48 years (Ashton
2000). Hunt et al. (2002) also reported that D. antarctica can maintain favourable water
relations during short periods of drought, if its habitat is limited to sheltered sites. This
apparent ameliorating effect of shade on drought might simply be due to reduced water
loss from soil and plants, rather than a direct effect of light intensity. Observational field
studies usually have to accept such confounding effects between shade, temperature and
(soil and air) humidity, because more shaded sites are also cooler and moister. Thus, it
often remains unresolved whether alleviation of drought stress is a direct effect of lower
irradiance – e. g. shading ameliorates drought-related photoinhibition and photo-
oxidative stress – or an indirect effect of greater water availability.
In this study, we compared interactive effects of short-term water deficit and high light
on the physiological performance of two tree ferns – one (D. antarctica) putatively less
drought/light tolerant than the other (C. australis) – by applying short-term but severe
water deficit treatment to potted individuals under semi-controlled conditions. We ana-
lysed key variables relevant to photosynthetic capacity (gas exchange, chlorophyll a
fluorescence), and plant water relations (water potential, osmolality, carbon isotope dis-
crimination) during a drought-rewatering cycle. Our research hypotheses were:

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-68-
The tree fern species will display contrasting physiological characteristics, owing
their different origins and microclimate preferences, where D. antarctica will be
more susceptible to water deficit;
Water deficit will be detrimental for high light exposed tree ferns, while shade
will ameliorate the negative effects of water deficit;
The tree ferns will be able to recover after short but severe water stress, with those
in shade recovering faster.
4.2. Materials and methods
4.2.1. Plant material
Twenty sporophytes of Dicksonia antarctica (Labill.) and twenty of Cyathea australis
(R.Br. Domin) (Fern Acres nursery, King Lake West, central Victoria, Australia) were
transplanted two months before the experiment into spacious 25-l pots, containing (%
volume) composted pine bark (30), gravel (45), coarse fern mulch (5), composted mulch
(14.5), fine fern mulch (5), „Dynamic lifter‟ (0.16, Yates, Padstow, NSW, Australia),
and two types of slow-release fertiliser (0.17 each) Osmocote Baulkham Hills, NSW,
Australia (18+4.8+8.3 mg) and (16+3.5+10+1.2 mg). Before the experiment plants grew
in an open-air nursery under the dense canopy, providing ca 70% of shade. All plants
were two and a half years-old at the start of the experiment.
4.2.2. Experimental design
The experiment ran from February to April 2008 under the prevailing summer to au-
tumn weather at Creswick Campus of the Melbourne School of Land and Environment,
The University of Melbourne, Victoria, Australia (143º 53‟ E, 37º 25‟ S; 392 m above
sea-level).
Ten plants of each species were randomly assigned to two light regimes – “moderate
light” (35% of ambient light intensity by a wavelength neutral shade-cloth) and “high
light” (70% of ambient light intensity). We choose 35% of ambient light as most repre-

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-69-
sentative for field conditions of south-eastern Australia (personal observations), while
70% of ambient light was the required minimum (derived from previous experiments,
unpublished) to protect plants from high light stress. Maximum photosynthetic photon
flux density (PPFD, 400 – 700 nm, measured with a Li-Cor PAR sensor, Li-Cor, USA)
at noon on clear sunny days was on average 600 µmol photons m-2
s-1
in moderate light
and 1000 µmol m-2
s-1
in high light at frond height, (ambient maximum PPFD ca
1800 µmol m-2
s-1
). Measurements with a Solarmeter (Ultraviolet Radiometer 5.0, So-
lartech Inc.) confirmed that UV-A and UV-B filtration was similar to that in the PAR
range (30% and 60% filtration, respectively). Weather conditions were recorded by a
weather station (Tain™ Electronics, Melbourne, Australia).
Within each light regime, 5 plants per species were randomly assigned to two water
treatments: - watered daily to field capacity for the entire experiment (control) and wa-
ter-deficit (deficit). Plants were divided into five blocks, each containing one plant per
species and four treatments (i.e. all combinations of “control” and “deficit” with “mod-
erate light” and “high light”). All measurements (as below) within one block were made
on the same day, with a one day lag in the application of water deficit treatments for
consecutive blocks. Plants were rotated randomly within their designated light regime
three times per week to minimise differences in light and temperature conditions.
Plants were measured:
(1) After 20 days of acclimation to designated light regimes, all plants regularly wa-
tered to field capacity – “pre-treatment” period;
(2) After 10 days of treatment – “water deficit” period (deficit plants not watered for
5 days then maintained at 10% of field capacity for next 5 days);
(3) Twenty days of watering to field capacity – “rewatering” period;
Additional shade cloth (to reach 50% of ambient light intensity) was placed over
high light plants to avoid damage in extremely hot weather (>35°C) during the
first 10 days of the rewatering period (Fig. 4.1).

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-70-
Each plant was measured for chlorophyll a fluorescence, predawn water potential and
gas exchange at the end of each of the three experimental periods. In addition, stomatal
conductance (gs) was measured daily for the first five days of water deficit period from
8 to 11.30 a.m. Samples for chlorophyll content, osmolality and stable isotopes were
taken in the morning at the end of each of the three periods. All measurements were
made on the mid-third of the youngest fully expanded fronds.
4.2.3. Maximum quantum yield of PSII (Fv/Fm)
Maximum quantum yield of PSII (Fv/Fm) was measured at predawn with a pulse modu-
lated fluorometer (OS-30p, Opti-Sciences, Hudson, USA). Ground fluorescence (F0)
was obtained with a low intensity modulated light (600 Hz, 650nm, PPFD<1 µmol m-2
s-
1). Maximum fluorescence (Fm) was induced by a saturating flash. Maximum quantum
yield of PSII was estimated as Fv/Fm = (Fm–F0)/Fm, nomenclature after Maxwell and
Johnson (2000).
4.2.4. Photosynthetic capacity
Gas exchange was measured with a Li-Cor 6400 portable photosynthesis measurement
device, equipped with a 2x3 cm broadleaf chamber with red-blue LEDs (Li-Cor, Lin-
coln, Nebraska, USA). All gas exchange measurements were conducted at the reference
frond temperature of 25ºC. For each plant, an A-Ci curve was generated after Long and
Bernacchi (2003) with some modifications: PPFD was 1000 µmol m-2
s-1
, frond tem-
perature was 25ºC, air flow rate was 400 µmol air s-1
, and relative humidity (RH)>60%.
CO2 mole fraction was increased in 13 successive steps from 50 to 2200 µmol mol s-1
with two measurements at each step. After finishing the A-Ci curve, illumination in the
leaf chamber was turned off, CO2 mole fraction was decreased to 400 µmol mol-1
and
respiration rate was recorded after 5 min in the dark. The frond area enclosed in the
chamber was marked, detached, scanned and calculated using Scion Image programme
(Scion Corporation 2000-2001, USA). Gas exchange measurements were then recalcu-
lated for real frond area.

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-71-
Intrinsic water use efficiency (WUEi) was calculated as Amax/gs; where Amax and gs were
measured at the conditions mentioned above and reference CO2 of 397±1 µmol mol s-1
.
Using the Farquhar model (Farqhuar et al. 1980), maximum carboxylation rate (Vcmax),
and maximum electron transport rate (Jmax) were evaluated by fitting A-Ci curves to the
model, as described in Dreyer et al. (2001). Triose phosphate use (TPU) limitation was
not included in the model, and corresponding points with decreased A at elevated Ci
were disregarded. The set of primary parameters of Rubisco kinetic properties used
herein (Kc=327µmol mol-1
, Ko=282.6 mmol mol-1
, Γ*=43.7 µmol mol-1
) are from von
Caemmerer et al. (1994).
A time course of gs versus days of water withheld was measured using the Li-Cor 6400
gas exchange system, with chamber conditions as above and a reference CO2 concentra-
tion of 397±1 µmol mol-1
.
For measurements of frond chlorophyll content, four frond discs (3.75 mm diameter)
were immediately immersed in liquid nitrogen and then stored at -80ºC until extraction.
Chlorophyll a and b were extracted using 1.8 ml of 100% dimethyl sulphoxide
(DMSO). Extracts were heated for 30 min at 65ºC in a dry block heater (Termoline
L+M, Northgate, Queensland, Australia). The supernatant was then transferred to a mi-
croplate reader (Tecan GmbH, Austria). Blank microplates were scanned and their ab-
sorbance was deducted from final measurements. Chlorophyll a, b and total were calcu-
lated according to Wellburn (1994). The absorbance of 200 μl of sample in a microplate
was converted into a 1 cm pathlength, and then corrected using correction coefficients
(Warren 2008). Correction coefficients for chlorophyll a and b were calculated from 20
samples using regressions of initial measurement against repeated measurement in a
spectrophotometer (Carry 300, Varian, The Netherlands) at the same wavelengths (all
regressions were highly significant P<0.001).
Leaf mass per area (LMA), needed to calculate total chlorophyll on a frond area, was
calculated as dry weight/frond area (g m-2
).

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-72-
4.2.5. Frond water relations
Predawn frond water potential (Ψ predawn) of each tree fern was measured using a pres-
sure chamber (PMS Corvallis, OR, USA) at the end of each of the three periods.
Concentrations of osmotically active solutes (osmolality) were measured via freeze-
point depression from fresh sap extracts using an OSMOMAT 030 cryoscopic osmome-
ter (Gonotec, Berlin, Germany), nomenclature by De Costa et al. (2007).
Relative water content (RWC) was determined as follows: pinnae were detached and
weighed (fresh weight), then floated in water for ca 5 h in the dark to reach full hydra-
tion. Pinnae were blotted dry with tissue paper and weighed (saturated weight), then
dried at 60ºC for 48 h and weighed again (dry weight). RWC (%) was calculated as:
(fresh weight – dry weight)/(saturated weight – dry weight) x 100.
4.2.6. Stable isotope analysis
Due to rapid turnover of the soluble carbon pool, we tracked differences in stable iso-
tope composition (δ13
C) of a hot water extract (e.g. Warren et al. 2007). Hot water ex-
tracts were prepared according to Callister et al. (2006). Extract sub-samples (100 µl)
were dried in tin capsules at 50ºC for 48 h, and δ13
C determined by IRMS (EuroVector,
IsoPrime Mass Spectometer, Manchester, UK) with Dumas flash combustion. Analysis
of δ13
C (in ‰ units) was against a tertiary standard (Acetanilide: δ13
C =33.44‰ and
C=71.09%), which was calibrated against a PDB (Pee Dee Belemnite) standard.
4.2.7. Relative extractable soil water, REW
Relative extractable soil water (REW), was calculated as the soil moisture content at
each day of water deficit divided by soil moisture content at field capacity minus soil
moisture content at permanent wilting point (after Bogeat-Triboulot et al. 2007). For

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-73-
this, each pot of water deficit treated plants was weighed at field capacity and at the end
of each day of water withheld. Potential differences in evaporation from the pot surface
between light regimes was taken into account by placing a plant-free pot in each of the
light regimes and weighed at the same time as treated plants (Merchant et al. 2006). Soil
moisture content at field capacity was determined gravimetrically by oven drying soils
at 105ºC for 24 h.
4.2.8. Statistical analysis
Effects of species (D. antarctica, C. australis), light (moderate, high), water treatment
(control, deficit), and their interactions on dependent variables were analysed using the
general linear models of SPSS (SPSS Inc. Chicago, USA). Variables were graphically
checked for deviations from normality, homogeneity of variances was tested using
Levene‟s test, and means and variances were not correlated across treatments. Each pe-
riod (pre-treatment, water deficit, rewatering) was analysed separately.
DOY
42 45 48 51 54 57 60 63 66 69 72 75 78 81 84 87 90 93 96
Ma
xim
al d
aily
air t
em
pe
ratu
re,o
C
0
5
10
15
20
25
30
35
40
45
50
55
Da
ily R
H a
t 9
a.m
., %
0
20
40
60
80
100Pre-treatment Drought RewateringRewatering
(extra shadeon high light plants)
(shade removal)
Fig. 4.1 Weather conditions during the experiment. Dotted grey line is maximum daily
air temperature (ºC), black line - daily relative humidity (RH, %) at 9 a.m. Vertical lines
indicate stages of the experiment for Block 1.

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-74-
4.3. Results
4.3.1. Maximum quantum yield of PS II (Fv/Fm)
Even during the pre-treatment period, both species had lower Fv/Fm than what is consid-
ered an optimum value for healthy fronds (0.83, Maxwell and Johnson 2000, Table 4.1).
Fv/Fm did not differ between species but was significantly higher (P<0.001) in moderate
light plants during the pre-treatment and the water deficit periods (Table 4.1). Fv/Fm was
not affected by water treatment in the water deficit period regardless of the light regime,
but was significantly greater in control than deficit plants in the rewatering period.
Drought stressed D. antarctica in moderate light had significantly lower Fv/Fm in the
rewatering period (Table 4.1).
4.3.2. Photosynthetic capacity
Light saturated net photosynthesis (Amax) and stomatal conductance (gs) at Amax were
significantly greater in C. australis than D. antarctica in the pre-treatment and the water
deficit periods, but not the rewatering period (Fig. 4.2). Water deficit resulted in a sig-
nificant reduction in Amax and gs in both species (water treatment P<0.001), with mar-
ginally higher, albeit non-significant, values under high light. Amax and gs recovered
with rewatering in both species, and under both light regimes (Fig. 4.2).
The maximum carboxylation rate, Vcmax, and the maximum rate of electron transport,
Jmax, did not differ significantly between species and light regimes in the pre-treatment
period (Table 4.1). However, water deficit caused significant decreases in both variables
for both species – an effect that may have been most pronounced for D. antarctica un-
der moderate light, although increasingly inaccurate Ci estimates at very low gs made
impossible to construct meaningful A-Ci curves (i.e. missing estimates for Vcmax Jmax;
Table 4.1). Vcmax and Jmax recovered to near control values with rewatering for both spe-
cies, although Jmax was significantly lower in C. australis in this period (Table 4.1).

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-75-
Total chlorophyll content per frond area (µmol m-2
) did not significantly differ between
species, light regimes, and water treatments across all experimental periods (Table 4.1).
Chlorophyll a/b ratios were significantly greater in D. antarctica than C. australis in the
pre-treatment period (P=0.01, Table 4.1). However, this difference was not detected for
the remainder of the experiment, with chlorophyll a/b ratios of both species largely in-
variant to light regimes and water treatments (Table 4.1).
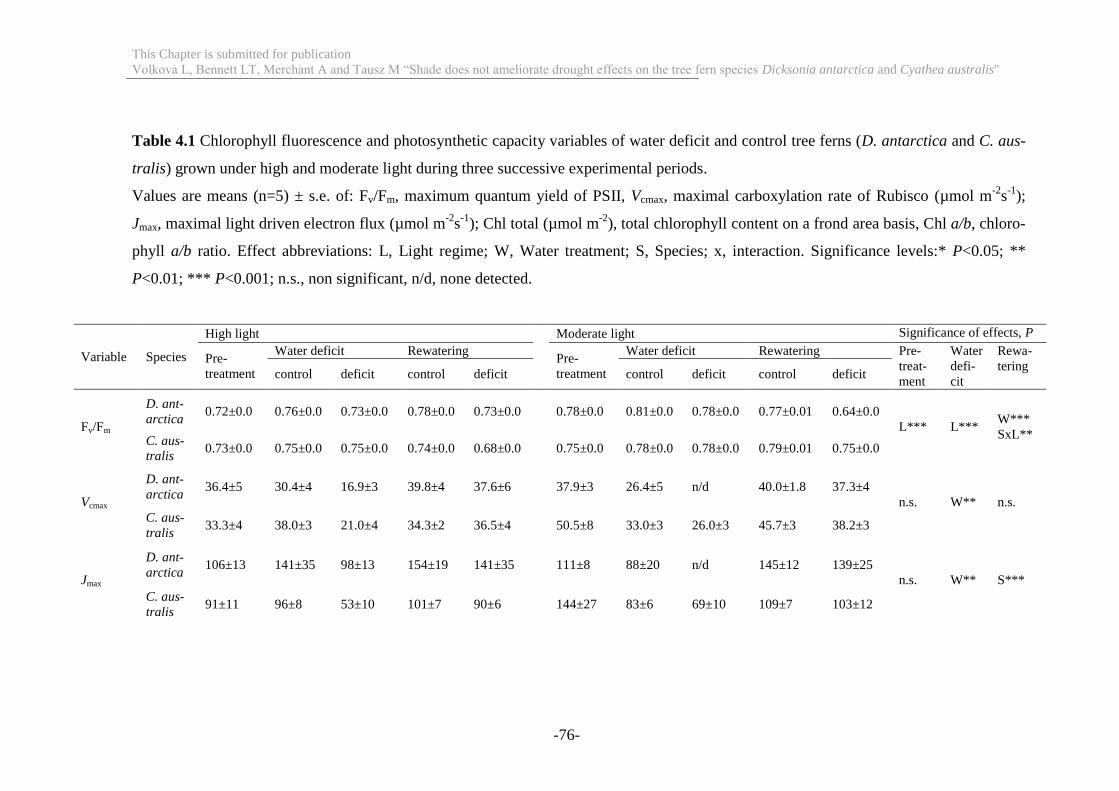
This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the tree fern species Dicksonia antarctica and Cyathea australis"
-76-
Table 4.1 Chlorophyll fluorescence and photosynthetic capacity variables of water deficit and control tree ferns (D. antarctica and C. aus-
tralis) grown under high and moderate light during three successive experimental periods.
Values are means (n=5) ± s.e. of: Fv/Fm, maximum quantum yield of PSII, Vcmax, maximal carboxylation rate of Rubisco (µmol m-2
s-1
);
Jmax, maximal light driven electron flux (µmol m-2
s-1
); Chl total (µmol m-2
), total chlorophyll content on a frond area basis, Chl a/b, chloro-
phyll a/b ratio. Effect abbreviations: L, Light regime; W, Water treatment; S, Species; x, interaction. Significance levels:* P<0.05; **
P<0.01; *** P<0.001; n.s., non significant, n/d, none detected.
Variable Species
High light Moderate light Significance of effects, P
Pre-
treatment
Water deficit Rewatering Pre-
treatment
Water deficit Rewatering Pre-
treat-
ment
Water
defi-
cit
Rewa-
tering control deficit control deficit
control deficit control deficit
Fv/Fm
D. ant-
arctica 0.72±0.0 0.76±0.0 0.73±0.0 0.78±0.0 0.73±0.0 0.78±0.0 0.81±0.0 0.78±0.0 0.77±0.01 0.64±0.0
L*** L*** W***
SxL** C. aus-
tralis 0.73±0.0 0.75±0.0 0.75±0.0 0.74±0.0 0.68±0.0 0.75±0.0 0.78±0.0 0.78±0.0 0.79±0.01 0.75±0.0
Vcmax
D. ant-
arctica 36.4±5 30.4±4 16.9±3 39.8±4 37.6±6 37.9±3 26.4±5 n/d 40.0±1.8 37.3±4
n.s. W** n.s.
C. aus-
tralis 33.3±4 38.0±3 21.0±4 34.3±2 36.5±4 50.5±8 33.0±3 26.0±3 45.7±3 38.2±3
Jmax
D. ant-
arctica 106±13 141±35 98±13 154±19 141±35 111±8 88±20 n/d 145±12 139±25
n.s. W** S***
C. aus-
tralis 91±11 96±8 53±10 101±7 90±6 144±27 83±6 69±10 109±7 103±12

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the tree fern species Dicksonia antarctica and Cyathea australis"
-77-
Variable Species
High light Moderate light Significance of effects, P
Pre-
treatment
Water deficit Rewatering Pre-
treatment
Water deficit Rewatering Pre-
treat-
ment
Water
defi-
cit
Rewa-
tering control deficit control deficit
control deficit control deficit
Chl total
D. ant-
arctica 382±30 277±14 218±12 288±12 219±13 372±31 275±40 253±31 251±36 268±25
n.s. n.s. n.s. C. aus-
tralis 349±15 285±66 242±18 252±14 222±23 354±23 256±13 219±18 328±65 261±25
Chl a/b
D. ant-
arctica 4.0±0.1 3.5±0.1 3.4±0.2 3.5±0.1 3.5±0.2 4.1±0.1 3.4±0.2 3.4±0.2 3.4±0.1 3.5±0.2
S** n.s. n.s. C. aus-
tralis 3.9±0.1 2.8±0.3 3.4±0.1 3.4±0.1 3.0±0.4 3.8±0.1 3.4±0.1 3.2±0.2 3.0±0.3 3.5±0.1

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the tree fern species Dicksonia antarctica and Cyathea australis"
-78-
Period
pre-treatment water deficit rewatering
Am
ax (
mol m
-2s-1
)
0
2
4
6
8
10
12
pre-treatment water deficit rewatering
High lightgs (
mm
ol m
-2s-1
)
0
50
100
150
200
250
300Moderate light
Period
S* S***W**
n.s.
Pre-treatment
Waterdeficit
Re-watering
S* S**W**
n.s.
Pre-treatment
Waterdeficit
Re-watering
Fig. 4.2. Stomatal conductance and light saturated net photosynthesis of water deficit (hatch) and control (solid) D. antarctica (white bars)
and C. australis (grey bars) under high and moderate light in three successive experimental periods (pre-treatment, water deficit and rewa-
tering). Values are means n=5 (± s.e.). Effect abbreviations: S, Species; W, Water treatment; none of interactions were significant. Signifi-
cance levels:* P<0.05; ** P<0.01; *** P<0.001; n.s., non significant

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-79-
4.3.3. Frond survival
Approximately 1/3 of fronds of water deficit plants survived the water deficit period – a
proportion that did not significantly differ between species and light regimes (high light:
33±7% for D. antarctica and 24±4% for C. australis; moderate light: 27±4% for D.
antarctica and 35±6% for C. australis).
4.3.4. Time course of stomatal conductance during 5 days without water
Stomatal response to withholding water did not differ between species but was affected
by light regime (Fig. 4.3). Water deficit plants under high light decreased gs by almost
50% relative to controls after one day without water, at relative extractable soil water
(REW) of ca 80% (Fig. 4.3). In contrast, plants under moderate light maintained initial
stomatal conductance for 1 day longer and decreased gs at lower REW (ca 65%;
Fig.4.3).

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-80-
REW, %
204060801000
20
40
60
80
100
Days without water
day 1day 2
day 3day 4
day 5
High light
gs (
% o
f contr
ols
)
0
20
40
60
80
100
Moderate light
5 days of water withheld
REW,%
20406080100
Days without water
day 1day 2
day 3day 4
day 5
C. australis C. australis
D. antarctica D. antarctica
gs control gs control
gs controlgs control
Fig. 4.3. Time course of stomatal conductance of water deficit D. antarctica and C. aus-
tralis grown under high and moderate light during first five days of water deficit. Where
gs, stomatal conductance, and REW, relative extractable soil water. Values are means
(n=5) ± s.e.

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-81-
4.3.5. Frond water relations
Significant differences in frond water relations were detected between species in the
pre-treatment period. D. antarctica had significantly lower Ψ predawn, RWC and signifi-
cantly greater osmolality (Table 4.2). With the exception of Ψ predawn, these species dif-
ferences were not detected during the water deficit period, although the difference in
osmolality was reinstated after rewatering.
Frond water relations of both species were significantly affected by water treatments.
Water deficit significantly decreased Ψ predawn and RWC of deficit relative to control
plants of both species under both light regimes (Table 4.2). These effects were reversed
in the rewatering period. Osmolality was not affected by water deficit; however, a sig-
nificant species by water treatment interaction was detected in the rewatering period
(i.e. increased osmolality of control D. antarctica relative to water deficit plants, and the
opposite trend in C. australis; Table 4.2).
Light regimes did not significantly affect frond water relations of either species, apart
from incomplete Ψ predawn recovery of deficit D. antarctica under moderate light in the
rewatering period (Light x Water; P=0.04; Table 4.2).

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the tree fern species Dicksonia antarctica and Cyathea australis"
-82-
Table 4.2. Frond water relations of water deficit and control tree ferns (D. antarctica and C. australis) grown under high and moderate
light during three successive experimental periods. Values are means (n=5) ± s.e. of: Ψ predawn, predawn frond water potential (MPa); osmo-
lality (osmol kg-1
); RWC, relative water content (%). Effect abbreviations: L, Light regime; W, Water treatment; S, Species; x, interaction.
Significance levels:* P<0.05; ** P<0.01; *** P<0.001; n.s., non significant.
Variable Species
High light Moderate light Significance of effects, P
Pre-
treatment
Water deficit Rewatering
Pre-
treatment
Water deficit Rewatering Pre-
treat-
ment
Wa-
ter
defi
cit
Re-
watering
control deficit control deficit
control deficit control deficit
Ψ predawn
D. antarc-
tica -0.3±0.0 -0.7±0.2 -1.3±0.2 -0.3±0.1 -0.3±0.1 -0.3±0.0 -0.5±0.1 -2.5±0.3 -0.2±0.1 -0.4±0.1
S ** S *
W*
**
LxW*
C.
australis -0.2±0.0 -0.5±0.0 -1.2±0.4 -0.4±0.1 -0.3±0.0 -0.2±0.0 -0.5±0.0 -1.3±0.4 -0.2±0.0 -0.3±0.1
Osmolality
D. antarc-
tica 0.69±0.0 0.62±0.1 0.61±0.0 0.79±0.1 0.68±0.0 0.71±0.0 0.60±0.0 0.68±0.1 0.79±0.0 0.72±0.0
S** n.s. S***
SxW**
C.
australis 0.64±0.0 0.63±0.0 0.60±0.0 0.56±0.0 0.66±0.0 0.62±0.0 0.58±0.0 0.62±0.0 0.63±0.0 0.65±0.0
RWC
D.
antarctica 95±0 94±1 92±2 97±0 96±1 94±1 95±2 93±1 96±0 96±1
S* W* n.s.
C. aus-
tralis 97±1 97±1 89±3 97±0 97±1 97±0 94±1 90±5 97±1 94±1

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-83-
4.3.6. Intrinsic water use efficiency (calculated as Amax/gs, WUEi) and stable
carbon isotope composition (δ13
C)
During the pre-treatment period, WUEi was significantly greater in D. antarctica and
was not affected by light regime. Water deficit treatment significantly increased WUEi
of deficit relative to control plants irrespective of the light regime and species. With re-
watering, WUEi was greater in C. australis than D. antarctica (Fig. 4.4).
δ13
C was significantly more negative in C. australis across the experiment (Fig. 4.4).
During water stress period, δ13
C of deficit plants under high light were comparable with
pre-treatment values, but δ13
C became more negative in control plants under moderate
light, particularly in D. antarctica (Light x Water interaction, P=0.02; Fig. 4.4). This
effect was not detected in the rewatering period, when δ13
C within species was compa-
rable across water treatments and light regimes (Fig. 4.4).

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the tree fern species Dicksonia antarctica and Cyathea
australis"
-84-
Moderate light
Period
Pre-treatment water deficit rewatering
WU
Ei (
mol m
ol-1
)
0
20
40
60
80
100
Period
Pre-treatment water deficit rewatering
High light
13C
(m
mol m
-2s-1
)
-30
-25
-20
-15
-10
-5
S * W ** S *
Pre-treatment
Waterdeficit
Re-watering
S *** S ***W **L x W *
S*
Pre-treatment
Waterdeficit
Re-watering
Fig. 4.4. Stable isotope composition, δ13
C and intrinsic water use efficiency, WUEi and of water deficit (hatch) and control (solid) D.
antarctica (white bars) and C. australis (grey bars). Values are means (n=5); ± s.e. Effect abbreviations: S, Species; W, Water treat-
ment; L, Light regime; L x W, light by water treatment interaction. Significance levels:* P<0.05; ** P<0.01; *** P<0.001

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-85-
4.4. Discussion
4.4.1. Pre-treatment period – species differences and effect of light
Photosynthetic capacity of both species were within range for tree fern species reported
in the literature: 6 – 10.8 µmol m-2
s-1
(Nobel et al. 1984) and 8.3 µmol m-2
s-1
(Hunt et
al. 2002) for D. antarctica, or even similar to some middle storey canopy species in
these types of forests, e.g. Nothofagus cunninghamii (Tausz et al. 2005). Maximum car-
boxylation rates, Vcmax (the maximal in vivo Rubisco activity), and the maximum rate of
electron transport, Jmax at the reference temperature of 25°C were within the lowest val-
ues among the large number of species reviewed by Wullschleger (1993).
D. antarctica had greater WUEi that together with its less negative δ13
C in the pre-
treatment period suggests that photo-assimilation may be comparatively less susceptible
to changes in water status.
Chlorophyll a/b ratios were also significantly higher in D. antarctica, a result inconsis-
tent with this species‟ putative preference for well shaded microhabitats. Normally,
chlorophyll a/b ratios are lower in shade acclimated than sun foliage because of greater
chlorophyll b contents (Hoober et al. 2007). For D. antarctica, we observed greater
chlorophyll a content while b was comparable between species (data not shown). How-
ever, our study involved young sporophytes and it must be noted that these species dif-
ferences might change at later stages in the tree fern life cycle.
Both species displayed low acclimation potential to changes in light regime - neither
photosynthetic capacity (except Fv/Fm) nor water relation variables were affected by a
nearly two-fold difference in maximum irradiance. Reduction in Fv/Fm usually indicates
down-regulation of PSII (i.e. photoinhibition, Savitch et al. 2000), yet because Fv/Fm
was measured at predawn, the down-regulation should be relaxed, indicating either
damage of PSII or sustained down-regulation of some type. Down-regulation or damage
of PSII in high light plants may affect maximum light use efficiency of photosynthesis,
and likely depress photosynthetic performance - yet effects of light regimes on Amax
were non significant, suggesting small differences in Fv/Fm under moderate and high

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-86-
light had minimal effect on Amax. These differences would become more relevant for
photosynthesis during low light periods and therefore integrated carbon gain (Zhu et al.
2004) – an issue not addressed in this study.
4.4.2. Water deficit and light interactions
Both species were susceptible to short-term water stress. Deeper shade did not amelio-
rate the negative effects of water deficit, and there is no evidence to suggest that water
deficit in combination with high light was more detrimental to plant function. The ab-
sence of significant water treatment by species interactions indicates that both species
responded to water deficit in a similar way. The only parameter insensitive to water
deficit was Fv/Fm - while this is in agreement with some studies (e.g. Epron and Dreyer
1992), others showed that drought reduced predawn Fv/Fm more in shade than in sun
(Valladares and Pearcy 2002). In our study, plants under moderate light had consistently
higher predawn Fv/Fm than high light plants even under water deficit, indicating full re-
covery of PSII overnight.
We detected contrasting stomatal behaviour under the different light regimes - plants
growing under moderate light closed stomata later and at lower relative extractable soil
water content. This indicated slower stomatal response under lower light, a result con-
sistent with the findings of Roberts et al. (1984) where stomata of bracken fern from the
most shaded understorey level were less sensitive to soil moisture treatments than those
from more irradiated levels. Surprisingly, even at REW of 30%, stomatal conductance
was not close to zero, possibly indicating limited stomatal control or high residual cu-
ticular conductance (which was not distinguished from stomatal conductance in our
measurements).
Decreases in gs by ca 70%, relatively to the pre-treatment values indicated that stress
was rather severe – the result we aimed at. However, a straightforward definition of
drought stress severity as given by Flexas and Medrano (2002) – that is, moderate stress
intensity at gs <150 and severe at <50 mmol H2O m-2
s-1
– appeared to be unsuitable in
our study. Ferns seem to have generally low stomatal conductance (Doi et al. 2006,
Hunt et al. 2002) and in our study, gs of D. antarctica rarely exceeded 150 mmol H2O

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-87-
m-2
s-1
even with adequate watering. The severity of drought stress was also confirmed
by the decrease in mesophyll capacity parameters of photosynthesis - both Vcmax and
Jmax decreased significantly after ten days of drought, confirming findings by Bota et al.
(2004) that impaired Rubisco activity and/or RuBP regeneration do not limit photosyn-
thesis until drought is severe and A and gs are strongly depressed.
Reduced values of gs and Amax in D. antarctica under moderate light resulted in in-
creased error in the computation of Ci, therefore it was not possible to accurately con-
struct A-Ci curves for these plants and measure Vcmax and Jmax values. This precluded a
formal significance test of the result. D. antarctica under moderate light also had greater
decreases in Ψ predawn which most probably reflect conditions close to the point of turgor
loss in some fronds, and it can be speculated that D. antarctica under moderate light
was most severely affected by water deficit treatment, possibly as a consequence of
greater water loss due to the slower stomatal closure under moderate light.
RWC can affect Rubisco activity and photochemistry of plants in response to drought.
According to Flexas et al. (2006) Rubisco activity remains essentially unaffected by wa-
ter stress until gs drops below 50 mmol H2O m-2
s-1
regardless of species. While our re-
sults on D. antarctica support this suggestion, gs of C. australis varied from less than 50
to above 100 mmol H2O m-2
s-1
, yet Rubisco activity (measured via Vcmax) decreased
significantly. Despite a significant decrease in RWC under water deficit, RWC of both
species remained around 90%. This corresponds to ca 30% of the cases reviewed by
Flexas and Medrano (2002), where decrease in Rubisco activity was associated with
high RWC between 90-100%. Possibly, our inability to detect significant osmotic ad-
justment in this study suggests that RWC is maintained in these species by a combina-
tion of alternative factors. High solute levels in frond tissues may provide „pre-emptive‟
protection against rapid changes in water status, a concept that would require additional
investigation given the lack of data on tree fern chemistry and physiology. If water con-
servation measures such as early and rapid stomatal closure fail, fronds may be left
without strong tissue level tolerance mechanisms. In our study, this was indicated by
necroses of two-thirds of fronds by the end of the water deficit period. In agreement

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-88-
with their ecological distribution, tree ferns may be unable to tolerate a major drop in
RWC instead relying on an „avoidance‟ strategy of frond death.
We observed significant increases in WUEi under severe water stress in line with the
stomatal responses. This was accompanied by significant reductions in photosynthetic
activity. For carbon isotopic composition, the general lack of significant changes be-
tween treatments may be due to low photosynthetic activity limiting the contribution of
new assimilates to the water extractable pool (Dawson et al. 2002). The observed sig-
nificant δ13
C shift in control plants under moderate light during the rewatering period
(when air temperatures increased for ca 10ºC) could be related to stomatal closure in
response to high air temperature while Rubisco remained active. Indeed, our other study
of heat and light interactions indicate that moderate light may have mediated effects of
heat on control plants (Volkova et al. 2009). An increase in air temperature can also ex-
plain relatively large changes for some variables within a species between pre-treatment
and control (Fv/Fm, Ψ predawn, RWC etc).
Overall, there were some indications that shade exacerbated drought stress effects in D.
antarctica (i.e. mesophyll capacity parameters, Ψ predawn). These findings are consistent
with results elsewhere indicating that shaded plants can be more susceptible to drought,
although the mechanisms explaining such an effect in other studies – such as increased
below-ground competition, greater reduction in predawn Fv/Fm (Valladares and Pearcy
2002) – were apparently not applicable to our study.
4.4.3. Rewatering period
Both species of tree fern recovered from short-term water deficit with rewatering (i.e.
treatment effect was non significant at this stage for all parameters but Fv/Fm). Recovery
of plants was mostly unaffected by light regime, however, due to an unexpected heat
wave we had to put an additional shade protection over high light plants, which resulted
in rather small differences in PPFD between light regimes during first 10 days of rewa-
tering, and probably smoothed potential differences during recovery.

This Chapter is submitted for publication
Volkova L, Bennett LT, Merchant A and Tausz M “Shade does not ameliorate drought effects on the
tree fern species Dicksonia antarctica and Cyathea australis"
-89-
Decreases in Fv/Fm in the rewatering period were presumably mediated by further in-
crease in ambient air temperatures (Galle et al. 2007). We speculate that the greater de-
crease in Fv/Fm of deficit plants was an after-effect of the treatment, which may have
made them more sensitive to the following conditions.
Species differences in Ψ predawn, RWC, Amax, gs, chlorophyll a/b during the pre-treatment
period were no longer evident in the rewatering period. This could be partly explained
by effects of more extreme temperatures combined with different cohorts of fronds, but
might also reflect some subtly different responses in frond physiology between the two
species triggered by the treatments.
4.5. Summary
D. antarctica and C. australis displayed contrasting physiological characteristics some-
times contrary of what would have be expected from species‟ different origin and mi-
cro-site preferences - greater WUEi, chlorophyll a/b ratios and less negative δ13
C for D.
antarctica would indicate this species as more drought and light tolerant. Results of our
study also suggest that shade does not ameliorate drought effects on C. australis and D.
antarctica. However, both species were resilient to short-term severe water stress, their
ability to restore physiology of surviving fronds and re-sprout after considerable loss of
fronds undoubtedly plays a role for both species exposed to more extreme changes in
environmental conditions. These findings are consistent with their distribution in tem-
perate forest systems that are subject to seasonal droughts and rapid changes in forest
canopy structure.

-90-

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-91-
Chapter 5. Seasonal variations in photosynthesis of the tree ferns
Dicksonia antarctica and Cyathea australis in wet sclerophyll forests
of Australia
(iv) Abstract
Steady state and dynamic responses of two tree fern species of contrasting origins,
Dicksonia antarctica (Gondwanan) and Cyathea australis (Pan-tropical), were studied
over two consecutive years under field conditions in wet sclerophyll forest of south-
eastern Australia. Irrespective of their different origins and micro-site preferences, there
were no significant differences in photosynthetic performance between the two species.
Growth irradiance (from open sites to dense canopy cover) had very little effect on pho-
tosynthetic rates of the tree ferns. Both species performed better in winter than in sum-
mer, when photosynthetic rates reached higher values under similar irradiance and leaf
temperatures. However, at the same leaf temperature, Fv/Fm was significantly lower in
winter than in summer, suggesting some cold-induced limitation in PSII efficiency indi-
cating persistent photoinhibition associated with cold winter mornings. Both species
displayed seasonal acclimation in a number of measured photosynthetic parameters and
frond traits. Acclimation of stomatal density to spatial variation in growth irradiance
seemed limited in both species, although stomatal pattern differed between species. Be-
cause there were no significant differences between the two species in photosynthetic
parameters, both species could be described by the same carbon gain and water use
models at the leaf-scale.
5.1. Introduction
The tree ferns Dicksonia antarctica (Labill) and Cyathea australis (R. Br.) Domin are
well known Australian representatives of the fern genera Cyathea and Dicksonia, which
include most tree fern species worldwide. Cyathea has a broad global distribution,

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-92-
whereas Dicksonia is diverse throughout Indonesia and New Guinea, and can be found
in isolated pockets including off the coast of Chile (Large and Braggins 2004).
Little is known about the photosynthesis and stomatal conductance of D. antarctica and
C. australis in their natural habitats. This knowledge gap effectively excludes tree ferns
from long-term carbon uptake and energy flux calculations, and limits the validity of
predictions of their survival under climate change. Yet, these tree ferns often form the
dominant understorey component of wet sclerophyll forests in south-eastern Australia
(Ough and Murphy 1996). These forests support as much as 1,053 tonnes carbon ha-1
in
above-ground living biomass (Keith et al. 2009), although the contribution of tree ferns
remains unknown. Tree ferns of the genus Cyathea account for 33% of above-ground
biomass in dwarf forests of Puerto Rico (Weaver 2008). Understorey species of ever-
green forests world wide contribute an average of 49% to ecosystem respiration (Mis-
son et al. 2007).
D. antarctica and C. australis are species of great ecological importance (Lindenmayer
et al. 1994, Roberts et al. 2005). Both species are broadly distributed in the wetter parts
of south-eastern Australia, with D. antarctica dominating wet, shady gullies, while C.
australis can also extend to forest margins (McCarthy 1998). Although D. antarctica
and C. australis can grow together in wet sclerophyll forests, previous studies have in-
dicated they have different micro-habitat preferences. Dignan and Bred (2003) found
greater probability of a tree fern being C. australis than D. antarctica with increasing
distance from a stream, suggesting greater drought tolerance of C. australis. The postu-
lated difference in origin of the two species – Gondwanan for D. antarctica compared
with pan-tropical for C. australis (Page and Clifford 1981) – also suggests different
physiological adaptation potential consistent with their observed distribution patterns
within forests.
During their lifetime, tree ferns of south-eastern Australia can be periodically exposed
to the harsh conditions of post-wildfire environments, characterised by increased irradi-
ance and leaf temperatures, and consequently stronger evapotranspiration and water

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-93-
loss. Sensitivity to high temperatures, high irradiance and decreased water could poten-
tially limit growth and distribution of tree ferns in the field, although there are few field
data to evaluate this theoretical model.
Ferns, as other plants (angiosperms or gymnosperms), have displayed a capacity to ac-
climate to changing irradiance. For example, leaf characteristics (frond surface area,
epidermis thickness, palisade/spongy mesophyll ratio, blade size, petiole length) of a
South American Cyathea species were found to be correlated with local irradiance
(Arens 1997). However, stomatal density, which might be expected to increase with in-
creasing light intensity (Casson and Gray 2008), was found to have limited plasticity in
the tree fern Cyathea caracasana in response to light environment (Arens 1997).
To evaluate relationships between physiological performance of the tree ferns and pre-
vailing environmental conditions, we analysed diurnal and seasonal trends in gas ex-
change during summer and winter over two consecutive years using pairs of D. antarc-
tica and C. australis at contrasting micro-habitats (shaded creek-side and more exposed
rocky knoll). The objectives of this study were:
To determine if D. antarctica and C. australis showed differences in morphologi-
cal and physiological frond traits among contrasting micro-habitats in the field.
The postulated difference in their origin in particular leads us to hypothesise that
the species would have different seasonal acclimation to temperature, with D.
antarctica having higher photosynthesis in winter, and C. australis favouring
summer;
To determine whether light, temperature and plant water status limited physio-
logical performance in the field;
To examine acclimation of stomatal density in the two tree ferns; that is, would
stomatal density differ between the two species and/or between high light exposed
and shaded habitats?

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-94-
5.2. Materials and methods
5.2.1. Study site and sampling design
Our study site was established in mountain ash (Eucalyptus regnans F. Muell) forest in
the Victorian Central Highlands (145‟42ºE 37‟35ºS, elevation 450 m). The average an-
nual rainfall of a representative weather station (Healesville, 145,53ºE 37.68ºS, eleva-
tion 131 m) is 1021 mm. October is the wettest (106 mm) and January is the driest (58
mm) month on average. Mean maximum temperature of the hottest month (February) is
26ºC and mean minimum of the coldest month (July) is 4ºC (Australian Bureau of Me-
teorology, 2009 www.bom.gov.au/climate /averages/cdo/about/about-stats.shtml, veri-
fied August 2009).
The experimental design involved tree ferns on the margins of a logging coupe that was
clearcut in late 2003 and early 2004, then slash-burnt in March 2004. The coupe is
bounded by the Acheron River to the south, and a creek to the east. Retained undis-
turbed vegetation buffer zones were maintained along the waterways in accordance with
the Code of Forest Practices (Department of Natural Resources and Environment,
1996), and were 200 m wide along the Acheron River and 20 m along the creek.
We selected eight mature individuals of each of two species, Dicksonia antarctica (La-
bill.) and Cyathea australis (R.Br.) Domin. To cover the full range of local environ-
mental conditions evenly for both species, tree ferns were selected in pairs (one from
each species) growing in close vicinity (Fig. 5.1). Such pairs were selected from the
most exposed sites at the fringe of the clearing on a rocky knoll (elevation up to 490 m
above sea level, proximity to the creek 49 - 100 m) to the most sheltered sites in buffer
zones (average elevation 450 m) near waterways (ca 2 m).

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-95-
Fig. 5.1. Location of the tree ferns at the study area. Where D is D. antarctica and C. is
C. australis
5.2.2. Tree fern measurement schedule
Each plant was measured for pre-dawn chlorophyll a fluorescence, predawn/midday
water potential, gas exchange measurements (diurnal and maximal net CO2 assimilation
rate Amax) in winter (August-September) and summer (December-January) over two
consecutive calendar years (2006-2008). Gas exchange measurements were made over
two consecutive days during each season per year. Diurnal measurements were made
once in summer (December 2006) and winter (August-September 2007). Frond samples
for nitrogen and chlorophyll content were collected at the same time as gas exchange
measurements. All measurements were made on the mid-third of the youngest fully ex-
panded fronds of similar north-facing orientation.
Road
Creek
C C
C
C
C; D
C
C C
D D
D
D
D
D
D
Unlogged area

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-96-
Samples for stomatal density determination were collected in January 2007. For this
analysis only, fronds that were most representative of the tree fern‟s light environment
were collected; that is, most light-exposed fronds of tree ferns growing in the open, and
most shaded fronds of shade-grown tree ferns. Orientation (e.g. north facing) was not
taken into account.
5.2.3. Mean irradiance on measured fronds
Mean daily photosynthetic photon flux density (daily PPFD) was estimated from hemi-
spherical photographs (Nikon 601 F-601, Japan, fisheye lens), which were taken from
the position of each measured frond. Black and white negatives were scanned and
evaluated using Winphot software (ter Steege 1996). Calculated irradiance (sum-
mer/winter) ranged from 40/17 mol m-2
d-1
for a tree fern growing in open habitat to
13/3 mol m-2
d-1
for a tree fern growing under dense canopy near the creek.
5.2.4. Maximal quantum yield of photochemistry (Fv/Fm)
Maximal quantum yield of photochemistry (Fv/Fm) was measured at predawn (5 a.m. in
summer, 6 a.m. in winter) with a pulse modulated fluorometer (OS-30p, Opti-Sciences,
Hudson, USA). Ground fluorescence (F0) was obtained with a low intensity modulated
light (600 Hz, 650nm, PPFD<1 µmol m-2
s-1
). Maximum fluorescence (Fm) was induced
by a saturating flash. Maximum efficiency of photosystem II (PSII) was estimated as
Fv/Fm = (Fm–F0)/Fm, nomenclature after Maxwell and Johnson (2000).
5.2.5. Gas exchange measurements
Light-saturated rates of net photosynthesis (Amax) and gs at Amax were measured using a
Li-Cor 6400 portable photosynthesis measurement system, equipped with a 2x3 cm
broadleaf chamber with red-blue LEDs (Li-Cor, Lincoln, Nebraska, USA). PPFD was
1500 µmol m-2
s-1
(determined as saturating by preliminary light response curves), air
flow rate was 400 µmol air s-1
, reference CO2 concentration 400 µmol mol-1
, and leaf

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-97-
temperature and relative humidity were kept at ambient values. The frond area enclosed
in the Li-Cor chamber was marked, photographed and calculated using imaging soft-
ware (UTHSCSA Image Tool Version 3, University of Texas, USA).
Diurnal courses of gas exchange under ambient conditions were measured using the Li-
Cor transparent chamber. Air/leaf temperature and water pressure deficit based on leaf
temperature (VPD) were also recorded.
5.2.6. Frond water potential
Predawn frond water potential was chosen as a measure of plant water status, because it
is an appropriate estimate for the water availability in the soil reached by the roots
(Jones 1992).
Predawn (Ψ predawn) and midday (Ψ midday) frond water potentials were measured using a
pressure chamber (PMS Corvallis, OR, USA) at predawn 5 a.m./6 a.m. and midday 12
p.m./1 p.m. (summer/winter, respectively).
5.2.7. Frond traits
Specific leaf area (SLA) was calculated as the ratio of frond area over frond dry weight
(m2 kg
-1 dry weight). Fresh frond samples were collected, the frond area scanned and calcu-
lated using Scion Image software (Scion Corporation 2000-2001, USA), and frond ma-
terial was then dried at 60ºC for 48 h for dry weight. Different cohorts of fully-
expanded fronds produced in summer and winter, were used for calculation of SLA.
Frond samples were dried as described above and ground to a fine powder, then ana-
lysed for total nitrogen and carbon content using an elemental analyser (LECO CHN-
1000, Michigan, USA). Photosynthetic nitrogen use efficiency (PNUE) was calculated
as Amax divided by frond nitrogen content (on a frond area basis).

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-98-
For measurements of frond chlorophyll content, four frond discs (diameter of 3.75 mm
each) were collected, immediately immersed in liquid nitrogen, and stored at -80ºC until
extraction. Chlorophyll a and b were extracted using 1.8 ml of 100% dimethyl sulphox-
ide (DMSO). Extracts were heated for 30 min at 65ºC in a dry block heater Termoline
L+M (Northgate, Queensland, Australia). The supernatant was then transferred to a
spectrophotometer Carry 300 (Varian, The Netherlands). A blank of pure DMSO was
used to calibrate the spectrophotometer at zero absorbance. Chlorophyll a, b and total
were calculated according to Wellburn (1994).
Fresh frond material (from 3-4 fronds per tree fern) was analysed for stomatal density
by variable pressure scanning electron microscopy (VP-SEM, model Leo 1450 VP; Leo
Electron Microscopy Inc., NY, USA). A flat cross-section of both sides of each frond
was scanned and stomatal density was calculated for four randomly chosen views (giv-
ing 12 to 16 stomatal density estimates per tree fern).
5.2.8. Statistical analysis
A general linear model of SPSS 15 (SPSS Inc. Chicago, USA) was used to analyse ef-
fects of species (D. antarctica, C. australis) and season (summer, winter) on each de-
pendent variable using the following covariates: estimated mean daily PPFD, leaf tem-
perature during gas exchange measurement, Tleaf, and predawn frond water potential Ψ
predawn (as a measure of plant water status).
Every variable was checked for normality using the Shapiro-Wilk test, and log trans-
formed if the assumption of normality was not satisfied. Relationships of covariates
with dependent variables were visually checked for linearity and differences in slopes
(see Fig. 5.2). In this analysis, the effect of season would only be significant if seasonal
differences were over and above the variation explained by seasonal differences in
growth irradiance, temperature, and water status as defined by the covariates.

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-99-
For diurnal course measurements we used boundary-line analysis in a scatter- plot of all
data points of each parameter against the environmental variable in question. Functions
for boundary line analysis were chosen according to González-Rodríguez et al. (2001)
and Larcher (2003), with accuracy of fits evaluated using r2 (in all fits, r
2 >0.85).
5.3. Results
5.3.1. Relationships between photosynthesis, growth irradiance and tempera-
ture
Predawn quantum yield of photochemistry Fv/Fm never decreased below 0.7. It was
negatively related to Tleaf (significant covariate effect in Table 5.1), but did not differ
between the two species. It was significantly higher in summer, despite a negative rela-
tionship with Tleaf, reflecting a flatter slope in their relationship in summer than in win-
ter (Fig 5.2).
Tleaf was positively correlated with the light-saturated rate of net photosynthesis Amax
and stomatal conductance gs, while growth irradiance was significantly correlated to
Amax, but not gs (Table 5.1, Fig. 5.2). Amax and gs were similar for both species (Table
5.1), and were significantly higher in winter than in summer (Fig. 5.2).
Nitrogen content per frond area, NA was correlated with growth irradiance and Tleaf (Ta-
ble 5.1). NA was significantly greater in D. antarctica than in C. australis, and in winter
than in summer (Fig. 5.2). Photosynthetic nitrogen use efficiency PNUE (Amax/NA) did
not vary significantly between species and seasons and was not significantly affected by
any of the tested covariates (Table 5.2).
SLA was inversely related to growth irradiance (Table 5.1). In addition, C. australis
displayed significantly greater specific leaf area, and SLA was significantly greater in
summer than in winter (Fig. 5.2; Table 5.1).

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-100-
None of the covariates had a significant effect on total chlorophyll or chlorophyll a/b
(Table 5.2). Total chlorophyll content (on an area basis) was significantly greater in D.
antarctica and in winter, while chlorophyll a/b ratios were comparable between species
and seasons (Table 5.2).
Table 5.1 Significance of the effect of fixed factors (species and season) and of covari-
ates (ANCOVA) on photosynthetic capacity parameters and frond traits of the tree ferns
D. antarctica and C. australis
Where: Fv/Fm, maximal quantum yield of photochemistry; Amax, light saturated rates of
net photosynthesis; gs, stomatal conductance at Amax; SLA, specific leaf area; NA, frond
nitrogen content on an area basis. Significance levels:*P<0.05; **P<0.01; ***P<0.001;
n.s., non significant; (+) positive relationship; (-) inverse relationship. No interactive
effects were significant
Effect and covari-
ates
Fv/Fm Amax gs SLA NA
Effect
Species n.s.(0.1) n.s (0.9) n.s. (0.1) * *
Seasons *** ** * ** **
Covariate
Mean daily PPFD
(mol m-2
d-1
) n.s. (0.08) (+)** n.s. (0.6) (-)* (+)**
Tleaf (ºC) (-)*** (+)** (+)* n.s. (0.2) (+)*
Ψ predawn (MPa) (+)* n.s. (0.4) n.s. (0.8) n.s. (0.08) n.s. (0.5)

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-101-
Tleaf (oC)
0 10 20 30 40
Fv/F
m
0.68
0.70
0.72
0.74
0.76
0.78
0.80
0.82
0.84
0.86
Daily mean PPFD (mol m-2
d-1
)
0 10 20 30 40
SLA
(m
2kg
-1)
0
5
10
15
20
25
Am
ax (
mol m
-2s-1
)
0
2
4
6
8
10
12
14
16A
max (
mol m
-2s-1
)
0
2
4
6
8
10
12
14
16
gs (
mol m
-2s
-1)
0.00
0.05
0.10
0.15
0.20
0.25
NA (
g m
-2)
0
1
2
3
4
5
6
7
Fig. 5.2. Relationships between photosynthetic capacity parameters and frond traits of
the tree ferns D. antarctica and C. australis and environmental variables
Where: Amax - light saturated rates of net photosynthesis; gs, stomatal conductance at
Amax; Fv/Fm, maximum quantum yield of photosystem II; SLA, specific leaf area; NA,
frond nitrogen content on an area basis. D. antarctica (triangle) and C. australis (circle)
in winter (open symbols) and summer (closed symbols). Regressions are indicated by
dashed lines in summer, and solid lines in winter. All regressions were highly signifi-
cant (P<0.01). See Table 5.1 for significance of effects and relationships.

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dicksonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae)
in wet sclerophyll forests of Australia
-102-
Table 5.2. Photosynthetic capacity and water relation parameters of the tree ferns D. antarctica and C. australis in summer and winter. Values
are means (n = 32) ± s.e. of: PNUE, photosynthetic nitrogen use efficiency, Chl total, total chlorophyll content on a frond area basis; Chl a/b,
chlorophyll a/b ratio; WUEi, intrinsic water use efficiency; Ψ predawn and Ψ midday, predawn and midday frond water potentials. Significance lev-
els: **P<0.01; ***P<0.001; n.s., non-significant; n/a, non applicable; (+) positive relationship (covariates tested were mean daily PPFD, Tleaf, Ψ
predawn). No interactive effects were significant.
Parameter Summer Winter Effects
D. antarctica C. australis D. antarctica C. australis Species Seasons Covariates
PNUE
(µmol CO2 mol-1
N-1
s-1
)
39±5 37±7 33±6 51±11
n.s. (0.3) n.s. (0.7) n.s.
Chl total (µmol m-2
) 390±51 279±27 725±88 496±58 ** ** n.s.
Chl a/b 3.0±0.1 3.1±0.2 3.5±0.4 3.1±0.1 n.s. (0.7) n.s. (0.7) n.s.
Ψ predawn (MPa) 1 -0.6±0.1 -0.5±0.1 -0.1±0.0 -0.1±0.0 n.s. (0.5) *** n/a
Ψ midday (MPa) -1.1±0.1 -1.2±0.1 -0.5±0.1 -0.3±0.1 n.s. (0.9) *** n.s.
WUEi
(µmol CO2 mol-1
H2O-1
) 125±16 109±12 62±6 64±12
n.s. (0.2) n.s. (0.8) PPFD* (+)
1-t-test

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-103-
5.3.2. Water status parameters
Predawn frond water potential (Ψ predawn) did not differ between species but was signifi-
cantly more negative in summer (t-test, P<0.001, Table 5.2). Fv/Fm was positively re-
lated to Ψ predawn (Table 5.1). Midday frond water potential (Ψ midday) was not related to
any of the tested covariates. It was similar between species but significantly more nega-
tive in summer than in winter (Table 5.2).
Growth irradiance had a significant positive effect on intrinsic water use efficiency
WUEi (Table 5.2). WUEi did not vary between species and seasons once covariates
were accounted for (Table 5.2).
5.3.3. Diurnal measurements
Instantaneous values of PPFD, leaf/air temperature, and leaf-to-air vapour pressure
deficit (VPD) at the measured fronds did not differ between species within season (t-test
P>0.1, data not shown). These variables differed significantly between seasons, with
mean daily PPFD, air temperature, and VPD in summer double that in winter (Fig.5.3).
Boundary-line analysis showed near-linear relationships between A and gs in summer
for both species, while in winter these relationships approached a saturation curve with
A reaching saturation at gs of ca 0.15 mol m-2
s-1
in both species (Fig. 5.4 a). VPD be-
came limiting to gs only at values greater than 1 kPa, which occurred only in summer
(Fig. 5.4 b). Optimal leaf temperature for photosynthesis in winter was on average 15ºC
for D. antarctica and 20ºC for C. australis, while in summer, A continued to rise at leaf
temperatures above 30ºC in both species (Fig. 5.4 c).
From light response curves fitted to the data of the diurnal measurements (Fig. 5.5),
Amax did not differ significantly between species but was significantly (P<0.05) higher
in summer than in winter (mean 17.4 vs. 8.6 µmol m-2
s-1
respectively). Light saturation
was reached at about 1100 in winter and nearly 1500 µmol photons m-2
s-1
in summer.

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-104-
Time (h)
9:00 11:00 1:00 3:00 5:00
PP
FD
(
mol m
-2s
-1)
0
500
1000
1500
2000
T a
ir (
oC
)
0
10
20
30
9:00 11:00 1:00 3:00 5:00
WinterSummerR
H (
%)
0
20
40
60
80
100
Fig. 5.3. Climate conditions during diurnal course measurements in summer and winter.
Lines indicate means.
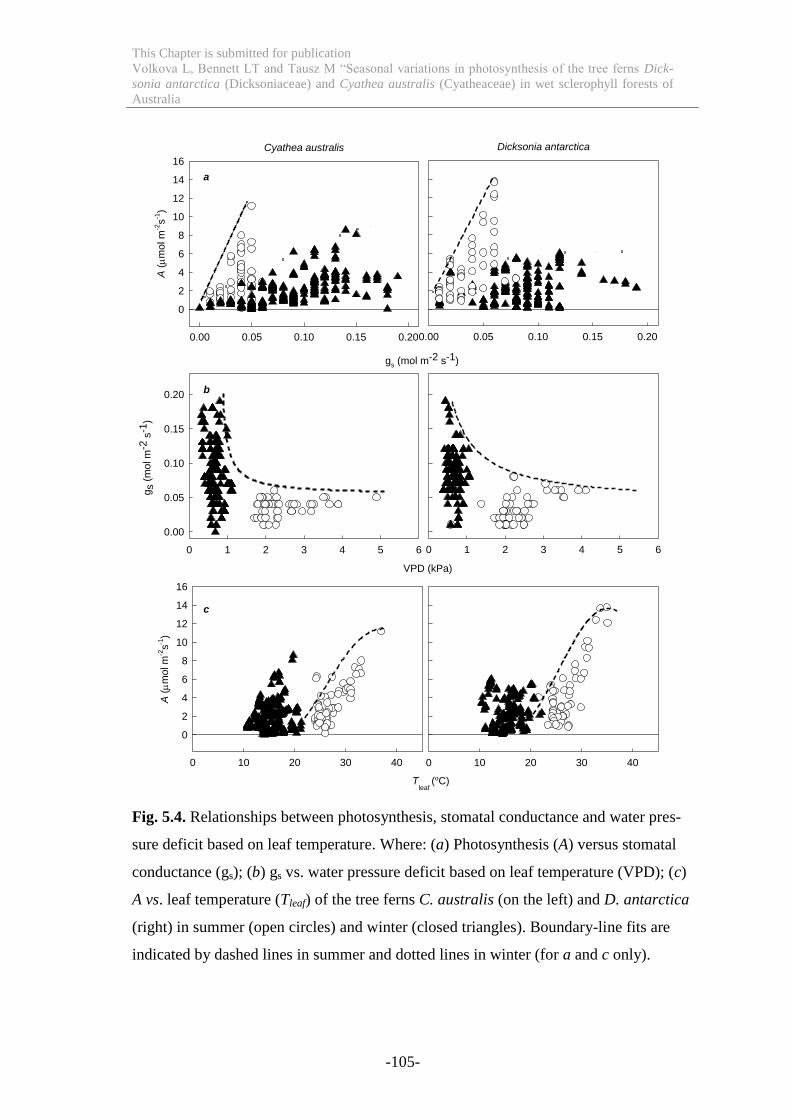
This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-105-
Tleaf
(oC)
0 10 20 30 40
A (
mo
l m
-2s
-1)
0
2
4
6
8
10
12
14
16
VPD (kPa)
0 1 2 3 4 5 6
gs (
mol m
-2 s
-1)
0.00
0.05
0.10
0.15
0.20
Cyathea australis
gs (mol m-2 s-1)
0.00 0.05 0.10 0.15 0.20
A (
mo
l m
-2s
-1)
0
2
4
6
8
10
12
14
16
a
b
c
Tleaf
,oC
5 10 15 20 25 30 35 40
gs, m
ol m
-2 s
-1
0.00
0.05
0.10
0.15
0.20
5 10 15 20 25 30 35 40
Dicksonia antarctica
0.00 0.05 0.10 0.15 0.20
0 1 2 3 4 5 6
0 10 20 30 40
Fig. 5.4. Relationships between photosynthesis, stomatal conductance and water pres-
sure deficit based on leaf temperature. Where: (a) Photosynthesis (A) versus stomatal
conductance (gs); (b) gs vs. water pressure deficit based on leaf temperature (VPD); (c)
A vs. leaf temperature (Tleaf) of the tree ferns C. australis (on the left) and D. antarctica
(right) in summer (open circles) and winter (closed triangles). Boundary-line fits are
indicated by dashed lines in summer and dotted lines in winter (for a and c only).

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-106-
PAR, mol photons m-2
s-1
0 500 1000 1500 2000 2500
A,
mo
lCO
2 m
-2s
-1
-4
0
4
8
12
16
Cyathea australis
0 500 1000 1500 2000 2500
Dicksonia antarctica
Fig. 5.5. Light response curves of the tree ferns D. antarctica and C. australis in sum-
mer and winter. Where: open circles – summer; closed circles – winter. Boundary-line
fits are indicated by dashed lines in summer and dotted lines in winter (r2 >0.9 in all
fits).
5.3.4. Stomatal density
Fronds of both species had no stomata on their adaxial (upper) surface. D. antarctica
displayed a more regular stomatal pattern in contrast to the random distribution of sto-
mata in C. australis (Fig. 5.6). Stomatal density did not differ significantly between
species (P=0.08) and was not correlated with growth irradiance (r =-0.007, P>0.9).
Leaf hairs were only observed on fronds of C. australis. Growth irradiance marginally
affected leaf hair density (P=0.06), which varied from 22 leaf hairs mm-2
on high light
exposed fronds to 11 leaf hairs mm-2
on shaded fronds, with an overall average of 13
hairs per mm-2
.
5.4. Discussion
5.4.1. Comparisons between the two tree fern species

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-107-
Regardless of their putatively different origins and distribution patterns within forests,
the majority of photosynthetic parameters did not differ between our two tree fern spe-
cies. Light saturated rates of net photosynthesis (Amax) were similar and within the range
reported previously for D. antarctica (Hunt et al. 2002, Volkova et al. 2009). Specific
leaf areas of D. antarctica and C. australis were in the range of SLAs for humid tem-
perate and tropical forests (Reich et al. 1999).
Both D. antarctica and C. australis performed better in winter, when photosynthetic
rates reached higher values under similar irradiance and leaf temperatures. Both higher
stomatal conductance, through less diffusive resistance to CO2, and greater frond N con-
tent, via increased carboxylation capacity dependent on N-rich proteins such as Rubisco
(Niinemets and Tenhunen 1997), could jointly enable greater assimilation rates.
Chlorophyll a/b ratios did not vary between seasons, suggesting low acclimation of both
tree ferns for this parameter, consistent with previous observations of ferns in the genus
Trichomanes (Johnson et al. 2000).
5.4.2. Light as a limiting factor to tree fern photosynthetic performance
Under our field conditions light-saturation of photosynthesis was recorded at above
1100 µmol m-2
s-1
, considerably higher than previously reported for D. antarctica (600
µmol m-2
s-1
for plants grown under natural illumination in a glasshouse, and 150 µmol
m-2
s-1
for plants grown in a gully, Hunt et al. 2002). This might reflect physiological
acclimation to open canopy conditions created after logging (Kursar and Coley 1999),
or differences between studies related to plant and frond age.
Growth irradiance (measured via mean daily PPFD) had stimulating effects on Amax and
NA confirming results with other tree species (e.g. González-Rodríguez et al. 2001,
Oliveira and Peñuelas 2004, Niinemets 2007). Increase in WUEi (Amax/gs) with increas-
ing growth irradiance possibly reflected the strong relationship between light and Amax.

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-108-
Linear relationships between A and gs, as observed in summer indicate that photosyn-
thesis was limited only by gs (Jones 1992), which, in turn, tended to decrease with in-
creases in VPD. In winter, A reached a plateau at ca gs 0.15 mol m-2
s-1
, indicating addi-
tional non-stomatal limitation of A (such as amount or activity of Rubisco or the rate of
electron transport, Massonnet et al. 2007).
In contrast to results from many other species (e.g. Romero and Botía 2006, Tazoe et al.
2009), gs at Amax (which reflects longer term response of gs to growth irradiance) was
insensitive to mean daily PPFD, indicating a lack of acclimation in gs to observed
growth irradiance for our tree fern species. In addition, there was no significant relation-
ship between instantaneous gs and instantaneous PPFD (as measured during the diurnal
measurements, P=0.9). This is in contrast to common knowledge that increasing light
induces gs until a saturation point is reached (Larcher 2003), a relationship commonly
used in models of stomatal responses. Such results may suggest poor responsiveness of
tree ferns‟ stomata to light and confirm observations of Doi and Shimazaki (2008),
when stomatal conductance of the fern Adiantum capillus-veneris showed much higher
light responsiveness when the light was applied to the lower leaf surface, where stomata
were situated, than when it was applied to the upper surface (as in our case). Similar
non-responsiveness of stomata to instantaneous light was found for Vicia faba (Mott et
al. 2008), leading to the conclusion that mesophyll signals play decisive roles in pre-
conditioning stomatal response to light.
The absence of significant relationship between the growth irradiance and Fv/Fm indi-
cates that PSII of both species fully recovered overnight regardless of received mean
daily PPFD.
Inverse relationship between the growth irradiance and SLA is consistent with the view
that sun leaves are more sclerophyllous (Groom and Lamont 1997, Niinemets 2007).
However, in contrast to the opinion that higher level of sclerophylly (i.e. decreased
SLA) results either from combinations of less water availability and high light intensi-
ties or from nutrient impoverished soils (for details see Groom and Lamont 1997), we

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-109-
found lower SLA in winter‟s than summer‟s frond cohorts, when light intensities were
lower and water availability was greater. In agreement with Turner (1994) we can only
conclude that increase in sclerophylly was not related to increased drought tolerance or
acclimation to drier conditions of the tree ferns.
Overall, the growth irradiance stimulated photosynthetic capacity and nitrogen alloca-
tion in fronds, yet, it had little effect on stomata or PSII in these tree fern species.
5.4.3. Temperature as a limiting factor to tree fern photosynthetic performance
Both tree fern species displayed broad temperature optima for net photosynthesis simi-
lar to a wide range of C3 plants reviewed by Kattge and Knorr (2007). Optimum tem-
peratures were higher in summer, confirming acclimation potential of the tree ferns to
higher growth temperatures, consistent with our previous study (Volkova et al. 2009).
Inverse relationship of Tleaf with Fv/Fm reflects the sensitivity of PSII to high tempera-
tures, which is often cited as the most heat-sensitive component of photosynthesis in
temperate species (e.g. Berry and Björkman 1980). Over and above the effect of sea-
sonal differences in Tleaf, season also had a significant additional effect on Fv/Fm – at the
same Tleaf, Fv/Fm was significantly lower in winter than in summer (Fig. 5.2) – suggest-
ing some cold-induced limitation in PSII efficiency. Reduction of Fv/Fm during winter is
confirmed by numerous other studies (see Wittmann et al. 2007), and may be related to
either chilling-induced photo-degradation of PSII components, or overnight retention of
de-epoxidised xanthophylls (Adams and Demmig- Adams 1994). Furthermore, Larcher
and Nagele (1992) demonstrated that photosynthetic capacity of Fagus sylvatica stems
decreased in winter and even short-term rewarming treatments could not restore it to
summer values. Thus, we may suggest that in winter, the tree ferns may experience per-
sistent photoinhibition of PSII induced by cold morning temperatures that are common
for this study site.

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-110-
Significant positive relations between NA and temperature might reflect changes in soil
nutrient mineralisation and thus N availability with temperature. In addition, a study by
Muller et al. (2009) indicated that temperature rather than irradiance primarily deter-
mined changes in NA in natural conditions for broad-leaved evergreen species.
Evaporative cooling through increased stomatal conductance and associated transpira-
tion implies a negative relationship between Tleaf and gs (see Snider et al. 2009). In con-
trast, we observed a positive relationship of Tleaf with gs, which may indicate that in our
study area gs is affected by low rather than high temperatures. Stomatal closure can also
occur when the water supply from the roots is restricted because of low temperatures in
the root zone (Davies et al. 1982).
5.4.4. Effects of plant water status and water relation parameters on tree fern
photosynthetic performance
Plant water status (measured via Ψ predawn) showed no significant relationship with any
of the measured parameters except Fv/Fm. Apparently, slightly, albeit significantly, more
negative Ψ predawn in summer (-0.05 MPa) relative to winter (-0.01 MPa) was not enough
to impose significant limitations on any of the measured variables. In agreement with
their ecological distribution in moist sites, tree ferns may be unable to tolerate a larger
drop in water potential, instead relying on an „avoidance‟ strategy involving frond loss,
as indicated in another open-air study by the first author (yet unpublished data). Sensi-
tivity of Fv/Fm to plant water status suggests that the efficiency of the photosystem
likely decreased due to reduced efficiency of the light-harvesting and antenna com-
plexes to deliver quanta to reaction centres (Wright et al. 2009).
No effect of season on intrinsic water use efficiency (WUEi) indicates prodigal or water
spending strategy by both species – the strategy, appropriate for plants that are subject
to droughts of short duration (Passioura 1982). Ferns are known for poor efficiency in
water transport through the leaf (Sack and Holbrook 2006), and together with the ob-

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-111-
served low acclimation potential of WUEi to seasonal fluctuations illustrates why many
fern species are confined to moist environments (Franks and Farquhar 1999).
5.4.5. Stomatal density
Stomatal patterns but not density differed between our two tree fern species. In contrast
to many angiosperms, where stomatal density is higher in leaves from sunny habitats
(Larcher 2003), stomatal density of both tree fern species did not correlate with the
growth irradiance. A similar lack of relationship between stomatal density and the
growth irradiance was observed for tree ferns of Cyatheaceae family by Arens (1997),
and for whole fern communities by Kessler et al. (2007). Our results indicate that tree
ferns (Pteridophyte) lack acclimation in terms of changing stomatal density in response
to their light environment.
Leaf hairs, known to reduce leaf temperature due to their reflection function (Lambers
et al. 2008) or to protect against UV-B radiation (Grammatikopoulos et al. 1994), were
found only in C. australis, possibly reflecting its microclimate preferences and assumed
Pan-tropical origin (i.e. adaptation to greater light intensities).
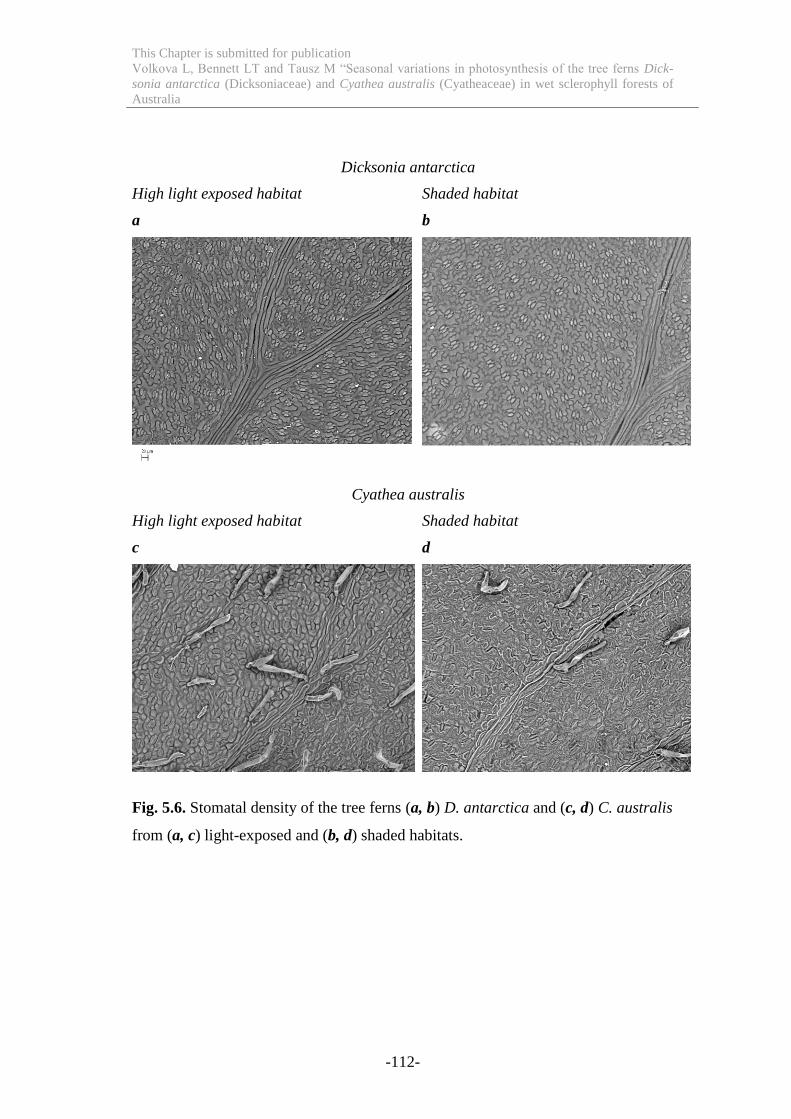
This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-112-
Dicksonia antarctica
High light exposed habitat Shaded habitat
a b
Cyathea australis
High light exposed habitat Shaded habitat
c d
Fig. 5.6. Stomatal density of the tree ferns (a, b) D. antarctica and (c, d) C. australis
from (a, c) light-exposed and (b, d) shaded habitats.

This Chapter is submitted for publication
Volkova L, Bennett LT and Tausz M “Seasonal variations in photosynthesis of the tree ferns Dick-
sonia antarctica (Dicksoniaceae) and Cyathea australis (Cyatheaceae) in wet sclerophyll forests of
Australia
-113-
5.5. Summary
Regardless of different origins and micro-site habitats, there were no significant differ-
ences in terms of photosynthetic performance between D. antarctica and C. australis,
and both tree fern species had greater photosynthetic capacity in winter. Low tempera-
tures appeared to be most limiting factor on plant performance under field conditions.
Plant water status did not vary markedly and had no effect on any of the measured pa-
rameters. Similar to other plants, both species of tree ferns displayed seasonal acclima-
tion in a number of measured photosynthetic parameters and frond traits (i.e. Fv/Fm,
Amax, gs, NA, chlorophyll total, SLA). Acclimation of stomatal density to spatial varia-
tion in growth irradiance among micro-sites seemed limited in both species, while
stomatal pattern differed between species.
Because there were no significant differences between the two species in photosynthetic
parameters, both species can probably be described by common carbon gain and water
use models at the leaf scale.

-114-
.

-115-
Chapter 6. Ecophysiology of two tree fern species and implications for
their future management. General discussion and conclusions
6.1. Species overview
The maximum amount of carbon fixed per unit leaf area and time or Amax (Taiz and
Zeiger 2002) is believed to be low in ferns compared with many other plants (Nobel et
al. 1984). In fact, both tree fern species in all of my studies had maximal photosynthetic
rates similar to a range of tree species in comparable ecosystems (e.g. middle-storey
canopy species Nothofagus cunninghamii Oerst (Tausz et al. 2005), Canarian laurel for-
est tree species Persea indica (González-Rodríguez et al. 2002), or Laurus azorica
(González-Rodríguez et al. 2001, Chapters 2-5).
According to von Caemmerer and Farquhar (1981), changes in Amax reflect changes in
both stomatal conductance (gs) and mesophyll capacity parameters (i.e. Vcmax and Jmax).
Stomatal conductance was greater in C. australis than D. antarctica at the beginning of
the water stress experiment (Chapter 3), but was comparable between two species in all
other experiments (Chapters 2 and 5, with an average of 100 mmol m-2
s-1
). The maxi-
mum carboxylation rate, Vcmax, or in vivo apparent Rubisco activity, was comparable in
both species across all experiments (Chapters 2, 4), while the maximum rate of electron
transport, Jmax, was consistently higher in D. antarctica than C. australis (Chapters 2,
4). Overall, both species had relatively low values of Vcmax and Jmax comparable with
some shade tolerant tree species, such as silver fir (Abies alba Mill, Robakowski et al.
2002, Chapters 2-5).
Stable isotope composition, δ13
C, is related to the ratio of internal to external CO2 con-
centrations (Ci/Ca), and therefore can be a useful tool in assessing intrinsic water use
efficiency, WUEi (Dawson et al. 2002). Lowest δ13
C assumes greater WUE (Farquhar
et al. 1989). δ13
C values of both tree fern species were within the ranges reported for C3
plants (-20 to -35‰, Dawson et al. 2002). However, despite lower δ13
C in C. australis‟s

-116-
fronds (closer to -30‰, Chapter 4), it was D. antarctica that had significantly greater
instantaneous (i. e. measured by gas exchange) WUEi. Values for WUEi in juveniles of
both species (range of 43-80µmol CO2 mol-1
H2O; Chapter 4) were comparable with
those recorded for mature tree ferns in the field study (Chapter 5), yet they were double
those recorded for an evergreen herbaceous fern of the family Adiantaceae Adiantum
reniforme var. sinensis (Liao et al. 2008).
Specific leaf area (SLA) is an index of sclerophylly (Lamont et al. 2002) and tends to be
higher in tropical and humid temperate forests (Reich et al. 1999). D. antarctica had
consistently lower SLA than C. australis across several experiments (e.g. 8.6 vs. 10.4
m2kg
-1 respectively). However, compared to a broad range of species, both species had
SLA similar to native Hawaiian tree ferns of the genus Cibotium (C. chamissoi), grown
in semi-wet to wet forests (Durand and Goldstein 2001).
Concentrations of chlorophylls were comparable between tree fern species in the first
glasshouse experiment (high light stress), but were greater in D. antarctica than C. aus-
tralis in other experiments involving older plants (Chapters 4, 5). Total chlorophyll
concentrations (per leaf area) of both species were comparable with a wide range of tree
species (e.g. shade grown Fagus sylvatica (Lichtenthaler 2007). Absolute values of total
chlorophyll concentrations only significantly differed in the second experiment, which
examined high light and heat interactions (Chapter 3). This could be due to the different
methods used to determine chlorophyll concentrations (HPLC vs. LECO), and possibly
due to plant age (Louis et al. 2009). Chlorophyll a/b ratio was comparable between the
two species in mature, field plants (Chapter 5), but was consistently higher in D. antarc-
tica than C. australis in juvenile plants (Chapters 2, 4) due to greater chlorophyll a con-
centrations.
Overall, my project found that the photosynthetic characteristics of D. antarctica and C.
australis were comparable and within reported ranges for a number of plant types. Thus,
the different origins of the two species, and their apparent preference for different mi-

-117-
cro-sites, do not appear to have resulted in a divergence of their physiological responses
to a range of environmental stresses.
6.2. Overview of light, temperature, and water availability as stresses on tree fern
physiology
My results in Chapter 2 (high light stress) indicated immediate down-regulation of pho-
tosynthetic capacity parameters by tree ferns in response to high light, but acclimation
to high light after prolonged (three month) exposure. These results are consistent with
the view that high light alone does not have detrimental effect on photosynthesis and
plants can often fully recover (Levitt 1980, Lovelock et al. 1994).
In Chapter 3, I examined interactive effects of high light and moderate heat on photo-
synthetic capacity of D. antarctica. Here, combination of high light and moderate heat
led to negative effects including chlorophyll bleaching and severe photoinhibition
(measured as a decrease in Fv/Fm<0.4). Given the comparable responses to stresses by
both tree fern species in all other studies (see above), it is reasonable to assume that
high light with moderate heat would be equally problematic for C. australis. This result
of detrimental effects of light by temperature interactions supports findings of many
other studies (e.g. Al-Khatib and Paulsen 1999, Montgomery et al. 2008), although it is
in contrast to Havaux et al. (1991), who stated that high light could alleviate negative
effects of high temperatures.
In Chapter 4, I studied interactive effects of high light and water deficit on photosyn-
thetic capacity and water relation parameters of both species D. antarctica and C. aus-
tralis. It appeared that both species were susceptible to water deficit either alone or in
interaction with high light. This funding was in contrast with Levitt (1980) stating that
interactive effect of high light and water deficit can be more detrimental than water
deficit alone. Stomatal response to water withheld was slower in shade-grown than in
high light exposed plants, confirming the findings with bracken fern where stomata
from the most shaded understorey level were less sensitive to soil moisture treatments

-118-
than those from more irradiated levels (Roberts et al. 1984). Probably as a result of it
there were some indications that shade did not ameliorate but rather intensified drought
effects on D. antarctica, the result confirming observation of Valladares et al. (2002)
who stated that the capacity to withstand severe drought was not enhanced in the shade
but decreased due to increased below-ground competition for water with established
trees.
The focus of Chapter 5 was on mature tree ferns growing in their natural environment,
the mountain ash forests of the Victorian Central Highlands, Australia. Measurements
of photosynthetic capacity as well as water relations and frond parameters over two
consecutive years indicated that both species, irrespective of their different origins and
apparent differences in micro-site preferences, performed better in winter than in sum-
mer. Low light and low temperatures were limiting factors for the tree ferns perform-
ance in the field. In contrast to many reports in the literature (e.g. Romero and Botía
2006, Tazoe et al. 2009), stomatal conductance of the tree ferns did not correlate with
growth irradiance but was positively correlated only with temperature. Plant water
status (Ψ predawn) had no effect on any of the measured parameters (except maximal
quantum yield of photochemistry, Fv/Fm), possibly because decreases in Ψ predawn to -
0.05 MPa were not enough to impose limitations. Another interpretation is that tree
ferns lack the ability to adjust to decreased water availability and cannot endure more
pronounced drops in tissue water potentials. This may also explain observations in both,
field and glasshouse, plants, where ferns shed fronds under more severe water deficit.
Both species showed characteristics of a water spending strategy irrespective of season
(summer or winter), i.e. WUEi in summer, when temperature was higher and less water
available, was comparable to WUEi in winter, a season characterised by lower tempera-
tures and greater rainfalls. Such prodigal strategy is common and effective for plants
that only occasionally experience drought (Passioura 1982). Using this interpretation,
results of this field Chapter were consistent with indications of low acclimation of the
tree ferns to water availability found in the controlled experiment involving juvenile
plants (Chapter 4). Tree ferns growing at open sites showed little difference in stomatal

-119-
density from those growing under dense canopy cover, indicating a lack of acclimation
of stomatal density in the tree ferns to growth irradiance. This is in contrast to observa-
tions on many other tree species (Larcher 2003), but consistent with findings of Arens
(1997) on the tree fern Cyathea caracasana.
6.3. Practical implications and future directions
Despite their ecological importance as dominant understorey species in Australia‟s wet
sclerophyll forests, the physiology of D. antarctica and C. australis has previously been
under-examined. Understorey plants like these two species may account for as much as
33% of living above-ground biomass (Weaver 2008), and, in evergreen forests world-
wide, contribute on average 49% of total ecosystem respiration (Misson et al. 2007).
Thus, the paucity of knowledge about tree fern physiology not only limits our ability to
predict potential impacts of climate change on their distribution but also excludes a sig-
nificant component of temperate forests from total carbon balance calculations (i.e. by
excluding a significant portion of potential carbon sinks and sources).
Two experiments and one observational field study (Chapters 2, 4 and 5) indicated that
D. antarctica and C. australis are fairly similar with respect to (eco)physiological and
morphological characteristics. Correspondingly, growth and carbon sequestration mod-
els could use a common set of parameters for both species, which would greatly sim-
plify the task of including tree ferns in process-based carbon accounting or environ-
mental risk assessment models. Moreover, because D. antarctica and C. australis be-
long to the two main tree fern families Dicksoniaceae and Cyatheaceae – arguably the
most important tree fern families worldwide – the knowledge created in this thesis will
be a sound starting point for understanding the ecophysiology of other tree ferns across
a potentially broad range of environmental conditions.
The problem of poor survival and ongoing decline of the tree fern numbers after clear-
cut logging (Ought and Murphy 1998) was one of the main inspirations for this study.
My experimental results, particularly Chapter 3 (high light and high temperature), indi-

-120-
cate detrimental effects of combined high light and moderate temperatures on tree fern
health, which should be considered when designing logging configurations. For exam-
ple, understorey islands should be designed to minimise tree fern numbers near edges
where they will be exposed to greater light intensities and consequently higher tempera-
tures. This could involve consideration of a minimum distance to edge for tree fern
populations of considerable numbers.
The results of Chapter 4 (high light and water deficit) reiterated the importance to tree
ferns of retaining buffer zones along waterways. This experiment also indicated that re-
tention of understorey islands in the middle of logging coupes away from waterways,
might not lead to substantial improvement in the protection of tree ferns in logging
coupes under climate change predictions of increasing drought frequency. Indeed, my
results indicated that shade led to slower stomatal responses in D. antarctica thereby
intensifying drought stress. Thus, design of understorey islands should take tree fern
needs for access to water into consideration, and should thus be placed on lower rather
than upper slopes.
Climate change predictions indicate that the severity and frequency of both droughts
and fire will increase in south-eastern Australia (Hennessy et al. 2007), leading to
changes in species distributions (see Fitzpatrick et al. 2008). Thus, species better
adapted to drought and fire could expand their range, while other elements of native
vegetation might be at risk (Watt et al. 2009). Findings from my study indicate that the
range of tree ferns in south-eastern Australia may contract under a more irradiated (i.e.
due to fire-induced canopy death), warmer, and drier climate. In addition, the current
divergence in micro-site preferences between the two studied species might become less
evident. That is, current localised extensions of C. australis to forest margins might
contract to more shaded, moister sites to be closer to the current distribution of D. ant-
arctica. This emphasises the need for buffer zones along waterways, which under future
climates could be considered refugia for tree ferns – that is, isolated areas of habitat that
retain the environmental conditions that were once widespread, Stewart and Lister
2001).

-121-
While my study provides useful indications of tree fern responses to environmental
stresses, it was limited to photosynthetic capacity parameters at the leaf-level scale. To
increase accuracy of carbon models, future studies should consider dynamics of frond
development, including frond longevity, and the potentially different physiology of
fronds of different age cohorts. In addition, precise micro-climate data will be another
essential component for improved modelling of tree fern ecophysiology and of under-
storey plants in general. Modelling at landscape to regional scale would be of particular
benefit to conservation goals, but scaling errors need to be considered.
Overall, my study revealed that both species of the tree ferns might be vulnerable in the
future because of their low acclimation potential to environmental stresses. These find-
ings highlight the need to develop more flexible conservation policies to maintain tree
fern communities under optimal conditions where they can be most resilient to both
predicted and unexpected future changes.

-122-

-123-
REFERENCES
Adams WW, Demmig-Adams B (1994) Carotenoid composition and down-regulation
of photosystem II in 3 conifer species during the winter. Physiologia Plantarum
92, 451-458.
Al-Khatib K, Paulsen GM (1999) High-temperature effects on photosynthetic processes
in temperate and tropical cereals. Crop Science 39, 119-125.
Arens NC (1997) Responses of leaf anatomy to light environment in the tree fern Cyat-
hea caracasana (Cyatheaceae) and its application to some ancient seed ferns.
Palaios 12, 84-94.
Ashton DH (2000) The Big Ash Forest, Wallaby Creek, Victoria - changes during one
lifetime. Australian Journal of Botany 48, 1-26.
Attiwill PM, Fewings JM (2001) 'Sustainable management of the Mountain ash (Euca-
lyptus regnans F. Muell) forests in Central Highlands, Victoria, Australia.'
(Blackwell science)
Bannister P, Wildish KL (1982) Light compensation points and specific leaf areas in
some New-Zealand ferns. New Zealand Journal of Botany 20, 421-424.
Barlow BA (1994) Phytogeography of the Australian region. In 'Australian vegetation'.
(Ed. RH Groves) pp. xvii, 562 p. (Cambridge University Press: Cambridge,
[England]; Oakleigh, Vic.)
Bennett LT, Adams MA (2004) Assessment of ecological effects due to forest harvest-
ing: approaches and statistical issues. Journal of Applied Ecology 41, 585-598.
Berry J, Björkman O (1980) Photosynthetic response and adaptation to temperature in
higher plants. Annual Review of Plant Physiology and Plant Molecular Biology
31, 491-543.
Blows MW, Schwarz MP (1991) Spatial-distribution of a primitively social bee - does
genetic population-structure facilitate altruism. Evolution 45, 680-693.
Bogeat-Triboulot MB, Brosche M, Renaut J, Jouve L, Le Thiec D, Fayyaz P, Vinocur
B, Witters E, Laukens K, Teichmann T, Altman A, Hausman JF, Polle A, Kan-
gasjarvi J, Dreyer E (2007) Gradual soil water depletion results in reversible
changes of gene expression, protein profiles, ecophysiology, and growth per-
formance in Populus euphratica, a poplar growing in arid regions. Plant Physiol-
ogy 143, 876-892.
Bota J, Medrano H, Flexas J (2004) Is photosynthesis limited by decreased Rubisco ac-
tivity and RuBP content under progressive water stress? New Phytologist 162,
671-681.
Brodribb TJ, Feild TS, Jordan GJ (2007) Leaf maximum photosynthetic rate and vena-
tion are linked by hydraulics. Plant Physiology 144, 1890-1898.
Brodribb TJ, Holbrook NM, Zwieniecki MA, Palma B (2005) Leaf hydraulic capacity
in ferns, conifers and angiosperms: impacts on photosynthetic maxima. New
Phytologist 165, 839-846.
Callister AN, Arndt SK, Adams MA (2006) Comparison of four methods for measuring
osmotic potential of tree leaves. Physiologia Plantarum 127, 383-392.

-124-
Campbell ML, Clarke PJ (2006) Response of montane wet sclerophyll forest understo-
rey species to fire: Evidence from high and low intensity fires. Proceedings of
the Linnean Society of New South Wales 127, 63-73.
Casson S, Gray JE (2008) Influence of environmental factors on stomatal development.
New Phytologist 178, 9-23.
Chaves MM, Maroco JP, Pereira JS (2003) Understanding plant responses to drought -
from genes to the whole plant. Functional Plant Biology 30, 239-264.
Davies S (2005) Harvesting of Manferns (Dicksonia antarctica) in Tasmania. Forest
Practices News 6 5-6.
Davies WJ, Rodriguez JL, Fiscus EL (1982) Stomatal behavior and water movement
through roots of wheat plants treated with abscisic acid. Plant Cell and Envi-
ronment 5, 485-493.
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in
plant ecology. Annual Review of Ecology and Systematics 33, 507-559.
De Costa W, Zörb C, Hartung W, Schubert S (2007) Salt resistance is determined by
osmotic adjustment and abscisic acid in newly developed maize hybrids in the
first phase of salt stress. Physiologia Plantarum 131, 311-321.
Demmig-Adams B, Adams WW (2006) Photoprotection in an ecological context: the
remarkable complexity of thermal energy dissipation. New Phytologist 172, 11-
21.
Department of Natural Resources and Environment (2002) Victorian tree fern manage-
ment plan. Department of Natural Resources and Environment, East Melbourne,
Australia.
Dieleman JA, Meinen E (2007) Interacting effects of temperature integration and light
intensity on growth and development of single-stemmed cut rose plants. Scientia
Horticulturae 113, 182-187.
Dignan P, Bren L (2003) A study of the effect of logging on the understorey light envi-
ronment in riparian buffer strips in a south-east Australian forest. Forest Ecology
and Management 172, 161-172.
Doi M, Shimazaki KI (2008) The stomata of the fern Adiantum capillus-veneris do not
respond to CO2 in the dark and open by photosynthesis in guard cells. Plant
Physiology 147, 922-930.
Doi M, Wada M, Shimazaki K (2006) The fern Adiantum capillus-veneris lacks
stomatal responses to blue light. Plant and Cell Physiology 47, 748-755.
Downton WJS, Berry JA, Seemann JR (1984) Tolerance of photosynthesis to high tem-
perature in desert plants. Plant Physiology 74, 786-790.
Dreyer E, Le Roux X, Montpied P, Daudet FA, Masson F (2001) Temperature response
of leaf photosynthetic capacity in seedlings from seven temperate tree species.
Tree Physiology 21, 223-232.
Ducruet JM, Peeva V, Havaux M (2007) Chlorophyll thermofluorescence and ther-
moluminescence as complementary tools for the study of temperature stress in
plants. Photosynthesis Research 93, 159-171.
Durand LZ, Goldstein G (2001) Photosynthesis, photoinhibition, and nitrogen use effi-
ciency in native and invasive tree ferns in Hawaii. Oecologia 126, 345-354.
Epron D, Dreyer E (1992) Effects of severe dehydration on leaf photosynthesis in
Quercus-petraea (Matt) Liebl - photosystem-II efficiency, photochemical and

-125-
nonphotochemical fluorescence quenching and electrolyte leakage. Tree Physi-
ology 10, 273-284.
Ethier GJ, Livingston NJ (2004) On the need to incorporate sensitivity to CO2 transfer
conductance into the Farquhar-von Caemmerer-Berry leaf photosynthesis model.
Plant Cell and Environment 27, 137-153.
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and pho-
tosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology
40, 503-537.
Farquhar GD, von Caemmerer S, Berry JA (1980) A biochemical model of photosyn-
thetic CO2 assimilation in leaves of C3 species. Planta 149, 78-90.
Fitzpatrick MC, Gove AD, Sanders NJ, Dunn RR (2008) Climate change, plant migra-
tion, and range collapse in a global biodiversity hotspot: the Banksia (Pro-
teaceae) of Western Australia. Global Change Biology 14, 1337-1352.
Flexas J, Escalona JM, Medrano H (1998) Down-regulation of photosynthesis by
drought under field conditions in grapevine leaves. Australian Journal of Plant
Physiology 25, 893-900.
Flexas J, Escalona JM, Medrano H (1999) Water stress induces different levels of pho-
tosynthesis and electron transport rate regulation in grapevines. Plant Cell and
Environment 22, 39-48.
Flexas J, Medrano H (2002) Drought-inhibition of photosynthesis in C3 plants: Stomatal
and non-stomatal limitations revisited. Annals of Botany 89, 183-189.
Flexas J, Ribas-Carbó M, Bota J, Galmés J, Henkle M, Martínez-Cañellas S, Medrano
H (2006) Decreased Rubisco activity during water stress is not induced by de-
creased relative water content but related to conditions of low stomatal conduc-
tance and chloroplast CO2 concentration. New Phytologist 172, 73-82.
Franks PJ, Farquhar GD (1999) A relationship between humidity response, growth form
and photosynthetic operating point in C3 plants. Plant Cell and Environment 22,
1337-1349.
Froux F, Ducrey M, Epron D, Dreyer E (2004) Seasonal variations and acclimation po-
tential of the thermostability of photochemistry in four Mediterranean conifers.
Annals of Forest Science 61, 235-241.
Galle A, Haldimann P, Feller U (2007) Photosynthetic performance and water relations
in young pubescent oak (Quercus pubescens) trees during drought stress and re-
covery. New Phytologist 174, 799-810.
García-Plazaola JI, Becerril JM (1999) A rapid high performance liquid chromatogra-
phy method to measure lipophilic antioxidants in stressed plants: Simultaneous
determination of carotenoids and tocopherols. Phytochemical Analysis 10, 307-
313.
Ghouil H, Montpied P, Epron D, Ksontini M, Hanchi B, Dreyer E (2003) Thermal op-
tima of photosynthetic functions and thermostability of photochemistry in cork
oak seedlings. Tree Physiology 23, 1031-1039.
Gildner BS, Larson DW (1992) Photosynthetic response to sunflecks in the desiccation-
tolerant fern Polypodium virginianum. Oecologia 89, 390-396.
González-Rodríguez AM, Morales D, Jiménez MS (2001) Gas exchange characteristics
of a Canarian laurel forest tree species (Laurus azorica) in relation to environ-
mental conditions and leaf canopy position. Tree Physiology 21, 1039-1045.

-126-
González-Rodríguez AM, Morales D, Jiménez MS (2002) Leaf gas exchange character-
istics of a Canarian laurel forest tree species [Persea indica (L.) K. Spreng.] un-
der natural conditions. Journal of Plant Physiology 159, 695-704.
Grammatikopoulos G, Karabourniotis G, Kyparissis A, Petropoulou Y, Manetas Y
(1994) Leaf hairs of olive (Olea europaea) prevent stomatal closure by Ultravio-
let-B radiation. Australian Journal of Plant Physiology 21, 293-301.
Groom PK, Lamont BB (1997) Xerophytic implications of increased sclerophylly: in-
teractions with water and light in Hakea psilorrhyncha seedlings. New Phytolo-
gist 136, 231-237.
Guo XR, Cao KF, Xu ZF (2006) Acclimation to irradiance in seedlings of three tropical
rain forest Garcinia species after simulated gap formation. Photosynthetica 44,
193-201.
Haldimann P (1999) How do changes in temperature during growth affect leaf pigment
composition and photosynthesis in Zea mays genotypes differing in sensitivity to
low temperature? Journal of Experimental Botany 50, 543-550.
Havaux M, Greppin H, Strasser RJ (1991) Functioning of photosystem I and photosys-
tem II in pea leaves exposed to heat-stress in the presence or absence of light -
analysis using in vivo fluorescence, absorbency, oxygen and photoacoustic
measurements. Planta 186, 88-98.
Havaux M, Gruszecki WI (1993) Heat-induced and light-induced chlorophyll a fluores-
cence changes in potato leaves containing high or low levels of the carotenoid
zeaxanthin - indications of a regulatory effect of zeaxanthin on thylakoid mem-
brane fluidity. Photochemistry and Photobiology 58, 607-614.
Havaux M, Tardy F (1996) Temperature-dependent adjustment of the thermal stability
of photosystem II in vivo: Possible involvement of xanthophyll cycle pigments.
Planta 198, 324-333.
Havaux M, Tardy F, Ravenel J, Chanu D, Parot P (1996) Thylakoid membrane stability
to heat stress studied by flash spectroscopic measurements of the electrochromic
shift in intact potato leaves: Influence of the xanthophyll content. Plant Cell and
Environment 19, 1359-1368.
Hennessy KB, Fitzharris BC, Bates N, Harvey SM, Howden L, Hughes J, Salinger and
R. Warrick (2007) Australia and New Zealand. In 'Climate Change 2007: Im-
pacts, Adaptation and Vulnerability. Contribution of Working Group II to the
Fourth Assessment Report of the Intergovernmental Panel on Climate Change'.
(Eds ML Parry, OF Canziani, JP Palutikof, PJ van der Linden and CE Hanson)
pp. 507-540. (Cambridge University Press, Cambridge, UK)
Herbinger K, Then C, Haberer K, Alexou M, Löw M, Remele K, Rennenberg H,
Matyssek R, Grill D, Wieser G, Tausz M (2007) Gas exchange and antioxidative
compounds in young beech trees under free-air ozone exposure and comparisons
to adult trees. Plant Biology 9, 288-297.
Hickey JE (1994) A floristic comparison of vascular species in Tasmanian old growth
mixed forest with regeneration resulting from logging and wildfire. Australian
Journal of Botany 42, 383-404.
Hoober JK, Eggink LL, Chen M (2007) Chlorophylls, ligands and assembly of light-
harvesting complexes in chloroplasts. Photosynthesis Research 94, 387-400.

-127-
Howard TM (1973) Studies in the ecology of Nothofagus Cunninghamii forest. Part 3.
Two limiting factors: light intensity and water stress. Australian Journal of Bot-
any 21, 93-102.
Hunt MA, Davidson NJ, Unwin GL, Close DC (2002) Ecophysiology of the Soft Tree
Fern Dicksonia antarctica Labill. Austral Ecology 27, 360-368.
Hüve K, Bichele I, Tobias M, Niinemets Ü (2006) Heat sensitivity of photosynthetic
electron transport varies during the day due to changes in sugars and osmotic po-
tential. Plant Cell and Environment 29, 212-228.
Johnson GN, Rumsey FJ, Headley AD, Sheffield E (2000) Adaptations to extreme low
light in the fern Trichomanes speciosum. New Phytologist 148, 423-431.
Jones DL, Clemesha SC (1993) 'Australian Ferns and fern allies.' (The Currawong
Press)
Jones HG (1992) 'Plants and microclimate: a quantitative approach to environmental
plant physiology.' (Cambridge University Press: Cambridge [England] ; New
York, NY, USA)
Kattge J, Knorr W (2007) Temperature acclimation in a biochemical model of photo-
synthesis: a reanalysis of data from 36 species. Plant Cell and Environment 30,
1176-1190.
Keith H, Mackey BG, Lindenmayer DB (2009) Re-evaluation of forest biomass carbon
stocks and lessons from the world's most carbon-dense forests. Proceedings of
the National Academy of Sciences 106, 11635-11640.
Kessler M, Siorak Y, Wunderlich M, Wegner C (2007) Patterns of morphological leaf
traits among pteridophytes along humidity and temperature gradients in the Bo-
livian Andes. Functional Plant Biology 34, 963-971.
Kirchgeßner HD, Reichert K, Hauff K, Steinbrecher R, Schnitzler JP, Pfündel EE
(2003) Light and temperature, but not UV radiation, affect chlorophylls and ca-
rotenoids in Norway spruce needles (Picea abies (L.) Karst.). Plant Cell and En-
vironment 26, 1169-1179.
Kursar TA, Coley PD (1999) Contrasting modes of light acclimation in two species of
the rainforest understory. Oecologia 121, 489-498.
Lambers H, Chapin FS, Pons TL (2008) 'Plant physiological ecology.' (Springer: New
York)
Lamont BB, Groom PK, Cowling RM (2002) High leaf mass per area of related species
assemblages may reflect low rainfall and carbon isotope discrimination rather
than low phosphorus and nitrogen concentrations. Functional Ecology 16, 403-
412.
Larcher W (2003) 'Physiological plant ecology: ecophysiology and stress physiology of
functional groups.' (Springer: New York)
Larcher W, Nagele M (1992) Changes in photosynthetic activity of buds and stem tis-
sues of Fagus-Sylvatica during winter. Trees-Structure and Function 6, 91-95.
Large MF, Braggins JE (2004) 'Tree ferns.' (CSIRO publishing: Collingwood, Vic.)
Law RD, Crafts-Brandner SJ (1999) Inhibition and acclimation of photosynthesis to
heat stress is closely correlated with activation of ribulose-1,5-bisphosphate car-
boxylase/oxygenase. Plant Physiology 120, 173-181.
Levitt J (1980) 'Responses of plants to environmental stresses.' (Academic Press: New
York)

-128-
Liao JX, Jiang MX, Huang HD (2008) Effects of soil moisture on ecophysiological
characteristics of Adiantum reniforme var. sinensis, an endangered fern endemic
to the Three Gorges region in China. American Fern Journal 98, 26-32.
Lichtenthaler HK (1987) Chlorophylls and carotenoids - pigments of photosynthetic
biomembranes. Methods in Enzymology 148, 350-382.
Lichtenthaler HK (2007) Biosynthesis, accumulation and emission of carotenoids, al-
pha-tocopherol, plastoquinone, and isoprene in leaves under high photosynthetic
irradiance. Photosynthesis Research 92, 163-179.
Lindenmayer DB, Cunningham RB, Donnelly CF, Triggs BJ, Belvedere M (1994) The
conservation of arboreal marsupials in the montane ash forests of the central
highlands of Victoria, South-Eastern Australia. 5. Patterns of use and the micro-
habitat requirements of the mountain brushtail possum Trichosurus caninus
ogilby in retained linear habitats (wildlife corridors). Biological Conservation
68, 43-51.
Llusià J, Peñuelas J, Munné-Bosch S (2005) Sustained accumulation of methyl salicy-
late alters antioxidant protection and reduces tolerance of holm oak to heat
stress. Physiologia Plantarum 124, 353-361.
Long SP, Bernacchi CJ (2003) Gas exchange measurements, what can they tell us about
the underlying limitations to photosynthesis? Procedures and sources of error.
Journal of Experimental Botany 54, 2393-2401.
Long SP, Humphries S, Falkowski PG (1994) Photoinhibition of photosynthesis in na-
ture. Annual Review of Plant Physiology and Plant Molecular Biology 45, 633-
662.
Louis J, Meyer S, Maunoury-Danger F, Fresneau C, Meudec E, Cerovic ZG (2009)
Seasonal changes in optically assessed epidermal phenolic compounds and chlo-
rophyll contents in leaves of sessile oak (Quercus petraea): towards signatures
of phenological stage. Functional Plant Biology 36, 732-741.
Lovelock CE, Jebb M, Osmond CB (1994) Photoinhibition and recovery in tropical
plant species: response to disturbance. Oecologia 97, 297-307.
Lovelock CE, Kursar TA, Skillman JB, Winter K (1998) Photoinhibition in tropical for-
est understorey species with short- and long-lived leaves. Functional Ecology
12, 553-560.
Magrath R, Lill A (1983) The use of time and energy by the crimson rosella in a tem-
perate wet forest in winter. Australian Journal of Zoology 31, 903-912.
Matsubara S, Krause GH, Aranda J, Virgo A, Beisel KG, Jahns P, Winter K (2009)
Sun-shade patterns of leaf carotenoid composition in 86 species of neotropical
forest plants. Functional Plant Biology 36, 20-36.
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence - a practical guide. Journal of
Experimental Botany 51, 659-668.
McCarthy PM (Ed.) (1998) 'Ferns, gymnosperms and allied groups.' Flora of Australia
(CSIRO Publications: Melbourne)
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry
J, West A, Williams DG, Yepez EA (2008) Mechanisms of plant survival and
mortality during drought: why do some plants survive while others succumb to
drought? New Phytologist 178, 719-739.

-129-
Merchant A, Tausz M, Arndt SK, Adams MA (2006) Cyclitols and carbohydrates in
leaves and roots of 13 Eucalyptus species suggest contrasting physiological re-
sponses to water deficit. Plant Cell and Environment 29, 2017-2029.
Misson L, Baldocchi DD, Black TA, Blanken PD, Brunet Y, Yuste JC, Dorsey JR, Falk
M, Granier A, Irvine MR, Jarosz N, Lamaud E, Launiainen S, Law BE, Longdoz
B, Loustau D, McKay M, Paw KT, Vesala T, Vickers D, Wilson KB, Goldstein
AH (2007) Partitioning forest carbon fluxes with overstory and understory eddy-
covariance measurements: A synthesis based on FLUXNET data. Agricultural
and Forest Meteorology 144, 14-31.
Montgomery RA, Goldstein G, Givnish TJ (2008) Photoprotection of PSII in Hawaiian
lobeliads from diverse light environments. Functional Plant Biology 35, 595-
605.
Montpied P, Granier A, Dreyer E (2009) Seasonal time-course of gradients of photosyn-
thetic capacity and mesophyll conductance to CO2 across a beech (Fagus sylva-
tica L.) canopy. Journal of Experimental Botany 60, 2407-2418.
Mott KA, Sibbernsen ED, Shope JC (2008) The role of the mesophyll in stomatal re-
sponses to light and CO2. Plant Cell and Environment 31, 1299-1306.
Muller O, Oguchi R, Hirose T, Werger MJA, Hikosaka K (2009) The leaf anatomy of a
broad-leaved evergreen allows an increase in leaf nitrogen content in winter.
Physiologia Plantarum 136, 299-309.
Munné-Bosch S (2005) The role of -tocopherol in plant stress tolerance. Journal of
Plant Physiology 162, 743-748.
Murata N, Takahashi S, Nishiyama Y, Allakhverdiev SI (2007) Photoinhibition of pho-
tosystem II under environmental stress. Biochimica Et Biophysica Acta-
Bioenergetics 1767, 414-421.
Niinemets Ü (2007) Photosynthesis and resource distribution through plant canopies.
Plant Cell and Environment 30, 1052-1071.
Niinemets Ü, Kull O, Tenhunen JD (1998) An analysis of light effects on foliar mor-
phology, physiology, and light interception in temperate deciduous woody spe-
cies of contrasting shade tolerance. Tree Physiology 18, 681-696.
Niinemets Ü, Tenhunen JD (1997) A model separating leaf structural and physiological
effects on carbon gain along light gradients for the shade-tolerant species Acer
saccharum. Plant Cell and Environment 20, 845-866.
Niyogi KK (2000) Safety valves for photosynthesis. Current Opinion in Plant Biology
3, 455-460.
Nobel PS, Calkin HW, Gibson AC (1984) Influences of PAR, temperature and water
vapor concentration on gas exchange by ferns. Physiologia Plantarum 62, 527-
534.
North HM, De Almeida A, Boutin JP, Frey A, To A, Botran L, Sotta B, Marion-Poll A
(2007) The Arabidopsis ABA-deficient mutant aba4 demonstrates that the major
route for stress-induced ABA accumulation is via neoxanthin isomers. Plant
Journal 50, 810-824.
Oikawa S, Hikosaka K, Hirose T (2006) Leaf lifespan and lifetime carbon balance of
individual leaves in a stand of an annual herb, Xanthium canadense. New Phy-
tologist 172, 104-116.

-130-
Oliveira G, Peñuelas J (2004) Effects of winter cold stress on photosynthesis and photo-
chemical efficiency of PSII of the Mediterranean Cistus albidus L. and Quercus
ilex L. Plant Ecology 175, 179-191.
Ottander C, Campbell D, Öquist G (1995) Seasonal changes in photosystem II organiza-
tion and pigment composition in Pinus sylvestris. Planta 197, 176-183.
Ough K (2001) Regeneration of Wet Forest flora a decade after clear-felling or wildfire
- is there a difference? Australian Journal of Botany 49, 645-664.
Ough K, Murphy A (1996) The effect of clearfell logging on tree-ferns in Victorian Wet
Forest. Australian Forestry 59, 178-188.
Ough K, Murphy A (1998) Understorey Island: a method of protecting understorey flora
during clearfelling operations. Department of Natural Resources and Environ-
ment, VSP Internal report # 29, Heidelberg.
Ough K, Murphy A (2004) Decline in tree-fern abundance after clearfell harvesting.
Forest Ecology and Management 199, 153-163.
Page CN, Clifford HT (1981) Ecological biogeography of Australian conifers and ferns.
In 'Ecological biogeography of Australia'. (Ed. A Keast) pp. 471-498. (Dr. W.
Junk bv Publishers: Hague-Boston-London)
Passioura JB (1982) Water in the soil-plant-atmosphere continuum. In 'Physiological
Plant Ecology II. Water Relations and Carbon Assimilation'. (Eds OL Lange, PS
Nobel, CB Osmond and H Ziegler) pp. 5–33. (Springer-Verlag: Berlin, Heidel-
berg, New York)
Pearcy RW (1988) Photosynthetic utilization of lightflecks by understory plants. Aus-
tralian Journal of Plant Physiology 15, 223-238.
Reich PB, Ellsworth DS, Walters MB, Vose JM, Gresham C, Volin JC, Bowman WD
(1999) Generality of leaf trait relationships: A test across six biomes. Ecology
80, 1955-1969.
Ribas-Carbo M, Taylor NL, Giles L, Busquets S, Finnegan PM, Day DA, Lambers H,
Medrano H, Berry JA, Flexas J (2005) Effects of water stress on respiration in
soybean leaves. Plant Physiology 139, 466-473.
Robakowski P, Montpied P, Dreyer E (2002) Temperature response of photosynthesis
of silver fir (Abies alba Mill.) seedlings. Annals of Forest Science 59, 163-170.
Roberts J, Wallace JS, Pitman RM (1984) Factors affecting stomatal conductance of
Bracken below a forest canopy. Journal of Applied Ecology 21, 643-655.
Roberts NR, Dalton PJ, Jordan GJ (2005) Epiphytic ferns and bryophytes of Tasmanian
tree-ferns: A comparison of diversity and composition between two host species.
Austral Ecology 30, 146-154.
Romero P, Botía P (2006) Daily and seasonal patterns of leaf water relations and gas
exchange of regulated deficit-irrigated almond trees under semiarid conditions.
Environmental and Experimental Botany 56, 158-173.
Rossetto M (2008) From populations to communities: understanding changes in rainfor-
est diversity through the integration of molecular, ecological and environmental
data. Telopea 12, 47-58.
Sack L, Holbrook NM (2006) Leaf hydraulics. Annual Review of Plant Biology 57, 361-
381.
Salvucci ME, Crafts-Brandner SJ (2004) Mechanism for deactivation of Rubisco under
moderate heat stress. Physiologia Plantarum 122, 513-519.

-131-
Savitch LV, Massacci A, Gray GR, Huner NPA (2000) Acclimation to low temperature
or high light mitigates sensitivity to photoinhibition: roles of the Calvin cycle
and the Mehler reaction. Australian Journal of Plant Physiology 27, 253-264.
Schreiber U, Berry JA (1977) Heat-induced changes of chlorophyll fluorescence in in-
tact leaves correlated with damage of photosynthetic apparatus. Planta 136, 233-
238.
Seidlova L, Verlinden M, Gloser J, Milbau A, Nijs I (2009) Which plant traits promote
growth in the low-light regimes of vegetation gaps? Plant Ecology 200, 303-318.
Senger H, Wagner C, Hermsmeier D, Hohl N, Urbig T, Bishop NI (1993) The influence
of light intensity and wavelength on the contents of -carotene and -carotene
and their xanthophylls in green algae. Journal of Photochemistry and Photobiol-
ogy B-Biology 18, 273-279.
Sinsawat V, Leipner J, Stamp P, Fracheboud Y (2004) Effect of heat stress on the pho-
tosynthetic apparatus in maize (Zea mays L.) grown at control or high tempera-
ture. Environmental and Experimental Botany 52, 123-129.
Smith T, Huston M (1989) A theory of the spatial and temporal dynamics of plant
communities. Vegetatio 83, 49-69.
Snider JL, Choinski JS, Wise RR (2009) Juvenile Rhus glabra leaves have higher tem-
peratures and lower gas exchange rates than mature leaves when compared in
the field during periods of high irradiance. Journal of Plant Physiology 166,
686-696.
Stewart JR, Lister AM (2001) Cryptic northern refugia and the origins of the modern
biota. Trends in Ecology & Evolution 16, 608-613.
Taiz L, Zeiger E (2002) 'Plant physiology.' (Sinauer Associates, Inc., Publishers: Sun-
derland, Mass.)
Tallon C, Quiles MJ (2007) Acclimation to heat and high light intensity during the de-
velopment of oat leaves increases the NADH DH complex and PTOX levels in
chloroplasts. Plant Science 173, 438-445.
Tausz M, Hietz P, Briones O (2001) The significance of carotenoids and tocopherols in
photoprotection of seven epiphytic fern species of a Mexican cloud forest. Aus-
tralian Journal of Plant Physiology 28, 775-783.
Tausz M, Warren CR, Adams MA (2005) Dynamic light use and protection from excess
light in upper canopy and coppice leaves of Nothofagus cunninghamii in an old
growth, cool temperate rainforest in Victoria, Australia. New Phytologist 165,
143-155.
Tausz M, Wonisch A, Grill D, Morales D, Jimenez MS (2003) Measuring antioxidants
in tree species in the natural environment: from sampling to data evaluation.
Journal of Experimental Botany 54, 1505-1510.
Tazoe Y, von Caemmerer S, Badger MR, Evans JR (2009) Light and CO2 do not affect
the mesophyll conductance to CO2 diffusion in wheat leaves. Journal of Experi-
mental Botany 60, 2291-2301.
ter Steege H (1996) Winphot 5: a Programme to analyze vegetation induces, light, and
light quality from hemispherical photographs. In 'Tropenbos Guyana Reports 95-
2. Tropenbos Guyana Programme'. (Georgetown, Guyana)
Thayer SS, Björkman O (1990) Leaf xanthophyll content and composition in sun and
shade determined by HPLC. Photosynthesis Research 23, 331-343.

-132-
Tingey DT, Evans RC, Bates EH, Gumpertz ML (1987) Isoprene emissions and photo-
synthesis in 3 ferns - the influence of light and temperature. Physiologia Planta-
rum 69, 609-616.
Turner IM (1994a) Sclerophylly: primarily protective. Functional Ecology 8, 669-675.
Valladares F, Niinemets Ü (2008) Shade tolerance, a key plant feature of complex na-
ture and consequences. Annual Review of Ecology, Evolution, and Systematics
39, 237-257.
Valladares F, Pearcy RW (2002) Drought can be more critical in the shade than in the
sun: a field study of carbon gain and photo-inhibition in a Californian shrub dur-
ing a dry El Niño year. Plant Cell and Environment 25, 749-759.
Volkova L, Tausz M, Bennett LT, Dreyer E (2009) Interactive effects of high irradiance
and moderate heat on photosynthesis, pigments, and tocopherol in the tree-fern
Dicksonia antarctica. Functional Plant Biology In press.
von Caemmerer S, Evans JR, Hudson GS, Andrews TJ (1994) The kinetics of ribulose-
1,5-bisphosphate carboxylase/oxygenase in vivo inferred from measurements of
photosynthesis in leaves of transgenic tobacco. Planta 195, 88-97.
von Caemmerer S, Farquhar GD (1981) Some relationships between the biochemistry
of photosynthesis and the gas exchange of leaves. Planta 153, 376-387.
Warren CR (2008) Rapid measurement of chlorophylls with a microplate reader. Jour-
nal of Plant Nutrition 31, 1321-1332.
Warren CR, Bleby T, Adams MA (2007) Changes in gas exchange versus leaf solutes
as a means to cope with summer drought in Eucalyptus marginata. Oecologia
154, 1-10.
Watling JR, Robinson SA, Woodrow IE, Osmond CB (1997) Responses of rainforest
understorey plants to excess light during sunflecks. Australian Journal of Plant
Physiology 24, 17-25.
Watt MS, Kriticos DJ, Manning LK (2009) The current and future potential distribution
of Melaleuca quinquenervia. Weed Research 49, 381-390.
Weaver PRL (2008) Dwarf forest recovery after disturbances in the Luquillo Mountains
of Puerto Rico. Caribbean Journal of Science 44, 150-163.
Wellburn AR (1994) The spectral determination of chlorophyll a and chlorophyll b, as
well as total carotenoids, using various solvents with spectrophotometers of dif-
ferent resolution. Journal of Plant Physiology 144, 307-313.
White RA, Weidlich WH (1995) Organization of the vascular system in the stems of
Diplazium and Blechnum (Filicales). American Journal of Botany 82, 982-991.
Wise RR, Olson AJ, Schrader SM, Sharkey TD (2004) Electron transport is the func-
tional limitation of photosynthesis in field-grown Pima cotton plants at high
temperature. Plant Cell and Environment 27, 717-724.
Wullschleger SD (1993) Biochemical limitations to carbon assimilation in C3 plants - a
retrospective analysis of the A/Ci curves from 109 species. Journal of Experi-
mental Botany 44, 907-920.
Yamamoto Y, Aminaka R, Yoshioka M, Khatoon M, Komayama K, Takenaka D, Ya-
mashita A, Nijo N, Inagawa K, Morita N, Sasaki T, Yamamoto Y (2008) Quality
control of photosystem II: impact of light and heat stresses. Photosynthesis Re-
search 98, 589-608.

-133-
Zhu XG, Ort DR, Whitmarsh J, Long SP (2004) The slow reversibility of photosystem
II thermal energy dissipation on transfer from high to low light may cause large
losses in carbon gain by crop canopies: a theoretical analysis. Journal of Ex-
perimental Botany 55, 1167-1175.
Zsófi Z, Váradi G, Bálo B, Marschall M, Nagy Z, Dulai S (2009) Heat acclimation of
grapevine leaf photosynthesis: mezo- and macroclimatic aspects. Functional
Plant Biology 36, 310-322.

Minerva Access is the Institutional Repository of The University of Melbourne
Author/s:
Volkova, Liubov Vladimirovna
Title:
Ecophysiology of the tree fern species Dicksonia antarctica Labill and Cyathea australis (R.
Br.) Domin
Date:
2009
Citation:
Volkova, L. V. (2009). Ecophysiology of the tree fern species Dicksonia antarctica Labill and
Cyathea australis (R. Br.) Domin. PhD thesis, Dept. of Forest and Ecosystem Science,
Melbourne School of Land and Environment, The University of Melbourne.
Persistent Link:
http://hdl.handle.net/11343/37893
File Description:
Ecophysiology of the tree fern species Dicksonia antarctica Labill and Cyathea australis (R.
Br.) Domin
Terms and Conditions:
Terms and Conditions: Copyright in works deposited in Minerva Access is retained by the
copyright owner. The work may not be altered without permission from the copyright owner.
Readers may only download, print and save electronic copies of whole works for their own
personal non-commercial use. Any use that exceeds these limits requires permission from
the copyright owner. Attribution is essential when quoting or paraphrasing from these works.