effect of alpha keto acids including sodium pyruvate on ... · effect of alpha keto acids including...
TRANSCRIPT

Effect of Alpha Keto Acids Including Sodium Pyruvate on Reducing and
Regulating the Inflammatory AgentsNeeded in the Healing of Infected and
Non-Infected Wounds
Draft/Confidential

Introduction
Wounds are internal or external bodily injuries or lesionscaused by mechanical, chemical, viral, bacterial or thermal means,which disrupt the normal continuity of structures. Such bodilyinjuries include contusions, which are wounds in which the skinis unbroken, incisions, i.e., wounds in which the skin is broken bya cutting instrument, and lacerations, which are wounds in whichthe skin is broken by a dull or blunt instrument.
Wound healing consists of a series of processes whereby injuredtissues are repaired, specialized tissue is generated, and new tissueis reorganized. Wound healing consists of three major phases: (a)an inflammation phase (0-3 days), (b) a cellular proliferation phase(3-12 days), and (c) a remodeling phase (3-6 months).
During the inflammation phase, platelet aggregation and clot-ting form a matrix which traps plasma proteins and blood cells toinduce the influx of various types of cells. Wounds produce oxy-gen radicals. Mammalian cells are continuously exposed to acti-vated oxygen species such as superoxide (02-), hydrogen peroxide(H2O2), hydroxyl radicals (OH), peroxynitrite (ONOO-) and sin-glet oxygen (1O2). In vivo, these reactive oxygen intermediatesare generated by cells in response to aerobic metabolism,catabolism of drugs and other xenobiotics, ultraviolet and x-rayradiation, and the respiratory burst of phagocytic cells (such aswhite blood cells) to kill invading bacteria and viruses introducedthrough wounds. The toxic by-products generated from thecatabolism of drugs to treat infected and cancerous cells activatethe inflammatory process generally through the activation of NF-Kappa B, which can delay healing. Hydrogen peroxide, for exam-ple, is produced during respiration of most living organisms espe-cially by stressed and injured cells.
It is at this time that peroxynitrite is produced and causes NF-kappa B to be over expressed, thus delaying the healing process.This over expression of peroxynitrite will also destroy drugs need-ed to treat various diseases including infected wounds. During thecellular proliferation phase, new connective or granulation tissueand blood vessels are formed. During the remodeling phase, gran-ulation tissue is replaced by a network of collagen and elastin fibersleading to the formation of scar tissue. Most wounds also producepain, swelling, itching, ischemia, crusting, erythema, and scarring,which is caused by the over expression of NF-kappa B from theover expression of peroxynitrite and the metabolites it produceswhen it reacts with drugs, all of which are undesirable.
Thus, the ability of pyruvate and other alpha-keto acids to upregulate or down regulate Nitric Oxide levels in the body and toprotect Nitric Oxide from other oxygen radicals, allows NitricOxide to deactivate NF-kappa B, which, in turn, will reduce NOlevels and reduce inflammation. In wounds and in HSV-1 infect-ed cells, sodium pyruvate along with alpha-keto isovalerate atthe higher concentrations, reduced viral loads, increased the syn-thesis of Nitric Oxide and in combination with antiviral agents,eliminated the virus completely from the infected cells.
NF-kappa B
Recent reports show that deactivating a protein called I-Kappa-Bkinase beta (IKK-beta) inside the cell stops cancer progression inits tracks along with inflammation. IKK-Beta normally plays a rolein healing.(37,38,64) During an injury or infection immune systemmolecules, including oxygen radicals, which deplete glutathione,activate IKK-beta.(37,64) Once stimulated, this protein keeps cellsalive and growing. IKK-beta also promotes inflammation in dam-aged tissues. IKK-beta is also known to be elevated in bronchialepithelial cells(37,64) and many other diseases, infected wounds,including cancer. IKK-beta also assists tumor growth in differenttypes of cells by subverting the programmed cell death that wouldotherwise prevent tumor formation, thus IKK-beta promotestumor development and growth through inflammation. IKK-betaworks by activating the transcription factor nuclear factor-kappa B(NF-kappa B).(39,40,64) NF-kappa B resides in the cytoplasm as aninactive dimer, consisting of two subunits, bound to an inhibitoryprotein.(44, 49,50) The inhibitory protein is degraded in response tovarious environmental stimuli, such as pro-inflammatorycytokines, viruses, and oxygen radicals. This allows NF-kappa B totranslocate to the nucleus where it activates genes that play a rolein the regulation of inflammatory responses, including genes thatencode pro-inflammatory cytokines IL-2, TNF, IL-6, IL-11, IL-17.NF-kappa B also regulates NO synthetase, and genes that inhibitapoptosis, which plays a major role in tumor growth and sur-vival.(44, 49,50) NF-kappa B when activated stops programmed cel-lular death, activates other inflammatory mediators, and increasesNitric Oxide synthesis and production.(38-40,44,64) In patients withinflammatory diseases including infections, lung diseases like cys-tic fibrosis, COPD, ILD, and cancer, NF-kappa B activation is exag-gerated as are other inflammatory components.(37,38,44) Hydrogenperoxide and other oxygen radicals, such as nitrogen dioxide and
2Draft/Confidential
Effect of Alpha-keto Acids Including Sodium Pyruvate onReducing and Regulating the Inflammatory Agents Hydrogen
Peroxide, Peroxynitrite, and Nitric Oxide To Reduce theInflammatory Process and Enhance the Healing of Infected
Wounds in Patients

peroxynitrite also activates NF-kappa B as does cellular glu-tathione depletion. (42-45,64) Oxygen radicals also damage p53, aprotein central to the inhibition of tumor growth. Its function is toensure that every time a cell divides, each of the two daughter cellsgets an undamaged copy of the original set of genes, free of muta-tions. If the cell contains damaged DNA, the p53 protein stops cel-lular division. Only when repairs are complete will p53 permitDNA replication to begin. But if the damage is too extensive torepair, p53 blocks the cell from dividing and commands the cell todie.(34) The p53 protein triggers the process of programmed celldeath. In at least 50 to 80 percent of all cancers, p53 is damagedand apparently useless.(34) Thus any molecule that deactivateseither IKK- beta or NF-kappa B would inhibit inflammation thatcould also lead to tumor development and survival.(41) Antioxi-dants have been shown to inhibit NF-kappa B thus inhibit inflam-mation and to protect DNA and proteins like p53 from oxidativedamage(75) thus facilitate the healing process. Antioxidants likevitamin C, vitamin A, acetylcysteine, vitamin E, glutathione, andpyruvate down regulate and inhibit NF-kappa B.(40-52, 55,64) Highlevels of Nitric Oxide also inhibit NF-kappa B. Thus a technologythat can reduce inflammation, and inhibit NF-kappa B fromincreasing inflammatory mediators would have tremendousadvantages in the therapeutic arena, especially in the treatement ofinfected wounds.
Nitric Oxide anti-inflammatory properties
Nitric Oxide (NO), an oxidation product of nitrogen, is pro-duced normally by many cell types, including endothelial cellsand macrophages.(1,2,3,12,15,16,17,26,27) Nitric Oxide has functionsranging from neurotransmission, anti-infectives, vasodilatation tobeing tumoricidal.(12-18,72) Nitric Oxide also possesses antiinflam-matory effects, which may be exerted via its ability to inhibit thetranscription factor, NF-kappa B(51) and other inflammatorycytokines.(73) The most commonly proposed mode of action forthe inhibition of NF-kappa B involves interference with NF-kappaB binding to DNA.(53) Because IKK-beta is subject to redox regu-lation, NO will inhibit NF-kappa B activation by the inactivationof IKK beta. High doses of NO also impaired the TNF-alpha-induced DNA binding activity of NF-kappa B.(55,64) High dosesof NO also repressed the TNF-alpha induced transactivation byNF-kappa B.(53-55) High doses of Nitric Oxide will inhibit NF-kappa B if available and is not transformed into peroxynitrite,which will activate NF-kappa B rather than deactivate it. Antioxi-dants, including pyruvate have been shown to protect NitricOxide from other oxygen radicals, which allows Nitric Oxide todeactivate NF-kappa B. Nitric Oxide also produces clinically use-ful bronchodilation(1) and is also used by the body to kill bacteria,fungal infections, viral infections, and tumors.(21,72) Nitric Oxidecan kill these cell types because bacterial, viral, and tumor cellshave no defenses against Nitric Oxide. Normal mammalian cellscan cope with normal levels of Nitric Oxide by using enzyme sys-tems to use or deactivate elevated cellular levels of NitricOxide.(21-25) Nitric Oxide is the main mediator of the tumoricidalaction of activated macrophages.(22-25,72) While innumerablepapers have been written on Nitric Oxide, the role of Nitric Oxide
in tumor biology is not completely understood. Nitric Oxideappears to have both tumor promoting and inhibiting effects.(24)
Recent reports have implicated the reactive oxygen species madefrom Nitric Oxide during the inflammatory process, particularlyperoxynitrite, and nitrogen dioxide as being the tumor promotingagents, not Nitric Oxide itself.(3,13,21,30) Nitric Oxide dose notmediate but inhibits transformation and tumor growth.(72)
Activation of the inflammatory process by Oxygen Radicals
Hydrogen peroxide is the oxygen radical that appears to have themajor effect on delaying the healing process.(1-4,64-67) Superoxideand hydrogen peroxide also cause vascular constriction.(1) H2O2markedly potentiates the cytotoxic effects of eosinophil derivedenzymes such as 5,8,11,14,17-eicosapentaenoic acid.(1-4,64-67) Excesssuperoxide anions and hydrogen peroxide, and their oxidationproducts specifically peroxynitrite, produced during the inflamma-tory phase of an injury, will destroy healthy tissue surrounding thesite and will mitigate the positive bronchodilation effect of NitricOxide.(19) Oxygen radicals can also initiate lipid peroxidationemploying arachidonic acid as a substrate producing prostaglandinsand leukotrienes. H2O2 can induce arachidonic acid metabolism inalveolar macrophages.(10,11,19) Hydrogen peroxide and other oxy-gen radicals, such as nitrogen dioxide and peroxynitrite also acti-vates NF-kappa B as does cellular glutathione depletion.(42-44,55,64,67-72) Oxygen radicals lower cellular levels of glu-tathione. Oxygen radicals also damage p53, a protein central to theinhibition of tumor growth and needed by cells to facilitate healingand DNA repair. Oxygen radicals also produce 8-isoprostanes,which are potent renal and pulmonary artery vasoconstrictors,bronchoconstrictors, and induce airflow obstructions.(19,20,64, 66-69)
Because oxygen radicals contribute to the instability of Nitric Oxide,the addition of superoxide dismutase (SOD) or catalase or VitaminE(21) protect Nitric Oxide to produce its desired bronchodilation andenhanced healing of wounds.(2) These active oxygen species caninjure cells. An important example of such damage is lipid peroxi-dation which involves the oxidative degradation of unsaturatedlipids. Lipid peroxidation is highly detrimental to membrane struc-ture and function and can cause numerous cytopathological effectsincluding the activation of NF-kappa B. Cells defend against lipidperoxidation by producing radical scavengers such as superoxidedismutase, catalase, and peroxidase. Injured cells have a decreasedability to produce radical scavengers. Excess hydrogen peroxideespecially peroxynitrite, can react with DNA to cause backbonebreakage of the DNA, produce mutations, as well as the alterationand liberation of the bases. Hydrogen peroxide can also react withpyrimidines to open the 5, 6-double bond, which inhibits the abilityof pyrimidines to hydrogen bond to complementary bases, Hallaen-der, et al. (1971). Such oxidative biochemical injury can result in theloss of cellular membrane integrity, reduced enzyme activity,changes in transport kinetics, changes in membrane lipid content,and leakage of potassium ions, amino acids, other cellular material,and the formation of excess keloid and scar formation.
In patients with infected or non infected wounds, and somecancers, NF-kappa B activation is exaggerated as are other
3Draft/Confidential

inflammatory components including oxygen radicals that keepNF-kappa B activated.(37,38,44) Hydrogen peroxide, and otheroxygen radicals, such as nitrogen dioxide and peroxynitrite canalso be generated by cancer drugs like bleomycin, activate NF-kappa B and deplete cellular glutathione levels.(42-45,64) NF-kappa B regulates the inflammatory responses, including genesthat encode pro-inflammatory cytokines IL-2, TNF, IL-6, IL-11,IL-17. NF-kappa B also regulates NO synthetase, and genes thatinhibit apoptosis, which plays a major role in tumor growth andsurvival.(44, 49,50)
Similarly, excess Nitric Oxide, which becomes peroxynitrite,has been implicated in airway inflammation caused by infections,which decreases FEV1 va lues and may worsen gas ex-change.(26,28,29,70) This is most likely due to the fact that NitricOxide can become a toxic oxidant when it reacts with excess oxy-gen radicals like hydrogen peroxide to produce nitrogen dioxide(NO2)(1-3) and peroxynitrite (ONOO). Oxygen radicals are pro-duced by many cell types including epithelial cells, macrophages,leukocytes, monocytes, and fibroblasts. Oxygen radicals, such assuperoxide (O2) and hydrogen peroxide, destroy Nitric Oxideand produce the toxic NO2 and peroxynitrite.(1-3) Nitrogen diox-ide is a major air pollutant and a deep lung irritant. Nitrogendioxide is formed in combustion processes, either directly orthrough secondary oxidation of Nitric Oxide.(7) Nitrogen dioxidecauses pulmonary in f lammat ion , lower leve ls o f lungantioxidants(9), including glutathione, deterioration of respiratorydefense mechanisms, and increases susceptibility to respiratorypathogens and cancer.(1,7) Nitrogen dioxide can also increase theincidence and severity of respiratory infections, can reduce lungfunction, and can aggravate the symptoms of asthmatics or sub-jects with COPD.(1,8)
Peroxynitrite ion and peroxynitrous acid, formed from theinteraction of Nitric Oxide and superoxide anions, specificallyhydrogen peroxide, are strong oxidant species that work againstNitric Oxide by inducing single-strand breaks in DNA, increas-ing the levels of inflammatory mediators and enhancing tumorformation and growth(21,64, 65, 71) rather than death. Peroxyni-trites, which cause bronchial constriction, are involved in lunginjury through the production of chemokines and contribute toviral pathogenesis and they enhance viral mutations.(2,3,13,30,65)
Nitric Oxide, when combined with superoxide anions and/orhydrogen peroxide to form peroxynitrite, can also generate thehighly reactive hydroxyl anion (OH), which also lower cellularlevels of glutathione. According to epidemiological studies, thepopulation group most susceptible to these adverse effects issmall children, or older people.(7) Even though children haveincreased amounts of Nitric Oxide at wound sites, there is per-suasive evidence that higher levels of Nitric Oxide are decreasedby the overproduction of oxygen radicals during the inflammato-ry process.(1-8) This becomes a problematic situation for whichthe only solution is denied by the circumstance inherent in theproblem. The underlying chronic inflammatory process in infec-tions, which induces Nitric Oxide synthesis, also produces excessoxygen radicals, which will destroy Nitric Oxide.(5,6) The use ofan irritant has been shown to enhance Nitric Oxide productionby alveolar macrophages in rats, which also produces an
increased level of oxygen radical that can react directly withNitric Oxide to produce NO2 and peroxynitrites.(1-3, 5,13)
Sodium Pyruvate
Sodium pyruvate is an antioxidant that reacts directly with oxy-gen radicals like hydrogen peroxide and peroxynitrite, neutraliz-ing them and thusly protecting DNA and other cellularcomponents, such as glutathione, lipids and proteins.(35,56-65,70,71)
In macrophages, as well as other cell lines, sodium pyruvate regu-lates the production and level of inflammatory mediators includ-ing oxygen radical production and also regulates the synthesis ofNitric Oxide.(8,49,50) Sodium pyruvate is administered to patientsfor a variety of medical disorders such as the treatment of Friedre-ich’s ataxia (83), and as a constituent in a therapeutic solutionused in open heart surgery.(84) It has been administered by sever-al routes included intravenous (85,86), topical (for hyperkeratoticdisorders) (87), and oral (dietary supplements). (87-89) In all cases,the administration of sodium pyruvate was shown to reduceinflammation and enhance healing. Low concentrations of pyru-vate decreased the expression of several proinflammatory genes,including NF-kappa B, activation of inducible NO synthasemRNA, TNF, cyclo-oxygenase, interleukin 6 and 10mRNA induc-tion.(32,33,44,49,50,64) Sodium pyruvate inhibited hydrogen perox-ide induced transcription of NF-kappa B while protecting cellularglutathione.(44,64) Sodium pyruvate blocked the p38 MAPK path-way and activated the ERK pathway which regulates the expres-sion of genes believed to prevent apoptosis and promote cellsurviva.(44,64) Sodium pyruvate also inhibited the hepatocytesNitric Oxide synthesis (27), and at higher concentrations causedup regulation of inducible Nitric Oxide synthase mRNA in intesti-nal cells and in cardiac monocytes.(8,26,28) Pyruvate specificallylowered the overproduction of superoxide anions, H2O2 and NOin white blood cells. (31,56-64)
Pyruvate also increased cellular levels of glutathione, a majorcellular antioxidant (8), needed to prevent activation of NF-kappaB, which activates the inflammatory process. It was recently dis-covered that glutathione is reduced dramatically in antigen-induced asthmatic patients (10) and inhaled glutathione does notreadily enter cells. Pyruvate does enter all cells via a transport sys-tem and can also cross the blood brain barrier. Oxygen radicals areinvolved in the induction and progression of malignancy andpyruvate, a known scavenger of oxygen radicals, has been impli-cated in cancer prevention. (32,33,57,64) Pyruvate inhibited thegrowth of implanted tumors and reduced lung metastases,decreased the number of DNA breaks caused by H2O2 by 40%.(32)
Excess sodium pyruvate beyond that needed to neutralize oxygenradicals will enter the bronchial and lung cells. All cells have atransport system that allow cells to concentrate pyruvate at higherconcentrations than serum levels. In monocytes cultures, the pro-duction of H2O2 was regulated by the level of sodium pyruvatesupplied in the culture medium. At 1mM and higher concentra-tions, the production of H202 was decreased by 30%. At 10mMconcentrations, the production of H2O2 was decreased by 60%. (64)
Pyruvate controls the positive and negative effects of NitricOxide at higher levels. Too high a level of Nitric Oxide is detrimen-
4Draft/Confidential

tal to cells. When higher levels of NO are produced, even by activa-tion of inducible NO synthase mRNA from higher levels of pyru-vate, it is kept at bay by pyruvate. Nitric Oxide affects cells byincreasing levels of cGMP and ADP, and requires an acid pH rangein which to work (12). Higher levels of pyruvate raises the pH level,increases levels of ATP, decreasing levels of ADP and cAMP, andincreases levels of GTP, while decreasing levels of cGMP.
Sodium pyruvate, and other alpha-keto acids are reactive oxy-gen radical antagonist, and other alpha-keto acids decreased theexpression of several pro-inflammatory genes, including NF-kappa B, activation of inducible NO synthase mRNA, TNF,cyclo-oxygenase, interleukin 6 and 10mRNA induction in numer-ous cell lines and in various animal models.(32,33,44,49,50,64) Sodi-um pyruvate, an effective scavenger of reactive oxygen species(ROS), acts to inhibit tumor necrosis factor-alpha production andNF-kB signaling pathways, reduces circulating levels of HMGB1(high mobility group b1), and decreases COX-2 (cyclo-oxyge-nase-2), iNOS (inducible Nitric Oxide synthesis), and IL-6 (inter-leukin-6) mRNA expression in liver, ileal mucosa, and colonicmucosa.(84) As reported, decreasing oxygen radicals, includinghydrogen peroxide, which reacts with Nitric Oxide to produceperoxynitrite, will deactivate NF-kappa B and reduce inflamma-tion. 1 Kappa-B-Kinase (IKK-beta) also promotes inflammation indamaged tissues. IKK-beta is known to be elevated in bronchialepithelial cells in cystic fibrosis(37,64) and in other lung diseases,infected cells, and in cancer. IKK-beta works by activating thetranscription factor nuclear factor-kappa B (NF-kappa B).(39,40,64)
Thus any molecule that deactivates either IKK- beta or NF-kappaB would inhibit inflammation (41). Nitric Oxide also possessesantiinflammatory effects, which can be exerted via its ability toinhibit the transcription factor, NF-kappa B (51) and other inflam-matory cytokines.(73) The most commonly proposed mode ofaction for the inhibition of NF-kappa B involves interference withNF-kappa B binding to DNA. (53) Because IKK-beta is subject toredox regulation, NO will inhibit NF-kappa B activation by theinactivation of IKK beta. Thus, the ability of pyruvate to up regu-late or down regulate Nitric Oxide levels in the body and to pro-tect Nitric Oxide from other oxygen radicals, allows Nitric Oxideto deactivate NF-kappa B, which, in turn, will reduce NO levelsand reduce inflammation. In HSV-1 infected cells, sodium pyru-vate at the higher concentrations, reduced viral loads, increasedthe synthesis of Nitric Oxide and in combination with antiviralagents, eliminated the virus completely from the infected cells.
Alpha-keto-isovalerate metabolic background
Alpha-keto-isovalerate (CH3)2CHCOCOOH is a fundamentalintermediate in protein synthesis, i.e., the biosynthesis of theamino acids leucine, valine and their metabolism. The formationof leucine begins by condensation of alpha-keto-isovaleric acid(which is also the precursor of valine) with acetyl CoA to yieldalpha-isopropylmalic acid. The subsequent steps are similar tothose leading from citric acid to keto-glutaric acid in the tricar-boxylic acid cycle. The two major sources of acetyl coenzyme Aare derived from the metabolism of glucose and fatty acids. Gly-colysis consists of a series of transformations wherein each glu-
cose molecule is transformed in the cellular cytoplasm into twomolecules of pyruvic acid. Pyruvic acid may then enter the mito-chondria where it is oxidized by coenzyme A in the presence ofenzymes and cofactors to acetyl coenzyme A. Pyruvic acid canalso be converted in several steps to keto-isovaleric acid leading tothe formation of valine. Keto-isovalerate inhibits pain, erythema,itching and swelling. It is known, to react with H2O2 to produceisobutyric acid and seems to reduce prostaglandins. In virallyinfected cells, keto isovalerate will inhibit amino acid up take bythe cells, inhibiting the virus from producing a protein coat.
Keto-butyrate is a superior moisturizing agent over all theother keto acids. It reacts with H2O2 to produce propionic acidwhich has been shown to be an anti-fungal agent and can be usedwith other keto acids to enhance the moisture in tissues.
Keto-glutarate is taken up by neurons and fibroblasts andtherein increases neuron survival and collagen deposition byfibroblasts. In combination with pyruvate, this keto acid pro-duced the greatest neuron survival and regeneration. It reactswith H2O2 to produce succinic acid which inhibits mast cellsfrom releasing histamines.
Keto-caproate will disrupt cellular membranes and mucus. Itreacts with H2O2 to produce valerate, a food source. It can beused with oxaloacetate to inhibit cancer cells from growing whileat the same time allowing normal cells to grow.
Keto-adipate dissolves excess mucus. It reacts with H2O2 toproduce glutaric acid.
Oxaloacetate is part of the Krebs citric acid cycle and isfound in the mitochondria of cells. We are the first to show thatoxaloacetate inhibits keloid formation, angiogenesis, andexcess infiltration of leukocytes. Its properties can be used inburns to prevent progressive burn ischemia due to thermalinjuries which causes delayed tissue loss in surroundinghealthy tissue. It reacts with H2O2 to produce malonate, acompetitive inhibitor of succinate dehydrogenase, which effec-tively inhibits respiration and the further production of H2O2.Succinate on the other hand increases the production of hydro-gen peroxide. Malonate is effectively transported out of themitochondria via glutathione. This is an protective method thatregulated the production of hydrogen peroxide. When weused it in combination with keto-isovalerate or pyruvate onburns, excess WBC infiltration is reduced and healing isenhanced. It can be used to treat patients undergoing radiationtherapy to inhibit regrowth of the tumor, while at the sametime healing normal cells.
Effect of Sodium pyruvate to reduce inflammation due to bleomycin
Bleomycin was administered to rats to cause inflammatoryinjury to the body, which then resulted in decreased healingtimes. Bleomycin enhances the production of oxygen radicalsin wounds, enhances the numbers of white blood cells thatinvade the various organs and activates NF-kappa B, whichincreases the inflammatory process. Sodium pyruvate, 5.0 mM,in 0.9% sodium chloride solution administered by injection ortopically to wounds to see if the drug regimen had any effect
5Draft/Confidential

on the progression of the injury. The rats were sacrificed 24hours, 72 hours, one week, and two weeks post injury. The ratssacrificed at two weeks were administered sodium pyruvatethe third and seventh day post bleomycin insult, while theother rats received only a single administration of sodiumpyruvate.
There was no effect on the progression of the injury in the ratssacrificed at 24 hours, 72 hours or one week. However, signifi-cant improvement was observed in the rats administered thesodium pyruvate and sacrificed two weeks after bleomycininsult, compared to the sodium chloride control. There was a sig-nificant (p<0.01) reduction in total white blood cells indicating areduction in airway inflammation. Healing times were alsoreduced by 40%.
Clinical Studies
Injury to human skin Epidermal keratinocytes are the specialized epithelial cells of
the epidermis which synthesize keratin, a scleroprotein which isthe principal constituent of epidermis, hair, nails, horny tissue,and the organic matrix of the enamel of teeth. Mammalian epi-dermal keratinocytes constitute about 95% of the epidermal cellsand together with melanocytes form the binary system of the epi-dermis. In its various successive stages, epidermal keratinocytesare also known as basal cells, prickle cells, and granular cells.
Monocytes are mononuclear phagocytic leukocytes whichundergo respiratory bursting and are involved in reactive oxygenmediated damage within the epidermis. Leukocytes are WBC orcorpuscles which may be classified into two main groups: granu-lar leukocytes (granulocytes) which are leukocytes with abun-dant granules in the cytoplasm and nongranular leukocytes(nongranulocytes) which are leukocytes without specific gran-ules in the cytoplasm and which include the lymphocytes andmonocytes. Phagocytic cells are cells which ingest microorgan-isms or other cells and foreign particles. Monocytes are alsoknown as large mononuclear leukocytes, and hyaline or transi-tional leukocytes.
Epidermal keratinocytic cells and monocytic cells have multi-ple oxygen generating mechanisms and the degree to which eachtype of mechanism functions differs in each type of cell. In mono-cytes, for example, the respiratory bursting process is more pro-nounced than in ep idermal kera t inocytes . Hence , thecomponents in the therapeutic wound-healing compositions ofthe present invention may vary depending upon the types ofcells involved in the condition being treated.
Human skin is know to produce the high amounts of NitricOxide and skin cells also produce hydrogen peroxide. This caus-es a problem in that high levels of peroxynitrite are also pro-duced. Burned, damaged or infected skin produces even higherlevels of peroxynitrite, which damages non damaged cells andimmune cells and drugs needed to treat the damaged skin. Anexample of this is the use of Acyclovir on cold sore lesions. Acy-clovir is effective when taken orally or intervenously, but topical-ly its effect has been minimal. Infections with herpes simplexvirus I (HSV-1) induces a persistent nuclear translocation of NF-
kappa B, which is dramatically enhanced by peroxynitrite. Theactivation of NF-kappa B promotes efficient replication by HSV.In epithelial cells HSV-1 induces NF-kappa B causing persistentactivation of NF-kappa B, which is a critical regulator of HSV-1replication in skin. In AIDs patients, HIV-1 also triggers and acti-vates NF-kappa B and AIDS patients have elevated levels of per-oxynitrite, which contributes to the etiology of AIDS relateddementia, persistent immunosuppression and Kaposi’s sarcoma.Peroxynitrite have also been shown to be very destructive to CD4and CD8 cells. We have discovered that alpha-keto acids candecrease the levels and production of peroxynitrite, while pro-tecting Nitric Oxide which can enhance the viral kill rate, whileprotecting drugs like Acyclovir. We discovered that oxaloacetate,found predominately in mitochondria, can react with excesshydrogen peroxide to produce malonate, which decreases cellu-lar respiration and inhibits the over synthesis of hydrogen perox-ide. This will lower the production of peroxynitrite. Pyruvateincreases the production of glutathione which is needed by thecells to eliminate malonate from the body. This mechanism of cel-lular control is used to control the production and level of perox-ynitrite needed to fight infections and protect drugs. Ketoisovalerate inhibits amino acid up take by virally infected cellswhich decreases the viruses ability to produce a protein coat.
The problem is two fold. One, people who need to reduce thelevels and production of peroxynitrite which destroys drugsand immune cells, to increase cellular levels of glutathione,reduce inflammation in their infected and noninfected woundsto minimize damage and DNA mutations that cause diseases,and to protect drugs needed to treat these conditions by pre-venting their destruction and the formation of toxic agents thatactivate the inflammatory process. Secondly, when drugs aredestroyed into toxic metabolites, one needs a method toremove them and inhibit their activation of NF-kappa B. Third-ly, if cancer has occurred a method to control its growth andenhance cancer cell death (apoptosis). It was recently shownthat deactivating a protein called I-Kappa-B kinase beta (IKK-beta) inside the cell stops inflammation and cancer progressionin its tracks. Once stimulated, this protein keeps cells alive andgrowing. IKK-beta also promotes inflammation in damaged tis-sues. IKK-beta works by activating the transcription factornuclear factor-kappa B (NF-kappa B).(39,40,64) NF-kappa Bwhen activated stops programmed cellular death, and activatesother inflammatory mediators.(38-40,64) Hydrogen peroxideand other oxygen radicals, such as nitrogen dioxide and perox-ynitrite also activates NF-kappa B as does cellular glutathionedepletion.(42-44,64) Oxygen radicals also damage p53, a proteincentral to the inhibition of tumor growth. When DNA damageis to extensive to repair, p53 blocks the cell from dividing andcommands the cell to die.(34) We discovered that toxic break-down products from cellular metabolites and drugs can alsoactivate NF-kappa B and that alpha-keto acids will eliminatethis activation process to facilitate healing. Thus any moleculethat can reduce or regulate the production of peroxynitrite,thus deactivates either IKK- beta or NF-kappa B woulddecrease inflammation at wound sites and inhibit tumor sur-vival. We have shown that these alpha-keto acids singly or in
6Draft/Confidential

combination will inhibit NF-kappa B thus inhibit inflammationand to protect DNA and proteins like p53 from oxidative dam-age. This was the first time that any one has shown that drugsused to treat diseases are themselves activators of NF-kappa Band that their by-products produced when treated with oxygenradicals, specifically peroxynitrite, and other inflammatorymediators will delay the healing process by the further activa-tion of NF-kappa B.
Example 1This experiment was designed to show that medicines used to
treat infections and cancer can be destroyed by the endogenous pro-duction of peroxynitrite, and that their by-products will cause theover production of peroxynitrite which will cause the over expres-sion of NF-kappa B, thus delay the healing process. Various antibac-terial, antifungals, anticancer and antiviral drugs were placed intosolutions containing peroxynitrite, then one hour later the solutionswere tested for peroxynitrite activity using the methods described inreferences 35, 65, 70. None of the peroxynitrite solutions containingthe drugs and their by products showed any peroxynitrite activityafter one hour. When the peroxynitrite treated drugs were placed ondermabraided skin they caused irritations do to the toxic break-down products that were produced. They also delayed healingwhen compared to peroxynitrite treated drugs that were tested withthe addition of alpha-keto acids (see tables 1-4 column 1 and 2).When peroxynitrite is tested alone on derabriaded skin it activatesinflammation and delays healing. Peroxynitrite drug by productsare detoxified by alpha-keto acids and do not increase inflammation.Healing was at least 40% better with the keto acids.
In the second experiment , various wound sites were made ona patient’s arm with strip tape which produces a shallow dermabrasion. Derm abrasion will activate the production of NitricOxide and hydrogen peroxide and generate peroxynitrite whichactivates NF-kappa B which will produce inflammatory agentsthat will increase erythema, swelling, and delay healing. The var-ious drugs were tested by them selves and in combination withthe five keto acids and compared (see tables 1-4 columns A andB). Time to healing was determined visually when redness and reepithelization occurred.
As can be seen from the results, drugs can activate the inflam-matory process through NF-kappa B. Drugs that are destroyedby peroxynitrite produce toxic by-products that will activate theinflammatory process as shown in the above experiment. It wasalso clear from the data, that alpha-keto acids will mitigate thedamage produced from the toxic drug byproducts and decreaseinflammatory process that will increase healing. The keto acidsreduced the time to healing in all cases. Unexpectedly, the com-bination of pyruvate and keto-isovalerate were synergistic andproduced the best results in all categories. Oxaloacetate pro-duced the best type of collagen deposition. Oxaloacetate whenmixed with pyruvate produced unexpected results when used incombination with peroxynitrite treated doxorubicin on der-mabraided skin. The results were superior to pyruvate alone.Oxaloacetate reduces the production of hydrogen peroxide pro-duced by doxorubicin, thus reduced the production of peroxyni-trite. Various keto acids could be mixed to varying effects.
What we discovered was that these alpha-keto acids, somehowthrough an new mechanism, mitigated the damage from drugsand the toxic metabolites of these drugs that appeared to haveactivated NF-kappa B through peroxynitrite.
There are a number of keto acids known in the art and whereasall are extracellular antioxidents. Each is unique in their interac-tion and effect on mammalian cells. Pyruvate enhances re-epithe-lization and angiogenesis of new capillaries. It has the ability toenter cells to protect cellular components including DNA.
Example 2Throughout medical history, there has been interest in shorten-
ing wound-healing by reducing the duration of associated symp-toms as well as diminishing resultant scars. This study looks atwound-healing after cosmetic surgery, but is potentially applica-ble to a broad spectrum of wounds. In jured or s t ressed ce l l sundergo free radical reactions more quickly than uninjuredhealthy cells. During wound healing, infiltrating leukocytesrelease activated oxygen species including O2, H2O2, OH-,ONOO-, and 1O2. Various agents such as burns, ultravioletlight, ionizing radiation, toxic agents and drugs, and invadingbacteria, viruses, cancer stimulate the release of these free radi-cals and activate NF-kappa B which if over expressed will delayhealing. Cells defend against free radicals by producing free rad-ical scavengers and antioxidants. Antioxidants are the main anti-dote to oxygen free radicals. Known antioxidants includevitamin E, pyruvate, vitamin C, and other compounds andenzymes. These antioxidants may be depleted resulting in “un-neutralized” oxidative biochemical injuries. These injuries leadto a final common pathway that impairs and delays wound heal-ing.
Laser skin resurfacing has become a very popular and preva-lent cosmetic procedure to rejuvenate photoaged skin. Candi-dates for this procedure generally have a greater than averageamount of photoaging and likely have depleted antioxidants. Inaddition, the greatest complaint surrounding these procedures isthe long period of wound-healing and prolonged erythema pain,swelling, and crusting. This is do to the increased levels of cellu-lar toxic agents produced from peroxynitrite that activate NF-kappa B. Alster, et al. recently reported application of stabilized10% topical L-ascorbic acid in aqueous formulation, which result-ed in a significant decrease in post-carbon dioxide (CO2) laserresurfacing erythema by the 8th postoperative week when com-pared with laser-irradiated skin that had not received topicalvitamin C.
The speed of wound-healing and the duration of erythema arethe primary complaints after laser skin resurfacing. Antioxidantshave been shown to enhance the healing of wounds by reducingfree radical damage. Reepithelialization is also enhanced by themoist environment produced by occlusive dressings. We wantedto determine if alpha-keto acids could regulate and decrease per-oxynitrite concentrations at the wound site. Peroxynitrite isresponsible for increased redness at injured sites.
This residual thermal injury creates the desirable tissue tighten-ing and collagen shrinkage, with minimal postoperative bleeding,but also results in the undesired postoperative pain, prolonged
7Draft/Confidential

erythema, and the increasingly troublesome hypopigmentationthat are characteristic of current CO2, resurfacing procedures.
Laser resurfacing of facial rhytids has become a popular treat-ment option for many patients with wrinkles, photoaging, andacne scarring. Laser wavelength options and optimization of
techniques continue to evolve in an attempt to shorten the heal-ing phase associated with laser skin resurfacing.
The purpose of the study was to compare the effects of 5 alpha-keto acids for their effects on the rate of cutaneous healing, andthe ability of alpha-keto acids to reduce inflammation, specifical-
8
NeomycinONOO Keto- keto- oxaloacetate keto- pyruvate pyruvatepretreated isovaleric butyrate glutarate keto-Neomycin isovaleri
1 2 A B A B A B A B A B A B
Days to healingre-epithelization 6 2.0 4.0 2.5 4.0 4.0 3.5 3.0 4.0 5.0 4.0 2.0 3.5 2.0
Days of redness crythema 7 6 6 5 7 6 6 3 60 5 7 6 6 3
Days for disappearance of swelling 6 3 3.5 2.5 4.0 3 3.5 2.5 4.0 3.5 3.5 2.5 3.5 1.0
1= without pyruvate 2= with pyruvate A = petrolatum with the neomycin B = petrolatum with the drug and alpha keto acids
TOTALSCORES 19 11 13.5 10 15 13 13 8.5 14 13.5 14.5 10.5 13 6
% Change from A Control +42% +26% +14% +35% +5% +27% +54%
AcyclovirONOO Keto- keto- oxaloacetate keto- pyruvate pyruvatepretreated isovaleric butyrate glutarate keto-Acyclovir isovaleric
1 2 A B A B A B A B A B A B
Days to healingre-epithelization 6 2 5 4 4.0 3.0 4 3.0 5.0 5.0 5.0 2.0 5.5 2.0
Days of redness crythema 8 6 6 5.5 6 6 6 4 6 3 6 6 5 3
Days for disappearance of swelling 4 2 4 3.5 5.0 3 3.5 2.5 4.0 3.5 3.5 2.5 3.5 2.0
C) 1= without pyruvate 2= with pyruvate A = vehicle with acyclovir B = vehicle with acyclovir and alpha keto acids
TOTALSCORES 18 10 15 11 15 13 13.5 9.5 15 11.5 14.5 10.5 13.5 7
% Change from A Control +45% +26% +13% +30% +23% +28% +48%
Draft/Confidential

ly peroxynitrite at the test sites. The wound model utilized was asplit upper lip design wherein the wound was created by a welldefined model of laser skin resurfacing. This model utilizedpulsed CO2 and Er: YAG lasers in a defined protocol (computer-ized scanner for CO2 and uniform pulses with Er: YAG) which
removes the entire stratum corneum and epidermis, as well as auniform amount of dermis. Patient diaries were maintained toassess erythema, crusting, pain, itching, swelling, pigmentarychanges, and the day of first make-up application. Blinded objec-tive grading of improvement was independently assessed by 4
9
DoxorubicinONOO Keto- keto- oxaloacetate keto- pyruvate pyruvatetreated isovaleric butyrate glutarate keto-Doxorubicin isovaleric
1 2 A B A B A B A B A B A B
Days to healingre-epithelization 8 4 7.0 3.5 7 4 8 3.0 6.5 5.0 7 3.0 7 2.5
Days of redness crythema 11 6 7 5 7 7 8 5 7 7 8 5 7 3
Days for disappearance of swelling 7 4 4 2.5 4.0 3 3.5 2.5 4.0 3.0 3.5 2.5 4 2.0
D) 1= without pyruvate 2= with pyruvate A = vehicle with Doxorubicin B = vehicle with the drug and alph keto acids
TOTALSCORES 26 14 18 11 18 14 19.5 10.5 17.5 15 18.5 10.5 18 7.5
% Change from A Control +44% +39% +24% +46% +14% +43% +58%
ClotrimazolONOO Keto- keto- oxaloacetate keto- pyruvate pyruvatetreated isovaleric butyrate glutarate keto-Doxorubicin isovaleric
1 2 A B A B A B A B A B A B
Days to healingre-epithelization 7 3 5.0 3 4.0 4.0 4.5 3.0 5.5 4.0 5.0 3.0 6.0 3.0
Days of redness crythema 8 6 6 5 8 6 8 5 7 6 7 6 6 3
Days for disappearance of swelling 4 3 4 2.5 4.0 3 4 2.5 4.0 3.5 3.5 2.5 3.5 2.0
E) 1= without pyruvate 2= with pyruvate A = vehicle with Clotrimazole B = vehicle with the drug and alph keto acids
TOTALSCORES 19 12 14 10.5 16 13 16 10.5 16.5 13.5 15.5 11.5 15.5 8
% Change from A Control +37% +25% +19% +34% +18% +25% +48%
Draft/Confidential

blinded observers at time intervals 3, 6, and 10 days, and 1, 2, and4 months. Chromometer measurements of erythema were alsoanalyzed and percentage moisture recorded
A control that was utilized consisted of the same alpha-keto-wound balm without the active alpha-keto acid (that is purepetroleum, vitamin E, and egg yolk fatty acids). The productcode is enclosed. Active agent were one of: alpha-keto-isovalericacid, sodium salt; alpha-keto-butyric acid sodium salt; oxalaceticacid, sodium salt; alpha-keto-glutaric acid, sodium salt; sodiumpyruvate.
Data analysis and diary information summaries:• Moisture analysis: Alpha-keto-butyric acid, sodium salt was by
far the most effective while others had little effect on skin. Thissuggests that the stratum corneum barrier layer of the skin wasless completely repaired than the other products (a negative).
• Crusting analysis: There is a tendency for decreased durationof crusting (that is speed of wound healing) to be related to thelength of the alpha-keto acids – that is it is possible that this isproportional to some molecular characteristic. This may alsobe mirrored in skin irritancy, and thus selected alpha-keto-iso-valeric acid, sodium salt for expanded study.
• Current laser skin resurfacing techniques: Averages 5 – 7 days toskin re-epithelialization (healing) whereas a few years ago, 10 –14 days was typical. For ethical reasons, the selected the cur-rent “standard of care” model and thus the “delta value” forwound-healing between active and control should begreater/more significant for other types of wounds. Also,inevitably, some active contacts the control side in the upperlip model. Finally, the control – if it were plain petroleum –would have shown a greater difference. In short, these activi-ties in all likelihood are more effective than these results showfor typical “real world” wounds.
• Redness: All laser wounds remain red for weeks to months, doto inflammation and the over production of peroxynitrite. Thestudy design did not address the final redness issue in thediaries, but rather we utilized a Minolta Chromometer to lookat relative values of erythema. The “LAB” color space modelwas used wherein the “A” value correlates with erythema.
• Irritation: Significant irritation would have been representedby a greater increase in the “A” value with one active com-pared to others. While significant differences were not notedduring the study, pilot open patch testing data on known sen-sitive skinned individuals did show differences – some sub-jects developed red skin irritations and welt-like reactions(which I do consider significant). These actives were: alpha-keto-glutaric acid, sodium salt and slight reaction in one per-son to alpha-keto-butyric acid, sodium salt.
• Pain: The increase in pain with alpha-ketoglutarate was anotable event.All 5 alpha-keto acid wound-healing formulations were effec-
tive in accelerating wound healing in the model and all reducedredness. Other antioxidants, such as vitamin C, vitamin E do notreduce redness and appear to be ineffective against peroxynitrite.The pyruvate formulation has proven in extended clinical usageto be an effective product. This pilot screening study provides
some insight into the relative efficacy of these 5 agents. Irritationwas the only adverse event. Active agent alpha-keto-isovalericacid, sodium salt was selected for further evaluation in more sub-jects based on my interpretation of this data, prior clinical experi-ence with the pyruvate product, prior career knowledge, ourown patch testing of the relative skin irritancy of some of theseketo acids, and finally on the relative molecular size. Other ofthe activities are worthy of further clinical evaluation – in partic-ular alpha-keto-butyric acid, sodium salt and possibly oxalaceticacid, sodium salt would be my choice.
Example 3Inhibition of Irritation and Cytotoxicity of
Therapeutic AgentsAll seven of the enumerated keto acids were placed into a
commercially available triple antibiotic ointment which wasmodified by also incorporating 4% vitamin E and 6% lecithin byweight. The keto acids were also placed individually into thesame formula and were compared to a control consisting of theantibiotic ointment alone. Each formula was tested for healingrates and any associated irritation caused thereby on a patch ofdermabraided skin. The triple antibiotic ointment without theketo acids produced an irritation of the skin from the antibiotics.The formulations with the keto acids did not. They inhibited theover production of peroxynitrite which causes the over expres-sion of NF-kappa B, which reduced healing times considerably.Time in which healing took place on the skin was enhanced bythree days with the keto acid formulations. The same antibioticsand Acyclovir were placed into solutions containing peroxyni-trite then one hour later the solutions were tested for peroxyni-trite, and found not to have any. When placed on dermabraidedskin they caused irritations do to the toxic breakdown productsproduced. When the keto acids were placed singly into the samesolutions containing the peroxynitrite treated antimicrobials,one hour later, then tested on dermabriaded skin, no irritationswere noted. The keto acids inhibited the toxic effects of perox-ynitrite treated antimicrobials, which reduces inflammation bydeactivating NF Kappa. When peroxynitrite is tested on derabri-aded skin it activates inflammation. When peroxynitrite is firstplaced in solutions containing alpha-keto acids, the peroxyni-trite is neutralized and does not increase inflammation.
b) The same formula was used to treat dry winter skin, whereit worked to prevent winter itch, cracking and pain. Hydrocorti-sone was also formulated with the keto acids to also treat dryskin. This formula reduced redness and pain. A commercialcold sore formula was purchased and the seven enumeratedketo acids were placed into it to treat cold sores, both as a com-bination and singly as well. The commercial formulation byitself was utilized as a control. The normal cold sore formulaswith phenol, an antiviral agent, did not work very well. Howev-er, when keto acids are placed in the cold sore formulation eithersingly or in combination with other keto acids both with andwithout anti-oxidants and fatty acids, they heal the cold sore at amuch faster rate when tested on a cold sore sufferer. The ketoacids reduced lesion size and duration by 40% when comparedto the control formula without alpha-keto acids.
10Draft/Confidential
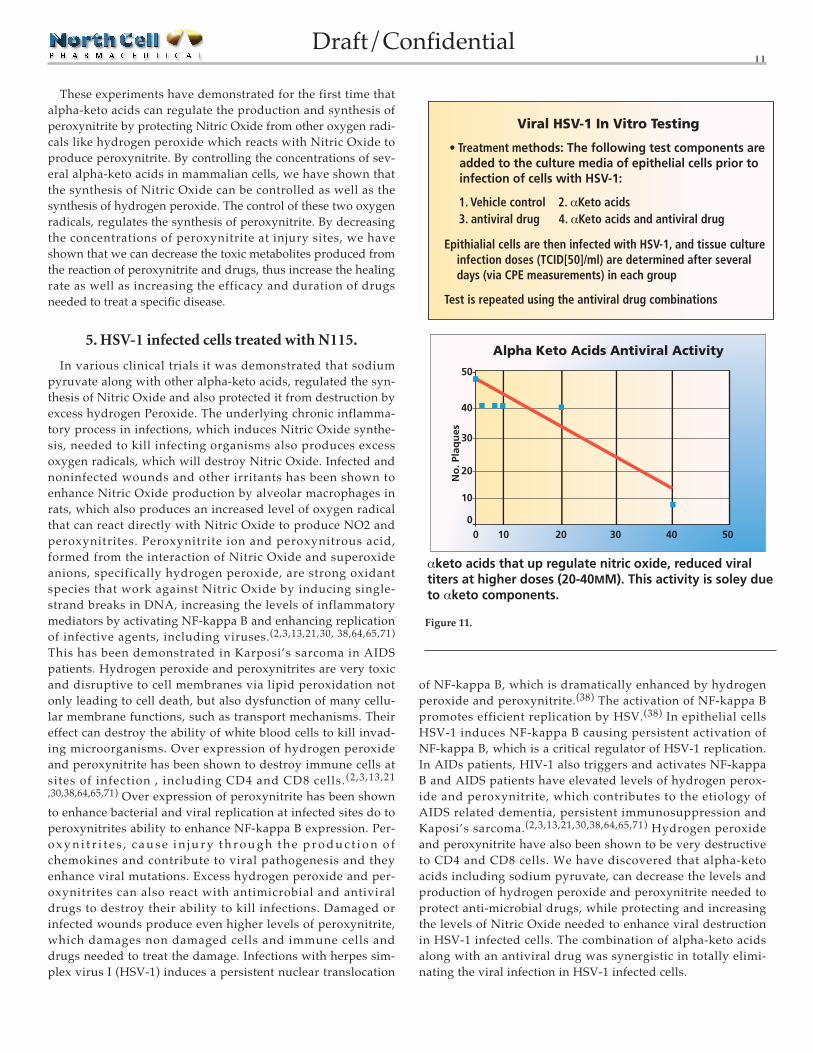
These experiments have demonstrated for the first time thatalpha-keto acids can regulate the production and synthesis ofperoxynitrite by protecting Nitric Oxide from other oxygen radi-cals like hydrogen peroxide which reacts with Nitric Oxide toproduce peroxynitrite. By controlling the concentrations of sev-eral alpha-keto acids in mammalian cells, we have shown thatthe synthesis of Nitric Oxide can be controlled as well as thesynthesis of hydrogen peroxide. The control of these two oxygenradicals, regulates the synthesis of peroxynitrite. By decreasingthe concentrations of peroxynitrite at injury sites, we haveshown that we can decrease the toxic metabolites produced fromthe reaction of peroxynitrite and drugs, thus increase the healingrate as well as increasing the efficacy and duration of drugsneeded to treat a specific disease.
5. HSV-1 infected cells treated with N115.
In various clinical trials it was demonstrated that sodiumpyruvate along with other alpha-keto acids, regulated the syn-thesis of Nitric Oxide and also protected it from destruction byexcess hydrogen Peroxide. The underlying chronic inflamma-tory process in infections, which induces Nitric Oxide synthe-sis, needed to kill infecting organisms also produces excessoxygen radicals, which will destroy Nitric Oxide. Infected andnoninfected wounds and other irritants has been shown toenhance Nitric Oxide production by alveolar macrophages inrats, which also produces an increased level of oxygen radicalthat can react directly with Nitric Oxide to produce NO2 andperoxynitrites. Peroxynitrite ion and peroxynitrous acid,formed from the interaction of Nitric Oxide and superoxideanions, specifically hydrogen peroxide, are strong oxidantspecies that work against Nitric Oxide by inducing single-strand breaks in DNA, increasing the levels of inflammatorymediators by activating NF-kappa B and enhancing replicationof infective agents, including viruses.(2,3,13,21,30, 38,64,65,71)
This has been demonstrated in Karposi’s sarcoma in AIDSpatients. Hydrogen peroxide and peroxynitrites are very toxicand disruptive to cell membranes via lipid peroxidation notonly leading to cell death, but also dysfunction of many cellu-lar membrane functions, such as transport mechanisms. Theireffect can destroy the ability of white blood cells to kill invad-ing microorganisms. Over expression of hydrogen peroxideand peroxynitrite has been shown to destroy immune cells atsites of infection , including CD4 and CD8 cells.(2,3,13,21
,30,38,64,65,71) Over expression of peroxynitrite has been shownto enhance bacterial and viral replication at infected sites do toperoxynitrites ability to enhance NF-kappa B expression. Per-o x y n i t r i t e s , c a u s e i n j u r y t h r o u g h t h e p r o d u c t i o n o fchemokines and contribute to viral pathogenesis and theyenhance viral mutations. Excess hydrogen peroxide and per-oxynitrites can also react with antimicrobial and antiviraldrugs to destroy their ability to kill infections. Damaged orinfected wounds produce even higher levels of peroxynitrite,which damages non damaged cells and immune cells anddrugs needed to treat the damage. Infections with herpes sim-plex virus I (HSV-1) induces a persistent nuclear translocation
of NF-kappa B, which is dramatically enhanced by hydrogenperoxide and peroxynitrite.(38) The activation of NF-kappa Bpromotes efficient replication by HSV.(38) In epithelial cellsHSV-1 induces NF-kappa B causing persistent activation ofNF-kappa B, which is a critical regulator of HSV-1 replication.In AIDs patients, HIV-1 also triggers and activates NF-kappaB and AIDS patients have elevated levels of hydrogen perox-ide and peroxynitrite, which contributes to the etiology ofAIDS related dementia, persistent immunosuppression andKaposi’s sarcoma.(2,3,13,21,30,38,64,65,71) Hydrogen peroxideand peroxynitrite have also been shown to be very destructiveto CD4 and CD8 cells. We have discovered that alpha-ketoacids including sodium pyruvate, can decrease the levels andproduction of hydrogen peroxide and peroxynitrite needed toprotect anti-microbial drugs, while protecting and increasingthe levels of Nitric Oxide needed to enhance viral destructionin HSV-1 infected cells. The combination of alpha-keto acidsalong with an antiviral drug was synergistic in totally elimi-nating the viral infection in HSV-1 infected cells.
11Draft/Confidential
Figure 11.
αketo acids that up regulate nitric oxide, reduced viral titers at higher doses (20-40MM). This activity is soley due to αketo components.
Alpha Keto Acids Antiviral Activity
50
40
30
20
10
00 10 20 30 40 50
No
. Pla
qu
es
Viral HSV-1 In Vitro Testing
• Treatment methods: The following test components are added to the culture media of epithelial cells prior to infection of cells with HSV-1:
1. Vehicle control 2. αKeto acids 3. antiviral drug 4. αKeto acids and antiviral drug
Epithialial cells are then infected with HSV-1, and tissue culture infection doses (TCID[50]/ml) are determined after several days (via CPE measurements) in each group
Test is repeated using the antiviral drug combinations

Summary
During an injury or infection immune system molecules, includ-ing oxygen radicals, which deplete glutathione, activate IKK-beta(37,64). IKK-beta also promotes inflammation in damaged tissues.IKK-beta is also known to be elevated in bronchial epithelial cellsin cystic fibrosis(37,64) and in other lung diseases, including cancer.IKK-beta works by activating the transcription factor nuclear fac-tor-kappa B (NF-kappa B).(39,40,64) NF-kappa B regulates theinflammatory responses, including genes that encode pro-inflam-matory cytokines IL-2, TNF, IL-6, IL-11, IL-17. NF-kappa B alsoregulates NO synthetase, and genes that inhibit apoptosis whichplays a major role in tumor growth and survival.(44,49,50)
Thus any molecule that deactivates either IKK- beta or NF-kappa B would inhibit inflammation.(41) In patients with lungdiseases including asthma, cystic fibrosis, COPD, ILD, and can-cer, NF-kappa B activation is exaggerated as are other inflamma-tory components.(37,38,44) Hydrogen peroxide and other oxygenradicals, such as nitrogen dioxide and peroxynitrite that can alsobe generated form cancer drugs like bleomycin, activate NF-kappa B and deplete cellular glutathione levels. (42-45,64) Sodiumpyruvate, an oxygen radical antagonist, was shown to decreasethe expression of several proinflammatory genes, including NF-kappa B, activation of inducible NO synthase mRNA, TNF,cyclo-oxygenase, interleukin 6 and 10mRNA induction in numer-ous cell lines and in various animal models.(32,33,44,49,50,64) Sodi-um pyruvate inhibited hydrogen peroxide induced transcriptionof NF-kappa B while protecting cellular glutathione.
Sodium pyruvate is a reactive oxygen species antagonist that hasbeen shown to neutralize oxygen radicals, specifically lower theoverproduction of superoxide anions, regulate the production andlevel of other inflammatory mediators, and control the synthesis ofNitric Oxide. Sodium pyruvate is an antioxidant that reacts directlywith oxygen radicals like hydrogen peroxide and peroxynitrite, to
neutralize them protecting DNA and other cellular componentssuch as glutathione.(56-65,70,71) In macrophages, and other cell lines,sodium pyruvate regulates the production and level of inflammato-ry mediators including oxygen radical production and also regu-lates the synthesis of Nitric Oxide.(8,49,50) Low concentrations ofpyruvate decreases the expression of several proinflammatorygenes, including NF-kappa B, activation of inducible NO synthasemRNA in various cel l l ines and in various animal mod-els.(32,33,44,49,50,64). Sodium pyruvate inhibited hydrogen peroxideinduced transcription of NF-kappa B while protecting cellular glu-tathione (44,64) Further sodium pyruvate blocked the p38 MAPKpathway and activated the ERK pathway, which regulates theexpression of genes believed to prevent apoptosis and promote cellsurvival.(44,64) Sodium pyruvate at lower concentrations inhibitedhepatocytes Nitric Oxide synthesis(27), and at higher concentrationscaused up regulation of inducible Nitric Oxide synthase mRNA inintestinal cells and in cardiac monocytes.(8,26,28) It can specificallylower the overproduction of superoxide anions, H2O2 and NO inwhite blood cells.(31,56-64) Sodium pyruvate also increases cellularlevels of glutathione, a major cellular antioxidant (8) which isreduced dramatically in antigen-induced lung disease patients.
Oxygen radicals are involved in cellular damage and excess oxy-gen radicals are involved in the induction and progression ofmalignancy. Pyruvate a known scavenger of oxygen radicals hasbeen shown to enhance cellular healing and implicated in cancerprevention.(32,33,57,64) Pyruvate inhibited the growth of implantedtumors and reduced lung metastases; also it decreased the numberof DNA breaks caused by H2O2 by 40%.(32) Pyruvate controls theproduction and the positive and negative effects of Nitric Oxide athigher levels. Sodium pyruvate also increases cellular levels of glu-tathione, a major cellular antioxidant, which is reduced dramatical-ly in antigen-induced lung disease patients. We have discoveredthat alpha-keto acids including sodium pyruvate, can decreasethe levels and production of hydrogen peroxide and while pro-tecting and increasing the levels of Nitric Oxide needed toenhance viral destruction in HSV-1 infected cells.
REFERENCES 1. Asthma , third edition, edited by Peter Barnes Chapter 21. 2. Moncada, S. et al, Nitric Oxide: physiology, pathophysiology, andpharmacology. 1991 Pharmacological Reviews Vol. 43 no pp 109-141.3. Nathan, C., Nitric Oxide as a secretory product of mammalian cells.FASEB journal vol. Sep. 6, 1992 pp 3051-3064. 4. Rossaint, R. et al, Inhaled Nitric Oxide: its effect on pulmonary circula-tion and airway smooth muscle cells. Euro Heart Jour. 1993 vol. 14 Supp.pp 133-140.5. Artlich, A. et al., Childhood asthma: exhaled Nitric Oxide in relation toclinical symptoms. Euro Respir. J. Vol. 13, pp 1395-1401.6. Jobsis, Q. et al. Hydrogen peroxide and Nitric Oxide in exhaled air ofchildren with cystic fibrosis during antibiotic treatment . Euro Respir.J.2000 Vol. 16, pp 95-100.7. Mukala, K. et al. Personally measured weekly exposure to NO.sub.2and respiratory health among preschool children. Euro. Respir. J. Vol. 13,pp 1411-1417.8. Stanko R., The power of Pyruvate 1999, Keats Publishing.9. Kelly, F. et al. Antioxidant kinetics in lung ravage fluid following expo-sure of humans to nitrogen dioxide. Am. J. Respir. Crit Med. Vol. 1541991 pp 1700-1705.10. Comhair S. at al. Rapid loss of superoxide dismutase activity during
12Draft/Confidential
% o
f vir
al p
laq
ues
αketo acids/antiviral drug combination more effective vs. either alone
Results
60
40
20
80
100
0
1 2
3
1
2
3
1
2
3
0.5X AD/5MM αKA 1X AD/10MM αKA 2X AD/20MM αKA
1 αketo acids (αKA) 2 antiviral drug (AD)
3 αketo acids/antiviral drug combinationFigure 12.

antigen induced asthmatic response. Lancet vol. 355 feb. 19 2000.11. Sporn P H, et al. Hydrogen peroxide induced arachidonic acidmetabolism in rat alveolar macrophage. Am Rev Respir Dis 1988 137: 49-56. 12. Alving, K. Methodological aspects of exhaled Nitric Oxide measure-ments Euro Respir Rev 1999: 9:68, 208-211.13. Kharitonov, S. Exhaled Nitric Oxide and carbon monoxide in asthma.Euro Respir. Rev. 1999, 9:68, 212-216.14. Gouw, P. et al. Stimuli affecting exhaled Nitric Oxide in asthma. EuroRespir. Rev. 1999; 9:68, 219-222.15. Kharitonov, S. Exhaled Nitric Oxide and carbon monoxide in respira-tory diseases. Euro Respir. Rev. 1999; 9:68, 223-226.16. Barnes, P. The effect of drugs on exhaled Nitric Oxide. Euro Respir.Rev. 1999; 9:68, 231-233.17. Baraldi, E. et al. Application of exhaled Nitric Oxide measurement inpediatrics. Euro Respir. Rev. 1999; 9:68, 234-240.18. Lundberg, J. Nitric Oxide in the nasal airways. Euro Respir. Rev. 1999;9:68, 241-245.19. Culpitt, S. The measurement of hydrogen peroxide in airways disease.Euro Respir. Rev. 1999; 9:68, 246-248.20. Montuschi, P. Isoprostanes and other exhaled markers in respiratorydiseases. . Euro Respir. Rev. 1999; 9:68, 249-253.21. Robertson, F M, Gene expression and cellular sources of inducibleNitric Oxide synthase during tumor promotion. Carcinogenesis Septem-ber; 1996 17 (9): 2053-9.22. Soler M N, et al, Gene therapy of rat medullary thyroid cancer bynaked Nitric Oxide synthase II DNA injection. J Gene Med September-October; 2000 2(5): 433-52.23. Wang H H, B 16 melanoma cell arrests in mouse liver induces NitricOxide release and sinusoidal cytotoxicity: a natural hepatic defenseagainst metastasis. Cancer Res Oct. 15, 2000 60(20): 5862-924. Brennan P A., The action and interactions of Nitric Oxide in solidtumors. Eur J Surg Oncol Aug. 26, 2000 (5): 434-7.25. Gossart S. et al. Reactive oxygen intermediates as regulators of TNF-alpha production in rat lung inflammation induced by silica . J ofImmunology 1996. 156 pp1540-1548.26. Oddis CV, et al. Glucose and pyruvate regulate cytokine inducedNitric Oxide production by cardiac monocytes. Am J Physiol. 1996Oct;271 (4 Pt 1):c1244-9.27. Liang JF. Et al. Role of metabolic intermediates in lipopolysaccharidecytokine mediated production of Nitric Oxide in isolated hepatocytes.Biochem Biophys Res Commun. 1997 Jul 18;236(2):379-82.28. Carvalho et al. Poster A35. Effects of Nitric Oxide on pulmonaryhyperreactivity and airway inflammation in allergic rat models. 97thinternational conference 1999. American Thoracic Society.29. Taskar V.S. et al Poster 515. Strategy for selective inhibition of humaninducible Nitric Oxide synthase by targeting a unique epitope. 97th inter-national conference 1999. American Thoracic Society.30. Okamoto, S. et al. Poster D42. Enhanced viral mutation by NitricOxide induced oxidative stress in sendai virus pneumonia in mice. 97thinternational conference. 1999. American Thoracic Society.31. Ferreira, I.M. et al. Poster 812. Effects of inhaled beclomethasone on exhaledNitric Oxide as a marker of lower respiratory inflammation in patients withCOPD. 97th international conference 1999. American Thoracic Society. 32. Stanko,R et al. Pyruvate inhibits Growth of Mammary adenocarcino-ma 13762 in rats. Research 54, 1004-1007, feb 15, 1994.33. Stanko R. et al Pyruvate inhibits clofibrate induced hepatic peroxiso-mal proliferation and free radical production in rats. Metabolism, vol 44,No2 (feb), 1995: pp 166-171.34. Rensberger R, Gardian angle protein molecule inside the cell identi-fied, Science, cell biology 1994.35. Herz H, et al. Multicomponent investigation of the hydrogen peroxideand hydroxyl radical scavenging antioxidant capacities of biofluids: the rolesof endogenous pyruvate and lactate. Free radical res. Vol.26, pp 19-35 1997. 36. Varkey, B, et al. Asbestosis. Medline Aug 18, 2004.37. Escotte,S. et al. Fluticasone reduces IL-6 and IL-8 production of cysticfibrosis bronchial epithelial cells via IKK-beta kinase pathway. EuroRespir J. 2003 Apr; 21(4): 574-81.38. Mogensen Th. et al. Activation of NF-kappa B in virus-infected
macrophages is dependent on mitochondrial oxidative stress and inter-cellular calcium: downstream involvement of kinases TGF-beta-activatedkinase 1, mitogen-activated kinase/extracellular signal-regulated kinase1, and I kappa B kinase. J immunol 2003 jun 15;170(12) : 6224-33.39. Li, ZW. Et al. IKK beta is required for peripheral B cell survival andproliferation. J Immunol. 2003 May 1; 170(9):4630-7.40. Macotela Y. et al. 16K prolactin induces NF-kappa B activation in pul-monary fibroblasts. J Endocrinol. 2002 Dec;175(3):R13-8.41. Conron M, et al. Nuclear factor kappa B activation in alveolarmacrophages requires I kappa B kinase-beta , but not nuclear factor kappaB inducing kinase. Am J Respir Crit Care Med. 2002 Apr 1 165(7):996-1004.42. Schoonbroodt S. et al. Critical role of the amino-terminal tyrosine residue42 and the carboxyl-terminal PEST domain of I kappa B alpha in NF-kappa Bactivation by an oxidative stress. J Immunol 2000 Apr. 15;164(8):4292-300.43. Torrie LJ. Hydrogen peroxide-mediated inhibition of lipopolysaccha-ride-stimulated inhibitory kappa B kinase activity in rat aortic smoothmuscle cells. Br. J Pharmacol 2001 sept;134(2):393-401.44. Lee YL. Et al. Enhanced survival effect of pyruvate correlates MAPKand NF-kappa B activation in hydrogen peroxide treated humanendothelial cells. J Appl Physiol. 2004 feb;96(2): 793-801.45. Deng L, et al. 2-acetylaminofluorene up regulates rat mdrib expres-sion through generating reactive oxygen species that activate NF-kappa Bpathway.J Biol Chem. 2001 Jan 5;276(1):413-20.46. Asehnoune K. et al. Involvement of reactive oxygen species in Toll-like receptor 4-dependent activation on NF-kappa B. J Immunol. 2004 Feb15;172(4): 2522-9.47. Lee SJ. Et al. Astaxanthin inhibits Nitric Oxide production and inflam-matory gene expression by suppressing I kappa B kinase-dependent NF-kappa B activation. Mol Cells. 2003 Aug 31; 16(1): 97-105.48. Carcamo JM. Et al. Vitamin C ia a kinase inhibitor: dehydroascorbicacid inhibits I kappa B alpha kinase beta. Mol Cell Biol. 2004 Aug;24(15):6645-52.49. Sappington PL. et al. Ethyl pyruvate ameliorates intestinal epithelialbarrier dysfunction in endotoxemic mice and immunostimulated caco-2enterocytic monolayers. J Pharmacol. Exp Ther. 2003 Jan;304(1):464-76.50. Yang R. et al. Ethyl pyruvate modulates inflammatory gene expres-sion in mice subjected to hemorrhagic shock. Am J Physiol Gastrointestliver Physiol. 2002 July;283 (1):G212-21.51. Spiecker M. et al. Differential regulation of endothelial cell adhesionmolecule expression by Nitric Oxide donors and antioxidants. J LeukocBiol. 1998 June;(6):732-9.52. Kang MK. et al. Protective effect of retinoic acid on interleukin-1 beta-induced cytotoxicity of pancreatic beta-cells. Mech Aging Dev. 2004Jul;125(7):480-90.53. Reynaert NL. et al. Nitric Oxide represses inhibitory kappa B kinasethrough S-nitrosylation. Proc Natl Acad Sci USA. 2004 June 15;101(24):8945-50.54. Ogino S. et al. Herbimycin A abrogates nuclear factor-kappaB activa-tion by interacting preferentially with the IkappaB kinase beta subunitMol Pharmacol. 2004June;65(6):1344-51.55. Umansky V. et al. Co-stimulatory effect of Nitric Oxide on endothelialNF-kappa B implies a physiological self-amplifying mechanism Eur JImmunol. 1998 Aug;28(8):2276-82.56. Giandomenico A. et al. The importance of sodium pyruvate in assess-ing damage produced by hydrogen peroxide. Free Radical Biol andMed.1997 Vol 23. no 3. pp426-434.57. Biri H. et al. Antioxidant potential of cancerous human kidney tissues.Cancer Biochem. Biophy. 1998 vol 16 pp. 265-272.58. Wang X. et al. Pyruvate released by astrocytes protects neurons fromcopper catalyzed cysteine neurotoxicity. The J.of Neuroscience may 15,2001. 21(10): 3322-3331.59. Venkataraman R. et al. Resuscitation with ethyl pyruvate solution prolongssurvival and modulates plasma cytokine and nitrite-nitrate concentrations in arat model of lipopolysaccharide-induced shock Shock. 2002 Dec;18(6):507-12.60. Nath K. et al. Effect of pyruvate on oxidant injury to isolated and cel-lular DNA. Kidney international. Vol 45 1994 pp 166-176.61. O’Donnell-Tormey J. et al. Secretion of pyruvate. J Exp. Med. TheRockefeller univ. press. Vol.165 feb 1987 pp 500-514.62. Andre B et al. Pyruvate reduces anoxic injury and free radical forma-
13Draft/Confidential

tion in perfused rat hepatocytes. The American Physiological society.1996 G535-G540.63. Bassenge E. et al. Antioxidant pyruvate inhibits cardiac formation ofreactive oxygen species through changes in redox state. Am J. Physiolheart Circ Physiol 2000 279: H2431-H2438.64. Fink M. Reactive oxygen species as mediators of organ dysfunctioncaused by sepsis, acute respiratory distress syndrome, or hemorrhagicshock: potential benefits of resuscitation with ringer’s ethyl pyruvatesolution. Curr Opin Nutr Metab Care vol 5(2) march 2002. pp167-174.65. McBride A. et al. Superoxide dismutase and hydrogen peroxide causerapid Nitric Oxide breakdown, peroxynitrite production and subsequentcell death. Biochemica et biophysica acta 1454 1999 pp 275-288.66. Dekhuijken R. et al. Increased exhalation of hydrogen peroxide inpatients with stable and unstable COPD Am J Respir Crit Care Med vol154. pp 813-16 1996..67. Kietzmann D. et al. Hydrogen peroxide in expired breath condensateof patients with acute respiratory failure and with ARDS. Intensive CareMed. 1993 19: pp 78-81.68. Hudson V. Rethinking cystic fibrosis pathology; the critical role ofabnormal reduced glutathione transport caused by CFRT mutation. FreeRad Biol and Med. 2001. Vol 30, no12 pp 1440-1461.69. Rahman I. Et al. Oxidative stress and regulation of glutathione in lunginflammation. Eur Respir J. 2000. 16: pp 534-554.70. vivar J V. et al. Peroxynitrite-mediated decarboxylation of pyruvate toboth carbon dioxide and carbon dioxide radical anion. Chem. Res. Toxi-col. 1997. 10, pp 786- 794.71. Schultz R. et al. Peroxynitrite impairs cardiac contractile function bydecreasing cardiac efficiency. The American Physiological Society. 1997pp H1212-H1219.72. Dhar A.et al. Nitric Oxide does not mediate by inhibits transformationand tumor phenotype. Mol Caner Ther. 2003 Dec;2(12): pp 1285-1293.73. Thomassen M. et al. Nitric Oxide regulation of asthmatic airwayinflammation and segmental allergen challenge. J Allergy Clin Innunolvol104, number 6 dec1999 pp 1174-1182.74. Puhakka A et al. Modulation of DNA damage by inhibition of NitricOxide synthase and gammaglutamycysteine synthase in lung cell exposed toasbestos fibers. 2000 meeting of the Amer Thoracic Society. D42 poster B6.75. Pan X. et al. Environmental Asbestos and Mesothelioma in California.C55 Poster E 77. 2001 Meeting of the Amer Thoracic Society.76. Abayasiriwardana, K.S. et al. The proline analog thiaproline inhibitsmalignant mesothelioma cell collagen production in vitro and tumorgrowth in mice. Poster A15. 2001 Meeting of the Amer Thoracic Society.77. Zhang J . et al. Asbestos inhalation induces DNA strand breaks andincreased p53, p21 and PARP expression in the rat lung. A26, Poster 813.2000 Meeting of the Amer Thoracic Society.78. Kamp D.W. et al. Asbestos augments A549 cell expression of trans-forming growth factor-beta 1: role of iron-catalyzed free radicals. A26poster 815. 2000 Meeting of the Amer Thoracic Society.79. Carter A.B. Crocidolite does not drive AP-1 dependent genes sue tothe lack of p38 MAP kinase activation. C98 poster 719. 2001 Meeting ofthe Amer Thoracic Society.80. Espiritu J.D. et al. COPD and TNF-alpha receptor gene polymorphismin asbestos exposed workers. A49 poster J61. 2001 Meeting of the Amer
Thoracic Society.81. Jiang Y. et al. Lung specific expression of a dominant negative mutantof p53 in transgenic mice increases spontaneous and benzo(A) pyreneinduced lung cancer . C48 poster A51. 2000 Meeting of the Amer ThoracicSociety.82. Natarajan R. et al. Pyrrolidine Dithiocarbamate has a biphasic effecton NF-kappa B induced IL-8 secretion in human lung fibroblasts. C57poster J28. 2001 Meeting of the Amer Thoracic Society. 83. Undurti. N. Pyruvate is an endogenous anti-inflammatory and antiox-idant molecule. Med Sci Monit, 2006; 12(5): RA79-84 pp 79-84.83. Dijkstra U, F Gabreels, E. Joosetn et al. Friedrich’s ataxia: Intravenouspyruvate load to demonstrate a defect in pyruvate metabolism. Neurolo-gy 1984; 34:1493-1497. 84. Giannelli S, JP McKenna, JM Bordiuk et al. Prevention of increasedhemoglobin-oxygen affinity in open-heart operations with inosine-phos-phate-pyruvate solution. Ann thoracic Surg 1976; 21:386-396. 85. Tsukiyama T, T Hara, M Lio et al. Preferential accumulation of 11C inhuman brain tumors after intravenous injection of 11C-1-pyruvate. Eur JNucl Med; 1986: 12(5-6):244-248. 86. Companacci L, GF Guarnieri, L Faccini et al. Pyruvate tolerance test inchronic uremic patients. Nephron 1983; 10:232-237. 87. Levy SB and LA Goldsmith. Sodium pyruvate treatment for hyperker-atotic disorders. South Med J 1979; 72(5):307-310. 88. Stanko RT, RJ Robertson, R-W Galbreath et al. Enhanced leg exerciseendurance with a high-carbohydrate diet and dihydroxyacetone andpyruvate. J Appl Physiol 1990; 69:1651-1656. 89. Stanko RT, HR Reynolds, KD Lonchar et al. Plasma lipid concentra-tions in hyperlipidemic patients consuming a high-fat diet supplementedwith pyruvate for 6 weeks. Am J Clin Nutr 1992: 56:950-954.90. Stanko RT, HR Reynolds, R Hoyson et al. Pyruvate supplementationof a low-cholesterol, low-fat diet: effects on lipid concentrations andbody composition in hyperlipidemic patients. Am J Clin Nutr 1994;59:423-427. 91. Stanko RT, HR Reynolds, R Hoyson et al. Pyruvate supplementationof a low-cholesterol, low-fat diet: effects on lipid concentrations andbody composition in hyperlipidemic patients. Am J Clin Nutr 1994; 59:423-427. 92. Horvath I, LE Donnelly, A Kiss et al. Combined Use of ExhaledHydrogen Peroxide and Nitric Oxide in Monitoring Asthma. Am J RespirCrit Care Med 1998;158:1042-1046.93. Dohlman AW, HR Black, JA Royall. Expired Breath Hydrogen Perox-ide Is a Marker of Acute Airway Inflammation in Pediatric Patients withAsthma. Am Rev Respir Dis 1993;148:955-60.94. Loukides S, I Horvath, T Wodehouse et al. Elevated Levels of ExpiredBreath Hydrogen Condensate in Bronchiectasis. Am J Respir Crit CareMed. 1998, 158:991-994. 95. Kietzmann D, R Kahl, M Muller et al. Hydrogen peroxide in expiredbreath condensate of patients with acute respiratory failure and withARDS. Intensive Care Med. 1993, 19:78-81. 96. Sznajder JI, A Fraiman, JB Hall et al. Increased Hydrogen Peroxide inthe Expired Breath of Patients with Acute Hypoxemic Respiratory Fail-ure. Chest 1989; 96:606-12.
14Draft/Confidential

15Notes
