effect of peg-induced drought stress on seed germination of four lentil genotypes
TRANSCRIPT

This article was downloaded by: [80.47.125.66]On: 08 July 2014, At: 22:57Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Plant InteractionsPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tjpi20
Effect of PEG-induced drought stress on seedgermination of four lentil genotypesAdele Muscoloa, Maria Sidaria, Umberto Anastasib, Carmelo Santonocetoa & AlbinoMaggioc
a Dipartimento di Agraria, Università “Mediterranea” di Reggio Calabria, Località Feo diVito, 89126 Reggio Calabria, Italyb Dipartimento di Scienze delle Produzioni Agrarie e Alimentari, Università di Catania,Via Valdisavoia 5, 95123 Catania, Italyc Dipartimento di Ingegneria Agraria e Agronomia del Territorio, Università di Napoli“Federico II”, Via Università 100, 80055 Portici, Napoli, ItalyAccepted author version posted online: 20 Aug 2013.Published online: 16 Sep 2013.
To cite this article: Adele Muscolo, Maria Sidari, Umberto Anastasi, Carmelo Santonoceto & Albino Maggio (2014) Effect ofPEG-induced drought stress on seed germination of four lentil genotypes, Journal of Plant Interactions, 9:1, 354-363, DOI:10.1080/17429145.2013.835880
To link to this article: http://dx.doi.org/10.1080/17429145.2013.835880
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. Taylor & Francis, our agents, and our licensors make no representationsor warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content.Versions of published Taylor & Francis and Routledge Open articles and Taylor & Francis and RoutledgeOpen Select articles posted to institutional or subject repositories or any other third-party website arewithout warranty from Taylor & Francis of any kind, either expressed or implied, including, but not limited to,warranties of merchantability, fitness for a particular purpose, or non-infringement. Any opinions and viewsexpressed in this article are the opinions and views of the authors, and are not the views of or endorsed byTaylor & Francis. The accuracy of the Content should not be relied upon and should be independently verifiedwith primary sources of information. Taylor & Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever causedarising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Terms & Conditions of accessand use can be found at http://www.tandfonline.com/page/terms-and-conditions It is essential that you check the license status of any given Open and Open Select article toconfirm conditions of access and use.

RESEARCH ARTICLE
Effect of PEG-induced drought stress on seed germination of four lentil genotypes
Adele Muscoloa*, Maria Sidaria, Umberto Anastasib, Carmelo Santonocetoa and Albino Maggioc
aDipartimento di Agraria, Universita ‘‘Mediterranea’’ di Reggio Calabria, Localita Feo di Vito, 89126 Reggio Calabria, Italy;bDipartimento di Scienze delle Produzioni Agrarie e Alimentari, Universita di Catania, Via Valdisavoia 5, 95123 Catania, Italy;
cDipartimento di Ingegneria Agraria e Agronomia del Territorio, Universita di Napoli ‘‘Federico II’’, Via Universita100, 80055 Portici, Napoli, Italy
(Received 24 July 2013; accepted 13 August 2013)
Seeds of four lentil genotypes (Castelluccio, Eston, Pantelleria, and Ustica) were subjected to five levels (0, 10, 15,
18, and 21%) of polyethylene glycol (PEG-6000). Germination percentage, root length, tissue water content(WC), a- and b-amylases, a-glucosidase activities, and osmolyte content were evaluated at 24, 48, and 72 h afterstarting the germination test. Water stress reduced seed germination percentage, root length, and seedling WC in
all cultivars to different extent. The increase in proline content and total soluble sugars was greater for Eston andCastelluccio compared to the other genotypes. The activity of the enzymes involved in the germination processdecreased in all cultivars; the activities of a-amylase and a-glucosidase were most negatively affected by osmoticstress, mainly in the drought sensitive Ustica and Pantelleria. Overall, Eston and Castelluccio were able to express
greater drought tolerance and consequently could be used as a valuable resource for breeding programs.
Keywords: amylase activity; drought; lentil; osmotic stress; proline; seed germination
1. Introduction
Worldwide agricultural productivity is subject toincreasing environmental constraints in the form ofabiotic stresses that adversely influence plants growthand development causing crop failure and decreasingaverage yields more than 50% (Buchanan et al. 2000;Bartels & Sunkar 2005; Mittler 2006; Wu et al. 2011).In semiarid environments where lentil is widespread,unfavorable soil moisture at sowing often conditionsseverely seed germination resulting in an irregularseedling emergence, which in turn affects the estab-lishment of a stand, with negative effects on the yield(Mwale et al. 2003; Okcu et al. 2005). For thesereasons, drought tolerance at the germination stagehas specific importance moreover in warm environ-ments most vulnerable to climate change (IPCC2007).
Lentil (Lens culinaris L.), one of the oldestdomesticated plants in the world, originated fromthe near East and central Asia, is traditionallycultivated in the Mediterranean basin (Zohary1972). Seeds of this species are an important sourceof protein for the human diet and the entire biomassof plant is a valued animal feed. Irrigation generallyincreases lentil yield (Salehi et al. 2008), improvingseed size, seed yield, biomass yield, and harvest index(Singh & Saxena 1990; Silim et al. 1993; Khourgamiet al. 2012). Thus, the successful crop establishment insemiarid areas depends on the rapid and uniform seedgermination, which is strictly associated to the abilityof seeds to germinate under low water availability
(Arjenaki et al. 2011). The sequence of events leading
to seed germination and root emergence is governed by
water uptake from the external medium (Kaur et al.
1998; Hodge et al. 2009). Water availability plays a
significant role in enzymatic reactions, solubilisation
and transportation of metabolites, and also as a
reagent in the hydrolytic breakdown of proteins,
lipids, and carbohydrates in the storage tissues of
germinating seeds (Bewley & Black 1994; Biaecka &
Kepczynski 2010). Amylase enzymes play an impor-
tant role during seed germination, hydrolyzing the
endosperm starch into metabolizable sugars, which
provide the energy for the growth of roots and shoots
(Nauriere et al. 1992). The activity of such enzymes is
reduced by water stress with negative effects on
carbohydrate metabolism (Kaur et al. 2000; Zeid &
Shedeed 2006).Selection of plants with a better drought tolerance
is critical in dry environments (Ashraf et al. 1992;
Tuberosa & Salvi 2006). However, controlled and
uniformly repeated simulation of drought in the field
cannot be easily achieved (Shaheen & Hood-Nowotny
2005). The slow progress in developing drought-
resistant cultivars also reflects the lack of a specific
method for screening the large numbers of genotypes
required in breeding for drought (Zeigler & Puckridge
1995). Using natural field conditions is difficult be-
cause rainfall can eliminate water deficits. However, in
vitro drought-screening methods are facilitating pro-
gress in our understanding of drought-resistance traits
and in our selection of drought-resistant genotypes.
*Corresponding author. Email: [email protected]
Journal of Plant Interactions, 2014Vol. 9, No. 1, 354�363, http://dx.doi.org/10.1080/17429145.2013.835880
# 2013 Taylor & Francis
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014

Richards (1978) suggested germination as a useful
criterion in screening for water stress tolerance. Khak-wani et al. (2011) demonstrated that among the sixvarieties of wheat tested, those who were tolerant todrought during in vitro germination tests were simi-
larly tolerant in field conditions. In addition,Agili et al.(2012) confirmed this finding with experiments onsweet potato. Thus, study of the influence of thedrought using osmotic solutions is one of the methods
in the evaluation of resistance during the germinationphase. Exposure to polyethylene glycol (PEG-6000)solutions has been effectively used to mimic drought
stress with limited metabolic interferences as thoseassociated to the use of low molecular weight osmo-lytes that can be taken up by the plant (Hohl &Schopfer 1991). PEG-based in vitro screening for
drought tolerance has been proven to be a suitablemethod to effectively screen large sets of germplasmwith good accuracy (Kulkarni & Deshpande 2007).
Understanding the biochemical mechanisms in-volved in plant drought stress tolerance is still a major
challenge in biology and agriculture to identify atearly stage suitable traits that would support plantbreeders in specific selection programs. The mainobjective of this study was to evaluate the influence of
drought stress on seeds of the four cultivars of lentil,which had previously shown to have diverse level oftolerance to NaCl stress (Sidari et al. 2008), in order
to select the best suitable parents for hybridization inbreeding patterns.
2. Materials and methods
2.1. Plant material, germination conditions, andexperimental design
The following lentil cultivars were studied in thisexperiment. Two salt stress tolerant landraces ‘Pan-
telleria’ and ‘Ustica’: native and cultivated in thehomonymous small islands close to Sicily (SouthernItaly), a local population ‘Castelluccio di Norcia’:
cultivated in Umbria region (Central Italy), and aCanadian commercial variety ‘Eston.’ Seven-month-old seeds (stored at 20918C and 95% R.U.) of eachlentil genotype were used. The seeds were selected for
size homogeneity, surface-sterilized for 20 min in 30%(v/v) H2O2, rinsed and soaked in distilled water for 1h. For each of four genotypes, five replicates of 50-seed were placed on a filter paper in 9-cm Petri dishes
containing 3 cm3 of distilled water or 10, 15, 18, and21% of PEG (MW 6000) concentration correspond-ing to final osmotic potentials of �0.30, �0.51,�0.58, and �0.80 MPa, respectively. We used 10,
15, 18, and 21% of PEG to have an osmotic potentialcomparable to that of NaCl at the concentrations of50, 100, 150, and 200 mM that we tested on seed
germination of the same cultivars in a previous work(Sidari et al. 2008), in order to evaluate similarity ordifferences in the metabolic traits of salt and droughtresistance. The Petri dishes were sealed with Parafilm
to prevent evaporation and kept accordingly to acompletely randomized design in a growth chamber ata temperature of 25918C in the dark with a relativehumidity of 70%. Seeds were considered germinatedwhen the radicle had extended for at least 2 mm. Thewater content (WC) was measured and expressed as apercentage according to the formula WC (%)�(Fresh Weight � Dry Weight/Fresh Weight)�100.Root length (cm) was also measured and for each offour genotypes, five replicates were used.
2.2. Enzyme activity
The activities of a-amylase, b-amylase, and a-gluco-sidase were determined in the crude extracts of eachcultivar. The seeds of each cultivar and for each PEGtreatment (0, 10, 15, 18, and 21%) were homogenizedin a chilled mortar with distilled water 1:4 (w/v) andcentrifuged at 14,000 g for 30 min. The supernatantswere filtered through a single layer of muslin clothand were used for a-amylase (EC 3.2.1.1) (Steup1988), b-amylase (EC 3.2.1.2) (Steup 1988), anda-glucosidase (EC 3.2.1.20) (Bergmeyer et al. 1983)activity determination.
For a-amylase, a mixture of 3 ml soluble starch(2% v/v) and 3 ml extract was incubated for 60 min at308C. After incubation, an equal volume of alkalinecolor reagent was added to 1-ml incubation mixture,mixed and heated for five min in a boiling water bath.The absorbance at 546 nm was measured against ablank (1 ml H2O plus 1 ml alkaline reagent). Thestandard curve was obtained by using different con-centrations of maltose in the range of 0�1.5 mmol l�1.The alkaline color reagent was prepared by dissolving1 g of 3,5-dinitrosalycylic acid in a mixture of 40 ml1 N NaOH solution and 30 ml H2O. Solid potassiumsodium tartrate was added and dissolved. The mixturewas brought to a final volume of 100 ml (Steup 1988).b-amylase was determined as described above butsoluble starch was replaced by amylopectin. Fora-glucosidase detection, the assay buffer consisted of50 mM Na-acetate, pH 5.2, containing 10 mM CaCl2.The substrate was 10 mmol l�1 maltose. The sampleswere incubated for 60 min. The release of glucose wasfollowed by measuring the changes in NADPH at340 nm in a coupled enzyme reaction of hexokinaseand glucose-6-P dehydrogenase. For each treatment,five replicates were used.
2.3. Osmolyte content
To detect free proline content samples (0.3 g) includ-ing 5 ml of 3% sulfosalicylic acid were homogenizedand centrifuged at 3000 rpm for 20 min. The super-natant was added to 2 ml of glacial acetic acid with2 ml acidic ninhydrin. The mixture was heated at1008C for 25 min. After the liquid was cooled, themixture was added to 4 ml toluene. The absorbance ofthe extracts was read at 520 nm (Bates et al. 1973). Thetotal soluble sugars were determined with the
Journal of Plant Interactions 355
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014
anthrone method (Yemn & Willis 1954). For eachtreatment, five replicates were used.
2.4. Measurement times and statistical analysis
Root length, water content (WC), enzyme activity,and osmolyte content were measured at 24, 48, and72 h after the start of the test. The data werestatistically analyzed separately for each time by atwo-way ANOVA according to the adopted experi-mental design combining PEG concentrations andgenotypes. The germination percentage data werepreviously subjected to arcsine transformation andwere reported in tables as untransformed values. Thedifferences between the means were compared by theleast significant difference (LSD) test (p50.05).
3. Results
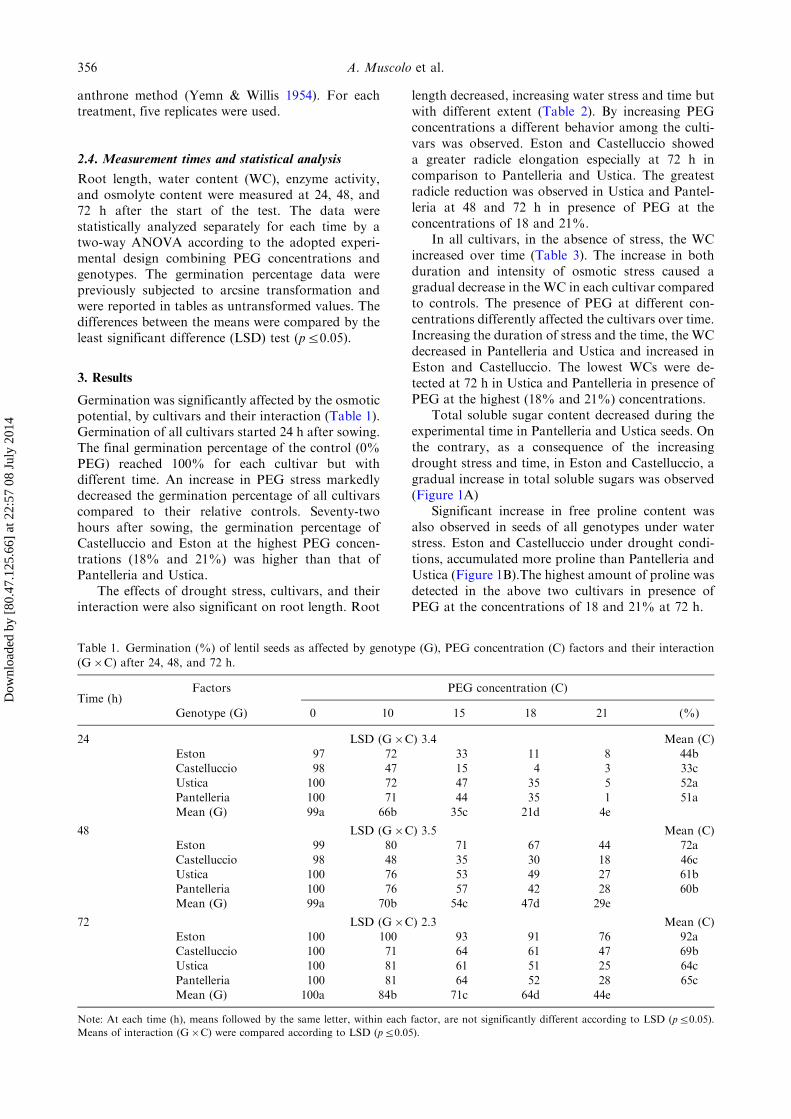
Germination was significantly affected by the osmoticpotential, by cultivars and their interaction (Table 1).Germination of all cultivars started 24 h after sowing.The final germination percentage of the control (0%PEG) reached 100% for each cultivar but withdifferent time. An increase in PEG stress markedlydecreased the germination percentage of all cultivarscompared to their relative controls. Seventy-twohours after sowing, the germination percentage ofCastelluccio and Eston at the highest PEG concen-trations (18% and 21%) was higher than that ofPantelleria and Ustica.
The effects of drought stress, cultivars, and theirinteraction were also significant on root length. Root
length decreased, increasing water stress and time butwith different extent (Table 2). By increasing PEGconcentrations a different behavior among the culti-vars was observed. Eston and Castelluccio showeda greater radicle elongation especially at 72 h incomparison to Pantelleria and Ustica. The greatestradicle reduction was observed in Ustica and Pantel-leria at 48 and 72 h in presence of PEG at theconcentrations of 18 and 21%.
In all cultivars, in the absence of stress, the WCincreased over time (Table 3). The increase in bothduration and intensity of osmotic stress caused agradual decrease in the WC in each cultivar comparedto controls. The presence of PEG at different con-centrations differently affected the cultivars over time.Increasing the duration of stress and the time, the WCdecreased in Pantelleria and Ustica and increased inEston and Castelluccio. The lowest WCs were de-tected at 72 h in Ustica and Pantelleria in presence ofPEG at the highest (18% and 21%) concentrations.
Total soluble sugar content decreased during theexperimental time in Pantelleria and Ustica seeds. Onthe contrary, as a consequence of the increasingdrought stress and time, in Eston and Castelluccio, agradual increase in total soluble sugars was observed(Figure 1A)
Significant increase in free proline content wasalso observed in seeds of all genotypes under waterstress. Eston and Castelluccio under drought condi-tions, accumulated more proline than Pantelleria andUstica (Figure 1B).The highest amount of proline wasdetected in the above two cultivars in presence ofPEG at the concentrations of 18 and 21% at 72 h.
Table 1. Germination (%) of lentil seeds as affected by genotype (G), PEG concentration (C) factors and their interaction
(G�C) after 24, 48, and 72 h.
Time (h)Factors PEG concentration (C)
Genotype (G) 0 10 15 18 21 (%)
24 LSD (G�C) 3.4 Mean (C)
Eston 97 72 33 11 8 44bCastelluccio 98 47 15 4 3 33cUstica 100 72 47 35 5 52a
Pantelleria 100 71 44 35 1 51aMean (G) 99a 66b 35c 21d 4e
48 LSD (G�C) 3.5 Mean (C)Eston 99 80 71 67 44 72a
Castelluccio 98 48 35 30 18 46cUstica 100 76 53 49 27 61bPantelleria 100 76 57 42 28 60bMean (G) 99a 70b 54c 47d 29e
72 LSD (G�C) 2.3 Mean (C)Eston 100 100 93 91 76 92aCastelluccio 100 71 64 61 47 69bUstica 100 81 61 51 25 64c
Pantelleria 100 81 64 52 28 65cMean (G) 100a 84b 71c 64d 44e
Note: At each time (h), means followed by the same letter, within each factor, are not significantly different according to LSD (p50.05).
Means of interaction (G�C) were compared according to LSD (p50.05).
356 A. Muscolo et al.
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014

The values of a-amylase, b-amylase, and
a-glucosidase activities were constitutively different
in the seeds of the four cultivars (Tables 4�6).Theactivity of these enzymes decreased in a dose depen-
dent manner, differing among the cultivars. The
activities of a-, b-amylase, and a-glucosidase in
stressed seeds of Eston and Castelluccio were higher,
compared to Ustica and Pantelleria with respect to
time and stress level. The activities of these enzymes
showed a greater decreasing trend in Pantelleria and
Ustica in presence of PEG at the concentrations of
18% and 21%, already 24 h after sowing. Among
the enzymes involved in the germination process,
a-amylase and a-glucosidase were the most negatively
Table 2. Root length (cm) of lentil seeds as affected by genotype (G), PEG concentration (C) factors and their interaction
(G�C) after 24, 48, and 72 h.
Time (h)Factors PEG concentration (C)
Genotype (G) 0 10 15 18 21 (%)
24 LSD (G�C) 0.31 Mean (C)
Eston 0.69 0.36 0.23 0.17 0.12 0.32aCastelluccio 0.40 0.24 0.14 0.07 0.02 0.17dUstica 0.52 0.27 0.20 0.15 0.08 0.25c
Pantelleria 0.69 0.39 0.24 0.11 0.09 0.30bMean (G) 0.57a 0.31b 0.20c 0.12d 0.08e
48 LSD (G�C) 0.06 Mean (C)Eston 2.51 1.34 1.00 0.68 0.24 1.15a
Castelluccio 1.10 0.83 0.76 0.52 0.20 0.68dUstica 2.03 0.86 0.65 0.34 0.18 0.81cPantelleria 2.85 0.92 0.77 0.47 0.18 1.04bMean (G) 2.12a 0.99b 0.80c 0.50d 0.20e
72 LSD (G�C) 0.06 Mean (C)Eston 4.31 2.22 1.74 1.02 0.83 2.02aCastelluccio 3.94 1.60 1.35 0.74 0.53 1.63bUstica 4.15 1.49 1.12 0.42 0.22 1.48d
Pantelleria 4.95 1.53 0.87 0.33 0.21 1.58cMean (G) 4.33a 1.71b 1.27c 0.63d 0.44e
Note: At each time (h), means followed by the same letter, within each factor, are not significantly different according to LSD (p50.05).
Means of interaction (G�C) were compared according to LSD (p50.05).
Table 3. WC (%) of lentil seeds as affected by genotype (G), PEG concentration (C) factors and their interaction (G�C)after 24, 48, and 72 h.
Time (h)Factors PEG concentration (C)
Genotype (G) 0 10 15 18 21 (%)
24 LSD p50.05 (G�C) 2.3 Mean (C)Eston 19 16 17 14 14 16c
Castelluccio 19 16 17 15 15 16cUstica 25 24 21 19 15 21bPantelleria 38 22 21 22 21 25a
Mean (G) 25a 20b 19b 17c 16c
48 LSD p50.05 (G�C) 2.5 Mean (C)Eston 39 27 28 27 21 29aCastelluccio 31 28 27 29 21 27bUstica 33 34 24 20 16 25c
Pantelleria 37 37 19 19 17 26cMean (G) 35a 32b 25c 24c 19d
72 LSD p50.05 (G�C) 2.9 Mean (C)Eston 40 34 30 26 24 31a
Castelluccio 42 26 25 25 22 28bUstica 35 31 14 12 10 20cPantelleria 45 35 13 7 7 21c
Mean (G) 40a 32b 20c 17d 16e
Note: At each time (h), means followed by the same letter, within each factor, are not significantly different according to LSD (p50.05).
Means of interaction (G�C) are compared according to LSD (p50.05).
Journal of Plant Interactions 357
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014

affected by drought stress especially in Ustica and
Pantelleria cultivars.
4. Discussion
Drought is a multifaceted stress condition that causes
serious crops yield limitations depending on plant
growth stage, stress duration, and severity. Germina-tion is the most critical and sensitive stage in the lifecycles of plants (Ahmad et al. 2009) and the seedsexposed to unfavorable environmental conditionssuch as drought may compromise the subsequentseedling establishment (Albuquerque & Carvalho2003; Soleymani et al. 2012). Genetic variability
Tot
al s
olub
re c
arbo
hydr
ates
[m
g g–
1 (d
w)]
0
20
40
60
80
PantelleriaUsticaEstonCastelluccio
0 h
Tot
al s
olub
le c
arbo
hydr
ates
[m
g g–
1 (d
w)]
0
20
40
60
80
24 h
Tot
al s
olub
le c
arbo
hydr
ates
[m
g g–
1 (d
w)]
0
20
40
60
80
48 h
PEG concentration %
0 5 10 15 20
Tot
al s
olub
le c
arbo
hydr
ates
[m
g g–
1 (d
w)]
0
20
40
60
80
72 h
Prol
ine
[µg
g–1
(dw
)]
0
200
400
6000 h
Prol
ine
[µg
g–1
(dw
)]
0
200
400
600 24 h
Prol
ine
[µg
g–1
(dw
)]
0
200
400
60048 h
PEG concentration %
0 5 10 15 20
Prol
ine
[µg
g–1
(dw
)]
0
200
400
60072 h
A B
Pantelleria
Ustica
Eston
Castelluccio
Figure 1. Total soluble carbohydrates (A) and proline content (B) in Pantelleria, Ustica, Eston, and Castelluccio lentilgenotypes seeds in 0, 10, 15, 18, and 21% PEG 6000 at 0, 24, 48, and 72 h. Vertical bars indicate SD (n�5).
358 A. Muscolo et al.
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014

within a species offers a valuable tool for studying
mechanism of drought tolerance. Our results high-
lighted significant differences among the cultivars
exposed to drought stress with a remarkably de-
creased and delayed germination in Ustica and
Pantelleria. These results are consistent with those
of other studies that have reported that high con-
centrations of PEG reduce the final germination
percentages of lentil (Siahsar et al. 2010; Jamaati-e-
Somarin & Zabihi-e-Mahmoodabad 2011). In Med-
iterranean semiarid regions, the topsoil WC during
the dry season is drastically reduced, sometimes to
less than 1%, which is very close to the permanent
wilting point estimated for xerophytes (Larcher 1995;
Table 4. a-amylase activity (mmoles of reducing sugars formed min�1 g�1 f.w.) of lentil seeds as affected by genotype (G),
PEG concentration (C) factors and their interaction (G�C) after 24, 48, and 72 h.
Time (h)Factors PEG concentration (C)
Genotype (G) 0 10 15 18 21 (%)
24 LSD p50.05 (G�C) 0.49 Mean (C)
Eston 16.12 17.42 17.04 15.49 12.13 15.64cCastelluccio 17.18 16.77 12.41 11.90 11.12 13.88dUstica 26.10 18.97 19.07 14.47 7.70 17.26b
Pantelleria 36.42 31.35 25.28 17.01 7.39 23.49aMean (G) 24.00a 21.12b 18.45c 14.71d 9.58e
48 LSD p50.05 (G�C) 0.21 Mean (C)Eston 16.41 15.31 14.09 13.22 11.13 14.03c
Castelluccio 15.52 14.06 13.37 12.06 11.88 13.37dUstica 27.06 20.09 15.10 10.47 5.69 15.68bPantelleria 36.05 21.07 13.91 11.22 6.64 17.78aMean (G) 23.76a 17.63b 14.11c 11.74d 8.84e
72 LSD p50.05 (G�C) 0.48 Mean (C)Eston 16.42 16.74 15.50 14.70 12.73 15.22cCastelluccio 12.11 11.97 11.52 11.46 11.06 11.62dUstica 24.93 22.47 16.40 11.47 7.70 16.59b
Pantelleria 36.14 25.20 21.12 18.60 6.73 21.56aMean (G) 22.40a 19.09b 16.13c 14.06d 9.55e
Note: At each time (h), means followed by the same letter, within each factor, are not significantly different according to LSD (p50.05).
Means of interaction (G�C) are compared according to LSD (p50.05).
Table 5. b-amylase activity (mmoles of reducing sugars formed min�1 g�1 f.w.) of lentil seeds as affected by genotype (G),
PEG concentration (C) factors and their interaction (G�C).
Time (h)Factors PEG concentration (C)
Genotype (G) 0 10 15 18 21 (%)
24 LSD p50.05 (G�C) 0.09 Mean (C)
Eston 0.75 0.65 0.54 0.43 0.38 0.55aCastelluccio 0.69 0.62 0.51 0.44 0.36 0.53aUstica 1.03 0.55 0.45 0.22 0.19 0.49b
Pantelleria 1.06 0.48 0.41 0.24 0.15 0.47bMean (G) 0.88a 0.58b 0.48c 0.33d 0.27e
48 LSD p50.05 (G�C) 0.33 Mean (C)Eston 0.91 0.88 0.84 0.80 0.64 0.81a
Castelluccio 0.80 0.71 0.65 0.60 0.56 0.67bUstica 1.40 0.75 0.45 0.34 0.23 0.63cPantelleria 1.37 0.73 0.43 0.32 0.22 0.61cMean (G) 1.12a 0.77b 0.60c 0.51d 0.41e
72 LSD p50.05 (G�C) 0.13 Mean (C)Eston 1.17 1.09 0.80 0.73 0.75 0.91aCastelluccio 1.02 0.86 0.72 0.62 0.55 0.75cUstica 1.55 1.35 0.60 0.45 0.24 0.83b
Pantelleria 1.47 1.09 0.52 0.43 0.21 0.75cMean (G) 1.30a 1.09b 0.66c 0.58d 0.44e
Note: At each time (h), means followed by the same letter, within each factor, are not significantly different according to LSD (p50.05).
Means of interaction (G�C) are compared according to LSD (p50.05).
Journal of Plant Interactions 359
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014
Padilla & Pugnaire 2007; Munodawafa 2012). Con-
sequently, plant traits related to water uptake are of
paramount importance for explaining plant persis-
tence in Mediterranean-type ecosystems (Valladares
et al. 2004). The ability to develop extensive root
systems contributes to differences among cultivars for
drought tolerance and root length is considered an
important trait in selection of drought resistant
cultivars (Turner 1997; Abd Allah et al. 2010).
Thus, root morphology and/or growth rate may be
instrumental to select drought tolerant varieties
(Wahbi & Gregory 1995; Malik et al. 2002). In our
study, total root length decreased with increasing
PEG concentrations more in Pantelleria and Ustica
than in the other two genotypes, indicating a higher
sensitivity to osmotic stress in these two landraces.
The root length reduction in Pantelleria and Ustica
under drought stress may be associated to a reduced
cellular division and elongation during germination
(Frazer et al. 1990). This hypothesis is supported
by the results of enzymatic activities involved in the
germination process and by the results of WC.
Water availability is usually the limiting factor for
the germination of non-dormant seeds, affecting
the percentage, speed, and uniformity of emergence
(Marcos-Filho 2005; Kaydan & Yagmur 2008). A
threshold level of hydration is required for the
synthesis of hydrolytic enzymes which are responsible
for the hydrolysis of stored substrates. The hydro-
lyzed products are utilized in seedling tissue synthesis
and radicle elongation (Canas et al. 2006). In lentil
seeds, the role of providing utilizable substrates is
taken over mainly by amylases. Inhibition of seed
germination (Haouari et al. 2013) is directly related to
reserve mobilization, energy production through
respiration, enzyme and hormonal activity, and
dilution of the protoplasm to increase metabolism
for successful embryonic growth (McDonald 2007).
The PEG treatment decreased WC in the seeds of the
studied lentil genotypes, as well documented also for
other species under similar experimental conditions
(Bajji et al. 2000; Guoxiong et al. 2002; Wu et al.
2011; Haouari et al. 2013). Seeds of Castelluccio and
Eston had higher WC compared to those of Ustica
and Pantelleria indicating that the former genotypes
have a more efficient water uptake/control system
that can be related to the increase in the free proline
content and total soluble carbohydrates, suggesting
that drought tolerance ability of these two last
landraces appears to be associated to the accumula-
tion of osmolytes which improved their water status.
The osmolyte content increase is one of the self-
defense reactions during water stress in seeds and
plants protecting the enzyme system. Drought and
salt stress have been reported to limit the mobilization
of starchy endosperm reserves in several species, as a
result of inhibition of different enzymatic activities
(Ashraf & Foolad 2005; Besma & Mounir 2010;
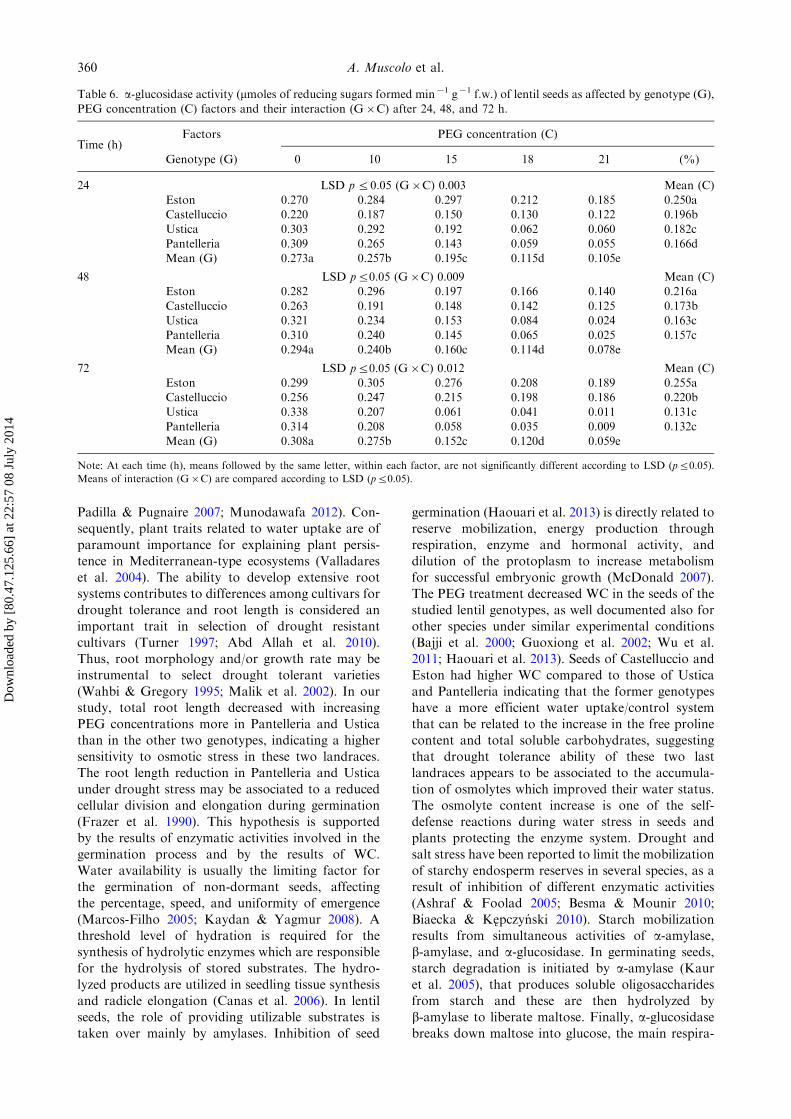
Biaecka & Kepczynski 2010). Starch mobilization
results from simultaneous activities of a-amylase,
b-amylase, and a-glucosidase. In germinating seeds,
starch degradation is initiated by a-amylase (Kaur
et al. 2005), that produces soluble oligosaccharides
from starch and these are then hydrolyzed by
b-amylase to liberate maltose. Finally, a-glucosidasebreaks down maltose into glucose, the main respira-
Table 6. a-glucosidase activity (mmoles of reducing sugars formed min�1 g�1 f.w.) of lentil seeds as affected by genotype (G),
PEG concentration (C) factors and their interaction (G�C) after 24, 48, and 72 h.
Time (h)Factors PEG concentration (C)
Genotype (G) 0 10 15 18 21 (%)
24 LSD p50.05 (G�C) 0.003 Mean (C)
Eston 0.270 0.284 0.297 0.212 0.185 0.250aCastelluccio 0.220 0.187 0.150 0.130 0.122 0.196bUstica 0.303 0.292 0.192 0.062 0.060 0.182c
Pantelleria 0.309 0.265 0.143 0.059 0.055 0.166dMean (G) 0.273a 0.257b 0.195c 0.115d 0.105e
48 LSD p50.05 (G�C) 0.009 Mean (C)Eston 0.282 0.296 0.197 0.166 0.140 0.216a
Castelluccio 0.263 0.191 0.148 0.142 0.125 0.173bUstica 0.321 0.234 0.153 0.084 0.024 0.163cPantelleria 0.310 0.240 0.145 0.065 0.025 0.157cMean (G) 0.294a 0.240b 0.160c 0.114d 0.078e
72 LSD p50.05 (G�C) 0.012 Mean (C)Eston 0.299 0.305 0.276 0.208 0.189 0.255aCastelluccio 0.256 0.247 0.215 0.198 0.186 0.220bUstica 0.338 0.207 0.061 0.041 0.011 0.131c
Pantelleria 0.314 0.208 0.058 0.035 0.009 0.132cMean (G) 0.308a 0.275b 0.152c 0.120d 0.059e
Note: At each time (h), means followed by the same letter, within each factor, are not significantly different according to LSD (p50.05).
Means of interaction (G�C) are compared according to LSD (p50.05).
360 A. Muscolo et al.
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014

tory substrate (Sticklen 2008), with release of theenergy required for essential metabolic functions(Nauriere et al. 1992). Consistently, the activity ofa- and b-amylases in germinating seeds is reduced bywater stress (Zeid & Shedeed 2006). Our resultsconfirmed that the amylase activities in lentil seedsdecreased under PEG induced drought stress, andsuggested that the variation in stress sensitivity ofcontrasting lentil genotypes may be linked to theirability to osmoregulate under stress, which causea strong decrease in WC affecting the hydro-lytic enzyme activities, particularly a-amylase anda-glucosidase levels highlighting the greatest decreasein the most drought sensitive Ustica and Pantelleriaseeds.
The germination of Ustica and Pantelleria, whichare previously identified as NaCl resistant genotypes(Sidari et al. 2007, 2008), was lower than Eston andCastelluccio at the same iso-osmotic PEG concentra-tions. These results indicate that mechanisms mediat-ing drought stress tolerance at germination stage aredifferent from those that mediate salt stress tolerance(Munns 2002). Since PEG does not enter to seeds(Khajeh-Hosseini et al. 2003; Mehra et al. 2003),these differences can be specifically associated tomechanisms that control ion homeostasis and toxicity(Bohnert et al. 1999). Identifying drought resistantcultivars of lentil at early growth stages is essential tocultivate this crop in arid and/or semiarid environ-ments where the survival of other species would bedifficult.
Taken together, these findings suggest that seedgermination, WC, and root length can be used astraits for rapid selection of drought tolerant cultivars.Eston and Castelluccio can be considered as valuabledrought tolerant germplasm. Our data highlighted anopposite response to salt and drought tolerancebecause the two genotypes resulting resistant todrought (Eston and Castelluccio) were found saltsensitive (Sidari et al. 2008) whereas the two drought-sensitive genotypes (Ustica and Pantelleria) werefound NaCl tolerant by the same authors. We canconclude that all these lentil genotypes could be usednot only in breeding programs to improve toleranceto both drought and salinity stress with the aim toincrease the probability of successful legume estab-lishments in arid or semiarid environments but also tobe cultivated in environments where water scarcity isa frequent constraint.
Acknowledgments
This research was supported by ‘‘Mediterranea’’ University
of Reggio Calabria-Italy, Programmi di Ricerca Scientifica
RDB-2011.
References
Abd Allah AA, Badawy Shimaa A, Zayed BA, ElGohary
AA. 2010. The role of root system traits in the drought
tolerance of rice (Oryza sativa L.). World Acad Sci Eng
Technol. 68:1378�1382.Agili S, Nyende B, Ngamau K, Masinde P. 2012. Selection,
yield evaluation, drought tolerance indices of orange-
flesh sweet potato (Ipomoea batatas Lam) Hybrid
Clone. J Nutr Food Sci. 2:138.Ahmad S, Ahmad R, Ashraf MY, Ashraf M, Waraich EA.
2009. Sunflower (Helianthus annuus L.) response to
drought stress at germination and growth stages. Pak J
Bot. 41:647�654.Albuquerque FMCD, Carvalho NMD. 2003. Effect of type
of environmental stress on the emergence of sunflower
(Helianthus annuus L.), soyabean (Glycine max (L.)
Merril) and maize (Zea mays L.) seeds with different
levels of vigor. Seed Sci Technol. 31:65�467.Arjenaki FG, Dehaghi MA, Jabbari R. 2011. Effects of
priming on seed germination of Marigold (Calendula
officinalis). Adv Environ Biol. 5:276�280.Ashraf M, Bokhari H, Cristiti SN. 1992. Variation in
osmotic adjustment of lentil (Lens culinaris, Medik) in
response to drought. Acta Bot Neerl. 41:51�62.Ashraf M, Foolad MR. 2005. Pre-sowing seed treatment �
a shotgun approach to improve germination growth
and crop yield under saline and non-saline conditions.
Advan Agron. 88:223�271.Bajji M, Lutts S, Kinet JM. 2000. Physiological changes after
exposure to and recovery from polyethylene glycol-
induced water deficit in roots and leaves of durum
wheat (Triticum durum Desf.) cultivars differing in
drought resistance. J Plant Physiol. 157:100�108.Bartels D, Sunkar R. 2005. Drought and salt tolerance in
plants. Critical Rev Plant Sci. 24:23�58.Bates LS, Waldren RP, Teare ID. 1973. Rapid determina-
tion of free proline for water stress studies. Plant Soil.
39:205�207.Bergmeyer HU, Grabl M, Walter HE. 1983. Enzymes. In:
Bergmeyer HU, Bergmeyer J, Grabl M, editors.
Methods of enzymatic analysis. Weinheim: Verlag
Chemie; p. 126�328.Besma BD, Mounir D. 2010. Salt stress induced changes in
germination, sugars, starch and enzyme of carbohy-
drate metabolism in Abelmoschus esculentus L.
(Moench.) seeds. Afr J Agric Res. 5:1412�1418.Bewley JD, Black M. 1994. Seeds: physiology of develop-
ment and germination. New York: Plenum Press.Biaecka B, Kepczynski J. 2010. Germination, a-, b-amylase
and total dehydrogenase activities of amaranthus
caudatus seeds under water stress in the presence of
ethephon or gibberellin A3. Acta Biol Cracov Ser Bot.
52:7�12.Bohnert HJ, Su H, Shen B. 1999. Molecular mechanisms
of salinity tolerance. In: Shinozaki K, Yamaguchi-
Shinozaki K, editors. Molecular responses to cold,
drought, heat and salt stress in higher plants. Austin,
TX: Landes Company; p. 29�60.Buchanan BB, Gruissem W, Jones RL. 2000. Biochemistry
and molecular biology of plants. Rockville, MD:
American Society of Plant Physiologists.Canas RA, Canovas FM, Canton FR. 2006. High levels of
asparagines synthetase in hypocotyls of pine seedlings
suggest a role of the enzyme in re-allocation of seed
scord nitrogen. Planta. 224:83�95.Frazer TE, Silk WK, Rost TL. 1990. Effect of low water
potential on cortical cell length in growing region of
maize roots. Plant Physiol. 93:648�651.
Journal of Plant Interactions 361
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014

Guoxiong C, Krugman T, Fahima T, Korol AB, Nevo E.
2002. Comparative study on morphological and phy-
siological traits related to drought resistance between
xeric and mesic Hordeum spontaneum lines in Israel.
Barley Gen Newsl. 32:22�33.Haouari CC, Nasraoui AH, Carrayol E, Gouia H. 2013.
Variations in a-, b-amylase and a-glycosidase activitiesin two genotypes of wheat under NaCl salinity stress.
Afr J Agricult Res. 8:2038�2043.Hodge A, Berta G, Doussan C, Merchan F, Crespi M.
2009. Plant root growth, architecture and function.
Plant Soil. 321:153�187.Hohl M, Schopfer P. 1991. Water relations of growing
maize coleoptiles. Comparison between mannitol and
polyethylene glycol 6000 as external osmotica for
adjusting turgor pressure. Plant Physiol. 95:716�722.IPCC. 2007. Climate change 2007: synthesis report, con-
tribution of working groups I, II and III to the fourth
assessment report of the intergovernmental panel on
climate change. In: Pachauri RK, Reisinger A, editors.
Core writing team. Geneva: IPCC; 104 p.Jamaati-e-Somarin S, Zabihi-e-Mahmoodabad R. 2011.
Evaluation of drought tolerance indices of lentil
varieties. Adv Environ Biol. 5:581�584.Kaur S, Gupta AK, Kaur N. 1998. Gibberellic acid and
kinetin partially reverse the effect of water stress on
germination and seedling growth. Plant Growth Regul.
25:29�33.Kaur S, Gupta AK, Kaur N. 2000. Effect of GA3, kinetin
and indole acetic acid on carbohydrate metabolism in
chickpea seedlings germinating under water stress.
Plant Growth Regul. 30:61�70.Kaur R, Liu X, Goerup O, Zhang A, Yuan X, Balk SP,
Schneider MC, Lu ML. 2005. Activation of p 21-
activated kinase 6 by MAP kinase 6 and p 38 MAP
kinase. J Bio Chem. 280:3323�3330.Kaydan D, Yagmur M. 2008. Germination, seedling
growth and relative water content of shoot in different
seed sizes of triticale under osmotic stress of water and
NaCl. Afr J Biotechnol. 7:2862�2868.Khajeh-Hosseini M, Powell AA, Bingham IJ. 2003. The
interaction between salinity stress and seed vigour
during germination of soybean seeds. Seed Sci Tech-
nol. 31:715�725.Khakwani AA, Dennett MD, Munir M. 2011. Drought
tolerance screening of wheat varieties by inducing
water stress conditions. Songklanakarin J Sci Technol.
33:135�142.Khourgami A, Maghooli E, Rafiee M, Bitarafan Z. 2012.
Lentil response to supplementary irrigation and plant
density under dry farming condition. Int J Sci Adv
Technol. 2:51�55.Kulkarni M, Deshpande U. 2007. In Vitro screening of
tomato genotypes for drought resistance using poly-
ethylene glycol. Afr J Biotechnol. 6:691�696.Larcher W. 1995. Physiological plant ecology. Berlin:
Springer.Malik AI, Colmer TD, Lambers H, Setter TL, Schorte-
meyer M. 2002. Short-term waterlogging has long-term
effects on the growth and physiology of wheat. New
Phytol. 153:225�236.Marcos-Filho JM. 2005. Fisiologia de sementes de plantas
cultivadas [Seed physiology of cultivated 480 plants].
Piracicaba: FEALQ.
McDonald MB. 2007. Seed moisture and the equilibrium
seed moisture content curve. J Seed Technol. 29:7�18.Mehra V, Tripathi J, Powell AA. 2003. Aerated hydration
treatment improves the response of Brassica juncea and
Brassica campestris seeds to stress during germination.
Seed Sci Technol. 31:57�70.Mittler R. 2006. Abiotic stress, the field environment and
stress combination. Trends Plant Sci. 11:15�19.Munodawafa A. 2012. The effect of rainfall characteristics
and tillage on sheet erosion and maize grain yield in
semiarid conditions and granitic sandy soils of Zim-
babwe. Appl Environ Soil Sci. 2012:1�8.Munns R. 2002. Comparative physiology of salt and water
stress. Plant Cell Environ. 25:239�250.MwaleS,HamusimbiC,MwansaK.2003.Germination, emer-
gence and growth of sunflower (Helianthus annuus L.) in
response to osmotic seed priming. Seed Sci Technol.
31:199�206.Nauriere C, Doyen C, Thevenot C, Dauffant J. 1992.
b-amylase in cereals: study of the maize b-amylase
system. Plant Physiol. 100:887�893.Okcu G, Kaya MD, Atak M. 2005. Effects of salt and
drought stresses on germination and seedling growth
of pea (Pisum sativum L.). Turk J Agric For. 29:237�242.
Padilla FM, Pugnaire FI. 2007. Rooting depth and soil
moisture control Mediterranean woody seedling survi-
val during drought. Funct Ecol. 21:489�495.Richards RA. 1978. Variation between and within species
of rapeseed (Brassica campestris and B. napus) in
response to drought stress. III. Physiological and
physicochemical characters. Aust J Agric Res. 29:
491�501.Salehi M, Haghnazari A, Shekari F, Faramarzi A. 2008.
The study of seed yield and seed yield components of
lentil (Lens culinarisMedik) under normal and drought
stress conditions. Pak J Biol Sci. 11:758�62.Shaheen R, Hood-Nowotny RC. 2005. Effect of drought
and salinity on carbon isotope discrimination in wheat
cultivars. Plant Sci. 168:901�909.Siahsar BA, Ganjali S, Allahdoo M. 2010. Evaluation of
drought tolerance indices and their relationship with
grain yield of lentil lines in drought-stressed and
irrigated environments. Aust J Basic Appl Sci.
4:4336�4346.Sidari M, Muscolo A, Anastasi U, Preiti G, Santonoceto C.
2007. Response of four genotypes of lentil to salt stress
conditions. Seed Sci Technol. 35:497�503.Sidari M, Santonoceto C, Anastasi U, Preiti G, Muscolo A.
2008. Variations in four genotypes of lentil under
NaCl-salinity stress. Am J Agric Biol Sci. 3:410�416.Silim SN, Saxena MC, Erskine W. 1993. Adaptation of
lentil to the Mediterranean environment. II. Response
to moisture supply. Exp Agric. 29:21�28.Singh KB, Saxena MC. 1990. Studies on drought tolerance:
Annual report. Aleppo, Syria: ICARDA.Soleymani A, Hesam M, Shahrajabian H. 2012. Study of
cold stress on the germination and seedling stage and
determination of recovery in rice varieties. Int J Biol.
4:23�30.Steup M. 1988. Starch degradation. In: Preiss J, editor. The
biochemistry of plants: a comprehensive treatise. San
Diego, CA: Academic Press; p. 255�296.
362 A. Muscolo et al.
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014

Sticklen MB. 2008. Plant genetic engineering for biofuelproduction: towards affordable cellulosic ethanol. NatRev Genet. 9:433�443.
Tuberosa R, Salvi S. 2006. Genomics-based approaches toimprove drought tolerance of crops. Trends Plant Sci.11:405�412.
Turner NC. 1997. Further progress in crop water relations.Adv Agron. 58:293�338.
Valladares F, Vilagrosa A, Penuelas J, Ogaya R, Camarero
JJ, Corchera L, Siso S, Gil-Pelegrın E. 2004. Estreshıdrico: ecofisiologıa y escalas de la sequıa [Waterstress: ecophysiology and scales of drought]. In:Valladeres F, editor. Ecologıa del BosqueMediterraneo
en un Mundo Cambiante. Madrid: EGRAF, SA;p. 163�190.
Wahbi A, Gregory PJ. 1995. Growth and development of
young roots of barley (Hordeum vulgare L.) genotypes.Ann Bot. 75:533�539.
Wu C, Wang Q, Xie B, Wang Z, Cui J, Hu T. 2011. Effects
of drought and salt stress on seed germination of three
leguminous species. Afr J Biotechnol. 10:17954�17961.Yemn EW, Willis AJ. 1954. The estimation of carbohy-
drates in plant extracts by anthrone. Biochem J.
57:508�514.Zeid IM, Shedeed ZA. 2006. Response of alfalfa to
putrescine treatment under drought stress. Biol Plant.
50:635�640.Zeigler RS, Puckridge DW. 1995. Improving sustainable
productivity in rice based rainfed lowland systems of
South and Southeast Asia. Feeding four billion people:
the challenge for rice research in the 21st century.
GeoJournal. 35:307�324.Zohary D. 1972. The wild progenitor and place of origin
of the cultivated lentil Lens culinaris. Econ Bot.
26:326�332.
Journal of Plant Interactions 363
Dow
nloa
ded
by [
80.4
7.12
5.66
] at
22:
57 0
8 Ju
ly 2
014