efficacy of lactobacillus acidophilus on the sperm quality...
TRANSCRIPT

International Journal of Chemical and Analytical Science ISSN: 0976-1206
Research Article
www.ijcas.info
Corresponding Author: Somnath Chakraborty, Faculty of Marine Sciences, Centre of Advanced Study in Marine Biology, Annamalai University,
Parangipettai – 608502, Tamil Nadu, India Received 22-11-2010; Accepted 21-07-2011
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1178
Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon (Fabricius, 1798)
MA Badhul Haq, R Manimurugan, AR Nazar, R Shalini, Upasana Ghosh, Somnath Chakraborty* Faculty of Marine Sciences, Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai – 608 502, Tamil Nadu, India
INTRODUCTION
The annual global production of farmed shrimp has grown at
the rate of 8% since 1989. In 1998, the production reac hed
1,113,887 mt, representing around 32% of world shrimp and
prawn catches [1]. T he industry has expanded mostly through
Southeast Asia and Latin America, where diverse penaeid
species are cultured. The most important species in Latin America is Penaeus (Litopenaeus) vannamei, followed by P.
(Litopenaeus) stylirostris and P. (Litopenaeus) schmitii. In Asia P. monodon, P. chinesis, P. (Marsupenaeus) japonicus, P. pencillatus,
P. merguiensis, and P. indicus are cultured [2]. The traditional
dependency on wild stock for naupii production will thus
change towards a more controlled production of genetically
improved animals, selected for impo rtant aquaculture traits
like growth rate and dis ease resistance [3]. However, t he
genetic improvement of shrimp requires the application of
breeding programs, which relay to excellent gamete quality,
controlled fertilization, and, if possible, cryogenic storage. For
this a proper understanding of basic shrimp reproductive
biology, and the developm ent of in vitro fertilization
techniques and cryogenic p rotocols for gametes, embryos,
and larvae are needed.
Female maturation is achieved by u nilateral eyestalk ablation.
This technique is applied to each female at intermolt, selecting
one of t he eyestalks. Few days after the p rocedu re, ovaries
will start to grow and first spawning will occu rs within a
week, with an increasing rate in t he subsequent days. Each
female generates various spawns, but eventually t he
spawning activity decreases, until around 2 to 3 months after
ablation, when t hey are rep laced by new females.
Eyestalk ablation affects all aspects of shrimp physiology [4]
and over time it has a deleterious effect on spawn quality and
quantity, as well as on survival [5, 6, 7]. This dependency on
eyestalk ablation is a major bottleneck for the adva nce of
controlled reproduction, and major research activities are
dedicated to develop hormonal treatments for controlled maturation. The reproductive cycle of the genus Penaeus,
consisting of two phases was desc ribed [8], as in the first
phase, pre-adults migrate from estuarine waters to a
reproduction zone in deeper a nd higher saline waters. After
spawning occurs, the postlarvae migrate to estuarine waters. This pattern is fol lowed by various species: P. duorarum, P. brasiliensis, P. aztecus, P. schmitii, P. stylirostris, P. vannamei, P.
setiferus and P. occidentalis [9 ]. The quality of wild
spermatophores during natural mating has been partially
documented. This type of analysis has been reported for
males under cu lture conditions (controlled reproduction) and wild P. setiferus broodstock [10, 1 1, 12].
The evaluations of reproductive quality in male penaeid
shrimp was first repo rted by resea rchers [13] who measured
spermatophores weight, sperm count, and p ercentages of live
and abnormal sperm for evaluating the effect of eyestalk ablation in P. vannamei. Using the same parameters, pioneer
researchers [10 ] studied a decline of sperm quality in P. setiferus
held in captivity a nd others [14] used t he same approach to evaluate the effect of water t emperature, EDTA, and Vibrio
bacterin o n sperm quality in captive P. setiferus. The effects of
dietary rancidity and Vitamin E on gonadal maturation of P.
setiferus were assessed [11].
In the present research, Penaeus monodon broodstock was collected f rom a Shrimp hatchery. After their acclimatization healthy
male P. monodon were selected based on size, general appearance, p resence of spermatophores from the culture tanks for the
probiotic test. Shrimp brooders were disinfected with formalin and maintained in the tanks for 3 days before beginning
experiments. All experimental animals were tra nsferred into epoxy coated rectangula r tank along with the control tank
maintained at identical relevant stocking density after treating t hem with intramuscular injection, containing Lactobacillus
acidophilus in different quantities (0.5, 1.0, 1.5 and 2.0 ml containing 3 × 109 cells/ml) into lateral su rface of the p receding segment of male brooder. Gonopores of male P. monodon were washed with sterile seawater and 70% ethanol prior to
collection of spermatophores. Manual ejaculation of spermatophores was carried out using elect ric stimulus on all males. Two
spermatophores were obtained from each male that were weighed and the sperm quality was examined morphologically
using light microscope. The length a nd weight of the shrimps were measured after subsequent ejaculation. Percentage of
viable sperm was calculated in triplicate from counting of a minimum number of 250 sperm cells under 1000-fold
magnification on each slide. Rep roductive routine was evaluated in the experimental animals inj ected on t he zeroth day of
initiation of the experiment with 0.5, 1.0, 1.5 and 2.0µl/g body weights of shrimp. During the four different treatments in 1.0µl
injected males performed differently. In the I extrusion, mean sperm counts in GM ranged from left vs right spermatophore
was 0.842, 0.912, 0.940 and 0.972; in II extrusion 0.521, 0.422, 0.988 and 0.591, in III extrusion 0.711,0.451, 0.910 and 0.740 and in
the IV extrusion 0.421, 0.422, 0.945 and 0.431 in the successive intervals respectively. Higher counts were recorded in the right halve of spermatophore of shrimps treated with 0.1µl. Among all the L. acidophilus treated broodstock shrimps, only 0.1µl/g
of body weight performed better after 7 days of administration. Therefore, this research depicts that the application of L.
acidophilus enha nces sperm quality and regenerates the spermatophores in male P. monodon brooders.
Key words – Penaeus Monodon, Spermatophores, Probiotics, Lactobacillus Acidophilus, Gonopores, Electric Stimulus

M. A. Badhul Haq, et al.: Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon
(Fabricius, 1798)
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1179
Researchers [10] evaluated spermatophores redevelopment time in wild P. stylirostris, P. vannamei, P. setiferus; and also
studied the effect of ascorbic acid on reproductive quality in P. vannamei using the previous mentioned quality indicato rs.
Eminent workers measured spermatophores weight, sperm
count, and percentage of abnormalities for evaluating
nutritional variations on t he onset of the Male Reproductive Tract Degenerative Syndrome (MRTDS) of P. setiferus [15].
Spermatophores of p enaeid s hrimp may vary considerably in
quality and have been evaluated in terms of spermatophores
weight, sperm count, and t he percentage incidence of
abnormalities [11, 13]. Recently the activation of an acrosome
reaction by the egg - water technique was developed [16] and
has been evaluated as a more accurate measure of sperm
quality [17, 18 ]. A new approach of inducing gonadal
maturation and spawning was defined [19]. This approach
relies in the roles t hat neurotransmitters play in regulating
gonadal maturation. Workers [20] established that 5-
hydroxtryptamine (5-HT, serotonin) induces ovarian maturation invivo and invitro, in Procambarus clarkia, by
acting to stimulate release of a gonad-stimulating hormone
(GSH) from t he brain and thoracic ga nglia; GSH is an abstract
entity, which has not been identified nor measured nor measured directly. The effect of Bacillus spp. used as
probiotics on digestive enzyme activity, survival and growth in the Indian white shrimp Fenneropenaeus indicus was proved [21]. Microbial flora of spermatophores from black tiger s hrimp
(Penaeus monodon) declines over long-term cryostorage was
also showed [22].
Serotonin also stimulates the release of other neurohormones,
including the crustacean hyp erglycaemic hormone (CHH),
red pigment-dispers ing hormone (RPDH), neurodepressing
hormone (NDH) and molt-inhibiting hormone (MIH) [20].
Methyl farnesoate (MF) is a factors that stimulates gonadal
maturation [23] and it is inhibited by 5-HT; on the other hand,
RPCH stimulates MF synthesis by the mandibular organ;
other roles played by 5- HT are migration of the proximal
retinal pigment, pericardial organ neurohormone,
stomatogastric ganglion neuromodu lator or nerohormone,
behavioural responses, osmoregulation, and
mechanoreception [19] as a effort to develop alternatives for
the comm ercial rep roduction of penaeid shrimp, this study
was conducted to evaluate the effect of two doses of 5-HT on P. vannamei maturation and spawning quality, compared to
unilateral eyestalk ablation.
The aim of the present investigation is to assess the effect of Lactobacillus acidophilus (different quantities) on the sperm
quality, and there was four different extrusions were
performed in 7 days intervals. Besides quality and quantity
present between the extrusions, an emphasis is also paid to
evaluate the regeneration interval of spermatophores.
OBJECTIVES 1. To study the efficacy of probiotics Lactobacillus acidophilus
on broodstock Penaeus monodon.
2. To find out the effectiveness of sp erm quality and regeneration using probiotics bacteria L. acidophilus in
different level dosage.
3. To determine the probiotics profiles in the different level of
ejaculations.
MATERIAL AND METHODS Penaeus monodon broodstock was collected from a Shrimp
hatchery collection centre in Marakkanam, Pondicherry. The
present study was carried out at the Faculty of Marine Sc ience
laboratory, CAS in Marine Biology, Annamalai University,
Tamil Nadu, India. The collection area is depicted in Fig.1. Experimental design
Fig. 1: Location of the shrimp hatchery in Pondicherry
After acclimatization at the laboratory for 5 days, uniform
size brooders were selected for probiotic testing (F ig. 2).
Fig. 2: P. monodon broodstock in experimental tank
Healthy male P. monodon were selected based on size, general
appearance, presence of spermatophores from the culture
tanks with the ABW of 65 – 80 g weight were stocked at the
density of 10 shrimps per tank (Fig. 3).

M. A. Badhul Haq, et al.: Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon
(Fabricius, 1798)
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1180
Fig. 3: View of length and weight measurement of P. monodon
Before stocking, the shrimps were disinfected with 20 ppm
formalin for 20 minutes. Initially the animals were stocked in
three different one tonned capacity annular synthetic tanks.
Shrimp were maintained in the tanks for 3 days before
beginning experiments. 100% water tune was done daily and
initial stocking of 40 brooders. After intramuscular injection,
all experimental animals were t ransferred in to 25 l capacity
epoxy coated rectangular tank (Fig. 4). Control tank were
maintained at identical relevant stocking density. Three
replicates were used for a total of 120 broodstock per
treatment.
Fig. 4: The view of experimental set up
Probiotic (Lactobacillus acidophilus) challenge test
Bacterial inoculums: L. acidophilus probiotic inocu lum was
prepared for int ramuscular injection in different dosage to the experimental animal P. monodon broodstock.
Microorganism: Among commonly used p robiotic organisms
in endothermic animals, only few can be compared to the
wealth of information available on probiotic bacteria; most
studies demonstrate the use of the same in aquaculture.
Consequently for t he p resent study, one benefic ial probionts
was commonly encountered as Lactobacillus acidophilus
strain (Fig. 5), that was obtained from the Marine
Microbiology Laboratory, Faculty of Marine Science, where it
was used as the probiotic organism. The authenticity of the
culture was verified by sub culturing and identification based
on t heir morphological and biochemical characteristics
(Bergey’s Manual of Systematic Bacteriology).
Fig. 5: Slant culture of Lactobacillus acidophilus
Production of bacterial biomass
L. acidophilus was grown aseptically in 10 ml of nutrient broth
for 24 hr at room temperature (28 ± 2 °C). 5 ml of log phase
culture was then transferred under aseptic conditions into 250
ml of MRS broth and p laced on a rotary shaker at 150 rpm for 24 hr at 28 ± 2°C. The cells of L. acidophilus strain were
harvested separately by cent rifuging at 10, 000 rpm under
sterile conditions for 15 min at 4°C. The liquid supernatant
was then discarded, and the pellet was suspended in sterile
saline solution. This process was rep eated again, and the cell
concentration in t he suspension was adjusted to an optical
density of 1.00 at 610 nm in a spectrophotometer (model DR-
2000; Hach, Loveland, Colo.). To estimate the bacterial
concentration achieved, the suspension was serially diluted in
sterile saline and spread plated in nutrient agar.
Brood stock conditioning and intramuscular injection
All the brood stock shrimps were injected the probiotic Lactobacillus acidophilus with different quantities (0.5, 1.0, 1.5
and 2.0 ml containing 3 × 109 cells/ml) into lateral surface of
the preceding segment of male brooder two days after
stocking into t he experimental tanks (Fig. 6) and (Table. 1 - 4).
Each experimental tank were stocked with 10 shrimps.
Experimental animals were fed with fresh frozen and
chopped squid, oyster and polycheate worms at 8:00, 13:00
and 18:30 hrs daily. Uneaten food was removed 3 hrs after
feeding. Feeds were given at the rate of 20% of the body
weight per day. Water quality remained constant between t he
treatments. No females were stocked, eliminating t he
possibilities of intervention. During stocking, the males were
marked with synthetic coloured tags.
Fig. 6: L. acidophilus for intramuscular injection
M. A. Badhul Haq, et al.: Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon
(Fabricius, 1798)
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1181
Spermatophores collection
Male P. monodon were washed with sterile seawater and 70%
ethanol was sprayed a round t he gonopores prior to collection
of spermatophores. Manual ejaculation of spermatophores
was performed on all males by stripping around the base of
the fifth pairs of walking legs (pereiopods) with forceps and
aseptic technique. Two spermatophores were obtained from
each male. The length and weight of the shrimps were
measured and t he shrimps were released into the tanks
initially and also after subsequent ejaculation.
Evaluation of sperm viability
Spermatophores were removed using a technique [24], in
which electrical stimulus was applied to the coxae of fifth pair
of pereiopods. The spermatophores were lifted from t he
gonop hores using forceps, dipped in Ca free saline solution
(NaCl- 21.63g; KCl-1.12g; H3BO3-0.53g; NaOH-0.19g and
MgSO4.7H2O-4.93g the pH of which was adjusted to7.4 with
1N HCl). The samples were prot ected from cross-
contamination.
All the extruded spermatophores were weighted to t he
nearest 0.01mg on an electronic bala nce, before
homogenization in 1 ml of Ca free saline as suggested [10], and
the sperm suspensions were prepared uniformly. Quality of
sperm was assayed using the met hod as described [18].
Percentage of morphologically normal sperm was determined
by recording the number of normal sp erm with a single spike
that project from an oral body or nucleus and abnormal
sperm with malformed bod y or bent or missing spike.
A sample of sperm suspension (50μl) was transferred to a
slide, mixed with 50 μL of Haematoxylin and Eosin, dried
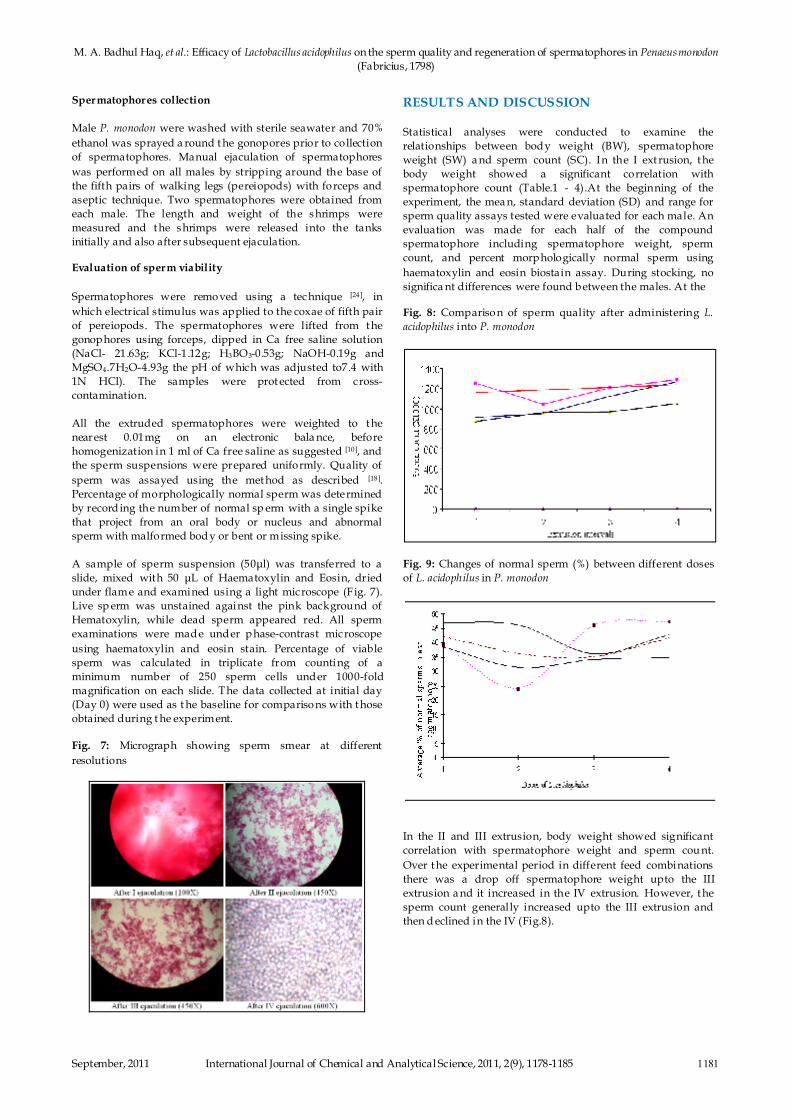
under flame and examined using a light microscope (Fig. 7).
Live sp erm was unstained against the pink background of
Hematoxylin, while dead sperm appeared red. All sperm
examinations were made under p hase-contrast microscope
using haematoxylin and eosin stain. Percentage of viable
sperm was calculated in triplicate from counting of a
minimum number of 250 sperm cells under 1000-fold
magnification on each slide. The data collected at initial day
(Day 0) were used as t he baseline for comparisons with t hose
obtained during t he experiment.
Fig. 7: Micrograph showing sperm smear at different
resolutions
RESULTS AND DISCUSSION
Statistical analyses were conducted to examine the
relationships between body weight (BW), spermatophore
weight (SW) a nd sperm count (SC). In the I ext rusion, t he
body weight showed a significant correlation with
spermatophore count (Table.1 - 4).At the beginning of the
experiment, the mea n, standard deviation (SD) and range for
sperm quality assays tested were evaluated for each male. An
evaluation was made for each half of the compound
spermatophore including spermatophore weight, sperm
count, and percent morphologically normal sperm using
haematoxylin and eosin biostain assay. During stocking, no
significa nt differences were found b etween the males. At the
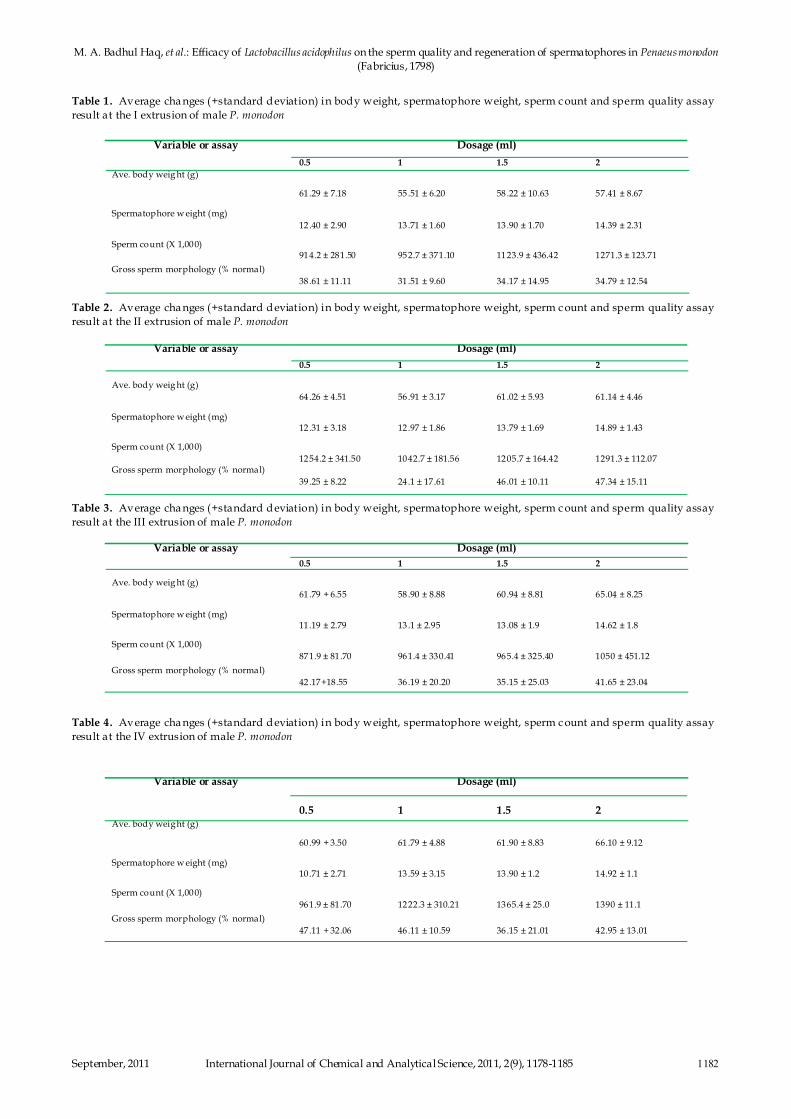
Fig. 8: Comparison of sperm quality after administering L.
acidophilus into P. monodon
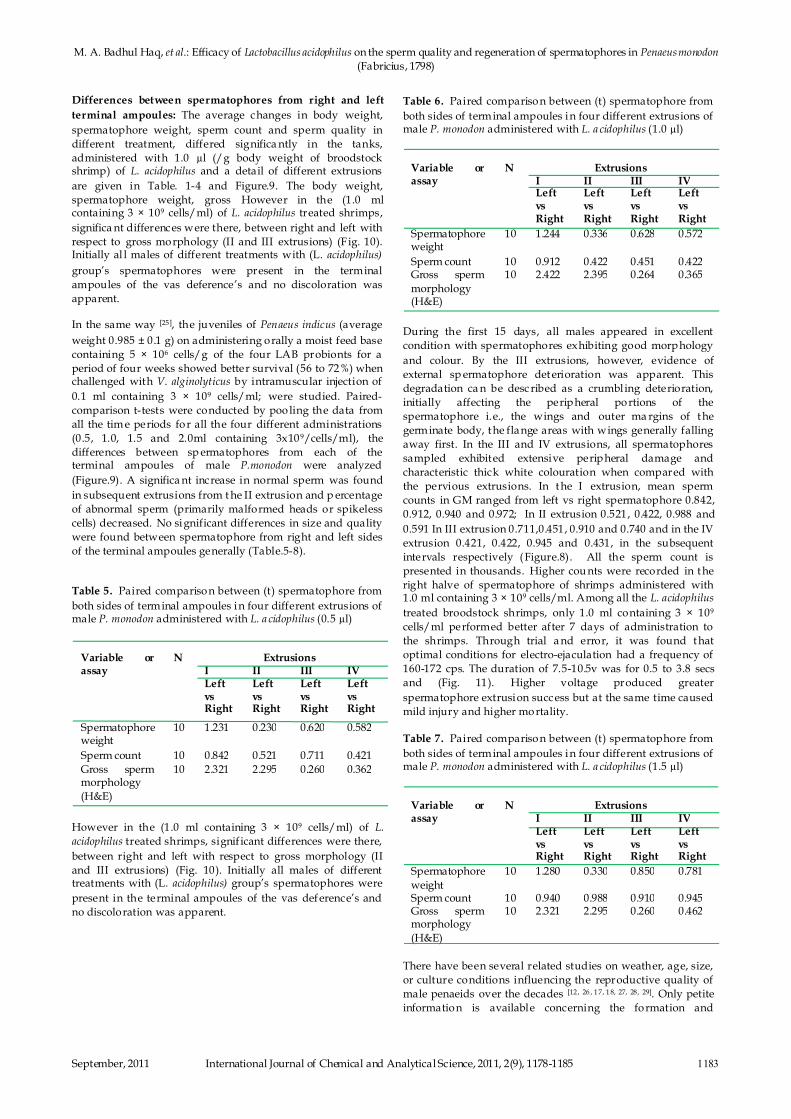
Fig. 9: Changes of normal sperm (%) between different doses
of L. acidophilus in P. monodon
In the II and III extrusion, body weight showed significant
correlation with spermatophore weight and sperm cou nt.
Over t he experimental period in different feed combinations
there was a drop off spermatophore weight upto the III
extrusion a nd it increased in the IV extrusion. However, t he
sperm count generally increased upto the III extrusion and
then d eclined in the IV (Fig.8).
M. A. Badhul Haq, et al.: Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon
(Fabricius, 1798)
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1182
Table 1. Average cha nges (+standard deviation) in body weight, spermatophore weight, sperm c ount and sperm quality assay
result at the I extrusion of male P. monodon
Table 2. Average cha nges (+standard deviation) in body weight, spermatophore weight, sperm c ount and sperm quality assay
result at the II extrusion of male P. monodon
Table 3. Average cha nges (+standard deviation) in body weight, spermatophore weight, sperm c ount and sperm quality assay
result at the III extrusion of male P. monodon
Variable or assay Dosage (ml)
0.5 1 1.5 2
Ave. body weig ht (g)
61 .79 + 6.55 58 .90 ± 8.88 60 .94 ± 8.81 65.04 ± 8.25
Spermatophore w eight (mg) 11 .19 ± 2.79 13 .1 ± 2.95 13 .08 ± 1.9 14.62 ± 1.8
Sperm count (X 1,000)
871.9 ± 81.70 961.4 ± 330.41 965.4 ± 325.40 1050 ± 451.12
Gross sperm morphology (% normal)
42 .17+18.55 36 .19 ± 20.20 35 .15 ± 25.03 41.65 ± 23.04
Table 4. Average cha nges (+standard deviation) in body weight, spermatophore weight, sperm c ount and sperm quality assay
result at the IV extrusion of male P. monodon
Variable or assay Dosage (ml)
0.5 1 1.5 2
Ave. body weig ht (g)
61 .29 ± 7.18 55 .51 ± 6.20 58 .22 ± 10.63 57.41 ± 8.67
Spermatophore w eight (mg)
12 .40 ± 2.90 13 .71 ± 1.60 13 .90 ± 1.70 14.39 ± 2.31
Sperm count (X 1,000) 914.2 ± 281.50 952.7 ± 371.10 1123.9 ± 436.42 1271.3 ± 123.71
Gross sperm morphology (% normal)
38 .61 ± 11.11 31 .51 ± 9.60 34 .17 ± 14.95 34.79 ± 12.54
Variable or assay Dosage (ml)
0.5 1 1.5 2
Ave. body weig ht (g)
64 .26 ± 4.51 56 .91 ± 3.17 61 .02 ± 5.93 61.14 ± 4.46
Spermatophore w eight (mg) 12 .31 ± 3.18 12 .97 ± 1.86 13 .79 ± 1.69 14.89 ± 1.43
Sperm count (X 1,000)
1254.2 ± 341.50 1042.7 ± 181.56 1205.7 ± 164.42 1291.3 ± 112.07 Gross sperm morphology (% normal)
39 .25 ± 8.22 24 .1 ± 17.61 46 .01 ± 10.11 47.34 ± 15.11
Variable or assay Dosage (ml)
0.5 1 1.5 2 Ave. body weight (g)
60 .99 + 3.50 61 .79 ± 4.88 61 .90 ± 8.83 66.10 ± 9.12
Spermatophore w eight (mg) 10 .71 ± 2.71 13 .59 ± 3.15 13 .90 ± 1.2 14.92 ± 1.1
Sperm count (X 1,000)
961.9 ± 81.70 1222.3 ± 310.21 1365.4 ± 25.0 1390 ± 11.1
Gross sperm morphology (% normal)
47 .11 + 32.06 46 .11 ± 10.59 36 .15 ± 21.01 42.95 ± 13.01
M. A. Badhul Haq, et al.: Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon
(Fabricius, 1798)
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1183
Differences between spermatophores from right and left
terminal ampoules: The average changes in body weight,
spermatophore weight, sperm count and sperm quality in
different treatment, differed significa ntly in the tanks,
administered with 1.0 µl (/g body weight of broodstock shrimp) of L. acidophilus and a detail of different extrusions
are given in Table. 1-4 and Figure.9. The body weight,
spermatophore weight, gross However in the (1.0 ml containing 3 × 109 cells/ml) of L. acidophilus treated shrimps,
significa nt differences were there, between right and left with
respect to gross morphology (II and III extrusions) (F ig. 10). Initially al l males of different treatments with (L. acidophilus)
group’s spermatophores were present in the terminal
ampoules of the vas deference’s and no discoloration was
apparent.
In the same way [25], the juveniles of Penaeus indicus (average
weight 0.985 ± 0.1 g) on administering orally a moist feed base
containing 5 × 106 cells/g of the four LAB probionts for a
period of four weeks showed better survival (56 to 72%) when challenged with V. alginolyticus by intramuscular injection of
0.1 ml containing 3 × 109 cells/ml; were studied. Paired-
comparison t-tests were conducted by poo ling the data from
all the tim e periods for all the four different administrations
(0.5, 1.0, 1.5 and 2.0ml containing 3x109/cells/ml), the
differences between sp ermatophores from each of the terminal ampoules of male P.monodon were analyzed
(Figure.9). A significa nt increase in normal sperm was found
in subsequent extrusions from t he II extrusion and p ercentage
of abnormal sperm (primarily malformed heads or spikeless
cells) decreased. No significant differences in size and quality
were found between spermatophore from right and left sides
of the terminal ampoules generally (Table.5-8).
Table 5. Paired comparison between (t) spermatophore from
both sides of terminal ampoules in four different extrusions of male P. monodon administered with L. a cidophilus (0.5 µl)
Variable or assay
N
Extrusions
I II III IV Left vs Right
Left vs Right
Left vs Right
Left vs Right
Spermatophore weight
10 1.231 0.230 0.620 0.582
Sperm count 10 0.842 0.521 0.711 0.421
Gross sperm morphology
(H&E)
10 2.321 2.295 0.260 0.362
However in the (1.0 ml containing 3 × 109 cells/ml) of L.
acidophilus treated shrimps, signif icant differences were there,
between right and left with respect to gross morphology (II
and III extrusions) (Fig. 10). Initially all males of different treatments with (L. acidophilus) group’s spermatophores were
present in the terminal ampoules of the vas deference’s and
no discoloration was apparent.
Table 6. Paired comparison between (t) spermatophore from
both sides of terminal ampoules in four different extrusions of male P. monodon administered with L. a cidophilus (1.0 µl)
Variable or assay
N
Extrusions
I II III IV Left vs Right
Left vs Right
Left vs Right
Left vs Right
Spermatophore weight
10 1.244 0.336 0.628 0.572
Sperm count 10 0.912 0.422 0.451 0.422 Gross sperm
morphology (H&E)
10 2.422 2.395 0.264 0.365
During the first 15 days, all males appeared in excellent
condition with spermatophores exhibiting good morphology
and colour. By the III extrusions, however, evidence of
external sp ermatophore det erioration was apparent. This
degradation ca n be desc ribed as a crumbling deterioration,
initially affecting the perip heral po rtions of the
spermatophore i.e., the wings and outer ma rgins of t he
germinate body, t he flange areas with wings generally falling
away first. In the III and IV extrusions, all spermatophores
sampled exhibited extensive perip heral damage and
characteristic thick white colouration when compared with
the pervious extrusions. In t he I extrusion, mean sperm
counts in GM ranged from left vs right spermatophore 0.842,
0.912, 0.940 and 0.972; In II extrusion 0.521, 0.422, 0.988 and
0.591 In III extrusion 0.711,0.451, 0.910 and 0.740 and in the IV
extrusion 0.421, 0.422, 0.945 and 0.431, in the subsequent
intervals respectively (Figure.8). All the sperm count is
presented in thousands. Higher cou nts were recorded in t he
right halve of spermatophore of shrimps administered with 1.0 ml containing 3 × 109 cells/ml. Among all the L. acidophilus
treated broodstock shrimps, only 1.0 ml containing 3 × 109
cells/ml performed better after 7 days of administration to
the shrimps. Through trial a nd error, it was found t hat
optimal conditions for electro-ejaculation had a frequency of
160-172 cps. The duration of 7.5-10.5v was for 0.5 to 3.8 secs
and (Fig. 11). Higher voltage produced greater
spermatophore extrusion success but at the same time caused
mild injury and higher mo rtality.
Table 7. Paired comparison between (t) spermatophore from
both sides of terminal ampoules in four different extrusions of male P. monodon administered with L. a cidophilus (1.5 µl)
Variable or assay
N
Extrusions
I II III IV Left vs Right
Left vs Right
Left vs Right
Left vs Right
Spermatophore
weight
10 1.280 0.330 0.850 0.781
Sperm count 10 0.940 0.988 0.910 0.945 Gross sperm morphology
(H&E)
10 2.321 2.295 0.260 0.462
There have been several related studies on weather, age, size,
or culture conditions influencing the reproductive quality of
male penaeids over the decades [12, 26 , 1 7, 1 8, 27, 28, 29]. Only petite
information is available concerning the formation and
M. A. Badhul Haq, et al.: Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon
(Fabricius, 1798)
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1184
development of spermatophores. Resea rchers [12] reported that pond-reared Penaeus stylirost ris were found to p roduce
spermatophores in individuals, larger that 23.6g, and
spermatophores were s ignificantly lower in weight for the 20-
30g shrimp that the 30-40 g shrimp. Similar scenario is noticed
in the p resent investigation.
Table 8. Paired comparison between (t) spermatophore from
both sides of terminal ampoules in four different extrusions of male P. monodon administered with L. a cidophilus (2.0 µl)
Variable or assay
N
Extrusions
I II III IV Left vs Right
Left vs Right
Left vs Right
Left vs Right
Spermatophore
weight
10 1.801 0.563 0.712 0.621
Sperm count 10 0.974 0.591 0.740 0.431 Gross sperm morphology
(H&E)
10 2.321 2.295 0.260 0.362
Fig. 10: Ejaculation of spermatophores with electric stimulus
at different time interval
Fig. 11: Electric stimulus applied on the spermatophore
The earliest time of spermatophore formation was different.
The age and size of males influ ence spermatophore
formation. The difference may be due to the growth rate and
culture conditions t hat differed for the t wo environments.
Similar result was repo rted for Penaeus merguiensis [30]; but
they suggested that the primary sexual maturity of prawns
depends upon size rather than age.
CONCLUSION
Reproductive performance was evaluated in sexually mature wild male P. monodon administered on the zeroth day of
initiation of the experiment with 0.5, 1.0, 1.5 and 2.0 µl /g
body weight of shrimp. Among the four d ifferent treatments
in 1.0 µl administered males p erformed in signif icantly
different.mannner. In the I extrusion, mean sperm counts in
GM ranged from left vs right spermatophore 0.842, 0.912,
0.940 and 0.972; in II extrusion 0.521, 0.422, 0.988 and 0.591, in
III extrusion 0.711,0.451, 0.910 and 0.740 and in the IV
extrusion 0.421, 0.422, 0.945 and 0.431 in the subsequent
intervals respectively. All the sperm counts are present ed in
thousands. Higher counts were recorded in the right halve of
spermatophore of shrimps administered with 0.1 µl. Among all the L. acidophilus treated broodstock s hrimps, only 0.1 µl/g
of body weight performed better after 7 days of
administration to t he shrimps. Invigoration of male for
ensuring effective fertilization and thereby p erfect hatching should also be attempted. Application of L. acidophilus in
enhancing sperm quality assay from this investigation would
immensely go to a longer way in maximizing the production
of shrimp seeds in hatchery facility which exists both east and
west coast of India.
ACKNOWLEDG MENT
The authors are tha nkful to the authorities of Annamalai
University, for providing t he facilities and support. Many
sincere t hanks to the fis hermen society for p roviding live
animals and their kind moral support towards the completion
of our research work.
REFERENCES [1] FAO; 2000. State of World Fisheries and Aquaculture. ISBN:
9251044929, X8002/E.
[2] Weidner D. and Rosenberry B. (1992) World Shrimp Farming. In: Wyban J. (Ed) Proceedings of the special session
on shrimp farming (May, 1992, Orlando, Florida, USA). World Aquaculture Society, Baton Rouge, Louisiana, USA pp.
1-21.
[3] Hedgecock, D., Malecha, S.R., 1991. Prospects for the
application of biotechnology to the development and improvement of shrimp and prawns. In: Sandifer, P.A. _Ed.., Shrimp Culture in North America and the Caribbean. World Aquaculture Society, Baton Rouge, LA, USA, pp. 161–200.
[4] Quackenbush, S.L., 1986. Yolk Synthesis in the Marine
Shrimp, Penaeus vannamei. Integrative and Comparative Biology.
41(3) - 458-464.
[5] Emmerson, W. D., 1980. Induced maturation of prawn Penaeus indicus. Mar. Ecol. Prog. Ser., 2: 121 – 131.
[6] Tsukimura, B., and F. I. Kamemoto. 1991. In vitro
stimulation of oocytes by presumptive mandibular organ secretions in the shrimp, Penaeus vannamei. Aquaculture, 9259-66.

M. A. Badhul Haq, et al.: Efficacy of Lactobacillus acidophilus on the sperm quality and regeneration of spermatophores in Penaeus monodon
(Fabricius, 1798)
September, 2011 International Journal of Chemical and Analytical Science, 2011, 2(9), 1178-1185 1185
[7] Benzie, J.A.H., 1995. Genetics in the domestication of the giant tiger prawn Penaeus monodon in Australia. Book of
Abstracts, Aquaculture ‘95. World Aquaculture Society, Baton
Rouge, LA, USA, p. 61.
[8] Boschi, E.E, 1979b. Geographic distribution of Argentinean marine decapod crustaceans. Bull. Biol. Soc. Wash. 3: 134 –
143.
[9] García, S. and L Le Reste. 1981. Life cycles, dynamics,
exploitation and management of coastal penaeids shrimp stocks. FAO Fisheries Technical Paper 203: 1-215.
[10] Leung-Trujillo, J.R. and A.L. Lawrence, 1987. Observations
on the decline in sperm quality of Penaeus setiferus under laboratory conditions. Aquaculture, 65:363 -370.
[11] Chamberlain, G.W., 1988. Stepwise investigation of
environmental and nutritional requirements for reproduction of penaeid shrimp. Ph.D. Thesis, Department
of Wildlife and Fisheries Sciences, Texas A & M University, USA, 45-63 pp.
[12] Alfaro, J., 1993. Reproductive quality evaluation of male
Penaeus stylirostris from a grow-out pond. J. World Aquacult.
Soc., 24: 6-11.
[13] Leung-Trujillo, J.R. and A.L. Lawrence, 1991. Spermatophore
generation times in Penaeus setiferus, P. vannamei, and P.
stylirostris. J. World Aquacult. Soc., 22 : 244 -251.
[14] Bray, W.A., Lawrence, A.L., Lester, L.J., Smith, L.L., 1990.
Hybridization of Penaeus setiferus (Linnaeus, 1767) and
Penaeus schmitti Burkenroad, 1936 (Decapoda). J. Crustac. Biol. 10 (2): 278–283.
[15] Talbot, P., D. Howard, D. Leung-trujillo, T.W. Lee, W.Y. Li,
H. Ro and A. L. Lawrence, 1989. Characterization of male reproductive-tract degenerative syndrome in captive penaeid shrimp (Penaeus-setiferus). Aquaculture ,78:365 -377.
[16] Griffin, F.J., C.A. Vines, M.C. Pillai, R Yanagimachi and G.N. Cherr, 1996. Sperm motility initiation factor is a minor component of the Pacific herring egg chorion. Development Growth and Differentiation, 38: 193–202.
[17] Pratoomchat, B., S.Piyatiratitivorakul, P. Menasveta and
A.W. Fast, 1993. Sperm quality of pond-reared and wild-caught Penaeus monodon in Thailand. J. World Aquacult. Soc.,
24: 530 -540.
[18] Wang, Q., Misamore, M., Jiang, C., Browdy, C.L., 1995. Egg
water induced reaction and biostain assay of sperm from marine shrimp, Penaeus vannamei: dietary effects on sperm quality. J. World Aquacult. Soc. 26, 261–271.
[19] Fingerman, S. W., and M. Fingerman. 1977. Effects of a
polychlorinated biphenyl and a polychlorinated
dibenzofuran on molting of the fiddler crab, Uca pugilator. Bull. Environ. Contam. Toxicol, 18138-142.
[20] Sarojini, R, R. Nagabhushanam, M. Devi and M. Fingerman
1995.Dopaminergic inhibition of Shydroxy-tryptaminestimulated testicular maturation in the fiddler crab, Uca pugilator. Camp. Biochem. Physiol. Vol. 111C. (2) 287-
292.
[21] Ziaei-Nejad S., G.A. Rezaei Takami, D.L. Lovett,
A.R.Mirvaghefi and M. Shakouri, 2006. The effect of Bacillus
spp. bacteria used as probiotics on digestive enzyme activity,
survival and growth in the Indian white shrimp Fenneropenaeus indicus. Aquaculture, 252: 516–524.
[22] Nimrat. S, A. N. Bart, A. Keatsaksit, V. Vuthiphandchai,
2008. Microbial flora of spermatophores from black tiger shrimp (Penaeus monodon) declines over long-term cryostorage. Aquaculture, 274:247–253.
[23] Laufer H. 1992. Method for increasing crustacean larval production. United States Patent 5, 161, 481.
[24] Sandifer P. A., A. I.awrence, S. Harris, G. H. Chamberlain, A.
D. Stokes, W. A. IIray, 1984. Electrical stimulation of spermatophore expulsion in marine shrimp, Prnuc,u.s spp. Aquaculture, 41, 18 1 - 187.
[25] Ajitha, S., M. Sridhar, N. Sridhar, I.S.B. Singh and V. Varghese, 2004. Probiotic Effects of Lactic Acid Bacteria Against Vibrio Alginolyticus in Penaeus (Fenneropenaeus) Indicus (H. Milne Edwards), Asian Fisheries Science 17:71-80.
[26] Gomez L.A. & Honculada-Primavera J.H. (1993)
Reproductive quality of male Penaeus monodon. Aquaculture, 112, 157 - 164.
[27] Ceballos-Vazquez, B.P., C. Rosas and I. S. Racotta, 2003.
Sperm quality in relation to age and weight of white shrimp Litopenaeus vannamei. Aquaculture, 28:141 -151.
[28] Ceballos-Vazquez, B.P., B. Aparicio-Simon, E. Palacios, and
I. S. Racotta, (2004). Sperm quality over consecutive spermatophore regenerations in the Pacific white shrimp Litopenaeus vannamei. J. World Aquacult. Soc., 35:178 -188.
[29] Meunpol O, Sandeman R, Sandeman D, Kapoor V. Stimulus-
coupled serotonin release from crayfish accessory lobes (C. destructor). Australian Soc Comp Physiol; Abstract, 1998
meeting; Melbourne, Australia. 1998.
[30] Hoang,T.,Lee, S.Y.,Keenan, C.P.,Marsden,G.E.,2002. Cold tolerance of the banana prawn, Penaeus merguiensis and its growth at different temperatures. Aquacult. Res. 33, 21–26.
Source of support: Nil, Conflict of interest: None Declared