electrophoresis of enzyme-monoclonal antibody complexes: studies
TRANSCRIPT

Proc. NatL Acad. Sci. USAVol. 78, No. 8, pp. 5061-5065, August 1981Genetics
Electrophoresis of enzyme-monoclonal antibody complexes:Studies of human placental alkaline phosphatase polymorphism
(hybridomas/isozymes/sialic acid)
KATHRYN J. GOGOLIN, CLIVE A. SLAUGHTER, AND HARRY HARRISDepartment of Human Genetics, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania 19104
Contributed by Harry Harris, May 4, 1981
ABSTRACT Enzyme-monoclonal antibody complexes formedbetween six different monoclonal antibodies and the six pheno-types of human placental alkaline phosphatase [orthophosphoric-monoester phosphohydrolase (alkaline optimum), EC 3.1.3.1] thatrepresent the homozygous and heterozygous combinations of thethree common alleles have been examined by electrophoresis instarch, acrylamide, and agarose gels. Since the complexes formedretain full enzyme activity, they could be detected after gel elec-trophoresis by an enzyme stain. Distinctive electrophoretic pat-terns were obtained with each monoclonal antibody. Differentialbinding of certain of the antibodies with the products of differentalleles produces clear discrimination of various homozygous andheterozygous phenotypes. This discrimination parallels the resultspreviously obtained by using a quantitative binding radioimmu-noassay. The results show that this general method should proveuseful in screening hybridoma fluids for the presence of mono-clonal antibodies to specific enzymes; in the detection of allelicvariation, even where this is not expressed by electrophoretic dif-ferences among the uncomplexed enzymes; and in discriminatingbetween homozygotes and heterozygotes. It could also prove to bea useful tool in the elucidation of the molecular structures of en-zyme-monoclonal antibody complexes.
Human placental alkaline phosphatase [ALPase, orthophos-phoric-monoester phosphohydrolase (alkaline optimum), EC3.1.3.1] is a highly polymorphic enzyme. Three common allelesgiving rise to six common electrophoretic phenotypes occur inmost human populations, and a considerable number of rarealleles have also been identified (1-3). We recently reporteda study in which a large panel of placental ALPases of variousphenotypes was tested against six different monoclonal anti-bodies raised against purified placental ALPase by the mousehybridoma technique (4). Relative binding of each of the AL-Pases to each of the monoclonal antibodies was determinedquantitatively by radioimmunoassay under standardized con-ditions. It was shown that the products of the three commonalleles could be differentiated by their reactivities with the dif-ferent antibodies. In this paper, we report the results of elec-trophoretic studies on the ALPase-monoclonal antibody com-plexes formed between the same six antibodies and each of thecommon placental ALPase polymorphic types. The studieswere made possible by the findings that, with these antibodies,the enzyme-monoclonal antibody complexes retain full enzymeactivity and that the complexes are soluble so that they can besubjected to gel electrophoresis. The findings confirm the re-sults of the quantitative binding assays by showing that essen-tially the same discrimination among the six common pheno-types is also achieved by electrophoresis of enzyme-antibodycomplexes formed with the various antibodies. They also dem-onstrate the potential power of the general method for studiesin enzyme genetics. In addition, certain unexpected featuresof the electrophoretic characteristics of the enzyme-antibodycomplexes have been identified.
MATERIALS AND METHODSHybridoma culture fluids and ascites fluids containing mono-clonal antibodies to purified placental ALPase were producedas described (4-6). They were the same as those used in ourearlier report (4) and will be referred to as P3/1, Sp2/2, Sp2/3, etc., indicating the parental plasmacytoma cell line P3 (P3/X63-Ag8) or Sp2 (Sp2/0-Agl4) and the serial number of thepositive hybridoma. Placental extracts were prepared, assayed,typed electrophoretically, and, where required, treated withneuraminidase as described (7, 8). ALPase-antibody complexeswere prepared by using ALPase extract diluted to 0.1 inter-national unit/ml with Tris-HCl, pH 7.4/0.15 M NaCl/0.02%sodium azide/0.05% Nonidet P-40 (British Drug House, Poole,England)/0.25% gelatin. One hundred microliters of ALPasewas incubated overnight at 50C with 20 1,u ofhybridoma culturefluid or ascites fluid.
Horizontal starch gel electrophoresis using 13% Electro-starch (Electrostarch, Madison, WI) was carried out as de-scribed (8). The gel buffer was 76 mM Tris citrate, pH 8.6, andthe bridge buffer was 0.3 M boric acid brought to pH 8.0 withNaOH. Electrophoresis was carried out for 6 hr at 7 V/cm atroom temperature. Fifty-microliter samples were inserted intothe gel by using saturated Whatman 17 paper strips. Verticalacrylamide slab gel electrophoresis was performed on 5% acryl-amide gels in 0.37 M Tris, pH 8.8/0.13% bis(N,N'-methylene-bisacrylamide)/0. 04% NN, N' ,N' -tetramethylethylene-diamine (Bio-Rad)/0.06% ammonium persulfate. The bridgebuffer was 0.1 M Tris/0. 1 M boric acid, pH 8.3. Electrophoresiswas for 16 hr at a constant 8-mA current. Horizontal agarose gelelectrophoresis was carried out in 1% agarose gels (Isogel,FMC, Rockland, ME) using 76 mM Tris/citrate, pH 8.6. Elec-trophoresis was for 5 hr at room temperature at a constant 25-mA current across the gel.The various gels were stained for ALPase by layering the
horizontal cut surface with Whatman 3M filter paper saturatedwith 1.2 mM 4-methylumbelliferyl phosphate (Sigma) in 0.2 Mborate buffer, pH 9.7. The fluorescent bands of free umbelli-ferone were viewed under UV light.
RESULTSStarch Gel Electrophoresis. Placental extracts subjected to
starch gel electrophoresis show two sets of ALPase isozymes.One set, referred to as the A isozymes, migrates relatively rap-idly toward the anode and represents the dimeric forms of theenzyme (1, 2, 9). The other set, referred to as the B isozymes,migrates relatively slowly, remains close to the origin, and rep-resents higher molecular weight forms that appear to be aggre-gates of the dimeric forms possibly associated with some as yetunidentified ligands derived from cellular membranes (9). The
Abbreviation: ALPase, alkaline phosphatase.
5061
Proc. Natl Acad. Sci. USA 78 (1981)
various polymorphic phenotypes and rare variants are routinelyidentified by the number and relative mobilities of the A iso-zymes. These phenotype differences cannot usually be recog-nized in the B isozymes, although they have been demonstratedafter prolonged electrophoresis at elevated temperatures (2).
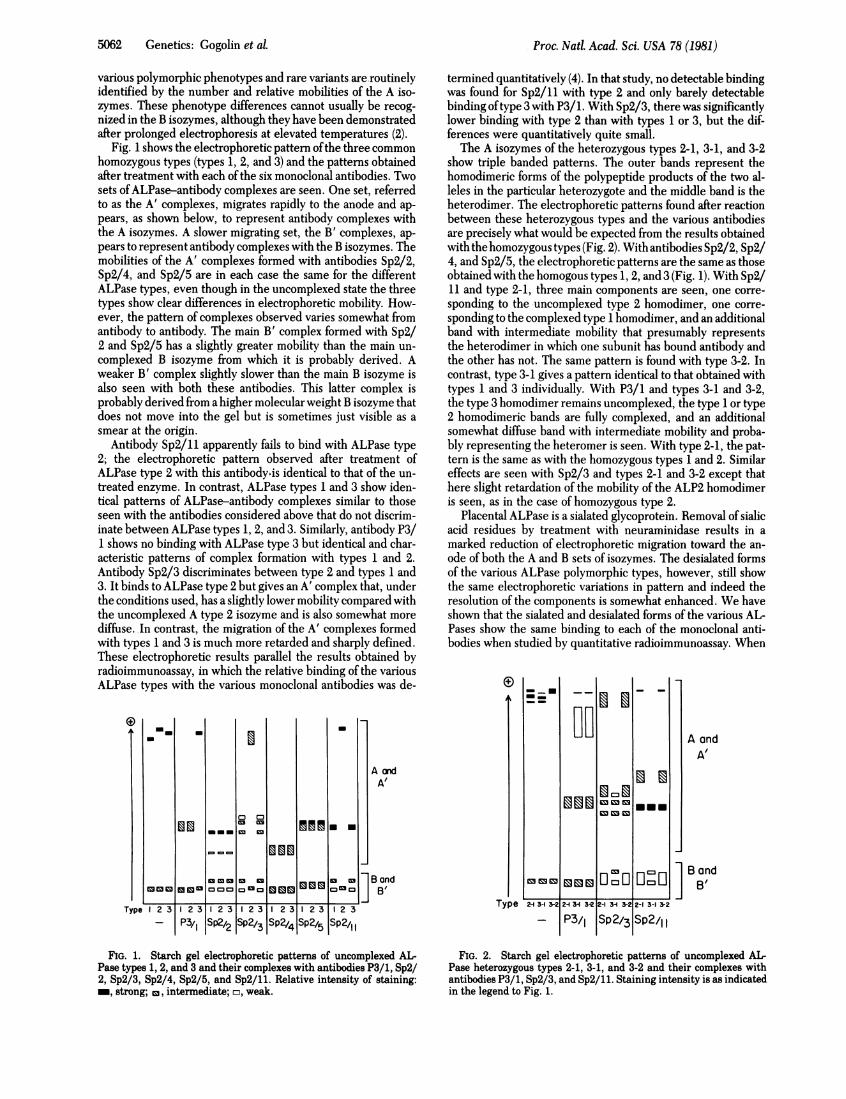
Fig. 1 shows the electrophoretic pattern ofthe three commonhomozygous types (types 1, 2, and 3) and the patterns obtainedafter treatment with each ofthe six monoclonal antibodies. Twosets ofALPase-antibody complexes are seen. One set, referredto as the A' complexes, migrates rapidly to the anode and ap-pears, as shown below, to represent antibody complexes withthe A isozymes. A slower migrating set, the B' complexes, ap-pears to represent antibody complexes with the B isozymes. Themobilities of the A' complexes formed with antibodies Sp2/2,Sp2/4, and Sp2/5 are in each case the same for the differentALPase types, even though in the uncomplexed state the threetypes show clear differences in electrophoretic mobility. How-ever, the pattern of complexes observed varies somewhat fromantibody to antibody. The main B' complex formed with Sp2/2 and Sp2/5 has a slightly greater mobility than the main un-complexed B isozyme from which it is probably derived. Aweaker B' complex slightly slower than the main B isozyme isalso seen with both these antibodies. This latter complex isprobably derived from a higher molecular weight B isozyme thatdoes not move into the gel but is sometimes just visible as asmear at the origin.
Antibody Sp2/11 apparently fails to bind with ALPase type2; the electrophoretic pattern observed after treatment ofALPase type 2 with this antibody is identical to that of the un-treated enzyme. In contrast, ALPase types 1 and 3 show iden-tical patterns of ALPase-antibody complexes similar to thoseseen with the antibodies considered above that do not discrim-inate between ALPase types 1, 2, and 3. Similarly, antibody P3/1 shows no binding with ALPase type 3 but identical and char-acteristic patterns of complex formation with types 1 and 2.Antibody Sp2/3 discriminates between type 2 and types 1 and3. It binds to ALPase type 2 but gives an A' complex that, underthe conditions used, has a slightly lower mobility compared withthe uncomplexed A type 2 isozyme and is also somewhat morediffuse. In contrast, the migration of the A' complexes formedwith types 1 and 3 is much more retarded and sharply defined.These electrophoretic results parallel the results obtained byradioimmunoassay, in which the relative binding of the variousALPase types with the various monoclonal antibodies was de-
Type 1 2 3
.mm
o Oas ,en IMMM. .
1 23 1 23 23 1 23 1 23 123
P3/1 Sp2/2 Sp2/3 Sp2/4 Sp2/5 Sp2/11
termined quantitatively (4). In that study, no detectable bindingwas found for Sp2/11 with type 2 and only barely detectablebinding oftype 3 with P3/1. With Sp2/3, there was significantlylower binding with type 2 than with types 1 or 3, but the dif-ferences were quantitatively quite small.
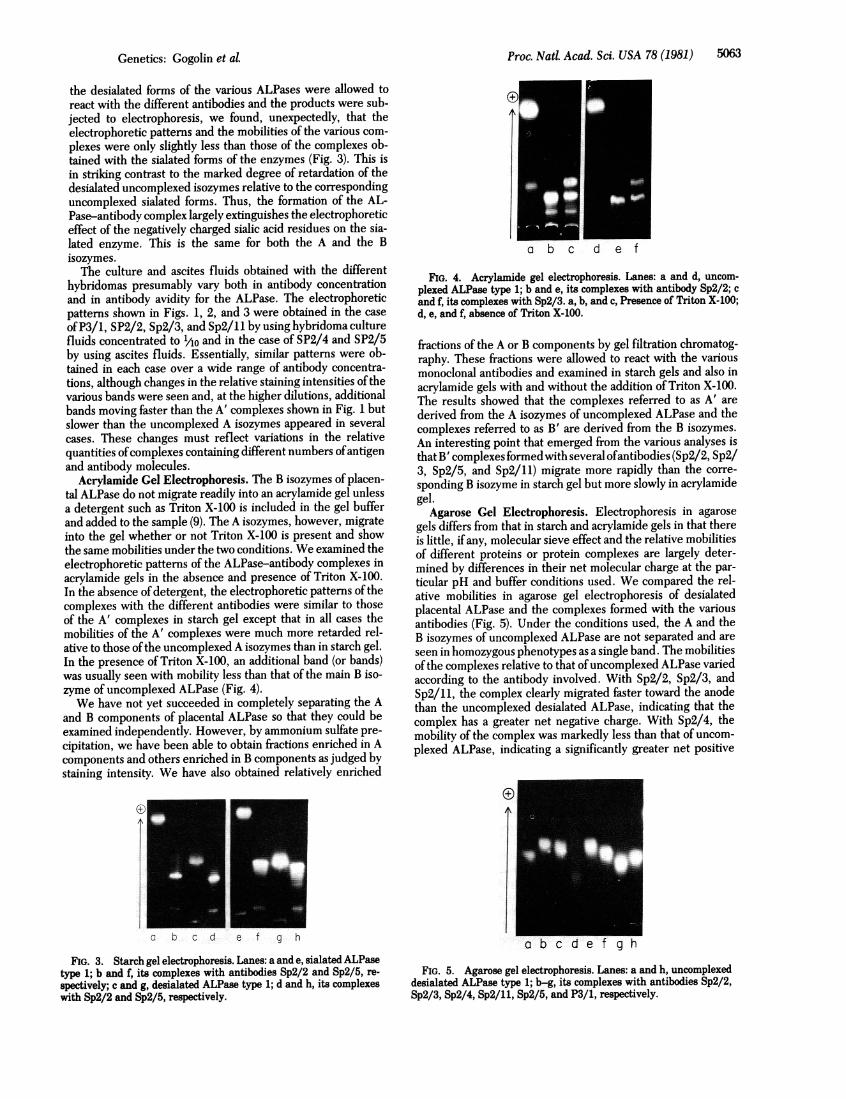
The A isozymes of the heterozygous types 2-1, 3-1, and 3-2show triple banded patterns. The outer bands represent thehomodimeric forms of the polypeptide products of the two al-leles in the particular heterozygote and the middle band is theheterodimer. The electrophoretic patterns found after reactionbetween these heterozygous types and the various antibodiesare precisely what would be expected from the results obtainedwith the homozygous types (Fig. 2). With antibodies Sp2/2, Sp2/4, and Sp2/5, the electrophoretic patterns are the same as thoseobtained with the homogous types 1, 2, and 3 (Fig. 1). With Sp2/11 and type 2-1, three main components are seen, one corre-sponding to the uncomplexed type 2 homodimer, one corre-sponding to the complexed type 1 homodimer, and an additionalband with intermediate mobility that presumably representsthe heterodimer in which one subunit has bound antibody andthe other has not. The same pattern is found with type 3-2. Incontrast, type 3-1 gives a pattern identical to that obtained withtypes 1 and 3 individually. With P3/1 and types 3-1 and 3-2,the type 3 homodimer remains uncomplexed, the type 1 or type2 homodimeric bands are fully complexed, and an additionalsomewhat diffuse band with intermediate mobility and proba-bly representing the heteromer is seen. With type 2-1, the pat-tern is the same as with the homozygous types 1 and 2. Similareffects are seen with Sp2/3 and types 2-1 and 3-2 except thathere slight retardation of the mobility of the ALP2 homodimeris seen, as in the case of homozygous type 2.
Placental ALPase is a sialated glycoprotein. Removal of sialicacid residues by treatment with neuraminidase results in amarked reduction of electrophoretic migration toward the an-ode of both the A and B sets of isozymes. The desialated formsof the various ALPase polymorphic types, however, still showthe same electrophoretic variations in pattern and indeed theresolution of the components is somewhat enhanced. We haveshown that the sialated and desialated forms of the various AL-Pases show the same binding to each of the monoclonal anti-bodies when studied by quantitative radioimmunoassay. When
A andA'
B andB'
FIG. 1. Starch gel electrophoretic patterns of uncomplexed AL-Pase types 1, 2, and 3 and their complexes with antibodies P3/1, Sp2/2, Sp2/3, Sp2/4, Sp2/5, and Sp2/11. Relative intensity of staining:_, strong; c, intermediate; m, weak.
Type
MEN.
u~u0 :
A andA'
BandB'
2A 3-1 3-2 2-I 31 3-2 2-1 3-I 3-2 2-1 3-1 3-2
- P3/1 Sp2/3 Sp2/11
FIG. 2. Starch gel electrophoretic patterns of uncomplexed AL-Pase heterozygous types 2-1, 3-1, and 3-2 and their complexes withantibodies P3/1, Sp2/3, and Sp2/11. Staining intensity is as indicatedin the legend to Fig. 1.
55-0162 Genetics: Gogolin et al.
__2~--
Proc. NatL Acad. Sci. USA 78 (1981) 5063
the desialated forms of the various ALPases were allowed toreact with the different antibodies and the products were sub-jected to electrophoresis, we found, unexpectedly, that theelectrophoretic patterns and the mobilities of the various com-plexes were only slightly less than those of the complexes ob-tained with the sialated forms of the enzymes (Fig. 3). This isin striking contrast to the marked degree of retardation of thedesialated uncomplexed isozymes relative to the correspondinguncomplexed sialated forms. Thus, the formation of the AL-Pase-antibody complex largely extinguishes the electrophoreticeffect of the negatively charged sialic acid residues on the sia-lated enzyme. This is the same for both the A and the Bisozymes.The culture and ascites fluids obtained with the different
hybridomas presumably vary both in antibody concentrationand in antibody avidity for the ALPase. The electrophoreticpatterns shown in Figs. 1, 2, and 3 were obtained in the caseofP3/1, SP2/2, Sp2/3, and Sp2/11 by using hybridoma culturefluids concentrated to '/Ao and in the case of SP2/4 and SP2/5by using ascites fluids. Essentially, similar patterns were ob-tained in each case over a wide range of antibody concentra-tions, although changes in the relative staining intensities ofthevarious bands were seen and, at the higher dilutions, additionalbands moving faster than the A' complexes shown in Fig. 1 butslower than the uncomplexed A isozymes appeared in severalcases. These changes must reflect variations in the relativequantities ofcomplexes containing different numbers ofantigenand antibody molecules.
Acrylamide Gel Electrophoresis. The B isozymes of placen-tal ALPase do not migrate readily into an acrylamide gel unlessa detergent such as Triton X-100 is included in the gel bufferand added to the sample (9). The A isozymes, however, migrateinto the gel whether or not Triton X-100 is present and showthe same mobilities under the two conditions. We examined theelectrophoretic patterns of the ALPase-antibody complexes inacrylamide gels in the absence and presence of Triton X-100.In the absence ofdetergent, the electrophoretic patterns of thecomplexes with the different antibodies were similar to thoseof the A' complexes in starch gel except that in all cases themobilities of the A' complexes were much more retarded rel-ative to those ofthe uncomplexed A isozymes than in starch gel.In the presence of Triton X-100, an additional band (or bands)was usually seen with mobility less than that of the main B iso-zyme of uncomplexed ALPase (Fig. 4).We have not yet succeeded in completely separating the A
and B components of placental ALPase so that they could beexamined independently. However, by ammonium sulfate pre-cipitation, we have been able to obtain fractions enriched in Acomponents and others enriched in B components as judged bystaining intensity. We have also obtained relatively enriched
c b c d e f g h
FIG. 3. Starch gel electrophoresis. Lanes: a and e, sialated ALPasetype 1; b and f, its complexes with antibodies Sp2/2 and Sp2/5, re-spectively; c and g, desialated ALPase type 1; d and h, its complexeswith Sp2/2 and Sp2/5, respectively.
FIG. 4. Acrylamide gel electrophoresis. Lanes: a and d, uncom-plexed ALPase type 1; b and e, its complexes with antibody Sp2/2; cand f, its complexes with Sp2/3. a, b, and c, Presence of Triton X-100;d, e, and f, absence of Triton X-100.
fractions of the A or B components by gel filtration chromatog-raphy. These fractions were allowed to react with the variousmonoclonal antibodies and examined in starch gels and also inacrylamide gels with and without the addition of Triton X-100.The results showed that the complexes referred to as A' arederived from the A isozymes of uncomplexed ALPase and thecomplexes referred to as B' are derived from the B isozymes.An interesting point that emerged from the various analyses isthat B' complexes formed with several ofantibodies (Sp2/2, Sp2/3, Sp2/5, and Sp2/11) migrate more rapidly than the corre-sponding B isozyme in starch gel but more slowly in acrylamidegel.
Agarose Gel Electrophoresis. Electrophoresis in agarosegels differs from that in starch and acrylamide gels in that thereis little, if any, molecular sieve effect and the relative mobilitiesof different proteins or protein complexes are largely deter-mined by differences in their net molecular charge at the par-ticular pH and buffer conditions used. We compared the rel-ative mobilities in agarose gel electrophoresis of desialatedplacental ALPase and the complexes formed with the variousantibodies (Fig. 5). Under the conditions used, the A and theB isozymes of uncomplexed ALPase are not separated and areseen in homozygous phenotypes as a single band. The mobilitiesofthe complexes relative to that ofuncomplexed ALPase variedaccording to the antibody involved. With Sp2/2, Sp2/3, andSp2/11, the complex clearly migrated faster toward the anodethan the uncomplexed desialated ALPase, indicating that thecomplex has a greater net negative charge. With Sp2/4, themobility of the complex was markedly less than that of uncom-plexed ALPase, indicating a significantly greater net positive
a b c d e f g h
FIG. 5. Agarose gel electrophoresis. Lanes: a and h, uncomplexeddesialated ALPase type 1; b-g, its complexes with antibodies Sp2/2,Sp2/3, Sp2/4, Sp2/11, Sp2/5, and P3/1, respectively.
Genetics: Gogolin et aL

Proc. Natl. Acad. Sci. USA 78 (1981)
charge. With SP2/5, the complex migrated slightly faster, andwith P3/1, the complex migrated slightly slower than the un-complexed ALPase. Similar experiments were carried out withsialated ALPase. In each case, the complex migrated slightlyfaster than the corresponding complex with the desialated en-zyme, but the difference in mobility between the two forms wasmuch less than the difference in mobility between the sialatedand desialated uncomplexed ALPase.
DISCUSSIONThese results show that electrophoresis of enzyme-monoclonalantibody complexes is potentially a powerful tool for studies inenzyme genetics. The finding that the electrophoretic patternsof the enzyme-antibody complexes are clearly different fromthose of the uncomplexed enzyme indicates that the methodcould be useful in screening hybridoma fluids for monoclonalantibodies to an enzyme under investigation. The results alsoshow that it is practical to discriminate among different allelicforms of an enzyme by this approach. We have used placentalALPase in this work because it shows extensive electrophoreticpolymorphism and thus provides a suitable test system to seewhether different monoclonal antibodies can discriminateamong the enzyme products of different alleles in homozygousand heterozygous states. The demonstration that such discrim-ination occurs indicates that the general method could also beused to screen individuals for allelic variation of an enzyme thatdoes not necessarily show electrophoretic polymorphism in theuncomplexed state. The method can also be applied to knownenzyme polymorphisms in the search for further allelic variationamong individuals with the same electrophoretic phenotype.Since it can detect heterozygotes, it can also be applied to thesearch for rare alleles that, as a rule, will occur with appreciablefrequency only in the heterozygous state. The electrophoreticmethod is clearly sensitive; it has detected the main differencespreviously identified by quantitative determinations of relativeantibody binding using radioimmunoassay. It should, however,be noted that we have not yet been able to differentiate by thisapproach certain rather subtle allelic differences that were iden-tified statistically in the quantitative binding data-e.g., thevery small differences between types 1, 2-1, and 2 detected byantibody P3/1 and small differences within the ALPase type 1phenotype detected by Sp2/4 (4).
Certain of the electrophoretic properties of the various AL-Pase-antibody complexes found in this study were unexpected.Perhaps the most surprising result emerged from comparisonof the antibody complexes of the sialated and desialated formsof the enzymes. In the absence of antibody, the sialated forms(both A and B isozymes) have a markedly greater anodal mo-bility than the desialated forms, presumably because of the ex-posed negatively charged sialic acid residues in the carbohy-drate moiety of the glycoproteins. These sialic acid residues donot appear to form part of the various antigenic determinantsto which the different monoclonal antibodies used here are di-rected; we have found no significant differences between thebinding of the antibodies to the sialated and desialated formsof the enzyme using our quantitative radioimmunoassay pro-cedure. Nevertheless, the ALPase antibody complexes ob-tained with the sialated and desialated enzymes show only slightdifferences in electrophoretic mobility. This is true for both theA and B isozymes in the same placental extract, in samples fromdifferent ALPase phenotypes with each of the monoclonal an-tibodies, and in each of the starch, acrylamide, and agarose gelsystems. It appears that the sialic acid residues normally ex-posed on the surface of the enzyme molecule are buried in theenzyme-antibody complex in such a way that their negativecharges are largely neutralized. It will be of interest to see
whether the same effect occurs when other sialoglycoproteinsare investigated in this way.The other unexpected result concerns the relative electro-
phoretic mobilities of the A isozymes of the three common ho-mozygous types 1, 2, and 3 in the uncomplexed and complexedstates. The characteristic mobility differences among the threetypes in the uncomplexed state are presumed to be due to aminoacid substitutions derived from the mutations by which the var-ious alleles originated. Two of the antibodies, Sp2/2 and Sp2/5, did not discriminate between the electrophoretic types in thequantitative radioimmunoassay binding study (4). This suggeststhat these antibodies, unlike the other antibodies studied, arenot directed to antigenic determinants that involve those sitesin the enzyme structures at which the mutational differencesoccur. Nevertheless, no electrophoretic differences betweenthe ALPase-antibody complexes of the three phenotypes witheither of these antibodies were detected. The amino acid dif-ferences from which the electrophoretic differences in the un-complexed state derive are apparently in these cases buried inthe enzyme-antibody complex.The electrophoretic mobilities of the various AL-
Pase-monoclonal antibody complexes in the different gelsystems may be influenced by at least three different generaltypes of effect (10). The first is the effect of the net charge ofthe complex at the particular pH used. This will depend inter-alia on the relative numbers of enzyme and antibody moleculesin the particular complex, on the charge characteristics of theenzyme and the immunoglobulin in the complex, and on theconfiguration ofthe enzyme and antibody molecules in the com-plex. The second is the effect of molecular sieving, which tendsto retard migration increasingly with increasing molecular size.The degree of molecular sieving is determined by the matrixstructure of the different types of gels-starch, acrylamide, oragarose-and by the gel concentration. Third is the effect ofpossible adsorption to the gel matrix. This effect probably ac-counts for the failure of the B isozymes and the B' complex tomigrate readily into acrylamide gels in the absence of detergent(9).
Both starch and acrylamide gels exert marked sieving effects.But sieving probably plays little or no part in agarose gel elec-trophoretic separations. Thus, with the same ALPase, differ-ences in electrophoretic mobility in agarose gel electrophoresisof complexes with different antibodies will be largely derivedfrom the charge characteristics of the antibody and its inter-action with the ALPase in the complex. From the results weobtained with agarose gel electrophoresis, we can infer, for ex-ample, that desialated ALPase complexes with antibodies Sp2/2, Sp2/3, and Sp2/11 have a greater net negative charge thanthose with the other antibodies and also a greater net negativecharge than uncomplexed desialated ALPase.
Both charge and size are important in determining electro-phoretic mobilities in starch and acrylamide gels. Since thedegree of retardation of the ALPase-antibody complex is muchmore marked in the acrylamide system than in the starch sys-tem, it appears that the effect of molecular size relative to netcharge is greater in the acrylamide system than in the starchsystem, even at the relatively low acrylamide concentration(5%) used here. This may account for the apparently anomalousfinding that complexes of antibodies Sp2/2, Sp2/3, and Sp2/11 with B isozymes migrate more rapidly than the correspond-ing uncomplexed isozymes in the starch system but are retardedrelative to the uncomplexed isozymes in the acrylamide system.The results from agarose gel electrophoresis indicate that com-plexes with these three antibodies have greater net negativecharges than uncomplexed ALPase. With the B isozymes, thisincrease in negative charge apparently overrides the sieving
5064 Genetics: Gogolin et al.

Genetics: Gogolin et al.
effect in the starch gel system but is overridden by the sievingeffect in the acrylamide gel system.
In general, one may expect that the use of electrophoresisin these various gel systems will prove to be a valuable tool inthe elucidation of the molecular structure of enzyme-monoclonalantibody complexes.
This work was supported by Grants GM 27018 and GM 20138 fromthe National Institutes of Health.
1. Robson, E. B. & Harris, H. (1965) Nature (London) 207,1257-1259.
2. Robson, E. B. & Harris, H. (1967) Ann. Hum. Genet. 30,219-232.
3. Donald, L. J. & Robson, E. B. (1974) Ann. Hum. Genet. 37,303-313.
Proc. Nat. Acad. Sci. USA 78 (1981) 5065
4. Slaughter, C. A., Coseo, M. C., Cancro, M. P. & Harris, H.(1981) Proc. Natl Acad. Sci. USA 78. 1124-1128.
5. Slaughter, C. A., Coseo, M., Abrams, C., Cancro, M. P. & Har-ris, H. (1980) in Monoclonal Antibodies, eds. Kennett, R. H.,McKearn, T. J. & Bechtol, K. B. (Plenum, New York), pp.103-120.
6. Slaughter, C. A., Coseo, M., Abrams, C., Cancro, M. P. & Har-ris, H. (1980) in Monoclonal Antibodies, eds. Kennett, R. H.,McKearn, T. J. & Bechtol, K. B. (Plenum, New York), pp.383-387.
7. Mulivor, R. A., Plotkin, L. I. & Harris, H. (1978) Ann. Hum.Genet. 42, 1-13.
8. Harris, H. & Hopkinson, D. A. (1976) Handbook of EnzymeElectrophoresis in Human Genetics (North-Holland, Amster-dam), Chap. 4, 3.1.3.1, pp. 1-4.
9. Doellgast, G. J., Spiegel, J., Guenther, R. A. & Fishman, W. H.(1977) Biochim. Biophys. Acta 484, 59-78.
10. Morris, C. J. 0. R. & Morris, P. (1976) Separation Methods inBiochemistry (Wiley, New York).