endo-p-1,4-glucanase gene of bacillus subtilis dlg - journal of
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, May 1987, p. 2017-2025 Vol. 169, No. 50021-9193/87/052017-09$02.00/0Copyright © 1987, American Society for Microbiology
Endo-P-1,4-Glucanase Gene of Bacillus subtilis DLGLORI M. ROBSONt AND GLENN H. CHAMBLISS*
Department ofBacteriology, University of Wisconsin, Madison, Wisconsin 53706
Received 10 November 1986/Accepted 9 February 1987
The DNA sequence of the BaciUus subtlis DLG endo-,B-1,4-glucanase gene was determined, and the in vivosite of transcription initiation was located. Immediately upstream from the transcription start site weresequences closely resembling those recognized by B. subtilis &43-RNA polymerase. Two possible ribosome-binding sites were observed downstream from the transcription start site. These were followed by a long openreading frame capable of encoding a protein of ca. 55,000 daltons. A signal sequence, typical of those presentin gram-positive organisms, was observed at the amino terminus of the open reading frame. Purification of themature exocellular 0-1,4-glucanase and subsequent amino-terminal protein sequencing defined the site ofsignal sequence processing to be between two alanine residues following the hydrophobic portion of the signalsequence. The probability of additional carboxy-terminal processing of the 0-1,4-glucanase precursor isdiscussed. Si nuclease protection studies showed that the amount of ,B-1,4-glucanase mRNA in cells increasedsignificantly as the culture entered the stationary phase. In addition, glucose was found to dramaticallystimulate the amount of ,B-1,4-glucanase mRNA in vivo. Finally, the specffic activities of purified B. subtilisDLG endo-0-1,4-glucanase and Trichoderma reesei QM9414 endo-f-1,4-glucanase (EC 3.2.1.4) were comparedby using the noncrystalline cellulosic substrate trinitrophenyl-carboxymethyl cellulose.
Many members of the family Bacillaceae produce extra-cellular ,B-glucanases. The linkage specificities of these en-zymes are varied, the most common being ,-1,3-1,4-glucanases and 13-1,3-glucanases (6, 20, 32, 41, 57). There arerelatively few reports of members of the family Bacillaceaeproducing ,B-1,4-glucanases (13, 21, 41, 44). The two rela-tively well characterized examples, the alkalophilic Bacillusstrain N-4 (21, 48) and Bacillus subtilis DLG (44, 45),produce cellulolytic activity in the form of a carboxymethylcellulase (CMC) (i.e., P-1,4-glucanase). Although incapableof degrading crystalline forms of cellulose individually,bacterial P-1,4-glucanases (most likely endo acting) may beable to act synergistically with cellulases of other specifici-ties, such as exo-acting P-1,4-glucanases or P-glucosidases,or both, to achieve the enzymatic bioconversion of celluloseto more commercially useful products. A Bacillus P-1,4-glucanase may also have a role to play in the brewingindustry (5, 11, 20).For B. subtilis DLG, ,B-1,4-glucanase production begins at
the onset of the stationary phase and is not repressed byeither glucose or cellobiose (44), as are many othercellulolytic systems (12, 30, 53, 55). In fact, glucose stimu-lates enzyme production in some fashion. Additionally, theenzyme is initially translated as a large (ca. 51,500-dalton),enzymatically active intracellular precursor in B. subtilisbefore undergoing efficient secretion to the exterior of thecell (45). The mature exocellular ,B-1,4-glucanase is 35,200 ±400 daltons.
Cloning the ,3-1,4-glucanase gene has made possible acloser study of some aspects of the regulation and transportof the enzyme. In this study, we describe the DNA sequenceof the P-1,4-glucanase gene and identify the in vivo start siteof the transcription and promoter region. We identify the siteof signal sequence processing and speculate as to the likeli-hood of additional and unusual carboxy-terminal processing.
* Corresponding author.t Present address: Department of Pharmacology and Experimen-
tal Therapeutics, The Johns Hopkins University School of Medi-cine, Baltimore, MD 21205.
The effect of glucose on the level of P-1,4-glucanase tran-scription is investigated. Finally, we directly compare thespecific activity of B. subtilis DLG P-1,4-glucanase with thatof Trichoderma reesei QM9414 endo-P3-1,4-glucanase on thenoncrystalline substrate trinitrophenyl (TNP)-CMC.
MATERIALS AND METHODSBacterial strains, plasmids, and bacteriophage. The Bacil-
lus strains used were B. subtilis DLG, a wild-type strainwhich produces P-1,4-glucanase (44, 45), and B. subtilisPSL1 (stp leuA8 arg-15 thrA recE4 hsrR hsrM) (38). The twostrains of Escherichia coli used were C600SF8 (thr-J leuB6thi-l recBC lop-il lacYl lig+/hsdM hsdR4 (54) and K-12JM101 [supE thi A(lac-proAB) (F' traD36 proAB laclIqZAM15)] (64). E. coli K-12 JM101 was used as a host forM13mpl8 and M13mpl9 (64). The B. subtilis-E. coli shuttleplasmid pLG4002 (Tcr Apr Cmr, ,B-1,4-glucanase+) (45) wasused.
Culture media and growth conditions. E. coli C600SF8 wasgrown in an L-broth medium containing the following (ingrams per liter): tryptone (Difco Laboratories, Detroit,Mich.), 10; yeast extract (Difco), 5; and NaCl, 5 (pH 7.2). E.coli K-12 JM101 was maintained on glucose-minimal me-dium agar containing the following (in grams per liter):Na2HPO4, 6; KH2PO4, 3; NH4Cl, 1; NaCl, 0.5;MgSO4 *7H20, 0.493; CaCl2 * 2H20, 0.0147; thiamine hy-drochloride 0.001; glucose, 0.2; and minimal agar, 15 (pH7.0). B. subtilis PSL1 and DLG were grown in nutrientsporulation medium (50) with or without 1% glucose. Selec-tive antibiotic media contained either ampicillin (30 jxg/ml) orchloramphenicol (7.5 ,ug/ml). Liquid cultures were grownwith vigorous aeration at 37°C. Growth was monitored byabsorbance measurements in a Klett-Summersoncolorimeter (Klett-Summerson, New York, N.Y.) fitted witha no. 66 red filter.
Plasmid isolation, restriction enzyme and ligation treat-ments, and transformation procedures. Plasmid and phagereplicative-form (RF) DNA were prepared by an alkalinelysis procedure (2) as modified by Maniatis et al. (31).Covalently closed, supercoiled DNA was purified by CsCl-
2017
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

2018 ROBSON AND CHAMBLISS
E
4-
.44 - 46--S 0
4-
H
200 400 600 600 000 1200 1400 1600 1800
FIG. 1. Relevant restriction sites and sequencing strategy of the 3-1,4-glucanase gene. , Coding sequence of P-1,4-glucanase; -,extent and direction of sequence reactions; rrrn , EcoRl-DdeI restriction fragment used in Si nuclease mapping. Distances are indicated inbase pairs starting at the EcoRI site. Abbreviations: E, EcoRI; S, SacI, D, DdeI; H, HindIII.
ethidium bromide equilibrium density gradient centrifuga-tion, n-butanol extractions, and dialysis against 10 mM Trishydrochloride-i mM EDTA (pH 8.0) (31, 42). Restrictionenzyme digestions, dephosphorylation reactions with calfintestinal alkaline phosphatase, and ligations with T4 DNAligase were carried out as specified by the manufacturers.Agarose gel electrophoresis was used to analyze plasmids,phage, endonuclease restriction fragments, and ligation re-actions. Agarose gels were made up and run in 40 mMTris-acetate-2 mM disodium EDTA containing ethidiumbromide (0.5 ,ug/ml). DNA was detected by illumination witha shot-wavelength UV transilluminator (Fotodyne, NewBerlin, Wis.). HindIII-digested lambda DNA and HincII-digested 4~X174 RF DNA were used as molecular sizemarkers. Low-melting-point agarose (FMC Corp.,Rockland, Maine) and Elutip-d columns (Schleicher &Schuell, Keene, N.H.) were used to recover and purify DNAfragments from preparative gels as specified by the manu-facturers. Transformation of B. subtilis PSL1 was perfomedby the method of Boylan et al. (4). Transformation of E. colistrains was carried out by the CaCl2 shock procedure (29;M13 Cloning and Sequencing Handbook, Amersham Corp.,Arlington Heights, Ill.).DNA sequence determination. A 2-kilobase (kb) EcoRI-
HindIII DNA fragment encoding the B. subtilis DLG P-1,4-glucanase gene was isolated and purified from pLG4002 byEcoRI and HindIII digestion and low-melting-point agarosegel electrophoresis. This 2-kb DNA fragment was thendigested with Sacl. The resulting EcoRI-SacI (ca. 400-base-pair [bp]) and SacI-HindIII (ca. 1,600 bp) restriction frag-ments were each cloned into appropriately digested anddephosphorylated M13mpl8 and M13mpl9 RF DNA. Anexonuclease digestion procedure (9) was used to generate asequential series of overlapping clones of the SacI-HindIIIDNA fragment in mpl8 and mp19. The dideoxynucleotidechain termination method of Sanger et al. (47), as describedin the M13 Cloning and Sequencing Handbook (AmershamCorp.), was used in the DNA sequence determination. DNAwas labeled with [a-35S]dATPaS (Amersham Corp.) duringthe sequencing reaction. Both strands were sequenced,except for ca. 100 bp at the extreme 5' end of the 2-kbfragment, for which the sequence was determined from onlyone strand.
Si nuclease mapping. RNA was isolated from E. coli andB. subtilis strains at various time points within the cell cycleby the procedure of Chambliss et al. (8) up to, but notincluding, the NaCl precipitation step. Instead, RNA wasprecipitated with 4 M LiCl as described by Cathala et al. (7),and their procedure was followed from that point on. A541-bp EcoRl-DdeI DNA restriction fragment (Fig. 1) en-compassing the putative 5' end of the 0-1,4-glucanase-codingregion was isolated from pLG4002 and used as a hybridiza-tion probe. 5'-end-labeling reactions were carried out with[-y-32P]ATP (specific activity, 3,000 Ci/mmol; Amersham
Corp.) and T4 polynucleotide kinase (31). Elutip-d columnswere used to separate end-labeled double stranded DNAfrom unincorporated label. Hybridization and SI digestionreactions were as described by Berk and Sharp (1) with thefollowing modifications: denaturation was performed for 10min at 72°C, hybridization was performed overnight at 45°C,samples were diluted 10-fold with 170 U of Si nuclease in350A Si digestion buffer, and Si digestion was for 30 min at45°C. After ethanol precipitation, pellets were suspended in5X gel loading buffer (80% deionized formamide, 0.01 MNaOH, 0.001 M EDTA, 0.03% bromphenol blue, 0.03%xylene cyanol). Samples were heated to 95°C for 2 min,chilled, and loaded onto 6% polyacrylamide gels containing8 M urea. In each reaction, 50 ,ug of RNA, 50 ,ug of carriertRNA, and 50,000 cpm of the 5' end-labeled EcoRI-DdeIDNA probe were used. A modification of the dideoxynu-cleotide chain termination procedure (25) was used to gen-erate the 541-bp EcoRI-DdeI DNA probe sequence ladder inthe Si mapping experiments. Owing to the use of thisprocedure, however, the EcoRI-DdeI DNA restriction frag-ment used for aligning the migration of the Si nuclease-resistant DNA bands in the sequencing ladder had four extrabases at the 3' end.
0-1,4-glucanase assay. ,-1,4-Glucanase was assayed bymeasuring the degradation of TNP-CMC (22). One unit ofTNP-CMC-degrading activity was defined as the amount ofenzyme that, in 1 min, released soluble color correspondingto a specific A344 of 0.01. Protein concentrations weredetermined by the method of Lowry et al. (28) with bovineserum albumin as a standard.
Purification of P-1,4-glucanase. B. subtilis DLG was grownfor 20 h at 37°C with vigorous aeration in 2 liters of nutrientsporulation medium plus 1% glucose. Cells were separatedfrom the culture fluid by centrifugation. Crystalline(NH4)2SO4 to 70% saturation was added to the fluid, and thesolution was stirred for 36 h at 4°C. Precipiated material wasremoved by filtration through a glass fiber filter (Schleicher& Schuell). The 70% (NH4)2SO4 filtrate was then brought tosaturation with crystalline (NH4)2SO4 and stirred gently for24 h at 4°C. Precipitated material, including exocellular,-1,4-glucanase, was collected by centrifugation and sus-pended in 15 ml of 0.05 M citrate-phosphate buffer (pH 5.5).Contaminating (NH4)2SO4 was removed by passing the prep-aration through a Bio-Gel P-6DG (Bio-Rad Laboratories,Richmond, Calif.) desalting column equilibrated and devel-oped with 0.01 M phosphate buffer (pH 6.6). The desaltedprotein was frozen at -80°C and lyophilized. Afterlyophilization, aliquots ofthe crude protein preparation weresuspended in 0.01 M phosphate buffer (pH 6.6) and loadedon a DEAE-5PW high-performance liquid chromatography(HPLC) ion exchange column (Beckman Instruments, Inc.,Berkeley, Calif.) equilibrated with 20mM Tris hydrochloride(pH 7.9). A gradient of increasing NaCl concentration in 20mM Tris hydrochloride (pH 7.9) was used to develop the
3, .0.
J. BACTERIOL.
I
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

ENDO-1-1,4-GLUCANASE GENE OF B. SUBTILIS DLG 2019
column. P-1,4-Glucanase came off the column when theeluant NaCl concentration reached approximately 0.03 M.A255 and A280 were monitored. Fractions containing P-1,4-glucanase were pooled and desalted in a Bio-Gel P-6DGcolumn equilibrated and developed with 0.01 M ammoniumacetate buffer (pH 5.6). The eluted protein was lyophilizedand suspended in 0.01 M ammonium bicarbonate (pH7.0)-0.02% sodium dodecyl sulfate SDS prior to amino acidsequence analysis. The purity of the final preparation waschecked by SDS-polyacrylamide gel electrophoresis (PAGE)(27). The HPLC portion of the work was performed at theBiotechnology Resource Center, University of Wisconsin,Madison.
Protein sequence determination. The NH2-terminal se-quence of mature exocellular 35,200-dalton P-1,4-glucanasefrom B. subtilis DLG was determined by stepwise Edmandegradation of ca. 100 jig of the enzyme with an automatedgas-phase protein sequencer (model 470A; AppliedBiosystems, Foster City, Calif.) (23). The resulting phenyl-thiohydantoin amino acid derivatives were identified byHPLC (49). This work was performed at the BiotechnologyResource Center, University of Wisconsin, Madison.
Chemicals and reagents. Ampicillin, chloramphenicol, sol-uble CMC, and Congo Red were obtained from SigmaChemical Co., St. Louis, Mo. TNP-CMC was prepared bythe method of Huang and Tang (22). Restriction endonu-cleases, T4 DNA ligase, T4 DNA polymerase, and T4polynucleotide kinase were purchased from Promega-Biotec, Madison, Wis. S1 nuclease was obtained from Be-thesda Research Laboratories, Inc., Gaithersburg, Md., andcalf intestinal alkaline phosphatase was purchased fromBoehringer Mannheim Biochemicals, Indianapolis, Ind. Adideoxy DNA sequencing kit was purchased fromAmersham Corp. Deletion subcloning primers (9) were ob-tained from International Biotechnologies, Inc., New Ha-ven, Conn. Purified Trichoderma reesei QM9414 endo-P-1,4-glucanase (EGI; EC 3.2.1.4) was a gift from R. Brown, Jr.All other chemicals were of standard reagent grade.
RESULTS
DNA sequence analysis. Through DNA sequence analysis,the ca. 2-kb EcoRI-HindIII DNA restriction fragment previ-ously found to encode P-1,4-glucanase in B. subtilis DLGwas determined to be 1,925 bp in length. A partial restrictionmap of the fragment and the strategy used for dideoxysequencing is shown in Fig. 1. The complete DNA sequenceis shown in Fig. 2. A computer analysis of this sequencerevealed only one open reading frame long enough to encodethe ca. 51,500-dalton protein detected by SDS-PAGE ofintracellular extracts of E. coli and B. subtilis transformantscarrying the ,B-1,4-glucanase gene on a high-copy-numberplasmid (45). This sequence begins at nucleotide 391 andends at nucleotide 1914. There are three ATG codons nearthe 5' end of the coding region which could serve as startsites of translation. Each of these is followed by an aminoacid sequence which resembles signal sequences found ingram-positive organisms. Such sequences consist of a rela-tively short hydrophilic region at the NH2 terminus followedby a longer stretch of hydrophobic residues (3, 40). There aretwo putative ribosome-binding sites upstream from the pos-sible start sites of translation (51). The first begins atnucleotide 378 (AGAAAGAG) and would display a calcu-lated free energy ofAG = -10.4 kcal/mol (ca. -43.5 kJ/mol)if bound to the 3' end of 16S rRNA (59). The first and secondin-frame ATG codons occur 5 and 8 bp downstream from
this sequence, respectively. The second likely ribosome-binding site begins at nucleotide 399 (AAGGAGG). If boundto the 3' end of 16S rRNA, this sequence would exhibit acalculated free energy of AG = -17.8 kcallmol (ca. -74.5kJ/mol). The third in-frame ATG occurs 12 bp downstreamfrom the second ribosome-binding site.The region upstream from the long open reading frame
was scanned by computer for sequences bearing homologyto various promoter consensus sequences used by B. subtilisRNA polymerases (24). A sequence bearing good homologyto the o43-type vegetative promoters of B. subtilis and thex70-type promoters of E. coli was found at nucleotides 334 to362. This sequence consists of TAGACA as the -35 regionand TACAAT as the -10 region; the two are separated by 17nucleotides. A likely start site of transcription was found atnucleotide 369 (G) or 370 (A). There was little homology tothe sequence RTRTG (R = purine) 1 nucleotide upstreamfrom the -10 region; such a sequence is believed to be acommon feature of strong B. subtilis vegetative promoters(34, 36). The region -57 to -35 bp was about 75% AT andcompares well with other Bacillus promoters in this respect(34). Upstream DNA (nucleotides 1 to 450) was alsosearched for complementary inverted repeat sequenceswhich, if transcribed, could be capable of forming hairpinsecondary structure. However, those found had calculatedfree binding energies of AG > -4.5 kcailmol (ca. -22.6kJ/mol) and so were not deemed particularly significant. Noinformation on possible transcription termination features ofthe P-1,4-glucanase gene was obtained, because the HindIIIsite used in cloning the gene for sequence analysis waspositioned very near the 3' end of the coding region.A hydropathicity profile (26) of the inferred amino acid
sequence of the enzyme is shown in Fig. 3. On the basis ofthis analysis, the protein as a whole appears to be slightlyhydrophilic, with the exception of the strongly hydrophobicportion of the putative signal sequence at the amino termi-nus. The codon usage of the open reading frame resembledthat of most other B. subtilis genes in that it was markedlyunbiased (37) (tabulated data not shown).
S1 nuclease mapping. High-resolution S1 nuclease map-ping studies were undertaken to locate the start site oftranscription of the B. subtilis DLG P-1,4,-glucanase gene invivo and to determine whether the temporal production ofthe enzyme after the onset of the stationary phase was due toregulation of the gene at the transcriptional level. Results areshown in Fig. 4. Alignment of the S1 nuclease-resistantfragment of the DNA probe with the corresponding bands inthe sequencing ladder showed that transcription did initiatein vivo at the predicted site, i.e., nucleotide 369 or 370.Additionally, it was observed that amounts of gene-specificmRNA were very low in mid- to late-exponential- andearly-stationary-phase cells (lanes 3 and 4) but that theamount of P-1,4,-glucanase mRNA increased substantiallyduring stationary phase (lanes 5 to 7). No other transcrip-tional start sites were used in vivo, at least up to 5 h after theonset of the stationary phase.When the start site of (3-1,4-glucanase transcription was
mapped by using RNA isolated from E. coliC600SF8(pLG4002) and B. subtilis PSL1(pLG4002), it wasobserved that the same unique initiation site was used ineach transformant strain as was used in B. subtilis DLG (Fig.5, lanes 2 to 4).
Effect of glucose on ft-1,4-glucanase transcription. Previ-ously, we had observed that although B. subtilis DLG didnot produce (3-1,4-glucanase when grown on nutrientsporulation medium, growth on nutrient sporulation medium
VOL. 169, 1987
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

2020 ROBSON AND CHi' fI 9''
NUT TCT GGC TGA ATC CCT A'r ':'.3),- 7i AACEcoRI 12 24 3t4.^C77 AAC CCG TAT TAA TCC GAA CCL ' . 1'.A!? ¼' CC?
66 7
CTT CTT ACA GCA ATC aTA AGA TY- Ai' .;lN..-'2T:C..') ACc120 13Z 4'
GTG TCT ACG TTC cAA ATA CAT T73 T, tvJ TCA1P'9':. TCTGT174 1 2'
AAA GTC CCC ATT CAc CCC GGT TS; . f;t , ;.'.) 13 31; TTA
228 2'.,w
TTC AAT AcA GAT AcA GCA AAT TC C'7: T.-i ?3'AC ,- .1;, 44AhA} .:AAA282 294
-;wTTA GCT TCA T ACA TC ATCAc,A ..t-' 7,FA T.TAA
336 34. '2)
- iiiie u,a, r .. Fit 110
GAA AGA CC AAAIATC ATAG.
AC C2 ATT
330 40~429
Ser 11. Ph. II. Thr Cys Laus T I., LauTCT ATT TTT ATT ACG TOT TTA t t 7A r; G A r TrC, CTG
444 45' F
Pro Ser Pro Al. Ser Al. Al -.iAl;,'.l'.. 0ClnCCT TCC CCC cCA TCA GCA GCA Cc; IRA I,' '.', .'GACA TCCA CA-T C
49 5- ? l.
Lou Ser Lys Cly Thr Gln La /. 1''r, LouCT aG ATA AAA COT ACA CAc Ct t v.;A 11 04 r A CAItCTC
O de- I 552 56i $ fi; ;Lys G1y II. Ser Ser His G1yL',\m , .. 4.;,^r.,31 Asp
AAA CCC ATC ACT TCA CAT GCAA'T-",i34T. 13 344% CAC
Ser Lau Lys Trp Leu Arg AspA - Cl-C) 31 Ii. 1 F - HtaCc TTA AAA TCC CtC AcA cAC CG T rC CT4.' G3; -r G ATG
650 674
Tyr Thr Ala Asp Cly G1 Tyr12) y'12' 'al1 Lys
TAT ACc cCA GAT CGC CGT TAT AT- A' ':A Aaa714 ;1.
0lu Ale Va1 Clu Ala Ale Lys GC . '.I I V.)-1.4eTI t ? rrp His
cAa CCC OTT cAA CCC GCA AAA GA . ?; T1.A "'i ','T 1'- t, .1. CAT
763 74C,c
I1. Las Asn Asp Cly Asn Pro As r, "'It' 4A' y I t G '-I FP's Ph.
ATT TTA aaT GAc CCC AAC CCA AAtC ('i A ii'A2 '"1 TTC
322 C33
Lys Glu Net Ser Ser Lau Tyr Cly A iT4r. o A VA) i T3.. Ale
AAc cAc at5 TCA AGT CTT TAC CCA A(' A C A '5A i.A ITT GCA876 89F ~ eev0;-(
Asn Clu Pso Asn 01 Asp Va1 As -p L3s ArS -1:l r Try .l GluAAC CAA CCA AAC CGT cAC GTG AAC T'71 Al '.33; 4I '. CCC
930 943 34
Clu V.1 II. Ser Val 11i Arg Ly- t r. : e Va1 017
GSA CTC ATT tCC CTT ATC CCC AA. r*3i h,5tA r, A - ,T CTC CCC
9 34 99 ;134
Thr Gly Thr Trp Ser CGl Asp Va1 Asn Asp Ala Ale Asp Asp Gln Lou Lys AspACC COT ACA 7CC ACC CAA CAT CTC AAT CAT cCA CCC cAT CAT CAC CTA AAA CAT
1035 1050 1062 1074
Al. Asn V1l Net Tyr Al. Lou His Ph. Tyr Ala Thr His 0ly Cln Ser LouGCA AAC CTC ATG TAC CCC CTT CAT TT7 TAT CCC CCC ACA CAT CCC CAA TCT TTA
1092 1104 1113 1123
Arg Asp Lys Al. Asn Tyr Al. Las Ser Lys Cly Al. Pro II& Ph. Va1 Thr CluCCC cAT MaA cCA AAC TAT cCA CTC ACT AAA ccA CCC CCT AT? TTC CTG ACc CAA
1146 1158 1170 1182
Trp Thr Ser Asp Al. Ser Asn 01y 01 Va1 Ph. Lau Asp Cln Ser argT70 ccA ACA AcC cAC CCC TCT GCA aaT CCC COT OTA TTC CTT GAC CAc TCC CCC
1200 1212 1224 1236
Gl Trp LuasAn Tyr Lou Asp Ser Lys Asn II* Ser Trp Va1 Asa Trp Asn LaucAA tcc CTC AAT TAT CTC cAC ACT Aac aaC ATC ACc TCC 0t7 AAC 7CC AAT CTT
1254 1266 1273 1290
Ser Asp Lys Gln Clu Ser Ser Ser Al. Lau Lys Pro Cly Al. Ser Lys Thr 1lyTCT cAT Mc CAc cMa TCA TCT TCC GCT TTA aMc CCC cca cCA TCT AAA ACA CCC
1308 1320 1332 1344
Gly Trp Pro Lau Thr Asp Las Thr Ale Ser Gly Thr Ph. Val Arg GCl Asn II.CCC TOC CCC CTT ACA cAT TTA ACT OCT TCA ccA ACA TTC CTA AcA cAA AAC 7TT
1362 1374 1386 1393
Arg Cly Thr Lys Asp Ser Thr Lys Asp Val Pro Clu Thr Pro Ale Cln Asp AsnCCC GCC ACT AAA CAT TCC ACO aac cAC CTC CCT GaA ACG CCA cCA CAA CAT AAC
1416 1428 1440 1452
Pro Thr Gls Cil Lys 0ly V.1 Ser Va1 Cla Tyr Lys Al. Gl0 Asp 0ly Arg VelCCC ACA CAC caA Aaa CCC CTT TCT CTA CAA TAC aaa Gca CCC CAT CCC COT CTC
1470 1482 1494 1506
Asn Ser Asn Gln 11 Arg Prc Cln Lau His Ile Lys Asn Asn Cly Asn Ale ThraaC ACC aaT CAA ATC CCC CCC CAc CTT CAC ATA aaA AaT ac CCC AAT CCC ACG
1524 1536 1548 1560
Vel Asp Lae Lys Asp Val Thr Ale Arg Tyr Trp Tyr Asn Val Lys Asn Lys GlyCGT cAT TTA AMa cAT CTC ACT CCC COT TAC tCC TAT MaC 0T7 AaA SAC AAA CCC
1578 1590 1602 1514
CGl Asn Ph. Asp Cys Asp Tyr Al. Gln Net 01y Cys 0ly Asn Lau Thr His LysCAA aAC TT7 cAC TOT cAC TAC CCC CAc ATG GCA TCC CCC aAT CTC ACC CAC AaG
1632 1544 1656 1668
Ph. Vel Thr Lau His Lys Pro Lys Gln Gl0 Ale Asp Thr Tyr Lau Glu Leu GlyT77 0TG ACO CTC CAT Aaa CCT Aac CAa CTCCcA cAT ACC TAT CTG cAa CTC CGG
1686 1698 1710 1722
Ph. Lys Thr Gly Thr Lau Ser Pro Cly Ale Ser Thr 0ly Asn Ile Cln Lau ArgaCA ccA ACc CTC TCA CCC CGA GCA AcC Aca CCC SAT aTT CAc CTT COT
1740 1752 1764 1776
Lau His Asn Asp asp Trp Ser Asn Tyr Ala Gln Ser Asp Tyr Ser Ph. Ph.CTT CAC AAT CAT cAC TCC AcC AaT TAT GCA CAa aSC CCC CAT TAT TCC TTT TTC
1794 1306 1313 1830
Gln Ser Asn Thr Ph. Lys Thr Thr Lys Lys I1e Thr Lau Tyr His Gln 0ly LysCAA TCA aaT aCG TT7 5Aa ACA ACc Aa MAA ATC aCA T7A TAT CAT CAA GcA AAa
1348 1860 1872 1334
Lau I1 Trp Gl0 Thr Olu Pro Asn *** Klind IIICTC aTT TOG GcA ACA Gaa CCC AaT TAO TTA ACc T7
1902 1914 1926
FIG. 2. DNA sequence of tL-. V "ti 'i ..'1 f-F,-'.- '!:canase gene. A nontranscribed strand is shown. The -35 and -10 regions of the
promoter are underlined and labt' (l; "ir. ' 't iucleotides 369 and 370 indicate the start site(s) of transcription. The most likelyribosome-binding site is labeled , t, ''r'l A a 1,ed double line indicates an alternative ribosome-binding site. The most likely startsite of translation is boxed with (l 1o ,) lie'i '.. enclose alternate translation initiation codons. An arrow indicates the site of signalsequence cleavage. The amino-torfriu31 (' j r)r3l,tre exocellular 0-1,4-glucanase which were identified by Edman degradation areoverlined: the remaining residue Kie ",NA sequence. Restriction endonuclease cleavage sites are bracketed and labeled.
4
3
2
c
0
>%
'iI Al AA AA. L
fFv YVt
50 100 5i? 200 250 300 350 400 450 500
Amino Acid Residues
FIG. 3. Hydropathic analysis of the Ted amino acid sequence of ,B-1,4-glucanase. Numbering of the residues is based on theassumption that translation initiates at the I fG codon located at nucleotides 418 to 420 of the DNA sequence shown in Fig. 2.
J. BACTERIOL.
t,1
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
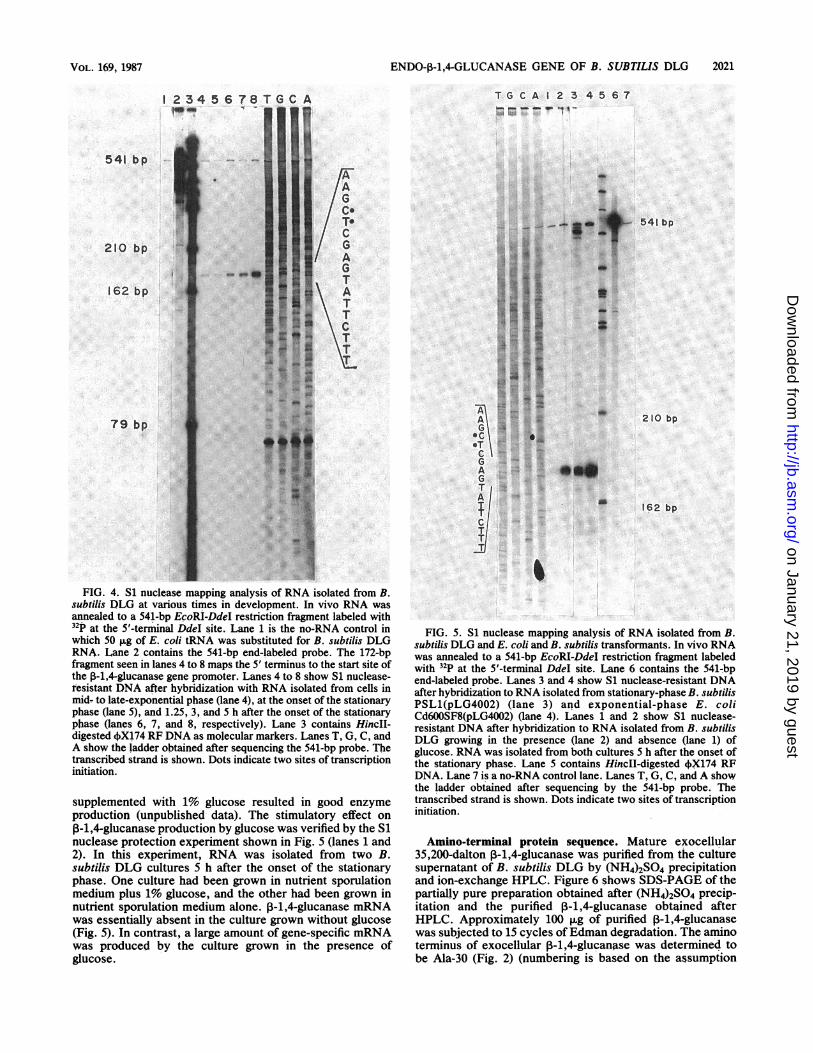
ENDO-P-1,4-GLUCANASE GENE OF B. SUBTILIS DLG 2021
T G CA I 2 3 4 5 6 7_- _.0 " IlAz Es
5 6 78
I79 bp
FIG. 4. Si nuclease mapping analysis of RNA isolated from B.
subtilis DLG at various times in development. In vivo RNA was
annealed to a 541-bp EpoRI-DdeI restriction fragment labeled with
tthe 5'-terminal Ddel site. Lane 1 is the no-RNA control in
which 50 iLg of E. coli tRNA was substituted for B. subtilis DLG
RNA. Lane 2 contains the 541-bp end-labeled probe. The 172-bpfragment seen in lanes 4 to 8 maps the 5' terminus to the start site of
the 0-1,4-glucanase gene promoter. Lanes 4 to 8 show Si nuclease-
tesistant DNA after hybridization with RNA isolated from cells in
mid- to late-exponential phase (lane 4), at the onset of the stationary
phase (lane 5), and 1.25, 3, and h after the onset of the stationary
phase (lanes 6, 7, and 8, respectively). Lane 3 contains HincII-digested 4OX174 RF DNA as molecular markers. Lanes T, G, C, and
A show the ladder obtained after sequencing the 541-bp probe. The
transcribed strand is shown. Dots indicate two sites of transcriptioninitiation.
supplemented with 1% glucose resulted in good enzyme
production (unpublished data). The stimulatory effect on
0-1,4-glucanase production by glucose was verified by the Si
nuclease protection experiment shown in Fig. 5 (lanes 1 and
2). In this experiment, RNA was isolated from two B.
subtilis DLG cultures 5 h after the onset of the stationary
phase. One culture had been grown in nutrient sporulationmedium plus 1% glucose, and the other had been grown in
nutrient sporulation medium alone. P-1,4-glucanase mRNA
was essentially absent in the culture grown without glucose
(Fig. 5). In contrast, a large amount of gene-specific mRNA
was produced by the culture grown in the presence of
glucose.
0
AGl
C
TA
1~TCl
_E
...
.: = i* _s a F
_ = ... 1 F.....X s ..| ig *
E.e. Ei: rE t wu L
S g[ |
|r | s[ w ....
|K :
541 bp
210 bp
0f
162 bp
FIG. 5. Si nuclease mapping analysis of RNA isolated from B.subtilis DLG and E. coli and B. subtilis transformants. In vivo RNAwas annealed to a 541-bp EcoRI-DdeI restriction fragment labeledwith 32p at the 5'-terminal DdeI site. Lane 6 contains the 541-bpend-labeled probe. Lanes 3 and 4 show Si nuclease-resistant DNAafter hybridization to RNA isolated from stationary-phase B. subtilisPSL1(pLG4002) (lane 3) and exponential-phase E. coliCd600SF8(pLG4002) (lane 4). Lanes 1 and 2 show Si nuclease-resistant DNA after hybridization to RNA isolated from B. subtilisDLG growing in the presence (lane 2) and absence (lane 1) ofglucose. RNA was isolated from both cultures 5 h after the onset ofthe station,ary phase. Lane 5 contains HinclI-digested',X174 RFDNA. Lane 7 is a no-RNA control lane. Lanes T, G, C, and A showthe ladder obtained after sequencing by the 541-bp probe. Thetranscribed strand is shown. Dots indicate two sites of transcriptioninitiation.
Amino-terminal protein sequence. Mature exocellular35,200-dalton P-1,4-glucanase was purified from the culturesupernatant of B. subtilis DLG by (NH4)2SO4 precipitationand ion-exchange HPLC. Figure 6 shows SDS-PAGE of thepartially pure preparation obtained after (NH4)2SO4 precip-itation and the purified P-1,4-glucanase obtained afterHPLC. Approximately 100 ,ug of purified ,-1,4-glucanasewas subjected to 15 cycles ofEdman degradation. The aminoterminus of exocellular j3-1,4-glucanase was determined tobe Ala-30 (Fig. 2) (numbering is based on the assumption
1 234
-4
541 bp |
210Ob
162 bp
VOL. 169, 1987
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

2022 ROBSON AND CHAMBLISS
1 2 A
$71194 a ....
(57am _ ...
60 _AW
43 .p s§
430
FIG. 6. Purification of exocellular endo-13-1,4-glucanase. SDS-
PAGE of purified endo-13-1,4-glucanase obtained after ion-exchange
HPLC (lane A) and partially purified endo-P3-1,4-glucanase prepara-
tion obtained after (NH4)2S04 precipitation (lane B). Lanes 1 and 2
contain molecular weight markers identified along the left side of the
figure.
that translation initiates at the ATG codon located 12 bpdownstream from the strongest putative ribosome-binding
site). The results of the following 14 cycles of Edman
degradation confirmed the amino acid sequence inferred
from the DNA sequence of the gene. The amino terminus of
intracellular precursor 13-1,4-glucanase was not determined;
therefore, the precise start site of translation is still un-
known.
Specific activity determination. The specific activity of
purified B. subtilis DLG 13-1,4-glucanase was compared with
that of purified EGI of T. reesei QM9414, a hypercellulolyticstrain. The assay measured the degradation of TNP-CMC, a
noncrystalline cellulosic substrate. The specific activity of B.
subtilis DLG 13-1,4-glucanase was found to be 1.8 times that
ofT. reesei QM9414 endo-P3-1,4-glucanase for this substrate,
i.e., 550 versus 300 U/mg.
DISCUSSION
DNA sequence analysis and nuclease mapping studies
of the 13-1,4-glucanase gene of B. subtilis DLG revealed
several features common to other Bacillus genes. Amongthese was a promoter, TAGACA (-35 region) and TACAAT
(-10 region), closely resembling the consensus promoter
recognized by or43-RNA polymerase, TTGACA (-35 region)and TATAAT (-10 region) (34). The gene also contained an
A+T-rich region upstream from the promoter between -57
and -35 bp and two potential ribosome-binding sites each
followed by an ATG codon correctly placed to initiatetranslation of a long open reading frame (34, 36). Thecalculated free binding energy associated with the firstribosome-binding site (AG - -10.4 kcal/mol) is lower thanusually associated with Bacillus ribosome-binding sites (18).In contrast, the second ribosome-binding site is associatedwith a free binding energy of -17.8 kcal/mol, well within thestandard range for Bacillus spp. Hence, the most likelytranslation start site is the ATG codon which lies 12 bpdownstream from the second ribosome-binding site.Like many other genes coding for a secreted product, the
P-1,4-glucanase gene codes for a signal sequence at theamino-terminal end of the protein. Such sequences arebelieved necessary for efficient export of a protein in mostorganisms and are usually composed of a short stretch ofcharged amino acids (2 to 11 residues) at the amino terminusfollowed by a longer stretch (14 to 20 residues) of stronglyhydrophobic amino acids (3, 40). Signal sequences fromgram-positive organisms have the same structure, but areoften slightly longer than those of other systems (18, 35).DNA sequencing studies also revealed that the P-1,4,-
glucanase gene has a coding capacity for a ca. 55,000-daltonprotein. In contrast, mature exocellular P-1,4-glucanase isonly 35,200 daltons. This result was not unexpected, since inearlier studies, a large form of P-1,4,-glucanase (ca. 51,500daltons by SDS-PAGE) had been observed in intracellularextracts of E. coli and B. subtilis transformants carrying theP-1,4-glucanase gene on a high-copy-number plasmid (45).As this large, and, presumably, precursor ,B-1,4-glucanasewas never detected in B. subtilis DLG intracellular extracts,it is likely that maintenance of the gene on a high-copy-number plasmid overloaded the transport-secretion systemof the transformed B. subtilis strain, so that a small amount(<5%) of precursor P-1,4-glucanase accumulated insidethese cells. A significant amount (ca. 60%) of precursor3-1,4-glucanase was present within E. coli transformants,
which probably reflected both the high copy number of thegene and the decreased ability of E. coli to correctly processthe precursor (45).The hypothesis was formed that the 3-1,4,-glucanase gene
might encode a region between the signal sequence and themature exocellular ,B-1,4-glucanase-coding region that isremoved in a similar fashion as some Bacillus proteasegenes. Such regions ranging from 77 to 206 residues havebeen found to be encoded by four protease genes of Bacillusorigin (56, 61, 62), including the subtilisin gene of B. subtilis(62). Like signal sequences, these regions are not containedin the mature exocellular forms of the proteases.
Purification and subsequent amino-terminal protein se-quence analysis of mature exocellular 35,200-dalton 3-1,4-glucanase showed that a region located between the signalsequence and the mature exocellular enzyme did not exist.Apparently, 13-1,4-glucanase precursor undergoes signal se-quence processing at the amino terminus at some pointbefore or during its secretion. The site at which the process-ing occurs is located between two alanine residues followingthe hydrophobic region. This correlates well with the accu-mulated data on signal sequence cleavage sites in membersof the family Bacillaceae and other organisms (33, 35). Theamino terminus of the mature, exocellular 35,200-daltonprotein is the second of the two alanines at the cleavage site.The apparent ca. 16,500-dalton difference between
exocellular f-1,4,-glucanase and the precursor form minusthe signal sequence could be due to one of several posttrans-lational modifications. One possibility is that the precursorundergoes significant carboxy-terminal processing as well as
J. BACTERIOL.
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

ENDO-P-1,4-GLUCANASE GENE OF B. SUBTILIS DLG 2023
amino-terminal processing of a typical signal sequence. Itwill be necessary to sequence the carboxy terminus of themature exocellular enzyme to substantiate this hypothesis.Examples of carboxy-terminal processing of other Bacil-
lus proteins have not yet been reported. However, theimmunoglobulin Al proteases of Haemophilus influenzaeand Neisseria gonorrhoeae do appear to undergo suchprocessing (43; F. Grundy and A. Wright, personal commu-nication). For the H. influenzae protein, there is someevidence that the carboxy-terminal portion of the proteaseprecursor has a role in the secretion of the matureexocellular protein, even though the immunoglobulin Alprotease gene also appears to encode an amino-terminalsignal sequence. Whether or not the carboxy-terminal por-tion of precursor f3-1,4,-glucanase plays a role in secretionhas yet to be elucidated.
In contrast, the ,-1,3-1,4-glucanase gene of B. subtilisC120 has been sequenced and appears to encode only themature enzyme and a standard signal sequence at the aminoterminus of the gene coding region (35). The same is true ofseveral a-amylase genes (39, 52, 63), a xylanase gene (15),and a levansucrase gene (14) of Bacillus origin. However,additional DNA and protein sequence data are needed topredict whether carboxy-terminal processing is a commonphenomenon among Bacillus proteins and extracellular en-zymes in particular.The finding that the ,-1,4-glucanase gene is transcribed in
vivo from a promoter resembling the consensus of promoterstranscribed by a43 RNA polymerase is noteworthy, since a43is the predominant a factor present in B. subtilis duringvegetative growth and persists at lower levels throughoutlater stages of growth (10). Other minor a factors are alsofound in B. subtilis; some are present during vegetativegrowth (r28) (16), others do not appear until later in devel-opment (a29) (60), and still others are found during severalgrowth phases [a37 (19) and &32 (24)]. Each of these a factorsappears to recognize and initiate transcription from a dif-ferent set of promoters in B. subtilis. Expression of certaingenes at different times in cell development has been postu-lated to be due, at least in part, to transcriptional regulationof these genes by a-factor recognition specificity (10). Thereis evidence to support this model (60), but the system is verycomplex.The Si nuclease mapping experiments performed in this
study suggest that the temporal expression of P-1,4-glucanase after the onset of the stationary phase is due toregulation at the level of transcription. P-1,4-GlucanasemRNA is just detectable in logarithmic-phase and earlystationary-phase cells but increases dramatically during thecourse of the stationary phase. As the gene is essentiallyunexpressed during logarithmic growth, yet appears to betranscribed from a a43-type vegetative promoter, it is likelythat there is some form of transcriptional regulation otherthan a-factor recognition of a promoter which is responsiblefor the temporal expression of the ,-1,4-glucanase gene. Analternative explanation, which cannot be ruled out at thistime, is that there is increased stability of 3-1,4-glucanasemRNA after the onset of stationary phase rather thantemporal transcription of the gene.Exoenzyme genes of B. subtilis have been one class of
genes thought to be transcribed by minor a RNA polymer-ases (62). This was because many of these enzymes areproduced only after logarithmic growth (6, 41, 46, 56, 61, 62).However, recent DNA sequence data on some exoenzymegenes indicate that several may be transcribed from a43-typepromoters (17, 35, 39), while others, such as some protease
genes, may be or are transcribed from promoters recognizedby some of the minor a factors (56, 61, 62). It can bespeculated that the mechanism regulating the temporal tran-scription of ,-1,4-glucanase in B. subtilis DLG may becommon to other exoenzyme genes of Bacillus origin whichseem to be transcribed from a43-type promoters, yet areexpressed only after vegetative growth. Clearly, more DNAsequence data and transcription initiation site mapping ofsimilar genes are needed to corroborate this possibility.Site-directed mutagenesis studies of exoenzyme gene pro-moter regions may help to define regions responsive to atemporal regulatory control.Another interesting aspect of ,B-1,4-glucanase regulation in
B. subtilis DLG is the possibility of induction by glucose. Inthis study, in vivo P-1,4-glucanase mRNA was observedonly when cells were grown in nutrient sporulation mediumplus glucose; almost no gene-specific mRNA was observedin cells grown on nutrient sporulation medium alone. Ascellulolytic enzyme systems of other organisms are generallyrepressed by glucose (12, 30, 53, 55), the effect observedhere is quite unusual. Thayer (58) has also reported in-creased ,B-1,4-glucanase activity by B. cereus in the presenceof glucose. It remains to be determined whether glucoseitself acts as an inducer of enzyme synthesis or whether thestimulatory effect on P-1,4-glucanase production by glucoseis more indirect.
Finally, it was shown that purified P--1,4-glucanase from B.subtilis DLG displayed almost twice the specific activity ofT. reesei EGI for degradation of TNP-CMC. However, thedifference may be greater, because we observed a decreaseof ca. 20% in the specific activity of the B. subtilis DLGP-1,4-glucanase preparation between its use in this assay andin a previous one. Crude P-1,4-glucanase preparations ap-pear relatively stable to several cycles of freezing andthawing, but purified and dilute P-1,4-glucanase preparationsmay not be as resistant. The enzyme will renature afterdenaturation by acetonitrile, isopropanol, and SDS (data notshown).The nature of B. subtilis DLG P-1,4-glucanase as an
endo-acting enzyme was confirmed in a separate assay,which measured the ability of the enzyme to reduce theviscosity of a CMC solution (R. Brown, Jr., and M. Gritzali,personal communication). Therefore, the P-1,4-glucanaseproduced by B. subtilis DLG can be referred to as anendo-,B-1,4-glucanase (EC 3.2.1.4). Interestingly, T. reeseiEGI showed the same specific activity as B. subtilis DLGendo-P-1,4-glucanase in this assay (Brown and Gritzali,personal communication). This illustrates the inadequacy ofusing only one assay system to compare the specific activityof two cellulases.At any rate, it appears that B. subtilis DLG endo-P-1,4-
glucanase is at least as active a cellulase as T. reesei EGI.Further studies are needed to show whether the B. subtilisDLG enzyme can work synergistically with exo-P-1,4-glucanases to effect the degradation of crystalline cellulose.
ACKNOWLEDGMENTS
We express our sincere appreciation to R. Brown, Jr., and M.Gritzali for providing us with purified T. reesei QM9414 endo-P-1,4-glucanase and for their other contributions to this study. We alsothank R. Niece and W. Strickland of the University ofWisconsin-Madison Biotechnology Resource Center for performingthe protein sequence analysis and for aid in the enzyme purificationprocess, respectively.
This research was supported by the College of Agricultural andLife Sciences, University of Wisconsin, Madison, Hatch grant no.
VOL. 169, 1987
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

2024 ROBSON AND CHAMBLISS
2603, and by Public Health Service grant no. GM34324 from theNational Institutes of Health. L.M.R. was supported on a NationalInstitutes of Health predoctoral training grant in cellular and molec-ular biology, fund no. 2 T32 GM07215.
LITERATURE CITED1. Berk, A. J., and P. A. Sharp. 1977. Sizing and mapping of early
adenovirus mRNAs by gel electrophoresis of S1 endonuclease-digested hybrids. Cell 12:721-732.
2. Birnboim, H. C., and J. Doly. 1979. A rapid alkaline extractionprocedure for screening recombinant plasmid DNA. NucleicAcids Res. 7:1513-1523.
3. Blobel, G., and B. Dobberstein. 1975. Transfer of proteins acrossmembranes. J. Cell Biol. 67:835-851.
4. Boylan, R. J., N. H. Mendelson, D. Brooks, and F. E. Young.1972. Regulation of the bacterial cell wall: analysis of a mutantof Bacillus subtilis defective in biosynthesis of teichoic acid. J.Bacteriol. 110:281-290.
5. Cantweli, B., G. Brazil, J. Hurley, and D. McConnell. 1985.Expression of the gene for the endo-13-1,3-1,4-glucanase fromBacillus subtilis in Saccharomyces cerevisiae, p. 259-266. InProceedings of the 20th European Brewing Convention Con-gress. IRL Press Ltd., Oxford.
6. Cantwell, B. A., and D. J. McConnell. 1983. Molecular cloningand expression of a Bacillus subtilis 3-glucanase gene in Esch-erichia coli. Gene 23:211-219.
7. Cathala, G., J.-F. Savouret, B. Mendez, B. L. West, M. Karin,J. A. Martial, and J. D. Baxter. 1983. A method for isolation ofintact translationally active ribonucleic acid. DNA 2:329-335.
8. Chambliss, G. H., T. M. Henkin, and J. M. Levanthal. 1983.Bacterial in vitro protein synthesizing systems. MethodsEnzymol. 101:598-605.
9. Dale, R. M. K., B. A. McClure, and J. P. Houchins. 1985. Arapid single-stranded cloning strategy for producing a sequentialseries of overlapping clones for use in DNA sequencing: appli-cation to sequencing the corn mitochondrial 18S rDNA. Plasmid13:31-40.
10. Doi, R. H. 1982. Multiple RNA polymerase holoenzymes exerttranscriptional specificity in Bacillus subtilis. Arch. Biochem.Biophys. 214:772-781.
11. Enari, T.-M.~ and P. H. Markkanen. 1975. Microbial -glucanases in brewing. Proc. Am. Soc. Brew. Chem. 33:13-17.
12. Fennington, G., D. Neubauer, and F. Stutzenberger. 1984.Cellulase biosynthesis in a catabolite repression-resistant mu-tant of Thermomonospora curvata. Appl. Environ. Microbiol.47:201-204.
13. Fogarty, W. M., and P. J. Griffin. 1973. Some preliminaryobservations on the production and properties of a cellulolyticenzyme elaborated by Bacillus polymyxa. Biochem. Soc. Trans.I:1297-1298.
14. Fouet, A., E. Aubert, M. Arnaud, A. Kler, G. Rapaport, and D.Le Coq. 1984. The sucrose system as a model of geneticregulation in Bacillus subtilis, p. 113-126. In J. A. Hoch andA. T. Ganesan (ed.), Genetics and biotechnology of the bacilli.Academic Press, Inc., New York.
15. Fukasaki, E., W. Panbangred, A. Shinmyo, and H. Okada. 1984.The complete nucleotide sequence of the xylanase gene (xynA)of Bacillus pumilus. FEBS Lett. 171:197-201.
16. Gilman, M. Z., and M. J. Chamberlin. 1983. Developmental andgenetic regulation of the Bacillus subtilis genes transcribed bysigma-28 RNA polymerase. Cell 35:285-293.
17. Gray, G. L., S. E. Mainzer, M. W. Rey, M. H. Lamsa, K. L.Kindle, C. Carmona, and C. Requadt. 1986. Structural genesencoding the thermophilic a-amylases of Bacillus stearothermo-philus and Bacillus licheniformis. J. Bacteriol. 166:635-643.
18. Hager, P. W., and J. C. Rabinowitz. 1985. Translational speci-ficity in Bacillus subtilis, p. 1-32. In D. A. Dubnau (ed.), Therholecular biology of the bacilli, vol. II. Academic Press, Inc.,New York.
19. Haldenwang, W. G., and R. L. Losick. 1980. Novel RNApolymerase cr factor from Bacillus subtilis. Proc. Natl. Acad.Sci. USA 77:7000-7004.
20. Hinchliffe, E. 1984. Cloning and expression of a Bacillus subtilis
endo-1,3-1,4-13-D-glucanase gene in Escherichia coli K12. J.Gen. Microbiol. 130:1285-1291.
21. Horikoshi, K., M. Nakao, Y. Kurono, and N. Sashihara. 1984.Cellulases of an alkalophilic Bacillus strain isolated from soil.Can. J. Microbiol. 30:774-779.
22. Huang, J. S., and J. Tang. 1976. Sensitive assay for cellulaseand dextranase. Anal. Biochem. 73:369-377.
23. Hunkapiller, M. W., R. M. Hewick, W. J. Dryer, and L. E.Hood. 1983. High-sensitivity sequencing with a gas-phasesequenator. Methods Enzymol. 91:399-413.
24. Johnson, W. C., C. P. Moran, Jr., and R. Losick. 1983. TwoRNA polyrnerase sigma factors from Bacillus subtilis discrimi-nate between overlapping promoters for a developmentallyregulated gene. Nature (London) 302:800-804.
25. Kudo, T., J. Yoshitake, C. Kato, R. Usami, and K. Horikoshi.1985. Cloning of a developmentally regulated element fromalkalophilic Bacillus subtilis DNA. J. Bacteriol. i61:158-163.
26. Kyte, J., and R. F. Doolittle. 1982. A simple method fordisplaying the hydropathic character of a protein. J. Mol. Biol.157:105-132.
27. Laemmll, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
28. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193:265-275.
29. Mandel, M., and A. Higa. 1970. Calcium dependent bacterio-phage DNA infection. J. Mol. Biol. 53:159-162.
30. Mandels, M. 1982. Cellulases, p. 35-78. In G. T. Tsao (ed.),Annual reports on fermenitation processes, vol. 5. AcademicPress, Inc., New York.
31. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
32. Martin, D. F., F. G. Priest, C. Todd, and M. Goodfellow. 1980.Distribution of 3-glucanases within the genus Bacillus. Appl.Environ. Microbiol. 40:1136-1138.
33. M6zes, P. S. F., and J. 0. Lampen. 1985. Secretion of proteinsby Bacilli, p. 151-183. In D. A. Dubnau (ed.), The molecularbiology of the bacilli, vol. II. Academic Press, Inc., New York.
34. Moran, C. P., Jr., N. Lang, S. F. J. LeGrice, G. Lee, M.Stephens, A. L. Sonenshein, J. Pero, and R. Losick. 1982.Nucleotide sequences that signal the initiation of transcriptionand translation in Bacillus subtilis. Mol. Gen. Genet. 186:339-346.
35. Murphy, N., D. J. McConnell, and B. A. Cantwell. 1984. TheDNA sequence of the gene and genetic control sites for theexcreted B. subtilis enzyme 1-glucanase. Nucleic Acids Res.12:5355-5367.
36. Murray, C. L., and J. C. Rabinowitz. 1982. Nucleotide se-quences of transcription and translation initiation regions inBacillus phage 4)29 early genes. J. Biol. Chem. 257:1053-1062.
37. Ogasawara, N. 1985. Markedly unbiased codon usage in Bacillussubtilis. Gene 40:145-150.
38. Ostroff, G. R., and J. J. Pene. 1983. Molecular cloning withbifunctional plasmid vectors in Bacillus subtilis: isolation of aspontaneous mutant of Bacillus subtilis with enhancedtransformability for Escherichia coli-propagated chimeric plas-mid DNA. J. Bacteriol. 156:934-936.
39. Palva, I., M. Sarvas, P. Lehtovaara, M. Sibakov, and L.Kaariainen. 1982. Secretion of Escherichia coli ,B-lactamasefrom Bacillus subtilis by the aid of a-amylase signal sequence.Proc. Natl. Acad. Sci. USA 79:5582-5586.
40. Perhnan, D., and H. 0. Halvorson. 1983. A putative signalpeptidase recognition site and sequence in eukaryotic andprokaryotic signal peptides. J. Mol. Biol. 167:391-409.
41. Priest, F. G. 1977. Extracellular enzyme synthesis in the genusBacillus. Bacteriol. Rev. 41:711-753.
42. Radloff, R., W. Bauer, and J. Vinograd. 1967. A dye-buoyant-density method for the detection and isolation of closed circularduplex DNA in HeLa cells. Proc. Natl. Acad. Sci. USA57:1514-1521.
J. BACTERIOL.
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

ENDO-P-1,4-;i ( AN Lj'; ( t. ';; OF B. SUBTILIS DLG 2025
43. Rahr, S., R. Halter, H. Muller, J. Pohlner, and T. F. Meyer.1985. Genetic analysis of neisserial immunoglobulin Aproteases, p. 157-163. In G. Schoolnik (ed.), The pathogenicneisseria. Proceedings of the 4th International Symposium.American Society for Microbiology, Washington, D.C.
44. Robson, L. M., and G. H. Chambliss. 1984. Characterization ofthe cellulolytic activity of a Bacillus isolate. Appl. Environ.Microbiol. 47:1039-1046.
45. Robson, L. M., and G. H. Chambliss. 1986. Cloning of theBacillus subtilis DLG ,-1,4-glucanase gene in Escherichia coliand B. subtilis. J. Bacteriol. 165:612-619.
46. Roncero, M. I. G. 1983. Genes controlling xylan utilization byBacillus subtilis. J. Bacteriol. 156:257-263.
47. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
48. Sashihara, N., T. Kudo, and K. Horikoshi. 1984. Molecularcloning and expression of cellulase genes of alkalophilic Bacillussp. strain N-4 in Escherichia coli. J. Bacteriol. 158:503-506.
49. Sathyamoorthy, V., and B. R. DasGupta. 1985. Partial aminoacid sequences of the heavy and light chains of botulinumneurotoxin type E. Biochem. Biophys. Res. Commun.127:768-772.
50. Schaeffer, P., J. Millet, and J. P. Aubert. 1965. Cataboliterepression of bacterial sporulation. Proc. Natl. Acad. Sci. USA54:704-711.
51. Shine, J., and L. Dalgarno. 1974. The 3' terminal sequence of E.coli 16S ribosomal RNA complementary to nonsense tripletsand ribosome-binding sites. Proc. Natl. Acad. Sci. USA71:1342-1346.
52. Stephens, M. A., S. A. Ortlepp, J. F. OWliigton, and D. J.McConnell. 1984. Nucleotide sequence of the 5' region of theBacillus licheniformis a-amylase gene: comparison with the B.amyloliquefaciens gene. J. Bacteriol. 158:369-372.
53. Stoppock, W., P. Rapp, and F. Wagner. 1982. Formation,location, and regulation of endo-1,4-,-glucanases and P-glucosidases from Cellulomonas uda. Appl. Environ. Micro-biol. 44:44-53.
54. StruhA. !', , and R. W. Davis. 1976. Functionalgenetic* t, ' c".'ii;yotic DNA in Escherichia coli. Proc.Natl. A,: -A >471-1475.
55. Stutzen')rr'; tegulation of cellulolytic activity, p.111-15 Pi '-; (ed.), Annual reports on fermentationprocess (temic Press, Inc., New York.
56. Takagi, ii. Aicka, and S. Aiba. 1985., Nucleotide se-quence a,iix 's egion for the neutral protease gene fromBacillus si iu,,philus. J. Bacteriol. 163:824-831.
57. Tanaka, H., r ;Y.ra, and K. Toda. 1978. Concerted inductionof 0-glucana f Bacillus circulans WL12 in response tovarious yeast L ;ans. Agric. Biol. Chem. 42:1631-1636.
58. Thayer, D. W. 1978. Carboxymethyl cellulase produced byfacultative bacteria from the hindgut of the termite Reticu-litermes herperus. J. Gen. Microbiol. 106:13-18.
59. Tinoco, I., P. N. Borer, B. Dengler, M. D. Levine, 0. C.Uhlenbeck, D. M. Crothers, and J. Gralla. 1973. Improvedestimation of secondary structure in ribonucleic acids. Nature(London) New Biol. 246:40-41.
60. Trempy, J. E., J. Morrison-Plummer, and W. G. Haldenwang.1985. Synthesis of oll, an RNA polymerase specificity determi-nant, is a developmentally regulated event in Bacillus subtilis. J.Bacteriol. 161:340-346.
61. Vasantha, N., L. D. Thompson, C. Rhodes, C. Banner, J. Nagle,and D. Filpula. 1984. Genes for alkaline protease and neutralprotease from Bacillus amyloliquefaciens contain a large openreading frame between the regions coding for signal sequenceand mature protein. J. Bacteriol. 159:811-819.
62. Wong, S.-L., C. W. Price, D. S. Goldfarb, and R. H. Doi. 1984.The subtilisin E gene of Bacillus subtilis is transcribed from a37 promoter in vivo. Proc. Natl. Acad. Sci. USA 81:1184-1188.
63. Yang, M., A. Galizzi, and D. Henner. 1983. Nucleotide sequenceof the amylase gene from Bacillus subtilis. Nucleic Acids Res.11:237-249.
64. Yanish-Perron, C., J. Vieira, and J. Messing. 1985. ImprovedM13 phage cloning vectors and host strains: nucleotide se-quences of the M13mpl8 and pUC19 vectors. Gene 23:103-119.
VOL. 169, 1987
on January 21, 2019 by guesthttp://jb.asm
.org/D
ownloaded from