essentials for atp synthesis by f1f0 atp synthases€¦ · essentials for atp synthesis by f 1f 0...
TRANSCRIPT

ANRV378-BI78-22 ARI 29 April 2009 18:5
Essentials for ATP Synthesisby F1F0 ATP SynthasesChristoph von Ballmoos,1 Alexander Wiedenmann,2
and Peter Dimroth3
1Department of Biochemistry and Biophysics, Arrhenius Laboratories for Natural Sciences,Stockholm University, SE-106 91 Stockholm, Sweden; email: [email protected] of Organic Chemistry, University of Zurich, CH-8057 Zurich, Switzerland;email: [email protected] of Microbiology, ETH Zurich, CH-8093 Zurich, Switzerland;email: [email protected]
Annu. Rev. Biochem. 2009. 78:649–72
The Annual Review of Biochemistry is online atbiochem.annualreviews.org
This article’s doi:10.1146/annurev.biochem.78.081307.104803
Copyright c© 2009 by Annual Reviews.All rights reserved
0066-4154/09/0707-0649$20.00
Key Words
ATP synthase assembly, coupling ion coordination, driving forces,torque generation
AbstractThe majority of cellular energy in the form of adenosine triphosphate(ATP) is synthesized by the ubiquitous F1F0 ATP synthase. Power forATP synthesis derives from an electrochemical proton (or Na+) gra-dient, which drives rotation of membranous F0 motor components.Efficient rotation not only requires a significant driving force (�μH+),consisting of membrane potential (�ψ) and proton concentration gradi-ent (�pH), but also a high proton concentration at the source P side. Invivo this is maintained by dynamic proton movements across and alongthe surface of the membrane. The torque-generating unit consists ofthe interface of the rotating c ring and the stator a subunit. Ion translo-cation through this unit involves a sophisticated interplay between thec-ring binding sites, the stator arginine, and the coupling ions on bothsides of the membrane. c-ring rotation is transmitted to the eccentricshaft γ-subunit to elicit conformational changes in the catalytic sites ofF1, leading to ATP synthesis.
649
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
ATP synthase: theenzyme synthesizingATP from ADP andphosphate
Mitochondria:membrane-enclosedorganelles ofeucaryotic cells, whichproduce most of theATP used as achemical energysource
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . 650Coupling ATP Synthesis to Ion
Cycling Across Membranes . . . . . . 650ATP Synthase Structure and
Function . . . . . . . . . . . . . . . . . . . . . . . 650THE F0 MOTOR . . . . . . . . . . . . . . . . . . . . 653
Assembly of the Rotor Complex . . . . 654c-Ring Stoichiometries . . . . . . . . . . . . . 655Structure of the Rotor Ring . . . . . . . . 656Ion Binding in Na+ and H+
ATP Synthases . . . . . . . . . . . . . . . . . . 656The Ion Path Through the
F0 Motor . . . . . . . . . . . . . . . . . . . . . . . 658Driving Forces for Na+- or
H+-Coupled F0 Motors . . . . . . . . . 662
INTRODUCTION
To support life, cells must be continuously sup-plied with external energy in the form of lightor nutrients and must be equipped with chem-ical devices to convert these external energysources into adenosine triphosphate (ATP).ATP is the universal energy currency of livingcells and as such is used to drive numer-ous energy-consuming reactions, e.g., biosyn-theses, mechanical motility, transport throughmembranes, regulatory networks, and nerveconduction. When performing work, ATP isusually converted to ADP and phosphate. Itmust therefore continuously be regeneratedfrom these compounds to continue the cell en-ergy cycle. The importance of this cycle canbe best illustrated by the demand for a dailyturnover of 50 kg of ATP in a human body onaverage. The majority of this ATP is synthe-sized by the ubiquitous F1F0 ATP synthase withenergy that has been supplied by the externalsources mentioned above.
Coupling ATP Synthesis to IonCycling Across Membranes
ATP synthases utilize the movement of pro-tons or Na+ ions across the membrane as they
flow down their electrochemical gradients todrive ATP synthesis. Conversely, enzymes act-ing as active proton or Na+ pumps must op-erate in tandem with the ATP synthase tore-energize the membrane (Figure 1). Exam-ples for the active transport of protons arethe respiratory-chain enzymes of mitochondriaand aerobic bacteria or the light-driven pro-ton pumps from plants or phototrophic bac-teria (1). A similar but Na+ ion-cycle-basedATP synthesis mechanism has been found insome fermenting bacteria. A prominent exam-ple is Propionigenium modestum, which growsby the fermentation of succinate to propionateand CO2. During metabolism, succinate is con-verted to methylmalonyl-coenzyme A (CoA),which is decarboxylated to propionyl-CoA by amembrane-bound decarboxylase coupling theexergonic decarboxylation to electrogenic Na+
transport. The electrochemical gradient ofNa+ ions generated subsequently drives ATPsynthesis by a Na+-translocating F1F0 ATPsynthase. It has been calculated that thefree energy of the decarboxylation reaction isapproximately −20 kJ/mol and that for ATPsynthesis it is approximately 60 kJ/mol in agrowing bacterium. Hence, three to four con-secutive Na+ export cycles, driven by decar-boxylation events, are required to accumulateenough energy for the synthesis of one ATP inthe Na+ import cycle. It has been found that oneNa+ ion is pumped per decarboxylation eventacross the membrane and that 3.7 Na+ ions flowbackward through the ATP synthase during thesynthesis of one molecule of ATP. Hence, thestoichiometries of the two Na+ pumps appearto be just right to balance the bioenergeticdemands (2, 3).
ATP Synthase Structure and Function
The F1F0 ATP synthase is a miniature en-gine composed of two opposing rotary motors,which are connected in series as described sep-arately below (Figure 2). The F1 motor is awater-soluble protein complex with the sub-unit composition α3β3γδε that catalyses ATPsynthesis or hydrolysis by a rotary mechanism.
650 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
Methylmalonyl-CoA
Propionyl-CoA + CO2
ADP + Pi
ATP O2
H2O
Δψ
Na+H+H+
H+/ Na+
Respiration in mitochondria
and many bacteria
Photosynthetic reactions in chloroplasts and
phototrophic bacteria
Decarboxylation reactions in some anaerobic bacteria
ATP synthesis by F-type ATP synthases in almost
all organisms
NADH
NAD+
H+/ Na+
P side
N side
Figure 1Ion cycling across biological membranes leading to ATP synthesis. In heterotrophic organisms, e.g., animals or bacteria, themetabolism of nutrients generates reducing equivalents (NADH, succinate). These are oxidized by the respiratory-chain enzymes,using oxygen as terminal electron acceptor. The free energy of the oxidation reactions is converted into an electrochemical gradient ofprotons (�μH+) across the membrane. Chloroplasts and phototrophic bacteria convert light energy into a �μH+ across themembrane. The anaerobic bacterium P. modestum couples the decarboxylation of methylmalonyl-CoA to electrogenic Na+ transport,which results in an electrochemical sodium gradient. The electrochemical H+ or Na+ gradients established by these membrane-boundcomplexes serve as energy sources for ATP synthesis from ADP and inorganic phosphate by an F1F0 ATP synthase. Abbreviations: Nside, the negatively charged side of a membrane; P side, the positively charged side of a membrane.
The F0 motor is membrane embedded and inits simplest bacterial form has the subunit com-position ab2c10−15. It catalyses ion translocationacross the membrane that is coupled to rotationof the c ring versus the stator subunits a andb2. The two motors are physically connectedby two stalks, a central one containing theγ- and ε-subunits and a peripheral one involv-ing the δ- and b-subunits. Mammals contain ad-ditional subunits, which are mostly in the stalkregion. Under mild dissociation conditions, theF1F0 complex disintegrates into the F1 and F0
complexes, which can be isolated and studiedindividually.
The crystal structure of F1 from bovineheart mitochondria revealed a hexameric
assembly of alternating α- and β-subunitsaround an eccentric α-helical coiled-coil γ-subunit (4). The catalytic nucleotide-bindingsites are located on each of the three β-subunitsat the interface with the α-subunits. As thesesites are in different nucleotide-bound statesduring catalysis, each β-subunit adopts a differ-ent conformation, causing a diverse interactionwith the central γ-subunit. On rotation of theγ-subunit, the conformations of the β-subunitschange sequentially such that each conforma-tion of varying nucleotide-binding affinity isadopted by each site during one rotationalcycle. Hence, the rotational model can ex-plain the binding change mechanism that waspredicted from numerous biochemical studies
www.annualreviews.org • Essentials of F1F0 ATP Synthases 651
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
α3β3
γε
δ
F1
c10-15
F0
b2
a
Cytoplasm
Periplasm
Figure 2The structural organization of an F1F0 ATP synthase. The structures ofindividual subcomplexes were taken from RCSB Protein Data Bank(http://www.rcsb.org/pdb/static.do?p=general information/web links/databases.html) and assembled by eye according to biochemical data. Thestructures used were from the c ring of Ilyobacter tartaricus (1CYE), the F1organization of Escherichia coli (1JNV), the δ-subunit of E. coli (2A7U), theperipheral stalk from bovine mitochondria (2CLY), and the membranous partof subunit b of E. coli (1B9U). No high-resolution structural data are availablefor subunit a and the hinge region of subunit b. The illustration was createdusing PyMOL (DeLano Scientific LLC).
(Reference 5 and references therein). The ro-tational catalysis was experimentally confirmedby a variety of biochemical and spectroscopictechniques. Most convincingly, the rotation ofa micrometer-sized fluorescent actin filamentattached to the central γ-subunit has been di-rectly visualized by video microscopy of singleF1 molecules (6). Rotation was similarly ob-served when the actin filament was attached tosubunit ε or to the c10−15 oligomer of F0 (7).
These studies are consistent with several cross-linking experiments, which have been impor-tant to define subunits γ, ε, and c10−15 as therotor and subunits α3, β3, δ, a, and b2 as thestator (8–11). A physical connection betweenthe c10 ring and the γ- and ε-subunits is simi-larly seen in the electron density map of an F1
complex from yeast with an attached c10 ring(12).
Single-molecule experiments with smallmagnetic or gold beads or with fluorescent dyesinstead of the large actin filament provided in-sights into mechanistic details of the enzyme.Rotation in the fully coupled F1F0 Na+ ATPsynthase was observed in both synthesis andhydrolysis mode (13). The rotor turns coun-terclockwise in the ATP hydrolysis directionand clockwise in the ATP synthesis direction,when viewed from the F0 domain, and per-forms up to 700 revolutions per second (14,15). In F1, the γ-subunit rotates in steps of 120◦
for each ATP molecule hydrolyzed (Figure 3).Each 120◦ step can be further divided into fourstages (Reference 16 and references therein). Inthe ATP-binding dwell, an ATP molecule bindsto the empty β1-site and elicits a rapid 80◦ sub-step of the γ-subunit rotation. In the followingcatalytic dwell, ATP is thought to be cleavedat the β2-site, and ADP and phosphate are re-leased from the β3-site (17). This initiates a 40◦
substep of the γ-subunit, completing its 120◦
rotation. The rotational behavior of F1F0 re-sembles that of F1, indicating that friction inthe F0 motor is negligible during ATP-drivenrotation (18). Tributyltin chloride, a specific in-hibitor of ion access through subunit a, inhibitsrotation by 96% in accordance with strictcoupling between mechanical and ion translo-cation events (15). When the performance ofthe F1 motor was probed by sophisticatedsingle-molecule experiments in femtoliter-sized chambers, the hydrolysis of three ATPmolecules per revolution was directly observedand showed very high mechanochemical cou-pling (19). If the γ-subunit of F1 was forcedto rotate in the ATP synthesis direction, ATPwas synthesized from ADP and phosphate. Themechanochemical coupling efficiency was low
652 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
ATP
ATP
ADPPi
ADPPi
ADP
Pi
ATP ATP ATP
ATP
ATP
ADPPi
ADPPi
ATP
80°
120°
200°
240°
320°
0°
0°
80°
120°
200°
240°
320°
0°
80°
120°
200°
240°
320°
0°
80°
120°
200°
240°
320°
0°
80°
120°
200°
240°
320°
Pi
I
II III IV
V0°
80°
120°
200°
240°
320°
β1
β2
β3
β1
β2
β3
β1
β2
β3
β1
β2
β3
ATP-binding dwell
ATP hydrolysis/Pi release dwell80° rotation
40° rotation
β1
β2
β3
Azide-inhibited structure of bovine F1
0° 24
320°
AMP-PNP
Pi
ADP
Power unitResting unit
Figure 3Model of the binding change mechanism during ATP hydrolysis (adapted from Reference 99). The F1 part is seen from the membraneand is divided in β1, β2, and β3, representing the α/β interface of each catalytic unit. The pink unit is thought to generate the powerstroke responsible for γ-rotation, and the resting units are shown in blue. The rotating, central γ-subunit is shown in orange. In states Iand V, when F1 is waiting for ATP, it is assumed that the catalytic β1, β2, and β3 sites are empty or contain ATP and ADP/Pi,respectively. The angular position of subunit γ in state I is set to be 0◦. (I–II) ATP-binding dwell: ATP is bound to the empty catalyticβ1 site. (II–III) A 80◦ rotation: ATP binding initiates rotation of γ by 80◦, accompanied by ADP release from β3. (III–IV) ATPhydrolysis/Pi-release dwell: ATP is cleaved into ADP and Pi at β2, and Pi is released from β3. The sequence of events is not yetdetermined. (IV–V) A 40◦ rotation: The subsequent rotation completes one 120◦ step, yielding an empty β3 site, which is ready forATP binding (similar to β1 in I). The catalytic turnover of a particular ATP molecule needs a 360◦ rotation of the γ-subunit, with ATPbinding to β1 at 0◦, ATP cleavage at 200◦, ADP-release at 240◦–320◦, and Pi release at 320◦. Also shown is the azide-inhibitedstructure of bovine F1 as seen from the membrane, with AMP-PNP (cyan), ADP (magenta), and Pi ( green) in the respectivecatalytic-binding sites, which somewhat represent state III (100).
for an F1 subcomplex lacking the ε-subunitbut reached more than 70% after reconstitu-tion with ε (18). The ε-subunit thus has animportant role in the synthesis of ATP, but themechanism for this function has not yet beenelucidated.
THE F0 MOTOR
The bacterial F0 motor is a membrane-embedded protein complex consisting of anoligomeric ring of c subunits, a single a sub-unit, and a dimer of b subunits, which flankthe c ring laterally. Each c subunit consists
www.annualreviews.org • Essentials of F1F0 ATP Synthases 653
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
YidC: a bacterialmembrane proteinassisting in theintegration of newlysynthesized proteinsinto the bacterialmembrane
of two membrane-spanning α-helices that areconnected by a hydrophilic loop at the cyto-plasmic side of the membrane. In the c ring,the loop regions form an extensive area ofcontact with the foot of the central stalk sub-units γ and ε (12). The a subunit is a very hy-drophobic protein that in most models is com-posed of five transmembrane helices (20, 21).Ion translocation takes place through subunita and its interface with subunit c. Torque gen-erated during this process is conferred to theγ-subunit to drive ATP synthesis in F1. Theb subunits are anchored within the membraneby an N-terminal α-helix and extend as the pe-ripheral stalk all the way to the head of the F1
domain (22). According to cross-linking stud-ies, the b subunits contact the C-terminal partof the c subunit and the loop between helices4 and 5 of the a subunit at the periplasmicsurface. In the solvent-exposed part of the bsubunit, the tether, dimerization, and δ-subunitinteraction domains have been defined (23).The δ-subunit forms a strong complex withthe latter domain and the α-subunit (24, 25).The connection of F1 and F0 by the periph-eral stalk is an important device to counter thetendency of the stator to follow the rotationof the rotor. In mitochondria, the peripheralstalk consists of more subunits (named OSCP,b, δ, and F6), and its high-resolution X-raystructure indicates that it forms a rigid com-plex in accord with the suggested function tocounteract the torque created through the rotor(26).
Assembly of the Rotor Complex
To date, there is only little information on theassembly of the ATP synthase or the subcom-plexes of this enzyme (27). It is remarkable,however, that a typical bacterial ATP synthaseconsists of eight different subunits, whereasnine open reading frames are present in the re-spective atp (or unc) operon with the uncI genepreceding the structural genes for the eight sub-units of the ATP synthase. The uncI gene ofEscherichia coli encodes a membrane protein of14 kDa that is not a component of the ATP syn-
thase. The protein copurifies with the enzymein substoichiometric amounts and might thushave a functional role in the assembly of theenzyme. Disruption of the uncI gene leads todiminished growth yields without significantlyaffecting the function of the ATP synthase andthus could not provide conclusive evidence foruncI gene function (28, 29). More recently, stud-ies with the UncI protein from P. modestumshowed that it plays a chaperone-like role inthe assembly of the c11 ring of its ATP synthase(30). When expressed in E. coli, UncI of P. mod-estum was required for the formation of the fullyassembled c11 rings, and in its absence, neitherc11 nor c1 protein products are observed, indi-cating degradation of the c subunit under theexperimental conditions employed. With an-other expression system, containing plasmidswith the corresponding c subunit genes undercontrol of the strong T7 promotor, however,small amounts of correctly assembled c11 ringsfrom P. modestum or Ilyobacter tartaricus weresynthesized in E. coli (31). As uncI from therespective organism was not present in theseexperiments, either the UncI protein fromE. coli or another protein must have taken therole to direct the correct assembly of these crings. These results are not unexpected giventhe formation of catalytically active E. coli ATPsynthase in the uncI deletion background andargue for an overlapping function of anotherprotein with UncI in promoting the assembly ofthe c ring. Indeed, YidC has been shown to assistin the formation of the c ring from monomers(32). The chaperone-like function of UncI ofP. modestum was corroborated by an in vitro sys-tem, which showed the oligomerization of c1
into c11 upon UncI addition (33).The interfaces of the rotor subunits can be
inferred from the medium-resolution crystalstructure of the F1–c10-ring complex from yeast(12). In this structure, the cytoplasmic loop ofthe c ring forms the contact site with the footof the γ- and ε-subunits. The in vitro assemblyof the rotor complex has been demonstratedwith the isolated c11 ring and a preformedcomplex of subunits γ and ε from I. tartar-icus (34). Using surface plasmon resonance
654 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
spectroscopy, association and dissociation ki-netics were recorded. The data yielded a KD
of about 7 nM, indicating the formation of avery stable complex. The value is similar to thebinding affinity determined for the ab2F1 sta-tor complex of E. coli (35). The high intrin-sic stability of the rotor and the stator com-plexes appears to be necessary to maintain theenzyme’s integrity under operation conditions.Structurally, the assembly of the rotor is basedprimarily on attachment of the γ-subunit to thec ring because a separate complex between thec ring and the ε-subunit is not observed. Theassembly is severely affected by mutagenesis ofconserved amino acids in the contact region ofall three subunits (34). Interaction kinetics be-tween the γε-complex and the c-ring mutantssuggest that at least two steps are involved inthe assembly of the complex. Following low-affinity binding between the polar loops of thec ring and the bottom part of the γ-subunit, thecomplex is stabilized by a high-affinity interac-tion involving the c ring and residues γE204and εH38 (Figure 4).
c-Ring Stoichiometries
The c rings of several ATP synthases have beenisolated and their subunit stoichiometries de-termined by various methods, including atomicforce microscopy, cryoelectron microscopy oftwo-dimensional crystals, mass spectroscopy,and sodium dodecyl sulfate-gel electrophoresis(36, 37). The different methods produced con-sistent results on the c subunit stoichiometryof a particular c ring but yielded stoichiometryvalues between 10 and 15 for c rings from dif-ferent sources.
These stoichiometries have important con-sequences on the bioenergetics of the respec-tive ATP synthases because the number of csubunits in the ring equals the number of ionstranslocated per 360◦ rotation during whicheach site takes part in the translocation of oneion (see References 38 and 39 for additionaldiscussion). The F1 part of all ATP synthaseshas a threefold symmetry and synthesizes orhydrolyzes three molecules of ATP during a
γ
ε
c11
cR45/ Q46
εH38
εE29/ E31
γF203/ E204
Figure 4Assembly of the rotor complex. Shown are thestructures of the c ring of I. tartaricus (1CYE) andthe truncated γε-complex of E. coli (1FS0). Criticalresidues for interaction with the γε-complex are atthe inner end of the loop of the c ring. Residuesinvolved in the docking of the γε-complex to thetop of the c ring are indicated.
Chloroplasts: plantorganelles that uselight energy for theproduction of ATPand for photosynthesis
360◦ rotation. Consequently, the ion-to-ATPratio of an ATP synthase can be calculated ifthe number of c subunits is divided by three.The ATP synthase of yeast mitochondria andthe aerobic bacteria of Bacillus PS3 and of E. colihave c-ring sizes of 10 subunits, thus operatingat an H+/ATP ratio of 3.3, whereas the ATPsynthase of chloroplasts with a c14 ring has anH+/ATP ratio of 4.7, and the cyanobacteriumSpirulina platensis contains a c15 ring, whichleads to an H+/ATP ratio of 5 (Reference 39 andreferences therein). The different c-ring stoi-chiometries and the resulting H+/ATP ratiosmight be adaptations to the different environ-mental conditions of the ATP-generating cellsor organelles. A comparatively low H+/ATP ra-tio is more economical and can be employedby mitochondria or aerobic bacteria where aconstantly high �μH+ with the main contri-bution from the �ψ is maintained. The highH+/ATP ratio of chloroplasts does not reflect a
www.annualreviews.org • Essentials of F1F0 ATP Synthases 655
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
V-type ATPases(V-ATPase): generatean electrochemicalproton gradient at theexpense of ATP andare prevalent ineucaryotic vacuoles
significantly reduced overall driving force(�μH+). This, however, exists mainly as �pH,and to operate at a low �ψ might be a challengethat benefits from a larger c-ring stoichiome-try. Alternatively, economic energy conversionsmay be less mandatory if light serves as theprimary energy source. The chloroplast ATPsynthase can therefore operate more distantfrom thermodynamic equilibrium, compared tothe mitochondrial enzyme, with the benefit ofhigher turnover rates. Although the c-ring sizevaries in different organisms, it appears to beconstant in a given organism. It has been pro-posed that the c-ring size can be varied by nutri-tional or genetic manipulations (40). However,attempts to verify this idea have failed or wererevoked (41) and argue for sequence-intrinsicfeatures of each particular c subunit, which de-termine assembly into a specific oligomeric ring(31, 41, 42).
Direct biochemical measurements ofH+/ATP ratios yielded values ranging mostlybetween 2 and 4 (reviewed in References 40 and43). At thermodynamic equilibrium, a constantH+/ATP ratio of 4 has been determined forthe ATP synthases from chloroplasts (c14),E. coli (c10), and Thermus thermophilus (c12),in spite of their different c-ring stoichiome-tries (44, 45). This situation with very lowturnover rates (�1 ATP/s), however, does notreflect physiological conditions with high ATPsynthesis rates (>70–200 ATP/s). Additionalinvestigations are therefore necessary todetermine H+/ATP ratios at the physiologicalworking conditions of ATP synthases.
Structure of the Rotor Ring
The structure of the c11 oligomer from theNa+-translocating F-ATP synthase of I. tartar-icus was first solved by two-dimensional crys-tallography and then by X-ray crystallographyto 2.3-A resolution (46, 47). This was comple-mented by the structure of the K ring fromthe Na+-translocating V-type ATPase from En-terococcus hirae (48). The I. tartaricus c11 ringstructure shows an hourglass-shaped hollowcylinder whose periplasmic cavity is filled with
phospholipids in the natural environment of themembrane (49). Each c monomer folds into ahelical hairpin with the loop at the cytoplasmicside and the termini at the periplasmic side ofthe membrane. The N-terminal helices forma tightly packed inner ring with no space forlarge side chains, which is accounted for by aconserved motif of four glycine residues (47).In addition, the tight packing of the glycinescauses the inner ring of helices to narrow fromthe cytoplasm toward the middle of the mem-brane. The outer ring consists of 11 C-terminalhelices, which pack into the grooves of the in-ner ring in the cytoplasmic half of the protein.All helices show a bend of about 20◦ in themiddle of the membrane (at P28 and E65 inthe N-terminal and C-terminal helices, respec-tively), marking the narrowest part of the hour-glass shape. Eleven Na+ ions are located hereat their binding sites, facing toward the outersurface of the c ring, which is consistent withcross-linking data (50).
Ion Binding in Na+ and H+
ATP Synthases
In the I. tartaricus c-ring structure, each of the11 Na+ ions is bound at the interface of one N-terminal and two C-terminal helices (46). Thecoordination sphere is formed by side chainoxygen atoms of Q32 and E65 of one sub-unit, the hydroxyl oxygen atom of S66, andthe carbonyl oxygen atom of V63 of the neigh-boring subunit (Figure 5a). A fifth coordina-tion site found in the K ring from E. hirae,but not clearly visible in the structure of thec ring (48), may be responsible for the higherNa+-binding affinity of the K ring (51). Thecoordination sphere is completed by a networkof hydrogen bonds. The side chain oxygen ofE65, which contributes to Na+ coordination,receives a hydrogen bond from the NH2 groupof Q32, and the other oxygen of E65 receiveshydrogen bonds from the OH groups of S66and Y70. These hydrogen bonds serve to keepE65 deprotonated at physiological pH in or-der to allow Na+ binding. This creates a stablelocked conformation of the binding site, which
656 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
AY70
V63
AS66
BE65
BQ32
a b
pH5 6 7 8 9 10 11
0
10
20
30
40
50
60
I. tartaricus E. coliH. salinarium
% la
belin
g
Na+
Figure 5Ion binding coordination in c rings from Na+- and H+-translocating ATP synthases. (a) Close-up of theNa+-binding site formed by two neighboring c subunits (A and B). Na+ coordination and selected hydrogenbonds are indicated with dashed lines. This structure represents the locked conformation (see text fordetails). (b) pH dependence for the modification of c subunit binding sites by N,N′-dicyclohexylcarbodiimide.Depicted are representatives from each class of proton-translocating enzymes. The ATP synthase ofHalobacterium salinarium and of I. tartaricus (in the absence of Na+) show the expected profile for theprotonation of a carboxylate, whereas the profile of the E. coli ATP synthase might reflect the coordination ofa hydronium ion.
is observed in the X-ray structure, from whichhorizontal transfer of the Na+ ion to subunit a isprevented (52). In the subunit a/c interface, theconformation of the binding site must thereforeconvert to an open one to allow ion transfer be-tween the binding site and subunit a, see below.
Structural data for an H+-binding site in thec ring of an H+-translocating enzyme are notavailable, and therefore, the mode of protonbinding is difficult to envisage. In most mod-els of the H+-dependent enzymes, the con-served acidic c-ring residues in the middle of themembrane are assumed to switch between pro-tonated and deprotonated states to allow pro-ton translocation across the membrane (53–55).If these models were correct, the protonationstate of the c-ring sites should depend on thepH, following a titration curve where the inflec-tion point corresponds to the pKa of the acidicresidues. This idea has recently been challengedby investigating the protonation state of c-ring
a/c interface: theproteinous interfacebetween the statorsubunit a and therotating c ring of theATP synthase
DCCD: N,N ′-dicyclohexylcarbo-diimide (inhibitor ofthe F0 sector of ATPsynthase)
binding sites from several H+-translocatingenzymes (56).The protonation state has beenprobed by labeling experiments with the spe-cific inhibitor N,N ′-dicyclohexylcarbodiimide(DCCD), which is known to covalently mod-ify the binding site carboxylate by consump-tion of a proton. The labeling reaction there-fore depends on the proton availability at theappropriate sites. When the pH dependence ofDCCD labeling was followed in the ATP syn-thase of Halobacterium salinarium or I. tartaricusin its H+-translocation mode (in the absenceof Na+), a typical titration curve was observedwith maximum labeling rates at pH 6 and be-low, half-maximal labeling around pH 7, and nodetectable labeling at pH 8 and above. Hence,these enzymes show the expected behavior fora group protonation mechanism of the bindingsite carboxylate (Figure 5b).
Surprisingly, bell-shaped pH profiles forDCCD labeling were observed for the majority
www.annualreviews.org • Essentials of F1F0 ATP Synthases 657
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
ATPase: the enzymehydrolyzing ATP toADP and phosphate
of H+ ATP synthases, including enzymes fromE. coli, spinach chloroplasts, and bovine mi-tochondria. Maximum labeling rates betweenpH 7.5 and 9 were flanked by decreasing ratesat lower or higher pH values (Figure 5b). De-creasing rates with decreasing pH values are notconsistent, however, with group protonation ofthe acidic residues. Instead, the proton couldbe coordinated as a hydronium ion, as alreadyproposed by Boyer (57), in order to strengthenthe analogy of ion binding in Na+- and H+-translocating enzymes. Efficient coordinationof a hydronium ion requires acceptors for threehydrogen bonds, e.g., both oxygens of the c-ring carboxylate and the carbonyl oxygen of thebackbone near the kink induced by breaking thehelix through a conserved proline residue. Inthe I. tartaricus c ring, the carboxylate oxygensalready receive three hydrogen bonds from in-teracting donor groups (see above), which ef-fectively prevents hydronium ion binding.
In the ATPases with presumed hydroniumion binding, where a high proton binding affin-ity extends well into the alkaline region, theATP hydrolysis pH profile is also shifted towardalkaline pH values. Conversely, in the ATP syn-thases from P. modestum and H. salinarium, ATPhydrolysis is only possible in a narrow pH rangearound pH 6 (56).
The Ion Path Through the F0 Motor
The rotational mechanism of the ATP syn-thase demands ingeniously designed interfacesbetween rotor and stator subunits, particularlybetween the rotating c ring and the laterallyabutted subunit a, because rotation speeds upto 500 Hz must be tolerated in the absence of astabilizing rotor axis. This proteinous interfacealso acts as the critical scaffold for torque gen-eration and ion translocation across the mem-brane. To prohibit charge translocation withoutrotation, ion leakage at the interface must beefficiently prevented.
Electron microscopy studies have shownthat presumably more than one c-ring sub-unit is covered by the laterally abutted ab2 sub-unit complex (58, 59). Conversely, it is unlikely
that the ab2 subcomplex consisting of maximaleight-transmembrane helices masks more thantwo binding sites at the same time.
Stable subcomplexes of subunit a and the cring have been purified and shown to mediateion translocation after reconstitution with thetransmembrane part of subunit b (60). The pre-cise localization of the two b subunits within theF0 complex is unclear, but the purification ofa stable ab2 subcomplex (61) and site-directedcross-linking experiments ( J. Zingg Ebneter,P. Dimroth, & C. von Ballmoos, unpublishedresults) indicate that within the membrane-spanning region subunit b preferentially inter-acts with subunit a.
Plasmid-borne F1F0 ATP synthase lackingsubunit a is efficiently integrated into the mem-brane, proving that subunit a is not essentialfor assembly of the holoenzyme (62). Becauseonly decreased ATP hydrolysis activity wasobserved, which was greatly stimulated by theaddition of purified subunit a, the authors pro-posed that, in the absence of the a subunit,subunit b interacts with the c ring like an an-chor rail and prevents rotation of the γεc10−15
complex.Clearly, the a/c interface represents the most
intriguing part of the F0 motor. The lackof structural information makes it particularlychallenging to elucidate the molecular detailsof its major tasks as denoted below:
� positions of entry and exit sites for thecoupling ions, including access pathwaysto the c-ring binding site from either sideof the membrane,
� specific interactions between subunit aand the c ring to provide tightness againstunspecific ion transfer,
� almost frictionless rotation of the c ringagainst the stator to ensure high efficiencyand high turnover rates, and
� ingenious interplay between the statorcharge and the binding site to allowhorizontal torque generation by verticaldriving forces.
Critical residues in the a/c interface. Dur-ing the past 20 years, several hundred mutations
658 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
were introduced in the E. coli a subunit and char-acterized in various laboratories. Generally, theimpact of the mutation on the ability to syn-thesize ATP was tested by growth on succinate,and its impact on the ability to translocate pro-tons was tested by ATP-driven ACMA quench-ing or by passive proton flux through F0 in in-verted vesicles that were energized with NADHoxidation. Second-site suppressor mutation ex-periments, in which an inactive or poorly activeenzyme could be rescued by a secondary mu-tation, provided insights into the spatial ar-rangements and functional requirements of therespective residues.
In spite of the impressive coverage of the in-vestigations, only a very small subset of residueswas found to be essential for function of theATP synthase. The most conserved residue is amembrane-embedded arginine (position 210,E. coli numbering), also known as the statorcharge (63). So far, every mutation of thisresidue in the E. coli enzyme led to a completeloss of activity, whereas in the P. modestum en-zyme, mutations retaining the positive chargewere tolerated under specified conditions (64).In current models, the arginine is believed toensure release of the coupling ion from thebinding site on the c ring when it enters the a/cinterface and to provide a seal between the twoaccess pathways to either side of the membrane(64–66). Recent experiments assign the statorcharge an active role in generation of therotational torque, as discussed in detail below.An additional set of critical residues, includingG218, E219, and H245, is proposed to packclosely together in the upper third of the mem-brane at the periplasmic side (67, 68). Althoughonly a few single mutations are lethal (e.g.,E219 → L or C, 245H → S, C, L, Y), these areparticularly interesting because the lethal effectcan successfully be counterbalanced by second-site mutants in one of the two other positions.There appears to be a remarkable connectionbetween the amino acid composition of thistriad and the environmental pH of the organ-ism, suggesting that the triad acts as an ion trapfor protons from the surrounding environment(for further considerations, see Reference 39).
Extensive cysteine scanning analysis of sub-unit a and accessibility studies with cysteine re-active chemicals (NEM, Ag+, and others) wereapplied to indentify the two proposed aqueoushalf channels on either side of the membrane(69). Not surprisingly, the amino acids of thetriad, described above, were sensitive targetsand were thus concluded to be a major part ofthe access pathway from the periplasmic sideof the membrane to the c-ring binding site. Forthe access pathway from the cytoplasmic reser-voir, present models locate most of the reactiveamino acids in the vicinity of the membraneborder (69, 70). Furthermore, several lines ofevidence indicate an access route for the ionfrom the cytoplasm to the binding site in the ab-sence of subunit a (71, 72). Whether this accessis sufficiently fast to meet physiological condi-tions is presently unknown. Nevertheless, thedata seem to indicate that the c ring contributesat least partially to the ion pathways.
The stator charge induces a conformationalchange of the binding site residues in the a/cinterface. The structure of the c ring from I.tartaricus and K ring from E. hirae revealed theprecise mode of Na+ binding outside of the a/cinterface (46, 48). However, it has to be assumedthat the c ring adopts alternative conformationsunder the influence of the stator charge.
Fillingame and coworkers (73) found bydisulfide cross-linking between subunits a and cof the E. coli ATP synthase that helix 2 of subunitc and helix 4 of subunit a are facing each other.These data were later processed in the view ofNMR structures of the monomeric c subunit inorganic solvent mixtures at different pH values.The resulting mechanism for ion translocationthrough F0 involved large swiveling motions ofthe whole outer helix of subunit c by 150◦ (55,74). This view could not be corroborated inlater studies, including homology modeling ofthe E. coli c ring from the I. tartaricus structureand NMR structures of the c-monomer of theH+-translocating ATP synthase of Bacillus PS3in organic solvents over a broad pH range (pH2–8) (75). Although in the latter study no sig-nificant conformational changes were observed
www.annualreviews.org • Essentials of F1F0 ATP Synthases 659
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
at different pH values, the homology model ofthe E. coli c ring revealed that no large confor-mational changes are necessary to explain thebiochemical data because all residues on the cring, which formed disulfide bridges with sub-unit a, are facing outward (46). Fillingame andcoworkers (76) retreated from their swivelingmodel and proposed that such a twinned con-formation of the c subunit is possible but doesnot necessarily contribute to the mechanism ofion translocation.
Strong evidence for a small but significantchange in the c-ring binding site structure uponcontact with the stator charge of subunit awas recently obtained by intersubunit Cys-Cyscross-linking studies between subunit a and cof the I. tartaricus ATP synthase (77). The mostremarkable results reflect cross-linking exper-iments between residues cT67C or cG68Cand aN230C on subunit a. Although cT67Cfrom one c subunit and cG68C from a neigh-boring c subunit occupy spatially similar po-sitions in the c-ring crystal structure towarda potent binding partner on subunit a, onlycT67C, but not cG68C, was capable of effi-cient cross-linking with aN230C. Upon mu-tagenesis of the stator charge arginine by anuncharged amino acid (R226 →A, N, S, H),the cross-linking efficiency of cT67C/aN230Cremained unchanged but that of cG68C/aN230C increased significantly. Furthermore,increasing cross-link yields between cG68C/aN230C in the aR226H background were onlyfound at higher pH values, where the histidineis supposed to be in its neutral form. Thesefindings were rationalized by proposing a sta-tor charge-induced conformational change inthe empty binding site, whereby cG68C is dis-connected from its cross-linking partner in sub-unit a. Relating these data to the c-ring struc-ture suggests a rearrangement of the bind-ing site residues, with the side chain of cE65being slightly pushed or rotated toward thec-ring center, thereby affecting the accessibil-ity of cG68 (which is on the same subunit) butnot of cT67 (which is on the neighboring he-lix). The spatial demand for this rearrangementis presumably allocated by a cavity between the
inner and the outer rings of helices in the re-gion of the two conserved residues cG25 andcG68 (G27 and G66 in the K ring of E. hirae)(Figure 6a). Accordingly, replacement of cG25by a bulky isoleucine increased the cross-linkingefficiency of the cG68C/aN230C pair to asimilar extent as the replacement of the sta-tor charge. No ATP synthesis was observed inthe cG25I mutant, emphasizing the functionalimportance of the small residue. The interpre-tations were supported by energy minimiza-tion calculations of the c-ring structure witha short strip of helix 4 of subunit a, contain-ing residues aI225 to aM231, which suggestedpossible modes for the interaction between thestator arginine and an unoccupied binding site(Figure 6b) (77).
Inhibitors of the F0 motor. The most com-mon inhibitors of the F0 motor can be groupedinto the two different classes described below.
Arginine mimicking inhibitors. The rationalefor the first class of inhibitors of the F0 motoris a structural similarity with the stator chargearginine (Figure 7).
1. 5-(N-ethyl-N-isopropyl)amiloride (EIPA)effectively inhibits Na+-dependent ATPsynthases but not their H+-translocatingcounterparts (77, 78). Inhibition of theI. tartaricus enzyme could be modulatedby varying the Na+ concentration, indi-cating that Na+ and EIPA compete forthe same binding site (77). The amiloridescontain a guanidino group attached to ahydrophobic tail, which mimics the sidechain of the conserved arginine of sub-unit a. The rationale for the inhibitiontherefore is a competition between EIPAand the stator arginine for binding to thec-ring sites (77). This (Figure 6b) alsoprovides an explanation of why only Na+-dependent ATP synthases are affectedby amiloride compounds. A well-definedhydrogen bonding network between thearginine (or EIPA) and the bindingsite residues is not possible in most
660 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
cT67
cE65
aN230
aR226
cQ32
cV63
cG25
cS66
cY70cG68
cE65
cQ32 cV63
cT67
2.622.88
2.68
2.75
2.622.54
2.57
a b
Figure 6The a/c interface. (a) Perspective view of the surface of the isolated c ring of I. tartaricus. The atom boundaries are displayed on thesurface to allow visualization of the cavity below the ion binding site. The residues of the ion binding site and the glycine residues cG25and cG68 around the cavity are depicted (see text for details). (b) Coordination of the stator arginine after energy minimizationcalculations of the c ring and a heptapeptide of helix 4 of subunit a. The illustration shows the calculated positions and possiblehydrogen bonds of the binding site residues on the c ring and the stator arginine. Putative hydrogen bond lengths are marked inangstroms. The figure was prepared using PyMOL (DeLano Scientific).
H+-translocating enzymes because sev-eral of the proposed binding partners aremissing.
2. DCCD inhibits the F0 part of ATP syn-thases by forming an N-acyl bond withthe conserved carboxylic acid of subunitc (79). A rationale for the recognitionof DCCD by the c-ring binding sitesis the similarity of its protonated tran-sition state to the guanidino group ofthe stator arginine. Several lines of ev-idence show that DCCD preferentiallyreacts with binding sites outside the a/cinterface and that inhibition is caused bysterically blocked rotation of the modi-fied c ring against subunit a (80). Success-ful Na+/Na+-exchange experiments withthe DCCD-modified P. modestum ATPsynthase support the view that the ion
translocation pathway within the a/c in-terface is not affected by the modification(50).
A diaquilirine compound, which inhibits theATP synthase of Mycobacterium tubercolusis inthe nanomolar range, has been proposed to actsimilarly by mimicking the stator charge guani-dine group (81).
Tributyltin chloride. Tributyltin chloride(TBT) and its derivatives are known as nonco-valent inhibitors of oxidative phosphorylationand ATP synthesis. Competition studiesbetween TBT and DCCD initially seemed toindicate that both compounds bind to the samesite (82), but this has been challenged. Withone of the butyl chains of TBT substituted bya photoaffinity label for covalent modification,
www.annualreviews.org • Essentials of F1F0 ATP Synthases 661
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
N
N
Cl
N
NH2
HN
O
NH2
NH2
c-HxN
C
Nc-Hx
c-Hx
NH
C
Nc-Hx
HN
NH2
NH2R
DCCD
Arginine
EIPA
H+
Figure 7Structural analogies in F0 inhibitors and the statorcharge. Shown are the structures of EIPA (top),arginine (middle), and DCCD (bottom) and itsproposed transition state after protonation.Abbreviations: c-Hx, cyclohexyl; DCCD,N,N ′-dicyclohexylcarbodiimide; EIPA,5-(N-ethyl-N-isopropyl)amiloride.
Δψ: membranepotential, i.e., theelectrical potentialdifference betweentwo bulk phasesseparated by amembrane
ΔpH: the pHdifference betweentwo bulk phases oneither side of amembrane
P side: the side of abiological membranethat is positivelycharged (e.g.,periplasm in bacteria)
the primary site of interaction was identified assubunit a, not the c ring (83). TBT inhibitedthe Na+-dependent ATP synthase, and thiswas impaired by high concentrations (>100mM) of Na+, demonstrating a competitionbetween TBT and Na+ at a site different fromthe c-ring binding site. In contrast to DCCD,TBT also impaired Na+/ Na+ exchange in theI. tartaricus enzyme, indicating blockage of theion transport pathway within the a/c interface(83). Proton transport through the purified F0
parts of the ATP synthases of E. coli and spinachchloroplasts was also inhibited by TBT (84).
Driving Forces for Na+- orH+-Coupled F0 Motors
In most organisms, ATP synthesis is energizedby the proton-motive force (�μH+), which is
the sum of the thermodynamically equivalentparameters membrane potential (�ψ) andproton concentration gradient (�pH). Someanaerobic bacteria couple ATP synthesis toNa+ ion translocation and therefore utilizethe electrochemical gradient of Na+ ions(sodium-motive force) consisting of �ψ andthe Na+ concentration gradient (�pNa) as adriving force.
Implicit in Mitchell’s chemiosmotic model(85) is the assumption that membrane potentialand transmembrane ion gradients are equiva-lent driving forces for ATP synthesis. First, invitro experiments with the chloroplast enzymeindicated that ATP synthesis can indeed be en-ergized by a �pH exclusively (86). This con-clusion has been challenged, however, becausethe method to establish the �pH through anacid/base transition with succinate as the acidicbuffer also creates a significant �ψ owing tothe membrane permeability of the succinatemonoanion (87). Early experiments with themitochondrial ATP synthase seemed to indicateequivalence between �pH and �ψ as drivingforces for ATP synthesis (88, 89). Accordingly,it was predicted that ATP synthesis occurs if thethermodynamic requirements are fulfilled, i.e.,if
�μH+ = �ψ + �pH > �Gp
= �G0, × log10([ATP]/([ADP]×[Pi])).
This prediction was questioned by ATP syn-thesis experiments in whole cells of Streptococcuslactis, revealing that not all combinations of �ψ
and �pH were able to elicit ATP synthesis, ir-respective of the total driving force (90).
Accordingly, the thermodynamically equiv-alent driving forces �ψ and �pH appear tobe not kinetically equivalent, suggesting thatdifferent kinetic barriers have to be overcomeby either �ψ or �pH. The idea of a nonperfectequivalence between �ψ and �pH (or �pNa)was supported by results with the purifiedenzymes from Bacillus PS3, E. coli, Wolinellasuccinogenes (91–93), and P. modestum (101).In all cases, a total driving force >180 mV,consisting of a minimal �ψ of ∼80 mV and a
662 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.
ANRV378-BI78-22 ARI 29 April 2009 18:5
P side NaCl (mM)0 50 100 150
0
20
40
60
80
100
% re
lati
ve a
ctiv
ity
% re
lati
ve a
ctiv
ity
25 75 125
P side pH5.75 6.00 6.25 6.50 6.75 7.00
0
20
40
60
80
100
a b
91 mV
122 mVATP synthesis
ATP hydrolysis
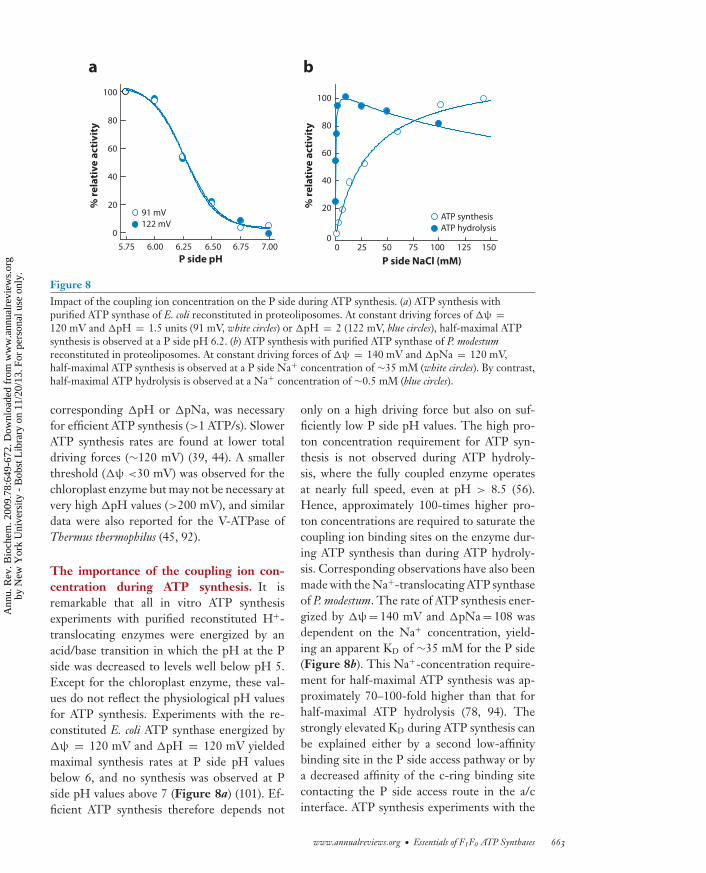
Figure 8Impact of the coupling ion concentration on the P side during ATP synthesis. (a) ATP synthesis withpurified ATP synthase of E. coli reconstituted in proteoliposomes. At constant driving forces of �ψ =120 mV and �pH = 1.5 units (91 mV, white circles) or �pH = 2 (122 mV, blue circles), half-maximal ATPsynthesis is observed at a P side pH 6.2. (b) ATP synthesis with purified ATP synthase of P. modestumreconstituted in proteoliposomes. At constant driving forces of �ψ = 140 mV and �pNa = 120 mV,half-maximal ATP synthesis is observed at a P side Na+ concentration of ∼35 mM (white circles). By contrast,half-maximal ATP hydrolysis is observed at a Na+ concentration of ∼0.5 mM (blue circles).
corresponding �pH or �pNa, was necessaryfor efficient ATP synthesis (>1 ATP/s). SlowerATP synthesis rates are found at lower totaldriving forces (∼120 mV) (39, 44). A smallerthreshold (�ψ <30 mV) was observed for thechloroplast enzyme but may not be necessary atvery high �pH values (>200 mV), and similardata were also reported for the V-ATPase ofThermus thermophilus (45, 92).
The importance of the coupling ion con-centration during ATP synthesis. It isremarkable that all in vitro ATP synthesisexperiments with purified reconstituted H+-translocating enzymes were energized by anacid/base transition in which the pH at the Pside was decreased to levels well below pH 5.Except for the chloroplast enzyme, these val-ues do not reflect the physiological pH valuesfor ATP synthesis. Experiments with the re-constituted E. coli ATP synthase energized by�ψ = 120 mV and �pH = 120 mV yieldedmaximal synthesis rates at P side pH valuesbelow 6, and no synthesis was observed at Pside pH values above 7 (Figure 8a) (101). Ef-ficient ATP synthesis therefore depends not
only on a high driving force but also on suf-ficiently low P side pH values. The high pro-ton concentration requirement for ATP syn-thesis is not observed during ATP hydroly-sis, where the fully coupled enzyme operatesat nearly full speed, even at pH > 8.5 (56).Hence, approximately 100-times higher pro-ton concentrations are required to saturate thecoupling ion binding sites on the enzyme dur-ing ATP synthesis than during ATP hydroly-sis. Corresponding observations have also beenmade with the Na+-translocating ATP synthaseof P. modestum. The rate of ATP synthesis ener-gized by �ψ = 140 mV and �pNa = 108 wasdependent on the Na+ concentration, yield-ing an apparent KD of ∼35 mM for the P side(Figure 8b). This Na+-concentration require-ment for half-maximal ATP synthesis was ap-proximately 70–100-fold higher than that forhalf-maximal ATP hydrolysis (78, 94). Thestrongly elevated KD during ATP synthesis canbe explained either by a second low-affinitybinding site in the P side access pathway or bya decreased affinity of the c-ring binding sitecontacting the P side access route in the a/cinterface. ATP synthesis experiments with the
www.annualreviews.org • Essentials of F1F0 ATP Synthases 663
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
N side: the side of abiological membranethat is negativelycharged (e.g.,cytoplasm in bacteria)
closely related enzyme of I. tartaricus, wherethe binding site was altered by mutagenesis,strongly favor the second scenario in which thebinding site has an altered affinity when ac-cessed from the P side instead of the N side(101). Furthermore, the apparent KD was resis-tant to variation of �ψ, implying that �ψ hasno direct effect on the ion uptake process fromthe P side.
Ion translocation studies with the isolatedF0 parts of these enzymes reinforced the asym-metric properties of the holoenzymes, showingdifferent ion affinities from either side. Further-more, it was particularly interesting to observethat �pNa, unlike �ψ, is only an effective driv-ing force for ion transport through F0 in syn-thesis, but not in the hydrolysis direction in theP. modestum enzyme, whereas �pH showed tobe an equivalent driving force to �ψ in the en-zymes of E. coli and spinach chloroplasts (84,95, 96, 101).
Model for torque generation in the F0 part.This paragraph describes a mechanistic modelfor torque generation in F0 that takes recentfindings, described in the previous sections, intoaccount (Figure 9). A key feature of the modelis the assumption that the binding site in theperiplasmic access channel (S1) forms a tightcomplex with the stator arginine, whereas allother c-ring sites are in an ion-bound state. Inorder to elicit rotation, the arginine must bereleased from S1 and form a new complex withthe next incoming binding site (Sn). A couplingion entering through the P side channel fromthe periplasm competes with the arginine forcomplex formation with the S1 site and, at suf-ficiently high ion concentrations, displaces thearginine (a → b) (Figure 9). As soon as thearginine becomes displaced, the interaction be-tween the rotor and the stator is temporarilyreleased, and S1, with its neutralized charge,is ready to move into the lipid interface. Toproceed into this direction, the ion from theSn site must be released into the N side chan-nel, and a new complex of the empty Sn sitewith the free arginine must be formed (b → c)(Figure 9). Rotation therefore requires ap-
propriate external conditions for the replace-ment of arginine from the S1(Arg) complex byH+/Na+ and for the dissociation of H+/Na+
from the Sn(H+/Na+) complex. Hence, the Pside H+/Na+ concentration must suffice effec-tive binding of the coupling ion to the S1 site,which is of low affinity, and simultaneously,dissociation of H+/Na+ from the Sn(H+/Na+)complex must be guaranteed. This reaction isaffected not only by the N side ion concentra-tion but also by the membrane potential. Thesubsequent switch of the arginine from S1 toSn generates torque and completes one step ofc-ring rotation and ion transport.
Generally speaking, the described modelis governed by dissociation/association equlib-riums of binding site/arginine and bindingsite/ion complexes at the two opposing ion ac-cess routes. These equlibriums are controlledby the ion concentrations on both sides ofthe membrane and by the membrane potential,leading under proper conditions to directed ro-tation and torque generation as follows:
S1(Arg) + Sn(H+/Na+) + H+/Na+out
� S1(H+/Na+) + Sn(Arg) + H+/Na+in.
Physiological implications. The present re-sults affect the mechanism of ATP synthesisin vivo. The driving forces �ψ and �pNa inP. modestum or �ψ and �pH in E. coli arecreated by the methylmalonyl-CoA decarboxy-lase Na+ pump or respiratory proton pumps,respectively. The high Na+ concentration re-quirement for ATP synthesis in P. modestum isreadily realized by its natural marine environ-ment. For E. coli, however, which grows op-timally at pH 7–8 but requires pH < 6.5 forATP synthesis in vitro, the underlying mecha-nism is more difficult to reconcile. An obvioussolution to this dilemma is a local increase ofthe proton concentration at the ATP synthase,as proposed in the long-standing hypothesis oflateral diffusion of the pumped protons alongthe membrane surface. The hypothesis is inaccordance with the observation that low pHvalues (<6.5) are only required if ATP synthe-sis is energized by an artificial K+/valinomycin
664 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
diffusion potential, whereas E. coli vesicles read-ily synthesize ATP at pH 7–8 if they are en-ergized by respiratory proton pumping (101).Similar differences, but with a shift to more al-kaline pH values, had previously been describedfor the different modes of energizing ATP syn-thesis in the alkaliphilic Bacillus firmus (97). ATPsynthesis by H. salinarium despite the presenceof uncouplers can be similarly explained by lat-eral proton diffusion (98).
It is important to note that effective ATPsynthesis requires contributions from the twodriving forces �ψ and �pH and a high pro-ton concentration at the source P side. Atfirst glance, all of these criteria seem to befulfilled rarely. The proton-motive force inmitochondria and neutrophilic aerobic bacte-ria, e.g., E. coli, primarily consists of a high �ψ
and only a small �pH. This scenario is exagger-ated in alkaliphilic bacteria, where the pH gra-dient is reversed, and the driving force is con-tributed entirely by the �ψ. Furthermore, theenvironmental pH barely meets the high protonconcentration requirements for ATP synthesis.A local acidification at the P side membrane
−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−→Figure 9Events in the rotor/stator interface during ATPsynthesis. (Rotation from left to right). Side view ofthe a/c interface of an F0 motor. Shown is the c ring(blue) with binding sites (red circles, S1 to Sn), subunita containing the stator charge (Arg), and the P sideand the N side access channels ( green). (a) In thestarting position, the stator arginine is tightly boundto binding site S1. Governed by chemicalequlibriums, which are influenced by the drivingforces �ψ and �pH/�pNa, an incoming ion fromthe P side is bound to site S1 (requiring a high ionconcentration), which displaces the Arg from the S1site. Simultaneously, the equlibriums have to allow arelease of an ion from the Sn site to the N side,leading to a transition state in which the Arg islocated between sites S1 and Sn, and the rotor isdisconnected from the stator (b). This allows theneutralized S1 site to move into the lipid bilayer andthe formation of an Arg-Sn complex in the interface,which generates unidirectional torque (c). Browianmotions are thought to bring the F0 motor furtherinto a new starting position (c → a). See text foradditional details.
border is therefore a tempting explanation forATP synthesis under these apparently inappro-priate conditions. In chloroplasts, by contrast,the proton-motive force consists of a large �pHand only a small �ψ. Efficient ATP production,
C-ringAccesschannel
Binding site
Ion
Arg
Arg
Arg
High affinity
Low affinity
a
b
c
P side
N side
Δψ
S2Sn S1
‡
S2Sn S1
S2Sn S1
Ion binding/ Arg release at S1
Ion release at Sn
Arg binding at Sn
Generation of rotation
www.annualreviews.org • Essentials of F1F0 ATP Synthases 665
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
under these conditions, may require specific
Ion motive force: anelectrochemical iongradient composed ofthe ion concentrationgradient and theelectrical membranepotential across amembrane
adaptations of the F0 motor itself, e.g., a largec ring or an adapted binding site with an atten-uated response to the �ψ. Although the highproton concentration demand for ATP synthe-sis is obviously met by dynamic processes at thecell membrane, the reverse operation of the en-zyme in fermenting bacteria relies on the pro-ton concentration in the cytoplasmic environ-ment, which usually is in the neutral to slightly
alkaline pH range. It is therefore essential thatATP synthases, like that of E. coli functioningduring fermentation as ATP-driven ion pumps,operate at near maximal rate at pH valuesbetween 7 and 8. Hence, the asymmetric pro-ton concentration requirement for ATP syn-thesis and hydrolysis is a consequence of theenzyme’s adaptation to the natural environmentand its additional function as a �ψ-generatingion pump.
SUMMARY POINTS
1. ATP is the major energy currency in living cells. Each cell performs an energy cycle inwhich ATP is split into ADP and phosphate to drive numerous endergonic reactions.The cleavage products are subsequently recombined to ATP by the F1F0 ATP synthase.The primary energy source for ATP synthesis is either light or an exergonic chemicalreaction. The primary energy sources are converted into an electrochemical ion gradient,which serves as the actual driving force for ATP synthesis, across the membrane.
2. Structure and function of the F1 part are highly conserved, most likely because similarconditions prevail in its environment inside the soluble compartment of the cell (pH,salt). In contrast, the F0 part faces highly diverse environments (pH, salt, driving forces),and its functionality thus diverges to adapt to specific conditions. Likewise, the F0 motorhas evolved different strategies to efficiently use the dominant component of the ionmotive force.
3. The mechanism of ATP formation in the F1 part is well described by the binding changemechanism. This has been developed in great detail by many techniques, and under-standing of the mechanism has been claimed to be “almost complete.”
4. In most bacteria, the unc/atp operon contains nine open reading frames. However, onlyeight different subunits are identified in the assembled enzyme. The missing subunit,the UncI protein, has been characterized in P. modestum as a chaperone assisting inthe assembly of the c ring from single subunits. This discovery not only provides anexplanation for the conservation of the uncI gene, but also sheds light on the intricatemechanism of ATP synthase assembly.
5. Kinetic studies show that the binding affinity between F1 and F0 subunits of the peripheraland central stalk are comparable and thus provide a robust scaffold to withstand the elasticdrag imposed during rotation.
6. Within the family of proton-translocating ATP synthases, different proton binding char-acteristics are evident. In one group of enzymes, the proton binding sites are characterizedby group protonation of the conserved c ring carboxylates. In the other group, the pro-ton binding characteristics are incompatible with a group protonation mechanism andinstead indicate coordination of a hydronium ion.
666 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
7. The total minimal driving forces for efficient ATP synthesis (>1 ATP/s) reported arevery similar (>180 mV). Thereby, �ψ and �pH/�pNa are not equivalent driving forces.While a �ψ may not be required in all organisms (e.g. chloroplasts), it certainly is themajor thermodynamic driving force in most species (e.g., bacteria, mitochondria).
8. A high coupling ion concentration on the P side was defined as a critical and so farunrecognized determinant for ATP synthesis. Concomitantly, this finding demonstratesthe asymmetry of ion binding to the c-ring binding site from either side of the membrane.In contrast to Na+-dependent enzymes, the demand of values below pH 6 at the P sidein E. coli is not compatible with its prevailing growth conditions and supports the long-standing hypothesis of lateral diffusion of translocated protons along membrane surfaces.
9. Cys-Cys cross-linking data indicate a conformational change in the c-ring binding siteupon contact with the stator charge in the a/c interface. This structural change allowsthe release of the coupling ion and tight interaction with the stator charge. Appropriatedriving forces and ion concentrations allow the subsequent dissociation of this complexby an incoming coupling ion as well as ion release from the neighboring rotor site inthe a/c interface. The interaction of the free arginine with the just generated unoccupiedbinding site elicits unidirectional rotation and generates torque for the synthesis of ATP.
FUTURE ISSUES
1. The assembly of the ATP synthase and the synthesis of a balanced stoichiometry of thevarious subunits are still unclear. Although interesting insights were gained in the recentyears, these questions have escaped from intense investigations so far.
2. The a/c interface.
a. The a/c interface is at the core of the F0 mechanism, generating torque at the ex-pense of ion translocation across the membrane. Until now this fundamental proteinassembly for the operation of the entire enzyme has escaped structural investigationswith high resolution. Such a structure is likely to make a significant contribution toour understanding of the F0 mechanism.
b. A possible structure displays only a snapshot and may not be sufficient to unravel thedynamic features that happen in the a/c interface, and thus, functional investigationsalong the ion pathway are of critical importance.
c. Molecular modeling investigations based on the crystal structure and biochemicaldata might provide interesting details, explaining how �ψ and �pH/ �pNa+ influ-ence the structure within the a/c interface.
3. The concept of lateral proton transfer and its impact on bioenergetics in general andATP synthesis in particular requires an in-depth investigation. Although several lines ofevidence exist for this scenario in vitro, its presence and importance in vivo are still elusive.Generally, the interplay between �μH+-generating and �μH+-consuming enzymes hasto be investigated not only on a structural (e.g., supercomplexes) but also on a functionallevel. Thereby, the impact of the lipid composition of biological membranes has to beconsidered. A pronounced impact of localized proton circuits between these proteinswould have wide consequences for the entire field of bioenergetics.
www.annualreviews.org • Essentials of F1F0 ATP Synthases 667
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
Work in the authors’ laboratory was supported by Swiss National Science Foundation. Christophvon Ballmoos and Alexander Wiedenmann were at the Institute of Microbiology, ETH Zurich,when this article was written.
LITERATURE CITED
1. Saraste M. 1999. Oxidative phosphorylation at the fin de siecle. Science 283:1488–932. Dimroth P, Cook GM. 2004. Bacterial Na+- or H+-coupled ATP synthases operating at low electro-
chemical potential. Adv. Microb. Physiol. 49:175–2183. Dimroth P, von Ballmoos C. 2008. ATP synthesis by decarboxylation phosphorylation. Results Probl. Cell
Differ. 45:153–844. Abrahams JP, Leslie AG, Lutter R, Walker JE. 1994. Structure at 2.8 A resolution of F1-ATPase from
bovine heart mitochondria. Nature 370:621–285. Boyer PD. 1993. The binding change mechanism for ATP synthase—some probabilities and possibilities.
Biochim. Biophys. Acta 1140:215–506. Noji H, Yasuda R, Yoshida M, Kinosita K Jr. 1997. Direct observation of the rotation of F1-ATPase.
Nature 386:299–3027. Panke O, Gumbiowski K, Junge W, Engelbrecht S. 2000. F-ATPase: specific observation of the rotating
c subunit oligomer of EF0EF1. FEBS Lett. 472:34–388. Capaldi RA, Aggeler R, Wilkens S. 1995. Conformational changes in the gamma and epsilon subunits
are integral to the functioning of the Escherichia coli H+-pumping ATPase (ECF1F0). Biochem. Soc. Trans.23:767–70
9. Duncan TM, Bulygin VV, Zhou Y, Hutcheon ML, Cross RL. 1995. Rotation of subunits during catalysisby Escherichia coli F1-ATPase. Proc. Natl. Acad. Sci. USA 92:10964–68
10. Gogol EP, Johnston E, Aggeler R, Capaldi RA. 1990. Ligand-dependent structural variations inEscherichia coli F1 ATPase revealed by cryoelectron microscopy. Proc. Natl. Acad. Sci. USA 87:9585–89
11. Sabbert D, Engelbrecht S, Junge W. 1997. Functional and idling rotatory motion within F1-ATPase.Proc. Natl. Acad. Sci. USA 94:4401–5
12. Stock D, Leslie AG, Walker JE. 1999. Molecular architecture of the rotary motor in ATP synthase.Science 286:1700–5
13. Kaim G, Prummer M, Sick B, Zumofen G, Renn A, et al. 2002. Coupled rotation within single F0F1
enzyme complexes during ATP synthesis or hydrolysis. FEBS Lett. 525:156–6314. Diez M, Zimmermann B, Borsch M, Konig M, Schweinberger E, et al. 2004. Proton-powered subunit
rotation in single membrane-bound F0F1-ATP synthase. Nat. Struct. Mol. Biol. 11:135–4115. Ueno H, Suzuki T, Kinosita K Jr, Yoshida M. 2005. ATP-driven stepwise rotation of F0F1-ATP synthase.
Proc. Natl. Acad. Sci. USA 102:1333–3816. Nishizaka T, Oiwa K, Noji H, Kimura S, Muneyuki E, et al. 2004. Chemomechanical coupling in F1-
ATPase revealed by simultaneous observation of nucleotide kinetics and rotation. Nat. Struct. Mol. Biol.11:142–48
17. Describessophisticatedexperiments, in whichcatalysis and rotationare monitored inparallel, elucidating thecomplete reaction cycleduring ATP hydrolysis.
17. Adachi K, Oiwa K, Nishizaka T, Furuike S, Noji H, et al. 2007. Coupling of rotation and catalysisin F1-ATPase revealed by single-molecule imaging and manipulation. Cell 130:309–21
18. Observation of ATPsynthesis bysingle-moleculeexperiments innanosized chemicalchambers.
18. Rondelez Y, Tresset G, Nakashima T, Kato-Yamada Y, Fujita H, et al. 2005. Highly coupled ATPsynthesis by F1-ATPase single molecules. Nature 433:773–77
19. Itoh H, Takahashi A, Adachi K, Noji H, Yasuda R, et al. 2004. Mechanically driven ATP synthesis byF1-ATPase. Nature 427:465–68
668 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
20. Valiyaveetil FI, Fillingame RH. 1998. Transmembrane topography of subunit a in the Escherichia coliF1F0 ATP synthase. J. Biol. Chem. 273:16241–47
21. Vik SB, Ishmukhametov RR. 2005. Structure and function of subunit a of the ATP synthase of Escherichiacoli. J. Bioenerg. Biomembr. 37:445–49
22. Dunn S, Revington M, Cipriano D, Shilton B. 2000. The b subunit of Escherichia coli ATP synthase.J. Bioenerg. Biomembr. 32:347–55
23. Greie JC, Deckers-Hebestreit G, Altendorf K. 2000. Subunit organization of the stator part of the F0
complex from Escherichia coli ATP synthase. J. Bioenerg. Biomembr. 32:357–6424. Weber J, Wilke-Mounts S, Nadanaciva S, Senior AE. 2004. Quantitative determination of direct binding
of b subunit to F1 in Escherichia coli F1F0-ATP synthase. J. Biol. Chem. 279:11253–5825. Weber J. 2007. ATP synthase–the structure of the stator stalk. Trends Biochem. Sci. 32:53–5626. Dickson VK, Silvester JA, Fearnley IM, Leslie AG, Walker JE. 2006. On the structure of the stator of
the mitochondrial ATP synthase. EMBO J. 25:2911–1827. Ackerman SH, Tzagoloff A. 2005. Function, structure, and biogenesis of mitochondrial ATP synthase.
Prog. Nucleic Acid Res. Mol. Biol. 80:95–13328. von Meyenburg K, Jorgensen BB, Nielsen J, Hansen FG. 1982. Promoters of the atp operon coding for
the membrane-bound ATP synthase of Escherichia coli mapped by Tn10 insertion mutations. Mol. Gen.Genet. 188:240–48
29. Gay NJ. 1984. Construction and characterization of an Escherichia coli strain with a uncI mutation.J. Bacteriol. 158:820–25
30. Investigations on therole of the UncI proteinas chaperone assistingin c-ring formation in aNa+-dependent ATPsynthase.
30. Suzuki T, Ozaki Y, Sone N, Feniouk BA, Yoshida M. 2007. The product of uncI gene in F1F0-ATP synthase operon plays a chaperone-like role to assist c-ring assembly. Proc. Natl. Acad. Sci.
USA 104:20776–8131. Meier T, Yu J, Raschle T, Henzen F, Dimroth P, Muller DJ. 2005. Structural evidence for a constant
c11 ring stoichiometry in the sodium F-ATP synthase. FEBS J. 272:5474–8332. Kol S, Nouwen N, Driessen AJ. 2008. Mechanisms of YidC-mediated insertion and assembly of multi-
meric membrane protein complexes. J. Biol. Chem. 283:31269–7333. Ozaki Y, Suzuki T, Kuruma Y, Ueda T, Yoshida M. 2008. UncI protein can mediate ring-assembly of
c-subunits of F0F1-ATP synthase in vitro. Biochem. Biophys. Res. Commun. 367:663–6634. Kinetic studies onthe interaction betweenrotor subunits of abacterial ATP synthase.
34. Pogoryelov D, Nikolaev Y, Schlattner U, Pervushin K, Dimroth P, Meier T. 2008. Probing therotor subunit interface of the ATP synthase from Ilyobacter tartaricus. FEBS J. 275:4850–62
35. Krebstakies T, Zimmermann B, Graber P, Altendorf K, Borsch M, Greie JC. 2005. Both rotor and statorsubunits are necessary for efficient binding of F1 to F0 in functionally assembled Escherichia coli ATPsynthase. J. Biol. Chem. 280:33338–45
36. Meier T, Morgner N, Matthies D, Pogoryelov D, Keis S, et al. 2007. A tridecameric c ring of theadenosine triphosphate (ATP) synthase from the thermoalkaliphilic Bacillus sp. strain TA2.A1 facilitatesATP synthesis at low electrochemical proton potential. Mol. Microbiol. 65:1181–92
37. Seelert H, Poetsch A, Dencher NA, Engel A, Stahlberg H, Muller DJ. 2000. Structural biology. Proton-powered turbine of a plant motor. Nature 405:418–19
38. Cook GM, Keis S, Morgan HW, von Ballmoos C, Matthey U, et al. 2003. Purification and biochemicalcharacterization of the F1F0-ATP synthase from thermoalkaliphilic Bacillus sp. strain TA2.A1. J. Bacteriol.185:4442–49
39. A recent reviewdescribing the diversityof ATP synthases indifferent organisms.
39. von Ballmoos C, Cook GM, Dimroth P. 2008. Unique rotary ATP synthase and its biologicaldiversity. Annu. Rev. Biophys. 37:43–64
40. Tomashek JJ, Brusilow WS. 2000. Stoichiometry of energy coupling by proton-translocating ATPases:a history of variability. J. Bioenerg. Biomembr. 32:493–500
41. Krebstakies T, Aldag I, Altendorf K, Greie JC, Deckers-Hebestreit G. 2008. The stoichiometry ofsubunit c of Escherichia coli ATP synthase is independent of its rate of synthesis. Biochemistry 47:6907–16
42. Meyer Zu, Tittingdorf JM, Rexroth S, Schafer E, Schlichting R, Giersch C, et al. 2004. The stoichiometryof the chloroplast ATP synthase oligomer III in Chlamydomonas reinhardtii is not affected by the metabolicstate. Biochim. Biophys. Acta 1659:92–99
43. Hinkle PC. 2005. P/O ratios of mitochondrial oxidative phosphorylation. Biochim. Biophys. Acta1706:1–11
www.annualreviews.org • Essentials of F1F0 ATP Synthases 669
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
44. Steigmiller S, Turina P, Graber P. 2008. The thermodynamic H+/ATP ratios of the H+-ATP synthasesfrom chloroplasts and Escherichia coli. Proc. Natl. Acad. Sci. USA 105:3745–50
45. Toei M, Gerle C, Nakano M, Tani K, Gyobu N, et al. 2007. Dodecamer rotor ring defines H+/ATPratio for ATP synthesis of prokaryotic V-ATPase from Thermus thermophilus. Proc. Natl. Acad. Sci. USA104:20256–61
46. High-resolutionstructure ofNa+-binding c ringfrom an F-type ATPase.
46. Meier T, Polzer P, Diederichs K, Welte W, Dimroth P. 2005. Structure of the rotor ring ofF-type Na+-ATPase from Ilyobacter tartaricus. Science 308:659–62
47. Vonck J, Krug von Nidda T, Meier T, Matthey U, Mills DJ, et al. 2002. Molecular architecture of theundecameric rotor of a bacterial Na+-ATP synthase. J. Mol. Biol. 321:307–16
48. High-resolutionstructure of aNa+-binding K ringfrom a V-type ATPase.
48. Murata T, Yamato I, Kakinuma Y, Leslie AG, Walker JE. 2005. Structure of the rotor of theV-Type Na+-ATPase from Enterococcus hirae. Science 308:654–59
49. Oberfeld B, Brunner J, Dimroth P. 2006. Phospholipids occupy the internal lumen of the c ring of theATP synthase of Escherichia coli. Biochemistry 45:1841–51
50. von Ballmoos C, Appoldt Y, Brunner J, Granier T, Vasella A, Dimroth P. 2002. Membrane topographyof the coupling ion binding site in Na+-translocating F1F0 ATP synthase. J. Biol. Chem. 277:3504–10
51. Murata T, Igarashi K, Kakinuma Y, Yamato I. 2000. Na+ binding of V-type Na+-ATPase in Enterococcushirae. J. Biol. Chem. 275:13415–19
52. Dimroth P, von Ballmoos C, Meier T. 2006. Catalytic and mechanical cycles in F-ATP synthases. Fourthin the Cycles Review Series. EMBO Rep. 7:276–82
53. Engelbrecht S, Junge W. 1997. ATP synthase: a tentative structural model. FEBS Lett. 414:485–9154. Vik SB, Antonio BJ. 1994. A mechanism of proton translocation by F1F0 ATP synthases suggested by
double mutants of the a subunit. J. Biol. Chem. 269:30364–6955. Fillingame RH, Angevine CM, Dmitriev OY. 2003. Mechanics of coupling proton movements to c-ring
rotation in ATP synthase. FEBS Lett. 555:29–3456. von Ballmoos C, Dimroth P. 2007. Two distinct proton binding sites in the ATP synthase family. Bio-
chemistry 46:11800–957. Boyer PD. 1988. Bioenergetic coupling to protonmotive force: Should we be considering hydronium
ion coordination and not group protonation? Trends Biochem. Sci. 13:5–758. Mellwig C, Bottcher B. 2003. A unique resting position of the ATP-synthase from chloroplasts. J. Biol.
Chem. 278:18544–4959. Rubinstein JL, Walker JE, Henderson R. 2003. Structure of the mitochondrial ATP synthase by electron
cryomicroscopy. EMBO J. 22:6182–9260. Greie JC, Heitkamp T, Altendorf K. 2004. The transmembrane domain of subunit b of the Escherichia
coli F1F0 ATP synthase is sufficient for H+-translocating activity together with subunits a and c. Eur. J.Biochem. 271:3036–42
61. Stalz WD, Greie JC, Deckers-Hebestreit G, Altendorf K. 2003. Direct interaction of subunits a andb of the F0 complex of Escherichia coli ATP synthase by forming an ab2 subcomplex. J. Biol. Chem.278:27068–71
62. Ono S, Sone N, Yoshida M, Suzuki T. 2004. ATP synthase that lacks F0a-subunit: isolation, properties,and indication of F0b2-subunits as an anchor rail of a rotating c-ring. J. Biol. Chem. 279:33409–12
63. Cain BD, Simoni RD. 1989. Proton translocation by the F1F0 ATPase of Escherichia coli. Mutagenicanalysis of the a-subunit. J. Biol. Chem. 264:3292–300
64. Wehrle F, Kaim G, Dimroth P. 2002. Molecular mechanism of the ATP synthase’s F0 motor probed bymutational analyses of subunit a. J. Mol. Biol. 322:369–81
65. Elston T, Wang H, Oster G. 1998. Energy transduction in ATP synthase. Nature 391:510–1366. Xing J, Wang H, von Ballmoos C, Dimroth P, Oster G. 2004. Torque generation by the F0 motor of the
sodium ATPase. Biophys. J. 87:2148–6367. Cain BD, Simoni RD. 1988. Interaction between Glu-219 and His-245 within the a subunit of F1F0-
ATPase in Escherichia coli. J. Biol. Chem. 263:6606–1268. Schwem BE, Fillingame RH. 2006. Cross-linking between helices within subunit a of Escherichia coli ATP
synthase defines the transmembrane packing of a four-helix bundle. J. Biol. Chem. 281:37861–6769. Angevine CM, Herold KA, Fillingame RH. 2003. Aqueous access pathways in subunit a of rotary ATP
synthase extend to both sides of the membrane. Proc. Natl. Acad. Sci. USA 100:13179–83
670 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
70. Angevine CM, Fillingame RH. 2003. Aqueous access channels in subunit a of rotary ATP synthase.J. Biol. Chem. 278:6066–74
71. Meier T, Matthey U, von Ballmoos C, Vonck J, Krug von Nidda T, et al. 2003. Evidence for structuralintegrity in the undecameric c-rings isolated from sodium ATP synthases. J. Mol. Biol. 325:389–97
72. von Ballmoos C, Meier T, Dimroth P. 2002. Membrane embedded location of Na+ or H+ binding siteson the rotor ring of F1F0 ATP synthases. Eur. J. Biochem. 269:5581–89
73. Jiang W, Fillingame RH. 1998. Interacting helical faces of subunits a and c in the F1F0 ATP synthase ofEscherichia coli defined by disulfide cross-linking. Proc. Natl. Acad. Sci. USA 95:6607–12
74. Rastogi VK, Girvin ME. 1999. Structural changes linked to proton translocation by subunit c of theATP synthase. Nature 402:263–68
75. Nakano T, Ikegami T, Suzuki T, Yoshida M, Akutsu H. 2006. A new solution structure of ATP synthasesubunit c from thermophilic Bacillus PS3, suggesting a local conformational change for H+-translocation.J. Mol. Biol. 358:132–44
76. Vincent OD, Schwem BE, Steed PR, Jiang W, Fillingame RH. 2007. Fluidity of structure and swiveling ofhelices in the subunit c ring of Escherichia coli ATP synthase as revealed by cysteine-cysteine cross-linking.J. Biol. Chem. 282:33788–94
77. Experimental andcomputational analysisof the a/c interface,describing possiblemodes for interaction ofthe c ring and the statorcharge.
77. Vorburger T, Zingg Ebneter J, Wiedenmann A, Morger D, Weber G, et al. 2008. Arginine-induced conformational change in the c-ring/a-subunit interface of ATP synthase. FEBS J.
275:2137–5078. Kluge C, Dimroth P. 1993. Kinetics of inactivation of the F1F0 ATPase of Propionigenium modestum by
dicyclohexylcarbodiimide in relationship to H+ and Na+ concentration: probing the binding site for thecoupling ions. Biochemistry 32:10378–86
79. Hoppe J, Schairer HU, Sebald W. 1980. Identification of amino-acid substitutions in the proteolipidsubunit of the ATP synthase from dicyclohexylcarbodiimide-resistant mutants of Escherichia coli. Eur. J.Biochem. 112:17–24
80. Hermolin J, Fillingame RH. 1989. H+-ATPase activity of Escherichia coli F1F0 is blocked after reactionof dicyclohexylcarbodiimide with a single proteolipid (subunit c) of the F0 complex. J. Biol. Chem.264:3896–903
81. de Jonge MR, Koymans LH, Guillemont JE, Koul A, Andries K. 2007. A computational model of theinhibition of Mycobacterium tuberculosis ATPase by a new drug candidate R207910. Proteins 67:971–80
82. Partis MD, Bertoli E, Griffiths DE, Azzi A. 1980. Interaction of the dibutylchloromethyltin chloridebinding site with the carbodiimide binding site in mitochondria. Biochem. Biophys. Res. Commun. 96:1103–8
83. von Ballmoos C, Brunner J, Dimroth P. 2004. The ion channel of F-ATP synthase is the target of toxicorganotin compounds. Proc. Natl. Acad. Sci. USA 101:11239–44
84. Wiedenmann A, Dimroth P, von Ballmoos C. 2008. �ψ and �pH are equivalent driving forces forproton transport through isolated F0 complexes of ATP synthases. Biochim. Biophys. Acta 1777:1301–10
85. Mitchell P. 1961. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotictype of mechanism. Nature 191:144–48
86. Jagendorf AT, Uribe E. 1966. ATP formation caused by acid-base transition of spinach chloroplasts.Proc. Natl. Acad. Sci. USA 55:170–77
87. Kaim G, Dimroth P. 1999. ATP synthesis by F-type ATP synthase is obligatorily dependent on thetransmembrane voltage. EMBO J. 18:4118–27
88. Mitchell P. 1966. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev.Camb. Philos. Soc. 41:445–502
89. Cockrell RS, Harris EJ, Pressman BC. 1967. Synthesis of ATP driven by a potassium gradient in mito-chondria. Nature 215:1487–88
90. Maloney PC, Wilson TH. 1975. ATP synthesis driven by a protonmotive force in Streptococcus lactis.J. Membr. Biol. 25:285–310
91. Sone N, Yoshida M, Hirata H, Kagawa Y. 1977. Adenosine triphosphate synthesis by electrochemicalproton gradient in vesicles reconstituted from purified adenosine triphosphatase and phospholipids ofthermophilic bacterium. J. Biol. Chem. 252:2956–60
www.annualreviews.org • Essentials of F1F0 ATP Synthases 671
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

ANRV378-BI78-22 ARI 29 April 2009 18:5
92. Fischer S, Graber P. 1999. Comparison of �pH- and �ϕ-driven ATP synthesis catalyzed by the H+-ATPases from Escherichia coli or chloroplasts reconstituted into liposomes. FEBS Lett. 457:327–32
93. Bokranz M, Morschel E, Kroger A. 1985. Phosphorylation and phosphate-ATP exchange catalyzed bythe ATP synthase isolated from Wolinella succinogenes. Biochim. Biophys. Acta 810:332–39
94. Laubinger W, Dimroth P. 1988. Characterization of the ATP synthase of Propionigenium modestum as aprimary sodium pump. Biochemistry 27:7531–37
95. Feniouk BA, Kozlova MA, Knorre DA, Cherepanov DA, Mulkidjanian AY, Junge W. 2004. The proton-driven rotor of ATP synthase: ohmic conductance (10 fS), and absence of voltage gating. Biophys. J.86:4094–109
96. Kluge C, Dimroth P. 1992. Studies on Na+ and H+ translocation through the F0 part of the Na+-translocating F1F0 ATPase from Propionigenium modestum: discovery of a membrane potential dependentstep. Biochemistry 31:12665–72
97. Guffanti AA, Fuchs RT, Schneier M, Chiu E, Krulwich TA. 1984. A transmembrane electrical potentialgenerated by respiration is not equivalent to a diffusion potential of the same magnitude for ATP synthesisby Bacillus firmus RAB. J. Biol. Chem. 259:2971–75
98. Michel H, Oesterhelt D. 1980. Electrochemical proton gradient across the cell membrane of Halobac-terium halobium: comparison of the light-induced increase with the increase of intracellular adenosinetriphosphate under steady-state illumination. Biochemistry 19:4615–19
99. Comprehensivereview on theregulatory mechanismsin F-ATP synthases.
99. Feniouk BA, Yoshida M. 2008. Regulatory mechanisms of proton-translocating F0F1-ATP syn-thase. Results Probl. Cell Differ. 45:279–308
100. Bowler MW, Montgomery MG, Leslie AG, Walker JE. 2006. How azide inhibits ATP hydrolysis by theF-ATPases. Proc. Natl. Acad. Sci. USA 103:8646–49
101. Experimentsdefining therequirements for ATPsynthesis by purifiedreconstituted ATPsynthases.
101. Wiedenmann A, Dimroth P, von Ballmoos C. 2009. Functional asymmetry of the F0 motor inbacterial ATP synthases. Mol. Microbiol. doi: 10.1111/j.1365-2958.2009.06658.x
672 von Ballmoos ·Wiedenmann · Dimroth
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

AR378-FM ARI 7 May 2009 15:43
Annual Review ofBiochemistry
Volume 78, 2009
ContentsPreface � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �v
Prefatory Articles
FrontispieceE. Peter Geiduschek � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �xii
Without a License, or Accidents Waiting to HappenE. Peter Geiduschek � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
FrontispieceJames C. Wang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �30
A Journey in the World of DNA Rings and BeyondJames C. Wang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �31
Biochemistry and Disease Theme
The Biochemistry of Disease: Desperately Seeking SyzygyJohn W. Kozarich � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �55
Biosynthesis of Phosphonic and Phosphinic Acid Natural ProductsWilliam W. Metcalf and Wilfred A. van der Donk � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �65
New Antivirals and Drug ResistancePeter M. Colman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �95
Multidrug Resistance in BacteriaHiroshi Nikaido � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 119
Conformational Pathology of the Serpins: Themes, Variations,and Therapeutic StrategiesBibek Gooptu and David A. Lomas � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 147
Getting a Grip on Prions: Oligomers, Amyloids, and PathologicalMembrane InteractionsByron Caughey, Gerald S. Baron, Bruce Chesebro, and Martin Jeffrey � � � � � � � � � � � � � � � � � 177
Ubiquitin-Mediated Protein Regulation
RING Domain E3 Ubiquitin LigasesRaymond J. Deshaies and Claudio A.P. Joazeiro � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 399
Regulation and Cellular Roles of Ubiquitin-SpecificDeubiquitinating EnzymesFrancisca E. Reyes-Turcu, Karen H. Ventii, and Keith D. Wilkinson � � � � � � � � � � � � � � � � � � � � 363
vi
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

AR378-FM ARI 7 May 2009 15:43
Recognition and Processing of Ubiquitin-Protein Conjugatesby the ProteasomeDaniel Finley � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 477
Degradation of Activated Protein Kinases by UbiquitinationZhimin Lu and Tony Hunter � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 435
The Role of Ubiquitin in NF-κB Regulatory PathwaysBrian Skaug, Xiaomo Jiang, and Zhijian J. Chen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 769
Biological and Chemical Approaches to Diseases of ProteostasisDeficiencyEvan T. Powers, Richard I. Morimoto, Andrew Dillin, Jeffery W. Kelly,and William E. Balch � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 959
Gene Expression
RNA Polymerase Active Center: The Molecular Engineof TranscriptionEvgeny Nudler � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 335
Genome-Wide Views of Chromatin StructureOliver J. Rando and Howard Y. Chang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 245
The Biology of Chromatin Remodeling ComplexesCedric R. Clapier and Bradley R. Cairns � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 273
The Structural and Functional Diversity of Metabolite-BindingRiboswitchesAdam Roth and Ronald R. Breaker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 305
Lipid and Membrane Biogenesis
Genetic and Biochemical Analysis of Non-Vesicular Lipid TrafficDennis R. Voelker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 827
Cholesterol 24-Hydroxylase: An Enzyme of Cholesterol Turnoverin the BrainDavid W. Russell, Rebekkah W. Halford, Denise M.O. Ramirez, Rahul Shah,and Tiina Kotti � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �1017
Lipid-Dependent Membrane Protein TopogenesisWilliam Dowhan and Mikhail Bogdanov � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 515
Single-Molecule Studies of the Neuronal SNARE Fusion MachineryAxel T. Brunger, Keith Weninger, Mark Bowen, and Steven Chu � � � � � � � � � � � � � � � � � � � � � � � 903
Mechanisms of EndocytosisGary J. Doherty and Harvey T. McMahon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 857
Contents vii
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.

AR378-FM ARI 7 May 2009 15:43
Recent Advances in Biochemistry
Motors, Switches, and Contacts in the ReplisomeSamir M. Hamdan and Charles C. Richardson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 205
Large-Scale Structural Biology of the Human ProteomeAled Edwards � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 541
Collagen Structure and StabilityMatthew D. Shoulders and Ronald T. Raines � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 929
The Structural and Biochemical Foundations of Thiamin BiosynthesisChristopher T. Jurgenson, Tadhg P. Begley, and Steven E. Ealick � � � � � � � � � � � � � � � � � � � � � � � � 569
Proton-Coupled Electron Transfer in Biology: Results fromSynergistic Studies in Natural and Model SystemsSteven Y. Reece and Daniel G. Nocera � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 673
Mechanism of Mo-Dependent NitrogenaseLance C. Seefeldt, Brian M. Hoffman, and Dennis R. Dean � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 701
Inorganic Polyphosphate: Essential for Growth and SurvivalNarayana N. Rao, Marıa R. Gomez-Garcıa, and Arthur Kornberg � � � � � � � � � � � � � � � � � � � � � 605
Essentials for ATP Synthesis by F1F0 ATP SynthasesChristoph von Ballmoos, Alexander Wiedenmann, and Peter Dimroth � � � � � � � � � � � � � � � � � � 649
The Chemical Biology of Protein PhosphorylationMary Katherine Tarrant and Philip A. Cole � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 797
Sphingosine 1-Phosphate Receptor SignalingHugh Rosen, Pedro J. Gonzalez-Cabrera, M. Germana Sanna, and Steven Brown � � � � 743
The Advent of Near-Atomic Resolution in Single-Particle ElectronMicroscopyYifan Cheng and Thomas Walz � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 723
Super-Resolution Fluorescence MicroscopyBo Huang, Mark Bates, and Xiaowei Zhuang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 993
Indexes
Cumulative Index of Contributing Authors, Volumes 74–78 � � � � � � � � � � � � � � � � � � � � � � � � � �1041
Cumulative Index of Chapter Titles, Volumes 74–78 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �1045
Errata
An online log of corrections to Annual Review of Biochemistry articles may be found athttp://biochem.annualreviews.org/errata.shtml
viii Contents
Ann
u. R
ev. B
ioch
em. 2
009.
78:6
49-6
72. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by N
ew Y
ork
Uni
vers
ity -
Bob
st L
ibra
ry o
n 11
/20/
13. F
or p
erso
nal u
se o
nly.