esterase variations between the 14 syngens · pdf file · 2003-07-24esterase...
TRANSCRIPT

ESTERASE VARIATIONS BETWEEN THE 14 SYNGENS OF PARAMECZUM AURELZA UNDER AXENIC GROWTH1
S . L. ALLEN, S. W. FARROW AND P. A. GOLEMBIEWSKI
Departments of Botany and Zoology, The University of Michigan, Ann Arbor, Michigan 48104
Manuscript received October 11, 1972 Revised copy received December 14, 1972
ABSTRACT
Under axenic growth all 14 syngens of Paramecium aurelia have 4 types of esterases. The three major types (A, B and cathodal C) vary independently in electrophoretic mobility among the syngens. Using these three esterases, stocks can be keyed to a syngen, except for the groupings 1-3-5 and 7-13. Using 5 other enzymes only syngens 1 and 5 cannot be distinguished. Most syngens differ from each other in 6 out of the 8 enzymes. An axenically-grown stock of Paramecium multimicronucleatum collected in Costa Rica has the same types of esterases as P. aurelia. Two of the types (A and C) are similar in mo- bility to those found in syngens 7 and 13, but its B esterase differs in mobility from all the known syngens of P. aurelia.
E have been concerned with the distribution of esterases present in the Wspecies complex Paramecium aurelia, with a description of the types of esterases and the degree of variation within a type within and between the 14 sibling species or syngens. Collections of P. aurelia have been made from ponds and streams throughout most of the land masses of the world by SONNEBORN, BEALE and others; a laboratory stock is established from a single autogamous individual and the stock is assigned to a particular syngen on the basis of its ability to exchange genes with other individuals in that syngen. Because of the procedure of selecting single autogamous cells, the laboratory stocks are homozy- gous for all their genes. All the stocks used in our study were homozygous.
The stocks can be grown on bacteria or in a bacteria-free medium which we refer to as “axenic medium.” Not surprisingly, bacteria contain esterases, and when paramecia are grown on bacteria additional esterases appear on the gels and effects on Paramecium esterases are observed (ALLEN and GOLEMBIEWSKI 1972; GIBSON and CAVILL 1972; ROWE, GIBSON and CAVILL 1971). In earlier reports we compared the esterases of all fourteen syngens in bacterized stocks (ALLEN, BYRNE and CRONKITE 1971) and in axenic stocks of six of the syngens (ALLEN and GIBSON 1971). Axenic stocks from the remaining eight syngens have now been prepared. This permits us to extend the analysis of the esterases from axenically-grown stocks to all fourteen syngens. We are able to show that
Supported by a research grant, GM-15879, from the National Institute of General Medical Sciences, Public Health Service.
Gene:ics 73: 561-573 April 1973.

5 62 S. L. ALLEN, S. W. FARROW AND P. A. GOLEMBIEWSKI
each syngen has four different types of esterases which are characterized on the basis of substrate specificity and sensitivity to the inhibitor, eserine sulfate, and that differences in mobility and/or activity occur between syngens for esterase types A, B and C. Each syngen or group of syngens has a characteristic array of subtype;.
MATERIALS AND METHODS
Stocks: The 39 axenic stocks fom syngens 1, 2, 4, 5, 6, and 8, previously tested, appear in Table 1 in ALLEN and GIBSON (1971). Those from syngens 3, 7, 9, 10, 11, 12, 13, and 14 are listed in Table I of this paper. Cultures of the latter set of stocks growing on bacteria were kindly sent by MYRTLE V. SCHNELLER at Indiana University. Samples of each of these stocks were rendered bacteria-free by handwashing in the antibiotic neamine* (0.2-1 .O mg/ml final con- centration) and introduced into axenic medium (SOLDO, GODOY and VAN WACTENDONK 1966). For some syngens (3, 9, 10, and 11) considerable variation i n the method was needed before we achieved success. Some of the stocks only tolerated low concentrations of the antibiotic and others required a long period of adaptation on diluted axenic medium (1: 1 with DRYL'S salts solution; DRYL 1959) before they would survive on undiluted axenic medium. The stocks from syngen 11 presented the most difficulty, and the one we managed to put on to axenic medium grows very poorly and requires a high surface-to-volume ratio for growth. Most of the other stocks now grow very well on the axenic medium, although some grew poorly initially. Tubed cultures of axenically-grown paramecia are maintained at 16°C with bimonthly subculture. The slow- growing stocks and cold-sensitive stocks (e.g. 6/225) are maintained at 23°C with weekly sub- culture.
Identification of esterases: For enzyme extracts all cultures were grown at 23"C, hanested by centrifugation, and extracts of the cell concentrates were made by freeze-thawing. Electro- phoresis of whole-cell extracts was carried Out in 12% starch gels using Connaught starch or Electrostarch, as described previously (ALLEN, BYRNE and CRONKITE 1971 ; ALLEN and GIBSON 1971; ALLEN and GOLEMBIEWSKI 1972). Two substrates were used: a-naphthyl propionate and a-naphthyl butyrate; and one inhibitor, eserine sulfate, at varying concentrations. All esterases are visualized in a reaction mixture which includes a-naphthyl propionate, while one class is missing (the A esterases) in the reaction mixture containing a-naphthyl butyrate. Details of the histochemical procedures may be found elsewhere (ALLEN and GIBSON 1971).
RESULTS
The following kinds of tests of the new stocks listed in Table 1 were made: (1) the esterases of each stock were compared with those of all other stocks in the same syngen; (2) where available, two stocks from each syngen were used in tests of the sensitivity of the esterases to four concentrations of eserine sulfate; (3) the esterases of one-two stocks from each of the new syngens were compared with those of two stocks from each of the other syngens, including the six syngens previously studied (ALLEN and GIBSON 1971). In this section we therefore con- sider the results obtained on all 14 syngens of P. aurelia.
In each of the fourteen syngens four classes of esterases are found, as described previously for six syngens (ALLEN and GIBSON 1971). The A esterases react specifically with a-naphthyl propionate and are inhibited by to NI eserine sulfate. The B esterases have a greater affinity for a-naphthyl butyrate than for a-naphthyl propionate and are only inhibited by eserine sulfate when
* A gift of the Upjohn Company, Kalamazoo, Mich.

ESTERASE VARIATIONS BETWEEN SYNGENS O F PARAMECIUM 563 TABLE 1
Axenic stocks of paramecium
P. aurelia syngen Stock Number’ Geographic origin
3 112 Illinois 152 Connecticut 161 Minnesota 181 Michigan 231 Vermont 283 Colorado
? 38 Florida 227 Florida
9 204 Scotland 317 France 323 Scotland 338 U.S.S.R.
10 223 Florida 11 319 Delaware 12 270 Florida
273 Louisiana 13 209 France
321 Mexico 14 328 Australia
P. multimicronucleatum 8M0 Costa Rica
* Stocks designated by numbers 1-4.99 were isolated by SONNEBORN, those numbered from 500 on by BEALE, and the He series by GIBSON (see Table 1, ALLEN and GIBSON 1971). Originally stocks 1-26 were designated A to Z; thus, stock P, listed in ALLEN and GIESON (1971) and in ALLEN, BYRNES and CRONKITE (1971) is the same as stock 16. Some of these stocks were obtained directly from the isolators; others were from people to whom they had been sent, as indicated when another place or name, or both, appears in the tables of the two previous papers under the heading “Source”. All of the stocks except 8M0 in the present paper were received directly from SONNEBORN’S laboratory at Indiana University; stock 8M0 was received from J. H. VANDERMEER at The University of Michigan.
the concentration is raised M. The C and D esterases are resistant to 10-1 M eserine sulfate and react with both substrates, the C esterases having a greater affinity for a-naphthyl propionate than for a-naphthyl butyrate and the D ester- ases having equal affinity for both substrates.
The A esterases are typically isozymic. Usually three isozymes are found, with the more cathodal isozyme being the most active. More than three isozymes may occur in stocks of syngen 2 with as many as five isozymes being observed in a single stock (ALLEN and GOLEMBIEWSKI 1972).
A single B esterase is found in each stock and in each syngen. An exception may be syngen 4 where two isozymes are resolved in Electrostarch. The cathodal C esterase varies in activity in different stocks and in certain stocks appears as a doublet. There are, in addition, some rapidly-migrating anodal C esterases which occur in certain syngens (2,8,10,11 and 12). The D esterases are erratic in their behavior but when present they migrate to a region about four cm from the anode in Connaught starch.
The migration of the esterases is affected by the brand of starch used (compare

564 S. L. ALLEN, S. W. FARROW A N D P. A. GOLEMBIEWSKI
-4
Cm (a) Esterases of Axenic S t o c k s -5 1
I
I_ - - - . - - - - -- - - - - - - - - - - _ _ _ _ -
m Y E!!
-1 -
4
1 r---7 I -
+7 I ~. , 1 2 3 4 5 6 7 8 9 1 0 11 1 2 1 3 1 4
S y n g e n s
FIGURE 1.-Diagrams of the esterases in axenic stocks of thet 14 syngens of P. aurelia based on measurements of hundreds of gels (a) Connaught starch (b) Electrostarch. Types of esterases are represented by the following symbols: dark bars-A type, diagonal slashes-B type, light bars-C type, and vertical slashe-D type. There is considerable variation between stocks of syngen 2 in the mobilities of the A esterases. Those shown are for stock 93. The star in (b) indi-
ESTERASE VARIATIONS B E T W E E N S Y N G E N S O F PARAMECIUM
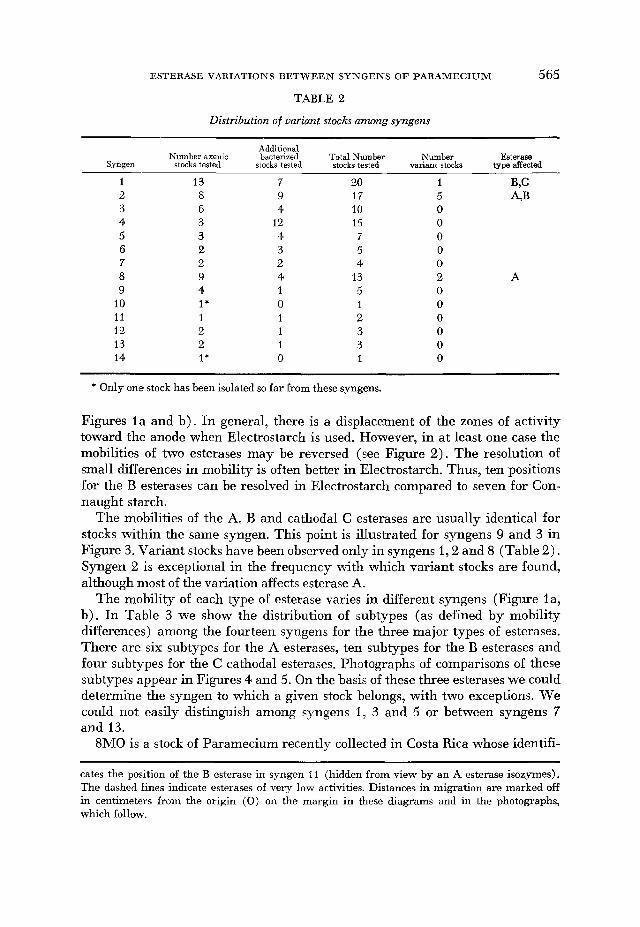
TABLE 2
Distribution of variant stocks a m n g syngens
565
Additional Number axenic bacterized Total Number Number Esterase
Syngen stocks tested stocks tested stocks tested variant stochs type affected
1 13 7 U) 1 B,C 2 3 4 5 6 7 8 9
IO 11 12 13 14
8 6 3 3 2 2 9 4 1* 1 2 2 I*
9 17 4 10
12 15 4 7 3 5 2 4 4 13 1 5 0 1 1 2 1 3 1 3 0 1
5 A,B 0 0 0 0 0 2 0 0 0 0 0 0
A
* Only one stock has been isolated so far from these syngens.
Figures l a and b). In general, there is a displacement of the zones of activity toward the anode when Electrostarch is used. However, in at least one case the mobilities of two esterases may be reversed (see Figure 2). The resolution of small differences in mobility is often better in Electrostarch. Thus, ten positions for the B esterases can be resolved in Electrostarch compared to seven for Con- naught starch.
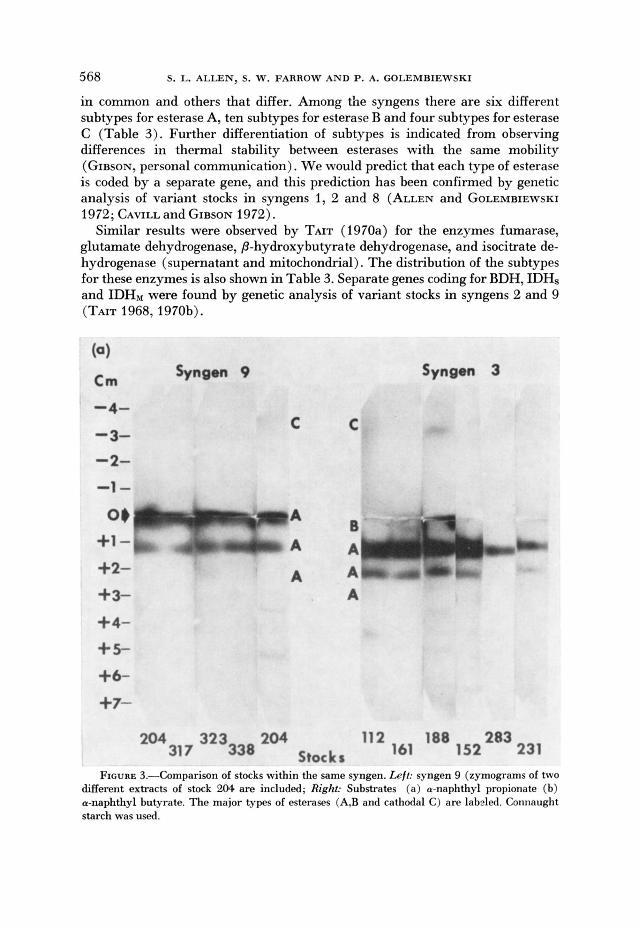
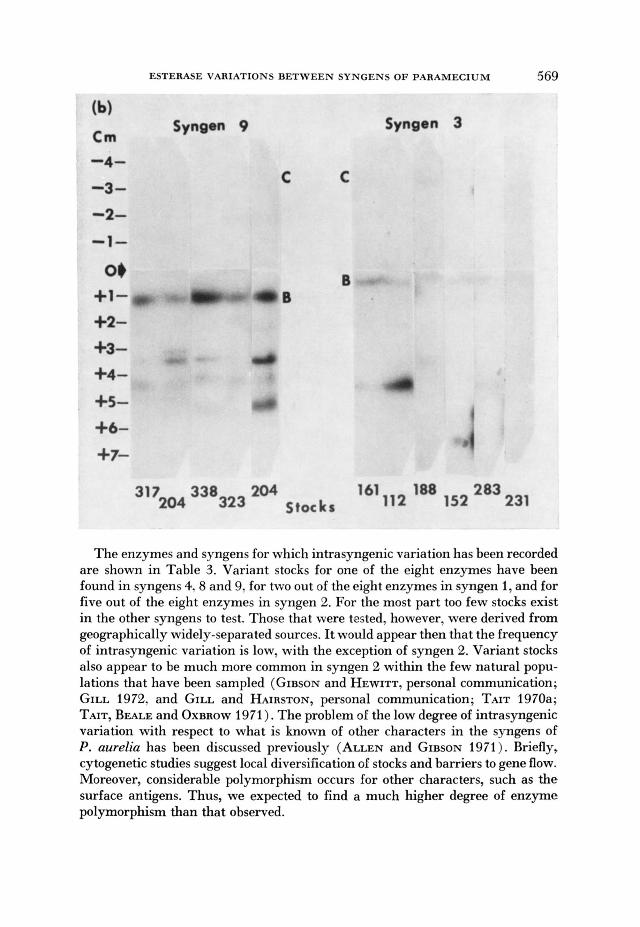
The mobilities of the A, B and catliodal C esterases are usually identical for stocks within the same syngen. This point is illustrated for syngens 9 and 3 in Figure 3. Variant stocks have been observed only in syngens 1,2 and 8 (Table 2). Syngen 2 is exceptional in the frequency with which variant stocks are found, although most of the variation affects esterase A.
The mobility of each type of esterase varies in different syngens (Figure la, b). In Table 3 we show the distribution of subtypes (as defined by mobility differences) among the fourteen syngens for the three major types of esterases. There are six subtypes for the A esterases, ten subtypes for the B esterases and four subtypes for the C cathodal esterases. Photographs of comparisons of these subtypes appear in Figures 4 and 5. On the basis of these three esterases we could determine the syngen to which a given stock belongs, with two exceptions. We could not easily distinguish among syngens 1, 3 and 5 or between syngens 7 and 13.
8MO is a stock of Paramecium recently collected in Costa Rica whose identifi- ~
cates the position of the B esterase in syngen 11 (hidden from view by a n A esterase isozymes). The dashed lines indicate esterases of very low activities. Distances in migration are marked off in centimeters from the origin (0) on the margin in these diagrams and in the photographs, which follow.

566 S. L. ALLEN, S. W. FARROW AND P. A. GOLEMBIEWSKI
I
FIGURE 2.-Comparison of two brands of starch in resolving the esterases in syngens 9 and 10. Note the displacement of the A and B esterases toward the anode in Electrostarch. The syngen difference in the mobility of the A esterases is similar in both starches, but note that the relative mobilities of the B esterases are reversed in the two syngens in thc different starches.

ESTERASE VARIATIONS BETWEEN SYNGENS O F P A R A M E C I U M
TABLE 3
Distribution of enzyme subtypes among syngens
567
Enzyme EsA EsB EsC Fumarase+ GDHt BDHt IDHSt IDH,t
* Using Electrostarch. t Data from TAIT (197Oa). 4 No migration into gel.
T I Variant stocks show multiple zones of activity unlike “normal”. ‘I Variant stock differs in activity. Abbreviations: EsA = Esterase A; EsB = Esterase B; EsC = Esterase C; GDH = glutamate
dehydrogenase; BDH = P-hydroxybutyrate dehydrogenase; IDH, = isocitrate dehydrogenase (supernatant) ; IDH, = isocitrate dehydrogenase (mitochondrial).
Variant stocks show change in mobility.
cation was unknown at the time we received it. Subsequent to our tests, it was identified as P. multimicronucleatum by SONNEBORN and DIPPELL (personal communication). This Paramecium is found exclusively in the brachts of Heli- conia imbricata, H . bahui and Caluthio insignis (JOHN VANDERMEER, personal communication). In this temporary environment it develops dense populations. Attempts to key it to syngen by mating type tests failed because it selfs vigorously. The animals are small and roughly similar in size to those found in syngens 4 and 8 of P. aurelia. In the laboratory it grows with vigor and develops enormous popu- lations in exenic medium. The identification of 8MO was tested by means of its esterases. It has the four classes of esterases of P. aurelia, as determined by sub- strate and inhibitor studies. Its types A and C esterases have mobilities similar to those found in syngen 13 of P. aurelia (Figure 6 ) . Its B esterase has a mobility which sets it apart from all the known syngens o i P. aure2ia.
DISCUSSION
Four types of esterases can be recognized in axenic stocks of all fourteen syn- gens of P. aurelia. These types differ in substrate specificity, sensitivity to the inhibitor eserine sulfate, and electrophoretic mobility. They also occur in P . multimicronucleatum (SMO) , a species closely related to P. aurelia ( SONNEBORN 1958). It is therefore of interest that the esterases of these two species are so similar. In fact, only one type of esterase distinguishes 8MO from syngens 7 and 13 in P. aurelia.
Clear-cut differences in electrophoretic mobility occur fo r each of the three major types of esterases (A, B and cathodal C) among the syngens of P. aurelia, and each type varies independently. Thus; some of the syngens have certain types
568 S. L. ALLEN, S. W. FARROW AND P. A. GOLEMBIEWSKI
in common and others that differ. Among the syngens there are six different subtypes for esterase A, ten subtypes for esterase B and four subtypes for esterase C (Table 3). Further differentiation of subtypes is indicated from observing differences in thermal stability between esterases with the same mobility (GIBSON, personal communication). We would predict that each type of esterase is coded by a separate gene, and this prediction has been confirmed by genetic analysis of variant stocks in syngens 1, 2 and 8 (ALLEN and GOLEMBIEWSKI 1972; CAVILL and GIBSON 1972).
Similar results were observed by TAIT (1970a) for the enzymes fumarase, glutamate dehydrogenase, /3-hydroxybutyrate dehydrogenase, and isocitrate de- hydrogenase (supernatant and mitochondrial). The distribution of the subtypes for these enzymes is also shown in Table 3. Separate genes coding for BDH, IDHs and IDH, were found by genetic analysis of variant stocks in syngens 2 and 9 (TAIT 1968,1970b).
I 2-
3-
4-
5-
6-
FIGURE 3.-Comparison of stocks within the same syngen. h j i : syngen 9 (zymograms of two merent extracts of stock u)4. are included; Right: Substrates (a) a-naphthyl propionate (b) a-naphthyl butyrate. The major types of esterases (A,B and cathodal C) are labeled. Connaught starch was used.
ESTERASE VARIATIONS BETWEEN SYNGENS OF PARAMECIUM 569
The enzymes and syngens for which intrasyngenic variation has been recorded are shown in Table 3. Variant stocks for one of the eight enzymes have been found in syngens 4,8 and 9, for two out of the eight enzymes in syngen 1, and for five out of the eight enzymes in syngen 2. For the most part too few stocks exist in the other syngens to test. Those that were tested, however, were derived from geographically widely-separated sources. It would appear then that the frequency of intrasyngenic variation is low, with the exception of syngen 2. Variant stocks also appear to be much more common in syngen 2 within the few natural popu- lations that have been sampled (GIBSON and HEWITT, personal communication; GILL 1972, and GILL and HAIRSTON, personal communication; TAIT 1970a; TAIT, BEALE and OXBROW 1971). The problem of the low degree of intrasyngenic variation with respect to what is known of other characters in the syngens of P. aurelia has been discussed previously (ALLEN and GIBSON 1971). Briefly, cytogenetic studies suggest local diversification of stocks and barriers to gene flow. Moreover, considerable polymorphism occurs for other characters, such as the surface antigens. Thus, we expected to find a much higher degree of enzyme polymorphism than that observed.

FIG
UR
E 4
.--C
ompa
rison
of
est
eras
es A
, B a
nd c
atho
dal C
in t
he fo
urte
en sy
ngen
s usi
ng E
lect
rost
arch
. The
subs
trat
e is
a-na
phth
yl p
ropi
onat
e w
hich
rev
eals
the
A a
nd C
est
eras
es w
ell b
ut th
e B
este
rase
s onl
y fa
intl
y. A
n at
tem
pt h
as h
een
mad
e to
ord
er th
e sy
ngen
s hy
thei
r A
est
eras
e m
obili
ties,
thos
e on
the
left
hav
ing
the
mor
e ca
thod
al po
sitions.

9 10
14
13
14
10
S
vnaa
01
FIGU
RE
5.--C
ompa
riso
n of
the
B e
ster
ases
in t
he fourteen
syng
ens using E
lect
rost
arch
. The
subs
trat
e is
a-na
phth
yl bu
tyra
te
for
whi
ch t
he B
esterases h
ave
a gr
eate
r aN
iity
. Her
e, an
atte
mpt
has
bee
n m
ade t
o or
der
the
syng
ens b
y th
e po
sitio
n of
the
ir
B es
tera
ses.
2

5 72 S. L. ALLEN, S. W. FARROW AND P. A. GOLEMBIEWSKI
FIGURE 6.-Comparison of 8M0 with syngen 13. Note the similarity in mobility of the A and C esterases and the difference in mobility of the B esterases. The mobility of the B esterases in 8M0 is 1.7 cm, which is faster than that in Syngen 13, with a mobility of 1.2 cm, and that in Syngen 11, with a mobility of 1.5 cm. Electrostarch was used.

ESTERASE VARIATIONS BETWEEN SYNGENS O F PARAMECIUM 5 73
All the syngens of P. uurelia can be distinguished on the basis of which sub- types of the eight enzymes are present, except lor syngens 1 and 5. Some syngens differ for all eight enzymes (2 and 13; 7 and 10; 11 and 12). Some differ for only three enzymes (1-5 and 3,9 and 14; 2 and 3). Most syngens differ in six out of the eight enzymes.
Intersyngenic mating has been observed for 1 and 5, but also fo r 1 and 3 , l and 7, 4 and 8, and 3 and 8 (SONNEBORN 1957). On the basis of several biological criteria (mating type inheritance, serotype similarities and differences, symbiont distribution, presence and absence of a strong cytoplasmic influence) SONNEBORN (1957, 1958) suggested that there was an ancient evolutionary division of the syngens of P. aurelia into two or three groups and that syngen 1 gave rise to syngens 3, 5, 7, 9 and 11 and syngen 4 to 8 and 10. The enzyme data suggest a more complicated relationship for the syngens. These data can be used to explore evolutionary distances mathematically and will be the subject of a future com- munication.
LITERATURE CITED
ALLEN, S. L. and I. GIBSON, 1971 Intersyngenic variations in the esterases of axenic stocks of Paramecium aurelia. Biochem. Genetics 5: 161-181.
ALLEN, S. L. and P. A. GOLEMBIEWSKI, 1972 Inheritance of esterases A and B in syngen 2 of Paramecium aurelia. Genetics 71 : 469473.
ALLEN, S. L., B. C. BYRNE and D. L. CRONKITE, 1971 Intersyngenic variations in the esterases of bacterized Paramecium aurelia. Biochem. Genetics 5 : 135-150.
CAVILL, A. and I. GIBSON, 1972 Genetic determination of esterases in Paramecium aurelia. Heredity 38: 31-37.
DRYL, S., 1959 Antigenic transformation in Paramecium aurelia after homologous antiserum treatment during autogamy and conjugation. J. Protozool. 6 (Suppl.) : 25 (Abstr.).
GIBSON, I. and A. CAVILL, 1973 The effects of bacterial products on a Paramecium esterase. Biochem. Genetics (in press).
GILL, D. E., 1972 Intrinsic rates of increase, saturation densities, and competitive ability. 1. An experiment with Paramec:um. Am. Nat. 106: 461-471.
ROWE, E., I. GIBSON and A. CAVILL, 1971 The effects of growth conditions on the esterases of Parameckm aurelia. Biochem. Genetics 5 : 151-159.
SOLDO, A. T., G. A. GODOY and W. J. VAN WAGTENDONK, 1966 Growth of particle-bearing and particle-free Paramecium aurelia in axenic culture. J. Protozool. 13 : 494-497.
SONNEBORN, T. M., 1957 Breeding systems, reproductive methods and species problems in protozoa. pp. 155-324. In: The Species Problem. Edited by E. MAYR. Am. Assoc. Adv. Sci. Symp., Washington, D.C. -, 1958 Classification of syngens of the Paramechm aurelia-multimicronucleatum complex. J. Protozool. 5 (Suppl.) : 17 (Abstr.) .
Genetic control of P-hydroxybutyrate dehydrogenase in Paramecium aurelia. Nature 219: 941. - , 1970a Enzyme variation between syngens in Paramecium aurelia. Biochem. Genetics 4: 461470. - , 1970b Genetics of NADP-dependent iso- citrate dehydrogenase in Paramecium aurelia. Nature 225: 181-182.
Enzyme polymorphism in a population of Paramecium aurelia in S.E. England. J. Protozool. 18 (Suppl.) : 26 (Abstr.).
TAIT, A., 1968
TAIT, A., G. H. BEALE and A. R. OXBROW, 1971