eukaryotic gene regulation: role of chromatin lecture 7.pdf · by covalent modification of histone...
TRANSCRIPT

Eukaryotic gene regulation: Role of chromatin

Recap……………..
• Eukaryotic RNA polymerases
• Core promoter elements
• General transcription factors
• Enhancers and upstream activation sequences
• Transcriptional activators: DNA binding, transactivation

In eukaryotes, the cellular DNA is tightly wrapped up around histones and is organized in the form ofnucleosomes.
How do transcription factors gain access to the promoter DNA?
How are histones displaced from the promoter during transcription initiation?
Nucleosome core particle 147 bp DNA + octamerDNA
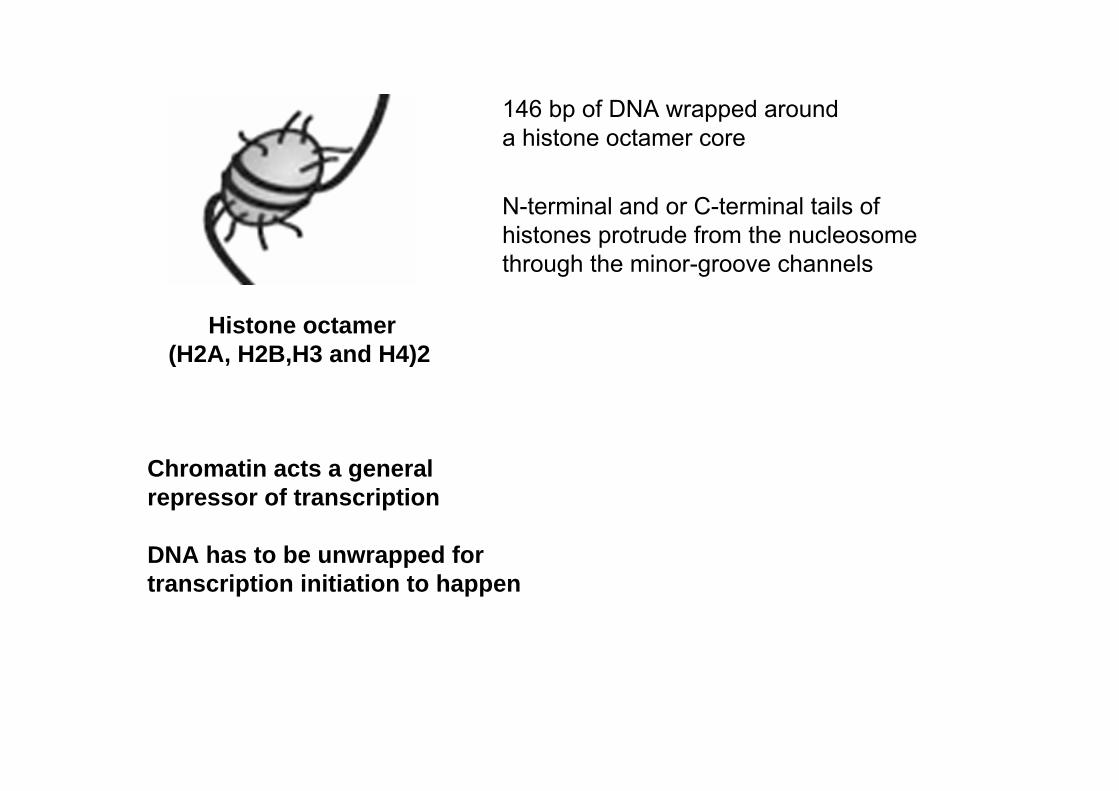
N-terminal and or C-terminal tails of histones protrude from the nucleosome through the minor-groove channels
Histone octamer (H2A, H2B,H3 and H4)2
146 bp of DNA wrapped arounda histone octamer core
Chromatin acts a general repressor of transcription
DNA has to be unwrapped for transcription initiation to happen

Chromatin compaction affects transcriptional activity in eukaryotic nuclei
Heterochromatin - transcriptionally silent
Euchromatin – transcriptionally active
Densely packed chromatin will block access of RNA pol II + General transcription factors to core promoters
Chromatin needs to be unpacked, unwrapped or opened before transcription initiation can happen

Transcription Activation in eukaryotes:
step 1: chromatin structure should be modified such that RNA polymerase II machinery can be recruited
step 2: Actual recruitment of RNA polymerase II machinery
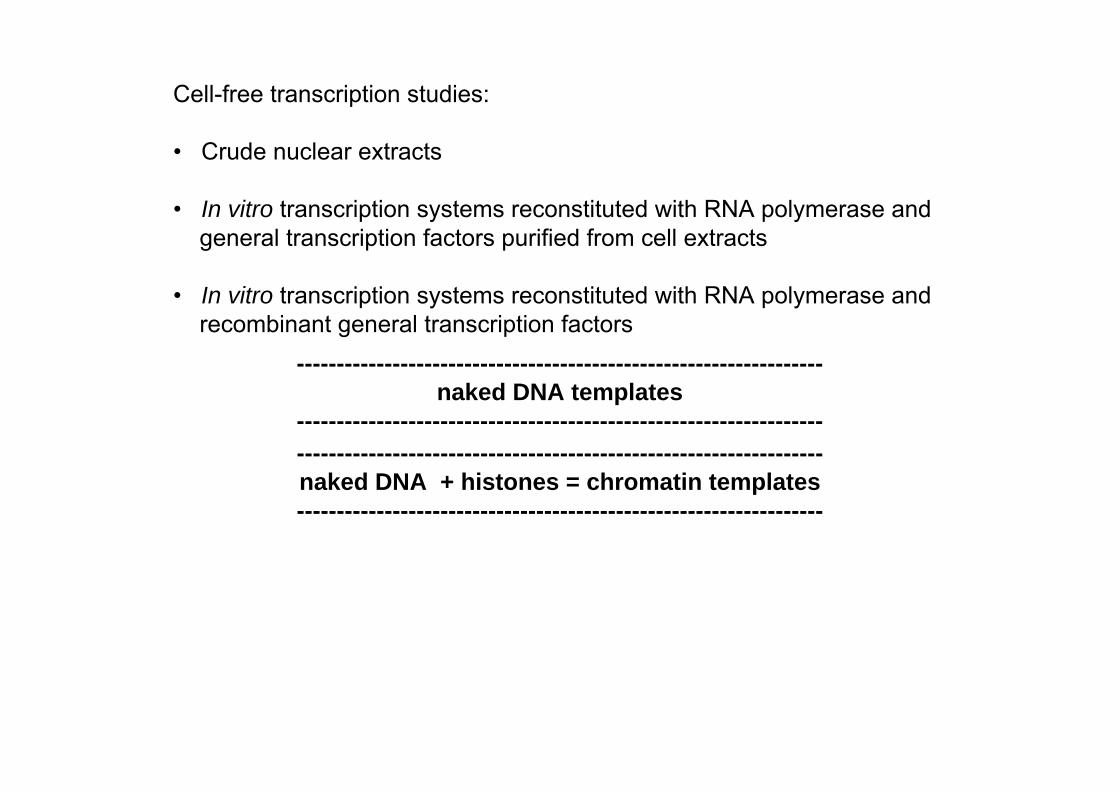
Cell-free transcription studies:
• Crude nuclear extracts
• In vitro transcription systems reconstituted with RNA polymerase and general transcription factors purified from cell extracts
• In vitro transcription systems reconstituted with RNA polymerase and recombinant general transcription factors
------------------------------------------------------------------naked DNA + histones = chromatin templates ------------------------------------------------------------------
------------------------------------------------------------------naked DNA templates
------------------------------------------------------------------

TBP cannot bind nucleosomal DNA
Nucleosomes prevent binding of TBP to core promoter elements in vitro
TBP does not associate with most core promoters in vivo in the absence of transcriptional activators
CHROMATIN TEMPLATES NEED TO BE MODIFIED IN ORDER FOR TRANSCRIPTION FACTORS TO BIND
HOW?
BY COVALENT MODIFICATION OF HISTONE TAILS

Histones can undergo several post translational modifications
ACETYLATION
METHYLATION
PHOSPHORYLATION
UBIQUITINYLATION
SUMOYLATION
ADP RIBOSYLATION
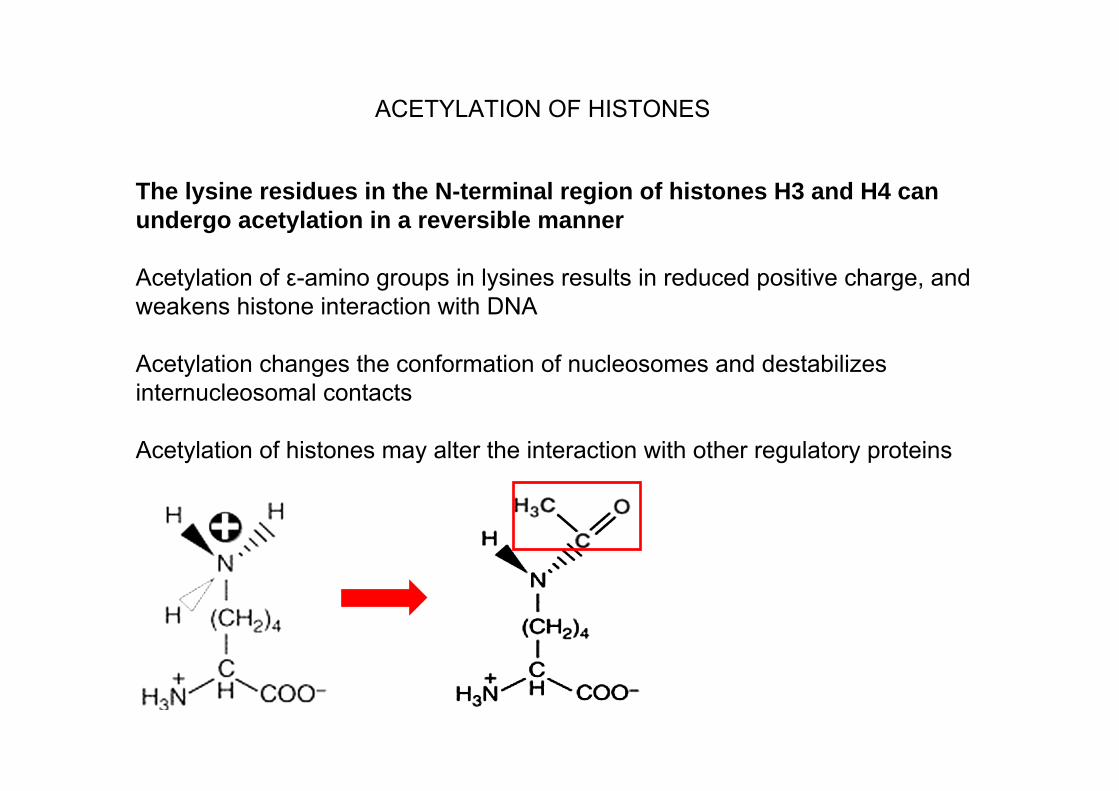
The lysine residues in the N-terminal region of histones H3 and H4 can undergo acetylation in a reversible manner
Acetylation of ε-amino groups in lysines results in reduced positive charge, and weakens histone interaction with DNA
Acetylation changes the conformation of nucleosomes and destabilizes internucleosomal contacts
Acetylation of histones may alter the interaction with other regulatory proteins
ACETYLATION OF HISTONES

Heterochromatin - transcriptionally silent
Euchromatin – transcriptionally active
Acetylated histones are enriched in transcribed chromatin
Acetylation of histones potentiates transcription by making nucleosomal DNA more accessible for transcription factor binding

How are histones acetylated?
Many proteins involved in transcriptional activation
(transcription factors/coactivators/TAFs)
are
histone acetyl transferases
(HATs)

Gcn5p, a well known transcriptional regulator in yeast cells was shown to possess HAT activity
The human homologue of Gcn5p is also a HAT
For the first time, researchers working on histones and chromatin structure got interested in transcriptional regulation
Similarly, researchers working on transcriptional regulation realized that they cannot ignore chromatin any longer
Gcn5, the first nuclear histone acetylase to be identified
Yeast Gcn5 was found in at least two distinct multiprotein complexes, Ada and SAGA, neither of which is tightlyassociated with TFIID or the Pol II
Brownell et al., 1996 Cell 84: 843-851

Histone acetyltransferases (HATs)
HAT- A HAT-B
A-Type HATs catalyze transcription-related acetylations
B-Type HATs are cytoplasmic. They catalyze acetylations linked to transport of newly synthesized
histones from the cytoplasm to the nucleus

GNAT familyGcn5-related N-acetyltransferase (bromodomains)
MYST familyMOZ, Ybf2/Sas3, Sas2, Tip60 (chromodomains)
CBP/p300
GTF-HATsTAFII250
Nuclear receptor linked HATs SRC1, ACTR
HAT FAMILIES
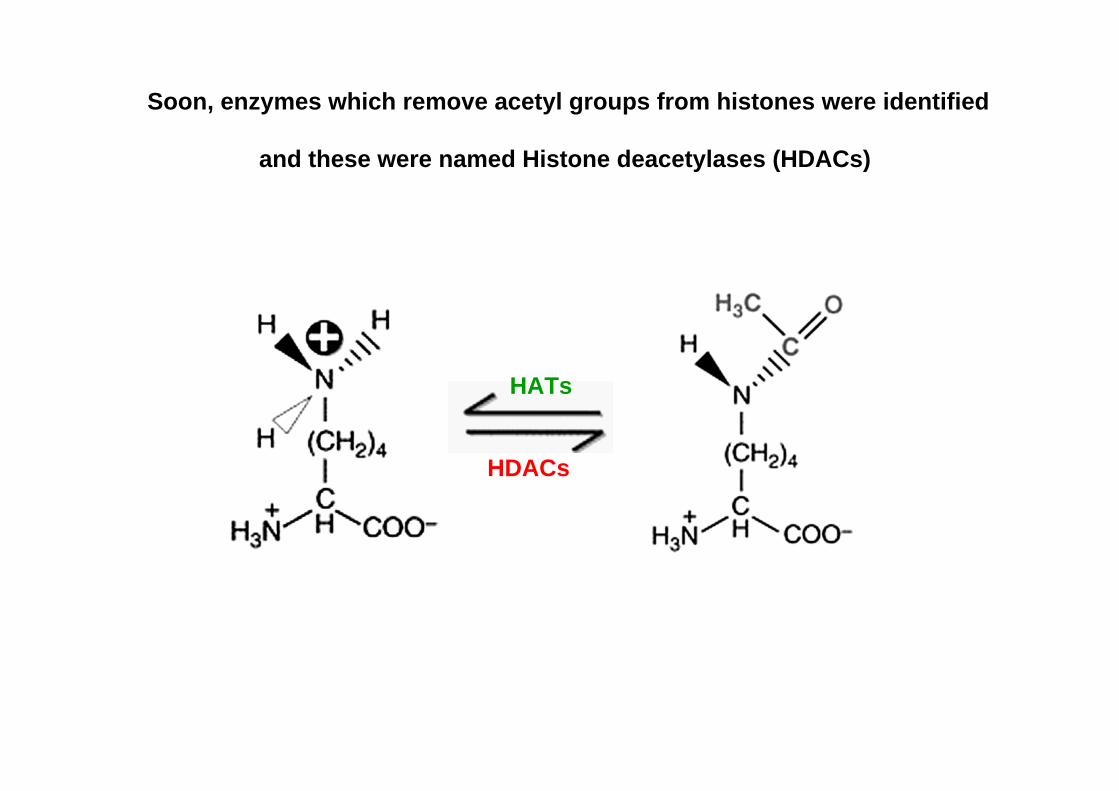
Soon, enzymes which remove acetyl groups from histones were identified
and these were named Histone deacetylases (HDACs)
HATs
HDACs

HATs such as Gcn5p are present as huge multi protein complexes
SAGA (Spt-Ada-Gcn5-Acetyltransferase)
Human Gcn5p was also found in a similar complex STAGA
These complexes were found to Interact with both TBP and acidic activators
The GNAT family of HATs include not only Gcn5p but also PCAF, TFTC (TBP-free TAF-containing complex) etc.
Free Gcn5 acetylated only free histones, but as part of the SAGA-complex it couldacetylate nucleosomes

Several coactivators have HAT activity
hTAF250, dTAF230, yTAF130p300/CBPACTR and SRC-1P/CAF
MICE IN WHICH THESE HATs ARE KNOCKED OUTEXHIBITED SEVERAL DEVELOPMENTAL DEFECTS
Each of these HATs acetylated specific lysine residues of H3 and H4

HAT-activity is recruited to promoters through specfic interactions with activators, resulting in local acetylation of nucleosomesaround promoters.
Activation or repression is determined bythe equilibrium between HATs and HDACs
On average, 13 of the 30 lysine residues in a histone octamer are acetylated and this steady-state level of acetylation is maintained by the opposing actions of HAT and HDAC complexes.
PROMOTER
TF
HAT
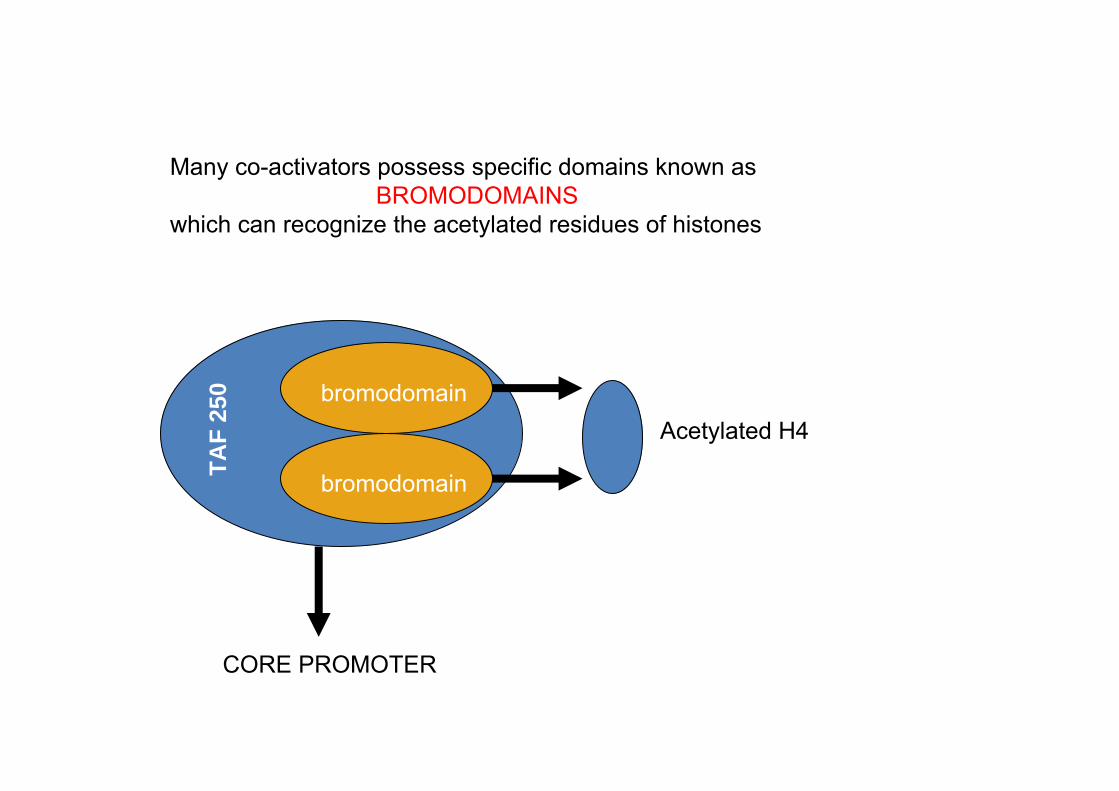
Many co-activators possess specific domains known as BROMODOMAINS
which can recognize the acetylated residues of histones
bromodomainTAF
250
CORE PROMOTER
Acetylated H4bromodomain

The best described HDACs are members of a common family that includes the founding member from human, HDAC1 and yeast Rpd3.
HISTONE DEACETYLASES (HDACs)
The HAT–HDAC system functions as a key regulatory switch of gene expression.
HATHDAC

Human HDACs belong to four classes based on homology to the yeast HDACs:
Rpd3 (class I—HDAC1, HDAC2, HDAC3 and HDAC8),
Hda1 (class II—HDAC4, HDAC5, HDAC6, HDAC7, HDAC9 and HDAC10) and
Sir2 (class III—human sirtuin proteins SIRT1-7)
HDAC11 (class IV HDAC)

HDAC1: Over expressed in prostate cancers (hormone-refractory), gastric, colorectal cancers.
HDAC2: Over expressed in colorectal and gastric cancers.
HDAC3: Over expressed in lung cancer and several solid tumors.
HDAC8: Knock down inhibits cell growth in several human tumor cells
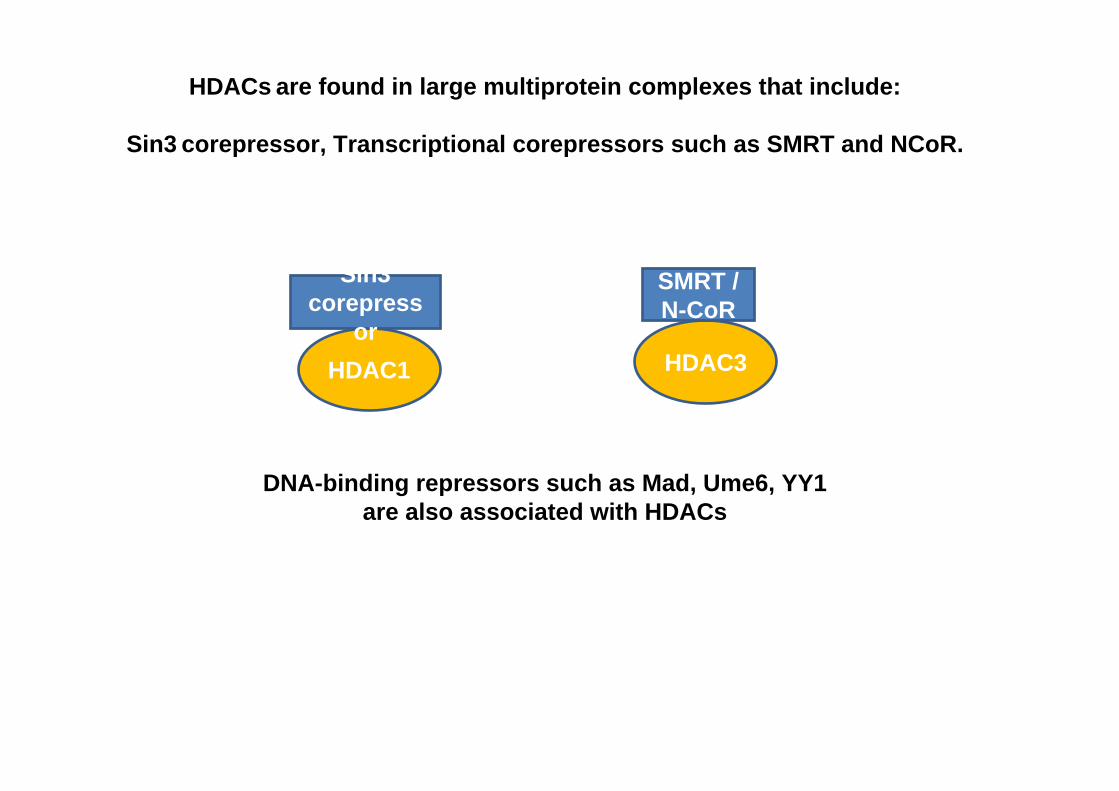
HDACs are found in large multiprotein complexes that include:
Sin3 corepressor, Transcriptional corepressors such as SMRT and NCoR.
HDAC1 HDAC3
SMRT / N-CoR
Sin3 corepress
or
DNA-binding repressors such as Mad, Ume6, YY1 are also associated with HDACs
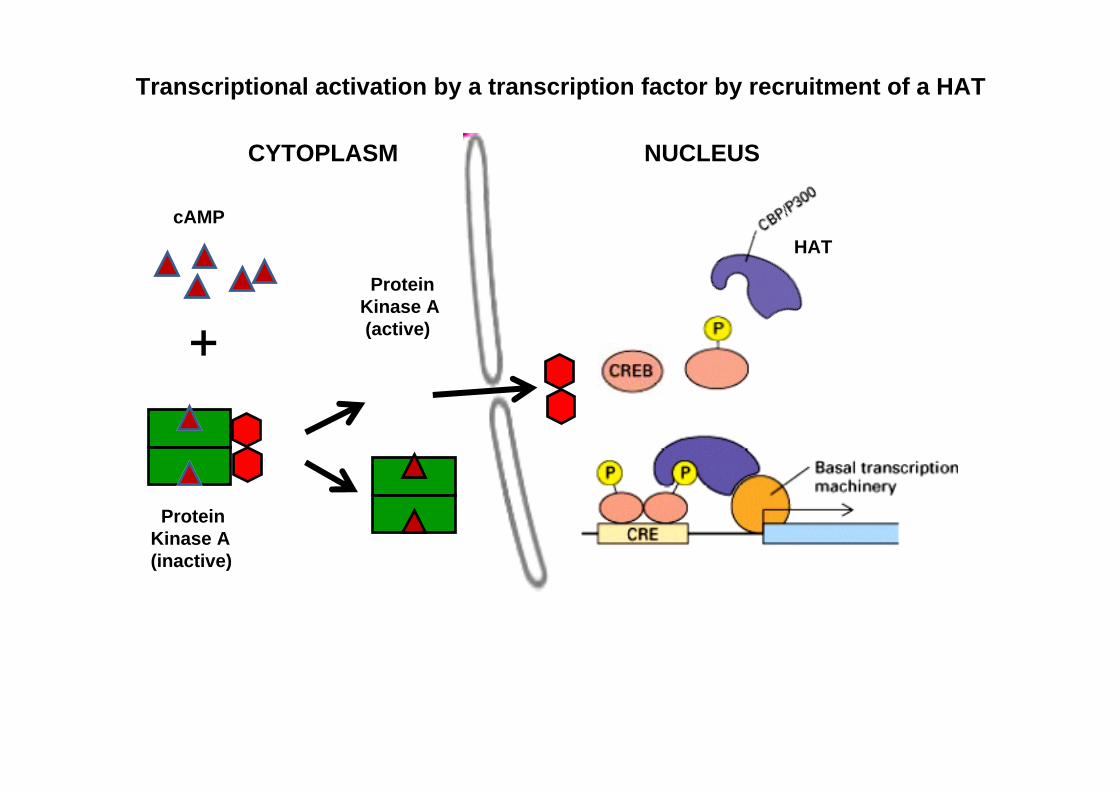
CYTOPLASM NUCLEUS
Protein Kinase A(inactive)
Protein Kinase A(active)
HAT
+
cAMP
Transcriptional activation by a transcription factor by recruitment of a HAT
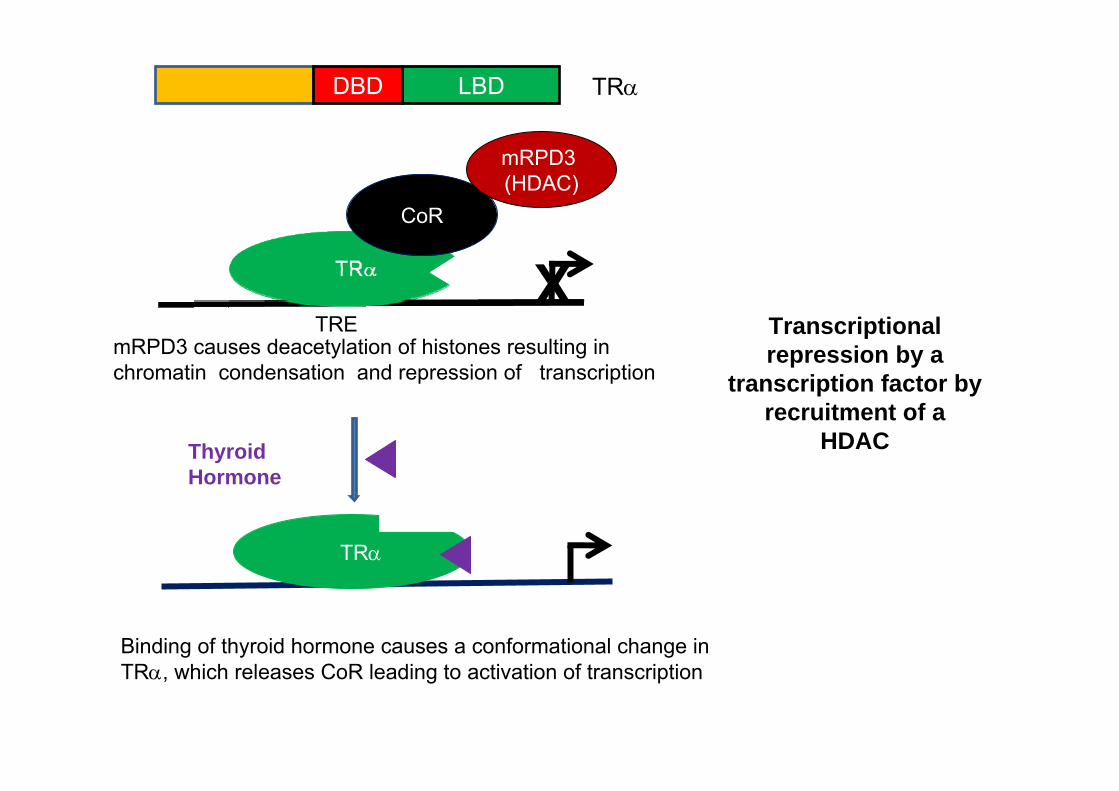
CoR
mRPD3 (HDAC)
mRPD3 causes deacetylation of histones resulting in chromatin condensation and repression of transcription
TRE
Thyroid Hormone
TRα
Binding of thyroid hormone causes a conformational change in TRα, which releases CoR leading to activation of transcription
X
DBD LBD TRα
Transcriptional repression by a
transcription factor by recruitment of a
HDAC

Inhibitors of HDACs, such as:
sodium butyrate, trichostatinA, suberoylanilide hydroxamic acid (SAHA), valproic acid,
induce cell cycle arrest, differentiation, and apoptosis in colon cancer cell lines in vitro and have demonstrated anti-cancer efficacy in clinical trials.

A HDAC inhibitor was approved for the treatment of cutaneous T-cell
lymphoma
Zolinza was approved on October 6th of 2006 by the United Stated Food and Drug Administration for the treatment of a type of skin cancer, cutaneous T cell lymphoma (CTCL), and Sezary's disease
http://www.zolinza.com

Non-histone substrates of HDAC
In addition to deacetylation of histones, HDACs regulate gene transcription by another mechanism wherein they deacetylate several transcription factors such as p53, E2F, andSp3.
In these cases, deacetylation results in reduced DNA binding or transcriptional activity
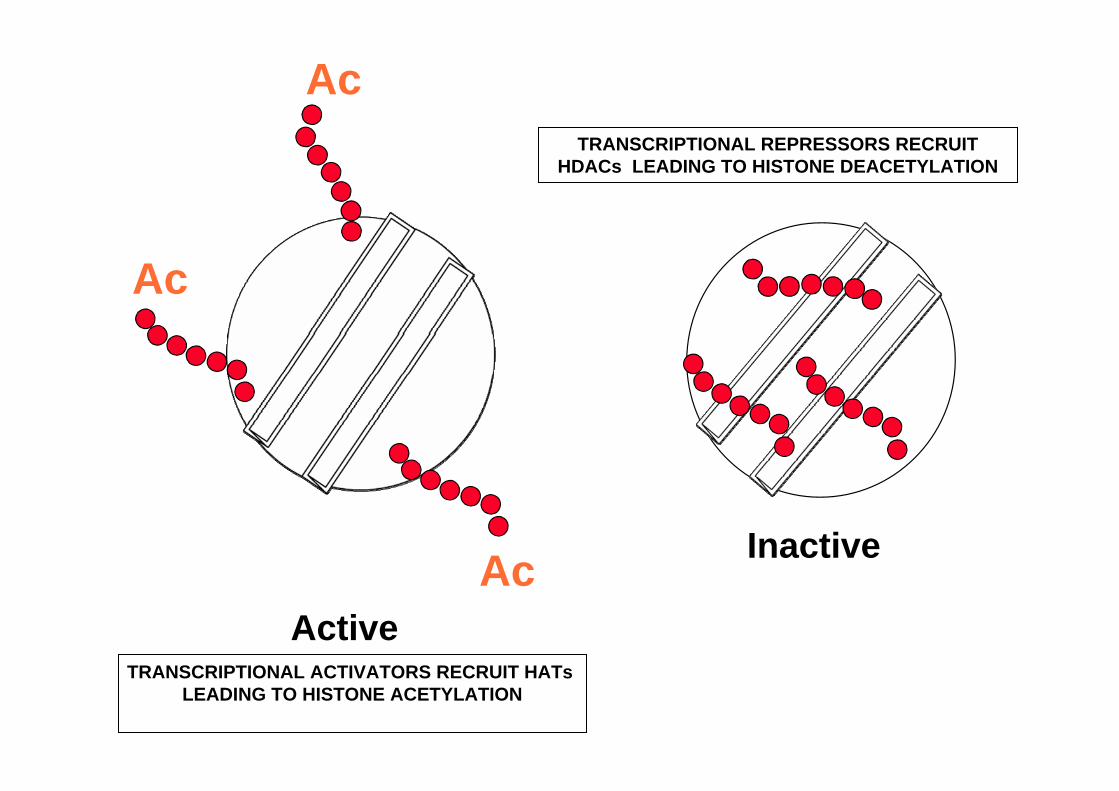
Ac
Ac
AcActive
Inactive
TRANSCRIPTIONAL ACTIVATORS RECRUIT HATs LEADING TO HISTONE ACETYLATION
TRANSCRIPTIONAL REPRESSORS RECRUIT HDACs LEADING TO HISTONE DEACETYLATION

Histone acetylation was first linked to transcriptional activation by Allfrey and colleagues (Proc. Natl Acad. Sci. USA (1964) 51, 786-794).
Acetylated core histones were shown to preferentially associatewith transcriptionally active chromatin (Nucleic AcidsRes.(1978) 5: 1863-1876; Proc. Natl. Acad. Sci. (1978) 75:2239-2243)
Ingram et al. dedmonstrated that n -butyrate can alter cellular differentiation by inhibiting histone deacetylation (Nature(1977)268, 462-464).
Acetylation occurs at lysine residues on the amino-terminal tails of the histones, thereby neutralizing the positive charge of the histone tails and decreasing their affinity for DNA (J. Biol. Chem.(1993) 268: 305-314)
Histone acetylation alters nucleosomal conformation (Norton et al. 1989) so that transcriptional regulatory proteins can access chromatin templates (EMBO J. (1996) 15: 2508-2518).
HISTONE ACETYLATION / DEACETYLATION Historical Aspects

Chromatin structure and modification can not be viewed as a process that is independent of transcriptional initiation.
Chromatin is not simply a structure that serves to compact DNA in the nucleus.
Histone acetylases and deacetylases provide a critical link between chromatin structure and transcriptional output.
Nature (1997) 389:349-352; Cell (1997) 89:325-328; Trends Biochem Sci.(1997) 22: 128-132.
The language of covalent histone modifications BRIAN D. STRAHL AND C. DAVID ALLIS Nature 403, 41 - 45 (2000)