evidence for rolling-circle replication in a major ... · pdf fileevidence for rolling-circle...
TRANSCRIPT

Evidence for Rolling-Circle Replication in a Major Satellite DNA from the South American Rodents of the Genus Ctenomys’
Marfa Susana Rossi, * 9-t Osvaldo Alfred0 Reig,“f and Jorge Zorz6pulos * *BioSidus S-A., Buenos Aires; and TGIBE, Department of Biology, FCEyN, University of Buenos Aires
A major PvuII satellite DNA has been cloned from a South American octodontid rodent of the genus Ctenomys (C. porteousi). The satellite monomer, termed RPCS, is 337 bp in size and 42% G+C. Analysis of the nucleotide sequence demonstrates that RPCS is not composed of a series of shorter repeats. RPCS-related sequences were found in 11 of 12 Ctenomys species analyzed by hybridization under high- stringency conditions. The only negative species, C. opimus, was reactive under low-stringency conditions. RPCS-related sequences were not found under high- or low-stringency conditions in Calomys musculinus and A4us musculus. However, under low-stringency conditions, RPCS-related sequences were found in the oc- todontid Octodontomys gliroides, which is thought to have diverged from the genus Ctenomys more than 10 Myr ago. The pattern of periodicities observed, by restriction analysis, between Ctenomys species in the satellite array can be mainly accounted for by a rolling-circle amplification mechanism but cannot be solely accounted for by unequal crossing-over.
Introduction
South American caviomorph rodents of the genus Ctenomys are the most nu- merous species of all fossorial rodents. This genus is a typical example of an explosive cladogenesis that took place during the Pleistocene and which led to about 55 living species. It has been suggested that this explosive speciation process was triggered by chromosomal rearrangements (Reig 1989; Reig et al. 1990). Species of Ctenomys have the highest rate of karyotypic differentiation shown by a mammalian genus, with diploid number varying from 10 to 72. Therefore, Ctenomys is an attractive model to study molecular mechanisms of speciation. On the other hand, there exists a growing recognition that much of the evolutionary history of eukaryotic genomes reflects the operation of turnover processes involving repetitive DNA sequences and that speciation may be a consequence of such turnover (Rose and Doolittle 1983 ) .
Taking these considerations into account, we initiated a study on the putative role of repetitive DNA sequences in the speciation process of the genus Ctenomys. The present paper describes the isolation and characterization of a major family of repetitive DNA in these rodents.
1. Key words: repetitive DNA Octodontidae, speciation process.
rolling-circle replication, South American subterranean rodents,
Address for correspondence and reprints: Dr. Jorge Zorz6pulos, Biosidus S.A., Constituci6n 4234, 1254 Buenos Aires, Argentina.
Head Department Molecular Genetics,
Mol. Biol. Evol. 7(4):340-350. 1990. 0 1990 by The University of Chicago. All rights reserved. 0737-4030/90/0704-0005$02.00
340

A Major Satellite DNA from Ctenomys 341
Material and Methods Specimens
We studied the DNA of an individual from each of 12 species of Ctenomys and from one specimen each of Octodontomys gliroides, Calomys musculinus, and Mus musculus for comparison. With the exception of M. muscuhs, obtained from a lab- oratory colony, all specimens were caught in the wild by using live traps. Skin and skull voucher specimens of all of them were deposited in the Collection of Mammals of the Municipal Museum of Natural History of Mar de1 Plata. Geographic locality of the collected specimens, all from Argentina, are as follows (t.1. = type locality): Ctenomys azare (Luan Toro, La Pampa), Ctenomys australis (Necochea, Buenos Aires, t.l.), Ctenomys argentinus (Colonia Ramirez, Chaco, t.l.), Ctenomys latro (Tapia, Tucuman, t.1.) , Ctenomys talarum (Necochea, Buenos Aires), Ctenomys tuconax (El Infiernillo, Tucuman ) , Ctenomys occultus ( Monteagudo, Tucuman ) , Ctenomys opi- mus (Tres Cruces, Tucuman), Ctenomys cf: perrensi, 2n = 54 (Salada, Corrientes), Ctenomys cf perrensi, 2n = 58 (7 km ENE of Salada, Corrientes), Ctenomys porteousi ( Bonifacio, Buenos Aires, t.l.), and 0. gliroides ( Tilcara, Jujuy ) .
DNA Extraction
Liver pieces were extracted by excision and were suspended in a buffer containing 100 mM Tris-HCl, 10 mM ethylenediaminetetraacetate (EDTA) , 100 mM NaCl, 1% sodium dodecyl sulfate (SDS), and 150 pg of proteinase K/ml, pH 7.8. This suspension was incubated for 6 h at 55°C. After this, half a volume of 7.5 M ammonium acetate was added. The DNA was precipitated from this solution by adding two volumes of ethanol. After centrifugation, the pellet was vacuum dried and was resuspended in sterile water. This ammonium acetate-ethanol precipitation was repeated twice. Finally, the pellet was resuspended in sterile water and stored at -20°C until use. Plasmid DNA was purified according to a method described by Bimboim and Doly ( 1979).
Enzyme Digestion and Gels
The DNA was digested with restriction enzymes (Bethesda Research Laboratories) according to the instructions of the manufacturer. The DNA fragments were separated in 1% agarose gels made up in Tris-borate-EDTA buffer (Maniatis et al. 1982).
Blotting and Hybridization
Agarose gels were denatured, neutralized, and blotted according to the method described by Southern ( 1975a), by using nylon membranes (Pall Byodine). Prehy- bridization and hybridization were performed in a solution containing 6 X standard saline citrate (SSC) ( 1 X SSC = 0.15 M NaCl, 0.0 15 M trisodium citrate, pH 7)) 100 pg of salmon DNA (Sigma, USA)/ml, 0.5% SDS, and 0.3% nonfat dry milk. Labeled DNA probes were hybridized with the DNA immobilized on the membranes for 18 h at 60°C. Then the membranes were washed five times at 60°C in 6 X SSC (low stringency) or 0.1 X SSC (high stringency) and 1% SDS. Hybrid DNA was detected by autoradiography, using 3M X-ray films and Du-Pont Lightening Plus screens.
Labeling of DNA
The DNA was labeled with (alpha-32P) to a specific activity of 10 * cpm/ kg by using the nick-translation procedure (Rigby et al. 1977).



10 20 30 40 50 60 70 80 PVUII NInC I nda I -- i?i&GCTATT CTCTATTCCA GACTCACTCT TAGACAACAT GATGGAGCAC AAGAAATTCC ATCTTGAACT ACCCTGCATA CATACCAGGG - Alu I Dde I Dde I
=TTCCCTT AGGAATGAAA GGGAGAGAAA ACATGTGCCT GTATCTCTTT TGCACTAAGA ACTGGAAAGC ACCAACTTCG CCTATACTTT
ACTGAACGAG TTGCAACACT CACAACCTCA CACTTCTGAA TTGGTTTGGA ACCAATGTGA CACACATTAG CCAAAATGCT GTATTTTGCT
ACAGGCATTG CAGTTCCCTG CTTCCCAATT TGATCAG - Alu I
FIG. 3.-Nucleotide sequence of the RFCS. Recognition sites for the enzymes used are marked


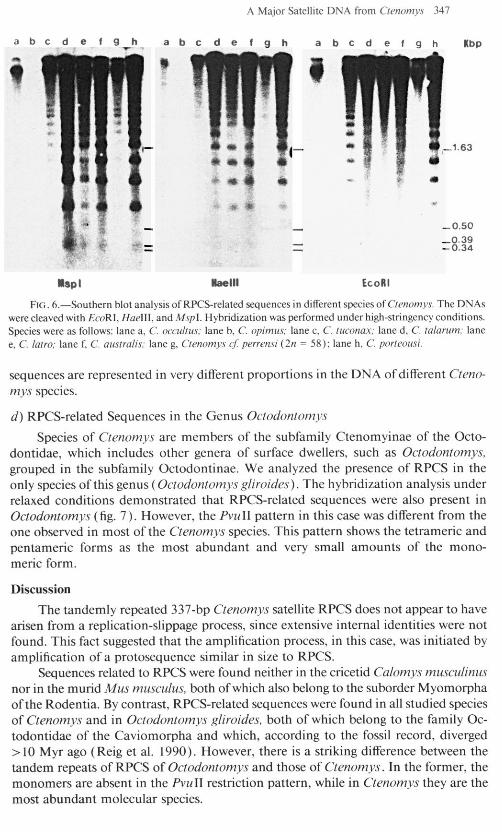
lSPl llaelll EcoRl
no. h.-Southern blot analysis of RPCS-related sequences in different skes of Clmomys. The DNAs were cleaved with EcoRI, Ha&l, and Mspl. Hybridization was performed under high-stringency conditions. Specie were as follows: lane a, C. occulrus: lane b, C. opimus; lane c, C. U~~nar: lane d, C. lolarum: lane e, C. lawo: lane f, C. ousrralis; lane g, Clenomvs c/Y perrensi (278 = 58); lane h, C. poneousi.
sequences are represented in very different proportions in the DNA ofdifferent Ctmo- mys species.
d) RPCS-related Sequences in the Genus Octodontomys
Species of Ctenomys are members of the subfamily Ctenomyinae of the Octo- dontidae, which includes other genera of surface dwellers, such as Octodontomys, grouped in the subfamily Octodontinae. We analyzed the presence of RPCS in the only species of this genus ( Octodontomys ghides), The hybridization analysis under relaxed conditions demonstrated that RPCS-related sequences were also present in Octodontomys (fig. 7). However, the PvuII pattern in this case was different from the one observed in most of the Ctenomys species. This pattern shows the tetrameric and pentameric forms as the most abundant and very small amounts of the mono- meric form.
Discussion
The tandemly repeated 337-bp Cfenomys satellite RPCS does not appear to have arisen from a replication-slippage process, since extensive internal identities were not found. This fact suggested that the amplification process, in this case, was initiated by amplification of a protosequence similar in size to RPCS.
Sequences related to RPCS were found neither in the cricetid Calomys mumdinus nor in the murid Mus musculus, both ofwhich also belong to the suborder Myomorpha ofthe Rodentia. By contrast, RPCS-related sequences were found in all studied species of Ctenomys and in Oclodontomys gliroides, both of which belong to the family Oc- todontidae of the Caviomorpha and which, according to the fossil record, diverged t 10 Myr ago (Reig et al. 1990). However, there is a striking difference between the tandem repeats of RPCS of Octodontomys and those of Ctenomys. In the former, the monomen are absent in the PvuII restriction pattern, while in Ctenomys they are the most abundant molecular species.


A Major Satellite DNA from Ctenomys 349
A proto-monomer h
-@4-D-
1
mutation
J- Pvu II sit*
1 unequal cross-over
-~-b+WS++,+,~
1
l ’ P >A-
1
introstrand exchange
4 -.-.U +
1
C.3
rolling cwcle rcplicatton
FIG. 8.-Scheme explaining the RPCS amplification process, based on the model proposed by Walsh ( 1987). A, Ancestral sequence with a SVUII site amplified by unequal crossing-over. This tandem array undergoes an intrastrand exchange resulting in the formation of a circular plasmid carrying monomers with the PvuII site. This plasmid is replicated through a rolling-circle process which therefore amplifies a particular section of the original tandem. Finally, the amplified sequence is reinserted into the chromosomal DNA into the original tandem. B, Periodicities generated by local amplification of monomers containing mutations which afIii restriction patterns.
ancestor of the Ctenomys. This amplification continued after the initiation of the explosive cladogenesis which led to the multifarious speciation of Ctenomys (Reig 1989). However, the rate of amplification was different for different Ctenomys lineages. We think that most of the amplification was a consequence of the creation of circular plasmids by intrastrand exchange followed by rolling-circle replication and reinsertion, mostly at homologous sites (for a discussion of why the unequal crossing-over process may not be sufficient for subsequent amplifications, see Walsh 1987 ) . However, the reinsertion occasionally occurred at nonhomologous sites which became hot spots for future reinsertions. Satellite periodicities were the result of the amplification of mono- mers containing mutations that suppressed the PvuII sites (fig. 8B). Expansion of monomers containing mutations that create other restriction sites (i.e., EcoRI or HaeIII sites) were also the result of this same local amplification process (also see Hiirz and Zachau 1977). This amplification model has been previously proposed by Walsh ( 1987), and we think that it is the one that best fits our results.
Ctenomys are the mammals with the highest level of chromosomal differentiation. Karyotype numbers for Ctenomys species vary from 2n = 10 to 2n = 72. On the basis of the known fossil history of Ctenomys, it can be confidently estimated that the highly extensive speciation of this genus occurred during the Pleistocene, within a span of 1.8 Myr (Reig et al. 1990). This is also the span of the high rate of chromosomal evolution observed in this genus. This extremely high rate of chromosomal repatterning

350 Rossi et al.
related to the explosive species diversification is only partially explained by the known population structure of Ctenomys. Present theories of chromosomal evolution (see Sites and Moritz 1987 ) are insufficient to explain multiple fixation of chromosomal changes under the known rates of chromosomal mutation. A dynamic process of DNA amplification, as suggested by our present results, may have acted as a triggering factor for high rates of chromosomal repatterning. However, direct evidence of this is still lacking.
Acknowledgments
We thank Miriam Denegri for help in the preparation of the manuscript. We also wish to thank Drs. John A. W. Kirsch and Pablo A. Galdi for reviewing the manuscript.
LITERATURE CITED
BIRNBOIM, H. C., and J. DOLY. 1979. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 7: 15 13- 1523.
HANAHAN, D. 1983. Studies on transformation of E. coZi with plasmids. J. Mol. Biol. 166:557- 580.
H&z, W., and H. G. ZACHAU. 1977. Characterization of distinct segments in mouse satellite DNA by restriction nucleases. Eur. J. Biochem. 73:383-392.
MANL~TIS, T., E. F. FRITSCH, and J. SAMBROOK. 1982. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory. Cold Spring Harbor, N.Y.
PROSSER, J., M. FROMMER, C. PAUL, and P. C. VINCENT. 1986. Sequence relationship of three human satellite DNAs. J. Mol. Biol. 187:145-155.
REIG, 0. A. 1989. Chromosomal repatterning as a causative factor in cases of explosive speciation. Pp. 246-289 in A. FONTDEVILA, ed. Evolutionary biology of transient, unstable populations. Springer, Berlin.
REIG, 0. A., C. BUSCH, M. 0. ORTELLS, and J. CONTRERAS. 1990. An overview of evolution, systematics, population biology and speciation in Ctenomys. Pp. 7 l-96 in E. NEVO and 0. A. REIG, eds. Evolutionary biology of subterranean mammals. Alan R. Liss, New York.
RIGBY, P. W., M. DIECKMANN, C. RHODES, and P. BERG. 1977. Labeling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I. J. Mol. Biol. 113:236-25 1.
ROSE, M. R., and W. F. DOOLITTLE. 1983. Molecular biological mechanisms of speciation. Science 220: 157- 16 1.
SANGER, F., S. NICI&EN, and A. R. COULSON. 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
SITES, J., and C. MORITZ. 1987. Chromosomal evolution and speciation. Syst. Zool. 36: 153- 174.
SOUTHERN, E. M. 1975a. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98:503-5 17.
- 1975b. Long range periodicity in mouse satellite DNA. J. Mol. Biol. 94:5 l-69. . WALSH, J. B. 1987. Persistence of tandem arrays: implications for satellite and simple-sequence
DNAs. Genetics 115:553-567.
WALTER M. FITCH, reviewing editor
Received January 14, 1989; revision received February 12, 1990
Accepted February 12, 1990