expression of murine ell-associated factor 2 (eaf2) is developmentally regulated
TRANSCRIPT

PATTERNS & PHENOTYPES
Expression of Murine ELL-Associated Factor 2 (Eaf2)Is Developmentally RegulatedMin Li,1 Xiangbing Wu,1 Fengfeng Zhuang,1 Shaoyun Jiang,1 Meisheng Jiang,3 and Yi-Hsin Liu1*
Eaf2, ELL-associated factor 2, encodes a protein that is homologous to the human EAF1, which was shown to interactwith the transcriptional elongation factor MEN/ELL. During mouse embryogenesis, Eaf2 is preferentially expressed inthe central nervous system and in sensory and neuroendocrine organs, including the brain, spinal cord, cranial andspinal ganglia, developing otocyst, the retina, and the pituitary. Eaf2 transcripts were also found in sites where activeepithelium–mesenchymal interactions are occurring. These included the invaginating tooth buds, mammary glandanlage, submandibular glands, the lung, the pancreas, and the kidney. Other sites of expression included bladder andintestine. In the developing lens, Eaf2 transcripts were absent in the proliferating anterior lens epithelial cells but werepresent in the terminally differentiated primary lens fiber cells and also in nonproliferating lens fiber cells in theequatorial zone where lens epithelial cells withdraw from cell cycle and terminally differentiate into secondary lensfiber cells. This spatially restricted pattern of Eaf2 expression in the developing lens suggests that Eaf2 may play animportant role in regulating lens maturation. Developmental Dynamics 228:273–280, 2003. © 2003 Wiley-Liss, Inc.
Key words: embryogenesis; epithelial–mesenchymal interaction; eye; CNS; Eaf2; ELL; in situ hybridization
Received 3 June 2003; Accepted 24 June 2003
INTRODUCTION
During embryonic development aswell as during tissue regenerationand repair, cell proliferation and dif-ferentiation are highly regulatedand well coordinated events that re-quire de novo transcription of tissue-specific or cell-specific genes. Acti-vation of target gene transcription isinitiated by tissue- or cell-specifictranscriptional factors and theirchromatin remodeling coactivatorsin contact with the RNA polymerase
II (RNAP II) initiation complex (Emer-son, 2002). During the elongationphase, RNAP II encounters numerousstructural obstacles causing RNAP IIto either pause or terminate tran-scription prematurely. In mammaliancells, several factors have beenidentified that can enhance RNAP IIprocessivity in vitro (Shilatifard, 1998;Conaway and Conaway, 1999).Among these factors that processthe elongation activity are Eleven-nineteen Lysine-rich Leukemia (ELL)
family of proteins (Thirman et al.,1994, 1997; Mitani et al., 1995; Shilati-fard et al, 1996; Shilatifard et al.,1997; Miller et al., 2000).
The ELL gene was initially identifiedon chromosome 19p 13.1, which un-dergoes frequent translocation withthe MLL gene on chromosome11q23 in acute myeloid leukemia(AML; Thirman et al., 1994). ELL mayplay an essential role in regulatingdevelopmental processes as its ex-pression is spatially and temporally
1Center for Craniofacial Molecular Biology, Division of Craniofacial Sciences and Therapeutics, School of Dentistry, University of SouthernCalifornia, Los Angeles, California2Department of Molecular and Medical Pharmacology and Anesthesiology, David Geffen School of Medicine, University of California, LosAngeles, CaliforniaGrant sponsor: NIDCR, NIH; Grant number: DE12779 Grant number: DE12941.Drs. Li and Wu contributed equally to this work.Dr. Wu’s present address is University of British Columbia, Department of Anatomy, Friedman Building, 2177 Wesbrook Mall, Vancouver BCV6T 1Z3, Canada.*Correspondence to: Yi-Hsin Liu, Center for Craniofacial Molecular Biology, School of Dentistry, University of Southern California, 2250Alcazar Street, CSA103, Los Angeles, CA 90033. E-mail: [email protected]
DOI 10.1002/dvdy.10367
DEVELOPMENTAL DYNAMICS 228:273–280, 2003
© 2003 Wiley-Liss, Inc.

restricted during embryogenesis(Thirman et al., 1997) and inactiva-tion of murine Ell by gene targetingresulted in embryonic lethalityaround the time of gastrulation (Mi-tani et al., 2000). In Drosophila, loss-of-function mutations in dELL locuscause recessive lethality and seg-mentation defects in developinglarva (Eissenberg et al., 2002). In cul-ture, overexpression of ELL inhibitscell division and induces apoptosis(Johnstone et al., 2001). Remark-ably, the growth-suppression anddeath-promoting activities of ELLcan be assigned to discrete func-tional domains that possess elonga-tion-enhancing activities and im-mortalizing functions, respectively(Kanda et al., 1998; DiMartino et al.,2000; Johnstone et al., 2001). Thisfinding further suggests the exis-tence of distinct interaction surfaceson ELL and through which ELL couldbe differentially regulated.
Previously, several ELL-interactingpartners have been identified andhave been shown to mask the inhib-itory activity of ELL in transcription ini-tiation and to stimulate RNA elonga-tion (Schmidt et al., 1999; Kamura etal., 2001) or transcription (Simone etal., 2001, 2003). Human EAF1 andEAF2 are two of these ELL-associ-ated factors that had been shownto enhance transcription (Simone etal., 2001, 2003). ELL and EAF1 colo-calize with p80 coilin and may func-tion as components of Cajal bodies,a nuclear compartment involved insnRNP biogenesis (Gall, 2000; Polaket al., 2003). Further studies showedthat EAF-interacting domain in ELL isessential for the transformation ac-tivity of the MLL–ELL fusion protein(Luo et al., 2001; Simone et al., 2003).EAF1 or EAF2 could replace ELL in itsability to immortalize hematopoieticprogenitor cells when it was fused toMLL (Luo et al., 2001; Simone et al.,2003). Together, these data suggestthat EAFs may function as oncopro-teins if recruited by the MLL/ELL com-plex. However, very little is knownabout the expression pattern ofthese ELL-interacting molecules dur-ing embryonic development andtheir normal cellular functions.
In this report, we describe thecloning and expression of a newmember of ELL-associated factors,
murine Eaf2. Sequence compari-sons revealed a high degree ofamino acid sequence conserva-tion among rodent and humanEAF2 proteins. Eaf2 is expressedpreferentially in the central nervoussystem (CNS) and sensory and neu-roendocrine organs and in siteswhere active epithelium–mesen-chymal interactions are occurring.Together these results suggest thatEaf2 may be an important regula-tor of morphogenesis, cell growth,and differentiation.
RESULTS
Identification of cDNAEncoding EAF2
In performing a yeast two-hybridscreen to identify proteins that maypotentially interact with murineMsx1, we isolated a novel cDNAclone. This cDNA produced a pre-dicted open reading frame of 262amino acids with a predicted mo-lecular weight of 29.187 kDa (Fig.1A). Charged residues constitute27.86% of overall amino acid con-tent and 17.6% of amino acids arecomposed of serine residues.
BLASTp search of the GenBanknonredundant sequence databaseidentified two human sequences(GenBank accession nos. AAH14209and AAF67627) that are virtuallyidentical to that of the mouse Eaf2(Fig. 1B). Although the carboxyl ter-minus showed significant diver-gence, these proteins share 79%amino acid identities. In addition,we identified a rat ortholog of Eaf2(GenBank accession no. AAL12225)that has 91% amino acids in com-mon with the mouse Eaf2 (Fig. 1B).Furthermore, we identified a relatedhuman gene, EAF1 (GenBank ac-cession no. AAK58687), that was pre-viously cloned and characterizedand its mouse Eaf1 ortholog (Gen-Bank accession nos. BAC32058 andBAC31806; Fig. 1C). Although theoverall amino acid homology be-tween the murine Eaf2 and humanEAF1 is only 53%, ClusterW alignmentof protein sequences between Eaf2and EAF1 identified two regions ofsignificant homology: one at the N-terminus spanning from amino acids
10 to 127 (79% amino acid identities)and another at the extreme C-termi-nus between amino acids 243 and262 (80% amino acid identities; Fig.1C). A unique proline-rich domainfound in the EAF1 distinguishes itselffrom the Eaf2 members of the family(Fig. 1C).
BLASTp search also identified aprototype sequence for the mam-malian Eafs in the Drosophila ge-nome encoded by the CG11166gene (GenBank accession no.AAF59266). We tentatively named itdEaf. ClusterW alignment showedhigh degree of amino acid se-quence similarities (48%) betweenthe N-terminus of dEaf and the N-termini of mammalian Eafs (Fig. 1D).In addition, two other domains ondEaf exhibited considerable homol-ogy with mammalian Eafs; thesewere the serine-rich domain and the20 amino acids at the extreme car-boxyl terminus (40% similarities and45% similarities, respectively; datanot shown).
Expression Profile of Eaf2 inCNS
To gain a better understanding ofsites where Eaf2 may exert its func-tion alone or together with ELL, wedecided to determine the expres-sion pattern of Eaf2 during embry-onic development by performingin situ hybridization on cryosectionsof developing mouse embryos. Atembryonic day (E) 10, Eaf2 tran-scripts were found mainly in the de-veloping brain and spinal cord(data not shown). In the mid-ges-tation embryo from E12 to E15, Eaf2was expressed at very high levelsthroughout the developing CNS, in-cluding the brain, spinal cord, andcranial and spinal ganglia (Fig.2A,B,E). Of interest, intense hybrid-ization signals were also observedin sensory organs and neuroendo-crine structures. These structures in-cluded the lens and retina of thedeveloping eye, the cochlea ofthe embryonic ear, the olfactoryepithelium, and the pituitary (Figs.2A–D,F, 3A). In the developing eye,Eaf2 transcripts were absent fromthe developing optic vesicle be-fore the appearance of retinal
274 LI ET AL.
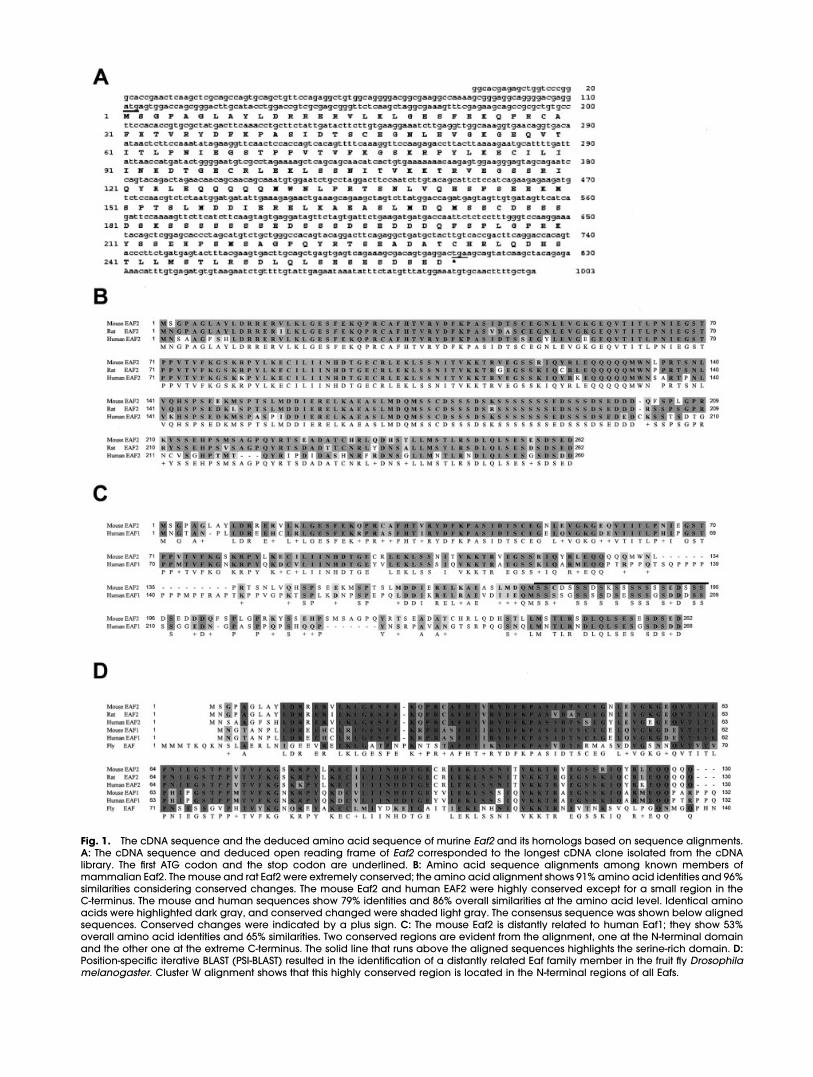
Fig. 1. The cDNA sequence and the deduced amino acid sequence of murine Eaf2 and its homologs based on sequence alignments.A: The cDNA sequence and deduced open reading frame of Eaf2 corresponded to the longest cDNA clone isolated from the cDNAlibrary. The first ATG codon and the stop codon are underlined. B: Amino acid sequence alignments among known members ofmammalian Eaf2. The mouse and rat Eaf2 were extremely conserved; the amino acid alignment shows 91% amino acid identities and 96%similarities considering conserved changes. The mouse Eaf2 and human EAF2 were highly conserved except for a small region in theC-terminus. The mouse and human sequences show 79% identities and 86% overall similarities at the amino acid level. Identical aminoacids were highlighted dark gray, and conserved changed were shaded light gray. The consensus sequence was shown below alignedsequences. Conserved changes were indicated by a plus sign. C: The mouse Eaf2 is distantly related to human Eaf1; they show 53%overall amino acid identities and 65% similarities. Two conserved regions are evident from the alignment, one at the N-terminal domainand the other one at the extreme C-terminus. The solid line that runs above the aligned sequences highlights the serine-rich domain. D:Position-specific iterative BLAST (PSI-BLAST) resulted in the identification of a distantly related Eaf family member in the fruit fly Drosophilamelanogaster. Cluster W alignment shows that this highly conserved region is located in the N-terminal regions of all Eafs.

ganglion cells (RGCs; data notshown). In the mouse, RGC matu-ration initiates around E12.5 andpeaked by E14.5 (Young, 1985;Brown et al., 2001). Expression of
Eaf2 coincided remarkably withthe birth of RGCs, and intense hy-bridization signal was preferentiallyfound in the RGC layer of the de-veloping retina (Fig. 2C,D). Of inter-
est, Eaf2 transcripts were absentfrom actively cycling retinal cells inthe retinal marginal zone and lowlevel Eaf2 gene expression was de-tected in dividing cells in the resid-
Fig. 2.
Fig. 3.
276 LI ET AL.

ual portion of the neuroretina, ex-cluding RGCs (Fig. 2C,D). In thedeveloping lens, Eaf2 transcriptswere found exclusively in the pri-mary lens fiber cells but were absent in the anterior lens epithelium(Fig. 2C). As anterior lens epithelialcells move into the equatorial zoneof the lens and exit cell cycle, Eaf2was preferentially transcribed bythe lens equatorial epithelial cells(Fig. 2D).
Expression Profile of Eaf2During Epithelium–Mesenchymal Induction
Furthermore, Eaf2 transcripts weredetected in structures that undergoextensive branching morphogene-sis. Extensive expression was ob-served in the epithelia of develop-ing nephrons, in the bronchialepithelium of the embryonic lung,in the secretory epithelium of sub-mandibular glands, and in the tu-bular epithelium of the epididymis,although a greatly reduced levelof Eaf2 expression was detected inthe mesenchyme of these organs(Fig. 3A–E). In the ectodermal in-vaginations of mammary buds andvibrissae follicles, Eaf2 transcripts
were also found (Fig. 3F,G). Of in-terest, Eaf2 transcripts were alsodetected in the developing incisorsand molars (Fig. 3H–J). Again, Eaf2expression was more intense in thedental epithelium, although a re-duced amount of Eaf2 transcriptswas also found in the dental mes-enchyme and dental papilla (Fig.3I,J). Other sites of expression in-cluded intestine and bladder en-dothelium (Fig. 3A; data notshown). In the developing liver andheart, Eaf2 transcript was absent orbelow the sensitivity of our detec-tion method (Figs. 2A, 3A).
Expression Profile of Eaf2 inAdult Tissues
To determine the expression patternof Eaf2 in adult tissues, we per-formed Northern blot hybridizationon poly(A�) RNAs or total RNAs iso-lated from mouse adult tissues. Sev-eral adult tissues that perform repro-ductive functions, including theovary, uterus, mammary glands,and testis, were found to produceEaf2 transcripts (Fig. 4A,B). Eaf2 tran-scripts were also detected in theadult brain, spleen, liver, lung, thy-mus, and kidney (Fig. 4A). Skeletal
muscle and skin expressed the Eaf2gene at very low levels (Fig. 4B) asindicated by the relative intensity ofhybridization signal in comparison tothe control glucose-6-phosphate di-hydrogenase probe. Eaf2 transcriptswere undetectable in the adultheart (Fig. 4A).
Three transcripts with different mo-lecular weights were detectedbased on Northern blot analysis. Amajor and the most abundant tran-script is approximately 1.4 kb inlength. A second minor transcriptthat was detected only in the brain,lung, and ovary was approximately2.6 kb long. A third minor RNA spe-cies was detected in the spleensample and corresponds to a tran-script size of approximately 1 kb.These minor RNA species are mostlikely products of splicing variantsnot due to cross-hybridization withrelated genes, such as Eaf1, be-cause Eaf1 and Eaf2 shared only35% similarities at the nucleic acidlevel and BLASTn search of themouse genome database and theGenBank nonredundant nucleicacid database did not produceadditional DNA sequences with sig-nificant homology (data notshown).
Fig. 2. Expression profile of Eaf2 in the central nervous system and sensory organs during mouse embryonic development. A: Aparasagittal section of an embryo at embryonic day (E) 11 shows hybridization signal (purple stain) for Eaf2 transcripts in the brain (br),spinal cord (sc), spinal ganglia (white arrows), mandible (mb), and the gut (g). Eaf2 expression was not detected in the liver (li) and heart(h). B: In this frontal section through the head of an embryo at E12, stronger hybridization signal is seen in the diencephalon and in theneuroretina. Interestingly, Eaf2 expression in the ependymal layer of the brain was substantially less in intensity (arrow). C: Highermagnification view of a frontal section through the eye of an embryo at E12 shows selective hybridization in the presumptive retinalganglion cell layer (white arrow) and primary lens fiber cells. The optic stalk (os) is also shown to produce Eaf2 transcripts. Interestingly,in the anterior lens epithelium cells (star) and the pigmented ciliary margins (arrowheads), where lens and retina progenitor cells arefound, respectively, no Eaf2 transcripts were detected. D: In this section of the eye from an embryo at E15, Eaf2 expression in the retinacould be separated into two zones based on expression level, the expression level in the outer layer is relatively weak in comparison tothe inner layer where differentiated RGCs are located. In the lens, hybridization signal in the equatorial epithelium (arrow) is more intense.E: In the developing spinal cord of an embryo at E12, Eaf2 expression level is relatively low in the ependymal layer of dorsal half of thespinal cord (sc) in comparison to its expression in the mantle layer. Intense hybridization signal is seen in dorsal root ganglia (sg). F: Eaf2transcripts were also detected in the olfactory epithelium (oe) of the nasal pits (black arrows) and a pair of vomeronasal organs (whitearrows) in this frontal section of an embryo at E12. G: Eaf2 gene expression was found in the pituitary gland (pit) and cochlear (ch) of anembryo at E14.Fig. 3. Eaf2 expression in epithelially derived organs. A: This parasagittal section of an embryonic day (E) 15 embryo shows hybridizationof Eaf2 transcripts in the brain (br), cochlea (ch), eye (e), lung (lu), intestine (int), seminiferous tubules of the testis (te), kidney (ki), spinalganglia (sg), and brown fat (bf). B: In the kidney of an embryo at E15, Eaf2 transcripts were found in the developing nephrons (whitearrows). C: In the developing lung of an embryo at E15, intense hybridization signal was detected in bronchial epithelium, although lowerexpression level was also found in the lung mesenchyme. D: In the submandibular gland of an embryo at E15, strong hybridization signalwas seen in the ductal epithelium. E: At E15, high levels of Eaf2 expression were seen in the tubular epithelium of the epididymis (blackarrows). F: Eaf2 transcripts were detected (arrows) in the mammary buds from an embryo at E15. G: Eaf2 expression (arrows) was seenin vibrissae follicles of an embryo at E15. H,I: Eaf2 transcripts were found in the developing incisors and molars of an embryo at E13.Hybridization signals were more intense in the dental epithelium (white arrows) in contrast to the expression level in the dentalmesenchyme (black arrows). J: At E15, Eaf2 expression continues as the tooth progresses through the bell stage. Hybridization signals weremore intense in the dental epithelium (white arrow) in comparison to that in the mesenchyme (black arrow). ins, incisors; mb, mandible;p, palate; t, tongue; h, heart; li, liver; hl, hindlimb.
CLONING AND EXPRESSION ANALYSIS OF MOUSE Eaf2 277

DISCUSSION
We have isolated and character-ized murine Eaf2. It showed remark-able amino acid conservation withits human ortholog; 79% amino acidswere identical and homology ap-proaches 86% if conserved changeswere taken into consideration. By us-ing the BLASTp algorithm, we havealso identified the Drosophila proto-type of mammalian Eafs in additionto the rat Eaf2 and the mouse Eaf1.Together with previously identifiedhuman EAFs, these genes constitutea new gene family. Moreover, byperforming sequence alignmentsbetween the Drosophila Eaf andmammalian Eafs, we identified threehighly conserved protein domains.At least two of these conserved pro-tein domains were shown previouslyto be essential for binding to ELL oractivating transcription, respectively(Simone et al., 2001, 2003), suggest-ing possible functional conservation.Although no loss-of-function muta-tion has been reported, dEaf en-coded by the Drosophila CG11166gene appeared to functionally inter-
act with pannier (pnr), a GATA-typezinc-finger transcriptional factor. In-duced overexpression of CG11166by a promoter-specific insertion re-sulted in the enhancement of pnrmutant phenotypes (Pena-Rangelet al., 2002).
Of interest, Eaf2 expression ap-pears to be tightly regulated duringembryonic development. Its expres-sion is spatially and temporally re-stricted. Eaf2 transcripts were de-tected in organs whose developmentrequire series of reciprocal tissue–tis-sue inductions between the epithe-lium and the underlying mesen-chyme. These included the tooth,mammary placodes, vibrissae folli-cles, submandibular glands, lung,pancreas, and kidney. This preferen-tial spatial distribution of Eaf2 tran-scripts in branching structures sug-gests that Eaf2 may be activelyinvolved in their inductive processes.
Furthermore, Eaf2 may play a keyrole in regulating growth and differ-entiation of the nervous system asdemonstrated by its extensive ex-pression in the embryonic brain, spi-nal cord, and cranial and spinal
ganglia. The expression of Eaf2 in theCNS during early stages of mouseembryogenesis is very similar to thereported expression profile of Ell (Thir-man et al., 1997). In the sensory or-gans, such as olfactory epitheliumand the sensory epithelium in the co-chlea, high levels of Eaf2 expressionwere found, suggesting its possiblefunction in facilitating the matura-tion process of these specialized ep-ithelia. In the developing retina, theappearance of Eaf2 transcripts co-incides with the birth of retinal gan-glion cells (RGCs) and RNA in situshowed intense hybridization signalin the RGC layer. This finding sug-gests that Eaf2 may be an activeplayer in regulating the differentia-tion process of RGCs perhaps bypromoting or stabilizing the state ofdifferentiation. Further studies areneeded to define its role in control-ling neurogenesis.
In the developing lens, Eaf2 tran-scripts were absent in the proliferat-ing anterior lens epithelial cells butwere present aplenty in the termi-nally differentiated primary lens fibercells and also in nonproliferating lensfiber cells in the equatorial zonewhere lens epithelial cells withdrawfrom the cell cycle and terminallydifferentiate into secondary lens fi-ber cells (Menko, 2002). This spatiallyrestricted pattern of Eaf2 expressionin the developing lens provides agood indication that Eaf2 may playan important role in regulating thedifferentiation program of lens fibercells or their withdraw from the cellcycle. The functional significance ofthese observations awaits corrobo-rating results from genetic perturba-tion experiments.
EXPERIMENTAL PROCEDURES
Isolation of Eaf2 and PlasmidConstructions
Eaf2 was identified as a putativepositive-interacting target for Msx1while performing a yeast two-hybridscreening using a mouse two-hybridcDNA library prepared from E14.5embryos (Stratagene, La Jolla, CA).The plasmid that contained Eaf2cDNA was isolated from yeast cellsand used to transform Escherichiacoli (XL-Blue). This resulted in the
Fig. 4. Expression profile of Eaf2 in the adult mouse. A: Two micrograms of polyA-selectedmRNA from each selected adult mouse tissue and E14.5 embryos were blotted onto themembrane and hybridized to radiolabeled random-primed Eaf2 probe. Three transcriptswith different molecular weights were detected based on Northern blot analysis. A majorand the most abundant transcript is approximately 1.4 kb in length. A second minortranscript that was detected only in the brain, lung, and ovary was approximately 2.6 kblong (black arrowhead). A third minor RNA species was detected in the spleen sample thatcorresponds to a transcript size of approximately 1 kb (black arrow). Lane labels: M, RNAsize marker; H, heart; B, brain; Li, liver; S, spleen; K, kidney; E, embryonic day 14 embryo; Lu,lung; T, thymus; Te, testis; O, ovary. B: Total RNAs were isolated from a 10-week-old femalemouse. Twenty to 50 �g of RNA per lane was blotted onto nylon membrane and hybridizedto 32P-labeled random-primed Eaf2 probe. The membrane was stripped and rehybridizedwith glucose-6-phosphate dehydrogenase (GPDH) to show relative amount of RNA load-ing. Lane labels: MG, mammary gland; Skn, skin; Mus, skeletal muscle.
278 LI ET AL.

cloning of Eaf2. However, we werenot able to demonstrate its interac-tion with Msx1 by immunoprecipita-tion.
Nonradioactive In SituHybridization
Embryos were fixed in 4% parafor-maldehyde overnight. Fixed em-bryos were then transferred into 30%sucrose and followed by equilibra-tion in 15% sucrose in OCT beforeembedment in OCT. Cryosections of20 microns thick were collectedonto Histoplus slides (Fisher Scien-tific). Sense and antisense RNAprobes were generated by incorpo-rating digoxigenin–UTP (Roche Bio-chemicals), according to the manu-facturer’s recommendation. Beforehybridization, sections were treatedwith 25 �g/ml proteinase K and post-fixed in 0.2% glutaraldehyde and4% paraformaldehyde. Hybridizationwas carried out at 65°C with a probeconcentration of 1–2 �g/ml in 50%formamide, 5� standard saline ci-trate (SSC), 0.1% Tween-20, 0.1%CHAPS, 0.2 mg/ml yeast tRNA,0.005M ethylenediaminetetraaceticacid, 50 �g/ml Haperin, and 1%blocking reagent (Roche Biochemi-cals). Posthybridization washes werecarried out once in 2 � SSC/0.1%CHAPS at 65°C, once with 1 �g/mlRNAse A in 2� SSC at 37°C, once in2� SSC/0.1% CHAPS at 65°C, andthe final wash in 0.2� SSC/0.1%CHAPS at 65°C. Sections were thenblocked with 20% heat-inactivatedgoat serum and incubated over-night with mouse anti-digoxigeninantibody that carries alkaline phos-phatase conjugate at 4°C. After ex-tensive washes with phosphate-buff-ered saline, sections were subjectedto colorimetric development in thepresence of nitroblue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate(NBT/BCIP) solution (Roche Bio-chemicals). Washed sections werethen post-fixed in 4% paraformalde-hyde, washed, and cover-slidemounted. Images were collected byusing a Spot digital cameramounted on an Olympus BX-50 lightmicroscope.
Northern Blot Analysis
A multiple-tissue Northern blot thatcontained 2 �g of poly(A) RNA from10 mouse tissues was hybridized andwashed according to the manufac-turer’s recommendations (Ambion,Austin, TX). The mice used for RNAisolation were of mixed sex, 8–10weeks old. The embryonic RNA wasextracted from E14.5 embryos. Addi-tional Northern blot that containedapproximately 20–50 �g of totalRNA per lane was probed andwashed as previously described (Liuet al., 1999). The 32P-labeled probefor hybridization was generated byrandom-priming using a purified 1-kbEaf2 cDNA fragment as the tem-plate.
ACKNOWLEDGMENTWe thank Dr. Cheng-Ming Chuongand Dr. Ping Wu for providing helpon in situ hybridization. The Eaf2 se-quence has been deposited in theGenBank and can be accessed byusing GenBank accession no.AY034479.
REFERENCES
Brown NL, Patel S, Brzezinski J, Glaser T.2001. Math5 is required for retinal gan-glion cell and optic nerve formation.Development 128:2497–2508.
Conaway JW, Conaway RC. 1999. Tran-scription elongation and human dis-ease. Annu Rev Biochem 68:301–319.
DiMartino JF, Miller T, Ayton PM, LandeweT, Hess JL, Cleary ML, Shilatifard A. 2000.A carboxy-terminal domain of ELL is re-quired and sufficient for immortaliza-tion of myeloid progenitors by MLL-ELL.Blood 96:3887–3893.
Eissenberg JC, Ma J, Gerber MA, Chris-tensen A, Kennison JA, Shilatifard A.2002. dELL is an essential RNA polymer-ase II elongation factor with a generalrole in development. Proc Natl AcadSci U S A 99:9894–9899.
Emerson BM. 2002. Specificity of generegulation. Cell 109:267–270.
Gall JG. 2000. Cajal bodies: the first 100years. Annu Rev Cell Dev Biol 16:273–300.
Johnstone RW, Gerber M, Landewe T,Tollefson A, Wold WS, Shilatifard A.2001. Functional analysis of the leuke-mia protein ELL: evidence for a role inthe regulation of cell growth and sur-vival. Mol Cell Biol 21:1672–1681.
Kanda Y, Mitani K, Kurokawa M, Yama-gata T, Yazaki Y, Hirai H. 1998. Overex-
pression of the MEN/ELL protein, anRNA polymerase II elongation factor,results in transformation of Rat1 cellswith dependence on the Lysine-rich re-gion. J Biol Chem 273:5248–5252.
Kamura T, Burian D, Khalili H, Schmidt SL,Sato S, Liu WJ, Conrad MN, ConawayRC, Conaway JW, Shilatifard A. 2001.Cloning and characterization of ELL-as-sociated proteins EAP45 and EAP20 arole for yeast EAP-like proteins in regu-lation of gene expression by glucose. JBiol Chem 276:16528–16533.
Liu YH, Tang Z, Kundu RK, Wu L, Luo W, ZhuD, Sangiorgi F, Snead ML, Maxson R.1999. Msx2 gene dosage influences thenumber of proliferative osteogeniccells in growth centers of the develop-ing murine skull: a possible mechanismfor MSX2-mediated craniosynostosis inhumans. Dev Biol 205:260–274.
Luo RT, Lavau C, Du C, Simone F, PolakPE, Kawamata S, Thirman MJ. 2001. Theelongation domain of ELL is dispens-able but its ELL-associated Factor 1 in-teraction domain is essential for MLL-ELL-induced leukemogenesis. Mol CellBiol 21:5678–5687.
Menko AS. 2002. Len epithelial cell differ-entiation. Exp Eye Res 75:485–490.
Miller T, Williams K, Johnstone RW, Shilati-fard A. 2000. Identification, cloning, ex-pression, and biochemical character-ization of the testis-specific RNApolymerase II elongation factor ELL3.J Biol Chem 275:32052–32056.
Mitani K, Kanda Y, Ogawa S, Tanaka T,Inazawa J, Yazaki Y, Hirai H. 1995. Clon-ing of several species of MLL/MEN chi-meric cDNAs in myeloid leukemia witht(11:19)(q23:p13.1) translocation. Blood85:2017–2024.
Mitani K, Yamagata T, lida C, Oda H,Maki K, Ichikawa M, Asai T, Honda H,Kurokawa M, Hirai H. 2000. Nonredun-dant roles of the elongation factorMEN in postimplantation develop-ment. Biochem Biophys Res Commun279:563–567.
Pena-Rangel MT, Rodriguez I, Riesgo-Es-covar JR. 2002. A misexpression studyexamining dorsal thorax formation inDrosophilamelanogaster.Genetics160:1035–1050.
Polak PE, Simone F, Kaberlein JJ, Luo RT,Thirman MJ. 2003. ELL and EAF1 are Ca-jal body components that are dis-rupted in MLL-ELL leukemia. Mol BiolCell 14:1517–1528.
Schmidt AE, Miller T, Schmidt SL, Shiekhat-tar R, Shilatifard A. 1999. Cloning andcharacterization of the EAP30 subunitof the ELL complex that confers dere-pression of transcription by RNA poly-merase II. J Biol Chem 274:21981–21985.
Shilatifard A. 1998. Identification and pu-rification of the Holo-ELL complex. Evi-dence for the presence of ELL-associ-ated proteins that suppress thetranscriptional inhibitory activity of ELL.J Biol Chem 273:11212–11217.
Shilatifard A, Lane WS, Jackson KW, Con-away RC, Conaway JW. 1996. An RNA
CLONING AND EXPRESSION ANALYSIS OF MOUSE Eaf2 279

polymerase II elongation factor en-coded by the human ELL gene. Sci-ence 271:1873–1876.
Shilatifard A, Haque D, Conaway RC,Conaway JW. 1997. Structure andfunction of RNA polymerase II elonga-tion factor ELL. Identification of twooverlapping ELL functional domainsthat govern its interaction with poly-merase and the ternary elongationcomplex. J Biol Chem 272:22355–22363.
Simone F, Polak PE, Kaberlein JJ, Luo RT,Levitan DA, Thirman MJ. 2001. EAF1, anovel ELL-associated factor that is de-localized by expression of the MLL-ELLfusion protein. Blood 98:201–209.
Simone F, Luo RT, Polak PE, Kaberlein JJ,Thirman MJ. 2003. ELL-associated fac-tor 2 (EAF2), a functional homolog ofEAF1 with alternative ELL binding prop-erties. Blood 101:2355–2362.
Thirman M, Levitan D, Kobayashi H, SimonM, Rowley J. 1994. Cloning of ELL, a
gene that fuses to MLL in a t(11;19)(q23;p13.1) in acute myeloid leuke-mia. Proc Natl Acad Sci U S A 91:12110–12114.
Thirman MJ, Diskin EB, Bin SS, Ip HS, MillerJM, Simon MC. 1997. Developmentalanalysis and subcellular localization ofthe murine homologue of ELL. Proc NatlAcad Sci U S A 94:1408–1413.
Young RW. 1985. Cell differentiation inthe retina of the mouse. Anat Rec 212:199–205.
280 LI ET AL.