expression of peg1 (mest) in the developing mouse heart: involvement in trabeculation
TRANSCRIPT

BRIEF COMMUNICATIONS
Expression of Peg1 (Mest) in the Developing Mouse Heart:Involvement in TrabeculationTIM KING,1* YVETTE BLAND,1 SANDRA WEBB,1 SHEILA BARTON,2 AND NIGEL A. BROWN1
1Department of Anatomy and Developmental Biology, St. George’s Hospital Medical School, University of London,London, United Kingdom2The Wellcome Trust/Cancer Research UK, Institute of Cancer and Developmental Biology, Cambridge, United Kingdom
ABSTRACT Peg1 (Mest) is an imprintedgene of unknown function widely expressed inthe mouse embryo, predominantly in cells ofthe mesodermal lineage. We have revealed a re-stricted expression pattern within the developingheart. Initial uniform expression throughout thelinear heart tube subsequently becomes restricted,primarily to the developing myocardial trabeculaeof both the atria and ventricles, where it persistsinto late development. Expression in the atrialappendage myocardium precedes the emergenceof trabeculae (pectinate muscles), and occursearlier and to a greater extent on the right thanon the left, reflecting the spatial and temporalpattern of trabeculation. Analysis of myocardialmorphology in mice lacking the Peg1 gene, whichare viable and appear grossly normal, reveals asubtle alteration in the pattern of trabeculation:an increase in thickness and reduction in densityof the compact myocardium, similar to that seenin the human cardiomyopathy ventricular non-compaction. © 2002 Wiley-Liss, Inc.
Key words: Peg1; Mest; trabeculae; heart; myo-cardium; atrial appendage; pecti-nate muscles; left right asymmetry;ventricles; noncompaction; imprint-ing
INTRODUCTION
The lumen of the vertebrate heart is ramified bymyocardial projections or trabeculae. These structuresform early in embryonic development and their partialcompaction provides the bulk of outer compact myocar-dium (reviewed in Sedmera et al., 2000). In a subtrac-tive screen designed to identify genes differentiallyexpressed between the left and right atrial appendagesof 30–35 somite mouse embryos, we isolated a frag-ment of the paternally imprinted gene Peg1 or Mest(Sado et al., 1993; Kaneko-Ishino et al., 1995). Peg1encodes a protein of unknown function that is widelyexpressed during development (Kaneko-Ishino et al.,1995; Lefebvre et al., 1998). Although mice lacking
Peg1 are viable (Lefebvre et al., 1998), we describe herea potential role for Peg1 in development of trabeculaeand suggest that mice lacking Peg1 function may pro-vide a model for isolated left ventricular noncompac-tion (Jenni et al., 2001).
RESULTS AND DISCUSSION
We have examined expression of Peg1 within theheart and find a dynamic expression pattern associatedwith the finger-like myocardial protrusions termed tra-beculae, which ramify the lumen of both the ventriclesand atria.
Before looping, expression of Peg1 is found uniformlythroughout the heart tube (data not shown). As loopingproceeds during days 9 and 10, expression becomesconcentrated in the ventricular myocardium, beingstrongest in the trabeculae, which have started project-ing into the ventricular lumen. Some endocardial ex-pression is also apparent in the ventricles (Fig. 1A–C).In the common atrium at the 12 somite stage (late day9), in which trabeculae (pectinate muscles) have not yetformed, expression is absent from myocardium but isapparent in endocardium (Fig. 1A,A�). Expression isabsent from the myocardial component of the atrioven-tricular junction (AVJ) and the outflow tract (OFT)(Fig. 1A–C). By 15 somites (day 10), expression startsto appear in the myocardium of the future right atrialappendage (Fig. 1B,C) but remains absent in the leftappendage, OFT, and AVJ. By day 11, expression isfound in the myocardium of both the atrial appendages,being, however, significantly stronger on the right thanthe left (Fig. 1E,H). Expression remains absent fromthe OFT myocardium and both myocardium and endo-cardial cushions of the AVJ. Endocardial cells them-
Grant sponsor: British Heart Foundation; Grant number:RG/1998004
*Correspondence to: Tim King, Department of Anatomy and Devel-opmental Biology, St. George’s Hospital Medical School, CranmerTerrace, London, SW17 ORE, UK. E-mail: [email protected]
Received 16 April 2002; Accepted 4 July 2002DOI 10.1002/dvdy.10142Published online 28 August 2001 in Wiley InterScience (www.
interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 225:212–215 (2002)
© 2002 WILEY-LISS, INC.
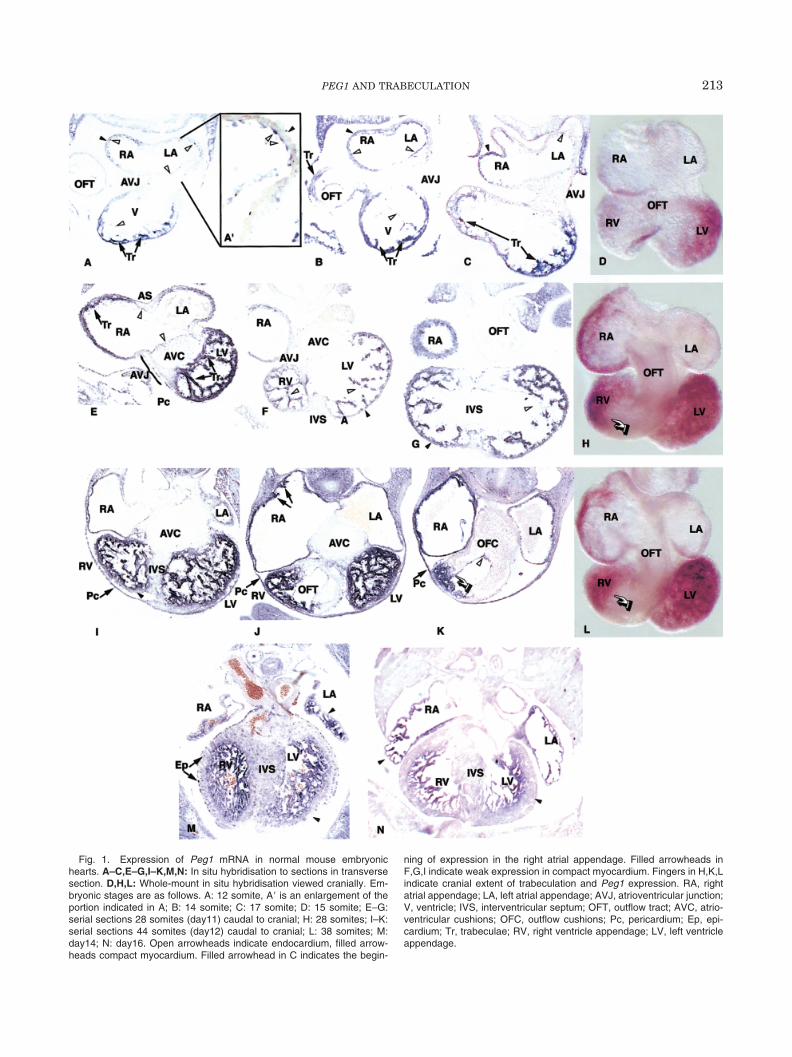
Fig. 1. Expression of Peg1 mRNA in normal mouse embryonichearts. A–C,E–G,I–K,M,N: In situ hybridisation to sections in transversesection. D,H,L: Whole-mount in situ hybridisation viewed cranially. Em-bryonic stages are as follows. A: 12 somite, A� is an enlargement of theportion indicated in A; B: 14 somite; C: 17 somite; D: 15 somite; E–G:serial sections 28 somites (day11) caudal to cranial; H: 28 somites; I–K:serial sections 44 somites (day12) caudal to cranial; L: 38 somites; M:day14; N: day16. Open arrowheads indicate endocardium, filled arrow-heads compact myocardium. Filled arrowhead in C indicates the begin-
ning of expression in the right atrial appendage. Filled arrowheads inF,G,I indicate weak expression in compact myocardium. Fingers in H,K,Lindicate cranial extent of trabeculation and Peg1 expression. RA, rightatrial appendage; LA, left atrial appendage; AVJ, atrioventricular junction;V, ventricle; IVS, interventricular septum; OFT, outflow tract; AVC, atrio-ventricular cushions; OFC, outflow cushions; Pc, pericardium; Ep, epi-cardium; Tr, trabeculae; RV, right ventricle appendage; LV, left ventricleappendage.
213PEG1 AND TRABECULATION
selves continue to express throughout the heart (Fig.1E–G) and do so at all stages examined. Expression isnot detected in the compact myocardium of the devel-oping interventricular septum (IVS). Even in the verythin outer layer of compact myocardium, expression islower than in trabeculae, and nonexpressing patchesare detected (Fig. 1F,G).
By day 12 it is clear that expression in the ventricu-lar compact outer myocardium is either absent or muchlower than in the trabeculae (Fig. 1I). There is still noexpression in the IVS, OFT, AV myocardium, and theendocardial cushions. There is a sharp boundary ofexpression between the future right ventricle and OFT,corresponding to the cranial extent of trabeculation(Fig. 1H,K,L). Trabeculae are now becoming more ap-parent in the atrial appendages, particularly the right,
and this finding correlates with increased levels of ex-pression (Fig. 1J,K). We have examined embryos up today 16 of gestation and find that the pattern of expres-sion then remains essentially unchanged. The cleardistinction between compact and trabeculated myocar-dium becomes more apparent and is visible now in theatria as well as ventricles (Fig. 1M,N). Expression isalso found in the epicardium at day 11 and 12 (Fig.1E,I). The punctate staining apparent in the compactmyocardium at day 14 (Fig. 1M) may correspond to theepicardium that is now migrating into the myocardiumto contribute to the coronary vasculature (Mikawa andGourdie, 1996). We have failed to detect significantexpression levels in adult heart, although low levelshave been detected by Northern blot (Kaneko-Ishino etal., 1995; Lefebvre et al., 1998).
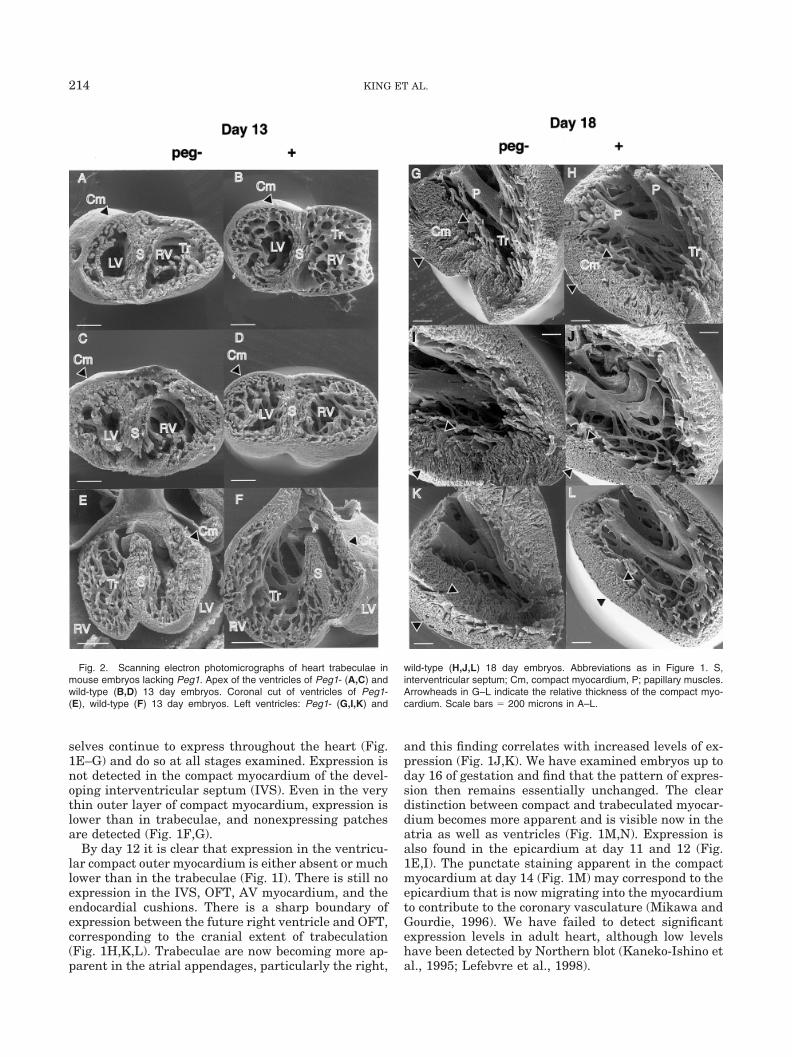
Fig. 2. Scanning electron photomicrographs of heart trabeculae inmouse embryos lacking Peg1. Apex of the ventricles of Peg1- (A,C) andwild-type (B,D) 13 day embryos. Coronal cut of ventricles of Peg1-(E), wild-type (F) 13 day embryos. Left ventricles: Peg1- (G,I,K) and
wild-type (H,J,L) 18 day embryos. Abbreviations as in Figure 1. S,interventricular septum; Cm, compact myocardium, P; papillary muscles.Arrowheads in G–L indicate the relative thickness of the compact myo-cardium. Scale bars � 200 microns in A–L.
214 KING ET AL.

Although mice lacking Peg1 function are viable, withno reported cardiac abnormality (Lefebvre et al., 1998),our expression analysis led us to examine the heartmorphology of such embryos. At days 12, 13, 17, and18, we can detect no gross morphologic abnormality inPeg1 mutant embryos, aside from growth retardation.However, examination by scanning electron micros-copy revealed a subtle but consistent alteration to thepattern of trabeculae of the mutant hearts. At day 13,there appears to be a reduction in the extent of trabec-ulae compared with the volume of the lumen within theventricles, and the appearance of the trabeculae them-selves are slightly aberrant, being rather thicker andmore irregular. There also appears to be an increase inthe thickness of the compact myocardium and the IVS(Fig. 2A–F). At day 18, a similar reduction in the extentand regularity of trabeculation is apparent (Fig. 2G–L).Again, the outer layer of myocardium appears thicker,but also less dense (compare Fig. 2I,K with 2J,L) thanin wild-type hearts. There also appears to be a similareffect on atrial morphology in Peg1 mutants, but be-cause normal atrial walls are thin and trabeculationmuch less pronounced, we cannot be certain on thispoint.
The widened but less dense ventricular compactmyocardium is reminiscent of a human cardiomyopa-thy termed “noncompaction” (Jenni et al., 2001). Al-though Peg1 mutant mice are viable and examinationof the male colony history reveals normal longevity andfecundity, we have not yet carried out experiments totest cardiovascular performance under stress. Thispursuit, together with a more detailed histologic anal-ysis of the myocardial structure, will determine thevalidity of Peg1 mutants as a model for noncompaction.
The mechanism of Peg1 action in myocardial mor-phogenesis is unclear. Although there is no absoluterequirement for Peg1 in the formation of trabeculae, itdoes in some way contribute to their normal growthand organisation. Mice lacking Peg1 do exhibit interu-terine growth retardation with a birth weight of ap-proximately 85% of normal littermates (Lefebvre et al.,1998). This finding suggests a role for Peg1, as for mostimprinted genes, in control of growth. Indeed, trabec-ulae are a site of rapid differential growth, but lack ofan understanding of the function of the Peg1 protein,however, prohibits further speculation.
EXPERIMENTAL PROCEDURES
Peg1 is expressed only from the paternal allele, somice lacking Peg1 function were generated as heterozy-
gotes carrying one maternal copy and one copy of atransgenic targeted mutation, Peg1tm1Lef, which haslacZ replacing exons 3–8 of Peg1 (Lefebvre et al., 1998).This was achieved by crossing of Peg1 �/ tm1Lef malesto wild-type females. Transgenic embryos (designatedPeg1 �/- for maternal Peg1/paternal Peg1tm1Lef) ex-press no wild-type Peg1 mRNA but express �-galacto-sidase. These embryos, thus, were detected by X-galstaining for �-galactosidase activity in yolk sac andlimb bud mesenchyme, by using previously describedmethods (Lefebvre et al., 1998). Scanning electron mi-croscopy was carried out as described previously (Webbet al., 1998). In situ hybridisation on whole-mounts andsections were carried out as previously described (Hen-rique et al., 1995; Moorman et al., 2001) with the mod-ification of using Hybri-Well chambers (Grace BioLabs)to ensure that the sections were hermetically sealedduring hybridisation. A fragment comprising bases1470-2330 of the mouse Peg1 gene (accession no. NM-008590) was used as a probe for both whole-mountsand sections.
REFERENCES
Henrique D, Adam J, Myat A, Chitnis A, Lewis J, Ish-Horowicz D.1995. Expression of a Delta homologue in prospective neurons inthe chick. Nature 375:787–790.
Jenni R, Oechslin E, Schneider J, Jost C, Kaufmann PA. 2001. Echo-cardiographic and pathoanatomical characteristics of isolated leftventricular non-compaction: a step towards classification as a dis-tinct cardiomyopathy. Heart 86:666–671.
Kaneko-Ishino T, Kuroiwa Y, Miyoshi N, Kohda T, Suzuki R,Yokoyama M, Viville S, Barton SC, Ishino F, Surani M.A. 1995.Peg1/Mest imprinted gene on chromosome 6 identified by cDNAsubtraction hybridisation. Nat Genet 11:52–59.
Lefebvre L, Viville S, Barton SC, Ishino F, Keverne EB, Surani MA.1998. Abnormal maternal behaviour and growth retardation asso-ciated with loss of the imprinted gene Mest. Nat Genet 20:163–169.
Mikawa T, Gourdie RG. 1996. Pericardial mesoderm generates apopulation of coronary smooth muscle cells migrating into the heartalong with ingrowth of the epicardial organ. Dev Biol 174:221–232.
Moorman AF, Houweling AC, de Boer PA, Christoffels VMJ. 2001.Sensitive nonradioactive detection of mRNA in tissue sections:novel application of the whole-mount in situ hybridization. Histo-chem Cytochem 49:1–8.
Sado T, Nakajima N, Tada M, Takagi N. 1993. A novel mesoderm-specific cDNA isolated from a Mouse embryonal carcinoma cell line.Dev Growth Differ 35:551–560.
Sedmera D, Pexieder T, Vuillemin M, Thompson RP, Anderson RH.2000. Developmental patterning of the myocardium. Anat Rec 258:319–337.
Webb S, Brown NA, Anderson RH. 1998. Formation of the atrioven-tricular septal structures in the normal mouse. Circ Res 82:645–656.
215PEG1 AND TRABECULATION