external diffusion effects on the kinetic constants of immobilized enzyme systems
DESCRIPTION
External Diffusion Effects on the Kinetic Constants ofImmobilized Enzyme SystemsTRANSCRIPT

f. theor. Biol. (1980) 84, 259 -279
External Diffusion Effects on the Kinetic Constants of Immobilized Enzyme Systems
SUN BOK LEE AND DEWEY D. Y. RYU
The Korea Advanced Institute of Science, P.O. Box 150, Chung- Ryang-Ri, Seoul, Korea
(Received 29 May 1979, and in revised [orm 29 November 1979)
In order to further our understanding of immobilized enzyme reaction kinetics, the effects of external diffusion on the kinetic constants are studied for various reaction systems. It is shown that the variations of apparent kinetic constants with diftusional limitations are not the same as those of Michaelis-Menten kinetics, although both the inhibition kinetics and the two substrate reaction kinetics can be expressed in the form of simple Michaelis-Menten type by using apparent rate parameters. We found that there will be changes in the apparent kinetic constants depending not only on the types of enzyme reaction kinetics but also on the relative rate of diffusion or flow-through in the microenvironment. For substrate inhibition kinetics, both the apparent maximum reaction rate and the apparent Michaelis constant decrease while the apparent maximum reaction rate for product inhibition kinetics is increased and the apparent Michaelis constant decrease as mass transfer limitation is reduced. In the case of two substrate enzyme reaction kinetics, as the diffusio'nal limitation is reduced the apparent maximum reaction rate increase but the apparent Michaelis constant can increase, decrease or remain nearly constant depending on the values of relative affinity and on the fixed substrate concentrations. The results of theoretical analyses are compared with the experimental data obtained and reported previously, and a very good agreement was found.
1. Introduction
When the enzymes are immobilized, such changes as structural con- formation, microenvironment , the partitioning or electrostatic effect and mass transfer resistance could be brought about (Goldstein, 1976). A m o n g these the effects of mass transfer limitation on the reaction kinetics have been widely studied by many workers from the viewpoint of heterogeneous catalysis.
The effects of internal diffusion in the reaction kinetics have been studied experimentally as well as theoretically (Lee, Fratze, Wun & Tsao, 1976; Moo-Young & Kobayashi, 1972; Engasser & Horvath , 1973). In the case of external diffusion, however, the results of several workers (Lilly etal., 1966;
259 0022-5193/80/100259+21 $02.00/0 �9 1980 Academic Press Inc. (London) Ltd.

260 S . B . L E E A N D D. D. Y. RYLI
Wilson, Kay & Lilly, 1968a, 1968b; Sharp et al., 1969; Tosa, Mori & Chibata, 1971; Kobayashi & Moo-Young, 1973; Gellf, Thomas, Broun & Kerneves, 1974: Cho & Swaisgood, 1974; Toda, 1975; Hirano, Karube, Matsunaga & Suziki, 1977; Paul, Coulet, Gautheron & Engasser, 1978; Kim, Lee & Ryu, 1978) appear to be contradictory and conflicting.
Most of these authors cited made some endeavors to show the important factors that affect the kinetic constants of immobilized enzymes. Many more works also attempted to analyze theoretically the effects of external diffusion on the kinetic constants. For irreversible Michaelis-Menten kinetics, Hornby, Lilly & Crook (1968), Kobayashi & Moo-Young (1971), Shuler, Aris & Tsuchiya (1972), Hamilton, Stockmeyer & Colton (1973), Kobayashi & Laidler (1974), and Toda (1975) derived the equations for apparent kinetic parameters. For reversible Michaelis-Menten kinetics, Lee, Kim & Ryu (1979) showed that the extent the apparent kinetic :constants changes as a function of superficial velocity depends on the value of l l Vm/Km parameter for forward reaction and for reverse reaction, respectively. For other enzyme reaction kinetics, the effects of external diffusion on the kinetic constant have not been clearly established up to the present.
In view of these developments and some confusion, our study on the effects of external diffusion was undertaken in order to further our under- standing of reaction kinetics of immobilized enzymes. In this paper, the external diffusion effects on the kinetic constants were studied theoretically and the confusion due to conflicting results was clarified.
2. The System
It is assumed that the system is at a steady state, the enzymes are immobilized On a nonporous support material, and the diffusion coefficient is constant.
For the reaction system that a substrate, s, is converted to a product p, the diffusive fluxes can be expressed as
D ds Js = S-~y (I)
4 =Dpd~ (2) with the boundary conditions
s=S, ! p = P
s=S, p = P
aty = 0
a tY = 8

DIFFUSION EFFECTS ON ENZYME KINETICS 2 6 1
where the mass fluxes toward the surface of the immobilized enzyme are positive and the origin of the co-ordinate system is at the surface. Under steady state, J, = -Jp =3", and the Equations (1) and (2) yield the following expression
J = k L s ( S - - S ) = k r e C P - P ) (3)
where kLs = D s / 8 and kt .p = D p / & For the two substrate enzyme reaction system the diffusive flux can be
expressed as
J = k u ( S , - S,) = kLi(Si -- 4 ) (4)
which is analogous to equation (3).
3. Theoretical Examination of the Mass Transfer Effect on the Kinetic Constants
(A) M E T H O D OF ANALYSIS
Some workers defined the apparent Michaelis constant as that cor- responds to a value at whicl~ the rate of enzyme reaction is half of the maximum reaction rate (Goldman, Kedem & Katchalski, 1971; Sundaram et al., 1972; Kobayashi & Laidler, 1974), although it cannot describe the overall reaction rate exactly (Horvath & Engasser, 1974; Kobayashi & Laidler, 1974). In general, for the homogeneous enzyme reaction system, the maximum reaction rate is defined as the reaction rate at a high substrate concentration or S-~ co and the Michaelis constant is the substrate concen- tration at which the reaction rate is half the maximal. If these concepts are applied to the heterogeneous system, then we can define that
VJtrn(app ) = lim R (5) $ ~ o 0
I !
gm(app ) ---- S l a t R =0.5 V:,.pp, (6)
where R represents the reaction rate of immobilized enzymes. This type of analysis was found to be very useful if the trends or variations of kinetic constants with diffusional limitations are to be determined, especially for the complex enzyme reaction system.
It is now convenient to introduce the following dimensionless variables,
R s v " , O r = - - / 2 , =
= V'. , K' . , . ' kLsK '~
where, ~" represents the dimensionless reaction rate, o" the dimensioniess substrate concentration, and Ix the mass transfer modulus.

2 6 2 s . B . LEE AND D. D. Y. RYLI
Equations (5) and (6) becomes
a * = lim ~" (7) o - - ~ oO
/3* = r s o* (8)
where a* and/3* represent the dimensionless apparent maximum reaction rate and the apparent Michaelis constant, respectively.
We will now examine the effects of diffusional limitations on the kinetic parameters for various enzyme reactions by using equations (7) and (8). The purpose of this examination is to find how a* and/3* can be affected as the mass transfer modulus, it, changes.
(B) ONE SLIBSTRATE ENZYME R E ACTI ON SYSTEMS
( i) Michaelis-Menten kinetics
When the enzymes are immobilized, the enzyme reaction takes place at the surface and the reaction rate for this heterogeneous enzyme reaction system can be expressed as
v ' g g = K " +-----ff (9)
where, R is the observed global rate of heterogeneous reaction per unit area. Equation (9) may be rewritten in terms of measurable bulk substrate concentration instead of surface concentration
/ /
V,nS R = K ~ +S" (10)
From equation (3), S = S - ( J / k ~ ) and substitution into equation (9) yields
~" = ( 1 1 ) 1 + o . - ~rtx
in dimensionless form since the mass transfer flux, J, will be equal to the rate of reaction, R, under steady state. Equation (11) yields
= ~ {( 1 +/.t + o.) - [(1 + tt + o.)2 _ 4bto.] 1/2} (12a)
= 2o'{(1 +/z +o-)+[(1 +/z + o.)2- 4/zo.]1/2}-1 (12b)
since 0-< s r-< 1. From equation (12), for Michaelis-Menten kinetics
a * = l : (13)
/3* = 1+0.5/~ (14)

DIFFUSION EFFECTS ON E N Z Y M E KINETICS 263
From these equations one finds that the apparent maximum reaction rate, a*, is constant and the apparent Michaelis constant, /~*, increases with diffusional limitations (i.e. with increasing t~). An expression similar to equation (14) was previously defined as apparent Michaelis constant by Kobayashi & Laidler (1974), although the relationship holds only for the one substrate Michaelis-Menten type enzyme reaction system.
(ii) Substrate inhibition
The substrate inhibition kinetics expressed as
v ' g R =
K " + S + ~ ' S /Ki~
can be rewritten in a dimensionless form
(15)
where,
~" = 1 +,7-( tL +Ks(o"- ~'tz) 2 (16)
K " K,=
K~,"
Equation (16) yields a cubic equation for ~" and its analytical solution is complex. !n Fig. 1 the Lineweaver-Burk plot is shown at Ks = 0.05 using the result of computer simulation. There is little change in K~', with the variation of t* while V~, and K~, vary similarly to those of Michaelis-Menten kinetics.
(iii) Product inhibition
There are, in general, three types of product inhibition, that is, competi- tive, non-competitive, and anti-competitive inhibitions (Laidler & Bunting, 1973). For competitive product inhibition kinetics,
v ' g R Kin(1 + W K , p ) + g (17)
can be expressed as
o ' - 6 ~" = (18)
(1 + ~: -t- gd~ ' ) + o- - ~'tz
where
P K " k ~ ~ = ~',.p, andKp =K~p kLp
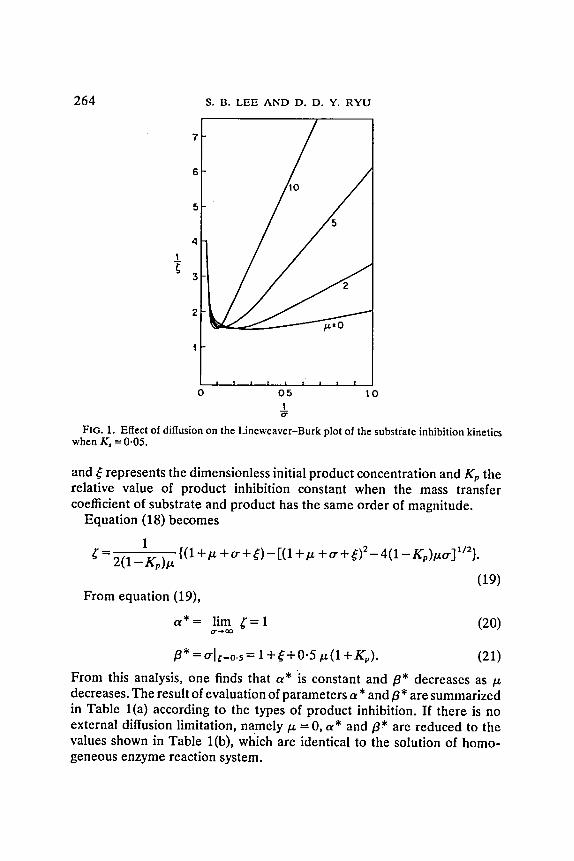
264
4
&
3
S. B. LEE AND D. D. Y. RYU
t ! = i 0 8 5 f t t t 0 10
o"
Flo. I. Effect of diffusion on the Lineweaver-Burk plot of the substrate inhibition kinetics when K, = 0.05.
and ~ represents the dimensionless initial product concentration and Kp the relative value of product inhibition constant when the mass transfer coefficient of substrate and product has the same order of magnitude.
Equation (18) becomes
1 ~" = 2(1 - Kp)/z {(1 +/.t + t r + so)-[(1 +/z +tr +sr -Kp)l.ttr]]/2}.
(19) From equation (19),
a * = lim ~'= 1 (20) o - ~ o o
~* = o'l~=o.s = 1 +~+0.5 / . t (1 + K~,). (21)
From this analysis, one finds that a* is constant and fl* decreases as /z decreases. The result of evaluation of parameters a * and fl* are summarized in Table l(a) according to the types of product inhibition. If there is no external diffusion limitation, namely/z = 0, a* and fl~' are reduced to the values shown in Table l(b), which are identical to the solution of homo- geneous enzyme reaction system.

DIFFUSION EFFECTSON ENZYME KINETICS
TABLE 1
Values of or* and [3" for product inhibition kinetics
265
(a) With diffusional limitations
Inhibition type a* /3'
Competitive
Non-competitive
Anti-competitive
1 1 +6+0.5/,t(1 +Kp)
1 2 Kpp,2X3+(I +,~-Kp)p.X2- (l +6+p.)X I'4(1+6) +4Kptz-(l +6)] Ktd.tX2 +(l + 6)X - 1
2_K_~_~ 1-4(1 + 6)2 +4Kp~_ (1 + 6)] Kp ~2X3+(l+6)v'X2-(I+~)X KplzX2+ (1 +6 )X- 1
(b) Without diftusional limitations �9 .
Inhibition type a*l,,.o /3"1,=o
Competitive 1 1 + 6 Non-competitive (1 + 6) -t 1 Anti-competitive (1 + 63-1 . (l + 6)-I
P K~ Where, 6 = K-~.p ' Kp = K~p' X = 0-5 a*
For non -compe t i t i ve p roduc t inhibit ion kinetics, c~* and [3* are p lo t ted against tz in Fig. 2. F r o m this figure we find tha t a * will increase and fl* will decrease as Ix decreases . In addi t ion, we find that a * increases m o r e sharply as Kp increases (or inhibi t ion cons tant is smaller) but [3* decreases m o r e sharply as Kp decreases . Similar results are ob t a ined for an t i - compet i t ive inhibition react ion kinetics.
(C) TWO SUBSTRATE ENZYME REACTION SYSTEM
For two subs t ra te e n z y m e r e a c t i o n s y s t e m s equa t ion (22) can be used (Laidler & Bunt ing, 1973; Paul et al., 1978),
V,.S;$~ R K , , , i K , , q + K , , , f i , + K ' . ~ , . + ~ (221
and such type of two subs t ra te e n z y m e reac t ion kinetics as p ing -pong bi-bi react ion mechan i sm can be ana lyzed by the m e t h o d shown below. F r o m
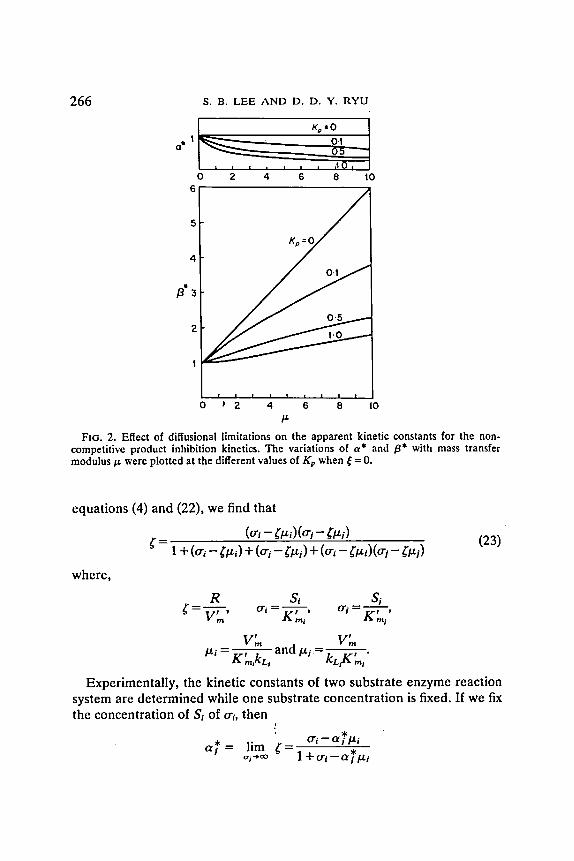
266
a~ , I
0 6
5
4
2
I
S. B. L E E A N D D . D. Y. R Y U
2 4 6 8 10
0.1
�9 0 . 5
! | ! i ! . l , I
0 # 2 4 6 8 tO
FIG. 2. Effect of diffusionai limitations on the apparent kinetic constants for the non- competitive product inhibit ion kinetics. The variations of a * and /3* with mass transfer modulus p, were plotted at the different values of K'p when ~ = 0.
equations (4) and (22), we find that
(o-, - ~ 3 (o'j - g 'ui )
~" = 1 + (o-, - ~'t~i) + (o-j - ( m ) + (o-~ - r - t ' m )
where,
(23)
R Sl S~ ~" = V " cri K ' , ' o ' / = k " '
v 2 v " IX~=K~,kL~ and /z i= k , �9 LrK,.j
Experimentally, the kinetic constants of two substrate enzyme reaction system are determined while one substrate concentration is fixed. If we fix the concentration of Si of o'z, then
o ' i - - ~ ~ i a ~ = lim ~=

D I F F U S I O N E F F E C T S O N E N Z Y M E K I N E T I C S 267
giving
a T = 2o'i{(1 +/zi+o'~)+, (1 +/xt +o'~)2-4/z,.o'i]'/2} -1. (24)
From equation (23) and (24),/3" can be obtained by eliminating o-~.
/3? = (rj
2(1 - - Ot ~)2/Xi = 1+0 .5 a* tq ( 1 - a ] ' ) ( 2 - a * ) / z , + 2 " (25)
The expressions for a* and/3* can also be derived simply by changing the subscript.
Now, we introduce the another dimensionless parameter , K, which is defined as
#, k_~ K ' , 6) K = - - = 0--<K-<I. (2 tzl kL, K~,'
If the mass transfer coefficient of two substrates are the same order of magnitude (in most cases, this assumption is reasonable), K means the relative affinity of two substra~es. In equation (26), the subscript i denotes one of two substrate species having less affinity (or high K " values) and the subscript/" denotes the other having higher affinity (or low K " values) because x is defined as smaller than unity.
Since #~ = Kl.t i, at constant o-j
or* =20"~{(l +KlXl+Ori)+[(l +tctZi+~ri)2--4KIZjO'l]l/2} -~ (27)
2(1 - a*)2Ktzi (28) /37 = 1+0 .5 a*/zi ( 1 - ~ , ~ ) ( 2 - a * ) K ~ i + 2
and at constant o-~
a * = 2o' i ( ( l + # j + o ' l ) + [ ( l + t . t i+o ' i )2-4~,o ' t ]~ /2} -~ ( 2 9 )
/37 = 1 + 0 . 5 (1 - a * ) ( 2 - a* )~ i + 2 (30)
From these results we find that: (1) a~" always increases as/~ decreases regardless of cr and g. (2) /3* increases as p. decreases regardless of K when cr I is smaller than
unity, but 13" can be increased, decreased, or nearly constant depending on the values of K when o- I is greater than..unity.
(3) a * is constant at K = 0 as in the Michaelis-Menten kinetics, and it always increases as/z decreases.
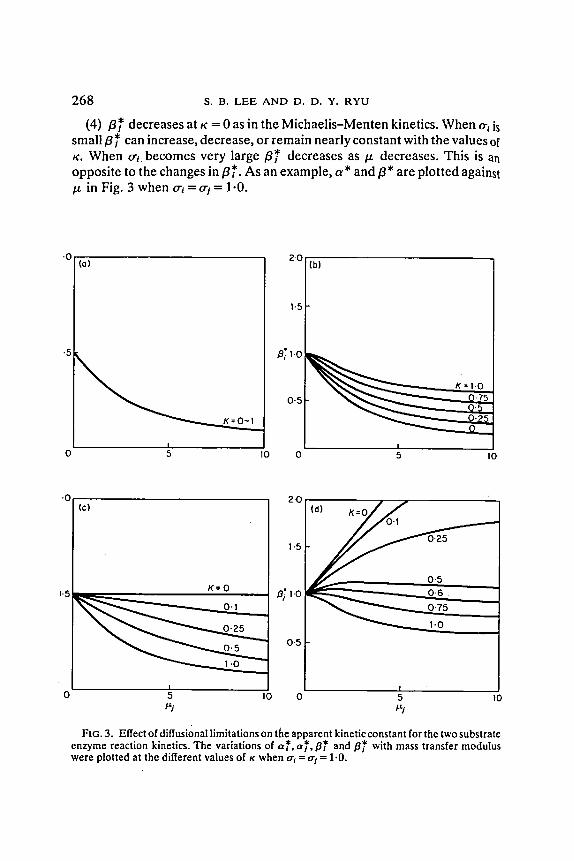
2 6 8 s . B . LEE AND D. D. Y. RYU
(4) f l~ d e c r e a s e s at K = 0 as in t h e M i c h a e l i s - M e n t e n k ine t ics . W h e n o-~ is
sma l l f l* can i n c r e a s e , d e c r e a s e , o r r e m a i n n e a r l y c o n s t a n t w i t h t h e v a l u e s of
K. W h e n o ' t b e c o m e s v e r y l a r g e / ~ * d e c r e a s e s a s / . t d e c r e a s e s . T h i s is an
o p p o s i t e to t h e c h a n g e s in f l* . A s an e x a m p l e , a * a n d / ~ * a r e p l o t t e d against
p. in Fig. 3 w h e n cr~ = tr i = 1"0.
~ I .5
(a) 2-0
1.5
~', o
0-5
I 0 5 10 0
(b)
5 10
"01 2.0 (e)
1.5
,5 K=o ~;,o
0.5
(d)
0.5 o ~ O. 7.~5" 1.0
o 5 10 o 5 I0
FIG. 3. Effect of diffusional limitations on the apparent kinetic constant for the two substrate enzyme reaction kinetics. The variations of a~, a} ~,/3* and fl~' with mass transfer modulus were plotted at the different values of K when o-~ = o- t = 1.0.
DIFFUSION EFFECTS ON ENZYME KINETICS 2 6 9
4. Discussion
Based on the results of this study we found that, when a soluble enzyme is immobilized, there will be changes in the kinetic constants which may either go up or down depending on the types of enzyme kinetics as well as on the relative rate of diffusion or flow-through in the microenvironment.
In the case of one substrate enzyme reaction, from equations (9) and (10) we find that the observed rate parameters are
V~ = V',. (31)
K '.. = K ~ ( S / g ) = K'~ (1 - J / k c s S ) - t (32)
and similarly,
K ,", = K } , ( S / g) = K}, (1 - Y/ kLsS) -1 (33)
K}~, = g~p (P/P) = K ~ (1 +Y/kLpP) - t (34)
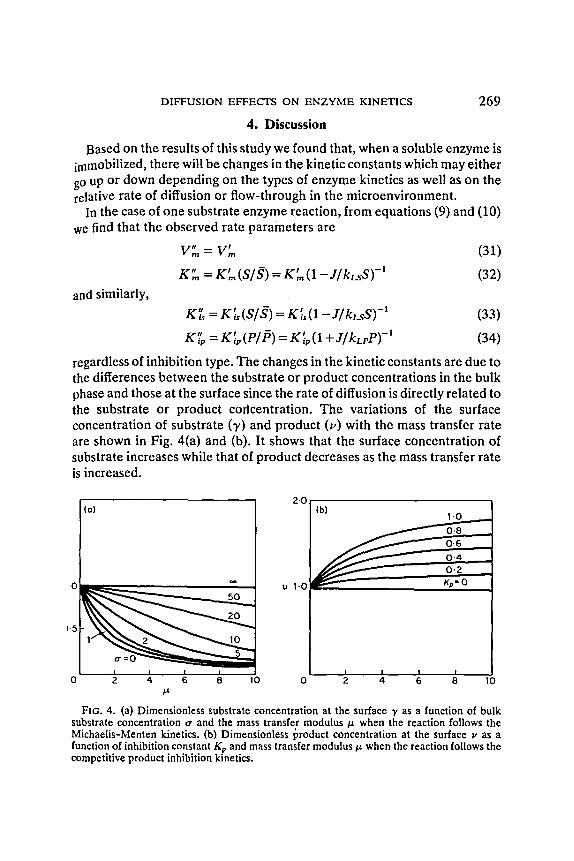
regardless of inhibition type. The changes in the kinetic constants are due to the differences between the substrate or product concentrations in the bulk phase and those at the surface since the rate of diffusion is directly related to the substrate or product corlcentration. The variations of the surface concentration of substrate (7') and product (v) with the mass transfer rate are shown in Fig. 4(a) and (b). It shows that the surface concentration of substrate increases while that of product decreases as the mass transfer rate is increased.
'0 l(a)
0 2 4 6 8 I0
2-0 lb)
1.0
I ! I I O 2 4 6 8 10
FIG. 4. (a) Dimensionless substrate concentration at the surface y as a function of bulk substrate concentration tr and the mass transfer modulus t.t when the reaction follows the Michaelis-Menten kinetics. (b) Dimensionless product concentration at the surface v as a function of inhibition constant Kp and mass transfer modulus/1, when the reaction follows the competitive product inhibition kinetics.

270 s .B. LEE AND D. D. Y. RYLI
From equations (31) and (32) one can find that V~ is constant and K~ is decreased as the transport rate of substrate f rom the bulk solution to the surface is increased. For substrate inhibition kinetics, on the other hand, the increase in the transport rate of substrate will cause the increase in the affinity of substrate to the immobilized enzyme and at the same time the inhibition effect is increased by the substrate. The effect of mass transfer rate on K " values would be to a certain extent similar to that on K ~ values. But the overall diffusion effect is likely to be less owing to the higher range of substrate concentrations involved, as shown in Fig. 1.
For product inhibition kinetics, the changes in Ki'~, are caused by the difference between the product concentration at the bulk phase and that at the surface. Thus, K~, may be increased as mass transfer rate increases. The increase in the transport rate of product from the immobilized enzyme surface to the bulk solution may cause the reduction or elimination of inhibition effect by the product. This phenomenon is different f rom the substrate inhibition kinetics. If the apparent kinetic parameters that reflect the inhibition effects are to be evaluated, the effects of mass transfer rate must be considered. That is, the reduction of inhibition effect with the increasing mass transfen rate may cause an apparent increase in the maxi- mum reaction rate. For competi t ive type, however, V " ( a p p ) is constant since the inhibitor concentration will not affect the maximum reaction rate.
The results of analysis on the effects of external diffusion on the kinetic constants for one-substrate enzyme reaction are summarized in Table 2.
TABLE 2
Summary of external diffusion effects on the observed and apparent rate parameters for one-substrate enzyme reaction system
Reaction type V~ K~, K~ K~'p V~( .pp ) K~t.pp)
Michaelis-Mentenf C - - C(NC-) Substrate inhibition C - - NC- Product inhibition C - - +
competitivet C(NC ~') non-competitive + anti-competitive +
C, constant; NC-, nearly constant, but decreases as kt. (mass transfer rate) increases or vice versa; NC +, nearly constant, but increases as kL (mass transfer rate) increases or vice versa; --, decreases as kL (mass transfer rate) increases or rice versa., +, increases as kL (mass transfer rate) increases or vice versa. 1 When the apparent rate parameters are'determined by conventional graphical method, the apparent maximum reaction rate, V~c,pp), can be varied with the diffusional limitations, as shown in the parenthesis.

DIFFUSION EFFECTS ON E N Z Y M E KINETICS 271
In the case of two substrate enzyme reaction system, the results of theoretical analysis are somewhat complex due to the subtle interactions between two substrates. V~(app) always increases as mass transfer rate increase (except for K = 0), but K~(~pp) can be increased, decreased, or nearly constant depending on (1) the values of relative affinity between the two substrates and (2) on the substrate concentrations where the experi- ments are performed. Recently, Paul etal . , (1978) analysed the effect of two substrates on kinetic constants when the relative affinity is significantly different. If the fixed substrate concentration is very large so that the one substrate is saturated, the changes in the apparent rate constants are identical with those of Michaelis-Menten kinetics. On the other hand, when the fixed substrate concentration is very low, both /3* and /3* can be increased with increasing tz. In any case the changes in /3* (apparent Michaelis constant of the species with less affinity) are always smaller than fl*, and in a limited range of tt,/3* is nearly constant.
Although the rate equations of the inhibition kinetics or two substrate reaction kinetics can be expressed as the simple Michaelis-Menten type of equation using apparent rate parameters, the changes in the apparent kinetic constants as an effect df diffusional limitations are not the same as those in Michaelis-Menten kinetics. For this reason, the results of several workers appear to be contradictory and conflicting.
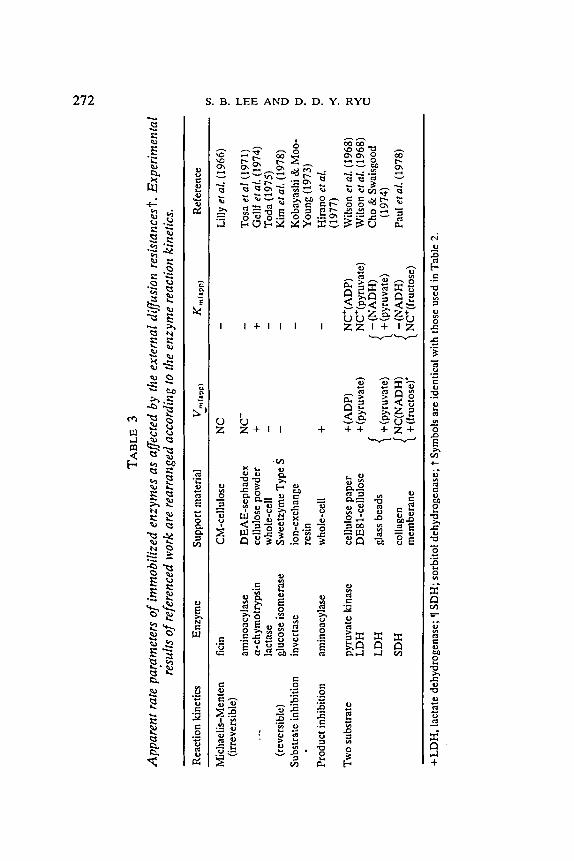
Based on our theoretical analysis, the experimental results of previous workers reported are classified according to the reaction kinetics as shown in Table 3. All experimental data cited in this paper were determined by graphical method and all are apparent rate parameters. The analysis of kinetic data for enzymes, in homogeneous solution and in immobilized form, is often carried out by using a graphical technique such as a Lineweaver- Burk plot. However, this type of representation no longer yields a straight line due to the diffusional limitations. Thus, it should be noted that V~ (app) even for Michaelis-Menten kinetics and for competitive product inhibition kinetics can be varied with the changes in the mass transfer rate as reported previously (Lee, Kim & Ryu, 1979). For Michaelis-Menten kinetics, V~ (app) decreases as the mass transfer rate increases if the rate parameters are determined by the graphical method, but not as greatly as K~ (app). (Refer to Appendix B.) In view of these findings and observations, the experimental data of Tosa et al. (1971), Toda (1975), and Kim et al. (1978) are consistent with our theoretical analysis. Lilly et aL (1966) reported V~ (app) was constant with the flow rate, and this may be due to the high substrate concentration. From ,the equation (B11) one can find that V~ (app) may be nearly constant if/z << 1 +tr, or high values of tr. Under their experimental conditions the substrate concentration was about 30
272
~
~. ".~.
~ ~
t J t , l
~ . m
8
c~
E
E
0
;an
.o
S. B. LEE AND D. D. Y. RYU
= o o ~
I I + I I I bb z~ z b
I Z Z l + I Z
o b Z Z + I I +
0 ~3. U
<3"8 ~ ~.E "8
~E ~zE ++ +Z+
= = -8
e,3
. - y . ~ ~ . -
0 ~' :~ .o
> ,-. ~ . ~ o
ra~
0
0
E
4- -
R
.8
C~ c~
o
+

DIFFUSION EFFECTS ON ENZYME KINETICS 273
times the K " value (Lee & Ryu, 1979). The result of Gellf et al. (1974) is, however, contradictory to our analysis. This conflicting result may be due to incorrectness of the enzyme reaction kinetics that they used. If the a- chymotrypsin follows the Michaelis-Menten kinetics, V"(app) and K,~(app) should be decreased as the flow rate increases. Various types of inhibition behavior have been observed with a-chymotrypsin depending on the substrate used and the rate determining step for that substrate (Laidler & Bunting, 1973). However, Gellf et al. reported that there were neither inhibitory nor activation effects.
For substrate inhibition and product inhibition" kinetics, the results of Kobayashi etal . (1973) and Hirano etal . (1977) were found to be consistent with the result of our theoretical analysis.
In the case of two substrate enzyme reaction kinetics, the results of Wilson et al. (1968 a, b), Cho et al. (1974) and Paul et al. (1978) all agree with the results of our theoretical analysis. Paul et al. reported that the intrinsic Michaelis constant of N A D H was 0.035 mM and that of fructose was 0.10 M for sorbitol dehydrogenase system. From these values, one finds that the ratio of affinity, K, is about 3.5 x 10 -4 or nearly zero. When the relative affinity is significantly different,~namely K = 0, both a* and/3* increase as tt decreases, a * is constant, and/31.* decreases as/z decreases (Fig. 3). These results are consistent with the experimental results reported. The rate parameters calculated using equations (27)-(30) are also in good agreement with the experimental data as shown in Table 4 (see Appendix A).
TABLE 4
Comparison o f experimental data with theoretical values
Agitation speed V~tNADH) 2 V~(Fr) 2 KNADll K~-r (rpm) (n mol/min cm ) (n mol/min cm ) (mM) (M)
570 experimental - 7 0-11 0.03 calculated 22-4 7.2 0-110 0-034
13 experimental - 4 0.17 0.02 calculated 22.4 4.5 0.168 0.020
Experimental data m:e from Paul et aL (1978) for sorbitol dehydrogenase system and the theoretical values are calculated using equations (27)-(30). Detailed calculations are also shown in the Appendix A.
Our analytical solutions can be use'd to predict the apparent kinetic constants when the intrinsic kinetic constants are known.

274 S. B. LEE AND D. D. Y. RYU
REFERENCES
CHO, I. C. & SWAISGOOD, H. (1974). Biochem. Biophys, Acta 334, 243. ENGASSER, J-M. & HORVATH, C. (1973). J. theor. Biol. 42, 137. GELLF, G., THOMAS, D., BROUN, G. & KERNEVEZ, J. F. (1974). Biotech. Bioengng. 16, 315. GOLDMAN, R., KEDEM, O. & KATCHALSKI, E. (1971). Biochemistry 10, 165. GOLDSTEIN, L. (1976). In Methods in Enzymology, Vol. 44, (K. Mosbach, ed.). New York &
London: Academic Press. HAMILTON, B. K., S T O C K M E Y E R , L. J . & C O L T O N , C . K. (1973), J. theor. Biol., 41, 547. HIRANO, K., KARUBE, I., MATSUNAGA, T. & SUZUKI, S. (1977). J. Ferment. TechnoL, 55,
401. HORNBY, W. E., LILLY, M. D. & CROOK, E. M. (1968). Biochem. J. 107, 669. HORVATH, C. & ENGASSER, J-M. (1974). Biotech. Bioengng. 16, 909. KIM, S. M., LEE, S. B. & RYU, D. D. Y. (1978). In Enzyme Engineering Vol. 4, (G. B. Broun,
G. Manecke & L. B. Wingard, eds), p. 101. New York: Plenum. KOBAYASHI, T. & Moo-YOUNG, M. (1971). Biotech. Bioengng. 13, 893. KOBAYASHI, T. & Moo-YoUNG, M. (1973). Biotech. Bioengng., 15, 47. KOBAYASIII, T. & LAIDLER, K. J. (1974). Biotech. Bioengng. 16, 77. LAIDLER, K. J. & BUNTING, P. S. (1973). The ChemicaIKinetics o[ Enzyme Action, 2nd edn.
Oxford: Clarendon Press. LEE, S. B., KIM, S. M. & RYU, D. D. Y. (1979). Biotech. Bioengng. 21, 2023. LEE, S. B. & RYU, D. D. Y. (1979). Biotech. Bioengng. 21, 1499. LEE, Y. Y., FRA'rzE, A. R., WEN, K. & TSAO, G. T. (1976). Biotech. Bioengng., 18, 389. LILLY, M. D., HORNBY, W. E. & CROOK, E. M. (1966). Biochem. J. 100, 718. Moo-YOUNG, M. & KO~YASHI, T. (1972). Can. J. Chem. Eng. 50, 162. PAUL, F., COULET, P. R., GAUTHERON, D: C. & ENGASSER, J-M. (1978). Biotech.
Bioengng. 20, 1785. SHULER, M. L., ARIS, R. & TSffCtIIYA, H. M. (1972). J. theor. Biol. 35, 67. 8UNDARAM, P. V., PYE, E. K., KATCHALSKY, E., LEVIN, Y., LILLY, M. D., ~,IANECKE, G.,
MOSBACiI, K., PATCHORNIK, A., PORATH, J., WEETALL, H. H. & WINC3ARD, L. B. Jr. (1972). In Enzyme Engineering, Vol. 2, (L. B. Wingard, Jr. ed). p. 15. New York: Wiley-Interscience.
TODA, K. (1975), Biotech. Bioengng. 17, 1729. TOSA, T., MORI, T. & CHIBATA, I. (1971), J. Ferment. Technol., 49, 522. WILSON, R. J. H., KAY, G. & LILLY, M. D. (1968a), Biochem. J., 108, 845. WILSON, R. J. H., KAY, G. & LILLY, M. D. (1968b), Biochem. 3". 109, 137.
APPENDIX A
The Calculation oi Rate Parameters in Table 4
From the data of Paul et al. (1978), the intrinsic rate parameters for sorbitol dehydrogenase are: V " = 28 n mol/min cm2; K~-, 0.1 M; ' = KNADH = 0.035 mM and the mass transfer coefficients for N A D H are: kNADH =
2 " 5 x l 0 - 3 c m / s at 570rpm; k N A D H = I ' 4 x l 0 - 3 c m / s at 13rpm. They measured activity at constant fructose concentration of 0.4 M and at constant N A D H concentration of 0.06 raM. Using our dimensionless
I I I parameters, m = Si/K'~i = 4.0; o- i = Si/Kmj = 1.71; tz i = V'~K,,ikLi = 5.34 at 570 rpm; and tz i = 9.53 at 13 rpm. In this case i and/" represent the fructose and N A D H , respectively, since i denotes the one of two species of substrate having less affinity and / the other having more affinity.

D I F F U S I O N E F F E C T S O N E N Z Y M E K I N E T I C S 275
Since the ratio of affinity is approximately equal to x, K - ~ K ' j / K ' ~ =
3.5 • 10-a or nearly zero. When i< = 0, equations (27)-(30) reduces to
,~" = o-l/(1 + o ' 3
3~ = 1 +0"5 o~'/z i
a* = 2o'j{(1 + tz i + o'j) + [(1 + tzi + o'j) 2 - 4tz,o'j] 1/2}-1
3,* = 1 - 2 ( 1 - o,*)2uj/[(1- o,*)(2- ~')~j + 2].
(A1)
(A2)
(A3)
(A4)
Using equations (A1)-(A4) one finds that a} ~ = 0.8; 3* = 3-14; oe* = 0.26; /3~*=0.34 at 570 rpm; and a ~ = 0 . 8 ; 3 ~ ' = 4 . 8 1 ; a * = 0 . 1 6 ; 3 * = 0.20 at 13 rpm. Hence we have V,~(r~hDm" ---- 0"8 V,,' = 22"4 n mol / nain cm2; KI~ADH = 3" 14 KNADH 0" 11 mM; " -- ' ' = V[~(F,) -- 0"26 Vm = 7"2 n rnol /mincm2;K~,=0"34K~- , = 0 . 0 3 4 M at 570rpm, and similarly/ Vr~(NADH) = 22"4 n mol /min 2 .... " cm ; ~NAtn, = 0" 17 mM; V~(F,) = 4-5 n mol/. nain cm2; K~, = 0.020 M at 13 rpm.
1, APPENDIX B
So far, several workers have derived the equations for apparent kinetic constants using Michaelis-Menten kinetics (Hornby et a l . , 1968; Kobayashi & Moo-Young, 1971; Shuler et al . , 1972; Hamilton et a l . , 1973; Kobayashi & Laidler, 1974), and they all reported that the apparent maximum reaction rate would not be affected by the diffusional limitations and remain constant. Here, we will show that the apparent rate parameters can be, in fact, affected by the diffusional limitations, when the apparent kinetic constants are determined by the conventional graphical method.
If we introduce the following dimensionless variables,
a = V , , 3 = K , , and T = ~ ,
equations (9) and (10) become
and
O"
~" = ( l / y ) + o" (B1)
O~O-
~" = 3 +---~" (B2)

2 7 6 s . B . LEE AND D. D. Y. RYU
From equations (B1) and (B2), one finds that
a = 1 (B3)
fl = 1 = ~{(1 +/z + o') + [(1 +/z + o')2-4#o']1/2}- o" (B4) ,y
since 3' can be expressed as [from equation (12)]
r = 1 - Oz/o"
1 = 1 -~--~ {(1 +/x + t r ) - [ ( 1 +/.t + o')2-4t.to']'/2}. (BS)
The two limiting cases for the solution in equation (B4) are"
t ~ = l , f l ~ l + l z a s t r ~ 0 (B6)
a = 1, fl ~ 1 as tr ~ ~ . (B7)
Now, we see that the equation derived by Hornby et al. (1968) is identical to equation (B6) and thp results of Shuler et al. (1972) and Hamilton et al. (1973) are approximate solutions of equation (B4) for special cases when
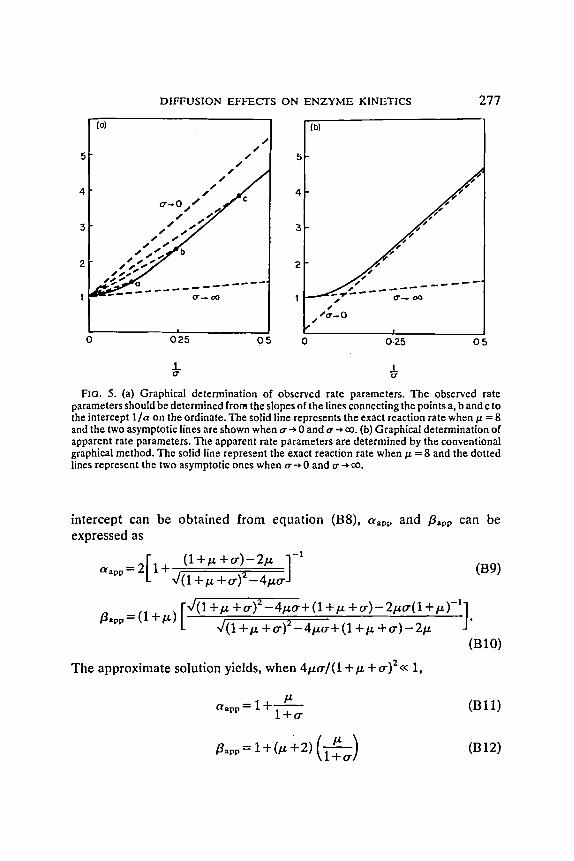
<< 1 + tr and/.t >> 1 + o', respectively. Two lines corresponding to equations (B6) and (B7) are shown in Fig. 5(a)
in the plot of 1/~" vs. 1/tr. In the first order region (tr ~ 0), however, the line is only parallel to its true one (solid line). This implies that fl should be determined from the individual slope of the lines connecting the points a, b, or c to the intercept 1/a on the ordinate, when and if the theoretical equations derived by previous workers are to be used as shown in Fig. 5(a). The apparent Michaelis constant, determined by the method described here will be highly dependent on the substrate concentrations.
Conventionally, the apparent rate parameters a/e determined from the slope of the line passing through the data points a, b, and c and from the intercept on the ordinate which can be obtained by the extrapolation of the line [line corresponds to o-~ 0 in Fig. 5(b)]. In order to determine the apparent values by the conventional method the slope and the intercept must be determined from the Lineweaver-Burk expression of equation (B2)
1 s (Bs)
The slope and the intercept are equal to f l /a and I /a , respectively. Since the slope can be obtained by differentiating 1/~" with respect to 1/o- and the
DIFFUSION EFFECTS ON E N Z Y M E KINETICS 277 (o)
/ /
/
/ /
c r ~ O / , / / c
J S ~'
0 0~25 0 5
(b)
s ~7
�9 "a'- 0 / " , ]
0 0-25 0 5
_1_ .1_ O" o"
FIG. 5. (a) Graphical determination of observed rate parameters. The observed rate parameters should be determined from the slopes of the lines connecting the points a, b and c to the intercept 1/a on the ordinate. The solid line represents the exact reaction rate when/1 = 8 and the two asymptotic lines are shown when tr ~ 0 and tr --~ co. (b) Graphical determination of apparent rate parameters. The apparent rate parameters are determined by the conventional graphical method. The solid line represent the exact reaction rate when/.t = 8 and the dotted lines represent the two asymptotic ones when o--* 0 and tr ~ co.
intercept can be expressed as
211+ (l+At+tr)-2/z ]-'
r,/(1 + . + 0 - ) 2 - 4 / / . 0 - + (1 +At + o ' ) - 2Ato'(1 + At)-'] (1 + A t ) [
obtained from equation (B8), aapp and /3~pp can be
The approximate solution yields, when 4Attr/(1 +At +0-)2<< 1,
( B 9 )
(BIO)
At (BII) a~pp=lq l + g
(B12)

278 s . B . L E E A N D D. D. Y. R Y U
and for the two limiting cases of the solution (Bl l ) and (B12),
aapp~ (1 +~) , /3app~ (1 +~ ) 2 as tr--~0 (B13)
otapp ~ 1, fla~p~ 1 as o--* oo. (B14)
The two asymptotic lines corresponding to equations (B13) and (B14) are shown in Fig. 5(b). In the first order region, the slope of the Lineweaver- Burk plot, fl/a, is the same as that of previous analysis [equation (B6)] but the intercept is altered. It should be noted when the apparent values are determined by the conventional graphical method the apparent maximum reaction rate can be varied with the change of mass transfer rate although the variations in the apparent maximum reaction rate are not so significant as these in the apparent Michaelis constant. The values of 1/ct could vary within the range of 1.0 and 1/( l+~t) depending on the mass transfer modulus,/x.
The equations for apparent kinetic constants derived by previous workers should be improved or modified if the apparent rate parameters are to be determined by the conventional graphical method. Equations (B11)-(B 14) are recommended as good approximate expressions to be used for deter- mination of apparent kinetic constants.
D J kt., kLs, kLp,} kL~, kLi
K~s K7
K" K~,(app)
Ks Porp P R S o r s g
Nomenclature
ditlusion coefficient (cm2/s) mass flux (mol/cm2-s)
mass transfer coefficient (s -~)
intrinsic product inhibition constant (mol/1) observed product inhibition constant (mol/1) intrinsic substrate inhibition constant (mol/1) observed substrate inhibition constant (mol/1) intrinsic Michaelis constant (mol/1) observed Michaelis constant (mol/1) apparent Michaelis constant (mol/1) K'/K~p, relative product inhibition constant (dimensionless) K',,/K~s, relative substrate inhibition constant (dimensionless) product concentration in bulk solution (mol/1) product concentration at the surface reaction site (mol/1) reaction rate in heterogeneous enzyme reaction (mol/cm2-s) substrate concentration in bulk solution (mol/1) substrate concentration at the surface reaction site (tool/l)

v" v" V~(app)
X
Y t /
t~ /3" y 6
K
/z b'
Or
D I F F U S I O N E F F E C T S O N E N Z Y M E K I N E T I C S 2 7 9
intrinsic maximum reaction rate (mol/cm2-s) observed maximum reaction rate (mol/cm2-s) apparent maximum reaction rate (mol/cm2-s)
1 [x/(1 + ~:)2 + 4Kpp.- (1 + •)] (dimensionless) 4Kp~
axis, or distance in linear dimension (cm) V~] V', relative maximum reaction rate (dimensionless) dimensionless maximum reaction rate, defined by equation (7) K".,]K', relative Michaelis-Menten constant (dimensionless) dimensionless Michaelis constant, defined by equations (8) S]S, relative substrate concentration (dimensionless) thickness of diffusion boundary layer (cm) R] V', reaction rate (dimensionless) ~i/tz s, relative affinity of two substrates (dimensionless) V'/K'kL, mass transfer modulus (dimensionless) P/P, relative product concentration (dimensionless) P/K~p, initial product concentration (dimensionless) S/K',,,, substrate concentration (dimensionless)