factor b and the mitochondrial atp synthase complexa,b grigory i. belogrudov and
TRANSCRIPT

FACTOR B AND THE MITOCHONDRIAL ATP SYNTHASE COMPLEXa,b
Grigory I. Belogrudov and Youssef Hatefic
Division of BiochemistryDepartment of Molecular and Experimental Medicine
The Scripps Research InstituteLa Jolla, California 92037
Running Title: Factor B
aSupported by the United State Public Health Service Grant DK08126
bThis is publication number 14385-MEM from The Scripps Research Institute
cTo whom correspondence should be addressed.Tel: 858-784-8092; Fax: 858-784-2054; E-mail: [email protected]
Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on December 14, 2001 as Manuscript M111256200 by guest on A
pril 12, 2019http://w
ww
.jbc.org/D
ownloaded from

2
ABSTRACT
Factor B is a subunit of the mammalian ATP synthase complex, whose existence has been
controversial. This paper describes the molecular and functional properties of a recombinant human
factor B, which when added to bovine submitochondrial particles depleted of their factor B restores
the energy coupling activity of the ATP synthase complexes. The mature human factor B has 175
amino acids and a molecular mass of 20,341 Da. The preparation is water-soluble, monomeric, and
is inactivated by monothiol and especially dithiol modifying reagents, probably reacting at its
cysteine residues C-92 and C-94. A likely factor B gene composed of 5 exons has been identified on
chromosome 14q21.3, and the functional role of factor B in the mammalian ATP synthase complex
has been discussed.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3
It is generally considered that the mammalian mitochondrial ATP synthase complex is
composed of 16 unlike subunits (1-3). These subunits are α, β, γ, δ and ε in the catalytic F1 domain;
OSCP, a, b, c, d, e, f, g, F6 and A6L in FO and stator; and the ATPase inhibitor protein, IF1, which
binds reversibly to F1 to inhibit ATP hydrolysis. In 1967, Sanadi and coworkers (4) showed that
submitochondrial particles prepared by sonication from bovine mitochondria suspended in 0.25 M
sucrose and 0.6 mM EDTA1 and adjusted to pH 9.0 with ammonium hydroxide lost considerable
activity for respiration-driven ATP synthesis and ATP hydrolysis-driven electron transfer from
succinate to NAD. Addition to the ammonia-EDTA treated particles (AE-SMP) of a partially
purified soluble protein extracted from mitochondrial acetone powders partially restored these
activities (4,5). Sucrose density-gradient centrifugation suggested a molecular mass of 32 kDa for
the active peak of the soluble preparation, which was designated factor B (4,5). Using bovine
mitochondrial acetone powder extracts, we isolated a pure and monodisperse protein, which restored
ATP synthase-coupled activities to AE-SMP (6). Its molecular mass as estimated from
sedimentation equilibrium and gel filtration experiments was 11-12 kDa, and it was
immunoprecipitated by Sanadi’s anti-factor B antiserum in an Ouchterlony double diffusion
experiment (7). Sanadi and coworkers revised the molecular mass of their preparation to 29.2 kDa
and suggested that it is a dimer of monomer molecular mass 14.6 kDa (5,8). They also obtained
preparations of relative molecular mass 13 – 15 and 47 kDa, which exhibited factor B-like activity
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

4
(5,9,10), and in 1990 published the sequence of the 55 amino terminal amino acids of a factor B
preparation with a relative molecular mass of 22 kDa (11).
The existence of factor B as a component of the ATP synthase complex remained
controversial, however. Whereas Sanadi claimed that the ATP synthase complex prepared in his
laboratory contained factor B (5,12), a thorough analysis of the polypeptide composition of an
ATPase complex prepared in Walker’s laboratory demonstrated the existence of the 16 unlike
polypeptides mentioned above, but no factor B (1,2). The multiplicity of Sanadi’s factor B-like
preparations with relative molecular masses ranging from 13 – 15 to 47 kDa, the small yield of our
preparation, which precluded antibody production and chemical analyses, and the compelling
extensive data of Walker’s laboratory discouraged further interest in the pursuit of factor B.
More recently, two nucleotide sequences corresponding in part to Sanadi’s 55 amino acid
sequence of factor B amino terminus were detected by our colleagues in the human genome. The
shorter frame corresponding to 96 amino acid residues was expressed with a histidine tag. The
expressed protein was found in inclusion bodies, and when extracted with 8 M urea and dialyzed
exhibited no factor B-like activity. The longer frame corresponding to 175 amino acid residues was
expressed once with a histidine tag and a second time fused to thioredoxin. Both were recognized
after purification by polyclonal antibodies raised to the shorter polypeptide. The histidine-tagged
preparation was also inactive, but the other, after removal of thioredoxin, exhibited all the functional
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

5
features previously described by Sanadi and coworkers (4,5) and by ourselves (6,7) for factor B.
This paper describes the molecular and functional properties of this human factor B preparation.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

6
MATERIALS AND METHODS
Materials – NAD, NADH, ATP, oligomycin, and 2,4-dithiothreitol were obtained from
Calbiochem; DCCD, venturicidin, FCCP, tributyltin chloride, NEM, pCMB, sodium succinate, and 2-
mercaptoethanol were from Sigma; DEAE-Sepharose was from Amersham Pharmacia Biotech;
polyacrylamide was from Bio-Rad; Tris was from ICN; and oxonol VI was from Molecular Probes. All
other chemicals were reagent grade. AE-SMP were prepared from heavy beef heart mitochondria (13)
according to (14). After a final wash with 0.25 M sucrose containing 10 mM Tris-HCl, pH 7.8, the
particles were suspended in the same medium at 17 mg of protein/ml and stored at –80°C in small
aliquots. F1-ATPase was prepared according to Senior and Brooks (15) and stored in 50% saturated
ammonium sulfate at 4°C. The catalytic activity of the enzyme was in the range of 90-100 µmol ATP
hydrolyzed (min•mg)-1. OSCP was prepared by Dr. A. Matsuno-Yagi as before (16).
Assays – ATP-driven reverse electron transfer from succinate to NAD was assayed at 38° C
essentially according to Joshi and Sanadi (14), except that after 1-2 min of incubation of AE-SMP with
factor B, succinate, ATP, MgCl2, KCN, and dithiothreitol, the reaction was started by the addition of
NAD. The progress of the reaction was monitored at 340 nm in a Milton Roy 1201 spectrophotometer,
and the rate of NAD reduction was calculated using an extinction coefficient of 6.22 mM-1. Inactivation
of factor B with thiol modifying reagents was performed by incubating factor B for 5 min on ice in 50 mM
Tris-sulfate, pH 7.8, with increasing concentration of CdSO4 dissolved in water, or of phenylarsine oxide,
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

7
pCMB or NEM dissolved in dimethylformamide. The incubation mixtures containing CdSO4 and
phenylarsine oxide also contained 0.25 mM 2-mercaptoethanol. The inhibitor treated samples of factor B
were assayed for reconstitution of the ATP-driven reverse electron transfer activity of AE-SMP in the
absence of dithiothreitol. The ATPase activities of F1 and AE-SMP were measured
spectrophotometrically by the coupled pyruvate kinase/lactate dehydrogenase method as before (17).
SDS-polyacrylamide gel electrophoresis and immunoblotting were performed as described
elsewhere (18). The immune complexes were visualized using the SuperSignal West Pico
chemiluminescence detection kit from Pierce. Protein concentration was determined, using the BCA kit
from Pierce. Standard molecular biology techniques, including cloning, restriction enzyme analysis,
plasmid preparation, and agarose gel electrophoresis were as described in (19). Amino terminal analysis
of purified recombinant human factor B was performed at the Protein and Nucleic Acid Core Facility of
this institution. Tryptic peptide mass fingerprinting and matrix-assisted laser desorption/ionization time-
of-flight (MALDI-TOF) mass spectrometry were performed at the Scripps Center for Mass Spectrometry.
Preparation of recombinant human factor B – The I.M.A.G.E. clone 2663774 containing EST
AW173068 was purchased from Research Genetics, AL. The sequence of the insert was confirmed by
DNA sequence analysis. The forward primer P1: 5’-TTCTGGGGCTGGTTGAATGCA-3’ and reverse
primers P2: 5’-TTACTTCAATTGTAATTTTAGTTC-3’ and P3: 5’-
CTTCAATTGTAATTTTAGTTCCAG-3’, with and without termination codon, respectively, were
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

8
designed to amplify the full length of mature factor B by PCR using Taq DNA polymerase (Qiagen). The
amplicons were subcloned into pBAD/Thio-TOPO plasmid (Invitrogen) according to manufacturer’s
recommendations, and clones containing plasmids, designated pBAD-FB1 and pBAD-FB2, with the
correct sequence of the inserts were selected after DNA sequencing. In both pBAD-FB plasmids, the
factor B nucleotide sequence was located downstream of the nucleotide sequence coding for His-Patch
thioredoxin under regulation of araBAD promoter.
In the present study, a single colony of TOP10 cells transformed with pBAD-FB1 plasmid from a
fresh LB plate containing 100 µg of ampicillin/ml was used to inoculate 5 ml of 2 x YT liquid medium in
the presence of 100 µg of ampicillin/ml and grown overnight in a shaker at 37°C. Then, 0.5 liter of the 2
x YT liquid medium plus 100 µg of ampicillin/ml were placed in a 2 liter flask and inoculated with 3 ml
of the overnight culture and grown in a shaker (250 rpm) at 37°C until OD600 reached 0.5-0.6. The
temperature in the shaker was shifted to 30°C, and the protein expression was induced by adding 20%
arabinose to 0.02% final concentration. The incubation was continued for 4 h, and the culture was
harvested by centrifugation and stored at –80°C until further processing.
The bacterial cells were disrupted by 4 cycles of freeze-thawing, followed by sonication on ice
slurry, using a Branson sonifier. The soluble fraction was recovered by centrifugation for 1 hr at 15,000
rpm in a Beckman SS-34 rotor, and affinity-purified on His-Bind resin (Novagen) preloaded with NiCl2.
After wash with a pH 8.0 buffer containing 50 mM sodium phosphate, 0.3 M NaCl, and 20 mM imidazole,
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

9
the fusion protein was eluted with 50 mM imidazole in the above buffer. The fractions containing the
fusion protein were concentrated using the Centriprep YM-10 centrifugal filter devise (Millipore) and
subjected to size-seiving chromatography on Ultragel AcA 44 (column size 1.5 cm x 90 cm) equilibrated
in a pH 8.0 buffer containing 50 mM Tris-HCl, 0.1 M NaCl, and 5 mM 2-mercaptoethanol.
To remove the thioredoxin moiety, the recombinant protein was cleaved overnight at room
temperature with recombinant enterokinase (Novagen) and purified by chromatography on DEAE-
Sepharose Fast Flow, using a pH 8.0 buffer containing 50 mM Tris-HCl and 5 mM 2-mercaptoethanol.
The fractions were assayed for restoration of ATP-driven reverse electron transfer from succinate to NAD
as catalyzed by AE-SMP and combined based on activity and purity after analysis by SDS-PAGE. The
protein was concentrated on YM-10 and stored in small aliquots at –80°C.
Preparation of cDNA for the ORF of human factor B – To obtain a cDNA encoding the full-
length ORF of human factor B, the oligonucleotide primers P1 and P2 (see above) as well as the
forward primer P4: 5’-CCTGCTCTGTACTCGACCCG-3’ and the reverse primer P5: 5’-
AAGAGTCGGTGGCGTCGAT-3’ were used to amplify by PCR the nucleotide sequences encoded
by inserts in I.M.A.G.E. clones 2663774 (GenBank accession number AW173068) and 24431
(GenBank accession number AF052186) obtained from Research Genetics, AL. The PCR
amplifications were performed in 50 µl volumes, each containing 5 µl of High Fidelity PCR buffer,
2 mM MgSO4, 0.2 mM dNTPs, 0.2 µM of each primer, ~10 ng of template, and 1 unit of
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

10
PLATINUM Taq High Fidelity DNA polymerase (Life Technologies). The PCR conditions used
were: initial denaturation at 94°C for 2 min followed by 30 cycles each of 94°C for 30 s, 55°C for 1
min, 72°C for 1 min, and a final extension at 72°C for 7 min. In a second round, 5 µl from each of
the PCR reaction mixtures were combined and subjected to amplification with primers P2 and P4 as
above. The final PCR product was purified on agarose gel and subcloned into the pDrive vector
(Qiagen). The nucleotide sequence of the insert was verified in both directions by ABI 327
sequencer using BigDye terminator sequencing.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

11
RESULTS
Molecular properties of factor B – The predicted amino acid sequence of human factor B is
shown in Fig. 1. In the amino terminal 55 residues, there are three differences between the mature
human factor B shown in Fig. 1 and the sequence published by Sanadi and coworkers for the bovine
protein. The human amino acid residues Y-14, C-33 and E-43 are, respectively, H, G and Q in the
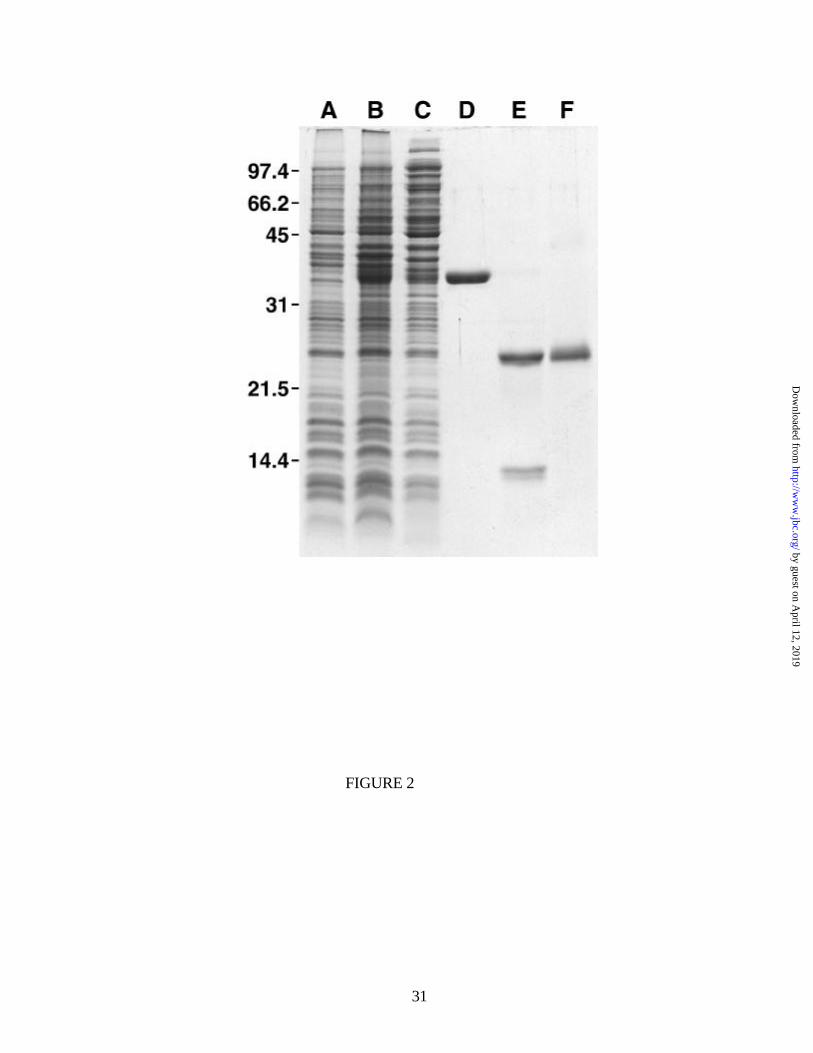
Sanadi partial sequence (11)2. Figure 2 shows on SDS polyacrylamide gel the polypeptide pattern of
Escherichia coli total cell lysate before (A) and after (B) induction of fused thioredoxin-factor B
synthesis. Lane C is the pattern of the soluble proteins of the induced cells, and lane D is the
purified thioredoxin-factor B fused preparation. Lane E shows two protein bands due to factor B
(upper) and thioredoxin (lower) after treatment of the fused protein with enterokinase, and lane F
shows a single protein band due to purified factor B. As determined by amino terminal sequence
analysis, this preparation of factor B contained at its amino terminus an extra tripeptide, L-A-L,
which had been retained after cleavage of thioredoxin. Otherwise, mass spectrometric analysis of
tryptic peptide masses of our recombinant factor B matched the calculated masses of the
corresponding peptides expected from its tryptic degradation (data not shown). Thus, the mature
factor B shown in Fig. 1 is composed of 175 amino acids with a calculated molecular mass of 20,
341 Da. The molecular mass derived from MALDI-TOF mass spectrometry of the factor B
preparation containing the amino terminal L-A-L tripeptide was 20,607 Da (calculated value: 20,638
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

12
Da), and gel exclusion suggested a mass of ~ 22 kDa, which indicated that the recombinant human
factor B is monomeric (data not shown). Hydropathy analysis of the human factor B sequence
revealed no hydrophobic clusters of amino acids for possible membrane anchorage.
Functional properties of factor B – The term factor B has been applied to several protein
preparations derived from bovine heart mitochondria with molecular masses of 11 – 12 to 47 kDa,
which partially or largely restored to AE-SMP preparations ATP synthesis and energy-linked
reactions driven by ATP hydrolysis. It was, therefore, assumed that extraction of mitochondria with
ammonia-EDTA partially removed one or more factor B-like molecules from the mitochondrial ATP
synthase complexes. However, the effectiveness and the specificity of this extraction procedure for
factor B removal were unclear. Figure 3 depicts immunoblots of mitochondria (column A), SMP
(column B) and AE-SMP (column C) showing their relative contents of factor B (row 1), F1α
subunit (row 2), OSCP (row 3) and IF1 (row 4). The results clearly show that ammonia-EDTA
extraction is highly effective for the efficient removal of factor B3. The levels of F1 as represented
by its α subunit, and OSCP, which is not anchored to the membrane, were essentially unchanged by
the extraction. The level of IF1 was decreased in AE-SMP (Fig. 3, bottom row) which was
understandable, because of the alkaline pH of the extraction medium (24). Consistent with these
results, the ATPase activity of AE-SMP was shown to be about 40% higher than that of SMP.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

13
Figure 4 shows on the ordinate the activity of AE-SMP for ATP hydrolysis-driven electron
transfer from succinate to NAD as affected by addition of increasing amounts of recombinant human
factor B. The activity of AE-SMP in the absence of factor B was 4 nmol NADH formed (min • mg
protein)-1. Addition of factor B increased this activity to 225, which was 87% of the activity of the
SMP before ammonia-EDTA extraction. The abscissa intercept of the double reciprocal plot of
these data in the inset indicated half-maximal activation of AE-SMP at 0.33 nmol of factor B/mg of
AE-SMP. Assuming that AE-SMP have roughly the same amount of ATP synthase as SMP, i.e., 0.4
– 0.45 nmol/mg of protein (25), the factor B half-saturation amount of 0.33 nmol/mg of AE-SMP
indicated that this water-soluble recombinant human protein bound specifically and with high
affinity to the site from which its bovine counterpart had been removed. Addition to AE-SMP of F1
plus OSCP, instead of factor B, resulted in a marginal increase of reverse electron transfer activity
(10-12% as compared to 87% when factor B was added), indicating in agreement with the data of
Fig. 3 that the poor coupling activity of AE-SMP was not due to the loss of F1 and/or OSCP.
It has long been known that addition of low concentrations of oligomycin to SMP treated
with EDTA or ammonia-EDTA partially restores their energy-coupled activities. Apparently, when
added at low concentrations oligomycin preferentially reacts with and seals the proton leakiness of
the fraction of the ATP synthase complexes that have been rendered defective, thereby allowing the
remaining intact ATP synthases to function normally (26). Further addition of oligomycin would
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

14
then begin to inhibit the intact ATP synthases, resulting in a decrease of the ATP synthase-coupled
activity tested. Similar results are shown in Fig. 5. It is seen that addition to AE-SMP of oligomycin
up to about 150 ng (0.3 nmol) /mg protein increased the ATP-driven reverse electron transfer
activity of the particles from about 5 to about 54 nmol NADH formed (min • mg protein)-1, and that
further increase in oligomycin concentration resulted in inhibition. As was shown previously (6),
addition of low concentrations of oligomycin (<120 ng/mg protein) to factor B-replenished AE-SMP
caused no activation, but only partial inhibition of ATP-driven reverse electron transfer activity. We
have examined the effect of other FO inhibitors with regard to activation of AE-SMP for ATP driven
electron transfer from succinate to NAD. At concentrations that they caused inhibition of ATPase
activity of AE-SMP from 10 to 70%, venturicidin and tributyltin chloride had no stimulating effect
on its reverse electron transfer activity. Incubation of AE-SMP at 0°C with 6 µM DCCD caused a
time-dependent inhibition of its ATPase activity , as expected. At 10 and 20 min of incubation,
when ATPase activity was inhibited by 30% and 43% respectively, ATP-driven reverse electron
transfer activity of the particles was increased from 4 nmol NADH formed (min • mg)-1 to 18 at 10
min and to 27 at 20 min of incubation. Further incubation of AE-SMP with 6 µM DCCD lowered
this stimulated activity to 22 at 30 min and to 7 at 50 min.
Previous preparations of bovine factor B were shown to be inhibited by thiol and dithiol
modifiers (5-7). The effects of these modifiers on recombinant human factor B are shown in Fig. 6.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

15
It is seen that factor B was inhibited when treated with the compounds shown, with the dithiol
modifiers phenylarsine oxide and Cd2+ causing 50% inhibition at < 10 µM concentration. As seen in
Fig. 1, human factor B contains 6 cysteine residues, two of which are located at positions 92 and 94.
The strong inhibitory effects of phenylarsine oxide and Cd2+ suggest that these dithiol modifiers bind
to cysteine residues 92 and 94 of factor B.
Effect of factor B on membrane potential – Figure 7 shows the formation of a membrane
potential in SMP (traces A and B), AE-SMP (traces C and D) and AE-SMP plus factor B (traces E
and F) as monitored by the absorbance change of oxonol VI at 630 minus 603 nm. Traces A and B
are controls with SMP, showing membrane energization upon addition of ATP or NADH, and
deenergization by inhibition of ATP hydrolysis by oligomycin, inhibition of NADH oxidation by
rotenone, or by uncoupling with FCCP. Traces C and D show that AE-SMP could not develop a
high membrane potential upon addition of either ATP or NADH, but that addition of oligomycin
repaired this defect and allowed high membrane potential formation upon subsequent addition of
NADH. Similar results were obtained when oligomycin was replaced with venturicidin or tributyltin
chloride, or when the AE-SMP were treated for 60 min at 0°C with 10 µM DCCD. Like oligomycin,
these reagents inhibited ATP hydrolysis, but allowed membrane potential formation, as in Fig. 7C,
upon addition of NADH (data not shown). These results suggested, therefore, that removal of factor
B created a proton leak in the FO of AE-SMP, which could be blocked by the specific FO inhibitors
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

16
mentioned. Traces E and F show that the addition of factor B to AE-SMP repaired the defect shown
in traces C and D, and made it possible for the particles to develop a high membrane potential upon
addition of either ATP or NADH. As seen in traces E and F, the development of the membrane
potential to a static head level in factor B supplemented AE-SMP was a slow process as compared to
the results shown in traces A and B for SMP. It is possible that factor B binding to AE-SMP
requires an energized conformation of FO-F1, apparently best achieved with the addition of ATP.
This possibility would be somewhat analogous to the binding of IF1 to F1, which occurs during ATP
hydrolysis by the enzyme (24).
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

17
DISCUSSION
Molecular properties of factor B – According to the data reported here, mature human factor
B is composed of 175 amino acid residues with a molecular mass of 20,341 Da and the amino acid
sequence shown in Fig. 1. The active, recombinant human factor B purified and used in the studies
reported here contained an extra L-A-L tripeptide at its amino terminus. It was water-soluble,
monomeric, and stable when stored at –80°C in a pH 8.0 buffer containing 5 mM 2-
mercaptoethanol. In its amino terminal 55 residues and molecular mass, this recombinant human
factor B is similar to the latest of a number of bovine mitochondrial factor B preparations reported
previously by Sanadi and coworkers (11). Because rigorous data on the purity and the amino
terminal sequences of the other factor B-like preparations of Sanadi and coworkers are not available,
it is not possible to discuss them further. Nor can we compare our pure protein of molecular mass
11-12 kDa, which was isolated from bovine heart mitochondria and exhibited a factor B-like activity
(6,7), with the recombinant factor B described here. Its low yield precluded in 1976 any attempts at
amino terminal sequencing, but its sensitivity to thiol and dithiol modifiers suggests that it might
have been an active proteolytic fragment of the bovine factor B. This possibility is supported by the
fact that immunoblots of bovine heart mitochondria with our polyclonal antibody used in the present
work recognized only a single protein band corresponding in mobility on SDS-gels to that of our
recombinant human factor B.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

18
Factor B gene – Figure 8 shows a segment of the human genome on chromosome 14q21.3.
This segment spans ~13 kb and contains 5 exons, of which the nucleotide sequences of the areas
depicted in Fig. 8 by the filled boxes from ATG in exon 1 to TAA in exon 5 correspond to the
sequence of the human factor B cDNA shown in Fig. 1, except that in the draft sequence of the
Human Genome Project (27) T168 is given as C, which would result in the substitution of Pro for Leu
in the corresponding position of the presequence of factor B. As seen in Fig. 8, there are 2 stop
codons and 2 polyadenylation signals in this DNA segment. An mRNA transcript arising from the
processing of pre-mRNA at poly (A) site ATTAAA would encode a hypothetical protein HSU79253
(LocusID 27109). Reverse transcription PCR, using multiple tissue cDNA panels I and II
(Clontech), showed that mRNAs corresponding to both the short (exons 1-3) and the long (exons 1-
5) sequences were present in 16 different human tissues that we examined. As mentioned above, the
short polypeptide containing a His tag was expressed in our laboratory as inclusion bodies and used
to raise polyclonal antibodies. This antibody preparation recognized in bovine heart mitochondria a
single protein with Mr of 22 kDa, which could be extracted from SMP by the procedures used to
remove factor B (Fig. 3). The short polypeptide, expected to have an Mr ~11 kDa , was not detected
in bovine heart mitochondria. This consideration suggests that this short “isoform” of factor B, if
translated, either does not enter mitochondria or is degraded after entering this organelle.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

19
A second human homologue, the hypothetical protein FLJ10241 (LocusID 55101) encoded
by a gene located on chromosome 19q13.2, with sequence similarity of 49% to human factor B was
identified by BLASTP (28) search of the GenBank database. This search also revealed the existence
of two mouse orthologues (GenBank Accession Nos. NP_080812 and BAB26107) with sequence
similarities of 83 and 47%, respectively, as well as two gene products in Drosophila melanogaster
(GenBank Accession Nos. AAF58055 and AAF51634) and Caenorhabditis elegans (GenBank
Accession Nos. AAK95868 and AAF59541) exhibiting 44-52 % sequence similarity to human factor
B. Neither a prokaryotic homologue nor a counterpart in Saccharomyces cerevisiae has been
identified.
Role of factor B – The results presented here clearly show that factor B is necessary for the
energy transduction activity of the ATP synthase complex. Treatment of well-coupled SMP with
ammonia/EDTA at pH 8.8 resulted in the specific removal of a protein of Mr 22 kDa, and in the
inability of the AE-SMP to develop and maintain a membrane potential as a result of ATP hydrolysis
or NADH oxidation (Fig. 7). This defect could be repaired by addition to the AE-SMP of a
recombinant human protein of Mr 22 kDa at a molar concentration nearly stoichiometric to the ATP
synthases of the particles. The recombinant human protein was homologous in molecular mass and
amino terminal sequence to the purified bovine factor B of Sanadi and coworkers (11), and like all
factor B preparations was sensitive to treatment with monothiol and especially dithiol modifiers.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

20
The bovine ATP synthase complex contains at least 7 subunits of totally unknown function,
namely subunits ε, F6, A6L, d, e, f, and g. This complexity makes it difficult to employ the E. coli
ATP synthase with only 8 unlike subunits (29) as a model into which to build a role for factor B.
However, the basic operational design of the E. coli ATP synthase, which is composed of a catalytic,
a rotor, and a stator domain (30,31) applies to the more complicated ATP synthases (32,33), and we
will discuss the role of factor B in the light of this basic design. The fact that factor B is not a
component of the catalytic domain is clear and needs no further consideration. It cannot be a
necessary component of the stator either, because unlike the systems lacking OSCP the ATPase
activity of AE-SMP (5 µmol/min/mg) is completely inhibited by FO inhibitors, including DCCD
whose mechanism of inhibition is well known. The sensitivity of the ATPase activity of AE-SMP to
FO inhibitors also indicates that in these particles the rotating part of F1, i.e., γ and δ, is not
disengaged from FO. These considerations allow the conclusion, therefore, that factor B is a
component of FO.
As seen in Fig. 7, AE-SMP are uncoupled and incapable of forming a membrane potential as
a result of respiration or ATP hydrolysis. This defect can be repaired by addition of factor B or by
addition of FO inhibitors at concentrations that these reagents inhibit ATP hydrolysis. Repair by
factor B allows membrane potential formation as a result of ATP hydrolysis or respiration. Repair
by FO inhibitors allows only respiration-dependent membrane potential formation, which indicates
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

21
that the proton leak of AE-SMP involves, at least in part, the normal proton channel of FO. This
leads to the following important question. Since the proton leak of AE-SMP can be blocked by FO
inhibitors, especially DCCD, does the proton leak from the cytosolic (positive, P) side to the matrix
(negative, N) side of the membranes in respiring AE-SMP require the rotation of the c ring? Based
on the generally accepted basic design and operation of the ATP synthase complex, the answer to
this question is no. This is because in respiring AE-SMP the proton leak, which can be blocked by
DCCD, occurs in the absence of F1 substrates. These considerations allow the conclusion that the FO
of mammalian ATP synthase is capable of uncoupled transmembrane proton translocation via a
second path that involves, at least in part, its normal proton channel, possibly including Glu-58 of
subunit c. Proton translocation via this second path does not require the operation of the rotor of the
ATP synthase, and can be blocked by the water-soluble factor B which appears to bind to FO on the
matrix side.
Whether the FO subunits F6, A6L, d, e, f and g, which like factor B do not have prokaryotic
counterparts, are involved in this second proton path remains to be seen. Another mammalian FO-F1
subunit that does not have a prokaryotic counterpart is the ATPase inhibitor protein, IF1, which binds
to F1 β subunits and prevents futile ATP hydrolysis when the protonmotive force is low (34-36).
The fact that factor B can also be easily and reversibly removed from FO-F1, and the fact that its
displacement results in dissipation of membrane potential may be indicative of a regulatory function
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

22
for factor B as well. It is generally assumed that the state 4 rate of respiration is due to a slow proton
leak through the mitochondrial inner membrane at high protonmotive force. However, it is well
known that proteoliposomes of sonicated phospholipids plus a proton pump such as cytochrome
oxidase or nicotinamide nucleotide transhydrogenase are not as proton leaky at high protonmotive
force as SMP. Therefore, considering that over-reduction of the respiratory chain results in an
increased rate of superoxide anion production, which leads to the formation of toxic H2O2 and
hydroxyl radicals (37,38), it is possible that factor B acts as a pressure valve for maintaining the
protonmotive force (hence the reduced level of the respiratory chain) below a damaging threshold
(see also the concept of “mild uncoupling” in 38).
Finally, another point that deserves consideration is the designation “factor B” for the protein
discussed here. Since more than two decades ago, the word “factor” has correctly been replaced by
“subunit,” and already there are among the ATP synthase subunits a β subunit and a b subunit.
Continuing the alphabetical sequence beyond e, f, and g would result in confusion with the yeast
ATP synthase subunit designations. Therefore, it may be appropriate to designate the protein under
consideration here as subunit s, after its original discoverer.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

23
Acknowledgments
We thank Drs. Akemi Matsuno-Yagi and Mutsuo Yamaguchi for their interest and help in
this work, and C. Munoz for the preparation of bovine heart mitochondria. Synthesis of
oligonucleotides was supported in part by the Sam and Rose Stein Endowment Fund.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

24
REFERENCES
1. Walker, J.E., Lutter, R., Dupuis, A., and Runswick, M.J. (1991) Biochemistry 30, 5369-5378.
2. Collinson, I.R., Runswick, M.J., Buchanan, S.K., Fearnley, I.M., Skehel, J.M., Van Raaij, M.J.,Griffiths, D.E., and Walker, J.E. (1994) Biochemistry 33, 7971-7918.
3. Belogrudov, G.I., Tomich, J.M., and Hatefi, Y. (1996) J. Biol. Chem. 271, 20340-20345.
4. Lam, K.W., Warshaw, J.B., and Sanadi, D.R. (1967) Arch. Biochem. Biophys. 119, 477-484.
5. Sanadi, D.R. (1982) Biochim. Biophys. Acta 683, 39-56.
6. You, K.-S., and Hatefi, Y. (1976) Biochim. Biophys. Acta 423, 398-412.
7. Stiggall, D.L., Galante, Y.M., Kiehl, R., and Hatefi, Y. (1979) Arch. Biochem. Biophys. 196,638-644.
8. Lam, K.W., Swann, D., and Elzinga, M. (1969) Arch. Biochem. Biophys. 130, 175-182.
9. Joshi, S., Hughes, J.B., Shaikh, F., and Sanadi, D.R. (1979) J. Biol. Chem. 254, 10145-10152.
10. Shankaran, R., Sani, B.P., and Sanadi, D.R. (1975) Arch. Biochem. Biophys. 168, 394-402.
11. Kantham, L., Raychowdhury, R., Ogata, K.K., Javed, A., Rice, J., and Sanadi, D.R. (1990)FEBS Lett. 277, 105-108.
12. Joshi, S., Kantham, L., Kaplay, S., and Sanadi, D.R. (1985) FEBS Lett. 179, 143-147.
13. Hatefi, Y., and Lester, R.L. (1958) Biochim. Biophys. Acta 27, 83-88.
14. Joshi, S., and Sanadi, D.R. (1979) Methods Enzymol. 55, 384-391.
15. Senior, A.E., and Brooks, J.B. (1979) Arch. Biochem. Biophys. 140, 257-266.
16. Matsuno-Yagi, A., and Hatefi, Y. (1984) Biochemistry 23, 3508-3514.
17. Stiggall, D.L., Galante, Y.M., and Hatefi, Y. (1979) Methods Enzymol. 55, 308-315.
18. Belogrudov, G., and Hatefi, Y. (1994) Biochemistry 33, 4571-4576.
19. Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.C., Smith, J.A., Struhl, K.,Albright, L.M., Coen, D.M., Varki, A., and Chanda, V.B. (Eds.) (1997) Current Protocols inMolecular Biology, Wiley, New York.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

25
20. Tzagoloff, A., Byington, K.H., and MacLennan, D.H. (1968) J. Biol. Chem. 243, 2405-2412.
21. Lutter, R., Saraste, M., van Walraven, H.S., Runswick, M.J., Finel, M., Deatherage, J.F., andWalker, J.E. (1993) Biochem. J. 295, 799-806.
22. Groth, G., and Walker, J.E. (1996) Biochem. J. 318, 351-357.
23. McEnery, M.W., Buhle, E.L., Aebi, U., and Pedersen, P.L. (1984) J. Biol. Chem. 259, 4642-4651.
24. Galante, Y.M., Wong, S.-Y., and Hatefi, Y. (1981) Biochemistry 20, 2671-2678.
25. Matsuno-Yagi, A., and Hatefi, Y. (1988) Biochemistry 27, 335-340.
26. Lee, C.-P., and Ernster, L. (1965) Biochem. Biophys. Res. Commun. 18, 523-529.
27. Lander, E.S., Linton, L.M., Birren, B., Nusbaum, C., Zody, M.C., Baldwin, J., Devon, K., et al.(2001) Nature 409, 860-921.
28. Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., and Lipman, D.J.(1997) Nucleic Acids Res. 25, 3389-3402.
29. Fillingame, R.H. (1990) The Bacteria (T.A. Krulwich, ed.) Vol. XII, pp. 345-391, AcademicPress, New York.
30. Hutcheon, M.L., Duncan, T.M., Ngai, H., and Cross, R.L. (2001) Proc. Natl. Acad. Sci. U.S.A.98, 8519-8524.
31. Junge, W., Pänke, O., Cherepanov, D.A., Gumbiowski, K., Müller, M., and Engelbrecht, S.(2001) FEBS Lett. 504, 152-160.
32. Stock, D., Leslie, A.G.W., and Walker, J.E. (1999) Science 286, 1700-1705.
33. Stock, D., Gibbons, C., Arechaga, I., Leslie, A.G.W. and Walker, J.E. (2001) Curr. Opin. Struct.Biol. 10, 672-679.
34. Schwerzmann, K., and Pedersen, P.L. (1986) Arch. Biochem. Biophys. 250, 1-18.
35. Hashimoto, T. Yoshida, Y., and Tagawa, K. (1990) J. Bioenerg. Biomembr. 22, 27-38.
36. Ichikawa, N., Yoshida, Y., Hashimoto, T., Ogasawara, N., Yoshikawa, H., Imamoto, F., andTagawa, K. (1990) J. Biol. Chem. 265, 6274-6278.
37. Chance, B., Sies, H., and Boveris, A. (1979) Physiol. Rev. 59, 527-605.
38. Papa, S., and Skulachev, V.P. (1997) Mol. Cell. Biochem. 174, 305-319.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

26
Footnotes
1The abbreviations used are: SMP, bovine heart submitochondrial particles; EDTA,
ethylenediaminetetraacetic acid; AE-SMP, SMP depleted of factor B; OSCP, oligomycin-sensitivity-
conferring protein; DCCD, N,N’-dicyclohexylcarbodiimide; FCCP, carbonyl cyanide p-
trifluoromethoxyphenylhydrazone; NEM, N-ethylmaleimide; pCMB, p-chloromercuribenzoate;
ORF, open reading frame; PCR, polymerase chain reaction; and SDS, sodium dodecyl sulfate.
2The residues G-33 and Q-43 in the partial sequence of the bovine factor B published by Sanadi and
coworkers (11) may in fact be C-33 and E-43 as in the human protein (see GenBank accession
number AV611504, which might be a partial nucleotide sequence of the bovine factor B cDNA).
3The ease with which factor B can be dissociated from the membrane may be related in part to the
fact that most mammalian FO-F1 preparation reported in the literature exhibited low or no ATP-32Pi
exchange activity (see, for example 20-23).
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

27
LEGENDS TO FIGURES
Fig. 1. The nucleotide and the deduced amino acid sequences of human factor B. Arrow
indicates the start of the sequence of the mature human factor B shown in bold letters and numbered
on the right. Star indicates the termination codon. For details see Materials and Methods. This
nucleotide sequence has been submitted to the GenBankTM/EB1 Data Bank with accession number
AY052377.
Fig. 2. Expression and purification of mature human factor B as depicted on SDS-
polyacrylamide gel. Lane A, uninduced total E. coli cell lysate; lane B, total cell lysate after
induction; lane C, soluble E. coli proteins after induction; lane D, purified thioredoxin-factor B fused
preparation; lane E, fused protein of lane D after treatment with enterokinase; lane F, purified human
factor B containing an extra amino terminal L-A-L tripeptide. Lanes A, B and C received 20 µg of
protein each, and lanes D, E and F 5 µg of protein each. The gel (SDS-15% polyacrylamide) was
stained with Coomassie Brilliant Blue. Numbers on the left show an Mr scale.
Fig. 3. Immunoblots of intact bovine heart mitochondria (column A), SMP (column B), and
AE-SMP (column C) blotted with subunit-specific polyclonal antibodies to recombinant human
factor B (row 1), α subunit of bovine F1 (row 2), bovine OSCP (row 3), and bovine ATPase inhibitor
protein IF1 (row 4). For other details, see Materials and Methods.
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

28
Fig. 4. Effect of increasing concentrations of recombinant human factor B on the ATP-
driven reverse electron transfer (succinate to NAD) activity of AE-SMP. Inset shows a double-
reciprocal plot of the data indicating on the abscissa intercept half-maximal activation at 0.33 nmol
of factor B/mg of AE-SMP.
Fig. 5. Effect of increasing concentrations of oligomycin on the ATP-driven reverse electron
transfer activity of AE-SMP. AE-SMP in 0.25 M sucrose, 10 mM Tris-HCl, pH 7.8, were
preincubated for 5 min on ice with oligomycin dissolved in ethanol at the concentrations indicated,
then sampled into the reaction mixture. Ethanol concentration in the reaction mixture was < 1%.
Fig. 6. Inhibition of factor B with NEM, pCMP, phenylarsine oxide (PAO), and CdSO4.
Factor B was preincubated with the indicated reagents as described in Materials and Methods, then
added to the reaction mixture for reconstitution of the ATP-driven reverse electron transfer activity
of AE-SMP. The factor B concentration in the reaction mixture was 17.6 µg of factor B/mg of AE-
SMP. 100% activity was 160 nmol NADH formed (min • mg AE-SMP)-1.
Fig. 7. Effects of factor B and oligomycin on membrane potential formation by AE-SMP.
The reaction mixture at 30°C contained 0.25 M sucrose, 50 mM Tris-HCl, pH 7.5, 2 mM MgCl2, 2
µM oxonol VI, and 0.1 mg of SMP (traces A and B) or AE-SMP (traces C-F) per ml. Where
indicated 4.5 mM ATP, 0.25 mM NADH, 5 µg of oligomycin/ml, 5 µM rotenone, and 5 µM FCCP
were added. In experiments E and F, the reaction mixtures contained 6 µg of factor B. In Trace D,
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

29
oligomycin was added 7.5 min after the addition of NADH (see the 4 min gap shown) to allow
NADH to become completely oxidized. Membrane potential formation was monitored by the
absorbance change of oxonol VI at 630 minus 603 nm in an SLM DW-2000 dual wavelength
spectrophotometer.
Fig. 8. Segment of the human genome on chromosome 14q21.3 containing in exons 1-5
(filled boxes) the nucleotide sequence shown in Fig. 1 for the human factor B cDNA, except for
substitution of T168 for C in the draft sequence of the Human Genome Project (27). The start codon
(ATG), the two stop codons (TGA and TAA), and the two polyadenylation signals are marked. This
figure includes in part (exons 1-3) the DNA sequence encoding the hypothetical protein HSU79253
(see text).
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

30
CCTGCTCTGTACTCGACCCGGCACTGGGTACCGGAAGAACCAGACAGCTCGGTTTTTGCC
ACCATTTATAAGTCTGTGTCCTTTTCTTGAGTACTGAACCGAGATACAGTTTTAAATGTG M C
CTGTGCGGTCTCTGAGCAGCGACTCACCTGTGCAGATCAAATGATGCTGTTTGGAAAAAT C A V S E Q R L T C A D Q M M L F G K I
TTCCCAGCAGTTGTGTGGCGTAAAGAAACTCCCATGGTCATGTGACTCCAGATACTTCTG S Q Q L C G V K K L P W S C D S R Y ↓↓F W 2
GGGCTGGTTGAATGCAGTGTTTAATAAGGTGGATTATGATCGCATCAGGGATGTTGGCCC G W L N A V F N K V D Y D R I R D V G P 22
TGACAGGGCGGCATCCGAGTGGTTGCTGCGCTGTGGGGCCATGGTGCGCTACCATGGCCA D R A A S E W L L R C G A M V R Y H G Q 42
GGAGAGGTGGCAGAAGGACTACAACCACCTTCCAACAGGCCCTCTGGACAAATACAAGAT E R W Q K D Y N H L P T G P L D K Y K I 62
TCAGGCGATCGACGCCACCGACTCTTGTATCATGAGCATTGGATTTGATCACATGGAGGG Q A I D A T D S C I M S I G F D H M E G 82
CCTAGAGCATGTTGAAAAAATAAGGCTGTGCAAGTGTCATTATATCGAGGATGACTGTTT L E H V E K I R L C K C H Y I E D D C L 102
GCTGAGACTTAGTCAACTTGAAAATTTACAAAAAACCATATTGGAAATGGAAATAATATC L R L S Q L E N L Q K T I L E M E I I S 122
CTGTGGGAATATCACAGACAAAGGCATCATTGCTTTGCGTCATTTAAGAAACCTCAAATA C G N I T D K G I I A L R H L R N L K Y 142
TTTGTTGTTAAGTGATCTTCCTGGAGTAAGAGAAAAAGAAAATCTTGTCCAAGCCTTTAA L L L S D L P G V R E K E N L V Q A F K 162
GACAGCACTGCCTTCTCTGGAACTAAAATTACAATTGAAGTAA T A L P S L E L K L Q L K * 175
FIGURE 1
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from







Grigory I. Belogrudov and Youssef HatefiFactor B and the mitochondrial ATP synthase complex
published online December 14, 2001J. Biol. Chem.
10.1074/jbc.M111256200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from