flavone and isoflavone phytoestrogens are agonists of...
TRANSCRIPT

Flavone and Isoflavone Phytoestrogens Are Agonistsof Estrogen-Related Receptors
Masatomo Suetsugi,1 Leila Su,2 Kimberly Karlsberg,2 Yate-Ching Yuan,2 and Shiuan Chen1
1Department of Surgical Research and 2Division of Information Sciences, Beckman Research Institute of theCity of Hope, Duarte, CA
AbstractWhile estrogen-related receptors (ERRA, ERRB, and
ERR;) share a high amino acid sequence homology with
estrogen receptors (ERs), estrogens are not ligands of
ERRs. Structure-function studies from this and other
laboratories have revealed that ERRs have small
ligand-binding pockets and have provided evidence
to show that these receptors can activate gene
transcription in a constitutive manner. To address the
question as to whether there is any agonist for ERRs,
our laboratory recently performed virtual ligand
screening on ERRA that predicted flavone and
isoflavone phytoestrogens to be ligands of this receptor.
Our mammalian cell transfection and mammalian
two-hybrid experiments revealed that three isoflavones
(genistein, daidzein, and biochanin A) and one flavone
(6,3V,4V-trihydroxyflavone) behaved as agonists of ERRs.
These phytoestrogens induced the activity of ERRA at
concentrations that are comparable to those for the
activation of ERA and ERB. In this study, we also used
the results of ERRA ligand-binding site mutant, F232A,
to verify our ERRA hypothetical computer model. Our
recent ERR research has determined for the first time
that flavone and isoflavone phytoestrogens are agonists
of ERRs. In addition, our studies have demonstrated that
an approach that combines structure-based virtual
screening and receptor functional assays can identify
novel ligands of orphan nuclear receptors.
IntroductionThere are three members in the estrogen-related receptor
(ERR) family, that is, ERRa, ERRh, and ERRg. The cDNAs
for ERRa and ERRh were first isolated by screening cDNA
libraries using probes corresponding to the DNA-binding
domain of the human estrogen receptor a (ERa; 1). ERRg
was identified during an analysis of the critical region of type
IIa Usher syndrome (2), and was also identified by yeast two-
hybrid screening, using the transcriptional coactivator gluco-
corticoid receptor interacting protein 1 (GRIP1) as bait (3).
While ERRs share a high amino acid sequence homology with
ERs, estrogens are not ligands of ERRs. In fact, ERRs are
transcriptionally active in the absence of exogenous ligand. In a
recent study, we generated results that lead us to propose that
Phe-2321 (4) in ERRa (analogous to Ala-350 in ERa) plays an
important role for the constitutive activity of ERRa (5). The
ERRa mutant F232A lost the transactivation activity and acted
as a dominant negative mutant. On the other hand, like wild-
type ERRa, the ERa mutant A350F was found to be
constitutively active (5). Our previous molecular model of
ERRa revealed that the side chain of Phe-232 might be able to
mimic bound ligand because it partially fills the binding pocket.
As a result, the receptor is constitutively active. The X-ray
structure of ERRg has been recently published (6). The
structure reveals that the ligand-binding pockets of ERRs are
very small and provide additional structural information to
support the conclusion that ERRs are constitutively active.
During the last several years, four compounds have been
shown to act as antagonists of ERRs. Using yeast-based assays
and mammalian transient transfection assays, we have
previously found that two organochlorine pesticides, toxa-
phene and chlordane, can act as antagonists of ERRa,
suppressing its constitutive activity (7). Diethylstilbestrol
(DES) and 4-hydroxytamoxifen (4-OHT) have also been
found to be antagonists of ERRs by two laboratories (8, 9).
Because ERRs are constitutively active, it is easier to search
for antagonists that suppress the basal activity than to identify
agonists, which have to be able to augment the basal activity.
To address the important question as to whether there are any
agonists of ERRs, we used an approach that combines
structure-based virtual screening (SVS) and receptor functional
assays to search for agonists. Our studies have revealed that
flavone and isoflavone phytoestrogens can act as agonists of
ERRs (Fig. 1). Results from these studies will be presented
and discussed.
ResultsComputer Modeling of the Human ERRa Ligand-Binding Domain
Virtual ligand screening by docking requires a good
representative three-dimensional structure of the considered
Received 8/25/03; revised 10/13/03; accepted 10/16/03.The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.Grant support: National Institutes of Health Grants ES08258 and CA44735.Note: Masatomo Suetsugi and Leila Su contributed equally to this work.Requests for reprints: Shiuan Chen, Department of Surgical Research, BeckmanResearch Institute of the City of Hope, 1450 East Duarte Road, Duarte, CA91010. Phone: (626) 359-8111, ext. 63454; Fax: (626) 301-8972.E-mail: [email protected] D 2003 American Association for Cancer Research.
1Please note that ERRa was numbered according to the protein sequence recentlyupdated in GenBank (NM_004451; 4). The ERRa numbering will be usedthroughout the text, if not otherwise mentioned.
Vol. 1, 981–991, November 2003 Molecular Cancer Research 981
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

target. To screen for agonists of ERRa, it would be advantageous
to use an agonist-bound ERRa crystal structure. While an ERRa
ligand-bound structure has not yet been reported, a ligand-free
human ERRg structure (PDB code 1KV6) was reported by
Greschik et al. (6) that shares 57% amino acid sequence identity
with the human ERRa ligand-binding domain (LBD). We
prepared a homology model of ERRa based on the ERRg free
protein X-ray structure and found that it possessed a very small
‘‘ligand’’-binding pocket. More specifically, the side chain of
Phe-399 on helix 11 protruded into the ligand-binding pocket,
partially filled the cavity, and interfered with the formation of
hydrogen bonds between His-398 and the ligand. The position of
this Phe-399, together with the pull-in of Leu-305, Phe-286, as
well as helices 3, 5, 7, 11, and 12, led to a tightly packed pocket
that is only about half of the size of ERa as indicated by Greschik
et al. (6). Due to the small size of the pocket, we decided not to
use this model for ligand screening.
A homology model of ERRa using the DES-bound agonist
form of human ERa LBD as the template was previously
generated from our laboratory and was used to explain the
constitutive activity of ERRa (5). Since then, several high-
resolution X-ray crystal structures of the agonist-bound human
ERa have been published. Sequence homology between the
human ERRa LBD (235 residues) and the human ERa LBD is
34% amino acid identity and 55% similarity. The homology is
better conserved in the ligand-binding pocket formed by
22 residues (10), where the two receptors share amino acid
sequence identity of 45% and similarity of 74%. For the
purpose of generating an accurate, diversified, and unbiased
ligand-binding pocket for the screening of agonists of ERRa,
three human ERa crystal structures with different agonist
complexes [i.e. , DES, (R ,R)-5,11-cis-diethyl-5,6,11,12-tetrahy-
drochrysene-2,8-diol, and 17h-estradiol] were used as tem-
plates (PDB codes 3ERD, 1L2I, and 1GWR, respectively). This
second-generation structure model has the common classical
three-dimensional features of the nuclear receptor family. The
22 residues forming the ligand-binding cavity are mostly
hydrophobic (Fig. 2). The model was minimized as described in
‘‘Materials and Methods’’ and was used to screen for ERRa
agonists. Our previous study suggested that Phe-232 on helix 3,
one of the amino acids in the ligand-binding pocket of ERRa, is
responsible for the constitutive activity of ERRa (5). The
mutant F232A, noted as ERRaM, has a significantly lower
activity than the wild-type receptor. A homology model of
ERRaM was built by the same procedure as described for
ERRa and used to investigate the role of Phe-232 in the protein
activity and ligand binding. It is interesting to note that, like
ERa and ERh, ERRh and ERRg also have an alanine residue
in this site. Phe-232 is a unique structural feature in the ligand-
binding site of ERRa.
The quality of our ERRa and ERRaM models was
evaluated by checking the stereochemistry, local geometry,
solvent accessible surface areas, and side chain conformational
probabilities with the ProTable module from SYBYL (11).
Analysis of the Ramachandran plot showed that 97.5% of the
residues in the two models have the U/w dihedral angles in the
most favored or allowed regions. Only Thr-191 at the NH2
termini and Glu-343 in the loop region connecting H8 and H9,
which are far from the ligand-binding cavity, sit in disallowed
regions of the Ramachandran plot, and were corrected later by
energy minimization. The overall average energies of the
models calculated by MatchMaker energy plot (11) were below
zero, indicating that there is no major problem with the
structures and the models could be used for further analysis.
Structural Features of the Ligand-Binding Pocket of ERRaCompared with the ER family, members of the ERR family
have ligand-binding pockets that contain bulkier residues
(Fig. 2). As a result, human ERRg (ligand-binding pocket vol-
ume of 220 A3, 6) and human ERRa (295 A3 from our modeling
study, Fig. 3B) have smaller ligand-binding pockets than human
ERa (450 A3) and human ERh (390 A; 11). Our new ERRa
model based on ERa ligand-bound structures indicates that three
phenylalanine residues, Phe-232 from H3, Phe-399 from
H11, and Phe-414 from H12, line up on one side of the ERRa
FIGURE 1. Chemical structures of17h-estradiol (E2), 5,7,4V-trihydroxyi-soflavone (genistein), 7,4V-dihydroxyi-soflavone (daidzein), 5,7-dihydroxy-4V-methoxyisoflavone (biochanin A),and 6,3V,4V-trihydroxyflavone (fla-vone). Carbons of E2 and genisteinwere numbered.
Flavones and Isoflavones Are Agonists of ERRs982
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

ligand-binding cavity and stack with each other (Fig. 3, B and C).
These stacked aromatic side chains fill the upper portion of the
h-face of the cavity and change the pocket from the ‘‘wedge’’
shape in ER (Fig. 3A) to a more flattened shape (Fig. 3B).
A conserved water molecule has been indicated to be
important for ligand binding to human ERa and ERh(12–14), and is also structurally conserved among other
steroid receptors (15). The water molecule was manually
added to our ERRa model by superimposing the modeled
structure with the ERa template, followed by unrestrained
energy minimization. This water resides at the narrow end of
the h-face of the ligand-binding cavity of ERRa receptors and
could stabilize the ligand-receptor complex by forming
hydrogen bonds with hydroxyl groups of the ligand and
Glu-235 and Arg-276 of the receptor. The hydrogen bonding
between the water and the ligand could help the ligand dock
into the pocket (Fig. 3, B, D, E, and F).
Screening of a Virtual Database for Agonists of ERRaWhile our previous computer model of ERRa had revealed
that the side chain of Phe-232 might be able to mimic bound
ligand in the free protein, our new model suggests that in the
presence of ligand, Phe-232 took a different rotamer to stack
with Phe-414 and opened up the pocket for ligand binding. This
new model was used in the search for ERRa agonists. A virtual
screening of 603 compounds in the Indofine catalog was
performed (see ‘‘Materials and Methods’’). Each compound in
the Indofine database was flexibly docked into the modeled
ERRa agonist-binding pocket with the conserved water present.
Those ligands that failed to dock into the pocket were filtered
out using a TRIPOS SPL script. To verify our findings from the
virtual screening, 50 compounds in the hit list were purchased
and tested. Mammalian cell transfection and mammalian two-
hybrid functional assays were used to test these compounds
(discussed in the next section). Four ligands, ranked within the
top 30 in the hit list, including three isoflavones (genistein,
daidzein, and biochanin A) and a flavone (6,3V,4V-trihydroxy-flavone), demonstrated the ability to enhance ERRa activity
and were thus identified as ERRa agonists (Fig. 1). Two known
antagonists of ERRs, 4-OHT and DES, and the ERa agonist,
E2, failed to dock in our virtual screening. E2 is known not to
be a ligand of ERRs. 4-OHT and DES have been shown to be
antagonists of ERRh and ERRg, thus they should not bind to
the agonist-bound form of ERRa (16).
Confirmation of the Interaction of Phytoestrogens WithERRa by Mammalian Transfection and MammalianTwo-Hybrid Experiments
Our computer docking analyses have predicted that flavone
and isoflavone phytoestrogens are ligands of ERRa. Using
mammalian transfection and mammalian two-hybrid experi-
ments, we have found that three isoflavones (genistein,
daidzein, and biochanin A) and one flavone (6,3V,4V-trihydrox-yflavone; Fig. 1) can act as agonists of ERRa. The mammalian
transfection experiments demonstrated that these four com-
pounds also act as agonists of ERRh (Fig. 4). However, the
induction of the activation of ERRg by genistein and daidzein
were not statistically significant, suggesting that these two
isoflavones are relatively poor ligands of ERRg. Dose-response
studies to evaluate the binding of these compounds to ERRa
were performed, and the results were compared to those
generated with ERa and ERh (Fig. 5). Although ERRa has a
high constitutive activity, a dose-dependent increase of the
reporter activity in the presence of phytoestrogens was
observed. In addition, the maximal activity after phytoestrogen
treatment was similar among ERRa, ERa, and ERh (Fig. 5).
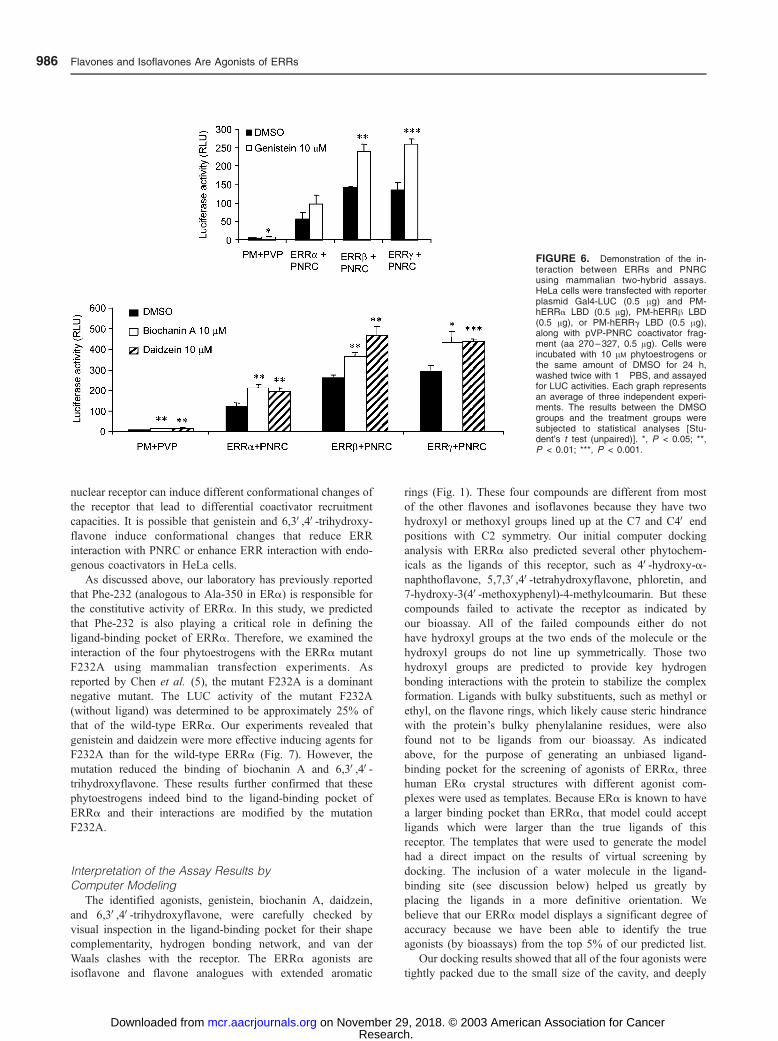
The interaction of phytoestrogens with ERRs was further
examined using mammalian two-hybrid analysis. The three
isoflavones were shown to enhance the interaction between
ERRs and the coactivator PNRC (Fig. 6). These results
FIGURE 2. Schematic comparison of amino acidresidues that form the ligand-binding pocket in hERRa
with the corresponding residues of hERa (underlined ).Genistein is shown in the binding pocket as in thecrystal structure 1KQM, where it takes the orientationas the phenyl ring sits in the narrow end of the bindingpocket and forms hydrogen bonds with the glutamatein helix 3, the arginine in helix 5, and a water molecule.Two distal hydroxyl groups (7-OH and 4V-OH ) arelabeled. The bulky phenylalanine residues couldcontribute to the constitutive activity of ERRa and areindicated by the box .
Molecular Cancer Research 983
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

confirmed that the compounds tested here are indeed agonists
of ERRs. It is recognized that the increase in the interaction
between ERRa and PNRC by genistein is not statistically
significant. Furthermore, while 6,3V,4V-trihydroxyflavone was
shown to be an agonist of ERRs using the mammalian
transfection assays, this compound was not able to enhance
the interaction between ERRs and PNRC (results not shown).
Several studies (17–19) have found that different ligands of a
FIGURE 3. The binding pocket of DES-bound ERa (A), the binding pocket of genistein-bound ERRa (B), and ribbon representation of the ligand-bindingsite (C). H3, H5, H7, and H12 are shown in yellow . F414 from H12 forms aromatic stacking interactions with F232 from H3 and F399 from H11. D. Thebinding pocket of ERRaM with genistein. E. The binding pocket of 6,3V,4V-trihydroxyflavone-bound ERRa. F. The binding pocket of ERRaM with 6,3V,4V-trihydroxyflavone. The pose of each ligand was chosen by CScore. Molecular surfaces of the ligand-binding cavity have been rendered with a translucentvan der Waals surface (1.4 A probe) and colored in green in A, B, D, E, and F. Proteins are shown as stick representations and colored by atom type(oxygen colored red , hydrogen in cyan , and carbon in white ). Side chains of key residues (E235, R276, H398, F/A232, F286, F399) in the active site aredepicted and labeled. The conserved water molecule is shown in ball-and-stick representation . Hydrogen bonds (colored in yellow ) are shown between theligands and the proteins, as well as the water.
Flavones and Isoflavones Are Agonists of ERRs984
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from
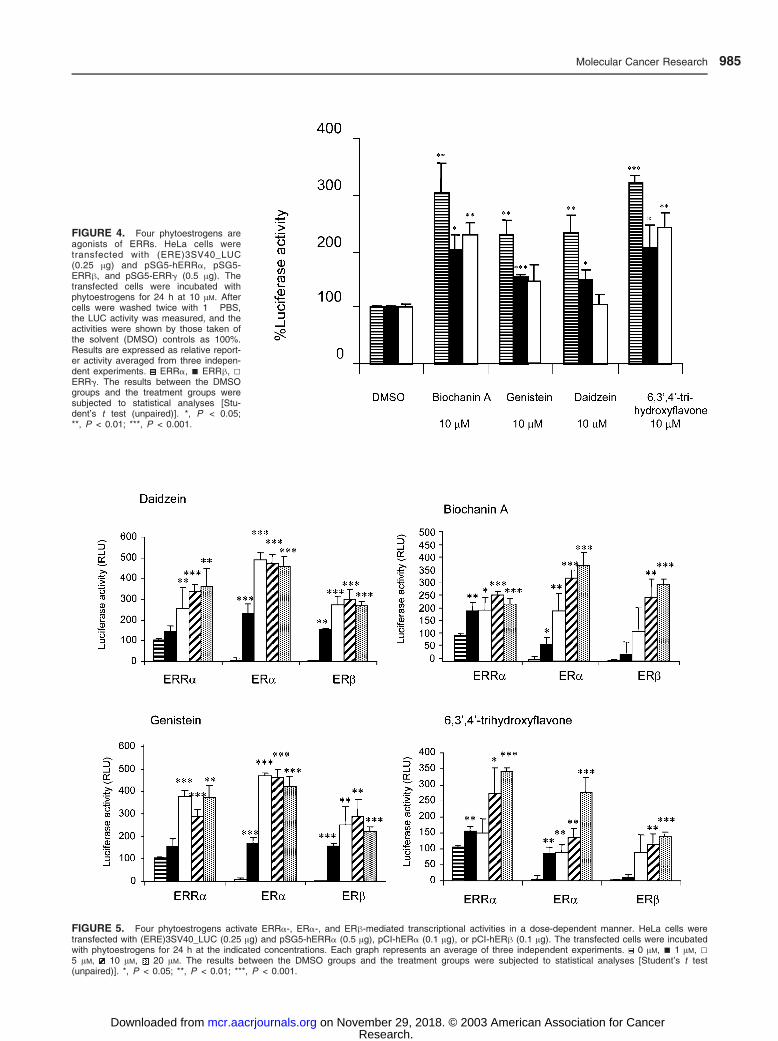
FIGURE 5. Four phytoestrogens activate ERRa-, ERa-, and ERh-mediated transcriptional activities in a dose-dependent manner. HeLa cells weretransfected with (ERE)3SV40_LUC (0.25 Ag) and pSG5-hERRa (0.5 Ag), pCI-hERa (0.1 Ag), or pCI-hERh (0.1 Ag). The transfected cells were incubatedwith phytoestrogens for 24 h at the indicated concentrations. Each graph represents an average of three independent experiments. 0 AM, n 1 AM, 55 AM, 10 AM, 20 AM. The results between the DMSO groups and the treatment groups were subjected to statistical analyses [Student’s t test(unpaired)]. *, P < 0.05; **, P < 0.01; ***, P < 0.001.
FIGURE 4. Four phytoestrogens areagonists of ERRs. HeLa cells weretransfected with (ERE)3SV40_LUC(0.25 Ag) and pSG5-hERRa, pSG5-ERRh, and pSG5-ERRg (0.5 Ag). Thetransfected cells were incubated withphytoestrogens for 24 h at 10 AM. Aftercells were washed twice with 1� PBS,the LUC activity was measured, and theactivities were shown by those taken ofthe solvent (DMSO) controls as 100%.Results are expressed as relative report-er activity averaged from three indepen-dent experiments. ERRa, n ERRh, 5ERRg. The results between the DMSOgroups and the treatment groups weresubjected to statistical analyses [Stu-dent’s t test (unpaired)]. *, P < 0.05;**, P < 0.01; ***, P < 0.001.
Molecular Cancer Research 985
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from
nuclear receptor can induce different conformational changes of
the receptor that lead to differential coactivator recruitment
capacities. It is possible that genistein and 6,3V,4V-trihydroxy-flavone induce conformational changes that reduce ERR
interaction with PNRC or enhance ERR interaction with endo-
genous coactivators in HeLa cells.
As discussed above, our laboratory has previously reported
that Phe-232 (analogous to Ala-350 in ERa) is responsible for
the constitutive activity of ERRa. In this study, we predicted
that Phe-232 is also playing a critical role in defining the
ligand-binding pocket of ERRa. Therefore, we examined the
interaction of the four phytoestrogens with the ERRa mutant
F232A using mammalian transfection experiments. As
reported by Chen et al. (5), the mutant F232A is a dominant
negative mutant. The LUC activity of the mutant F232A
(without ligand) was determined to be approximately 25% of
that of the wild-type ERRa. Our experiments revealed that
genistein and daidzein were more effective inducing agents for
F232A than for the wild-type ERRa (Fig. 7). However, the
mutation reduced the binding of biochanin A and 6,3V,4V-trihydroxyflavone. These results further confirmed that these
phytoestrogens indeed bind to the ligand-binding pocket of
ERRa and their interactions are modified by the mutation
F232A.
Interpretation of the Assay Results byComputer Modeling
The identified agonists, genistein, biochanin A, daidzein,
and 6,3V,4V-trihydroxyflavone, were carefully checked by
visual inspection in the ligand-binding pocket for their shape
complementarity, hydrogen bonding network, and van der
Waals clashes with the receptor. The ERRa agonists are
isoflavone and flavone analogues with extended aromatic
rings (Fig. 1). These four compounds are different from most
of the other flavones and isoflavones because they have two
hydroxyl or methoxyl groups lined up at the C7 and C4V end
positions with C2 symmetry. Our initial computer docking
analysis with ERRa also predicted several other phytochem-
icals as the ligands of this receptor, such as 4V-hydroxy-a-naphthoflavone, 5,7,3V,4V-tetrahydroxyflavone, phloretin, and
7-hydroxy-3(4V-methoxyphenyl)-4-methylcoumarin. But these
compounds failed to activate the receptor as indicated by
our bioassay. All of the failed compounds either do not
have hydroxyl groups at the two ends of the molecule or the
hydroxyl groups do not line up symmetrically. Those two
hydroxyl groups are predicted to provide key hydrogen
bonding interactions with the protein to stabilize the complex
formation. Ligands with bulky substituents, such as methyl or
ethyl, on the flavone rings, which likely cause steric hindrance
with the protein’s bulky phenylalanine residues, were also
found not to be ligands from our bioassay. As indicated
above, for the purpose of generating an unbiased ligand-
binding pocket for the screening of agonists of ERRa, three
human ERa crystal structures with different agonist com-
plexes were used as templates. Because ERa is known to have
a larger binding pocket than ERRa, that model could accept
ligands which were larger than the true ligands of this
receptor. The templates that were used to generate the model
had a direct impact on the results of virtual screening by
docking. The inclusion of a water molecule in the ligand-
binding site (see discussion below) helped us greatly by
placing the ligands in a more definitive orientation. We
believe that our ERRa model displays a significant degree of
accuracy because we have been able to identify the true
agonists (by bioassays) from the top 5% of our predicted list.
Our docking results showed that all of the four agonists were
tightly packed due to the small size of the cavity, and deeply
FIGURE 6. Demonstration of the in-teraction between ERRs and PNRCusing mammalian two-hybrid assays.HeLa cells were transfected with reporterplasmid Gal4-LUC (0.5 Ag) and PM-hERRa LBD (0.5 Ag), PM-hERRh LBD(0.5 Ag), or PM-hERRg LBD (0.5 Ag),along with pVP-PNRC coactivator frag-ment (aa 270 – 327, 0.5 Ag). Cells wereincubated with 10 AM phytoestrogens orthe same amount of DMSO for 24 h,washed twice with 1� PBS, and assayedfor LUC activities. Each graph representsan average of three independent experi-ments. The results between the DMSOgroups and the treatment groups weresubjected to statistical analyses [Stu-dent’s t test (unpaired)]. *, P < 0.05; **,P < 0.01; ***, P < 0.001.
Flavones and Isoflavones Are Agonists of ERRs986
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

docked into the ligand-binding site, partially due to the termini
water-bridged hydrogen bonds between hydroxyl group(s) of
the ligands and the conserved water molecule. The hydrogen
bonding groups on the two ends of the agonists form hydrogen
bonds with residues Glu-235, Arg-276, and His-398 at the
corresponding ends of the ligand-binding cavity (Fig. 3, B, D,
E, and F). Phe-286 on sheet 1 was fixed in an orientation by its
aromatic ring current interaction with the flat aromatic rings on
the agonists. The three phenylalanines, Phe-232, Phe-399, and
Phe-414, sit at the h face, while Phe-286 sits at the opposite a
face of the cavity to sandwich the agonists (Fig. 3, B, D, E, and
F). The hydrogen bonding, aromatic, and hydrophobic
interactions between the ligands and helices 3, 5, 7, 11, and
12 of the receptor stabilized the complexes of ERRa with the
phytoestrogens.
In summary, our modeling study indicated that the ERRa
agonist-binding cavity is rather flat due to the phenylalanines
in the ligand-binding site. This suggests that ligands with
extended aromatic structures make more effective agonists. Our
receptor functional analysis further indicates that an effective
agonist should be a molecule without bulky substituents in the
middle and with a pair of lined-up hydrogen bond forming
groups at the ends. The derived pharmacophore information
will enable us to identify additional agonists of ERRa by
screening larger commercial three-dimensional chemical data-
bases and verifying by mutagenesis and functional assays.
DiscussionVirtual Screening of the ERRa Agonists
SVS using a homology model has been proven to be a
valuable technique for nuclear hormone receptors, an important
therapeutic target family (20). Here, we identified for the first
time that flavone and isoflavone phytoestrogens are agonists of
all three isoforms of ERR through SVS and verified by
mammalian cell transfection and two-hybrid functional assays.
An important element of determining the success of the SVS is
the choice of the specific agonist conformation of nuclear
receptors used for screening. Our functional assay results
validate the effectiveness of using the homology model
generated from agonist-bound ERa X-ray complex structures
to automatically screen more than 600 structurally diverse
phytoestrogens and score using an effective consensus scoring
approach. Our studies have provided key structural information
FIGURE 7. Modification of the binding affinity of phytoestrogens by ERRa mutant F232A. HeLa cells were transfected with (ERE)3SV40_LUC (0.25 Ag)and pSG5-hERRa (0.5 Ag) or pSG5-hERRa F232A (0.5 Ag). Cells were incubated with 1 or 10 AM phytoestrogens, or the same amount of DMSO for 24 h. Allother experimental conditions are identical to those described in Fig. 5. The relative LUC activities were shown by those taken of the solvent (DMSO) controlsas 100%. n ERRa WT, 5 ERRa F232A. *, P < 0.05; **, P < 0.01; ***, P < 0.001.
Molecular Cancer Research 987
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

to identify additional agonists of ERRa. We believe that our
study represents an excellent example of an approach that
combines SVS and receptor functional assays to identify novel
ligands of orphan nuclear receptors.
This study was aimed at the identification of ERRa novel
agonists and we presented a successful strategy to meet that goal.
An in-depth evaluation of the agonist binding mode and
structure-activity relationship requires extended work, such as
molecular dynamics and mutagenesis experiments, and that is
beyond the scope of this discussion. Enormous advances in
genomics have identified a significant number of orphan nuclear
receptors as potential therapeutic targets. We would like to place
our emphasis on aspects of combining the reliable and
inexpensive SVS technique with nuclear receptor functional
assays for lead discovery.
Functional Significance of Phe-232 of ERRaOur previous modeling study indicated that Phe-232 is re-
sponsible for the constitutive activity of ERRa (5). The current
modeling study suggested that the Phe-232 could contribute to the
protein activity by stacking with Phe-414 of H12 to help position
and stabilize H12 in the agonist position and uphold the
constitutive activity of ERRa. It was shown in the ERRa model
that Phe-414 was sandwiched by stacking to Phe-232 of H3 and
Phe-399 of H11 on each side (Fig. 3, B and C). A close inspection
of the free ERRg structure (1KV6, 6) indicates that the
phenylalanine on H12 (Phe-450 in ERRg) and the phenylalanine
on H11 (Phe-435 in ERRg), the two conserved phenylalanines in
all three ERR isoforms, sit very close to each other and fall in the
range of van der Waals interactions (about 2.5 A proton-proton
distance). A similar type of aromatic interaction has been observed
recently from the structural analysis of the constitutively active
orphan nuclear receptor Nurr1 (21). Together with results from
previous studies (5), we hypothesize that the constitutive activity
of ERRa results from (a) the side chain of Phe-232 mimicking
bound ligand, and (b) helix 12 being kept at the agonist-bound
conformation through an aromatic interaction among Phe-232
(H3), Phe-399 (H11), and Phe-414, which is in helix 12.
The receptor functional assays carried out in our laboratory
clearly indicated that the mutation of one of the three stacked
phenylalanines of ERRa, F232A, could significantly reduce
the constitutive activity of the receptor. Furthermore, genistein
and daidzein were found to be better agonists for the mutant
F232A, while biochanin A and 6,3V,4V-trihydroxyflavone were
more effective to activate wild-type ERRa (Fig. 7). This
raised the question of how the mutation, F232A, affected the
binding of each agonist and the protein activity in the
presence of the agonist. Unlike the ER-E2 complex, in the
ERRa-agonist system, ERRa has more bulky aromatic
residues at its ligand-binding site (Fig. 2), and the agonists
are molecules with extended aromatic ring structures (Fig. 1).
These new structure features imply that aromatic interactions
between the receptor and its agonists could play a very
important role in ligand binding and receptor activity.
Modeling of the mutant F232A (i.e. , ERRaM) indicated that
the side chain of Phe-414 took a different rotamer from the
stacking position in ERRa to fill part of the space left out by
Phe-232 in the mutant (Fig. 3, D and F). Phe-414 was then
stabilized by aromatic interactions with Phe-399 of H11
(Fig. 3, D and F). H12 could be further stabilized by
interacting with the agonists. The protein activity changes
induced by the mutation are the consequences of the adjusted
protein and ligand conformations. One of the many factors
accountable for the changes could be the orientation change of
the agonists in the ligand-binding site.
A detailed structure and activity analysis requires further
work considering the very similar structural features of the four
agonists. However, there are a few points that can be addressed
from this study. It was observed that the agonists of ERRa were
able to take two different docking orientations in the binding
pocket. One is the classical orientation of the nuclear receptor
ligand, where the phenolic ring sits at the A-ring end (Fig. 3, E
and F) as in the E2-ERa complex (22). The alternate orientation
is the flavone ring, instead, residing at this end (Fig. 3, B and
D). It was shown that each agonist had its favored orientation in
the binding site, depending on the specific structural character-
istics of the agonist. Genistein and daidzein, which have only
one functional group, the 4V-OH, on the phenolic ring, favor theflavone ring at the A-ring end in both the wild type and the
mutant (Fig. 3, B and D) and are better agonists for the mutant.
On the other hand, 6,3V,4V-trihydroxyflavone, which has an
extra hydroxyl group on the phenolic ring and is a better agonist
for the wild-type protein, tended to favor the phenolic ring at
the A-ring end in both the wild-type and the mutant receptors
(Fig. 3, E and F). Unlike 6,3V,4V-trihydroxyflavone, biochaninA, which acts as a better agonist for the wild-type protein and
has a bulkier methoxyl group lacking the favored hydrogen
donor for the A-ring end (23), kept its orientation with the
flavone ring at the A-ring end in both the mutated and wild-type
proteins. The preference of different orientations under specific
circumstances was not well understood.
Implication of Physiological Significance of Our Findingsof Phytoestrogens as Ligands of ERRa
As indicated in the introduction, it is not easy to identify
agonists for a receptor that is constitutively active, because they
have to be able to augment the basal activity. This work
suggested that ERRa stays at a stable agonist conformation
through the unique Phe-232 that mimics ligand and contributes
to aromatic interactions with Phe-399 in helix 11 and Phe-414
in helix 12. It is important to point out that the binding of
flavone and isoflavone ligands to ERRa increases its activity to
the levels of ERa and ERh in the presence of the same
compounds at similar concentrations. These results suggest that
ERRa has been stimulated to its maximal level in the presence
of the ligands.
ERRa is expressed in breast tissue and its expression has
recently been shown to associate with unfavorable bio-
markers in breast cancer (24). The action of ERRa and
enhancement of its activity by phytoestrogens in breast
cancer should not be overlooked. In addition, ERRa mRNA
has been found to be more highly expressed in rat calvaria
(RC) cell cultures than either ERa and ERh (25), and a
decrease in the synthesis of ERRa (through the use of
antisense oligonucleotides) led to an inhibition of RC
proliferation and bone nodule formation in vitro (26). The
Flavones and Isoflavones Are Agonists of ERRs988
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

interaction of phytoestrogens with ERRa can play important
roles in breast and bone, a newly identified action of
phytoestrogens. The effect of phytoestrogens on ERRa may
not be easily observed when functionally active ERa or ERhis present because these phytochemicals are also ligands of
ERs. The effect of phytoestrogens (through the interaction
with ERRa) in breast and bone in women could be detected
when the function of ERs is suppressed, such as through the
use of antiestrogens.
Our study has provided a novel approach to identifying
environmental chemicals that interact with ERRa as agonists
through the use of SVS and cell culture-based functional
analyses. We will further screen larger commercial chemical
databases for both agonists and antagonists of ERRa and verify
these results by mutagenesis and functional assays. This
research has a translational impact because the results generated
can be used for designing prevention strategies against breast
cancer. One should avoid exposure to those chemicals shown to
act as agonists of ERRs that may induce breast cancer
development. On the other hand, chemicals that act as
antagonists of these receptors may have value as preventative
agents against breast cancer. However, through the interaction
with ERRa in bone, agonists of ERRa may prevent bone loss
in women who use antiestrogens.
Materials and MethodsMaterials
5,7-Dihydroxy-4V-methoxyisoflavone (biochanin A), 7,4V-dihydroxyisoflavone (daidzein), 5,7,4V-trihydroxyisoflavone(genistein), and 6,3V,4V-trihydroxyflavone were purchased from
Indofine Chemical Co., Inc. (Somerville, NJ). The structures of
these compounds are shown in Fig. 1. hERRh cDNA and HeLa
cervix adenocarcinoma cells were purchased from American
Type Culture Collection (ATCC, Manassas, VA). HeLa cells
were maintained in Eagle’s MEMwith nonessential amino acids,
sodium pyruvate, and 10% fetal bovine serum at 37jC and 5%
CO2. Charcoal/dextran-treated serum was obtained from Gemini
Bio-Products (Woodland, CA). Lipofectin was purchased from
Invitrogen Life Technologies (Palo Alto, CA).
PlasmidAll recombinant DNA and plasmid construction experi-
ments were performed according to standard procedures. The
sequence and orientation of inserted DNA fragments in
plasmid constructs were verified by standard DNA sequenc-
ing. The expression plasmids, pSG5-ERRa and pSG5-ERRa
mutant F232A, were constructed by Chen et al. (5). The full-
length coding region of human ERRh was generated by PCR
using forward primer 5V-GCTGGAATTCATGTCGTCCGAAGACAGGCA-3V and reverse primer 5V-TGCGGAATTCT-CACACCTTGGCCTCCAGCA-3V, with the template cDNA,
hERR2 (ATCC). The PCR product was digested with EcoRI
and subcloned into vector pSG5 through the EcoRI sites. The
full-length coding region of human ERRg was generated by
PCR using forward primer 5V-CCGGGAATTCATG-
GATTCGGTAGAACTTTG-3V and reverse primer 5V-GAGC-GAATTCTCAGACCTTGGCCTCCAACA-3V, with the
template cDNA prepared from H295 cells (ATCC). The
PCR product was digested with EcoRI and subcloned into
vector pSG5 through the EcoRI sites. The luciferase reporter
plasmid, pGL3 (ERE)3-Luciferase, which contains three
copies of the ERE sequence, was constructed by Chen et al.
(5). The pSG5-GRIP1 plasmid was kindly provided by Dr.
Michael R. Stallcup (University of Southern California, Los
Angeles, CA), and pCI-ERa and pCI-ERh were kindly
provided by Dr. Y. Kinoshita (Beckman Research Institute
of the City of Hope, Duarte, CA).
Generation of the expression plasmids for mammalian two-
hybrid analysis, that is, PM-ERRa LBD, PM-ERRh LBD, PM-
ERRg LBD, and pVP-PNRC coactivator fragment (aa 270–
327), is briefly described below: The cDNA fragments of
human ERRa LBD were generated by PCR using forward
primer 5V-CCCCGAATTCACAGCAGCCCCCAGTGAATGC-3V and reverse primer 5V-ACCCGGATCCTCAGTCCAT-CATGGCCTCGA-3V, with the template DNA, pSG5 ERRa.
The cDNA fragment of human ERRh LBD was generated by
forward primer 5V-TCCCGACGCTAAAAAGCCATTGAC-TAA-3V and reverse primer 5V-TGCGGGATCCTCA-
CACCTTGGCCTCCAGCA-3V with the template DNA,
pSG5 ERRh. Finally, the cDNA fragment of human ERRg
LBD was generated by forward primer 5V-GGTTGAATTCGC-CAAAAAGCCATATAACAA-3V and reverse primer 5V-GAGCGGATCCTCAGACCTTGGCCTCCAACA-3V, with
the template DNA, pSG5 ERRg. The PCR products were
subcloned into vector pM through the EcoRI and BamHI sites.
For pVP-PNRC270–327, the cDNA fragment of PNRC270–327
(27) was generated by PCR using forward primer 5V-GCCGGATCCTAATGACTGAAGTGAGCCAAAAGGAA-3Vand reverse primer 5V-CGCTCGGATCCCTAAGTTT-
GAACTTTTGAGGAG-3V, and the PCR product was inserted
in proper reading frame into pVP16 activation domain vector
(Clontech Laboratories, Inc., Palo Alto, CA) at the BamHI site.
Mammalian Cell Transfection and Luciferase AssaysHeLa cells were cultured in MEM Earle’s salts medium
supplemented with 5% charcoal/dextran-treated fetal bovine
serum. Cells were divided and cultured in six-well plates until
80% confluent. The cells were transfected with 4 Ag Lipofectin,
and an equal amount of total DNAwas used in all transfections
by including appropriate amounts of the empty vector, pSG5, in
addition to specific amounts of the test plasmids indicated in each
experiment. After overnight incubation, medium containing
Lipofectin and DNAwas removed, and the cells were cultured in
growth medium containing 5% charcoal/dextran-treated fetal
bovine serumwith or without ligands. After a 24-h incubation, the
cells were harvested from the plates by scraping, and the luciferase
activities in the cell lysates (with the same amounts of protein)
were measured according to the manufacturer’s instructions
(Promega, Madison, WI). All experiments were performed in
triplicate.
Mammalian Two-Hybrid AssaysHeLa cells were transiently transfected with 0.5 Ag reporter
plasmid, Gal4-luciferase (Clontech), and 0.5 Ag PM-ERRa
LBD, PM-ERRh LBD, or PM-ERRg LBD along with 0.5 AgpVP-PNRC270–327.
Molecular Cancer Research 989
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

Building the Models of ERRa LBDThree-dimensional homology modeling and SVS were
performed using the SYBYL program package (11), version
6.9 (Tripos, Inc., St. Louis, MO) on a Silicon Graphics O2+
workstation with the IRIX 6.5 operating system. The sequence
of hERRa was obtained from GenBank (NM_004451). The
FASTA search option available in the Protein Data Bank (28)
helped to identify several structural templates on which to base
the homology model. The homology models of hERRa LBD
and its mutant F232A were generated using the SYBYL
COMPOSER module (29). Three different agonist-bound X-ray
complex structures of the human ERa LBD templates (PDB
codes 3ERD, 1L2I, 1GWR) were chosen because of their high
resolution (2.03, 1.95, and 2.40 A). The structures were then
refined by torsional minimization and a series of energy
minimization steps first with ligand-binding pocket side chains,
followed by all protein side chains and finally the entire protein.
All the energy minimizations were performed using the Tripos
force field with the cutoff of nonbonded (NB) interactions at
8.0 A and the distance dielectric constant set at 4.0 following
the gradient termination of the Powell method with RMS of
0.005 kcal/mol A or the maximum 1000 iterations. The volume
of the ERRa ligand-binding pocket was estimated using
SYBYL MOLCAD separated surfaces with a grid width of
1.0 and a probe radius of 1.4 A.
Database PreparationThe Flavonoid and Coumarin Catalog was obtained from
Indofine Chemical Company. Three-dimensional structures of
the Indofine compounds were imported from the Available
Chemicals Directory (ACD; Molecular Design Limited, San
Leandro, CA) using ISIS/Base (MDL). For the known ligands of
ERs and ERRs not already included in the Indofine Catalog, such
as E2, DES, and 4-OHT, two-dimensional structures were
prepared by ISIS/Draw (MDL). Three-dimensional structures of
those molecules were generated using the Concord (Tripos)
conversion program. The new structures were then added to the
Indofine Catalog to create a final Indofine database of 603
molecules. Final coordinates were stored in a SYBYL database.
A subset of this database, containing 37 molecules (known ERa
agonists together with randomly chosen molecules), was created
as a test set to fine tune parameters for docking the Indofine
database and scoring the ligands for ERa (data not shown).
Receptor-Ligand DockingThe SYBYL FlexX program version 1.10 interfaced within
TRIPOS SYBYL 6.9 (11) was used to dock compounds to the
ligand-binding sites of ERRa and ERRaM. FlexX is a fast-
automated docking program that considers ligand conforma-
tional flexibility into a rigid protein structure by an
incremental fragment placing technique (30, 31). A structur-
ally conserved water molecule has been included in the
binding pocket for the docking. Standard parameters and
FlexX scores implemented in the program were used for
docking and scoring of FlexX poses. The ligand-protein
complex was relaxed by torsional minimization and a series of
constrained energy minimization steps. The ligand was first
minimized within the complex to an RMS of 0.001 kcal/mol
A to remove bad contacts. The side chains of amino acids in
the ligand-binding site and the entire complex were then
minimized respectively. During the minimization processes,
hydrogen bond constraints with 50 kcal/(mol A)2 force
constant were applied. Energy minimizations were carried
out using the Tripos force field with an NB cutoff of 8.0 A
and the distance dielectric constant set at 4.0 following
gradient termination using the Powell method to an RMS of
0.005 kcal/mol A or the maximum 1000 iterations.
Virtual Ligand Screening of the Indofine DatabaseEach flexible ligand of the Indofine databases composed of
flavonoids and coumarins was docked automatically into the
receptor. The FlexX score was used to guide the growing of the
ligand and was assigned to each successfully docked compound
to measure the goodness of its fit with the receptor. The FlexX
scoring function includes both polar (hydrogen bond and
charge-charge) and non-polar (hydrophobic) interactions that
are used to dock the ligand into the active site. The screening of
the Indofine database of 603 compounds took less than 5 h on a
Silicon Graphics O2+ workstation. Out of 603 screened
compounds, 426 were claimed to be ‘‘successfully docked’’
and up to 30 top scored poses of each docked compound were
saved. These 426 compounds were checked by an SPL (Sybyl
Programming Language) script to find out whether these
compounds were actually docked into the ligand-binding
pocket (flexx_pocket_mss.spl). Two hundred twenty-seven
out of these 426 ‘‘successful’’ compounds were found docked
into the pocket. ChemScore, Dock, FlexX, Gold, and PMF
scores were recalculated for the docked ligands using the
CScore module of SYBYL 6.9 for consensus scoring.
Consensus scoring has found to outperform any of the single
functions comparing the results from the test set. A hit list
containing the top scores from the three single scoring functions
(PMF, FlexX, and Gold) which were able to successfully
predict the non-binder such as E2 in the test set was generated.
The top 100 ligands in the hit list were visually inspected and
50 of them were selected, purchased, and experimentally tested.
AcknowledgmentsWe thank Dr. Dujin Zhou and TRIPOS Application Scientists for their commentsand suggestions to the research described here.
References1. Giguere, V., Yang, N., Segui, P., and Evans, R. M. Identification of a new classof steroid hormone receptors. Nature, 331: 91–94, 1988.
2. Eudy, J. D., Yao, S., Weston, M. D., Ma-Edmonds, M., Talmadge, C. B.,Cheng, J. J., Kimberling, W. J., and Sumegi, J. Isolation of a gene encoding anovel member of the nuclear receptor superfamily from the critical region ofUsher syndrome type IIa at 1q41. Genomics, 50: 382– 384, 1998.
3. Hong, H., Yang, L., and Stallcup, M. R. Hormone-independent transcriptionalactivation and coactivator binding by novel orphan nuclear receptor ERR3. J.Biol. Chem., 274: 22618–22626, 1999.
4. Yang, N., Shigeta, H., Shi, H., and Teng, C. T. Estrogen-related receptor,hERR1, modulates estrogen receptor-mediated response of human lactoferringene promoter. J. Biol. Chem., 271: 5795–5804, 1996.
5. Chen, S., Zhou, D., Yang, C., and Sherman, M. Molecular basis for theconstitutive activity of estrogen-related receptor a-1. J. Biol. Chem., 276:28465 –28470, 2001.
6. Greschik, H., Wurtz, J. M., Sanglier, S., Bourguet, W., van Dorsselaer, A.,Moras, D., and Renaud, J. P. Structural and functional evidence for
Flavones and Isoflavones Are Agonists of ERRs990
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

ligand-independent transcriptional activation by the estrogen-related receptor 3.Mol. Cell, 9: 303–313, 2002.
7. Yang, C. and Chen, S. Two organochlorine pesticides, toxaphene andchlordane, are antagonists for estrogen-related receptor a-1 orphan receptor.Cancer Res., 59: 4519–4524, 1999.
8. Tremblay, G. B., Kunath, T., Bergeron, D., Lapointe, L., Champigny, C.,Bader, J. A., Rossant, J., and Giguere, V. Diethylstilbestrol regulates trophoblaststem cell differentiation as a ligand of orphan nuclear receptor ERRh. GenesDev., 15: 833 –838, 2001.
9. Coward, P., Lee, D., Hull, M. V., and Lehmann, J. M. 4-Hydroxytamoxifenbinds to and deactivates the estrogen-related receptor g. Proc. Natl. Acad. Sci.USA, 98: 8880–8884, 2001.
10. Pike, A. C., Brzozowski, A. M., Hubbard, R. E., Bonn, T., Thorsell, A. G.,Engstrom, O., Ljunggren, J., Gustafsson, J. A., and Carlquist, M. Structure of theligand-binding domain of oestrogen receptor h in the presence of a partial agonistand a full antagonist. EMBO J., 18: 4608–4618, 1999.
11. SYBYL. 6.9 edition. St. Louis, MO: Tripos, Inc.
12. Pike, A. C., Brzozowski, A. M., Walton, J., Hubbard, R. E., Thorsell, A. G.,Li, Y. L., Gustafsson, J. A., and Carlquist, M. Structural insights into the mode ofaction of a pure antiestrogen. Structure (Camb), 9: 145– 153, 2001.
13. Shiau, A. K., Barstad, D., Loria, P. M., Cheng, L., Kushner, P. J., Agard,D. A., and Greene, G. L. The structural basis of estrogen receptor/coactivatorrecognition and the antagonism of this interaction by tamoxifen. Cell, 95: 927–937, 1998.
14. Gangloff, M., Ruff, M., Eiler, S., Duclaud, S., Wurtz, J. M., and Moras, D.Crystal structure of a mutant hERa ligand-binding domain reveals key structuralfeatures for the mechanism of partial agonism. J. Biol. Chem., 276: 15059–15065, 2001.
15. Tanenbaum, D. M., Wang, Y., Williams, S. P., and Sigler, P. B.Crystallographic comparison of the estrogen and progesterone receptor’s ligandbinding domains. Proc. Natl. Acad. Sci. USA, 95: 5998–6003, 1998.
16. Tremblay, G. B., Bergeron, D., and Giguere, V. 4-Hydroxytamoxifen is anisoform-specific inhibitor of orphan estrogen-receptor-related (ERR) nuclearreceptors h and g. Endocrinology, 142: 4572–4575, 2001.
17. Kraichely, D. M., Sun, J., Katzenellenbogen, J. A., and Katzenellenbogen,B. S. Conformational changes and coactivator recruitment by novel ligands forestrogen receptor-a and estrogen receptor-h: correlations with biologicalcharacter and distinct differences among SRC coactivator family members.Endocrinology, 141: 3534–3545, 2000.
18. Wong, C. W., Komm, B., and Cheskis, B. J. Structure-function evaluation ofER a and h interplay with SRC family coactivators. ER selective ligands.Biochemistry, 40: 6756– 6765, 2001.
19. Margeat, E., Bourdoncle, A., Margueron, R., Poujol, N., Cavailles, V., andRoyer, C. Ligands differentially modulate the protein interactions of the humanestrogen receptors a and h. J. Mol. Biol., 326: 77–92, 2003.
20. Schapira M, Abagyan R, and Totrov M. Nuclear hormone receptor targetedvirtual screening. J. Med. Chem., 46: 3045– 3059, 2003.
21. Wang, Z., Benoit, G., Liu, J., Prasad, S., Aarnisalo, P., Liu, X., Xu, H.,Walker, N. P., and Perlmann, T. Structure and function of Nurr1 identifies a classof ligand-independent nuclear receptors. Nature, 423: 555–560, 2003.
22. Brzozowski, A. M., Pike, A. C., Dauter, D., Hubbard, R. E., Bonn, T.,Engstrom, O., Ohman, L., Greene, G. L., Gustafsson, J. A., and Carlquist, M.Molecular basis of agonism and antagonism in the oestrogen receptor. Nature,389: 753 –758, 1997.
23. Anstead, G. M., Carlson, K. E., and Katzenellenbogen, J. A. The estradiolpharmacophore: ligand structure-estrogen receptor binding affinity relationshipsand a model for the receptor binding site. Steroids, 62: 268 –303, 1997.
24. Ariazi, E. A., Clark, G. M., and Mertz, J. E. Estrogen-related receptor a andestrogen-related receptor g associate with unfavorable and favorable biomarkers,respectively, in human breast cancer. Cancer Res., 62: 6510–6518, 2002.
25. Bonnelye, E. and Aubin, J. E. Differential expression of estrogen receptor-related receptor a and estrogen receptors a and h in osteoblasts in vivo and in
vitro . J. Bone Miner. Res., 17: 1392– 1400, 2002.
26. Bonnelye, E., Merdad, L., Kung, V., and Aubin, J. E. The orphan nuclearestrogen receptor-related receptor a (ERRa) is expressed throughout osteoblastdifferentiation and regulates bone formation in vitro . J. Cell Biol., 153: 971–983, 2001.2001.
27. Zhou, D., Quach, K. M., Yang, C., Lee, S. Y., Pohajdak, B., and Chen, S.PNRC: a proline-rich nuclear receptor co-regulatory protein that modulatestranscriptional activation of multiple nuclear receptors including orphan receptorsSF1 and ERR alpha 1. Mol. Endocrinol., 14: 986 –998, 2000.
28. Bernstein, F. C., Koetzle, T. F., Williams, G. J. B., Meyer, E. F., Brice, M. D.,Rodgers, J. R., Kennard, O., Shimanouchi, T., and Tasumi, M. The Protein DataBank: a computer-based archival file for macromolecular structures. J. Mol. Biol.,112: 535 –542, 1977.
29. Srinivasan, N. and Blundell, T. L. An evaluation of the performance of anautomated procedure for comparative modelling of protein tertiary structure.Protein Eng., 6: 501–512, 1993.
30. Rarey, M., Kramer, B., Lengauer, T., and Klebe, G. A fast flexible dockingmethod using an incremental construction algorithm. J. Mol. Biol., 261: 470 –489, 1996.
31. Kramer, B., Rarey, M., and Lengauer, T. Evaluation of the FLEXXincremental construction algorithm for protein-ligand docking. Proteins, 37:228 –241, 1999.
Molecular Cancer Research 991
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from

2003;1:981-991. Mol Cancer Res Masatomo Suetsugi, Leila Su, Kimberly Karlsberg, et al. to this work.
Masatomo Suetsugi and Leila Su contributed equallyNote:Grants ES08258 and CA44735.
National Institutes of Health11Estrogen-Related ReceptorsFlavone and Isoflavone Phytoestrogens Are Agonists of
Updated version
http://mcr.aacrjournals.org/content/1/13/981
Access the most recent version of this article at:
Cited articles
http://mcr.aacrjournals.org/content/1/13/981.full#ref-list-1
This article cites 29 articles, 11 of which you can access for free at:
Citing articles
http://mcr.aacrjournals.org/content/1/13/981.full#related-urls
This article has been cited by 11 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://mcr.aacrjournals.org/content/1/13/981To request permission to re-use all or part of this article, use this link
Research. on November 29, 2018. © 2003 American Association for Cancermcr.aacrjournals.org Downloaded from