folding mechanisms and intermediates for aggregation-prone native structures
DESCRIPTION
Folding Mechanisms and Intermediates for Aggregation-Prone Native Structures. Patricia L. Clark Department of Chemistry & Biochemistry University of Notre Dame, Notre Dame, Indiana Workshop on Biomolecules - Bedlewo, Poland May 14, 2004. The protein folding problem:. - PowerPoint PPT PresentationTRANSCRIPT

Folding Mechanisms and Intermediates for Aggregation-Prone
Native Structures
Patricia L. ClarkDepartment of Chemistry & Biochemistry
University of Notre Dame, Notre Dame, Indiana
Workshop on Biomolecules - Bedlewo, Poland May 14, 2004

The protein folding problem:
ensemble ofdenatured
states
native state
?
fold or aggregate?
?
misfolded, aggregated state
i

The folding of small globular single-domain proteins
Common proteins:
• 100-250 amino acids• Single structural domain• Rich in -helix structure• Monomeric
Common folding themes:
• Fast folding kinetics (sec-sec)• Few (if any) folding intermediates besides ‘molten globule’• Negligible competition from off-pathway aggregation
HEWL
RNaseA

Funnels for protein folding: energy landscapes
Folding funnel diagrams capture many of the features observed for the folding pathways of small, monomeric, single domain, helix-rich proteins

Benefits and caveats of energy landscapes/funnels:
• Folding funnels make it clear why proteins fold:- Energy difference between the unfolded ensemble and the native state
• Folding funnels have shifted focus to fast folding rates:- What is the barrier for folding?- What is the ‘speed limit’ for folding?
• But what about proteins that:(i) fold slowly, and/or(ii) are prone to aggregation?- How does this affect the energy landscape?

A folding funnel for many proteins in solution:

What kinds of proteins are prone to aggregation?
• Topology effects: Contact order? (D. Baker, U. Washington)
• Kinetic effects: Long-lived folding intermediates?
Plaxco et al. (1998) JMB 277:985

Non-local contacts = High contact order
contacts between residues in the primary sequence:
NEARBY FAR APART
A BBA
A
B A
B
ordering many more residues at once= selecting from more conformational
states-> How is aggregation avoidance
encoded?

Protein folding in the cell:
E. coli:• 200-400 mg/ml total protein• [nascent chains] = 30-50 M• ribosomes > 1/4 cell weight• chain synthesis ~ 20 aa/sec
David Goodsell: http://www.scripps.edu/pub/goodsell/illustration/public/
--> How are partially folded conformations protected from aggregation in this environment?

How do high CO structures form co-translationally?
in vitro:
BA
A
B
in vivo:
A
• What conformations does A adopt before B appears?• How much native structure can be formed co-translationally?
ribosome
ordering many more residues at once= selecting from more conformational
states-> How is aggregation avoidance
encoded?

Bordetella pertussis P.69 pertactin
• 60 kDa, single domain -helix• All parallel -sheet: no local contacts• Average rung-to-rung contact distance: 34 amino acids• No Cys, cofactors, etc.• C-terminal 59 residues disordered in structure; can be deleted with no effect on folding or stability
Cross-sectionof 7 central rungs(residues 140-357)

Spacefilling model of pertactin backbone structure
• Long loops are clustered on one face of structure• -helix backbone is remarkably regular

Pertactin far-UV CD spectra, thermal denaturation
• Three-state thermal unfolding• Partially folded state populated at 70ºC• 1.5 uM pertactin in 50 mM phosphate pH 8.8
Mirco Junker

Pertactin tryptophan fluorescence spectra: N and D
• Seven tryptophan residues (some solvent exposed) in native -helix structure, plus one in C-terminus• 0.5 uM pertactin in 50 mM TRIS pH 8.8, 25ºC
N
D
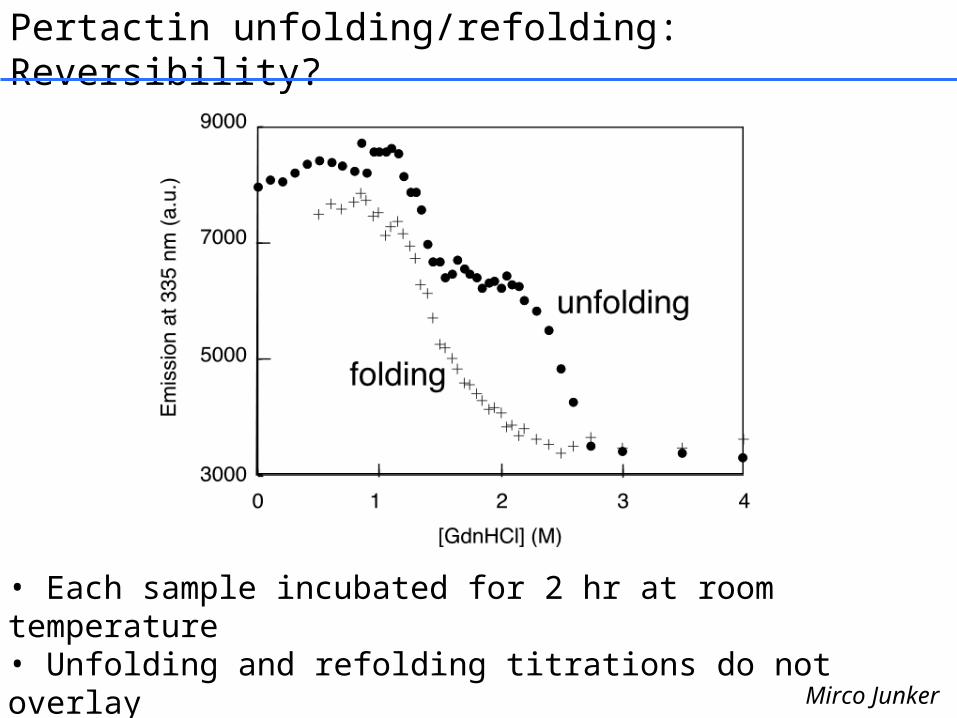
Pertactin unfolding/refolding: Reversibility?
• Each sample incubated for 2 hr at room temperature• Unfolding and refolding titrations do not overlay• No aggregation …microscopic reversibility?
Mirco Junker

Pertactin refolding IS reversible, but very slow:
• Similar results with urea, and when monitored by CD• GH2O = 46 kJ/mol (N-I) and 55 kJ/mol (I-D)• Partially folded structure forms extremely slowly• Origin of slow folding?
Mirco Junker

Models for the partially folded structure
• Trp fluorescence is halfway between N and D spectra• Half folded, Half unfolded…• Or: Half-folded?
Half folded/Half unfolded? Half-folded?

Testing the models: limited proteolytic digestion
Native pertactin:• Protease K resistant• Eventually degraded to 37 & 29 kDa fragments
Partially folded statein 1.4 M GdnHCl:• Less protease K resistant• Degraded to 29 kDa fragment• Stepwise: rung by rung?
Kelli Whiteman

MALDI-TOF mass spectrum of intact fragment
Proteinase K-resistant fragment:• harsher digestion results in 21 kDa band by SDS-PAGE, MALDI
Kay Finn & Elizabeth Klimek

MALDI-TOF: Tryptic digest of 21 kDa band
Trypsin digestion, followed by MALDI-TOF:• no fragments larger than 4 kDa• several peaks map to unique fragments
Kay Finn & Elizabeth Klimek

Identifying the partially folded structure
Mapping tryptic peptides onto the pertactin native structure:• RGD/PRR loop = red/blue (residues 226-262)• fragments cover residues 351-388, 395-435, 438-475, 480-509
Mirco Junker & Kay Finn
N C

Mapping pertactin slow folding/unfolding kinetics
• What occurs prior to 2 hr? How long does unfolding take?• How many events between 2 hr and 3 weeks?• How protect chromophores from bleaching/degradation?
Mirco Junker
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
30 min unfolding
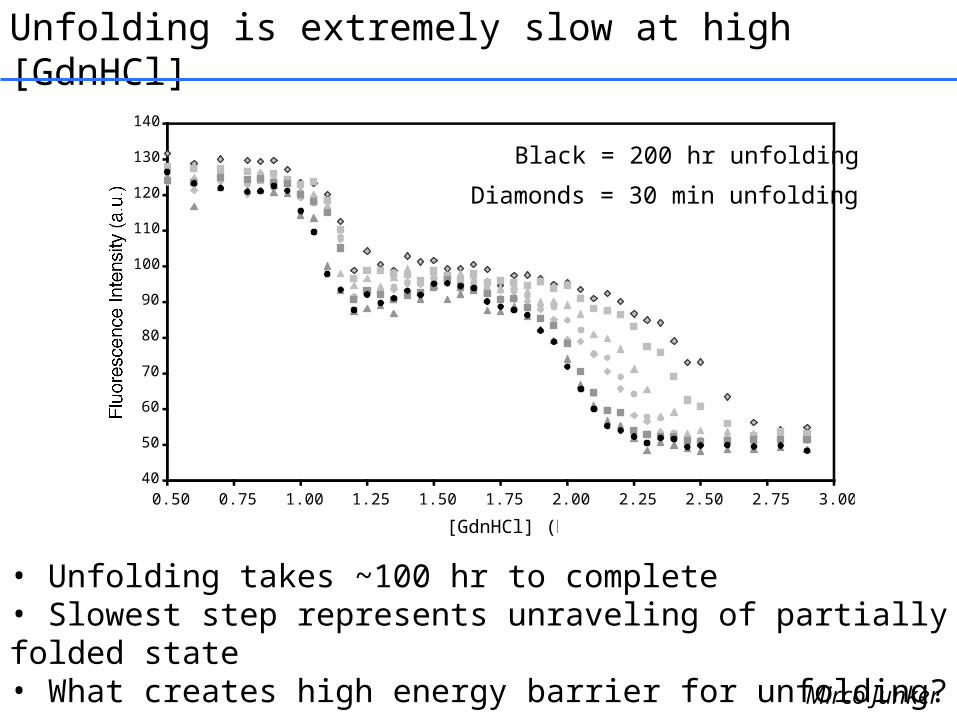
Unfolding is extremely slow at high [GdnHCl]
Mirco Junker
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 1 hr unfolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 2 hr unfolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 3 hr unfolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 4 hr unfolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 10 hr unfolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 100 hr unfolding
• Unfolding takes ~100 hr to complete• Slowest step represents unraveling of partially folded state• What creates high energy barrier for unfolding?
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 200 hr unfolding
Diamonds = 30 min unfolding

Spacefilling models of pertactin backbone structure
• -helix backbone is remarkably regular• Long loops are clustered on one face of structure

Refolding is even slower!
Chris Schuster & Katie O’Sullivan
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
30 min refolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 2 hr refolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 4 hr refolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 10 hr refolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 24 hr refolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 76 hr refolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 216 hr refolding
40
50
60
70
80
90
100
110
120
130
140
0.50 0.75 1.00 1.25 1.50 1.75 2.00 2.25 2.50 2.75 3.00
[GdnHCl] (M)
Fluorescence Intensity (a.u.)
Black = 312 hr refolding
Diamonds = 30 min refolding
• Refolding occurs over >200 hr• 0.5 M: fast events en route to native structure: HØ collapse?• 1.5 M: slow folding: conformational search?

Pertactin slow refolding kinetics:
• Refolding at 1.5 M GdnHCl; monitored by Trp fluor. emission• Multiple slow components Chris Schuster
40
45
50
55
60
65
70
75
80
85
90
0 20 40 60 80 100 120 140 160 180 200
Time (hr)
Fluorescence Intensity (a.u.)
Unfolded

Pertactin slow refolding kinetics:
• Refolding at 0.5 M GdnHCl; monitored by Trp fluor. emission• Fast and slow components Chris Schuster
40
60
80
100
120
140
0 20 40 60 80 100 120 140 160 180 200
Time (hr)
Fluorescence Intensity (a.u.)Unfolded

Pertactin slow refolding kinetics:
• Refolding at 0.5 M GdnHCl; monitored by Trp fluor. emission• Fast and slow components Mirco Junker

Slow formation of the partially folded structure:
Large conformational searchto form the native -helix ?
Fast formation of trapped, non-native structure ?
OR:

A folding funnel for many proteins in dilute solution:

Summary & Future directions
• Pertactin folding/unfolding is reversible, but equilibrium established very slowly
--> Large energy barrier to form partially folded state
--> A ‘template’ for -helix rungs?--> Selecting between energetically similar folded
andmisfolded states?
• Slow step at intermediate concentrations involves forming structure in C-terminal half of -helix
--> What parallel -sheet elements initiate folding?--> What rungs are more stable than others? Why?
• What cellular components regulate pertactin folding in vivo?

Acknowledgements
Thomas Clarke Neil Isaacs, U. GlasgowMichael EvansMirco Junker Andre Palmer, NDKrastyu UgrinovChris SchusterKatie O’Sullivan Bill BoggessElizabeth Klimek ND Mass Spec FacilityKelli Whiteman Kay Finn
NSF • AHAClare Boothe Luce Program, Henry Luce Foundation
University of Notre Dame

Pertactin partially folded state is monomeric
• Static light scattering detection of gel filtration eluate:
Kay Finn