fueling the heart: multiple roles for cardiac metabolism
TRANSCRIPT

1157
Fueling the Heart: Multiple Roles for
Cardiac MetabolismHeinrich Taegtmeyer
Key Points
• The heart requires an uninterrupted supply of energy for contraction, and this energy is derived from oxidative metabolism of fuels. The product of oxidative metabo-lism is adenosine triphosphate (ATP).
• The heart makes energy by oxidative phosphorylation of adenosine diphosphate (ADP).
• In the postprandial state, and under resting conditions, long chain fatty acids and glucose are the main fuels for respiration.
• Inborn errors in myocardial fatty metabolism are increas-ingly recognized as causes of cardiomyopathy and sudden death in children.
• Myocardial metabolism is an integral part of the function of the heart, as both a consumer and a provider of energy.
• Energy transfer is impaired in the failing heart.
Few cardiologists consider the heart a metabolic organ (the biochemistry of myocardial ischemia is a notable exception). Yet, energy substrate metabolism provides the energy cur-rency needed for normal contractile function of the heart, and metabolic dysregulation is considered both cause and consequence of contractile dysfunction of the heart. This chapter reviews several metabolic principles that are relevant for the practicing cardiologist and are useful in the diagnosis and treatment of heart disease.
Like any organ of the mammalian body, the heart con-sists of a complex system of interactive proteins, purine bases, energy-providing compounds, membranes, and signal molecules, all of which are in a constant state of fl ux. In this
dynamic system, the heart has retained its ability to adapt to a multitude of environmental changes, either by acutely changing fl ux through existing metabolic pathways to main-tain its energy supply,1,2 or by chronically altering the syn-thesis of specifi c proteins of cardiac structure, function, and metabolism.3–8
Between Gene Expression and Energetics: Fuel Metabolism in Perspective
The heart requires an uninterrupted supply of energy for contraction. This energy is derived from the oxidative metab-olism of fuels. Fuel metabolism, in turn, is regulated by the expression of enzymes and controlled by the energy require-ments of the heart. The principle of metabolic fl ux has long been recognized. Early in the 20th century, substrate metab-olism became the subject of intense research.9
The product of oxidative metabolism is adenosine tri-phosphate (ATP). Like in any other organ of the body, in the heart it is impossible to separate function from metabolism. As shown in the simple scheme (Fig. 50.1), a decrease in the fl ux of metabolic energy (e.g., with ischemia) results in a decrease in ATP production and, consequently, a decrease in contractile function. In contrast, an increase in contractile function (e.g., with inotropic stimulation) results in an increase in ATP turnover and an increase in oxygen con-sumption, which is a measure of energy utilization by the heart.
By measuring arteriovenous differences in the heart–lung preparation of the dog or in the human heart through
50
Between Gene Expression and Energetics: Fuel Metabolism in Perspective . . . . . . . . . . . . . . . . 1157
Heart Muscle: Consumer and Provider of Energy . . . . 1158Metabolic Pathways: The Highways of
Energy Transfer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1159Substrate Competition and Metabolic Flexibility. . . . . 1160Tracing Metabolic Pathways: Molecular Imaging
of the Heart. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1161
Metabolism of the Major Energy-Providing Substrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1162
Metabolic Signals of Cardiac Growth and Gene Expression. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1167
The Metabolic Syndrome . . . . . . . . . . . . . . . . . . . . . . . . 1167Metabolic Targets to Improve Cardiac Function . . . . . 1169Summary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1169
CAR050.indd 1157CAR050.indd 1157 11/24/2006 12:14:10 PM11/24/2006 12:14:10 PM

115 8 c h a p t e r 5 0
cannulation of the coronary sinus, early investigators10,11 have laid the foundation for our current understanding of myocardial substrate metabolism. The more recent reviews survey the knowledge gained through isolated heart prepara-tions, isolated cell preparations, and isolated cell organ-elles.5,12–17 Because factual information can be gleaned from these reviews, only principles of cardiac metabolism as they may be relevant to the patient with heart disease are reviewed here.
Since the classical biochemical era, our understanding of fuel metabolism in the heart has been greatly aided by intro-duction of new methods to assess metabolic activity in the intact, beating heart, such as nuclear magnetic resonance spectroscopy or magnetic resonance imaging (MRI), or in the heart in vivo, such as positron emission tomography (PET). A major feature of these new technologies is that they may permit a distinction between heart muscle that is reversibly dysfunctional and heart muscle that is irreversibly damaged.
Heart Muscle: Consumer and Provider of Energy
The goal of cardiac metabolism is to maintain a dynamic state of equilibrium for effi cient energy transfer in this highly specialized organ. In so doing, heart muscle converts chemical energy into physical energy. The physical energy of the heart consists of pump work (mechanical energy) that, in turn, distributes energy in the form of substrates and oxygen both to the heart itself and to the rest of the body. Here two important concepts must be considered. First, the heart is a “hot spot” of metabolic activity, continuously making and breaking ATP, the chemical energy available for conversion to mechanical energy at the contractile site. The greater the work output, the higher the rate of ATP turnover, the higher the rate of oxygen consumption, and the higher the rate of substrate utilization (Fig. 50.1). Second, when the
heart’s ability to convert chemical into mechanical energy is impaired for any reason,18 the consequences manifest themselves as functional and metabolic derangements in the rest of the body, commonly referred to as heart failure. Because heart failure is a systemic disease that begins and ends with the heart, understanding the regulation of energy transfer in the heart provides important clues for under-standing and treating heart failure.19
The rate of ATP turnover in the heart is greater than the rate of ATP turnover in any other organ of the body, and this is often underestimated. A simple calculation, based on mea-surements of myocardial oxygen consumption, indicates that during the course of a 24-hour period, the human heart produces (and uses) 5 kg of ATP (i.e., more than 10 times its own weight and 1000 times the amount of ATP stored in the heart). Another comparison comes to mind. Although the human heart makes up only 0.5% of total body weight, it claims 4% of total cardiac output and 10% of the body’s oxygen consumption. Lastly, it is important to remember that it is the rate of energy turnover, and not the tissue content of ATP, that determines myocardial energy metabolism.14,20–22
The heart makes energy by oxidative phosphorylation of adenosine diphosphate (ADP). Because of the high oxygen requirement, it is not surprising that the capillary density in heart muscle is far greater than in other organs (2500 capil-laries/mm2 vs. 400 capillaries/mm2 in skeletal muscle)23 and that heart muscle cells are fi lled with mitochondria, the cell organelles possessing the enzymes of oxidative metabolism (Fig. 50.2). A close correlation exists among mitochondrial volume fraction, heart rate, and total body oxygen consump-tion, with mitochondrial volume fractions ranging from 25% in humans to 38% in mice.24 Heart muscle mitochondria are not only abundant, they also contain a far greater number of
FIGURE 50.1. Adenosine triphosphate (ATP) cycle. Metabolism and function are linked through the synthesis and utilization of ATP. ADP, adenosine diphosphate; Pi, inorganic phosphate.
ADP+Pi ATP
ADP+Pi ATP
Fuelmetabolism
Contractilefunction
FIGURE 50.2. Ultrastructure of normal dog myocardium. The transmission electron micrograph shows, in addition to the regu-larly arranged contractile elements (sarcomeres), an abundance of mitochondria. Mitochondria are characterized by densely arranged cristae and constitute about 25% of the cell volume. A direct, posi-tive correlation exists between the number of mitochondria, resting heart rate, and myocardial oxygen consumption (×1800.)
CAR050.indd 1158CAR050.indd 1158 11/24/2006 12:14:10 PM11/24/2006 12:14:10 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 115 9
cristae (the location of respiratory chain enzymes) than do mitochondria of other organs, such as liver, brain, or skeletal muscle.25
Energy transfer in biologic systems obeys the fi rst and second laws of thermodynamics as defi ned by Helmholtz in 1845. The fi rst law of thermodynamics states that within a closed system, energy can only be converted from one form into another. The second law of thermodynamics states that a process occurs spontaneously only if it is associated with an increase in randomness (or entropy) of the system. Defi n-ing the bewildering array of metabolic pathways in the cell depicted in wall charts and textbooks is the result of many years of painstaking biochemical research.
Metabolic Pathways: The Highways of Energy Transfer
In heart muscle, complex chemical reactions proceed rapidly at a relatively low temperature and at low substrate concen-trations through enzyme-catalyzed pathways, an essential part of which are cycles (e.g., the citric acid cycle, the ATP cycle), because cycles have evolved as the most effi cient form of energy transfer.13,26 In addition, a few defi nitions may be useful.
A metabolic pathway is defi ned as a series of enzyme-catalyzed reactions beginning with a fl ux-generating step (usually a reaction catalyzed by a nonequilibrium reaction or transport of the metabolite across a membrane) and ending with the removal of a product.27,28 Characteristic of most metabolic pathways is that once fl ux has been initiated, there is a rapid and concerted response of the entire pathway. In this system of fl ux, metabolite levels control enzyme activi-ties and, in turn, enzyme activities control metabolite levels. It is important to distinguish between control and regulation of metabolism. Metabolic control is the power to change the state of metabolism in response to an external signal, whereas metabolic regulation is geared toward maintaining a con-stant internal state.29 In such a system, large changes in the fl ux through metabolic pathways correspond to only very small changes in myocardial metabolite concentration.30 Regulatory sites of metabolism, or pacemaker enzymes,31 may become targets for the manipulation of metabolism with drugs.32,33
In the heart, the purpose of most enzyme-catalyzed reac-tions is catabolic. Substrates of high potential energy are broken down to products of lower potential energy. Synthetic or anabolic reactions, such as those serving protein, glyco-gen, or triglyceride synthesis, are quantitatively of lesser importance. In other words, heart muscle is endowed with an effi cient system of energy transfer that liberates energy locked in chemical bonds by the generation of reducing equivalents and their reaction with molecular oxygen in the mitochondrial electron transport chain. The main purpose of intermediary metabolism in normal heart muscle, there-fore, is the production of reducing equivalents for ATP syn-thesis, in the course of which fuels are turned into carbon dioxide and water.
The breakdown of substrates can be divided into three stages (Fig. 50.3). The fi rst stage consists of substrate uptake by the cell and of the metabolic pathways leading to acetyl
coenzyme A (acetyl-CoA). The second stage consists of the oxidation of acetyl-CoA in the Krebs cycle. The third stage consists of the reaction of reducing equivalents with molecu-lar oxygen in the respiratory chain, where electron transfer is coupled to rephosphorylation of ADP to ATP. Adenosine triphosphate production is tightly coupled to ATP utiliza-tion, and in the normal heart substrate oxidation is also tightly coupled to cardiac work.1,34,35 An acute increase in workload of the isolated working rat heart results in an acute increase in myocardial oxygen consumption (Fig. 50.4, mea-sured) and CO2 production (Fig. 50.4, predicted), and the heart switches from fatty acids (oleate) to carbohydrates (glycogen, glucose, and lactate) as the main fuel for respiration (Fig. 50.5). Reduced fl avin adenine dinucleotide (FADH2) and reduced nicotinamide adenine dinucleotide (NADH) are gen-erated, and the reducing equivalents enter the electron trans-port chain, producing an electrochemical gradient across the inner mitochondrial membrane that drives ATP synthesis in the presence of molecular oxygen.
In isolated mitochondria the rate of ATP production is driven by the extramitochondrial concentration of ADP (state 3 respiration). The exact mechanism by which respira-tion is coupled to energy expenditure in vivo, however, is not known.36–38 But the effi cacy of oxidative phosphorylation for energy production is well established. For example, 1 mol of glucose, when oxidized, yields 36 mol of ATP, whereas the same amount of glucose yields only 2 mol of ATP when metabolized to lactate under anaerobic conditions.
There are two further aspects of cardiac metabolism that are often overlooked: the nutrition of the heart and endogenous substrates. Nutritional experiments (in the
Substrateuptake
Lactate Glucose Free fatty acids
TriglyceridesGlycogenG-6-P
GLUT
PDC
CO2 FADH2
H2O½O2
Lactate Pyruvate
Pyruvate β Oxidation
β-MHCα-MHC
FFAACS
CPT
Acyl-CoA
NAD+
NADH H+
NADH H+
Ca2+
Ca2+
Respiratory chain
Acyl-CoA
FAD
ADP + Pi
ADP + Pi
ATP
ATP
Acetyl-CoA
Krebscycle
Oxidation
Energyutilization
ATPADP ATP
ADP
ATP
ADP SERCASarcoplasmicreticulum
FIGURE 50.3. The main fuels for respiration in heart muscle uptake and oxidation of glucose and of long-chain fatty acids are tightly regulated to meet the energy needs for contraction of the heart. Acetyl-CoA, acetyl coenzyme A; Acyl-CoA, acyl-coenzyme A; ADP, adenosine diphosphate; ATP adenosine triphosphate; CPT, carni-tine-palmitoyl transferase; FAD, fl avine adenine dinucleotide; FADH2, fl avine adenine dinucleotide, reduced form; FFA, free fatty acid; G-6-P, glucose-6-phosphate; GLUT, glucose transporter; MHC, myosin heavy chain; NAD+, nicotinamide adenine dinucleotide; NADH, nicotinamide adenine dinucleotide, reduced form; Pi, inor-ganic phosphate; SERCA, sarcoendoplasmic reticulum ATPase. See text for further detail.
CAR050.indd 1159CAR050.indd 1159 11/24/2006 12:14:10 PM11/24/2006 12:14:10 PM

116 0 c h a p t e r 5 0
levels in the blood is still debated.40 Furthermore, the focus on cholesterol has shifted attention away from the fact that the heart requires large amounts of foodstuffs to meet its energy requirements. Both quantity and quality of these foodstuffs vary greatly in the course of a day. Metabolically speaking, the heart does not differ from the body as a whole. It is an omnivore (i.e., an organ that has developed or retained its ability to oxidize a variety of different substrates).41 The dietary advice for feeding the heart is the same as for feeding the body. Just as patients should not restrict fat and eat unre-stricted carbohydrates, patients should not overly restrict carbohydrates and eat unrestricted amounts of fat.42
The heart stores endogenous substrates such as glycogen and triglycerides in response to changes in the dietary state43,44 and the heart continuously synthesizes and degrades its own constituent proteins,45,46 a process that requires energy47,48 and is signifi cantly slowed by myocardial ischemia.
Substrate Competition and Metabolic Flexibility
In the postprandial state and under resting conditions, long-chain fatty acids and glucose are the main fuels for respira-tion (Fig. 50.6, top). Fatty acids, glucose, lactate, ketone bodies, and, under certain circumstances, even amino acids compete with one another as fuels for respiration. As an omnivore, the heart readily switches from one substrate to another when it is stressed and carbohydrates become the main fuel for respiration (Fig. 50.6, bottom). Metabolic fl exi-bility is an inherent property of the normal heart. The
90
80
Epi
neph
rine
70
60
Measured
Predictedfrom oxidation ofExogenous lipid +Endogenous lipid +Exogenous glucose +Endogenous glucose +Exogenous lactate +Pyruvate releaseO
xyge
n co
nsum
ptio
n, m
easu
red
pred
icte
d (μ
mol
.min
/g d
ry w
t.)
50
40
3050
Perfusion time (min)
55 60 65 70 75
FIGURE 50.4. Measured and predicted oxygen consumption in the isolated working rat heart. Predicted values are calculated from the sum of the measured oxidation rates of glucose, glycogen, lactate, oleate, triglycerides, and release of pyruvate.
4.5
4.0
3.5
2.5
1.5
0.5
3.0
2.0
1.0
0.0
–0.550 55 60
Perfusion time (min)
Oxi
datio
n ra
te (
μmol
/min
/g d
ry w
t.)
70
Glycogen
Glucose
Oleate
Lactate
Epi
neph
rine
7565
FIGURE 50.5. Rates of substrate oxidation following inotropic stimulation of the isolated working rat heart. Note the switch from oleate (fatty acids) to glycogen, glucose, and lactate as primary fuels for respiration.
microorganism Neurospora crassa) are at the beginning of the fundamental discovery that genes provide the blueprints for enzymes.39 Today, nutrition is often confi ned to a single issue, and that is obesity and cholesterol. “Heart-healthy” diets have also focused heavily on cholesterol because of its role in the development of coronary artery disease, but even the correlation between dietary cholesterol and cholesterol
Normal
Stress
Fatty acids
Fatty acids
Malonyl-CoA
Glucose
Glucose
Malonyl-CoA
FIGURE 50.6. Metabolic fl exibility. During periods of stress, the heart shifts from fatty acid oxidation to glucose oxidation. An increase in malonyl-CoA inhibits fatty acid transport into the mito-chondria, and promotes the oxidation of glucose.
CAR050.indd 1160CAR050.indd 1160 11/24/2006 12:14:10 PM11/24/2006 12:14:10 PM
f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 1161
relative predominance of a fuel for respiration depends on the arterial substrate concentration (which in the case of fatty acids, lactate, and ketone bodies can vary over a wide range) and on hormonal infl uences, workload, and oxygen supply. Stated simply, for a given environment the heart oxidizes the most effi cient substrate. When Bing11 cannulated the coro-nary sinus and measured aortocoronary sinus differences in substrate concentrations across the heart, he observed a pro-portional relationship between arterial substrate concentra-tion and substrate uptake by the heart under steady-state conditions, which changed with changes in the physiologic environment. Subsequent work by others49 has established that the relative contribution of a substrate to the fuel for respiration depends on the physiologic state of the entire body. In the stressed heart in vivo the effi ciency of glucose as substrate exceeds the effi ciency of fatty acids as substrate by as much as 40%.50 The normal mammalian heart readily switches from one substrate to another.
Tracing Metabolic Pathways: Molecular Imaging of the Heart
Tracing of metabolic pathways can be accomplished qualita-tively owing to the development of new, nondestructive imaging techniques such as nuclear magnetic resonance spectroscopy and PET. Both techniques permit the assess-ment of regional metabolic processes in the beating heart both in vitro and in vivo.51–58
Analysis of energy-rich phosphates in the beating heart by nuclear magnetic resonance spectroscopy of phosphorus 31 (31P) lends further support to the view that over a relatively wide range of work output, tissue content of ATP is not cor-related with ATP turnover, as assessed by oxygen consump-tion or contractile performance of the heart.59 The recent adaptation of isotopomer analysis of carbon 13 (13C) natural abundance or 13C-labeled compounds allows the analysis of fl ux through specifi c pathways, especially the Krebs citric acid cycle and glycogen turnover, to be studied quantitatively as serial spectra are obtained.58,60–62
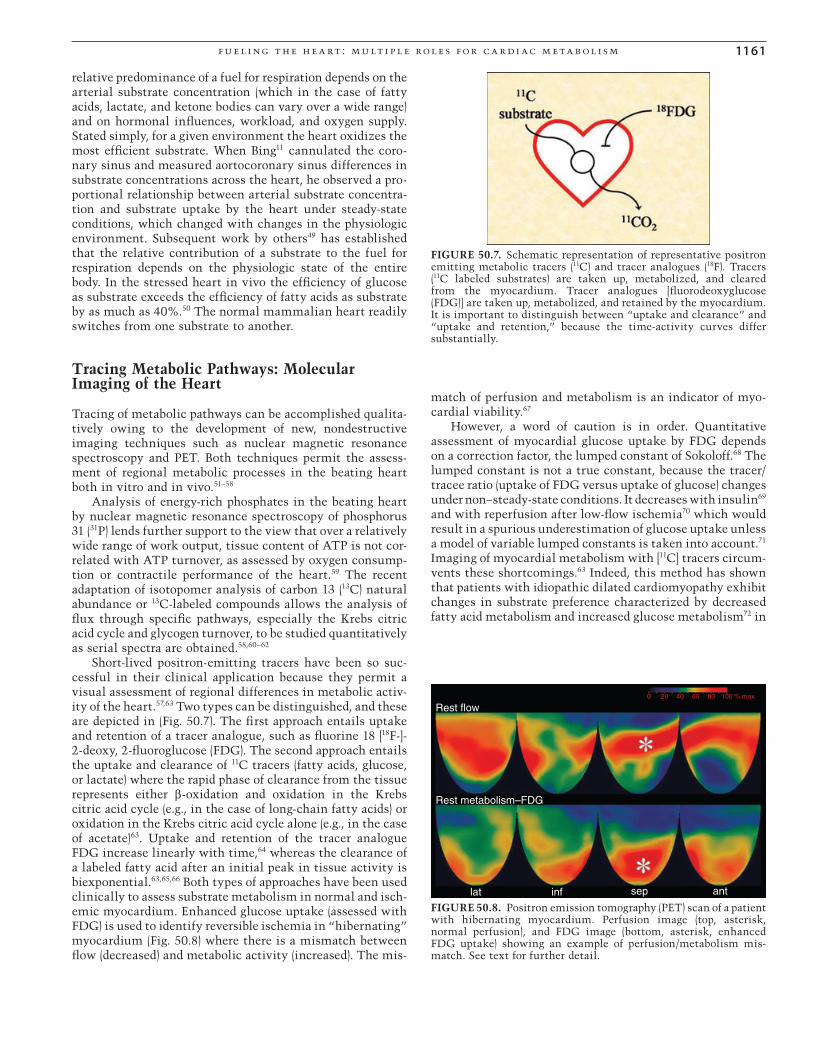
Short-lived positron-emitting tracers have been so suc-cessful in their clinical application because they permit a visual assessment of regional differences in metabolic activ-ity of the heart.57,63 Two types can be distinguished, and these are depicted in (Fig. 50.7). The fi rst approach entails uptake and retention of a tracer analogue, such as fl uorine 18 [18F-]-2-deoxy, 2-fl uoroglucose (FDG). The second approach entails the uptake and clearance of 11C tracers (fatty acids, glucose, or lactate) where the rapid phase of clearance from the tissue represents either β-oxidation and oxidation in the Krebs citric acid cycle (e.g., in the case of long-chain fatty acids) or oxidation in the Krebs citric acid cycle alone (e.g., in the case of acetate)63. Uptake and retention of the tracer analogue FDG increase linearly with time,64 whereas the clearance of a labeled fatty acid after an initial peak in tissue activity is biexponential.63,65,66 Both types of approaches have been used clinically to assess substrate metabolism in normal and isch-emic myocardium. Enhanced glucose uptake (assessed with FDG) is used to identify reversible ischemia in “hibernating” myocardium (Fig. 50.8) where there is a mismatch between fl ow (decreased) and metabolic activity (increased). The mis-
match of perfusion and metabolism is an indicator of myo-cardial viability.67
However, a word of caution is in order. Quantitative assessment of myocardial glucose uptake by FDG depends on a correction factor, the lumped constant of Sokoloff.68 The lumped constant is not a true constant, because the tracer/tracee ratio (uptake of FDG versus uptake of glucose) changes under non–steady-state conditions. It decreases with insulin69 and with reperfusion after low-fl ow ischemia70 which would result in a spurious underestimation of glucose uptake unless a model of variable lumped constants is taken into account.71 Imaging of myocardial metabolism with [11C] tracers circum-vents these shortcomings.63 Indeed, this method has shown that patients with idiopathic dilated cardiomyopathy exhibit changes in substrate preference characterized by decreased fatty acid metabolism and increased glucose metabolism72 in
FIGURE 50.7. Schematic representation of representative positron emitting metabolic tracers (11C) and tracer analogues (18F). Tracers (11C labeled substrates) are taken up, metabolized, and cleared from the myocardium. Tracer analogues [fl uorodeoxyglucose (FDG)] are taken up, metabolized, and retained by the myocardium. It is important to distinguish between “uptake and clearance” and “uptake and retention,” because the time-activity curves differ substantially.
Rest flow
Rest metabolism–FDG
lat inf sep ant
0 20 40 60 80 100 % max
FIGURE 50.8. Positron emission tomography (PET) scan of a patient with hibernating myocardium. Perfusion image (top, asterisk, normal perfusion), and FDG image (bottom, asterisk, enhanced FDG uptake) showing an example of perfusion/metabolism mis-match. See text for further detail.
CAR050.indd 1161CAR050.indd 1161 11/24/2006 12:14:11 PM11/24/2006 12:14:11 PM

116 2 c h a p t e r 5 0
keeping with the constitutive expression of the fetal meta-bolic gene program in the failing human heart.73
Metabolism of the Major Energy-Providing Substrates
Why does the heart use different fuels to make energy? Why is it important to know metabolic pathways in detail? The following analogy may be useful to answer these questions. Metabolic pathways can be likened to a power grid, and the heart can be likened to a light bulb. Fuels are the various sources of energy (coal, water, natural gas, nuclear fi ssion) that are all converted to electricity. A simple explanation for this redundancy is that fuels are used whenever they are most readily available. All fuels are converted to the same form of energy. In short, for a given environment the heart uses the most effi cient source of energy, and this is the result of tight regulation in metabolic pathways.
Carbohydrates
Carbohydrate fuels for the heart are glucose, lactate, and glycogen. Although long-chain fatty acids are normally the predominant fuel for energy production in the mammalian heart, carbohydrates are the fuel for the fetal heart74 and for the stressed adult heart in the state of exercise.75 Glucose serves as the heart’s “backup” fuel. The reasons for such a “hierarchy” of fuels can be found in a number of observa-tions: (1) In the normal, that is, the nondiabetic, mammalian organism, glucose levels in the blood are tightly regulated at around 5 mM or (90 mg%). (2) When we exercise and blood lactate levels rise, lactate replaces all other substrates as fuel for respiration of the heart.75 (3) When the normal heart is stressed, it oxidizes fi rst glycogen, then glucose and lactate.1,76 (4) Glucose is an anaplerotic substrate for the Krebs cycle,35,77 and glucose is essential for the initiation of fatty acid oxida-tion in heart.78 Hypoglycemic newborns develop cardiomeg-aly and heart failure, which are both completely reversible with the administration of glucose.79 Furthermore, isotopic tracer studies have demonstrated that the normal human heart produces lactate at the same time it oxidizes lactate,1,80 and that it takes up glucose to form glycogen when in the fasted, resting state.77,81 It appears that a large portion of exogenous glucose is shunted fi rst into glycogen before it is oxidized.82,83 Glucose and lactate extraction by the heart in vitro and in vivo increases with an increase in workload, even in the presence of competing substrates.35,84 This obser-vation is of interest in view of the importance of pyruvate, which is the common metabolic product of glucose, glyco-gen, and lactate. The main point is that pyruvate provides both acetyl-CoA (C2 units) and oxaloacetate (C4 units) for the Krebs citric acid cycle.
Regulatory Sites of Glucose Metabolism
There are three energy-yielding stages of glucose metabo-lism: the glycolytic pathway leading to pyruvate, oxaloace-tate, and lactate; the Krebs cycle; and the respiratory chain. Each state is regulated by its own set of checkpoints, so that overall fl ux through the pathways (which may be assessed
externally on a second-by-second time scale with the glucose tracer analogue FDG)64 proceeds at a rate just suffi cient to satisfy the heart’s beat-to-beat needs for the fi nal product, ATP. We discuss the regulatory sites of metabolism in more detail because dysregulation of one of these steps can affect the rate and effi cacy of energy transfer and, hence, contractile function of the heart.
Glucose Transport and Phosphorylation
The fi rst major regulatory site of the glycolytic pathway is glucose uptake. Glucose uptake is stereospecifi c and satura-ble. It follows classical Michaelis-Menten kinetics, and early investigators already deduced that active glucose transport-ers are membrane-associated proteins. The transport of glucose occurs along a steep concentration gradient and is regulated by specifi c transporters.85 Stereospecifi city of the transporter for sugars of the carbon confi guration is not matched by the same degree of selectivity, and various tracer analogues, including 2-deoxyglucose and FDG, are trans-ported in the same way as glucose.
Glucose transport, the rate-limiting step in myocardial glucose utilization,86 is carried out by facilitative glucose transporters. The family of glucose transporters (GLUTs) is conserved over a wide range of organisms, suggesting a common evolutionary origin.87–90 GLUT-1 and GLUT-4 are the major glucose transporter isoforms expressed in the heart.73,91 GLUT-4, the insulin-sensitive transporter, is also found in skeletal muscle and in adipose tissue.87
Heart muscle is an insulin-sensitive organ.13 Insulin is considered an “endogenous cardioprotector”92 because, in addition to promoting glucose uptake, insulin has a multi-tude of cellular and hemodynamic effects.93 Recruitment of GLUT-4 from a microsomal cytosolic pool to the sarcolemma by insulin (or ischemia or adrenergic stimulation)94–97 increases the maximal velocity of glucose transport. α-Adrenergic stimulation uses the same signaling pathway as insulin to promote glucose uptake,98 whereas the effects of ischemia, β-receptor stimulation, and insulin on glucose uptake are additive.99
In addition to GLUT-4, the cardiomyocyte expresses the GLUT-1 transporter isoform, which is presumably indepen-dent of insulin regulation and predominates in fetal, hyper-trophied, atrophied, and failing myocardium.73,88,100 GLUT-1 is the fi rst gene whose transcription is dually stimulated in response to hypoxia and inhibition of oxidative phosphoryla-tion,101 and overexpression of GLUT-1 prevents the functional decline of the hypertrophied heart.102 Both transporters have a Michaelis’ constant (Km) for glucose (i.e., the concentration at which the rate of glucose transport is half maximal) that is in the range of plasma glucose concentrations under fasting conditions.103 The normal heart also expresses a low amount of GLUT-3, which has a Km below the normal plasma glucose concentration.104 A number of novel GLUTs have been identi-fi ed in heart muscle, including GLUT-8, GLUT-11, and GLUT-12.105 The role of these proteins in heart muscle is not yet defi ned.
Cardiac work and substrate availability, as well as plasma glucose and insulin concentrations, are important factors determining glucose uptake.106 For example, myocardial glucose uptake is also determined by the dietary state per
CAR050.indd 1162CAR050.indd 1162 11/24/2006 12:14:12 PM11/24/2006 12:14:12 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 116 3
se,107 by oxygen availability82,108,109 and by hormones, such as catecholamines, growth hormone, and cortisol, which, in addition to insulin and glucagons, may affect either Km or maximal velocity (Vmax) of glucose transport. The role of insulin signaling in the heart is complex and is intertwined with cardiac growth and development.110
Phosphorylation of glucose by hexokinase becomes rate limiting for glycolysis at high rates of glucose transport. Rates of glucose phosphorylation measured in vitro are more than twice as high as the maximal measured rates of glucose utilization by the heart at a physiologic workload and with glucose as the only substrate.111 Some evidence suggests that glucose, once inside the cell and phosphorylated, may pref-erentially enter the glycogen pool rather than the glycolytic pathway.13,83,112 The intracellular glucose concentration of heart muscle rises during starvation, in diabetes, and with the concomitant oxidation of fatty acids, ketone bodies, or lactate, which indicates inhibition of the phosphorylation step. This is most likely to be due to accumulation of glucose-6-phosphate.
Glucose-6-phosphate is the substrate for several distinct enzyme systems. These systems include (1) degradation via the Embden-Meyerhof pathway (also termed the glycolytic pathway when it entails metabolism of glucose to lactate only); (2) conversion to glycogen via the glycogen synthase reaction; (3) metabolism (oxidation) via the pentose-phosphate pathway, which yields ribose and the reduced form of nicotinamide adenine dinucleotide phosphate (NADPH)113; and (4) entry into the hexosamine biosynthetic pathway.114 The latter two pathways are of quantitatively lesser impor-tance in heart muscle than are the former two. Of note is that most of these pathways are not available to the tracer analogue FDG 6-phosphate, which may, however, become a substrate for glycogen synthesis.115 Hence, the commonly used tracer analogue FDG traces glucose uptake and phos-phorylation, but it lacks specifi city with regard to the meta-bolic fate of glucose inside the cell.
Glycogen
The study of the pathways of glycogen synthesis and degrada-tion in liver and muscle has contributed more to the under-standing of enzyme regulation and the molecular basis of hormone action than any other known system of metabolic control.116 It was in this system that the fi rst example for the control of enzyme activity by an allosteric regulator [activa-tion of phosphorylase by adenosine monophosphate (AMP)] was described,117 enzyme regulation by covalent modifi ca-tion was discovered,118 and the molecular basis of hormone action by signal transduction was elucidated through the discovery of cyclic adenosine monophosphate (cAMP).119 The role of glycogen in the heart is still not completely under-stood.120 The vast amount of glycogen in fetal cardiac muscle allows the heart to maintain its contractile activity in the face of severe hypoxia121 during birth. One may speculate that the high concentration of glycogen in the specialized conduction system of the heart, fi rst described by Aschoff122 at the beginning of the 20th century, may exert a protective effect against ischemia. Lastly, glycogen and glycogen phos-phorylase are also closely associated with the sarcoplasmic reticulum,123 and in skeletal muscle decreased glycogen
content is closely associated with a reduction in force, Ca2+ release from the sarcoplasmic reticulum, and contractile protein function.124
Glycogen synthesis and degradation occur in two sepa-rate pathways. The combined effects of protein phosphoryla-tion and dephosphorylation on glycogen synthase and phosphorylase provide an interlocking system by which hor-mones (such as epinephrine) and mechanical activity (through Ca2+) can control the net fl ux of glucose 1-P into and out of the intracellular glycogen stores.62,125 Epinephrine-induced cAMP formation promotes protein phosphorylation and simultaneously inhibits glycogen synthesis while stimulat-ing glycogen breakdown, whereas stimulation of protein dephosphorylation shifts the balance toward glycogen syn-thesis. Although the interaction of the enzymes glycogen breakdown and synthesis is more complex than this simple mechanism suggests,126 the concept of a continuous turnover of the myocardial glycogen pool is relatively easy to grasp.
Excessive rates of glycogen breakdown during ischemia are thought to be deleterious for the recovery of contractile function because of the accumulation of lactate. However, the heart subjected to low-fl ow ischemia continues to syn-thesize glycogen,127,128 and enhanced glycogen stores (and with them, high rates of glycogen breakdown) improve con-tractile function in the ischemic and reperfused myocar-dium,129–133 in the stressed myocardium,134 and in the ischemic heart.135–137 The role of glycogen in conveying ischemia toler-ance is seen most dramatically in amphibian myocardium, which is particularly rich in this compound.138
Glycolysis
Glucose is unique among substrates in its ability to provide a small, but signifi cant, amount of ATP through substrate-level phosphorylation in the glycolytic pathway. This occurs especially in the setting of hypoxia and ischemia when fl ux through the glycolytic pathway is enhanced, resulting in the formation of lactate as well as alanine.139 Other stimulants of fl ux through the glycolytic pathway are increases in cardiac work, either acutely with exercise,1,84 or chronically with pressure overload without140 or with hypertrophy.141,142 Both hypertrophy and atrophy are associated with increased glucose oxidation rates but decreased insulin responsiveness of the heart.143
Enhanced glycolytic fl ux lessens ischemic tissue damage.136,144 Although it is thought that the accumulation of glycolytic products may worsen the effects of ischemia145 and acute hyperglycemia may abolish ischemic precondi-tioning in vivo,146 provision of glucose together with insulin and potassium improves contractile function in the acutely ischemic, reperfused myocardium.147–149 The inotropic effect of insulin in the postischemic heart is additive to epineph-rine and coupled to enhanced glucose uptake.150
Glycolysis from extracellular glucose (and no other meta-bolic pathway) protects cardiac myocytes from hypoxic injury and subsequent apoptosis.151 The fi rst step committing glucose to the glycolytic pathway is 6-phospho-fructo-1-kinase (PFK-1), which catalyzes the phosphorylation of fruc-tose 6-phosphate to fructose 1,6-bisphosphate. Because of the complex allosteric regulation of PFK, this is a rate-limiting step (pacemaker enzyme) for glycolysis.152 Adenosine
CAR050.indd 1163CAR050.indd 1163 11/24/2006 12:14:12 PM11/24/2006 12:14:12 PM

116 4 c h a p t e r 5 0
triphosphate, citrate, and protons are negative allosteric effectors, whereas AMP and fructose 1,6-bisphosphate are positive effectors.153–155 Fructose 2,6-bisphosphate is the main activator of PFK-1 in normoxic heart.
Further down in the glycolytic pathway, the oxidation of the triose-phosphate glyceraldehydes 3-phosphate to 1,3-diphosphoglycerate couples in the energy-conserving step in the glycolytic pathway that leads to the nonoxidative forma-tion of ATP. The reaction is thought to be at near equilib-rium. However, under conditions of high cardiac work156 or ischemia,157 when PFK becomes strongly activated, glycolysis is controlled further downstream, at the triose-phosphate dehydrogenase step.
Pyruvate at the Crossroads: Oxidation Carboxylation, Reduction, and Transamination
Just as the fi rst committed intermediate of glycolytic pathway, glucose-6-phosphate, is at a branch point where several enzymes compete for it as substrate, so is the last committed intermediate, pyruvate. In heart muscle, pyru-vate can be reduced to lactate (which completes the glyco-lytic pathway), transaminated to alanine,139 carboxylated to oxaloacetate or malate,158,159 or, most importantly, oxidized to acetyl-CoA. Lactate and most of the alanine are formed in the cytosol by near-equilibrium reactions, and both metabo-lites may be washed out from the cell. In well-oxygenated, working heart muscle, however, the bulk of pyruvate enters the mitochondrion. A specifi c carrier exists for transport of pyruvate into the mitochondrial matrix, as it has been most clearly demonstrated with the inhibitor 4-hydroxy-α-cyanocinnamate.160
Inside the mitochondrial matrix, pyruvate can be either decarboxylated to acetyl-CoA or carboxylated to oxaloace-tate.159 The capture of metabolically produced carbon dioxide from the pyruvate dehydrogenase reaction to form oxaloace-tate provides one example of the effi cient use of one substrate supplying two precursors for citrate synthesis and effi cient recycling of carbon dioxide.
Oxidative decarboxylation of pyruvate assumes a central position in the regulation of fuel supply to the heart. A system of intricate control mechanisms governs both activa-tion and inactivation of the pyruvate dehydrogenase complex (PDC).161 The conversion of pyruvate to acetyl-CoA requires the sequential action of three different enzymes: pyruvate dehydrogenase, dihydrolipoyl transacetylase, and dihydroli-poyl dehydrogenase. The reaction also requires fi ve different coenzymes or prosthetic groups: thiamine pyrophosphate, lipoic acid, uncombined sulfhydryl coenzyme A (CoA-SH), fl avin adenine dinucleotide (FAD+), and nicotinamide adenine dinucleotide (NAD+). These enzymes and coenzymes are organized into a multi-enzyme cluster.
Much work was done in the 1970s on the regulation of PDC by covalent modifi cation through a phosphorylation and dephosphorylation cycle. Multisite phosphorylation of the pyruvate dehydrogenase component of the complex provides an indirect means by which the entire complex is regulated by the relative activities of the PDC kinase and phosphatase reactions. Like most mammalian tissues, heart muscle possesses both active (dephosphorylated) and inactive (phosphorylated) dehydrogenase. Most of the known effectors
of the phosphorylation-dephosphorylation cycle are also effectors of the kinase reaction. Of the metabolite pairs ATP–ADP, acetyl-CoA–CoA-SH, NADH–NAD+, and lactate–pyru-vate, the fi rst member either activates the kinase or serves as substrate, and the second member inhibits the enzyme. Ca2+ and Mg2+ both inhibit the kinase and activate the phos-phatase reaction (i.e., lead to PDC activation). The effects of fatty acids or ketone body oxidation are likely to be mediated by the increase in acetyl-CoA because of the primary effect or the inhibition-inactivation of PDC by fatty acids or ketone bodies in the acetyl-CoA–CoA-SH ratio.162 Conversely, an increase in cardiac work may inhibit the PDC kinase owing to a decrease in NADH, acetyl-CoA, and ATP, leading to activation of PDC.163
Fatty Acids
Fatty acids, esterifi ed as triglycerides, represent the body’s main fuel reserve. Because oxidation of long-chain fatty acids releases more than six times as much energy as the oxidation of an equal mass of glucose, fatty acids represent the pre-dominant energy reserve for the body and fuel for respiration in heart muscle. In spite of their preeminence as a source for energy, fatty acids are the only fuel capable of uncoupling oxidative phosphorylation.164,165 This mechanism lowers the effi ciency of fatty acids as energy substrates. In contrast to glucose and glycogen, fatty acids require oxygen for their catabolism.
The predominant forms of fatty acids in the bloodstream are the monounsaturated long-chain fatty acid oleate (C18:1) and the saturated fatty acid palmitate (C16:0). The pathway of long-chain fatty acid oxidation starts with the liberation of fatty acids from triglycerides and ends with the entry of acetyl-CoA into the citric acid cycle. Free fatty acids (FFAs) of any chain length cross the plasma membrane with the help of carrier proteins, which have been cloned and expressed in a variety of systems.166 In the cytosol, long-chain fatty acids are bound by a heart-specifi c fatty acid-binding protein (h-FABP),167–169 which ushers long-chain fatty acids from the cytosol toward the oxidation pathway.170 Bound fatty acids are activated on the outer mitochondrial membrane by the enzyme fatty acyl-CoA synthetase and esterifi cation with CoA-SH to form fatty-acyl coenzyme A (acyl-CoA). Metabolism of long-chain fatty acids continues with fatty acyl transfer to carnitine, transport into the mito-chondria in exchange for carnitine, reesterifi cation with CoA-SH, β-oxidation, and fi nally, oxidation of acetyl-CoA. A variable amount of FFA taken up by heart muscle is ester-ifi ed with glycerol in the cytosol to form triglycerides. Therefore, the fatty acyl-CoA formed on the outer mito-chondrial membrane has two possible fates: either oxidation via the citric acid cycle in the mitochondria or conversion to triglycerides in the cytosol. Which pathway is taken depends on the rate of transfer of long-chain fatty acyl-CoA across the inner mitochondrial membrane and on the rate of esterifi cation in the cytosol. Increased net triglycer-ide synthesis by the heart has been observed with starva-tion, diabetes, ischemia, and in heart failure.171 It is not known whether increased triglyceride levels are the result of increased rates of esterifi cation or a decreased rate of lipolysis.172
CAR050.indd 1164CAR050.indd 1164 11/24/2006 12:14:12 PM11/24/2006 12:14:12 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 116 5
Carnitine Palmitoyl Transferase, Malonyl-CoA
Before the acyl group can be oxidized it must be moved from the outer mitochondrial membrane into the mitochondrion. Transfer on long-chain fatty acyl-CoA into mitochondria is rate limiting for β-oxidation of long-chain fatty acids and requires a three-step membrane transport process. The fi rst step in this sequence is the transfer of the acyl group from CoA to carnitine, catalyzed by the enzyme carnitine palmi-toyl transferase I (CPT I).
The CPT I system is inhibited by physiologic concentra-tions of malonyl-CoA, the product of acetyl-CoA carboxyl-ation, and was fi rst characterized in the liver, where malonyl-CoA serves as a signal regulating the relative rates of fatty acid oxidation, ketogenesis, and triglyceride synthe-sis.173 In a beautiful system of checks and balances malonyl-CoA also is the fi rst step in the pathway for de novo fatty acid synthesis. Although the heart is not a lipogenic organ, it contains malonyl-CoA,174 and the inhibition of cardiac CPT I by malonyl-CoA has been well documented.175 A tissue-specifi c acetyl-CoA carboxylase has been identifi ed for heart muscle,176 and so has malonyl-CoA decarboxylase.177–179 There is at least indirect evidence that malonyl-CoA plays an impor-tant role in substrate competition between fatty acids and glucose. Malonyl-CoA levels are controlled by its rates of synthesis [via acetyl-CoA carboxylase (ACC)] and its rate of degradation [via malonyl-CoA decarboxylase (MCD)]. Because of the importance of malonyl-CoA for the regulation of fatty acid oxidation, the regulation of ACC and MCD has been intensely investigated. Specifi cally, ACC is inhibited by phosphorylation through AMP kinase, a protein kinase that is activated when ATP levels fall and AMP levels rise in the myocyte. This results in lower malonyl-CoA levels and increased rates of fatty acid oxidations; hence, AMP kinase has also been termed the “fuel gauge” of the cell.180 Also, in the isolated, perfused, working rat heart, glucose suppresses the oxidation of oleate.35 It is also worth noting that ischemia reverses the inhibition of CPT I by malonyl-CoA, most prob-ably through a modifi cation in protein folding, thus making available increased amounts of palmitoyl-carnitine for oxida-tion on reperfusion.181
The carnitine-acyl unit traverses the inner mitochondrial membrane and is transferred to CoA-SH inside the mito-chondrial matrix by a second carnitine acyl-CoA transferase (CPT II) located in the inner surface of the inner mitochon-drial membrane. Once inside the mitochondria, acyl-CoA is committed to oxidation by the system of β-oxidation. The β-oxidation pathway is controlled by fl uctuations in the concentrations of substrates (acyl-CoA, NAD+, and FAD+)182 or, expressed in more physiologic terms, by workload and oxygen supply of the heart. It is not yet clear how the rate of oxidation of FFA in the intact heart is related to citric acid cycle activity and the rate of oxidative phosphorylation. However, it is clear that fatty acid oxidation is arrested by ischemia and that the accumulation of long-chain acyl-CoA esters in ischemia inhibits the adenine nucleotide translo-case in mitochondria.183 This inhibition provides a possible explanation for contractile dysfunction.
In the liver, and most probably also in heart muscle, CPT I is the regulatory site for fatty acid oxidation, with high concentrations of malonyl-CoA acting as an inhibitor of the
enzyme during the carbohydrate-fed state. To explore the gene regulatory mechanisms involved in the metabolic control of long-chain fatty acid oxidation in the heart, the expression of muscle-type CPT-1 (mCPT-1) was characterized in primary cardiac myocytes after incubation with oleate, which regulates mCPT-1 expression via the peroxisome proliferators-activated receptor-α (PPAR-α)184; PPAR-α is a nuclear receptor family with multiple functions that is cur-rently attracting the attention of many investigators and the pharmaceutical industry.185 Malonyl-CoA decarboxylase (MCD) is transcriptionally regulated by PPAR-α,179 and MCD inhibition protects the ischemic heart most likely by inhibit-ing fatty acid oxidation.186
Transcriptional Regulation of Myocardial Fatty Acid Metabolism
One way by which the heart responds to increased plasma fatty acid levels is by increasing the expression of various proteins involved in fatty acid utilization.179,184,187–189 The mechanism by which fatty acids activate the transcription of these genes is through activation of the nuclear receptor PPAR-α, which heterodimerizes with 9-cis retinoic acid receptor (RXR) and activates the transcription of genes whose promoter contains the PPAR response element (PPRE) (Fig. 50.9). Of the cofactors that bind to the PPAR-α/RXR het-erodimer, PCG-1α is highly expressed in the heart190,191 and plays a key role in mitochondrial biogenesis.7
Defective Fatty Acid Metabolism as a Cause of Heart Failure
Inborn errors in myocardial fatty acid metabolism are increasingly recognized as causes of cardiomyopathy and sudden death in children.192–194 Defects at nearly every step of the fatty acid oxidation pathway have been identifi ed.195 In recent years the advent of tandem mass spectrometry has greatly advanced the screening for metabolic cardiomyopa-thies.194 The analysis of acylcarnitines in a dried blood spot rapidly provides a diagnosis in patients in whom an inherited
Long-chainfatty acids 9-cis retinoic
acid
Coactivators
RXR
PPAR-α
AGGNCA A AGGTCA Target gene
PPRE mRNA
FIGURE 50.9. Transcription of PPAR-α regulated genes. Peroxi-some proliferators-activated receptor-α (PPAR-α) heterodimerizes with the 9-cis retinoic acid receptor, and interacts with different coactivators to begin transcription of PPAR-α regulated genes. See text for further detail.
CAR050.indd 1165CAR050.indd 1165 11/24/2006 12:14:12 PM11/24/2006 12:14:12 PM

116 6 c h a p t e r 5 0
metabolic disease is considered, and algorithms exist for the laboratory diagnosis of metabolic diseases presenting with cardiomyopathy.196 The most common group of inherited defects involves the acyl-CoA dehydrogenase enzyme family, which catalyzes the initial reactions in mitochondrial β-oxidation of fatty acids.197,198 In general, enzymatic defects involving long- and very-long-chain fatty acids (C14 or greater) or defects in cellular carnitine import of enzymes involved in the carnitine shuttle cause more severe clinical diseases, including cardiomyopathy, heart failure, and sudden death than with defects involving the metabolism of shorter-chain fatty acids.199 Impaired β-oxidation and accumulation of tri-glycerides in cardiomyocytes are hallmarks of contractile failing of the heart from obese Zucker rats.200 In Zucker dia-betic fatty rats, lipotoxicity and contractile dysfunction are reversed by the administration of insulin sensitizing agents.201,202
Clinical manifestations of inherited defects in fatty acid metabolism range from fasting hypoglycemia to frank car-diomyopathy or sudden death, most likely due to the accu-mulation of long-chain acyl-carnitines.203,204 A defi nitive diagnosis is not always easy because there is no correlation between in vitro enzyme assays in fi broblasts and clinical manifestations.205 Although of rare incidence, systemic car-nitine defi ciency presents as familial endocardial fi broelas-tosis, which is associated with mitochondrial substrate deprivation and myocardial triglyceride accumulation.206 As in experimental diphtheric cardiomyopathy,207 the abnor-mality is reversible with the administration of L-carnitine, and it has therefore been recommended that plasma carni-tine levels in all patients who present with familial cardio-myopathy be determined.206 A chronic cardiomyopathy has been described in children who have a defect in carnitine uptake.208
Ketone Bodies
The ketone bodies, acetoacetate and β-hydroxybutyrate, are produced by the liver when β-oxidation of FFAs exceeds the liver’s capacity for complete oxidation in the citric acid cycle. When released into the bloodstream, “predigested fatty acids” have ready access to the enzymes of the citric acid cycle in the heart. Thus, ketone bodies share in the supply of acetyl-CoA for energy production in heart muscle,209 but compared with the complexity of glucose and long-chain fatty acid metabolism, the metabolism of ketone bodies is simple and not subject to regulation.
Features of ketone body metabolism include their con-centration-dependent linear uptake in vivo210 and the activa-tion of acetoacetate to acetoacetyl-CoA by succinyl-CoA, which itself is a citric acid cycle intermediate. The enzymes that degrade β-hydroxybutyrate and acetoacetate to acetyl-CoA, β-hydroxybutyrate dehydrogenase, 3-oxoacid-CoA transferase, and acetoacetyl-CoA thiolase are of high activity in heart.209 The ability of the heart to oxidize ketone bodies may constitute a buffer against qualitative changes in sub-strate supply when the relative proportions of oxidizable fuel are shifting as a result of changes in the dietary or hormonal state of the body. Even in physiologic situations, such as short-term starvation or exercise, ketone body concentra-tions in the plasma can rise by up to 50-fold.211
When present as the only exogenous substrate for rat heart, ketone bodies do not sustain the full work output of the heart in vivo,212 and their rate of oxidation in the citric acid cycle is insuffi cient to meet the energy needs of the heart in vitro.35 This relative inhibition of the citric acid cycle occurs at the level of 2-oxoglutarate dehydrogenase and provides the fi rst demonstration of a defect in metabolism that causes reversible contractile dysfunction in the heart.35,213 The inhibition is caused by sequestration of the enzyme’s cofactor CoA SH, which is bound in the form of its thioester acetyl-CoA and as acetoacetyl-CoA, and is therefore not available for the reaction in the citric acid cycle.77
Observations in the isolated working rat heart perfused with ketone bodies have identifi ed hitherto unknown mech-anisms of the control of cardiac function by substrate metab-olism. These include the reversible inhibition of contractile function by inhibited fl ux through the Krebs citric acid cycle, the operational control of substrate fl ux through citric acid cycle spans, and the importance of anaplerosis (i.e., the provi-sion of oxaloacetate or other citric acid cycle intermediates in addition to acetyl-CoA for citrate synthesis for normal contractile function of the heart).159 The importance of anaplerosis of the Krebs cycle in heart muscle is also sug-gested by improved cardiac function with propionyl-L-carnitine214 and by stable isotope experiments using 13C-nuclear magnetic resonance spectroscopy,215 supporting the principle that the heart functions best when it oxidizes several substrates simultaneously. Pyruvate carboxylation in the in vivo heart accounts for at least 3% to 6% of citric acid cycle fl ux despite considerable variations in fl ux through pyruvate decarboxylase.216
Amino Acids
In addition to their role as “building blocks” of myocardial proteins, amino acids are also an integral part of myocardial energy metabolism. Most amino acids are either degraded or synthesized by heart muscle (e.g., glutamate, glutamine, aspartate, alanine, leucine), whereas others are not metabo-lized at all (e.g., phenylalanine, tyrosine).
The release of transaminases serves as biomarker of irre-versible myocardial injury and the diagnosis of myocardial infarction. Under physiologic circumstances one of the main functions of transaminases in heart muscle is the provision of carbon skeletons for the citric acid cycle.217 Furthermore, the amino acids aspartate and glutamate play an important role in the transfer of reducing equivalents across the mito-chondrial membrane for oxidation of cytosolic NADH by the mitochondrial electron transport chain. In the malate-aspartate cycle, cytosolic NADH, which cannot cross the inner mitochondrial membrane, is oxidized by the reduction of oxaloacetate to malate. Malate enters the mitochondrion and is oxidized to oxaloacetate, which is transaminated with glutamate to form 2-oxoglutarate and aspartate. The aspar-tate and 2-oxoglutarate leave the mitochondrion. Transmis-sion of these metabolites regenerates oxaloacetate and glutamate in the cytosol, the net effect being a transfer of hydrogen ions across the mitochondrial membrane.218–220 A second postulated function for the malate-aspartate shuttle is the delivery of intermediates (malate) to the citric acid cycle, serving as a modulator of cycle activity.221
CAR050.indd 1166CAR050.indd 1166 11/24/2006 12:14:13 PM11/24/2006 12:14:13 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 1167
Myocardial amino acid metabolism during hypoxia, isch-emia, and reperfusion has been the subject of a number of studies in the intact and isolated heart muscle.222–224 The amino acid alanine is like lactate, an end product of anaero-bic glucose breakdown, arising through transamination of glutamate in this reaction:
Glutamate + Pyruvate → 2-oxoglutarate + Alanine
Whereas alanine leaves the cell, 2-oxoglutarate is further metabolized and decarboxylated to succinate via substrate level phosphorylation in the citric acid cycle225:
2-oxoglutarate + NAD+ + GDP + Pi →Succinate + CO2 + NADH2 + GTP
where NAD+ is nicotinamide adenine dinucleotide, GDP is guanosine diphosphate, Pi is inorganic phosphate, and NADH2 is reduced nicotinamide adenine dinucleotide. Since guanosine triphosphate (GTP) is readily transphosphorylated to ATP, this reaction is a source of anaerobic energy indepen-dent of lactate formation. Enhanced glutamate uptake in vivo has been shown by nitrogen 13 (13N) accumulation from labeled glutamate in ischemic heart muscle,226 and gluta-mate enrichment has become a successful strategy to lessen the effects of ischemia with blood cardioplegia.227 However, the neurotoxicity of glutamate precludes its systemic administration.
Metabolic Signals of Cardiac Growth and Gene Expression
The heart’s response to changes in its environment is linked to a complex network of interconnecting signal transduction cascades.228 At fi rst glance it seems that signals arising from intermediary metabolism may have no place in such a scheme. However, we have reviewed evidence in support of the hypothesis that alterations in metabolic fl ux within the cardiomyocyte create essential signals for the adaptation of the heart,229 because many of the acute changes in metabolic fl ux are brought about by the same signal transduction cas-cades believed to be involved in the adaptation to the heart’s environment, e.g., phosphatidylinositol 3-kinase,98 diacylg-lycerol, Ca2+, and protein kinase C (PKC).126 Thus, metabolic signals are an integral part of cardiac adaptation.229 Here we briefl y mention some of the basic concepts of fatty acid and of glucose regulated gene expression.
As already discussed in detail, fatty acids are the pre-dominant fuel for respiration in the postnatal heart. In addi-tion to their role as energy providing substrates, fatty acids serve as mediators of signal transduction and as ligands for the nuclear transcription factor PPAR-α. Changes in sub-strate supply to the heart are refl ected in changes of fl ux through fatty acid metabolizing pathways. Levels of intracel-lular long-chain fatty acids are tightly regulated, so that there is a balance between transport of fatty acids from the interstitial space into the cell and transport from the cytosol into mitochondria and subsequent oxidation. This balance is regulated mainly by malonyl-CoA, which inhibits transport of fatty acids into mitochondria. Prolonged inhibition of CPT I promotes intramyocellular lipid accumulation and insulin resistance in rats.230 Conversely, leptin inhibits malonyl-
CoA synthesis in skeletal muscle, leading to greater rates of fatty acid oxidation.231 Metabolic dysregulation, especially in the situation where the rate of fatty acid uptake exceeds the rate of fatty acid oxidation,232,233 leads to a wide range of disturbances that have been attributed to mitochondrial dysfunction,234,235 and which culminate in triglyceride accumulation.172,236
Glucose and its metabolites also have multiple functions in the cardiac myocyte that extend beyond the role of glucose as an energy source. Failure to adequately control levels of intracellular glucose metabolites has been implicated in the development of insulin resistance and in the generation of reactive oxygen species (ROS). Compared to the liver,237 rela-tively little is known about the effects of glucose metabolites on gene expression in the heart. Through investigations of the glucose/carbohydrate response elements (GlRE/ChoRE) in the promoter regions of various glucose-regulated genes, a number of candidate transcription factors have been identi-fi ed that are believed to be involved in glucose-mediated gene expression. In the liver and in fat cells upstream stimulatory factor (USF), stimulatory protein-1 (Sp1), and sterol regulatory element binding protein-1 (SREBP1) play a role in glucose sensing,238 and it is reasonable to assume that the same tran-scription factors play similar roles in the heart.229
The Metabolic Syndrome
A discussion of cardiac metabolism is incomplete without a discussion of the metabolic syndrome (MS). The metabolic syndrome is not a disease but rather a constellation of physi-ognomic, physiologic, and pathologic features that refl ect the altered metabolic state of insulin resistance—a “metabolic axis of evil.” The constellation exhibits three characteristics: (1) it clusters within individuals; (2) it predicts the future development of type 2 diabetes mellitus; and (3) it predicts the development of atherosclerosis and the occurrence of ischemic events. The components of the MS may then be grouped into three categories: (1) disordered energy metabo-lism; (2) infl ammation; and (3) abnormalities of the circula-tion (including an increase in systemic vascular resistance and induction of a prothrombotic state).
Traditionally, the MS is a way of viewing the association between insulin resistance, altered fuel metabolism, and ath-erosclerotic cardiovascular disease, particularly coronary heart disease. To the epidemiologist, the metabolic syndrome is the set of attributes that share the property of being risk factors both for accelerated atherosclerosis and for develop-ment of type 2 diabetes mellitus.239 To the pathophysiologist, the metabolic syndrome consists of those metabolic and pathophysiologic consequences of visceral obesity and insulin resistance that are related to development of acceler-ated atherosclerosis. The fact that the components of the MS cluster within individuals implies that they are physio-logically related, although the underlying causes are not exactly known. To the clinician it has long been known that the MS, like type 2 diabetes mellitus, increases the risk for coronary, peripheral, and cerebral atherosclerotic/occlusive events. The new concept of cardiometabolic risk underscores that it precedes the appearance of hyperglyce-mia, at least for individuals who later develop type 2 diabetes
CAR050.indd 1167CAR050.indd 1167 11/24/2006 12:14:13 PM11/24/2006 12:14:13 PM

116 8 c h a p t e r 5 0
mellitus. Consequently, these individuals will benefi t from aggressive management of all known risk factors.
Obesity, type 2 diabetes, and the metabolic syndrome are multifactorial risk factors for cardiovascular diseases of con-siderable heterogeneity.240 However, whereas the diagnostic criteria for obesity and for type 2 diabetes are defi ned by large epidemiologic studies, the criteria for the metabolic syn-drome are not as clear-cut.241,242 The only consensus is that cardiovascular disease (CVD) is a major clinical outcome of the metabolic syndrome.242
Over the years there have been a number of attempts to develop standardized criteria for the diagnosis of MS. In 1998 the World Health Organization (WHO) proposed a defi nition stating that individuals with MS show evidence of insulin resistance plus at least two of four other factors: hyperten-sion, hyperlipidemia, obesity, and microalbuminuria.243 In 2001, the National Cholesterol Education Program’s (NCEP) Adult Treatment Panel III (ATP III) suggested a more practical defi nition for the metabolic syndrome (Table 50.1). At least three of the following fi ve factors need to be present for the diagnosis of the metabolic syndrome: increased waist cir-cumference, hypertriglyceridemia, low high-density lipopro-tein (HDL) cholesterol, hypertension, and a fasting glucose of 110 mg/dL (now 100 mg/dL) or higher. This defi nition is easier to use in clinical practice than the WHO guidelines. No matter what the defi nition, the term refers to a state of meta-bolic and circulatory dysregulation of the body. The term metabolic syndrome is somewhat of a misnomer because it fails to refl ect the profound metabolic dysregulation occur-ring at the organ level (e.g., muscle and liver). More appropri-ate would be the term dysmetabolic syndrome, which we prefer because it includes the connotation of dysregulated fuel supply due to profound changes in fuel homeostasis. As the result of an oversupply of substrate in the heart, the heart fails in the midst of plenty. For the heart in the pathophysio-logic state of diabetes mellitus, we have proposed three possible mechanisms for the metabolic maladaptation and contractile failure of the heart114,229—lipotoxicity, glucotoxic-ity, and glucolipotoxicity—none of which has been completely elucidated, although certain pharmacologic agents, especially
insulin sensitizing agents, hold the promise of reversing the adverse effects of metabolic dysregulation.201,202,244
We have shown that the progression of diabetes is, para-doxically, associated with a decline in messenger RNA (mRNA) transcript levels of PPAR-α.165,245 It is reasonable to assume that the continued exposure to high fatty acid levels will accelerate lipid accumulation of triglycerides (akin to a spillover of excess fuel). Like a sentinel, lipid accumulation also is a feature of the failing human heart171 and is associ-ated with a broad range of cellular and metabolic derange-ments collectively called lipotoxicity.172,246
Possible mechanisms causing a decline in contractile function of the “lipotoxic heart” include the inhibition of the adenine nucleotide translocator by fatty-acyl-CoA, the upregulation of tumor necrosis factor-α (TNF-α), increased ceramide levels, ROS accumulation, inducible nitric oxide synthase (iNOS) induction, chronic activation of PKCs, myosin isoform switches,247 downregulation of MEF2C regulated genes,248 cytochrome C release,249 and apoptosis.201 Interestingly, the heart’s ability to secrete lipoproteins is inversely related to pathologic lipid accumulation.250 In addi-tion, the ability to secrete lipoproteins protects the heart against the development of cardiomyopathy in a diabetes model.251 Lastly, in rodent models of lipotoxicity, the admin-istration of leptin decreases nonadipose tissue lipid accumu-lation and ameliorates lipotoxicity.252 Thus, a wide array of potentially reversible disturbances of fatty acid and lipid metabolism is involved in the phenomenon of lipotoxicity.
Excessive glucose metabolic accumulation is also associ-ated with various cardiac pathologies. Although studied mainly in the vasculature, the four main mechanisms pro-posed for hyperglycemia-induced diabetic complications252 are likely to be operative in the cardiomyocyte as well. The four hypotheses are increased polyol pathway fl ux (via aldose reductase), increased intracellular formation of advanced gly-cation end-products (AGE), and AGE-induced ROS genera-tion, activation of PKC mostly through activation by the lipid second messenger diacylglycerol (DAG), and increased fl ux through the hexosamine biosynthetic pathway.252 As men-tioned, many of these hypotheses have thus far been tested only in vascular tissue and await further confi rmation in cardiomyocytes. However, it is already known that O-linked β-N-acetylglucosamine (O-GlcNAc) modifi es many different nuclear and cytoplasmic proteins and that O-GlcNAc plays an important role in signal transduction.253 Young et al.229 have proposed a “spillover” of intermediates of glucose metabolism into the hexosamine biosynthetic pathway and into the pentose phosphate pathway. The concept of meta-bolic regulation of cell signaling is attractive in light of pos-sible crosstalk between glucose signaling and fatty acid signaling involving dysregulation of the 5′AMP-activated protein kinase AMPK/malonyl-CoA fuel sensing and signal-ing network.254 In this context Randle et al.’s255 “glucose-fatty acid cycle” comes again into play. The phenomenon describes the inactivation of pyruvate dehydrogenase by fatty acids, which is greater than the inhibition of glycolysis and glucose uptake in heart.
The “metabolic axis of evil” is a multifactorial, hetero-geneous group of risk factors for the development of cardio-vascular disease. Metabolic dysregulation of glucose and fatty acid metabolism involves many organs of the body,
TABLE 50.1. Components of cardiometabolic risk
Energy metabolism High triglycerides Low HDL cholesterol Small, dense LDL particles Increased apolipoprotein BInfl ammation Increased markers for infl ammation Directly adipose derived Leptin and adiponectin Indirectly adipose derived Tumor necrosis factor-α C-reactive proteinCirculation Hypertension Endothelial dysfunction Albuminuria Increased blood viscosity Tendency to thrombosis Increased fi brinogen levels Increased plasminogen activator inhibitor-1 (PAI-1) levels
CAR050.indd 1168CAR050.indd 1168 11/24/2006 12:14:13 PM11/24/2006 12:14:13 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 116 9
including the heart. Lipid metabolism does not stop in the arterial wall. In spite of its remarkable adaptability, the heart fails in the midst of plenty. We propose the following sce-nario: Elevated circulatory fatty acid levels result in activa-tion of PPAR-α in the cardiomyocyte, increased fatty acid oxidation, and decreased pyruvate oxidation (the Randle phe-nomenon). The dissociation of glycolytic fl ux and pyruvate oxidation results in the accumulation of glycolytic interme-diates that activate glucose sensitive transcription factors. The heart loses its capacity to switch from one substrate to another, and with additional stresses placed on the heart (e.g., hypertension, ischemia), metabolic maladaptation will occur. A vicious cycle is set into motion when substrate availability exceeds substrate oxidation by the heart. In principle, this phenomenon was already recognized by Rudolph Virchow’s256 textbook on cellular pathology when he described “fatty atrophy” as a “true metamorphosis of the heart muscle cell.” It appears that Virchow already recognized morphologic cor-relates of insulin resistance in the heart beyond the arterial wall.
Metabolic Targets to Improve Cardiac Function
Myocardial energy substrate metabolism has recently received the attention of the pharmaceutical industry, espe-cially with studies of the group of agents that are supposed to shift the heart’s energy supply from oxidation of fatty acids to the energetically more effi cient oxidation of glucose and lactate in the postischemic heart. This shift can be brought about by restoring insulin sensitivity of the heart, by inhibiting fatty acid oxidation at various levels, or by activating the pyruvate dehydrogenase complex. A case in point is glucose-insulin-potassium (GIK) in the treatment of cardiogenic shock after hypothermic ischemic arrest of the heart.147 Other examples include drugs such as etomoxir, ethyl-2-tetradecyl glycidate, and oxfenicine, drugs that inhibit long-chain fatty acid oxidation by inhibiting the entry of long-chain fatty acids into the mitochondria at the level of carnitine palmitoyl transferase-I (CPT-I).257,258 CPT-I has become a target for pharmacologic intervention in the postischemic reperfused heart in which rates of fatty acid oxidation are relatively high and contractile function is (reversibly) impaired. Etomoxir has also generated interest as a drug that may improve energy effi ciency in heart failure.259 However, the therapeutic window of etomoxir seems narrow, because in skeletal muscle, CPT-I inhibition leads to excess triglyceride accumulation and lipotoxicity.230 The piperazine compound trimetazidine belongs to the group of partial fatty acid oxidation (PFox) inhibitors and is widely used in France as an antianginal drug.260 Trimetazidine is thought to inhibit long-chain fatty acid oxidation by inhibiting one of the ter-minal steps in the β-oxidation pathway,261 resulting in a switch of energy substrate preference and improved exercise duration in patients with chronic, stable angina who were already receiving other antianginal therapy.262 Ranolazine decreases fatty acid oxidation, promotes glucose oxidation, and acts indirectly by increasing pyruvate dehydrogenase complex activity.263 The exact mechanism of action of PFox inhibitors, however, is still unknown.
The list of drugs targeting intermediary metabolism for the treatment of cardiac dysfunction also includes insulin-sensitizing agents such as thiazolidinediones, glucogen-like peptide-1 (GLP1),264 as well as propionyl L-carnitine, an anaplerotic agent that also raises the level of free coenzyme A. The opportunities for regeneration of normal cardiac myocyte function through metabolic interventions seem at present unlimited.
Summary
Myocardial metabolism is an integral part of the function of the heart as consumer and provider of energy. The bulk of the energy for contraction of the heart comes from oxidative phosphorylation of ADP. However, “energy reserves” in the form of ATP or phosphocreatine are very small. A vast network of highly regulated metabolic pathways regulates energy substrate metabolism and matches demand and supply with precision. Energy transfer is impaired in the failing heart, and restoration of effi cient energy substrate metabolism is an emerging target for the treatment of the failing heart.
Acknowledgments
I thank Roxy Ann Tate and Rebecca L. Salazar for help with the preparation of this manuscript as well as the past and present members of my laboratory for many discussions. Work in my lab is supported by the National Heart, Lung, and Blood Institute of the U.S. Public Health Service.
References
1. Goodwin GW, Taylor CS, Taegtmeyer H. Regulation of energy metabolism of the heart during acute increase in heart work. J Biol Chem 1998;273:29530–29539.
2. Taegtmeyer H, Golfman L, Sharma S, Razeghi P, Van Arsdall M. Linking gene expression to function: metabolic fl exibility in normal and diseased heart. Ann N Y Acad Sci 2004;1015:202–213.
3. Katz AM. Molecular biology in cardiology, a paradigmatic shift. J Mol Cell Cardiol 1988;20:355–366.
4. Taegtmeyer H. Genetics of energetics: transcriptional responses in cardiac metabolism. Ann Biomed Eng 2000;28:871–876.
5. Barger PM, Kelly DP. PPAR signaling in the control of cardiac energy metabolism. Trends Cardiovasc Med 2000;10:238–245.
6. Huss JM, Kelly DP. Nuclear receptor signaling and cardiac energetics. Circ Res 2004;95:568–578.
7. Huss JM, Kelly DP. Mitochondrial energy metabolism in heart failure: a question of balance. J Clin Invest 2005;115:547–555.
8. Taegtmeyer H, Wilson CR, Razeghi P, Sharma S. Metabolic energetics and genetics of the heart. Ann NY Acad Sci 2005; 1047:208–218.
9. Holmes FL. Between Biology and Medicine: The Formation of Intermediary Metabolism. Berkeley, CA: University of Califor-nia at Berkeley, 1992.
10. Evans CL. The metabolism of cardiac muscle. In: Newton W, ed. Evan’s Recent Advances in Physiology, vol 7, 6th ed. Phila-delphia: Blakiston’s, 1939:157–182.
11. Bing RJ. The metabolism of the heart. Harvey Lect 1955;50:27–70.
12. Randle PJ, Tubbs PK. Carbohydrate and fatty acid metabolism. In: Berne R, Sperelakis V, Geiger S, eds. Handbook of Physiol-
CAR050.indd 1169CAR050.indd 1169 11/24/2006 12:14:13 PM11/24/2006 12:14:13 PM

1170 c h a p t e r 5 0
ogy. The Cardiovascular system. Bethesda, MD: American Physiological Society, 1979:805–844.
13. Taegtmeyer H. Energy metabolism of the heart: from basic concepts to clinical applications. Curr Prob Cardiol 1994;19:57–116.
14. Balaban RS. Cardiac energy metabolism homeostasis: role of cytosolic calcium. J Mol Cell Cardiol 2002;34:1259–1271.
15. Lopaschuk GD. Malonyl CoA control of fatty acid oxidation in the diabetic rat heart. Adv Exp Med Biol 2001;498:155–165.
16. Chandler MP, Chavez PN, McElfresh TA, Huang H, Harmon CS, Stanley WC. Partial inhibition of fatty acid oxidation increases regional contractile power and effi ciency during demand-induced ischemia. Cardiovasc Res 2003;59:143–151.
17. Ingwall JS, Weiss RG. Is the failing heart energy starved? On using chemical energy to support cardiac function. Circ Res 2004;95:135–145.
18. Katz A. Is the failing heart energy depleted? Cardiol Clin 1998;16:633–644.
19. Taegtmeyer H, Razeghi P, Young ME. Alterations in Cardiac Metabolism in Heart Failure. In: Mann DC, ed. Heart Failure, a Companion to Braunwald’s Heart Disease. Philadelphia: WB Saunders, 2004:315–337.
20. Taegtmeyer H, Roberts AFC, Rayne AEG. Energy metabolism in reperfused rat heart: return of function before normalization of ATP content. J Am Coll Cardiol 1985;6:864–870.
21. Balaban RS, Kontor HL, Katz LA, Briggs RW. Relation between work and phosphate metabolite in the in vivo paced mamma-lian heart. Science 1986;232:1121–1123.
22. Kupriyanov VV, Lakomkin VL, Kapelko VI, Steinschneider AY, Ruuge EK, Saks VA. Dissociation of adenosine triphosphate levels and contractile function in isovolumic hearts perfused with 2–deoxyglucose. J Mol Cell Cardiol 1987;19:729–740.
23. Bassingthwaighte JB, Yipintsoi T, Harvey RB. Microvascula-ture of the dog left ventricular myocardium. Microvasc Res. Mar 1974;7:229–249.
24. Barth E, Stämmler G, Speiser B, Schaper J. Ultrastructural quantitation of mitochondria and myofi laments in cardiac muscle from 10 different animal species including man. J Mol Cell Cardiol 1992;24:669–681.
25. McNutt N, Fawcett D. Myocardial ultrastructure. In: Lauger G, Brady A, eds. The Mammalian Myocardium. New York: Wiley, 1974:1–50.
26. Baldwin JE, Krebs HA. The evolution of metabolic cycles. Nature 1981;291:381–382.
27. Newsholme EA, Start C. Regulation in Metabolism. London: Wiley, 1973.
28. Newsholme EA, Leech AR. Biochemistry for the Medical Sciences. Chichester: Wiley, 1983.
29. Fell D. Understanding the Control of Metabolism. Miami: Port-land Press, 1997.
30. Brown GC. Control of respiration and ATP synthesis in mam-malian mitochondria and cells. Biochem J 1992;284:1–13.
31. Krebs HA. Control of metabolic processes. Endeavour 1957;16:125–132.
32. Taegtmeyer H, King LM, Jones BE. Energy substrate metabo-lism, myocardial ischemia, and targets for pharmacotherapy. Am J Cardiol 1998;82(54K-60K):54K-60K.
33. Taegtmeyer H. Cardiac metabolism as a target for the treatment of heart failure [Editorial]. Circulation 2004;110:894–896.
34. Neely JR, Liebermeister H, Battersby EJ, Morgan HE. Effect of pressure development on oxygen consumption by isolated rat heart. Am J Physiol 1967;212:804–814.
35. Taegtmeyer H, Hems R, Krebs HA. Utilization of energy pro-viding substrates in the isolated working rat heart. Biochem J 1980;186:701–711.
36. Hochachka P, Somero G. Strategies of Biochemical Adapta-tions. Philadelphia: WB Saunders, 1973.
37. Balaban RS. Regulation of oxidative phosphorylation in the mammalian cell. Am J Physiol 1990;258:C377–C389.
38. Brown GC, Cooper CE. Control analysis applied to single enzymes: can an isolated enzyme have a unique rate-limiting step? Biochem J 1993;294(pt 1):87–94.
39. Beadle GW, Tatum EL. Genetic control of biochemical reactions in Neurospora. Proc Natl Acad Sci USA 1941;27:499–506.
40. Roseman R. Diet in haste; repent at leisure. The Biochemist 1992;14:6–10.
41. Jones B, Shan X, Park Y. Coordinated multisite regulation of cellular energy metabolism. Annu Rev Nutr 1992;12:327–343.
42. Parikh P, McDaniel MC, Ashen MD, et al. Diets and cardiovas-cular disease: an evidence-based assessment. J Am Coll Cardiol 2005;45:1379–1387.
43. Evans G. The glycogen content of the rat heart. J Physiol (Lond) 1934;82:468–480.
44. Denton RM, Randle PJ. Concentrations of glycerides and phos-pholipids in rat heart and gastrocnemius muscles. Biochem J 1967;104:416–422.
45. Gevers W. Protein metabolism of the heart. J Mol Cell Cardiol 1984;16:3–32.
46. Goldberg AL. Protein degradation and protection against mis-folded or damaged proteins. Nature 2003;426:895–899.
47. Lesch M, Taegtmeyer H, Peterson MB, Vernick R. Mechanism of the inhibition of myocardial protein synthesis during oxygen deprivation. Am J Physiol 1976;230:120–126.
48. Mitch WE, Goldberg AL. Mechanisms of muscle wasting. The role of the ubiquitin-proteasome pathway. N Engl J Med 1996;335:1897–1905.
49. Keul J, Doll E, Steim H, Homburger H, Kern H, Reindell H. Über den Stoffwechsel des menschlichen Herzens I. Pfl ügers Arch Ges Physiol 1965;282:1–27.
50. Korvald C, Elvenes OP, Myrmel T. Myocardial substrate metab-olism infl uences left ventricular energetics in vivo. Am J Physiol Heart Circ Physiol 2000;278:H1345–H1351.
51. Gadian D, Hoult D, Radda G, Seeley P, Chance B, Barlow C. Phosphorous nuclear magnetic resonance studies in normoxic and ischemic cardiac tissue. Proc Natl Acad Sci USA 1976;73:291–332.
52. Jacobus W, Taylor G, Hollis D, Nunnally R. Phosphorous nuclear magnetic resonance of perfused working rat hearts. Nature 1977;265:756–758.
53. Bottomley P. Noninvasive study of high energy phosphate metabolism in human heart by depth-resolved 31P NMR spec-troscopy. Science 1985;229:769–772.
54. McMillin-Wood J. Biochemical approaches in metabolism: application to positron emission tomography. Circulation 1985;72:IV145–IV150.
55. Schelbert HR. Assessment of myocardial metabolism by PET: A sophisticated dream or clinical reality? Eur J Nucl Med 1986;12:570–575.
56. Taegtmeyer H, Mossberg KA, Nguyen VTB. Positron labeled tracers: a window for the assessment of energy metabolism in heart and skeletal muscle. Acta Radiol Suppl 1991;376:40–44.
57. Schwaiger M, Hicks R. The clinical role of metabolic imaging of the heart by positron emission tomography. J Nucl Med 1991;32:565–578.
58. Lewandowski ED. Nuclear magnetic resonance evaluation of metabolic and respiratory support of work load in intact rabbit hearts. Circ Res 1992;70:576–582.
59. Russell RR, Cline GW, Guthrie PH, Goodwin GW, Shulman GI, Taegtmeyer H. Regulation of exogenous and endogenous glucose metabolism by insulin and acetoacetate in isolated working rat heart: a three-tracer study of glycolysis, glycogen metabolism and glucose oxidation. J Clin Invest 1997;100:2892–2899.
CAR050.indd 1170CAR050.indd 1170 11/24/2006 12:14:13 PM11/24/2006 12:14:13 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 1171
60. Malloy C, Sherry A, Jeffrey F. Carbon fl ux through citric acid cycle pathways in perfused heart by 13C NMR spectroscopy. FEBS Lett 1987;212:58–62.
61. Weiss RG, Gloth ST, Kalil-Filho R, Chacko VP, Stern MD, Gerstenblith G. Indexing tricarboxylic acid cycle fl ux in intact hearts by carbon-13 nuclear magnetic resonance. Circ Res 1992;70:392–408.
62. Laughlin MR, Fleming Taylor J, Chesnik AS, Balaban RS. Regu-lation of glycogen metabolism in canine myocardium: effects of insulin and epinephrine in vivo. Am J Physiol 1992;262:E875–E883.
63. Dence CS, Herrero P, Schwarz SW, Mach RH, Gropler RJ, Welch MJ. Imaging myocardium enzymatic pathways with carbon-11 radiotracers. Methods Enzymol 2004;385:286–315.
64. Nguyêñ VTB, Mossberg KA, Tewson TJ, et al. Temporal analy-sis of myocardial glucose metabolism by 18F-2–deoxy-2–fl uoro-D-glucose. Am J Physiol 1990;259:H1022–H1031.
65. Schelbert HR, Henze E, Sochor H, et al. Effects of substrate availability on myocardial C-11 palmitate kinetics by positron emission tomography in normal subjects and patients with ventricular dysfunction. Am Heart J 1986;111:1055–1064.
66. Brown MA, Marshall DR, Sobel BE, Bergmann SR. Delineation of myocardial oxygen utilization with carbon-11 labelled acetate. Circulation 1987;76:687–696.
67. Tillisch J, Brunken R, Marshall R, et al. Reversibility of cardiac wall-motion abnormalities predicted by positron tomography. N Engl J Med 1986;314:884–888.
68. Sokoloff L, Reivich M, Kennedy C, et al. The [14C] deoxyglucose method for the measurement of local cerebral glucose utiliza-tion: theory, procedure, and normal values in the conscious and anesthetized albino rat. J Neurochem 1977;28:897–916.
69. Hariharan R, Bray MS, Ganim R, Doenst T, Goodwin GW, Taegtmeyer H. Fundamental limitations of [18F] 2–deoxy-2–fl uoro-D-glucose for assessing myocardial glucose uptake. Circulation 1995;91:2435–2444.
70. Doenst T, Guthrie PH, Taegtmeyer H. Ischemic precondition-ing in rat heart: no correlation between glycogen content and return of function. Mol Cell Biochem 1998;180:153–161.
71. Bøtker HE, Goodwin GW, Holden JE, Doenst T, Gjedde A, Taegtmeyer H. Myocardial glucose uptake measured with fl uo-rodeoxyglucose: a proposed method to account for variable lumped constants. J Nucl Med 1999;40:1186–1196.
72. Davila-Roman VG, Vedala G, Herrero P, et al. Altered myocar-dial fatty acid and glucose metabolism in idiopathic dilated cardiomyopathy. J Am Coll Cardiol 2002;40:271–277.
73. Razeghi P, Young ME, Alcorn JL, Moravec CS, Frazier OH, Taegtmeyer H. Metabolic gene expression in fetal and failing human heart. Circulation 2001;104:2923–2931.
74. Fisher DJ, Heymann MA, Rudolph AM. Myocardial oxygen and carbohydrate consumption in fetal lambs in utero and in adult sheep. Am J Physiol 1980;238:H399–405.
75. Goodwin GW, Taegtmeyer H. Improved energy homeostasis of the heart in the metabolic state of exercise. Am J Physiol Heart Circ Physiol 2000;279:H1490–H1501.
76. Goodwin GW, Ahmad F, Doenst T, Taegtmeyer H. Energy pro-vision from glycogen, glucose and fatty acids upon adrenergic stimulation of isolated working rat heart. Am J Physiol 1998;274:H1239–H1247.
77. Russell RR, Taegtmeyer H. Coenzyme A sequestration in rat hearts oxidizing ketone bodies. J Clin Invest 1992;89:968–973.
78. Tirosh R, Mishor T, Pinson A. Glucose is essential for the initiation of fatty acid oxidation in ATP-depleted cultured myocytes. Mol Cell Biochem 1996;162:159–163.
79. Reid MM, Reilly BJ, Murdock AI, Swyer PR. Cardiomegaly in association with neonatal hypoglycaemia. Acta Paediatr Scand 1971;60:295–298.
80. Gertz EW, Wisneski JA, Neese RA, Bristow JD, Searle GL, Hanlon JT. Myocardial lactate metabolism: evidence of lactate release during net chemical extraction in man. Circulation 1981;63:1273–1279.
81. Goodwin GW, Ahmad F, Taegtmeyer H. Preferential oxidation of glycogen in isolated working rat heart. J Clin Invest 1996;97:1409–1416.
82. Wisneski JA, Gertz EW, Neese RA, Gruenke LD, Morris DL, Craig JC. Metabolic fate of extracted glucose in normal human myocardium. J Clin Invest 1985;76:1819–1827.
83. Wisneski JA, Stanley WC, Neese RA, Gertz EW. Effects of acute hyperglycemia on myocardial glycolytic activity in humans. J Clin Invest 1990;85:1648–1656.
84. Gertz EW, Wisneski JA, Stanley WC, Neese RA. Myocardial substrate utilization during exercise in humans. J Clin Invest 1988;82:2017–2025.
85. Gould GW. Facilitative Glucose Transporters. Georgetown, TX: Landes Co., Chapman & Hall, 1997.
86. Manchester J, Kong X, Nerbonne J, Lowry O, Lawrence J, Jr. Glucose transport and phosphorylation in single cardiac myo-cytes: rate limiting steps in glucose metabolism. Am J Physiol 1994;266:E326–E333.
87. Pessin JE, Bell GI. Mammalian facilitative glucose transporter family: structure and molecular regulation. Annu Rev Physiol 1992;54:911–930.
88. Gould GW, Holman GD. The glucose transporter family: struc-ture, function and tissue-specifi c expression. Biochem J 1993;295:329–341.
89. Mueckler M. Facilitative glucose transporters. Eur J Biochem 1994;219:713–725.
90. Shepherd PR, Kahn BB. Glucose transporters and insulin action–implications for insulin resistance and diabetes melli-tus. N Engl J Med 1999;341:248–257.
91. Olson AL, Pessin JE. Structure, function, and regulation of the mammalian facilitative glucose transporter gene family. Annu Rev Nutr 1996;16:235–256.
92. Das UN. Insulin: an endogenous cardioprotector. Curr Opin Crit Care 2003;9:375–383.
93. Baron AD. Hemodynamic actions of insulin. Am J Physiol 1994;267:E187–E222.
94. Cushman S, Wardzala L. Potential mechanism of insulin action on glucose transport in the isolated rat adipose cell. J Biol Chem 1980;255:4755–4762.
95. Wheeler TJ. Translocation of glucose transporters in response to anoxia in heart. J Biol Chem 1988;263:19447–19454.
96. Sun D, Nguyen N, Delgrado TR, Schwaiger M, Brosius FCI. Ischemia induces translocation of the insulin-responsive glucose transporter GLUT 4 to the plasma membrane of cardiac myocytes. Circulation 1994;89:793–798.
97. Russell RR, Yin R, Caplan MJ, Hu X, Ren J, Young LH. Ischemia on myocardial GLUT1 and GLUT4 translocation in vivo. Cir-culation 1998;98:2180–2186.
98. Doenst T, Taegtmeyer H. Alpha-adrenergic stimulation medi-ates glucose uptake through phosphatidylinositol 3–kinase in rat heart. Circ Res 1999;84:467–474.
99. Doenst T, Taegtmeyer H. Ischemia-stimulated glucose uptake does not require catecholamines in rat heart. J Mol Cell Cardiol 1999;31:435–443.
100. Depre C, Shipley GL, Chen W, et al. Unloaded heart in vivo replicates fetal gene expression of cardiac hypertrophy. Nat Med 1998;4:1269–1275.
101. Behrooz A, Ismail-Beigi F. Stimulation of glucose transport by hypoxia: signals and mechanisms. News Physiol Sci 1999;14:105–110.
102. Liao R, Jain M, Cui L, et al. Cardiac-specifi c overexpression of GLUT1 prevents the development of heart failure due to pres-sure-overload in mice. Circulation 2002;106:2125–2131.
CAR050.indd 1171CAR050.indd 1171 11/24/2006 12:14:13 PM11/24/2006 12:14:13 PM

117 2 c h a p t e r 5 0
103. Nishimura H, Pallardo FV, Seidner GA, Vannucci S, Simpson IA, Birnbaum MF. Kinetics of GLUT1 and GLUT4 glucose transporters expressed in Xenopus oocytes. J Biol Chem 1993;268:8514–8520.
104. Shepherd PR, Gould GW, Colville CA, McCoid SC, Gibbs EM, Kahn BB. Distribution of GLUT3 glucose transporter protein in human tissues. Biochem Biophys Res Commun 1992;188:149–154.
105. Joost HG, Bell GI, Best JD, et al. Nomenclature of the GLUT/SLC2A family of sugar/polyol transport facilitators. Am J Physiol Endocrinol Metab 2002;282:E974–976.
106. Barrett E, Schwartz R, Francis C, Zaret B. Regulation by insulin of myocardial glucose and fatty acid metabolism in the con-scious dog. J Clin Invest 1984;74:1073–1079.
107. Russell RR, Nguyêñ VTB, Mrus JM, Taegtmeyer H. Fasting and lactate unmask insulin responsiveness in the isolated working rat heart. Am J Physiol 1992;263:E556–E561.
108. Morgan HE, Henderson MJ, Regen DM, Park CR. Regulation of glucose uptake in muscle. I. The effects of insulin and anoxia on glucose transport and phosphorylation in the isolated per-fused heart of normal rats. J Biol Chem 1961;236:253–261.
109. Schwaiger M, Neese RA, Araujo L, et al. Sustained nonoxida-tive glucose utilization and depletion of glycogen in reperfused canine myocardium. J Am Coll Cardiol 1989;13:745–754.
110. Abel ED. Insulin signaling in heart muscle: lessons from genet-ically engineered mouse models. Curr Hypertens Rep 2004;6:416–423.
111. Newsholme EA, Crabtree B. Theoretical principles in the approaches to the control of metabolic pathways and their application to glycolysis in muscle. J Mol Cell Cardiol 1979;11:839–856.
112. Camici P, Ferrannini E, Opie LH. Myocardial metabolism in ischemic heart disease: basic principles and application to imaging by positron emission tomography. Progr Cardiovasc Dis 1989;32:217–238.
113. Goodwin GW, Cohen DM, Taegtmeyer H. [5–3H]glucose over-estimates glycolytic fl ux in isolated working rat heart: role of the pentose phosphate pathway. Am J Physiol Endocrinol Metab 2001;280:E502–E508.
114. Taegtmeyer H, McNulty P, Young ME. Adaptation and malad-aptation of the heart in diabetes: Part I: general concepts. Cir-culation 2002;105:1727–1733.
115. Doenst T, Taegtmeyer H. Profound underestimation of glucose uptake by [18F]2–deoxy-2–fl uoroglucose in reperfused rat heart muscle. Circulation 1998;97:2454–2462.
116. Cohen P. Control of Enzyme Activity. London: Chapman & Hall, 1976.
117. Cori G, Colowick S, Cori C. The action of nucleotides on the disruptive phosphorylation of glycogen. J Biol Chem 1938;123:381–389.
118. Krebs EG, Fischer EH. The phosphorylase b to a converting enzyme of rabbit skeletal muscle. Biochim Biophys Acta 1956;20:150–157.
119. Robison G, Butcher R, Sutherland E. Cyclic AMP. London: Academic Press, 1971.
120. Taegtmeyer H. Glycogen in the heart—an expanded view. J Mol Cell Cardiol 2004;37:7–10.
121. Johnson M, Everitt B. The high concentration of glycogen in fetal cardiac muscle probably explains why the heart can main-tain its contractile activity in the face of severe hypoxia. In: Essential Reproduction, 3rd ed. Oxford: Blackwell Scientifi c, 1988:275.
122. Aschoff L. Ueber den Glykogengehalt des Reizleitungssystems des Säugetierherzens. Verh Dtsch Pathol Ges 1908;12:150.
123. Entman ML, Kanike K, Goldstein MA, et al. Association of glycogenolysis with cardiac sarcoplasmic reticulum. J Biol Chem 1976;251:3140–3146.
124. Chin E, Allen D. Effects of reduced muscle glycogen concentra-tion on force, Ca2+ release and contractile protein function in intact mouse skeletal muscle. J Physiol (Lond) 1997;498:17–29.
125. Laughlin M, Petit W, Dizon J, Shulman R, Barrett E. NMR measurements of in vivo myocardial glycogen metabolism. J Biol Chem 1988;263:2285–2291.
126. Cohen P. The hormonal control of glycogen metabolism in mammalian muscle by multisite phosphorylation. Biochem Soc Trans 1979;7:459–480.
127. Bolukoglu H, Goodwin GW, Guthrie PH, Carmical SG, Chen TM, Taegtmeyer H. Metabolic fate of glucose in reversible low-fl ow ischemia of the isolated working rat heart. Am J Physiol 1996;270:H817–826.
128. Chen TM, Goodwin GW, Guthrie PH, Taegtmeyer H. Effects of insulin on glucose uptake by rat hearts during and after coronary fl ow reduction. Am J Physiol 1997;273:H2170–H2177.
129. Scheuer J, Stezoski SW. Protective role of increased myocardial glycogen stores in cardiac anoxia in the rat. Circ Res 1970;27:835–849.
130. Hearse DJ, Chain EB. The role of glucose in the survival and “recovery” of the anoxic isolated perfused rat heart. Biochem J 1972;128:1125–1133.
131. McElroy DD, Walker WE, Taegtmeyer H. Glycogen loading improves left ventricular function of the rabbit heart after hypothermic ischemic arrest. J Appl Cardiol 1989;4:455–465.
132. Schneider CA, Taegtmeyer H. Fasting in vivo delays myocar-dial cell damage after brief periods of ischemia in the isolated working rat heart. Circ Res 1991;68:1045–1050.
133. Schneider CA, Nguyêñ VTB, Taegtmeyer H. Feeding and fasting determine postischemic glucose utilization in isolated working rat hearts. Am J Physiol 1991;260:H542–548.
134. Conlee R, Tipton C. Cardiac glycogen depletion after exercise: infl uence of synthase and glucose 6–P. J Appl Physiol 1977;42:240–244.
135. Opie LH. The glucose hypothesis: relation to acute myocardial ischemia. J Mol Cell Cardiol 1970;1:107–115.
136. Apstein CS, Gravino FN, Haudenschild CC. Determinants of a protective effect of glucose and insulin on the ischemic myo-cardium. Effects on contractile function, diastolic compliance, metabolism, and ultrastructure during ischemia and reperfu-sion. Circ Res 1983;52:515–526.
137. Doenst T, Guthrie PH, Chemnitius JM, Zech R, Taegtmeyer H. Fasting, lactate, and insulin improve ischemia tolerance: a comparison with ischemic preconditioning. Am J Physiol 1996;270:H1607–H1615.
138. Wasser JS, Meinertz EA, Chang SY, Lawler RG, Jackson DC. Metabolic and cardiodynamic responses of isolated turtle hearts to ischemia and reperfusion. Am J Physiol 1992;262:H437–H443.
139. Taegtmeyer H, Peterson MB, Ragavan VV, Ferguson AG, Lesch M. De novo alanine synthesis in isolated oxygen-deprived rabbit myocardium. J Biol Chem 1977;252:5010–5018.
140. Taegtmeyer H, Overturf ML. Effects of moderate hypertension on cardiac function and metabolism in the rabbit. Hyperten-sion 1988;11:416–426.
141. Bishop S, Altschuld R. Increased glycolytic metabolism in cardiac hypertrophy and congestive heart failure. Am J Physiol 1970;218:153–159.
142. Allard MF, Emanuel PG, Russell JA, Bishop SP, Digerness SB, Anderson, PG. Preischemic glycogen reduction or glycolytic inhibition improves postischemic recovery of hypertrophied rat heart. Am J Physiol 1994;267:H66–H74.
143. Doenst T, Goodwin GW, Cedars AM, Wang M, Stepkowski S, Taegtmeyer H. Load-induced changes in vivo alter substrate fl uxes and insulin responsiveness of rat heart in vitro. Metabo-lism 2001;50:1083–1090.
CAR050.indd 1172CAR050.indd 1172 11/24/2006 12:14:14 PM11/24/2006 12:14:14 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 117 3
144. Eberli FR, Weinberg EO, Grice WN, Horowitz GL, Apstein CS. Protective effect of increased glycolytic substrate against sys-tolic and diastolic dysfunction and increased coronary resis-tance from prolonged global underperfusion and reperfusion in isolated rabbit hearts perfused with erythrocyte suspensions. Circ Res 1991;68:466–481.
145. Neely JR, Grotyohann LW. Role of glycolytic products in damage to myocardium: dissociation of adenosine triphosphate levels and recovery of function of reperfused canine myocar-dium. Circ Res 1984;55:816–824.
146. Kersten JR, Schmeling TJ, Orth KG, Pagel PS, Warltier DC. Acute hyperglycemia abolishes ischemic preconditioning in vivo. Am J Physiol 1998;275(2 pt 2):H721–725.
147. Gradinak S, Coleman GM, Taegtmeyer H, Sweeney MS, Frazier OH. Improved cardiac function with glucose-insulin-potassium after coronary bypass surgery. Ann Thorac Surg 1989;48:484–489.
148. Taegtmeyer H. The use of hypertonic glucose, insulin, and potassium (GIK) in myocardial preservation. J Appl Cardiol 1991;6:255–259.
149. Aasum E, Lathrop DA, Henden T, Sundset R, Larsen TS. The role of glycolysis in myocardial calcium control. J Mol Cell Cardiol 1998;30:1703–1712.
150. Doenst T, Richwine RT, Bray MS, Goodwin GW, Frazier OH, Taegtmeyer H. Insulin improves functional and metabolic recovery of reperfused working rat heart. Ann Thorac Surg 1999;67:1682–1688.
151. Brosius FC. How glucose and glucose transporters protect cardiac myocytes. J Mol Cell Cardiol 2003;35:1187–1189.
152. Passonneau JV, Lowry OH. Phosphofructokinase and the Pasteur effect. Biochem Biophys Res Commun 1962;7:10–15.
153. Uyeda K. Phosphofructokinase. Adv Enzymol 1979;48:193–244.
154. Kobayashi K, Neely J. Control of maximum rates of glycolysis in rat cardiac muscle. Circ Res 1979;44:166–175.
155. Hue L, Rider MH. Role of fructose 2,6-bisphosphate in the control of glycolysis in mammalian tissues. Biochem J 1987;245:313–324.
156. Rovetto M, Lamberton W, Neely J. Mechanisms of glycolytic inhibition in ischemic rat heart. Circ Res 1975;37:742–751.
157. Nuutila P, Koivisto VA, Knuuti J, et al. Glucose-free fatty acid cycle operates in human heart and skeletal muscle in vivo. J Clin Invest 1992;89:1767–1774.
158. Peuhkurinen KJ, Hassinen IE. Pyruvate carboxylation as an anaplerotic mechanism in the isolated perfused rat heart. Biochem J 1982;202:67–76.
159. Russell RR, Taegtmeyer H. Pyruvate carboxylation prevents the decline in contractile function of rat hearts oxidizing ace-toacetate. Am J Physiol 1991;261:H1756–H1762.
160. Denton RM, Halestrap AP. Regulations of pyruvate metabo-lism in mammalian tissues. Essays Biochem 1979;15:37–77.
161. Randle P. Regulation of glycolysis and pyruvate oxidation in cardiac muscle. Circ Res 1976;38(suppl 1):I8–I15.
162. Olson MS, Dennis SC, DeBuysere MS, Padma A. The regula-tion of pyruvate dehydrogenase in the isolated perfused rat heart. J Biol Chem 1978;253:7369–7375.
163. Kerbey AL, Randle PJ, Cooper RH, Whitehouse S, Pask HT, Denton RM. Regulation of pyruvate dehydrogenase in rat heart. Biochem J 1976;154:327–348.
164. Borst P, Loos JA, Christ EJ, Slater EC. Uncoupling activity of long chain fatty acids. Biochim Biophys Acta 1962;62:509–517.
165. Young ME, Patil S, Ying J, et al. Uncoupling protein 3 transcrip-tion is regulated by peroxisome proliferator-activated receptor (alpha) in the adult rodent heart. FASEB J 2001;15:833–845.
166. Schaffer JE. Fatty acid transport: the roads taken. Am J Physiol Endocrinol Metab 2002;282:E239–E246.
167. Bass NM. The cellular fatty acid binding proteins: aspects of structure, regulation, and function. Int Rev Cytol 1988;111:143–184.
168. van der Vusse G, Groot M. Interrelationship between lactate and cardiac fatty acid metabolism. Mol Cell Biochem 1992;116:11–17.
169. Binas B, Danneberg H, McWhir J, Mullins L, Clark AJ. Require-ment for the heart-type fatty acid binding protein in cardiac fatty acid utilization. FASEB J 1999;13:805–812.
170. Erol E, Cline GW, Kim JK, Taegtmeyer H, Binas B. Non-acute effects of H-FABP defi ciency on skeletal muscle glucose uptake in vitro. Am J Physiol Endocrinol Metab 2004;287:E977–982.
171. Sharma S, Adrogue JV, Golfman L, et al. Intramyocardial lipid accumulation in the failing human heart resembles the lipo-toxic rat heart. FASEB J 2004;18:1692–1700.
172. Schaffer JE. Lipotoxicity: when tissues overeat. Curr Opin Lipidol 2003;14:281–287.
173. McGarry JD, Mannaerts GP, Foster DW. A possible role for malonyl-CoA in the regulation of hepatic fatty acid oxidation and ketogenesis. J Clin Invest 1977;60:265–270.
174. McGarry JD, Mills SE, Long CS, Foster DW. Observations on the affi nity for carnitine and malonyl-CoA sensitivity of car-nitine palmitoyl transferase I in animal and human tissues. Demonstration of the presence of malonyl-CoA in non-hepatic tissues of the rat. Biochem J 1983;214:21–28.
175. Paulson D, Ward K, Shug A. Malonyl-CoA inhibition of carni-tine palmitoyl transferase. FEBS Lett 1984;176:381–384.
176. Bianchi A, Evans J, Iverson A, Nordlund A, Watts T, Wilters L. Identifi cation of an isozymic form of acetyl-CoA carboxylase. J Biol Chem 1990;265:1502–1509.
177. Dyck JR, Barr AJ, Barr RL, Kolattukudy PE, Lopaschuk GD. Characterization of cardiac malonyl-CoA decarboxylase and its putative role in regulating fatty acid oxidation. Am J Physiol 1998;275(6 pt 2):H2122–H2129.
178. Goodwin GW, Taegtmeyer H. Regulation of fatty acid oxida-tion of the heart by MCD and ACC during contractile stimula-tion. Am J Physiol 1999;277:E772–777.
179. Young ME, Goodwin GW, Ying J, et al. Regulation of cardiac and skeletal muscle malonyl-CoA decarboxylase by fatty acids. Am J Physiol Endocrinol Metab 2001;280:E471–E479.
180. Hardie DG, Carling D. The AMP-activated protein kinase—fuel gauge of the mammalian cell? Eur J Biochem 1997;246:259–273.
181. Pauly D, Kirk K, McMillin J. Carnitine palmitoyl transferase in cardiac ischemia. A potential site for altered fatty acid metabolism. Circ Res 1991;68:1085–1094.
182. Bremer J, Wojtzak A. Factors controlling the role of fatty acid beta oxidation in rat liver mitochondria. Biochem Biophys Acta 1972;280:515–530.
183. Shrago E, Shug AL, Sul H, Bittar N, Folts JD. Control of energy production in myocardial ischemia. Circ Res 1976;38(suppl 1):75–81.
184. Brandt JM, Djouadi F, Kelly DP. Fatty acids activate transcrip-tion of the muscle carnitine palmitoyltransferase I gene in cardiac myocytes via the peroxisome proliferator-activated receptor alpha. J Biol Chem 1998;273:23786–23792.
185. Vamecq J, Latruffe N. Medical signifi cance of peroxisome pro-liferator-activated receptors. Lancet 1999;354:141–148.
186. Dyck JR, Cheng JF, Stanley WC, et al. Malonyl coenzyme a decarboxylase inhibition protects the ischemic heart by inhib-iting fatty acid oxidation and stimulating glucose oxidation. Circ Res 2004;94:E78–84.
187. van Bilsen M, Van der Vusse GJ, Reneman RS. Transcriptional regulation of metabolic processes: implications for cardiac metabolism. Pfl ugers Arch 1998;437:2–14.
188. Gulick T, Cresci S, Caira T, Moore D, Kelly D. The peroxisome proliferator-activated receptor regulates mitochondrial fatty
CAR050.indd 1173CAR050.indd 1173 11/24/2006 12:14:14 PM11/24/2006 12:14:14 PM

1174 c h a p t e r 5 0
acid oxidative enzyme gene expression. Proc Natl Acad Sci USA 1994;91:11012–11016.
189. van der Lee K, Vork M, De Vries J, et al. Long-chain fatty acid-induced changes in gene expression in neonatal cardiac myo-cytes. J Lipid Res 2000;41:41–47.
190. Lehman JJ, Barger PM, Kovacs A, Saffi tz JE, Medeiros DM, Kelly DP. Peroxisome proliferator-activated receptor gamma coactivator-1 promotes cardiac mitochondrial biogenesis. J Clin Invest 2000;106:847–856.
191. Kelly DP, Scarpulla RC. Transcriptional regulatory circuits controlling mitochondrial biogenesis and function. Genes Dev 2004;18:357–368.
192. Ino T, Sherwood WG, Benson LN, Wilson GJ, Freedom RM, Rowe RD. Cardiac manifestations in disorders of fat and car-nitine metabolism in infancy. J Am Coll Cardiol 1988;11:1301–1308.
193. Wallace DC. Mitochondrial defects in cardiomyopathy and neuromuscular disease. Am Heart J 2000;139(2 pt 3):S70–85.
194. Bohles H, Sewell AC, ed. Metabolic Cardiomyopathy, revised 2nd ed. Stuttgart: Medpharm Scientifi c, 2004.
195. Coates PM, Stanley CA. Inherited disorders of mitochondrial fatty acid oxidation. Prog Liver Dis 1992;10:123–138.
196. Sewell A. Laboratory diagnosis of metabolic diseases present-ing with cardiomyopathy. In: Bohles H, Sewell AC, eds. Meta-bolic Cardiomyopathy, revised 2nd ed. Stuttgart: Medpharm Scientifi c, 2004:153–161.
197. Hale DE, Thorpe C, Braat K, et al. The L-3–hydroxyacyl-CoA dehydrogenase defi ciency. Prog Clin Biol Res 1990;321:503–510.
198. Kelly DP, Strauss AW. Inherited cardiomyopathies. N Engl J Med 1994;330:913–919.
199. Hale DE, Bennett MJ. Fatty acid oxidation disorders: a new class of metabolic diseases. J Pediatr 1992;121:1–11.
200. Young ME, Patil S, Davies PJA, Taegtmeyer H. Anaplerosis in the rat heart: decreased expression of pyruvate carboxylase with diabetes. Diabetes 2000;49(suppl 1):A278.
201. Zhou YT, Grayburn P, Karim A, et al. Lipotoxic heart disease in obese rats: implications for human obesity. Proc Natl Acad Sci U S A 2000;97:1784–1789.
202. Golfman LS, Wilson CR, Sharma S, et al. Activation of PPAR{gamma} enhances myocardial glucose oxidation and improves contractile function in isolated working hearts of ZDF rats. Am J Physiol Endocrinol Metab 2005;289:E328–E336.
203. Naylor I, Mosovich L, Guthrie R, et al. Intermittent non-ketotic dicarboxylic aciduria in two siblings with hypoglycemia: an apparent defect in beta-oxidation of fatty acids. J Inherit Metab Dis 1980;3:19–24.
204. Schwenk WF, Hale DE, Haymond MW. Decreased fasting free fatty acids with L-carnitine in children with carnitine defi -ciency. Pediatr Res 1988;23:491–494.
205. Amendt BA, Moon A, Teel L, Rhead WJ. Long-chain acyl-coenzyme A dehydrogenase defi ciency: biochemical studies in fi broblasts from three patients. Pediatr Res 1988;23:603–605.
206. Tripp ME, Katcher ML, Peters HA, et al. Systemic carnitine defi ciency presenting as familial endomyocardial fi broelasto-sis. N Engl J Med 1981;305:385–390.
207. Challoner D, Pols H. Fatty acid concentration and carnitine levels in diphtheric guinea pig myocardium. J Clin Invest 1972;51:2071–2076.
208. Stanley CA, DeLeeuw S, Coates PM, et al. Chronic cardiomy-opathy and weakness or acute coma in children with a defect in carnitine uptake. Ann Neurol 1991;30:709–716.
209. Williamson JR, Krebs HA. Acetoacetate as fuel of respiration in the perfused rat heart. Biochem J 1961;80:540–547.
210. Rudolph W, Haas D, Richter J, Masinger F, Hofmann H, Dohm P. Uber die Bedeutung von Acetoacetat und ß-Hydroxybutyrat
im Stoffwechel des menschlichen Herzens. Klin Wochenschr 1965;43:445–451.
211. Taegtmeyer H. Six blind men explore an elephant: aspects of fuel metabolism and the control of tricarboxylic acid cycle activity in heart muscle. Basic Res Cardiol 1984;79:322–336.
212. Zimmermann ANE, Meijler FL, Hülsmann WC. The inhibitory effect of acetoacetate on myocardial contraction. Lancet 1962;2:757–758.
213. Taegtmeyer H. On the inability of ketone bodies to serve as the only energy providing substrate for rat heart at physiological work load. Basic Res Cardiol 1983;78:435– 450.
214. Russell RR, Mommessin JI, Taegtmeyer H. Propionyl-L-carnitine-mediated improvement in contractile function of rat hearts oxidizing acetoacetate. Am J Physiol 1995;268:H441–H447.
215. Cohen DM, Bergman RN. Improved estimation of anaplerosis in heart using 13C NMR. Am J Physiol 1997;273(6 pt 1):E1228–1242.
216. Panchal AR, Comte B, Huang H, et al. Partitioning of pyruvate between oxidation and anaplerosis in swine hearts. Am J Physiol Heart Circ Physiol 2000;279:H2390–2398.
217. Krebs HA. Some aspects of the regulation of fuel supply in omnivorous animals. Adv Enzyme Regul 1972;10:397–420.
218. Yu X, White LT, Alpert NM, Lewandowski ED. Subcellular metabolite transport and carbon isotope kinetics in the intra-myocardial glutamate pool. Biochemistry 1996;35:6963–6968.
219. Lewandowski ED, Yu X, LaNoue KF, White LT, Doumen C, O’Donnell JM. Altered metabolite exchange between subcel-lular compartments in intact postischemic rabbit hearts. Circ Res 1997; 81:165–175.
220. O’Donnell JM, Kudej RK, LaNoue KF, Vatner SF, Lewandowski ED. Limited transfer of cytosolic NADH into mitochondria at high cardiac workload. Am J Physiol Heart Circ Physiol 2004;286:H2237–2242.
221. Lewandowski ED, Doumen C, White LT, LaNoue KF, Damico LA, Yu X. Multiplet structure of 13C NMR signal from gluta-mate and direct detection of tricarboxylic acid (TCA) cycle intermediates. Magn Reson Med 1996;35:149–154.
222. Taegtmeyer H, Ferguson AG, Lesch M. Protein degradation and amino acid metabolism in autolyzing rabbit myocardium. Exp Mol Pathol 1977;26:52.
223. Mudge GHJ, Mills RMJ, Taegtmeyer H, Gorlin R, Lesch M. Alterations of myocardial amino acid metabolism in chronic ischemic heart disease. J Clin Invest 1976;58:1185–1192.
224. Bittl J, Shine K. Protection of ischemic rabbit myocardium by glutamic acid. Am J Physiol 1983;245:H406–H412.
225. Taegtmeyer H. Metabolic responses to cardiac hypoxia: increased production of succinate by rabbit papillary muscles. Circ Res 1978;43:808–815.
226. Knapp W, Helus F, Ostertag H, Tilmanns H, Kuebler W. Uptake and turnover of L-[13N] glutamate in the normal human heart and patients with coronary artery disease. Eur J Nucl Med 1982;7:211–215.
227. Robertson J, Vinten-Johansen J, Buckberg G, Rosenkranz E, Maloney J. Safety of prolonged aortic clamping with blood car-dioplegia: glutamate enrichment in normal hearts. J Thorac Cardiovas Surg 1984;88:402–410.
228. Molkentin JD, Dorn GW. Cytoplasmic signaling pathways that regulate cardiac hypertrophy. Annu Rev Physiol 2001;63:391–426.
229. Young ME, McNulty P, Taegtmeyer H. Adaptation and malad-aptation of the heart in diabetes: Part II: potential mechanisms. Circulation 2002;105:1861–1870.
230. Dobbins RL, Szczepaniak LS, Bentley B, Esser V, Myhill J, McGarry JD. Prolonged inhibition of muscle carnitine palmi-toyltransferase-1 promotes intramyocellular lipid accumula-tion and insulin resistance in rats. Diabetes 2001;50:123–130.
CAR050.indd 1174CAR050.indd 1174 11/24/2006 12:14:14 PM11/24/2006 12:14:14 PM

f u e l i ng t h e h e a rt : m u lt i p l e rol e s f or c a r di ac m e ta b ol i s m 1175
231. Friedman JJ. Fat in all the wrong places. Nature 2002;415:268–269.
232. Chiu HC, Kovacs A, Ford DA, et al. A novel mouse model of lipotoxic cardiomyopathy. J Clin Invest 2001;107:813–822.
233. Yagyu H, Chen G, Yokoyama M, et al. Lipoprotein lipase (LpL) on the surface of cardiomyocytes increases lipid uptake and produces a cardiomyopathy. J Clin Invest 2003;111:419–426.
234. Shulman GI. Cellular mechanisms of insulin resistance. J Clin Invest 2000;106:171–176.
235. Petersen KF, Dufour S, Befroy D, Garcia R, Shulman GI. Impaired mitochondrial activity in the insulin-resistant off-spring of patients with type 2 diabetes. N Engl J Med 2004;350:664–671.
236. Unger RH, Orci L. Diseases of liporegulation: new perspective on obesity and related disorders. FASEB J 2001;15:312–321.
237. Ferre P. Regulation of gene expression by glucose. Proc Nutr Soc 1999;58:621–623.
238. Vaulont S, Vasseur-Cognet M, Kahn A. Glucose regulation of gene transcription. J Biol Chem 2000;275:31555–31558.
239. Reaven G. Banting lecture 1988. Role of insulin resistance in human disease. Diabetes 1988;37:1595–1607.
240. Zimmet P, Alberti KG, Shaw J. Global and societal implications of the diabetes epidemic. Nature 2001;414:782–787.
241. Haffner S, Taegtmeyer H. Epidemic obesity and the metabolic syndrome [Editorial]. Circulation 2003;108:1541–1545.
242. Grundy SM, Brewer HB Jr, Cleeman JI, Smith SC Jr, Lenfant C. Defi nition of metabolic syndrome: report of the National Heart, Lung, and Blood Institute/American Heart Association conference on scientifi c issues related to defi nition. Circula-tion 2004;109:433–438.
243. Alberti KG, Zimmet PZ. Defi nition, diagnosis and classifi ca-tion of diabetes mellitus and its complications. Part 1: diagno-sis and classifi cation of diabetes mellitus provisional report of a WHO consultation. Diabet Med 1998;15:539–553.
244. Nemoto S, Razeghi P, Ishiyama M, De Freitas G, Taegtmeyer H, Carabello BA. PPAR-gamma agonist rosiglitazone amelio-rates ventricular dysfunction in experimental chronic mitral regurgitation. Am J Physiol Heart Circ Physiol 2005;288:H77–82.
245. Depre C, Young ME, Ying J, et al. Streptozotocin-induced changes in cardiac gene expression in the absence of severe contractile dysfunction. J Mol Cell Cardiol 2000;32:985–996.
246. Unger RH. Longevity, lipotoxicity and leptin: the adipocyte defense against feasting and famine. Biochimie 2005;87:57–64.
247. Dillmann WH. Diabetes mellitus induces changes in cardiac myosin of the rat. Diabetes 1980;29:579–582.
248. Razeghi P, Young ME, Cockrill TC, Frazier OH, Taegtmeyer H. Downregulation of myocardial myocyte enhancer factor 2C and myocyte enhancer factor 2C regulated gene expression in diabetic patients with non-ischemic heart failure. Circulation 2002;106:407–411.
249. Ostrander DB, Sparagna GC, Amoscato AA, McMillin JB, Dowhan W. Decreased cardiolipin synthesis corresponds with cytochrome c release in palmitate-induced cardiomyocyte apoptosis. J Biol Chem 2001;276:38061–38067.
250. Bjorkegren J, Veniant M, Kim SK, et al. Lipoprotein secretion and triglyceride stores in the heart. J Biol Chem 2001;276:38511–38517.
251. Nielsen LB, Bartels ED, Bollano E. Overexpression of apolipo-protein B in the heart impedes cardiac triglyceride accumula-tion and development of cardiac dysfunction in diabetic mice. J Biol Chem 2002;277:27014–27020.
252. Lee Y, Wang MY, Kakuma T, et al. Liporegulation in diet-induced obesity. The antisteatotic role of hyperleptinemia. J Biol Chem 2001;276:5629–5635.
253. Zachara NE, Hart GW. O-GlcNAc a sensor of cellular state: the role of nucleocytoplasmic glycosylation in modulating cellular function in response to nutrition and stress. Biochim Biophys Acta 2004;1673:13–28.
254. Ruderman N, Prentki M. AMP kinase and malonyl-CoA: targets for therapy of the metabolic syndrome. Nat Rev Drug Discov 2004;3:340–351.
255. Randle PJ, Garland PB, Hales CN, Newsholme EA. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963;1:785–789.
256. Virchow R. Die Zellularpathologie und ihre Begründung auf physiologische und pathologische Gewebelehre. Berlin: Hirschwald, A; 1858.
257. Tutwiler GF, Brentzel HJ, Kiorpes TC. Inhibition of mitochon-drial carnitine palmitoyl transferase A in vivo with methyl 2–tetradecylglycidate (methyl palmoxirate) and its relationship to ketonemia and glycemia. Proc Soc Exp Biol Med 1985;178:288–296.
258. Rupp H, Jacob R. Metabolically modulated growth and pheno-type of the rat heart. Eur Heart J 1992;13(suppl D):56–61.
259. Bristow M. Etomoxir: a new approach to treatment of chronic heart failure. Lancet 2000;356:1621–1622.
260. Detry JM, Sellier P, Pennaforte S, Cokkinos D, Dargie H, Mathes P. Trimetazidine: a new concept in the treatment of angina. Comparison with propranolol in patients with stable angina. Trimetazidine European Multicenter Study Group. Br J Clin Pharmacol 1994;37:279–288.
261. Kantor PF, Lucien A, Kozak R, Lopaschuk GD. The antianginal drug trimetazidine shifts cardiac energy metabolism from fatty acid oxidation to glucose oxidation by inhibiting mitochon-drial long-chain 3-ketoacyl coenzyme A thiolase. Circ Res 2000;86:580–588.
262. Chaitman BR, Pepine CJ, Parker JO, et al. Effects of ranolazine with atenolol, amlodipine, or diltiazem on exercise tolerance and angina frequency in patients with severe chronic angina: a randomized controlled trial. JAMA 2004;291:309–316.
263. McCormack JG, Barr RL, Wolff AA, Lopaschuk GD. Ranolazine stimulates glucose oxidation in normoxic, isch-emic, and reperfused ischemic rat hearts. Circulation 1996;93:135–142.
264. Nikolaidis LA, Elahi D, Hentosz T, et al. Recombinant gluca-gon-like peptide-1 increases myocardial glucose uptake and improves left ventricular performance in conscious dogs with pacing-induced dilated cardiomyopathy. Circulation 2004;110:955–961.
CAR050.indd 1175CAR050.indd 1175 11/24/2006 12:14:14 PM11/24/2006 12:14:14 PM

CAR050.indd 1176CAR050.indd 1176 11/24/2006 12:14:14 PM11/24/2006 12:14:14 PM