functional diversification of cerato-platanins in ... · 1 functional diversification of...
TRANSCRIPT

Mario R. de O. Barsottini – MPMI
1
Functional diversification of cerato-platanins in 1
Moniliophthora perniciosa as seen by differential 2
expression and protein function specialization* 3
4
Mario R. de O. Barsottini a,b**
, Juliana F. de Oliveirab**
, Douglas Adamoskib**
, Paulo J. P. L. 5
Teixeiraa, Paula F. V. do Prado
a,b, Henrique O. Tiezzi
b, Mauricio L. Sforça
b, Alexandre Cassago
c, 6
Rodrigo V. Portugalc, Paulo S. L. de Oliveira
b, Ana C. de M. Zeri
b, Sandra M. G. Dias
b**, Gonçalo A. 7
G. Pereiraa,b and Andre L. B. Ambrosio
b** 8
9
aDepartamento de Genética e Evolução, UNICAMP, Campinas, SP, Brazil 13083-970 10 bLaboratório Nacional de Biociências, CNPEM, Campinas, SP, Brazil 13083-100 11
cLaboratório Nacional de Nanotecnologia, CNPEM, Campinas, SP, Brazil 13083-100 12
13
*Running title: Cerato-platanins function specialization in M. perniciosa 14
** These authors contributed equally to this work 15
16
To whom correspondence should be addressed: Andre L. B. Ambrosio or Sandra M. G. Dias, Laboratório 17
Nacional de Biociências, CNPEM, Rua Giuseppe Maximo Scolfaro 10000, Polo II Alta Tecnologia, 18
Campinas, SP, Brazil. CEP 13083-100. Tel.: (55) 19 3512 1115; Fax: (55) 19 3512 1004; E-mails: 19
[email protected]; [email protected]. 20
21
Nucleotide sequences were deposited at the GenBank under the accession numbers JX847578 (MpCP6), 22
JX422024 (MpCP7), EU250343 (MpCP8), JX422025 (MpCP9), JX847579 (MpCP10), JX422026 23
(MpCP11), JX422027 (MpCP12), JX422037 (SMpCP1), JX422028 (SMpCP2), JX422029 (SMpCP3), 24
JX422036 (SMpCP6), JX422030 (SMpCP7), JX422031 (SMpCP8), JX422032 (SMpCP9), JX422033 25
(SMpCP10), JX422034 (SMpCP11), JX422035 (SMpCP12), JX422041 (CcCP1), JX422043 (CcCP2), 26
JX422039 (CcCP3), JX422042 (CcCP4), JX422040 (CcCP5), JX422044 (CcCP6), JX422038 (MrCP1), 27
JX426104 (MrCP2), JX426107 (MrCP3), JX426108 (MrCP5), JX426106 (MrCP6), JX426113 (MrCP7), 28
JX426110 (MrCP8), JX426105 (MrCP9), JX426109 (MrCP10), JX426112 (MrCP11) and JX426111 29
(MrCP12). 30
31
The atomic coordinates and structure factors under PDB ID 3SUJ (MpCP1), 3SUK (MpCP2), 3SUL 32
(MpCP3) and 3SUM (MpCP5) are available at the Protein Data Bank. 33
34
35
Page 1 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
2
ABSTRACT 36
Cerato-platanins (CP) are small cysteine-rich fungal-secreted proteins involved in the various stages of 37
the host–fungus interaction process, acting as phytotoxins, elicitors and allergens. We identified 12 CP 38
genes (MpCP1 to 12) in Moniliophthora perniciosa’s genome, the causal agent of Witches’ Broom disease 39
in cacao, and showed that they present distinct expression profiles throughout fungal development and 40
infection. We determined the X-ray crystal structures of MpCP1, 2, 3 and 5, representative of different 41
branches of a phylogenetic tree and expressed at different stages of the disease. Structure-based 42
biochemistry, in combination with nuclear magnetic resonance and mass spectrometry allowed us to define 43
specialized capabilities regarding self-assembling and the direct binding to chitin and N-acetylglucosamine 44
(NAG) tetramers, a fungal cell wall building block, and to map a previously unknown binding region in 45
MpCP5. Moreover, fibers of MpCP2 were shown to act as expansin and facilitate basidiospore germination 46
while soluble MpCP5 blocked NAG6-induced defense response. The correlation between these roles, the 47
fungus life cycle and its tug-of-war interaction with cacao plants is discussed. 48
49
INTRODUCTION 50
The basidiomycete Moniliophthora perniciosa is the causal agent of the Witches’ Broom Disease 51
(WBD) and considered one of the major fungal plagues of cacao crops (Aime and Phillips-Mora 2005). 52
The WBD impacts severely the production of cocoa beans, the raw material of chocolate and other 53
derivatives, resulting in great agro-economic losses throughout the cultivating fields of Central and South 54
America (Evans 2007). In this regard it is urgent to gain knowledge on important proteins for the fungus 55
attack onto the plant. 56
Page 2 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
3
Defense responses in plants upon a pathogen attack usually result in deposition of callose, production 57
of reactive oxygen species, nitric oxide, phytoalexins, along with the transcription of defense-related genes 58
(Desender et al. 2007). These responses are promoted by elicitors, which are recognized by plant receptors. 59
They initiate the microbe-associated molecular pattern (MAMP)-triggered immunity or induce a second 60
layer of defense called the effector-triggered immunity (Jones and Dangl 2006). 61
One family of elicitors spread among the fungal kingdom is the cerato-platanin (CP) family. The 62
founding member of this family and the most studied protein is CP from Ceratocystis platani (Pazzagli et 63
al. 1999), hereon referred to as CpCP. CpCP, recently characterized as a MAMP, induces defense 64
responses and tissue necrosis in plants (Fontana et al. 2008, Lombardi et al. 2013, Pazzagli et al. 1999) and 65
may have a structural role in fungal cell wall formation (Baccelli et al. 2012, Boddi et al. 2004). The 66
tridimensional structure of CpCP was determined by NMR and is similar to proteins that bind to 67
polyssacharides (de Oliveira et al. 2011). The CpCP interacts with chitin (an unsoluble polymer of N-68
acetylglucosamine; NAG) and soluble NAG oligomers (Baccelli et al. 2013, de Oliveira et al. 2011), and 69
although does not bind to cellulose, CpCP loosens it, which could be important during host colonization 70
(Baccelli et al. 2013). Pop1, a CpCP homologue found in C. populicola, shares many of these features. 71
Indeed, it was demonstrated that both, CpCP and Pop1, form soluble ordered aggregates under mild 72
denaturing conditions, which are believed to be important for fungal development and virulence (Baccelli 73
et al. 2013, Carresi et al. 2006, Comparini et al. 2009, Lombardi et al. 2013, Martellini et al. 2012, Pazzagli 74
et al. 2009). Other cerato-platanins present diverse biological activities, such as the induction of systemic 75
acquired resistance (SAR) and induced systemic resistance (ISR) in plants, linked or not to tissue necrosis 76
(Djonović et al. 2007, Frías et al. 2011, 2013, Hall et al. 1999, Vargas et al. 2008, Wilson et al. 2002). Epl1 77
from the beneficial, root-colonizing fungus Trichoderma atroviride, was shown to be able to aggregate in a 78
quick and ordered fashion, although no biological role of such aggregates was presented (Frischmann et al. 79
Page 3 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
4
2013). The antigen CS-Ag from Coccidioides immitis, a human pathogen, has proteolytic activity (Pan and 80
Cole 1995). All these functions attributed to the CP proteins make them an attractive target to control the 81
pathogen attack. 82
The recent completion of M. perniciosa’s genome sequencing allowed us to identify an astonishing 83
high number of CP-coding genes, more precisely, 12 in total (hereon referred to as MpCP1 to 12). In this 84
manuscript, we show that they are clearly differentially expressed throughout the several stages of the 85
disease, as well as the different stages of the fungus development. Accordingly, the co-expressed proteins 86
clustered into distinct branches of an all-encompassing phylogenetic tree, indicating the presence of groups 87
with higher sequence similarity and likely functional proximity. In order to test this hypothesis, we 88
heterologoulsy expressed, purified and determine the crystallographic structure of four representative 89
members of the distinct phylogenetic clusters: MpCP1, which is exclusively expressed in basidiocarps, 90
MpCP2 and 3, both detected in the fast growing mycelium and necrotic infected seeds and fruits, and 91
MpCP5, majorly detected during the slow growth in the apoplast, where plant attack is set. Curiously, 92
MpCP5 structure was found to be essentially different from any other determined to date. The X-ray crystal 93
structures guided the subsequent investigation on MpCP’s capacity of binding to fungus and plant cell wall 94
components, as well as to self-aggregate into amyloid-like filaments. In this sense, NMR analysis showed 95
that MpCP3 and 5 distinctly bind to N-acetylglucosamine tetramers (NAG4) with different affinities and 96
allowed the description of an unforeseen binding interface in MpCP5. The MpCP5 presence was sufficient 97
to block NAG6-induced defense response in tobacco seedlings, representing the first CP shown to 98
counteract the chitin fragment plant sensitization. Finally, we show that MpCP2 and 3 are the most 99
responsive to self-aggregation into amyloid-like fibrils, with the fibrils of the former acting as expasins and 100
also promoting germination tube growth. MpCP5`s and MpCP2`s described biological activities are in 101
Page 4 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
5
accordance with their expression profile during the host-fungus interaction and exemplify the MpCP`s 102
protein function specialization during the Witches’ Broom disease of cacao. 103
RESULTS 104
M. perniciosa possesses multiple differentially expressed cerato-platanin genes - M. perniciosa has a 105
hemibiotrophic life cycle starting with meristem- and young fruits-infecting basidiospores that induce 106
hyperplasia and irregular branching of actively growing tissues of the cacao tree. During this stage, called 107
biotrophic phase, hyphae are formed in the host’s intercellular space. The disease then progresses to a 108
necrotic stage, during which the fungus grows intracellularly, resulting in the death of the plant tissues 109
(giving rise to the dry-broom aspect of the branches) and the development of fruiting bodies (necrotrophic 110
phase) (Meinhardt et al. 2008). 111
In addition to the five CPs originally described by Zaparoli and colleagues (2009) in the fungus M. 112
perniciosa (MpCP1 to MpCP5, NCBI Genbank accession codes EU250339, EU250340, EU250342, 113
EU250344 and EU250345, respectively), we present hereby seven new genes. It is yet worth mentioning 114
that the CERAT gene, previously identified by Rincones and colleagues (2008), corresponds to MpCP3. 115
An Aspf13 homologue was also described, and verified by us to match the MpCP2 gene (Leal et al. 2010). 116
Given the new apparent redundant number of isoforms (MpCP1 to 12), we first set out to analyze their 117
individual mRNA expression patterns throughout the different stages of the disease. RNA-seq data from 118
green-house grown cacao plants infected with M. perniciosa revealed that MpCPs 4, 5, 11 and 12 are 119
prominently expressed in the green parts-infecting fungus (biotrophic phase). Their gene expression levels 120
decrease as the disease progresses, until complete disappearance when the dry broom stage is reached (Fig. 121
1A). Secondly, MpCP2 and 3 are virtually the only two isoforms present in M. perniciosa from field-122
collected, infected fruits (Fig. 1B) and in the mycelium of in vitro lab-cultivated fungus (Fig. 1C). MpCP1 123
is basically the only isoform detected during basidiocarp formation, while MpCP4 and 11 dominated the 124
Page 5 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
6
basidiospore phase. MpCP6, 7, 8, 9 and 10 presented no significant gene expression throughout all the 125
samples and stages analyzed. Mass spectrometry analysis of M. perniciosa’s culture medium of in vitro 126
grown necrotrophic mycelium was further performed to allow the direct identification of the proteins. 127
Accordingly, MpCPs 2, 3 and 7 proteins (supplemental Table S1A) were identified, indicating that all other 128
MpCPs are likely only necessary during the fungus-cacao interaction. Glycosylation prediction using the 129
NetNGlyc 1.0 online server revealed that MpCPs 1, 2, 4, 5 and 11 contain one putative glycosylation site 130
(NxS/T), which is equivalent to the one described for the protein Sm1 as important to modulate protein 131
dimerization and plant recognition (29NGS31; Vargas et al. 2008). Additional putative glycosylation sites 132
were also identified for these MpCPs, which can be found in supplemental Table S1B. Two out of the three 133
spectrometry-identified MpCP2 peptides spaned the SM1-homologue glycosylation site indicating that, if 134
this protein is glycosylated on this segment, it is only partially. 135
Phylogenetic analysis of M. perniciosa’s cerato-platanin proteins - The clear differential expression 136
profiles of MpCP transcripts leaded us to speculate whether these isoforms might have distinct roles during 137
the fungus development or the plant infection process. In order to evaluate the evolutionary relationships 138
between the homologous sequences and gain insights on their specific roles, we reconstructed the cerato-139
platanin phylogenetic tree using deposited homologous sequences (Fig. 2). The group of sequences 140
included recently deposited genes from other cacao-infecting fungus, Moniliphthora roreri (MrCP1 to 10), 141
as well as the M. perniciosa biotype S, which infects plants of the Solanaceae family (Marelli et al. 2009) 142
(SMpCP1 to 3 and 6 to 12), and the saprotrophic Crinipellis campanella (CcCP1 to 6), representing a 143
saprotrophic non-pathogenic ancestral of the Moniliophthora genus (Aime and Phillips-Mora, 2005). The 144
sequences clustered in three major groups. One group contains CP homologues from ascomycetes and is in 145
accordance with previously published data (Seidl et al. 2006), regarding the clustering of the genes Sm1 (T. 146
virens), Epl1 (Hypocrea atroviridis), SP1 (Leptosphaeria maculans) and Snodprot1 (Phaeosphaeria 147
Page 6 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
7
nodorum), among others, within the so-called Epl1/Epl2 cluster (branches within light red box in Fig. 2). 148
MpCP4, 5 and 11, as well as homologues from the basiodiomycetes Pleurotus ostreatus and Coprinopsis 149
cinerea determined the MpCP Cluster 1 (Mper1; blue box in Fig. 2). On the other hand, MpCP2, 3, 7, 8, 10 150
and 12 grouped within the MpCP Cluster 2 (Mper2; green box in Fig. 2) along with saprotrophic C. 151
campanella’s CPs. 152
Mper clusters 1 and 2 are relatively distant in the evolutionary tree and, in accordance, comprise 153
MpCP proteins that are co-expressed during different stages of the fungus development. Specifically, 154
MpCP4, 5 and 11 (Mper1) are co-expressed during the fungus-cacao interaction (in the green/living parts 155
of the plant) as well as during basidiospore germination, while MpCP2 and 3 (Mper2) are highly expressed 156
only in the necrotic infected fruit and in vitro grown mycelium. Lastly, MpCP9, not significantly expressed 157
in any of the cases analyzed, clustered with several putative CP homologues from basidomycetes. MpCP1 158
and 6 did not significantly cluster into the cited groups. 159
This separation may suggest that MpCP4, 5 and 11 are important for the fungus-plant interaction 160
process and that MpCP2 and 3, as well as MpCP1, expressed in the basidiocarp, may have important 161
developmental roles for the growth in a nutrient-rich environment. The combination of the above 162
mentioned data led us to conclude that the CPs found in the M. perniciosa genome may have different roles 163
important for distinct aspects of the WBD. 164
X-ray structures - To unravel the different functions of the MpCPs, we solved the crystal structure of 165
selected recombinant MpCPs expressed during the infection of green parts (MpCP5) and fruits (MpCP2 166
and 3), as well as MpCP1, the major isoform during basidiocarp formation. Different constructs of a 167
recombinant MpCP4 were also tested for solubility after E. coli expression, but all failed in producing 168
soluble protein. The parameters and statistics for both data collection and processing and the final refined 169
models are presented in supplemental Table S2. 170
Page 7 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
8
First, MpCP1 structure was solved by single-wavelength anomalous dispersion using the signal from 7 171
intrinsic sulfur atoms (5 cysteine and 2 methionine residues). The obtained model was then employed as 172
search model for solving MpCP2 and MpCP3 structures by molecular replacement. MpCP5 could only be 173
solved by molecular replacement using the elicitor Sm1 from T. virens (PDB ID: 3M3G) as search model. 174
Regardless, the core fold of all MpCPs consists of a six-stranded antiparallel β-sheet flanked by six α-175
helices and loops, forming a double Ψβ-barrel (Fig. 3A). Also, all four structures presented the two 176
disulfide bridges expected for this family of proteins (supplemental Fig. S1). The overall MpCP1, 2 and 3 177
structures are highly conserved, with an average backbone r.m.s.d of 0.86 Å, and an average sequence 178
identity of 49.4%, as calculated by Coot (Emsley et al. 2010) (Fig. 3A). The major conformational 179
variability between these three structures reside in a β-hairpin loop between the strands 3 and 4, along with 180
some small divergences among other loops. MpCP5 sequence is only 25% identical in average to MpCP1, 181
2 and 3, and this is reflected in an overall higher core r.m.s.d. of about 1.33 Å. MpCP5 has an extra α-helix 182
(labeled as α2*), and the β-strand β6 previously identified on the other MpCPs breaks down into two 183
shorter β-strands (termed β6 and β6*). Moreover, MpCP5 presents a shorter β-hairpin between the strands 184
3 and 4 along with some other major conformational variability among the loops (Fig. 3B). Five strong 185
peaks (over 7σ in height) were observed in the Fourier difference electron density map for MpCP1, which, 186
with respect to their contributions to the dispersive component of the scattering factor, were interpreted as 187
zinc, sodium, and chloride ions (supplemental Fig. S2). All of these ionic species are found in high 188
concentration in the crystallization solution, with the exception of the chloride which was present in the 189
protein purification solution. No other extraneous density was found in the electron density maps of 190
MpCP2, 3 and 5. 191
All the structures were analyzed using the DALI server (Holm and Rosenström 2010), as a mean of 192
identifying similar folds and infer about their functions. Using a Z-score cutoff of 10, the best structural 193
Page 8 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
9
matches for all MpCPs were the elicitor Sm1 from T. virens (PDB ID: 3M3G) and CpCP from 194
Ceratocystis platani (PDB ID: 2KQA). Overall, all MpCPs superposed well to both Sm1 and CpCP, with 195
r.m.s. deviations ranging between 1 and 1.3 Å for the Cα, indicating very high structural conservation 196
between them. Still, by the same cutoff criteria, the DALI search identified two expansins, the bacterial 197
EXPB1 (Yennawar et al. 2006) (PDB ID: 2HCZ) and EXLX1 from corn plant (Kerff et al. 2008) (PDB ID: 198
3D30), as well as the pollen allergen Phlp1 from the grass Phleum pratense (unpublished data) (PDB ID: 199
1N10). CpCP was shown to interact with polymeric chitin oligomers of NAG, a sugar constituent of fungus 200
cell wall (Baccelli et al. 2013, de Oliveira el at. 2011). EXPB1 binds to maize cell walls, most strongly to 201
xylans, causing loosening of the plant cell wall (Yennawar et al. 2006). EXLX1 binds to cellulose and 202
peptidoglycan, but it lacked catalytic activity against these substrates (Kerff et al. 2008). Therefore, the 203
structural similarities clearly suggest that the MpCPs may bind sugar molecules and that this interaction 204
may be important for the plant-pathogen interaction. 205
NMR titration shows that NAG4 binds to distinct sites of MpCP3 and 5 - We next tested the ability of 206
the MpCP1, 2, 3 and 5 to binding NAG monomer and tetramer (NAG4), as well as determined the 207
dissociation constants of these sugars, by NMR spectroscopy. NAG monomer at 10 mM and a range of 208
NAG4 concentrations, from 100 µM to 10 mM, were titrated against 50 µM of 15N-labelled MpCP1, 2, 3 209
and 5, while monitoring the chemical shift and line width changes in the 15N-HSQC spectra. Even a 200 210
times molar excess of NAG did not cause any perturbations to the 15N-HSQC spectra of all tested MpCPs, 211
(supplemental Fig. S3A). However, when tested in the presence of NAG4, both 15N-MpCP3 and 15N-212
MpCP5 spectra were clearly perturbed. No significant changes were observed in the spectra of 15N-213
MpCP1, while 15N-MpCP2 was slightly disturbed in the presence of NAG4 (supplemental Fig. S3B). 214
By triple-resonance experiments we identified the NAG4-binding surface and calculated the binding 215
affinity to NAG4 for both MpCP3 and MpCP5. Peaks corresponding to residues Phe32, Thr34, Ala36, 216
Page 9 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
10
Cys37, Asp39, Trp68 and Asn69 in MpCP3, are those which have undergone the most perturbation upon 217
NAG4 binding (Fig. 4A, left panel; supplemental Fig. S3C), based on a chemical shift cut-off criteria of ∆δ 218
(15N+1H) ≥ 0.05 ppm. These residues are part of a region that is structurally similar to the one previously 219
described for CpCP (de Oliveira et al. 2011) and lies mainly within the helix α2 and the loops between 220
helices α2 and α3 and β-strands β2 and β3. The estimated Kd for MpCP3:NAG4 interaction, based on the 221
chemical shifts of eight consecutives residues, according to previously published methodology (Lehotzky 222
et al. 2010), was 24.7 ± 12.8 mM (Fig. 4B, upper panel). Strikingly, the MpCP5 peak assignment showed a 223
rather extended, completely unforeseen, NAG4-interacting surface on this protein. The mapped region, as 224
judged by chemically shifted and signal-faded (cut-off of -0.5) peak residues, is confined mainly at the C-225
terminus portion of MpCP5 (Fig. 4A left panel; supplemental Fig. S3D-E). Accordingly, NAG4 binds to 226
MpCP5 with a higher estimated affinity as judged by a Kd of 6.6 ± 1.7 mM (Fig. 4B, lower panel). These 227
results show that MpCP3 binds NAG4 through a conserved surface regarding the CpCP. MpCP5, on the 228
other hand, binds to NAG4 through an unforeseen surface and with a higher affinity. 229
To get a picture of the putative binding mode of NAG4 to MpCP3 and 5, we performed computational 230
blind docking. The MpCP3:NAG4 lowest energy model obtained showed a binding surface that matched 231
the residues disturbed by NAG4 interaction, as verified by the NMR experiments (Fig. 4A and 4C, left 232
panels). On the other hand, the MpCP5:NAG4 second lowest energy model indicated that NAG4 may bind 233
to part of the otherwise large disturbed surface measured by NMR. The results indicated that residues 234
Thr34, Ala36, Asp39, Trp68 from MpCP3 (Fig. 4C, left panel) and His92, Asn116 and Thr119 from 235
MpCP5, among others, (Fig. 4C, right panel) are likely directly involved in the sugar-protein interaction 236
through polar and non-polar contacts. To further confirm this finding, we double-mutated MpCP3 residues 237
Ala36 to a Thr and Asp39 to a Gly, as well as did a single Trp68 to Tyr mutation and tested the wild type 238
and mutant proteins for NAG4 binding capacity as judged by shifting on the proteins thermal melting 239
Page 10 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
11
points (supplemental Fig. S4). Accordingly, all the MpCP3 mutations affected their interaction to NAG4. 240
Similarly, we mutated MpCP5 Tyr88 to Ser, His92 to Ala, Asn116 to Lys and Thr119 to either Ser or Ala. 241
Only Asn116Lys mutation apparently increased protein affinity for NAG4, indicating that the 242
MpCP5:NAG4 binding surface is rather large and likely involves the simultaneous interaction of more than 243
one molecule. 244
MpCP5 blocked the perception of NAG6 by the plant. It has been reported that chitin fragments - 245
generated by the action of plant chitinases against the fungus cell wall during the early stages of infection - 246
can act as MAMPs, eliciting immune responses in plants (Boller 1995, de Jonge et al. 2010). MpCP5 is 247
expressed while the fungus is interacting with the living parts of the plant, when a host response to the 248
pathogen attack is expected. In order to evaluate the functional significance of the NAG oligomer-MpCP5 249
interaction we exposed tobacco plants to the chitin fragment NAG6 as well as NAG6 pre-incubated with 250
MpCP5, and investigated, by quantitative PCR, the expression of defense-related genes (Fig. 4D). As 251
expected, plant responded to the NAG6 presence by increasing expression of tpa1 (phenylalanine ammonia 252
lyase), involved on phytoalexin synthesis pathway and SAR activation (Gayoso et al. 2010), npr1 (non-253
expresser of PR genes 1), a transcription factor pivotal for the activation of SAR (Dong 2004), pr-4a and 254
pr-5 (pathogenesis-related) genes, both described as displaying antifungal activity (Guevara-Morato et al. 255
2010, van Loon et al. 2006). The pre-incubation of MpCP5 with the NAG6 fragments reduced the response 256
levels back to control (just buffer), suggesting that MpCP5 protein can help blocking plant defense 257
response by scavenging released chitin fragments. 258
The MpCPs neither have glycolytic activity against nor bind to plant cell wall components - Similarly 259
to CpCP and the bacterial expansin EXLX1, the MpCPs presents structural conservation with the catalytic 260
domain found in family 45 of glycosyl hydrolases and in the MltA family of lytic transglycosylases 261
(supplemental Fig. S5) (de Oliveira et al. 2011, Kerff et al. 2008). Nevertheless, tests for hydrolytic activity 262
Page 11 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
12
of MpCPs 1, 2, 3 and 5 against some common plant cell wall components and other polysaccharides, as 263
well as cacao cell wall extract (CCW), all proved negative (Fig. 5A). Given that MpCP3 and 5 were shown 264
to bind to the soluble tetrameric form of NAG, we questioned whether these proteins would be able to bind 265
to insoluble chitin polymers via pull-down assay, therefore mimicking a possible interaction with fungal 266
cell wall. The pull-down assays were also performed against the CCW. As described for the CpCP protein 267
(Baccelli et al. 2013), MpCP1, 2, 3 and 5 did not bind to the plant cell-wall (Fig. 5B) but they were all (on 268
both soluble - Fig. 5C - and aggregated forms - data not shown) pulled-down by chitin polymers, with 269
MpCP3 apparently binding the strongest. 270
MpCP2 and 3 have higher propensity to form soluble aggregates that induces Thioflavin T 271
fluorescence - The MpCPs-highly related protein CpCP resembles another class of fungal-secreted proteins 272
known as hydrophobins, which have many distinct roles in fungal development and in pathogen–host 273
interactions (Pazzagli et al. 2009, Whiteford and Spanu 2002). In this regard, we studied the MpCP1, 2, 3 274
and 5 self-assembling properties. A range of 0.1 mM up to 3 mM of each MpCP was incubated for 30 days 275
at 37°C under low pH treatment (pH 3.0), as previously described for the CpCP protein (Pazzagli et al. 276
2009), and the Thioflavin T (ThT) fluorescence signal monitored. The existence of a threshold 277
concentration below which aggregation does not occur is an important feature associated with amyloid 278
fibrils (Lomakin et al. 1996). As seen in Fig. 6A, the MpCP2’s ThT fluorescence readings drastically 279
increased (over 15 times in comparison to control) from 0.4 to 0.8 mM of protein concentration. MpCP3 280
and MpCP5, although to a smaller extent, also induced significant ThT fluorescence at 0.8 mM protein. In 281
comparison to the other analyzed MpCPs, MpCP1 showed low levels of ThT-positive aggregates at only 282
above 1.5 mM of protein concentration. The initial aggregation step is commonly known to start from a 283
partly unfolded monomeric intermediate which presents increased surface hydrophobicity (Murphy 2007). 284
Accordingly, the fluorescence intensities of the probes 1,8-ANS and bis-ANS, two hydrophobic dyes that 285
Page 12 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
13
bind to the solvent-exposed hydrophobic surfaces were greatly enhanced upon interaction with 0.8 mM 286
MpCP1, 2, 3 and 5 aggregates (Fig. 6B). 287
In light of these results we performed a time course study of protein aggregation of 0.8 mM of MpCP2 288
and MpCP3, at 37°C in low pH solution. The kinetic curves of the ThT fluorescence intensity are 289
consistent with a nucleation-dependent polymerization model commonly observed for proteins that can 290
form amyloid fibrils (Lomakin et al 1996). While MpCP2 took 3 days to reach a signal plateau (Fig. 6C), 291
MpCP3 showed similar pattern, but only after 25 days of incubation. 292
Biophysical and functional characterization of the soluble aggregates - The secondary structure 293
changes accompanying MpCPs aggregation (at a protein concentration of 0.8 mM) were analyzed by far-294
UV circular dichroism (CD) spectroscopy after 5, 10 and 20 days of low pH incubation at 37 °C. In as early 295
as 5 days, it was possible to detect a loss of the MpCP1 native conformation and the acquisition of a 296
random coil structure, with a negative band at 200 nm (Fig. 7A), suggesting protein unfolding, as further 297
confirmed by NMR spectroscopy (supplemental Fig. S6, left panel). Curiously, the highly ThT-responsive 298
MpCP2, as well as MpCP3 and 5, did not present characteristic random-coil spectra, neither a shift from α-299
helix to β-sheet content, as commonly found in β-amyloid fibers. This is somewhat in agreement with a 300
previous report, in which the CpCP did not display conformational changes corresponding to an increase in 301
β-structure, being rather unfolded or acquiring a new secondary structure pattern (Pazzagli et al. 2009). 302
Globular proteins under equilibrium conditions may have at least two different partially folded 303
conformations, the molten globule and its precursor, the pre-molten globule, both of which may potentially 304
play a role as the crucial amyloidogenic species (Uversky and Fink 2004). The most characteristic feature 305
of the amyloidogenic pre-molten globule CD spectra is a large negative ellipticity at 200 nm and low 306
ellipticity in the vicinity of 222 nm, with native/molten globule presenting combination of opposite 307
situations and the unfolded proteins showing the largest negative values at 200 nm and ellipticity in the 308
Page 13 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
14
vicinity of 222 nm close to zero (Uversky and Fink 2004). According to a survey of CD spectra of several 309
proteins captured on different states, the MpCP2, 3 and 5 CD profiling, as wells as their MRE[θ]200 x 310
MRE[θ]222 combination, shows that the mild denaturing treatment likely brought the protein to a molten 311
globule state (Fig. 7B), while MpCP1 positions closer to coordinates more characteristic of an unfolded 312
state. 313
Dynamic light scattering analysis of MpCP1, 2, 3 and 5 before and after 15 days of incubation in low 314
pH solution confirmed the formation of aggregates of protein (Fig. 7C). Since MpCP5 was one of the 315
proteins with the smallest ThT signal we further characterized its aggregation process by RMN. Two-316
dimensional 15N-HSQC spectra indicated that, after 4 days of incubation at 37oC at low pH, the great 317
majority of peaks either had a decreased intensity or disappeared, a clear indication of oligomerization 318
process (Cavanagh et al. 2007) (supplemental Fig. S6, right panel). The filamentous nature of the 319
aggregates was visually confirmed for MpCP2, by using transmission electron microscopy (Fig. 7D). 320
The Ceratocystis platani CP self-aggregation correlates to a fragmentation process, so as for some 321
amyloid-like proteins which undergo self-assemblage after a cleavage of the polypeptide chain (Mishra et 322
al. 2007, Pazzagli et al. 2009, Tsiroulnikov et al. 2006). The identification of the cleavage sites and 323
aggregating peptides of MpCP1, 2, 3 and 5 was done by mass spectrometry and are in clear accordance 324
with what was described for CpCP (Pazzagli et al. 2009) (supplemental Fig. S7, S8 and Table S3). 325
Finally, we further investigated the biological relevance of the aggregation processes by assessing the 326
capability of aggregated MpCP2 of loosening cellulose. Soluble CpCP and Pop1 were shown to fragment 327
cellulosic substrates through a non-enzymatic mechanism (Baccelli et. al, 2013). We verified that 328
aggregated MpCP2, but not its soluble monomeric form, promotes cellulose fragmentation which may 329
implicate a role of this protein’s aggregates during the interaction of M.perniciosa with the cocoa (fixation 330
to the substrate and nutrient acquisition) (Fig. 7E). Aggregated MpCP2 also potentiated the growth M. 331
Page 14 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
15
perniciosa basidiospore’s germination tube (Fig. 7F), a phenomenon which may be related to the fungus’ 332
hyphal development. Curiously, as shown in Fig 1B and C, MpCP2 is highly expressed in the in vitro-333
grown mycelium and in the fungus of infected fruits, situations where a fast grown is propitiated by the 334
nutrient rich environment. 335
336
DISCUSSION 337
M. perniciosa, a fungus with a hemibiotrophic lifestyle, presents twelve copies of CP-coding 338
sequences in its genome. Different genetic evolutionary models predict an expansion in a gene’s copy 339
number before their specialization or the rise of new functions, also accounting for an overlap of functions 340
due to a momentary intermediate state of specialization (Bergthorsson et al. 2007). In this work we show 341
that the MpCPs (1 to 12) are expressed at particular stages of the Witches’ Broom disease, and, based on 342
our data and on the literature above, we reason that M. perniciosa evolved different proteins to perform 343
different, specialized roles during its hemibiotrophic life cycle. 344
The biotrophic stage of M. perniciosa is unusually extended, lasting for 2–3 months in the plant’s 345
infected tissue and creating a nutrient sink to the infection site, so that the host is disadvantaged but not 346
killed (Scarpari et al. 2005). During this stage, preventing the elicitation of the host’s immune response is 347
key for the fungus survival. None of the purified MpCPs caused necrotic symptoms or phytoalexin 348
induction on tobacco leaves injected with these proteins (supplemental Fig. S9), in accordance with results 349
obtained with CtCP1, the CP from Colletotrichum truncatum, another hemibiotrophic pathogen, which 350
protein also did not induce defense responses in planta (Bhadauria et al. 2011). This may suggest an 351
evolutionary pressure to reduce the defense-eliciting ability of CP proteins in hemibiothrophic life styles. 352
However, our findings are in contrast with a previous report (Zaparoli et al. 2009) which showed MpCP1`s 353
necrotic activity on tobacco and cocoa leaves. It is possible that the observed differences are due to the 354
Page 15 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
16
presence of bacterial contaminants on their sample, since we added a further purification step in our protein 355
preparation. 356
Gene transcripts of MpCP4, 5 and 11, from the cluster Mper1 of the presented phylogenetic tree, were 357
exclusively detected in the slow-growing biothrophic phase, when the fungus develops in the apoplast of 358
the plant and where the pathogen-host survival battle is established. Using NMR, we have shown that 359
recombinant MpCP5 binds to NAG4 with the highest affinity via a previously unidentified binding site. 360
This interface, in MpCP5, is located around a much shorter β-hairpin formed between sheets β3 and β4, 361
when compared to MpCPs 1 and 2. The large protein surface disturbed by NAG4 opens the possibility that 362
either the protein goes through a profound conformational change due to the sugar interaction or multiples 363
binding sites are existent. Gene expression data showed that MpCP5 is capable of blocking the plant 364
response defense to the chitin fragment NAG6, a potent defense elicitor released during fungal colonization 365
(Boller 1995). Taking together, these results create a potential link between MpCP5`s biotrophic phase 366
expression, its NAG oligomer-binding property and its capacity of suppressing the plant immune response. 367
Likewise, MpCP4 and 11 may also be important in protecting the fungus against the cacao, since most of 368
the key residues we identified for NAG4 binding in MpCP5 are conserved on these proteins. Thus far, the 369
CPs have been implicated as fungal elicitors and/or effectors, either promoting the virulence of pathogenic 370
fungi, or enhancing the plant’s resistance against other microorganisms (Djonović et al. 2007, Frías et al. 371
2011, 2013, Jeong et al. 2007, Pazzagli et al. 1999 Vargas et al. 2008). Although de Oliveira and 372
colleagues (2011) have hypothesized that CPs might act as defense suppressors by scavenging free chitin 373
fragments, we present hereby a experimental evidence which suggest such thing, bringing a potential new 374
feature to this class of protein. 375
The MpCPs belonging to the phylogenetic cluster Mper2 (MpCP2, 3, 7, 8, 10 and 12) positioned 376
together with homologues from the saprothrophic C. campanella and their gene transcripts were detected 377
Page 16 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
17
during the late and necrotic stages of the cacao fruits infection, as well as in the in vitro grown mycelium. 378
Worth of note, MpCP3 was previously detected in our laboratory in an in vitro-grown biotrophic-like 379
mycelium (Rincones et al. 2008), and the gene fragment was identified as CERAT by then. Even though 380
the biotrophic-like mycelium used by Rincones and colleagues presented biotrophic morphological 381
characteristics (thick, monokaryotic hyphae without clamp connections), further studies in our laboratory 382
showed that it also expressed some of the genes typically found in the necrothrophic stage of the disease 383
(data not shown), probably due to the use of cocoa extract-containing conditions (nutrients which are 384
usually only available during necrotrophic phase). 385
NMR data showed that MpCP3 also binds to NAG4, however with a lower affinity when compared to 386
MpCP5. MpCP3 shares a similar NAG4-binding surface to the one previously described for CpCP as 387
confirmed by site-directed mutagenesis and thermal shift assays. Computational docking allowed the 388
determination of an interface involving the diametrically opposed loops between β2 and β3 (which is much 389
larger than its equivalent in MpCP5), and helices α1 and α2. The sequence composition of these loops in 390
MpCP3 is not only distinct from those in MpCP5, but it also diverges between MpCP1 and 2, thus 391
explaining why the later two did not bind significantly to NAG4. The three residues directly involved in 392
MpCP3 binding to NAG4 (Ala36, Asp39 and Trp68) are fully conserved on both MpCP7 and 9, suggesting 393
that these proteins also bind to NAG4. Worth of noting, similarly to MpCP3, MpCP7 and 9 are expressed 394
on the rotten infected fruits/plant and lab-cultivated fungus, both representative of the fast-growing 395
necrotrophic stage. 396
The CPs from C. platani and C. populicola have been shown to self-aggregate under mild-denaturing 397
conditions, such as acetic acid and TFE solution, as well as exposure to hydrophobic surfaces (Martellini et 398
al. 2012, Pazzagli et al. 2009). The protein Epl1 presented spontaneous quick assembling but no further 399
characterization of the aggregates was performed (Frischmann et al. 2013). Another protein family present 400
Page 17 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
18
in fungi that is able to enhance ThT fluorescence after its assemblage is the hydrophobins (HPB) (Wösten 401
and de Vocht, 2000). Rodlets formed by aggregated hydrophobins first lower the surface tension at the 402
water–air interface and are important for fungal development and virulence by creating amphipathic layers 403
that facilitate hyphal growth and adhesion, as well as the formation of haustoria and fruiting bodies. 404
Similarly to the CpCP and MpCPs, HPBs have intramolecular disulfide bonds, are secreted as monomers 405
and self-aggregate under favorable conditions (Whiteford and Spanu 2002). All four tested MpCPs were 406
capable of self-aggregation after incubation in low pH solution. MpCP1, the one with the slowest 407
aggregation kinetics, presents levels comparable to CpCP (Pazzagli at al. 2009), while MpCP2, followed 408
by MpCP3, are the most responsive, as qualitatively judged by the kinetics curves of ThT fluorescence. 409
In physiological environments, where many processes are dictated by interface phenomena, the 410
presence of a surface (such as the host cuticle, cell wall and/or membrane) may provide the necessary 411
perturbation to induce the conversion from the soluble conformation to the self-assembling conformation 412
(Martellini et al. 2012, Moores et al. 2011, Murphy 2007). The native globular structure of several proteins 413
has been shown to be destabilized to pre-molten/molten globule-like structures by a myriad of different 414
agents, e.g. high temperatures, organic solvents, low or high pH, low to mild concentrations of strong 415
denaturants, and others, which may significantly accelerate the rate of fibril formation (Uversky and Fink, 416
2004). Another aspect of CpCP and other aggregating proteins mirrored by the MpCPs is the self-cleavage 417
process as a prerequisite for the process (Mishra et al. 2007, Pazzagli et al. 2009, Tsiroulnikov et al. 2006), 418
probably enabling the exposure of hydrophobic regions of the protein that can then establish the first 419
intermolecular contacts needed for self-assembling. 420
We propose that the chitin binding property of both MpCP2 and MpCP3, along with their and MpCP1 421
self-aggregation characteristic, all proteins expressed during fast-growing phases of the fungus, very likely 422
facilitate the process of hyphal growth, fruiting body formation and substrate adhesion. Furthermore, 423
Page 18 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
19
MpCP2 aggregates, but not the soluble protein, revealed expansins characteristics as it was able to promote 424
cellulose fragmentation. Plant cell wall (rich in cellulose) loosening activity may be important for host 425
colonization during fungal infection, but also for growth on dead plant tissue and nutrient acquisition. 426
Moreover, MpCP2 aggregates were also shown to contribute to germination tube formation and are likely 427
to be important for the fungus development. Although not tested, MpCP3 binding surface to NAG4 (which 428
is distinct to MpCP5 surface) may be important for the chitin binding (this isoform was the one with higher 429
qualitative affinity to chitin), fungus cell wall remodeling and growth. 430
M. perniciosa presents a well-characterized, yet complex, lifestyle (Meinhardt et al. 2008). The 431
existence of a biotrophic phase, followed by the progression to a necrotrophic stage, requires the fungus to 432
thrive at the most diverse physiological environments inside its host. Consistently, we propose, for the first 433
time, a differentiation model for cerato-platanins in an organism, based on protein functional specialization. 434
M. perniciosa is the only basidiomycete phytopathogen among all previously studied CP-bearing 435
organisms and, overall, the results provided in this work may help constitute a useful platform for 436
compound design which can block the protein activity and help to compose a combat approach against the 437
disease. 438
439
MATERIALS AND METHODS 440
Genome survey - To identify Cerato-platanin orthologous genes in the M. perniciosa biotype C 441
genome (Mondego et al. 2008) we searched for sequences classified within the InterPro ID IPR010829 442
(Hunter et al 2012). Five genes (MpCP1 to 5) had been previously described (Zaparoli et al. 2009). Seven 443
new ones were further identified and named MpCP6 to MpCP12. The open reading frames (ORFs) of all 444
MpCP from M. perniciosa biotype C genes were confirmed by cDNA sequencing and deposited at the 445
GenBank (refer to the first page and the results for accession numbers). To identify the CP’s sequences 446
Page 19 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
20
from the isolate MCA2977 of M. roreri and the isolate APS1 of M. perniciosa biotype S, we used genomic 447
and RNA-seq data (unpublished data). CP’s introns were found through the alignment between RNA-seq 448
and genomic derived sequences of the M. perniciosa biotype S and M. roreri. Since there is no expression 449
data for the C. campanella, its CP`s genomic sequences were evaluated for intron presence with the aid of 450
the software Augustus Gene Finder v2.3 (Stanke et al. 2004) trained with genes of C. cinerea and ESTs 451
from M. perniciosa, as previously described (Mondego et al. 2008). The software SignalP 4.0 (Petersen et 452
al. 2011) was used to predict the presence of secretion signal peptide in all of the encoded proteins. 453
RNA-seq - The reproductive structures of M. perniciosa (basidiomata) were produced following the 454
procedures described (Pires et al. 2009) and were used to collect basidiospores directly in the collection 455
solution (16% glycerol, 0.01M MES, 0.01% Tween, pH 6.1) as described elsewhere (Frias et al. 1995). 456
Theobroma cacao var. Comum was cultivated in a greenhouse under controlled temperature and humidity 457
conditions (from 22 to 28 °C and above 50%, respectively). Two-month old seedlings were infected with 458
Moniliophthora perniciosa basidiospores as previously described (Frias et al. 1995) and RNA was 459
extracted from infected parts of the plant at representative time-points of the disease, from the green broom 460
to complete necrosis. Infected fruits from Theobroma cacao trees var. Comum were collected in a cacao 461
farm located in Ilhéus, Brazil. Three stages of the disease were analyzed: early stage (which is 462
characterized by disordered fruit ripening), partial necrosis stage and rotten fruit stage. The seeds and pods 463
of each fruit were collected independently for RNA extraction. For RNA extraction of M. perniciosa 464
basidiospores, 1 mL of a suspension containing approximately 107 spores/mL was gently centrifuged at 465
1000xg for 5 minutes to precipitate the basidiospores, which were resuspended in 1 mL collection solution 466
(which prevents the germination), or ressuspended in 1 mL of sterile water (to allow germination). Both 467
aliquots were transferred to small beakers, and incubated at 28°C and 120 rpm for 4 hours. The 468
germination of the basidiospores was microscopically verified prior to RNA extraction. In vitro grown 469
mycelium or mature basidiomata were pooled and used for RNA extraction. 470
Page 20 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
21
For each sample, 10 µg total RNA were used to prepare the mRNA-seq library according to the 471
protocol provided by Illumina. Library quality control and quantification were performed with the Experion 472
Automated Electrophoresis System (Bio-Rad) and the Qubit fluorometer (Invitrogen), respectively. Each 473
library was sequenced in one lane of an Illumina Genome Analyzer IIx sequencer to produce 36 bp single-474
end reads. The RNA-seq reads were aligned against a reference comprised of 17.008 gene models of M. 475
perniciosa using the program Bowtie (Langmead et al. 2009). The alignment was performed by allowing 476
up to two mismatches, and excluding reads that mapped to more than one position in the reference. The 477
number of reads mapped to each gene was divided by the size of the gene (in kilobases) and then 478
normalized by the total mapped reads of the library (in millions of reads). Thus, the expression value of 479
each gene was given in RPKM (mapped reads to a gene per kilobase per million of total mapped reads), so 480
that the level of expression of genes within the same library and in different libraries are comparable 481
(Mortazavi et al. 2008). This data is part of the WBD Transcriptome Atlas (unpublished data). 482
Heterologous MpCPs production - MpCP coding genes were amplified from a cDNA library 483
prepared from M. perniciosa as described elsewhere (Zaparoli et al. 2009). The constructs were cloned into 484
a modified version of pETSUMO (Invitrogen) and transformed into E. coli Origami 2 (Merck) chemically 485
competent cells. The final constructs spanned residues 20–145 (MpCP1), 17–140 (MpCP2), 15–135 486
(MpCP3), and 22–156 (MpCP5). For MpCPs purification, the transformed bacteria were lysed and the 487
clarified soluble fraction purified by a two-step procedure, starting with Immobilized Metal Ion Affinity 488
Chromatography (IMAC) using the Co2+-charged TALON resin (BD Biosciences), equilibrated with 50 489
mM Tris-HCl pH 8.5, 150 mM NaCl. The resin was washed extensively with this solution and then 490
incubated overnight at room temperature with appropriated amount of the protease ULP-1 for His-SUMO 491
tag removal. The cleaved protein was eluted from the resin and loaded onto the Superdex 75 HR 10/30 gel 492
filtration column (GE Healthcare) equilibrated in 50 mM Tris-HCl, pH 8.5 and 150 mM NaCl. Several 493
attempts to produce soluble MpCP4 did not succeed well. The eluted protein was concentrated to the 494
Page 21 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
22
required concentration as judged by UV280nm absorption and calculated coefficient extinction value as 495
estimated through the primary sequence and the ExPASy ProtParam tool (Gasteiger et al. 2005), and used 496
for crystallization screens as well as functional assays. 497
X-ray crystallography - Crystals were grown using the conventional sitting drop, vapor diffusion 498
method in the following conditions: MpCP1 - 0.1 M sodium acetate pH 4.5, 20% PEG 1000, 0.2 M zinc 499
acetate; MpCP2 - 0.1 M sodium acetate pH 4.6, 8% PEG 4000; MpCP3 - 0.1 M bis-tris propane pH 7.0, 500
1.5 M ammonium tartrate; MpCP5 - 0.1 M sodium acetate pH 5.5, 5% PEG 400, 18% PEG 3350. X-ray 501
diffraction datasets were collected at beamlines D03B-MX1 and W01B-MX2, at the Brazilian National 502
Synchrotron Laboratory, LNLS. All data were integrated using Mosflm (Leslie 1992) and scaled with 503
SCALA (Evans 2006). MpCP1 was solved by Sulfur-SAD using SHELX (Sheldrick 2010) and the first set 504
of phases of MpCP2 and MpCP3 was obtained by the molecular replacement with MrBUMP (Keegan and 505
Winn 2008), using the MpCP1 monomer as the search model. The initial set of phases of MpCP5 was 506
obtained using Phaser (McCoy et al. 2007) and the protein Sm1 from T. virens as the search model (PDB 507
ID: 3M3G). Positional and B-factor refinement cycles, as well as solvent modeling, were performed with 508
Refmac (Vagin et al. 2004) followed by visual inspection using COOT (Emsley et al 2010). 509
NMR spectroscopy - 15N- and 13C/15N-labelled MpCP proteins were produced and purified as 510
described in supplemental material, dialyzed against a solution containing 20 mM Na2HPO4-NaH2PO4 pH 511
7.2, 50 mM NaCl and then added of 10% D2O for NMR experiments. NMR spectra were recorded on a 512
Varian Inova 600-MHz spectrometer equipped with a cryogenic probe. The spectra were processed with 513
NMRPipe/NMRDraw (Delaglio et al. 1995) and analyzed with NMRView (Johnson and Blevins 1994). 514
The backbone assignment of MpCP3 and 5 were determined using the three-dimensional experiments 515
HNCACB, CBCA(CO)NH, HNCO, HN(CA)CO (Ikura et al. 1990, Grzesiek and Bax 1993, Wittekind and 516
Mueller 1993, Yamazaki et al 1994) and the graphical interface Smartnotebook (Slupsky et al. 2003). 517
Page 22 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
23
Titrations of 15N-labeled MpCPs proteins with NAG and NAG4 were followed by the recording of the two-518
dimensional 15N-HSQC spectra. For more details on the techniques describes and other techniques, see 519
supplemental material. 520
The following procedures are described in the Supplemental Material: Phylogenetic reconstruction; 521
Crystallization, X-ray Crystallography Data Collection and Processing; Structure Solution and Refinement; 522
NMR sample preparation; N-acetylglucosamine titration NMR experiment; N-acetylglucosamine docking 523
to MpCP3 and MpCP5, mutagenesis and fluorescence-based thermal shift assays; qPCR; Cacao cell wall 524
extraction; Pull-down assays; Thioflavin T, 1,8-ANS and bis-ANS fluorescence measurements; Cellulose 525
fragmentation; Spore germination; Circular dichroism spectroscopy; Identification of native MpCPs by 526
mass spectrometry; Transmission Electron Microscopy; Studies of protein cleavage associated to self-527
assembling by mass spectrometry; Injection of MpCPs in tobacco leaves. 528
ACKNOWLEDGEMENTS 529
This work was supported by FAPESP grant 2010/51884-8 (to ALBA), FAPESP fellowships 530
2010/14504-2 (to MROB) and 2010/51891-4 (to JFO) and CNPq fellowship 400796/2012-0 (to AC). We 531
thank LNBio for financial support and access to all facilities (MAS, LPP, LEC, Robolab and NMR). We 532
also thank LNNano for accessibility to LME, as well as the staff of the X-ray crystallography beamlines 533
(D03B-MX1 and W01B-MX2, at LNLS) used in this work. We thank Dr. Alessandra Girasole for technical 534
support and Gabriel Lorencini Fiorin for promptly providing fungal material used in this study. 535
536
AUTHOR CONTRIBUTIONS 537
MROB performed heterologous protein production and purification, polysaccharide interaction 538
assays, basidiospore germination assays, biophysical and biochemical characterizations, in planta defense 539
response experiments and edited the manuscript. JFO performed heterologous protein production and 540
Page 23 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
24
purification, X-ray crystallography and structure refinement, biophysical and biochemical 541
characterizations, NMR experiments, identified NMR spectra peaks and edited the manuscript. DAM 542
performed phylogenetic relationship construction and analysis, and executed qPCR experiments. PJPLT 543
and GAGP designed and performed RNAseq experiments. PFVP and HOT performed heterologous protein 544
production and purification. MLS and ACMZ designed, performed and interpreted NMR experiments. AC 545
and RVP performed transmission electron microscopy of MpCP2. PSLO performed docking experiments. 546
SMGD designed biophysical and biochemical characterizations, interpreted the data and edited the 547
manuscript. ALBA designed biophysical and biochemical characterizations, designed and performed X-ray 548
crystallography and structure refinement, interpreted the data and edited the manuscript. 549
550
LITERATURE CITED 551
Aime, M. & Phillips-Mora, W. (2005). The causal agents of Witches’ broom and frosty pod rot 552
of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycologia 97, 1012–553
1022. 554
Baccelli, I., Comparini, C., Bettini, P., Martellini, F., Ruocco, M., Pazzagli, L., Bernardi, R. & 555
Scala, A. (2012). The expression of the cerato-platanin gene is related to hyphal growth and 556
chlamydospores formation in Ceratocystis platani. FEMS Microbiol. Lett. 327, 155–163. 557
Baccelli, I., Luti, S., Bernardi, R., Scala, A., & Pazzagli, L. (2013). Cerato-platanin shows 558
expansin-like activity on cellulosic materials. Appl. Microbiol. and Biot. [Epub ahead of print]. 559
Bergthorsson, U., Andersson, D. & Roth, J. (2007). Ohno’s dilemma: evolution of new genes 560
under continuous selection. P. Natl. Acad. Sci. USA 104, 17004–17009. 561
Page 24 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
25
Bhadauria, V., Banniza, S., Vandenberg, A., Selvaraj, G., & Wei, Y. (2011). EST mining 562
identifies proteins putatively secreted by the anthracnose pathogen Colletotrichum truncatum. BMC 563
Genomics 12, 327. 564
Bhadauria, V., Banniza, S., Vandenberg, A., Selvaraj, G., & Wei, Y. (2011). EST mining 565
identifies proteins putatively secreted by the anthracnose pathogen Colletotrichum truncatum. BMC 566
Genomics 12, 327. 567
Boddi, S., Comparini, C., Calamassi, R., Pazzagli, L., Cappugi, G. & Scala, A. (2004). Cerato-568
platanin protein is located in the cell walls of ascospores, conidia and hyphae of Ceratocystis fimbriata 569
f. sp. platani. FEMS Microbiol. Lett. 233, 341–346. 570
Boller, T. (1995). Chemoperception of Microbial Signals in Plant Cells. Annu. Rev. Plant Phys. 571
46, 189–214. 572
Carresi, L., Pantera, B., Zoppi, C., Cappugi, G., Oliveira, A., Pertinhez, T., Spisni, A., Scala, A. 573
& Pazzagli, L. (2006). Cerato-platanin, a phytotoxic protein from Ceratocystis fimbriata: expression in 574
Pichia pastoris, purification and characterization. Protein Expres. Purif. 49, 159–167. 575
Cavanagh, J., Fairbrother, W., Palmer III, A., Rance, M. & Skelton, N. (2007). Protein NMR 576
Spectroscopy: Principles and Practice, Academic Press, San Diego, CA. 577
Comparini, C., Carresi, L., Pagni, E., Sbrana, F., Sebastiani, F., Luchi, N., Santini, A., Capretti, 578
P., Tiribilli, B., Pazzagli, L., Cappugi, G. & Scala, A. (2009). New proteins orthologous to cerato-579
platanin in various Ceratocystis species and the purification and characterization of cerato-populin from 580
Ceratocystis populicola. Appl. Microbiol. Biot. 84, 309–22. 581
Page 25 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
26
de Jonge, R., van Esse, H., Kombrink, A., Shinya, T., Desaki, Y., Bours, R., van der Krol, S., 582
Shibuya, N., Joosten, M. & Thomma, B. (2010). Conserved fungal LysM effector Ecp6 prevents chitin-583
triggered immunity in plants. Science 329, 953–955. 584
Delaglio, F., Grzesiek, S., Vuister, G., Zhu, G., Pfeifer, J. & Bax, A. (1995). NMRPipe: a 585
multidimensional spectral processing system based on UNIX pipes. J Biomol NMR 6, 277–293. 586
de Oliveira, A., Gallo, M., Pazzagli, L., Benedetti, C., Cappugi, G., Scala, A., Pantera, B., 587
Spisni, A., Pertinhez, T. & Cicero, D. (2011). The structure of the elicitor Cerato-platanin (CP), the first 588
member of the CP fungal protein family, reveals a double ψβ-barrel fold and carbohydrate binding. J. 589
Biol. Chem. 286, 17560–17568. 590
Desender, S., Andrivon, D. & Val, F. (2007). Activation of defence reactions in Solanaceae: 591
where is the specificity? Cell Microbiol. 9, 21–30. 592
Djonović, S., Vargas, W.A., Kolomiets, M. V, Horndeski, M., Wiest, A., & Kenerley, C.M. 593
(2007). A proteinaceous elicitor Sm1 from the beneficial fungus Trichoderma virens is required for 594
induced systemic resistance in maize. Plant Physiol. 145, 875–889. 595
Dong, X. (2004). NPR1, all things considered. Curr. Opin. Plant Biol. 7, 547–552. 596
Emsley, P., Lohkamp, B., Scott, W. & Cowtan, K. (2010). Features and development of Coot. 597
Acta Crystallogr. D 66, 486–501. 598
Evans, H. (2007). Cacao diseases-the trilogy revisited. Phytopathology 97, 1640–1643. 599
Evans, P. (2006). Scaling and assessment of data quality. Acta Crystallogr. D 62, 72–82. 600
Fontana, F., Santini, A., Salvini, M., Pazzagli, L., Cappugi, G., Scala, A., Durante, M. & 601
Bernardi, R. (2008). Cerato-Platanin Treated Plane Leaves Restrict Ceratocystis platani Growth and 602
Overexpress Defence-relates Genes. J. Plant Pathol. 90, 293–304. 603
Page 26 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
27
Frias, G.A., Purdy, L.H., & Schmidt, R.A. (1995). An Inoculation Method for Evaluating 604
Resistance of Cacao to Crinipellis perniciosa. Plant Dis. 79, 787–791. 605
Frías, M., González, C. & Brito, N. (2011). BcSpl1, a cerato-platanin family protein, contributes 606
to Botrytis cinerea virulence and elicits the hypersensitive response in the host. New Phytol. 192, 483–607
495. 608
Frías, M., Brito, N., & González, C. (2013). The Botrytis cinerea cerato-platanin BcSpl1 is a 609
potent inducer of systemic acquired resistance (SAR) in tobacco and generates a wave of salicylic acid 610
expanding from the site of application. Mol. Plant Pathol. 14, 191–196. 611
Frischmann, A., Neudl, S., Gaderer, R., Bonazza, K., Zach, S., Gruber, S., Spadiut, O., 612
Friedbacher, G., Grothe, H., & Seidl-Seiboth, V. (2013). Self-assembly at Air/Water Interfaces and 613
Carbohydrate Binding Properties of the Small Secreted Protein EPL1 from the fungus Trichoderma 614
atroviride. J. Biol. Chem. 288, 4278–4287. 615
Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S., Wilkins, M.R., Appel, R.D. & Bairoch, 616
A. (2005). Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics 617
Protocols Handbook, J.M. Walker, ed., Humana Press, pp. 571–607. 618
Gayoso, C., Pomar, F., Novo-Uzal, E., Merino, F., & De Ilárduya, O.M. (2010). The Ve-619
mediated resistance response of the tomato to Verticillium dahliae involves H2O2, peroxidase and 620
lignins and drives PAL gene expression. BMC Plant Biol. 10, 232. 621
Grzesiek, S. & Bax, A. (1993). Amino acid type determination in the sequential assignment 622
procedure of uniformly 13C/15N-enriched proteins. J. Biomol. NMR 3, 185–204. 623
Page 27 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
28
Guevara-Morato, M.A., De Lacoba, M.G., García-Luque, I., & Serra, M.T. (2010). 624
Characterization of a pathogenesis-related protein 4 (PR-4) induced in Capsicum chinense L3 plants 625
with dual RNase and DNase activities. J. Exp. Bot. 61, 3259–3271. 626
Hall, N., Keon, J. & Hargreaves, J.A. (1999). A homologue of a gene implicated in the 627
virulence of human fungal diseases is present in a plant fungal pathogen and is expressed during 628
infection. Physiol. Mol. Plant. P. 55, 69–73. 629
Holm, L. & Rosenström, P. (2010). Dali server: conservation mapping in 3D. Nucleic Acids 630
Res. 38, W545–W549. 631
Hunter, S., Jones, P., Mitchell, A., Apweiler, R., Attwood, T.K., Bateman, A., Bernard, T., 632
Binns, D., Bork, P., Burge, S., et al. (2012). InterPro in 2011: new developments in the family and 633
domain prediction database. Nucleic Acids Res. 40, D306–D312. 634
Ikura, M., Kay, L. & Bax, A. (1990). A novel approach for sequential assignment of 1H, 13C, 635
and 15N spectra of larger proteins: heteronuclear triple-resonance three-dimensional NMR 636
spectroscopy. Application to calmodulin. Biochemistry-US 29, 4659–4667. 637
Jeong, J.S., Mitchell, T.K., & Dean, R.A. (2007). The Magnaporthe grisea snodprot1 homolog, 638
MSP1, is required for virulence. FEMS Microbiol. Lett. 273, 157–165. 639
Johnson, B. & Blevins, R. (1994) NMR View: A computer program for the visualization and 640
analysis of NMR data. J. Biomol. NMR 4, 603–614. 641
Jones, J. & Dangl, J. (2006). The plant immune system. Nature 444, 323–329. 642
Keegan, R. & Winn, M. (2008). MrBUMP: an automated pipeline for molecular replacement. 643
Acta Crystallogr. D 64, 119–124. 644
Page 28 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
29
Kerff, F., Amoroso, A., Herman, R., Sauvage, E., Petrella, S., Filée, P., Charlier, P., Joris, B., 645
Tabuchi, A., Nikolaidis, N., et al. (2008). Crystal structure and activity of Bacillus subtilis YoaJ 646
(EXLX1), a bacterial expansin that promotes root colonization. P. Natl. Acad. Sci. USA 105, 16876–647
16881. 648
Langmead, B., Trapnell, C., Pop, M., & Salzberg, S.L. (2009). Ultrafast and memory efficient 649
alignment of short DNA sequences to the human genome. Genome Biol. 10, R25. 650
Leal, G.A, Gomes, L.H., Albuquerque, P.S.B., Tavares, F.C. a, & Figueira, A. (2010). 651
Searching for Moniliophthora perniciosa pathogenicity genes. Fungal Biol. 114, 842–854. 652
Lehotzky, R., Partch, C., Mukherjee, S., Cash, H., Goldman, W., Gardner, K. & Hooper, L. 653
(2010). Molecular basis for peptidoglycan recognition by a bactericidal lectin. P. Natl. Acad. Sci. USA 654
107, 7722–7727. 655
Leslie, A. (1992). Recent changes to the MOSFLM package for processing film and image plate 656
data. Joint CCP4 + ESF-EAMCB Newsletter on Protein Crystallography 26. 657
Lomakin, A., Chung, D., Benedek, G., Kirschner, D. & Teplow, D. (1996). On the nucleation 658
and growth of amyloid beta -protein fibrils: Detection of nuclei and quantitation of rate constants. P. 659
Natl. Acad. Sci. USA 93, 1125–1129. 660
Lombardi, L., Faoro, F., Luti, S., Baccelli, I., Martellini, F., Bernardi, R., Picciarelli, P., Scala, 661
A., & Pazzagli, L. (2013). Differential timing of defense-related responses induced by cerato-platanin 662
and cerato-populin, two non-catalytic fungal elicitors. Physiol. Plantarum. [Epub ahead of print]. 663
Marelli, J., Maximova, S., Gramacho, K., Kang, S. & Guiltinan, M. (2009). Infection Biology of 664
Moniliophthora perniciosa on Theobroma cacao and Alternate Solanaceous Hosts. Tropical Plant Biol. 665
2, 149–160. 666
Page 29 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
30
Martellini, F., Faoro, F., Carresi, L., Pantera, B., Baccelli, I., Maffi, D., Tiribilli, B., Sbrana, F., 667
Luti, S., Comparini, C., Bernardi, R., Cappugi, G., Scala, A., Luigia Pazzagli, L. (2012). Cerato-668
Populin and Cerato-Platanin, Two Non-Catalytic Proteins from Phytopathogenic Fungi, Interact with 669
Hydrophobic Inanimate Surfaces and Leaves. Mol. Biotechnol. Doi: 10.1007/s12033-012-9618-4. 670
Published online 671
McCoy, A., Grosse-Kunstleve, R., Adams, P., Winn, M., Storoni, L. & Read, R. (2007). Phaser 672
crystallographic software. J. Appl.Crystallogr. 40, 658–674. 673
Meinhardt, L., Rincones, J., Bailey, B., Aime, M., Griffith, G., Zhang, D. & Pereira, G. (2008). 674
Moniliophthora perniciosa, the causal agent of Witches’ broom disease of cacao: what's new from this 675
old foe? Mol. Plant. Pathol. 9, 577–588. 676
Mishra, R., Sörgjerd, K., Nyström, S., Nordigården, A., Yu, Y. & Hammarström, P. (2007). 677
Lysozyme amyloidogenesis is accelerated by specific nicking and fragmentation but decelerated by 678
intact protein binding and conversion. J. Mol. Biol. 366, 1029–1044. 679
Mondego, J.M.C., Carazzolle, M.F., Costa, G.G.L., Formighieri, E.F., Parizzi, L.P., Rincones, 680
J., Cotomacci, C., Carraro, D.M., Cunha, A.F., Carrer, H., et al. (2008). A genome survey of 681
Moniliophthora perniciosa gives new insights into Witches’ Broom Disease of cacao. BMC Genomics 682
9, 548. 683
Moores, B., Drolle, E., Attwood, S., Simons, J. & Leonenko, Z. (2011). Effect of surfaces on 684
amyloid fibril formation. PloS One 6, e25954. 685
Mortazavi, A., Williams, B.A., McCue, K., Schaeffer, L., & Wold, B. (2008). Mapping and 686
quantifying mammalian transcriptomes by RNA-Seq. Nat.Methods 5: 621-628. 687
Page 30 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
31
Murphy, R. (2007). Kinetics of amyloid formation and membrane interaction with 688
amyloidogenic proteins. Biochim. Biophys. Acta 1768, 1923–193. 689
Ortega, A., Amorós D. & García de la Torre, J. (2011). Prediction of hydrodynamic and other 690
solution properties of rigid proteins from atomic and residue-level models. Biophys. J. 101, 892-898. 691
Pan, S. & Cole, G. (1995). Molecular and biochemical characterization of a Coccidioides 692
immitis-specific antigen. Infect. Immun. 63, 3994–4002. 693
Pazzagli, L., Cappugi, G., Manao, G., Camici, G., Santini, A. & Scala, A. (1999). Purification, 694
characterization, and amino acid sequence of cerato-platanin, a new phytotoxic protein from 695
Ceratocystis fimbriata f. sp. platani. J. Biol. Chem. 274, 24959–24964. 696
Pazzagli, L., Zoppi, C., Carresi, L., Tiribilli, B., Sbrana, F., Schiff, S., Pertinhez, T.A., Scala, A. 697
& Cappugi, G. (2009). Characterization of ordered aggregates of cerato-platanin and their involvement 698
in fungus-host interactions. Biochem Biophys Acta 1790, 1334–1344. 699
Petersen, T.N., Brunak, S., Von Heijne, G., & Nielsen, H. (2011). SignalP 4.0: discriminating 700
signal peptides from transmembrane regions. Nat. Methods 8, 785–786. 701
Pires, A.B.L., Gramacho, K.P., Silva, D.C., Góes-Neto, A., Silva, M.M., Muniz-Sobrinho, J.S., 702
Porto, R.F., Villela-Dias, C., Brendel, M., Cascardo, J.C.M., et al. (2009). Early development of 703
Moniliophthora perniciosa basidiomata and developmentally regulated genes. BMC Microbiol. 9, 158. 704
Rincones, J., Scarpari, L.M., Carazzolle, M.F., Mondego, J.M.C., Formighieri, E.F., Barau, J.G., 705
Costa, G.G.L., Carraro, D.M., Brentani, H.P., Vilas-boas, L.A., et al. (2008). Differential Gene 706
Expression Between the Biotrophic-Like and Saprotrophic Mycelia of the Witches’ Broom Pathogen 707
Moniliophthora perniciosa. Mol. Plant Microbe In. 21, 891–908. 708
Page 31 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
32
Scarpari, L., Meinhardt, L., Mazzafera, P., Pomella, A., Schiavinato, M., Cascardo, J. & Pereira, 709
G. (2005). Biochemical changes during the development of Witches’ broom: the most important 710
disease of cocoa in Brazil caused by Crinipellis perniciosa. J. Exp. Bot. 56, 865–877. 711
Seidl, V., Marchetti, M., Schandl, R., Allmaier, G. & Kubicek, C. (2006). Epl1, the major 712
secreted protein of Hypocrea atroviridis on glucose, is a member of a strongly conserved protein family 713
comprising plant defense response elicitors. FEBS J. 273, 4346–4359. 714
Sheldrick, G. (2010). Experimental phasing with SHELXC/D/E: combining chain tracing with 715
density modification. Acta Crystallogr. D 66, 479–485. 716
Slupsky, C., Boyko, R., Booth, V. & Sykes, B. (2003). Smartnotebook: a semi-automated 717
approach to protein sequential NMR resonance assignments. J. Biomol. NMR 27, 313–321. 718
Stanke, M., Steinkamp, R., Waack, S., & Morgenstern, B. (2004). AUGUSTUS: a web server 719
for gene finding in eukaryotes. Nucleic Acids Res. 32, W309–W312. 720
Tsiroulnikov, K., Chobert, J. & Haertlé, T. (2006). Copper-dependent degradation of 721
recombinant ovine prion protein. Phosphatidylinositol stimulates aggregation and copper-driven 722
disappearance of prion protein. FEBS J. 273, 1959–1965. 723
Uversky, V. & Fink, A. (2004). Conformational constraints for amyloid fibrillation: the 724
importance of being unfolded. Biochem. Biophys. Acta 1698, 131–153. 725
Vagin, A., Steiner, R., Lebedev, A., Potterton, L., McNicholas, S., Long, F. & Murshudov, G. 726
(2004). REFMAC5 dictionary: organization of prior chemical knowledge and guidelines for its use. 727
Acta Crystallogr. D 60, 2184–2195. 728
van Loon, L.C., Rep, M., & Pieterse, C.M.J. (2006). Significance of inducible defense-related 729
proteins in infected plants. Annu. Rev. Phytopathol. 44, 135–162. 730
Page 32 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
33
Vargas, W., Djonović, S., Sukno, S. & Kenerley, C. (2008). Dimerization controls the activity 731
of fungal elicitors that trigger systemic resistance in plants. J. Biol. Chem. 283, 19804–19815. 732
Whiteford, J. & Spanu, P. (2002). Hydrophobins and the interactions between fungi and plants. 733
Mo.l Plant Pathol. 3, 391–400. 734
Wilson, L., Idnurm, A. & Howlett, B. (2002). Characterization of a gene (sp1) encoding a 735
secreted protein from Leptosphaeria maculans, the blackleg pathogen of Brassica napus. Mol. Plant 736
Pathol. 3, 487–493. 737
Wittekind, M. & Mueller, L. (1993). HNCACB, a High-Sensitivity 3D NMR Experiment to 738
Correlate Amide-Proton and Nitrogen Resonances with the Alpha- and Beta-Carbon Resonances in 739
Proteins. J. Magn. Reson. Ser. B 101, 201–205. 740
Wösten, H. & de Vocht, M. (2000). Hydrophobins, the fungal coat unravelled. Biochim. 741
Biophys. Acta 1469, 79–86. 742
Yamazaki, T., Lee, W., Arrowsmith, C., Muhandiram, D.R. & Kay, L.E. (1994). A Suite of 743
Triple Resonance NMR Experiments for the Backbone Assignment of 15N, 13C, 2H Labeled Proteins 744
with High Sensitivity. J. Am. Chem. Soc. 116, 11655–11666. 745
Yennawar, N., Li, L., Dudzinski, D., Tabuchi, A. & Cosgrove, D. (2006). Crystal structure and 746
activities of EXPB1 (Zea m 1), a beta-expansin and group-1 pollen allergen from maize. P. Natl. Acad. 747
Sci. USA 103, 14664–14671. 748
Zaparoli, G., Cabrera, O., Medrano, F., Tiburcio, R., Lacerda, G. & Pereira, G. (2009). 749
Identification of a second family of genes in Moniliophthora perniciosa, the causal agent of Witches’ 750
broom disease in cacao, encoding necrosis-inducing proteins similar to cerato-platanins. Mycol. Res. 751
113, 61–72. 752
753
Page 33 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
34
FIGURES CAPTIONS 754
FIGURE 1. Differential expression of MpCP1 to 12 throughout the stages of fungus development and 755
infection. (A) Out of 12 CP-coding genes identified in M. perniciosa, only transcripts from isoforms 4, 5, 756
11 and 12 are detected in the biotrophic fungus. A decrease in their expression levels is observed as the 757
disease progresses, until complete depletion when the dry-broom stage is reached. (B) Inversely 758
proportional levels MpCP2 and 3 coding transcripts are measured when rotten, field-collected seeds and 759
fruit pods are analyzed. (C) In samples extracted from lab-cultivated fungus, MpCP2 and 3 transcripts are 760
detected in the mycelium, whereas isoforms 4 and 11 are exclusively detected in germinated basidiospores. 761
MpCP1 is only detected in basidiocarps. Asterisks indicate isoforms that were detected at protein level, by 762
mass spectrometry, in the culture medium of in vitro grown necrotrophic mycelium. RPKM, Reads Per 763
Kilobase of exon model per Million mapped reads. 764
FIGURE 2. Phylogenetic reconstruction of the Cerato-platanin family. Comprehensive evolutionary tree 765
determined using deposited CP sequences. Four main clusters can be distinguished. Epl1/Epl2 (red box), in 766
accordance to previously published analysis (Seidl et al 2006); Mper1 and Mper2, delimited by blue, and 767
green boxes, respectively; and the sequences from other basidiomycetes, together with MpCP9 (not 768
highlighted). Numbers near branches are Bayesian posterior probabilities. 769
FIGURE 3. Crystal structures of MpCP1, 2, 3 and 5. Stereographic views in cartoon representations of 770
superposed MpCP1-3 (A) and MpCP1 and MpCP5 (B). All four structures present the canonical double 771
Ψβ-barrel, interspersed with helices. The major structural differences between MpCP5 and MpCP1, 2 and 3 772
is the length of the loop between sheets β2 and β3 and the hairpin connecting sheets β3 and β4. 773
FIGURE 4. MpCP3 and 5 bind to NAG4, though at distinct regions. (A) Overlay of regions of 15N-HSQC 774
spectra of 15N-labeled MpCP3 (left panels) and 5 (right panels), both proteins at 0.5 mM, without NAG4 775
(black) and titrated with 0.1 mM (red), 2.5 mM (green), 5 mM (purple) and 10 mM (pink) of NAG4, 776
Page 34 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
35
highlighting some of the shifted and diminished residue peaks. All spectra were acquired with the same 777
number of scans. (B) Curve fitting of the NAG4 concentration plotted against the ∆δ of specified residues 778
used for the Kd calculation. (C) Computational blind docking models for the interactions between MpCP3 779
and MpCP5 with NAG4. NAG4 is represented by sticks and NAG4-interacting residues are represented as 780
spheres. (D) Relative mRNA expression levels of defense-related tobacco genes after treatment with NAG6 781
(400 nM) or MpCP5 (40 uM) pre-incubated NAG6. 782
FIGURE 5. Sugar hydrolysis and cacao cell wall/chitin binding assays. (A) Colorimetric assay, with the 783
substrate 3,5-dinitrosalicylic acid (DNS), evaluating the glycolytic activity of the recombinant MpCP1, 2, 3 784
and 5 against different substrates. No significant activity was detected for any MpCP analyzed, when 785
compared to positive control TPE (termite total protein extract). (B) Similarly, no relevant interaction 786
between MpCPs and cocoa extracted cell-wall (CCW) was detected in a pull-down assay, when compared 787
to positive control CBM3 (family 3 Carbohydrate Binding Module), from B. subtillis. (C) All MpCPs 788
(soluble form) tested were pulled-down by chitin insoluble polymers, with MpCP3 showing the highest 789
levels of interaction. Chitin is one of the main components of fungal cell wall. S, supernatant fraction; P, 790
pellet fraction. 791
FIGURE 6. Evaluation of self-aggregation capability. (A) Detection of the fluorescence signal induced 792
upon the binding of Thioflavin T (ThT) against increasing concentrations of MpCPs incubated at 37oC, low 793
pH, for 30 days. The data allowed the determination of an amyloid-like self-aggregation behavior for 794
MpCPs 2, 3 and 5, at threshold concentrations between 0.4 and 0.8 mM protein. MpCP1 displays the same 795
behavior, although only at concentrations beyond 1.5 mM, similarly to CpCP (Pazzagli et al. 2009). (B) 796
The signal measured upon the binding of the dyes 1,8-ANS and bis-ANS to MpCP1, 2, 3 and 5 indicating 797
the hydrophobic nature of the aggregates. (C) Time course analysis of the self-assembly of 0.8 mM MpCP2 798
and 3, at 37 oC and low pH, as monitored by ThT fluorescence, shows a much quicker dynamics of 799
Page 35 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

Mario R. de O. Barsottini – MPMI
36
aggregation for MpCP2, which reaches equilibrium after only 50 hours, compared to 25 days required for 800
MpCP3. All the experiments were performed in triplicate. 801
FIGURE 7. Biophysical characterization of the aggregates. (A) Far-UV circular dichroism allowed the 802
tracking of the changes in secondary structure of the MpCPs required for self-assembling (at 0.8 mM 803
concentration). Upon incubation at low pH and high temperature, MpCP1 assumes a completely unfolded 804
structure (with a negative band at 200 nm), while MpCPs 2, 3 and 5 acquire a closer to zero band at 200 nm 805
and more negative ellipticity at 222 nm. No difference was observed in the CD spectra for the MpCPs, 806
between native initial state (0 days) and after 20 days of incubation at 4 °C (data not shown). (B) 807
MRE[θ]200 x MRE[θ]222 combination extracted from the 4°C pH 7.2, and 37°C at low pH spectra for all 808
proteins. The definition of native, pre-molten globule and molten globule regarding to the combined mean 809
residue ellipticity at 200 and 222 nm was taken as revised (Uversky and Fink 2004). (C) Dynamic light 810
scattering data showing the aggregation process as defined by a shift in the Stokes radius after 37°C, low 811
pH incubation. Theoretical values for Rs, calculated using Hydropro (Ortega et al 2011), are 1.83, 1.76, 812
1.73, and 1.95 nm, for monomeric MpCP1, 2, 3 and 5, respectively. (D) Micrographs 1 and 2, obtained by 813
negative stain electron microscopy, depict the filamentous nature of MpCP2 aggregates (0.8 mM, 20 days 814
of incubation at low pH). (E) Cellulose loosening activity was detected for aggregated MpCP2, but not its 815
soluble monomeric counterpart. (F) Aggregated MpCP2 also promotes M perniciosa`s basidiospores 816
germination tube growth. Aggregated n = 12; Soluble n = 37; Buffer n = 34. 817
Page 36 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

139x40mm (300 x 300 DPI)
Page 37 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

133x175mm (300 x 300 DPI)
Page 38 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

71x99mm (300 x 300 DPI)
Page 39 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

125x126mm (300 x 300 DPI)
Page 40 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

125x69mm (300 x 300 DPI)
Page 41 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

91x92mm (300 x 300 DPI)
Page 42 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

170x166mm (300 x 300 DPI)
Page 43 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

1
SUPPLEMENTAL MATERIAL 1
2
Functional diversification of cerato-platanins in 3
Moniliophthora perniciosa as seen by differential 4
expression and protein function specialization 5
6
Mario R. de O. Barsottini a,b**
, Juliana F. de Oliveirab**, Douglas Adamoski
b**, Paulo J. P. L. 7
Teixeiraa, Paula F. V. do Prado
a,b, Henrique O. Tiezzi
b, Mauricio L. Sforça
b, Alexandre 8
Cassagoc, Rodrigo V. Portugal
c, Paulo S. L. de Oliveira
b, Ana C. de M. Zeri
b, Sandra M. G. 9
Diasb**, Gonçalo A. G. Pereira
a,b and Andre L. B. Ambrosio
b** 10
11
aFrom the Departamento de Genética e Evolução, UNICAMP, Campinas, SP 13083-970 12 bLaboratório Nacional de Biociências, CNPEM, Campinas, SP 13083-100 13
cLaboratório Nacional de Nanotecnologia, CNPEM, Campinas, SP 13083-100 14
15
Page 44 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

2
SM Text 16
Experimental Procedures 17
Phylogenetic reconstruction. The phylogenetic tree was estimated with protein sequences 18
aligned with MUSCLE (Edgar 2004). The model of amino acid substitution (WAG+G) was 19
chosen with ProtTest v3.0 (Darriba et al. 2011) and defined in MrBayes v3.2 (Ronquist and 20
Huelsenbeck 2003). The 148.000.000 generations were evaluated in TRACER v1.5 and 21
AWTY (Nylander et al. 2008) to burn-in 103.600.000 (70%) generations (after visual 22
inspections of LnL and cumulative splits plot of putative stationary phase, as well as 23
Effective Sample Size of the last 30% of the chain). The tree was assembled with SumTrees 24
from DendroPy package (Sukumaran and Holder 2010). The protein sequence IDs from the 25
previously described MpCP1 to MpCP5 (Zaparoli et al 2009) are EU250339 (MpCP1), 26
EU250340 (MpCP2), EU250342 (MpCP3), EU250344 (MpCP4) and EU250345 27
(MpCP5). The other sequences from M. perniciosa biotype C and biotype S, M. roreri and 28
C. campanella may be found in the main text. Protein sequences from other fungi were 29
obtained from NCBI and Mycocosm portal (DOE Joint Genome Institute) databases, and 30
their accession numbers are as follows. NCBI accession numbers: AAB0010140 (19 kDa 31
Antigen from Coccidioides posadasii), AAT1191162 (Aca1 from Taiwanofungus 32
camphoratus), EFE4297541 (Asp f13 from Trichophyton verrucosum), EAU3799938 (Asp 33
f15 from Aspergillus terreus), EDN3009454 (BC1G_02163 from Botryotinia fuckeliana), 34
ABM6351348 (Cerato-platanin from Ceratocystis platani), EIW85572 (Cerato-platanin 35
from Coniophora puteana), EDR1116852 (Cerato-platanin from Laccaria bicolor), 36
ACA1360043 (Ceratovariosporin from Ceratocystis variospora), ABE7369260 (Epl1 from 37
Hypocrea atroviridis), CAL8075357 (Epl1 from Trichoderma asperellum), EJD54183 38
Page 45 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

3
(Heat-stable 19 kDa antigen from Auricularia delicata), ABM6350642 (Pop1 from 39
Ceratocystis populicola), EAA5865739 (Predicted protein from Aspergillus nidulans), 40
CCD4351765 (Predicted protein from Botryotinia fuckeliana), EAU8647650 (Predicted 41
protein from Coprinopsis cinerea), EKM58222 (Predicted protein from Phanerochaete 42
carnosa), EED7983453 (Predicted protein from Postia placenta), XP_003032879 43
(Predicted protein from Schizophyllum commune), EGN96314 (Predicted protein from 44
Serpula lacrymans), AAZ8038858 (Sm1 from Hypocrea virens), AAV8379355 (Snodprot1 45
from Gibberella pulicaris), XM0037164 (Snodprot1 from Magnaporthe oryzae), 46
CAC2858561 (Snodprot1 from Neurospora crassa), AAC2687063 (Snodprot1 from 47
Phaeosphaeria nodorum), EEY2380156 (Snodprot1 from Verticillium alboatrum), 48
EAU8063649 (Snodprot1 A from Coprinopsis cinerea), EAU8196951 (Snodprot1 B from 49
Coprinopsis cinerea) and AAM3313059 (Sp1 from Leptosphaeria maculans). Mycocosm 50
protein IDs: 159980 (Predicted protein from Amanita muscaria), 34748 (Predicted protein 51
from Bjerkandera adusta), 266297 (Predicted protein from Gymnopus luxurians), 171081 52
(Predicted protein from Paxillus involutus), 22402 (Predicted protein from Pisolithus 53
microcarpus), 996983 (Predicted protein from Pisolithus tinctorius), 154741 (Predicted 54
protein A from Pisolithus tinctorius), 1069722 (Predicted protein from Pleurotus 55
ostreatus), 102212 (Predicted protein A from Pleurotus ostreatus), 45615 (Predicted 56
protein B from Pleurotus ostreatus), 1110827 (Predicted protein C from Pleurotus 57
ostreatus), 13911 (Predicted protein A from Serpula lacrymans), 61724 (Predicted protein 58
from Sphaerobolus stellatus), 506951 (Predicted protein from Suillus luteus), 73763 59
(Predicted protein A from Suillus luteus). 60
61
62
Page 46 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

4
Crystallization, X-ray Crystallography Data Collection and Processing. Crystallization 63
experiments were performed at 18oC using the conventional sitting drop vapor diffusion 64
technique. Drops were made by mixing equal parts of protein (at 10, 40, 20 and 40 mg/mL 65
for MpCP1, MpCP2, MpCP3 and MpCP5, respectively) and well solution. The 66
crystallization conditions are described in the main text. Before data collection at cryogenic 67
temperature, MpCP3 harvested crystal was cryoprotected with 10% ethylene glycol added 68
to the mother liquor, while MpCP1’s, MpCP2’ s and MpCP5’s crystal were collected 69
without additional cryo-protection. 70
71
Structure Solution and Refinement. The structure of MpCP1 was solved by Sulfur-SAD. 72
The redundant anomalous data allowed the unambiguous identification and phasing of 7 73
sulfur atoms using SHELXC/D/E (Sheldrick 2010). Using ARP/wARP (Perrakis et al 74
1999) it was possible to build 116 out of the 127 residues expected for the monomer. 75
Positional and B-factor refinement cycles were then made with Refmac (Vagin et al. 2004). 76
Manual building of the extra portions and real space refinement, including Fourier electron 77
density map inspection, were performed with Coot (Emsley et al. 2010). Solvent water 78
molecules, treated as oxygen atoms, were added using the appropriate Coot routine. The 79
first set of phases of MpCP2 and MpCP3 was obtained by the molecular replacement 80
technique as implemented in the program MrBUMP (Keegan and Winn 2008), using the 81
MpCP1 monomer (deposited in PDB ID: 3SUJ) as the search model. Real space and 82
reciprocal space refinements were performed with Coot and Refmac, respectively. The set 83
of phases of MpCP5 was obtained by the Phaser program (McCoy et al. 2007) using the 84
protein Sm1 from Trichoderma virens as the search model (deposited in PDB ID: 3M3G). 85
Following molecular replacement, density modification was performed using the program 86
Page 47 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

5
Parrot, which is part of the CCP4 suite (Winn et al 2011) and the improved map then 87
submitted to automated interpretation by the ARPwARP routine. Positional and B factor 88
refinement cycles were performed with Refmac. For calculation of Cα r.m.s.d values we 89
used the algorithm SSM (secondary structure matching) as implemented in Coot. 90
91
NMR sample preparation. To prepare 15N- and 13C/15N-labelled MpCP proteins, the E. 92
coli strain SHuffle (NEB) was cultivated in M9 minimal media supplemented with 1 g/L 93
15NH4Cl (Cambridge Isotope Laboratories) combined with 4 g/L D(+)-glucose (Sigma) to 94
obtain only 15N-labelled proteins or with [13C6]-D(+)-glucose (Cambridge Isotope 95
Laboratories) to obtain double-labeled proteins. The cultures were grown at 30°C until 600 96
nm absorbance of 1.0 followed by the protein expression induction with 0.5 mM isopropyl-97
β-D-thiogalactoside (IPTG) for further 14 hours at 30°C. The purification protocol was 98
performed essentially as described for the unlabelled protein. 99
100
N-acetylglucosamine titration NMR experiments. Titrations of 15N-labeled MpCP 101
proteins with the carbohydrate N-acetylglucosamine (NAG, Sigma-Aldrich, A8625) in its 102
tetrameric form (NAG4, Megazyme O-CHI4) were followed by the recording of the two-103
dimensional 15N-HSQC spectra. The titration series was performed by direct addition of 104
small aliquots (0.3 to 30 µL) of the carbohydrate (200 mM stock) to 600 µL of 15N-MpCPs 105
(50 uM) in order to obtain carbohydrate final concentrations of 0.1, 2.5, 5 and 10 mM. The 106
monomeric NAG was added to a final concentration of 10 mM. The temperature was 107
maintained at 25°C throughout the NMR experiment. The carbohydrates binding constant 108
was determined by spectral changes in the protein 15N-HSQC chemical shift values as a 109
function of the concentration of unlabeled ligand. The chemical shift perturbation of the 110
Page 48 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

6
15N-HSQC spectra was normalized according to the equation ∆δ(15N+1H) = 111
[(∆δ15N/10)2+(∆δ1H)2]1/2 (ppm) (Cavanagh el al. 2007). The dissociation constant values 112
(Kd) were calculated based on chemical shift changes of eight residues of each MpCP 113
(Phe32-Asp39 for MpCP3 and Val108-Lys115 for MpCP5), using the program xcrvfit. 114
115
N-acetylglucosamine docking to MpCP3 and MpCP5, mutagenesis and fluorescence-116
based thermal shift assays. NAG4 ligand structure in Mol format was obtained from 117
ChemicalBook website (CAS #2706-65-2). NAG4, MpCP3 and MpCP5 structures were 118
first submitted to energy minimization using YASARA (Krieger et al. 2009). For that, all 119
hydrogen atoms and other missing atoms from the model were created using force field 120
parameters obtained from YAMBER3. All receptor water molecules were removed. A short 121
steepest descent energy minimization was carried until the maximum atom speed dropped 122
below 2200 m/s. Then 500 steps of simulated annealing were performed with a temperature 123
of 0K. A simulation cell was centered upon the MpCPs structures and its extension defined 124
as the extreme coordinates of protein plus twice the size of the ligand radius. The size of the 125
docking grid for MpCP3 and MpCP5 complexes were 86 Å × 79 Å × 75 Å and 95 Å × 80 126
Å × 77 Å, respectively. Grid spacing used for MpCP3 and MpCP5 complexes were 0.685 127
and 0.766 Å, respectively. All docking simulations were performed by AutoDock 4.2.3 128
called from YASARA. The number of Lamarckian genetic algorithm (LGA) runs was 100 129
for each molecular complex. Population size was 150 individuals and 15 random torsions 130
for each individual. The 100 independent LGA runs were processed using the built-in 131
clustering analysis as implemented in YASARA with a 5.0 Å cutoff. 132
Mutants were obtained with the QuikChange Site-Directed Mutagenesis Kit 133
(Stratagene) following the manufacturer’s instructions and purified according to the 134
Page 49 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

7
protocol described for the wild-type proteins. To evaluate the ability of wild type and 135
mutants MpCPs 3 and 5 to bind to NAG4 we performed thermal shift assays in a 7500 Real 136
Time PCR System (Applied Biosystems). Fifty micromolar of protein and a 100-fold 137
dilution of a 5000-fold SYBRGreen stock solution (Life Technology) were incubated in the 138
absence and presence of 20 mM or 60 mM of NAG4 (for MpCP3 and MpCP5, 139
respectively), in 50 mM Tris-HCl, pH 8,5 and 150 mM NaCl solution. The fluorescence 140
signal was measured between 20°C and 100°C with a 1°C step per cycle. A sigmoidal 141
curve was fitted to the data points between 50oC and 80oC (90oC for MpCP5) using Origin 142
8.0 (OriginLab Corp., Northamptom, NA) and the inflection point taken as the melting 143
temperature (Tm) of the proteins. 144
145
qPCR. Three-week-old tobacco seedlings were thoroughly washed with distilled water, had 146
their roots removed with a scalpel and were left in distilled water for 5 h in order to recover 147
from the injury. Each seedling (approx. 100 mg of plant material) was transferred to plate 148
dish containing 300 µL of either 20 mM Na2HPO4-NaH2PO4 pH 7.2 solution, 400 nM of 6-149
acetylglucosamine (NAG6, Megazyme O-CHI6), or MpCP5 (40 µM) pre-incubated with 150
NAG6 (400 nM) for 24 hours. After 24 h, the seedlings were flash frozen in liquid nitrogen 151
and stored at -80 °C. RNA extraction was performed with RNeasy Plus Mini Kit (Qiagen) 152
following the manufacturer’s instructions. cDNA synthesis was performed with Superscript 153
III First-Strand Synthesis System for RT-PCR (Invitrogen), following the manufactures`s 154
instructions and the random hexamers primers. qPCR was performed in triplicate with 155
Power SYBR Green PCR Master Mix (Applied Biosystems) and the equipment ViiA7 156
Termocycler (Applied Biosystems). Data was evaluated following the 2-∆∆Ct (Livak and 157
Schmittgen 2001). Primers used for gene expression quantification were: 5’-158
Page 50 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

8
CCGTTGCTGCGATGATTCA-3’, 5’-GCTGCCTTCCTTGGATGTG-3’ (rRNA 18S; 159
Genbank accession number AJ236016.1), 5’-GACATCAGCGGAAGCAGTAGTAT-3’, 160
5’-CTTAGCGTCGGCGAAGTAGT-3’ (npr1; DQ837218.1), 5’-161
TGCTCTCCGAACATCTCCACAATG-3’, 5’-CCGATAGGAGTGCCTTGGAAGTTG-3’ 162
(pal1; M84466.1), 5’-GATGCCGACAAGCCTCTC-3’, 5’-163
TTGAGTTCCTGTTCCTGTGTTC-3’ (pr4a; X60281.1), 5’-164
GCTGTTACTCATGCTGCCACTT-3’ and 5’-AATGCGAGCCTGGACTGTTC-3’ (pr5; 165
X15224.1). 166
167
Cacao cell wall extraction. Extraction of cacao leaves cell wall was performed as 168
described elsewhere (Melton and Smith 2001) with minor modifications. Theobroma cacao 169
var. Comum was cultivated in a greenhouse under controlled temperature and humidity 170
conditions (from 22 to 28 °C and above 50%, respectively). Fifteen grams of healthy fully-171
expanded leaves from three months old plants were extirpated from their central nerves and 172
grinded with a mortar and pestle in the presence of liquid nitrogen. The obtained powder 173
was transferred to 300 mL of 50 mM Hepes pH 7.0 and 10 mM Dithiothreitol (DTT) and 174
quickly homogenized with a blender. The suspension was filtered through a filter paper and 175
extensively washed with 50 mM Hepes pH 7.0 followed by distilled water. The retentante 176
was lyophilized and stored at -80°C. 177
178
Glycolytic activity. MpCPs glycolytic activity was measured using a colorimetric assay 179
with the substrate 3,5-dinitrosalicylic acid (DNS) (Miller 1959), with fifty micrograms of 180
each MpCP and the positive control termite total protein extract (kindly donated by Dr. 181
Fábio Márcio Squina, CTBE, CNPEM, Campinas, SP, Brazil), and five hundred 182
Page 51 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

9
micrograms of various cell-wall polyssacharides. were incubated with 0.5% (w:v) of either 183
carboxymethyl cellulose, arabinan, arabinoxylan, laminarin, xylan, pectin, chitosan, 184
arabinogalactan, manan, starch, or CCW in 50 mM citrate buffer pH 5.0 at 37°C for 1h, 185
after which it was added to one volume of DNS solution (1% DNS, 0.2% phenol, 0.05% 186
Na2SO3 and 1% NaOH). The samples were heated at 95 °C during five minutes and the 540 187
nm absorbance measured in a 96-well plate (Eppendorf) with the Infinite 200 PRO Plate 188
Reader (Tecan). All reactions were made in triplicate. 189
190
Pull-down assays. The cocoa cell-wall extract (CCW) or chitin from shrimp shell (Sigma-191
Aldrich, C7170) interaction with MpCP1, MpCP2, MpCP3, MpCP5 (either soluble or 192
aggregated protein) and the positive control purified recombinant CBM3 (family 3 193
carbohydrate-binding module) from Bacillus subtillis (kindly donated by Dr. Mario 194
Murakami, LNBio, CNPEM, Campinas, SP, Brazil) was assessed by pull-down assay. Fifty 195
micrograms of each protein were incubated with either 20 mg of CCW or 5 mg of chitin in 196
a final volume of 1 mL binding solution (50 mM Tris-HCl pH 8,5, 150 mM NaCl). After 197
overnight incubation at room temperature, the insoluble fraction was separated by 198
centrifugation, the pellet washed three times with binding solution and then subjected to 199
SDS-PAGE 15 % analysis. 200
201
Thioflavin T, 1,8-ANS and bis-ANS fluorescence measurements. To study MpCP1, 202
MpCP2, MpCP3 and MpCP5 capacity of forming ordered amyloid-like aggregates we 203
incubated 0.1, 0.2, 0.4, 0.8, 1.5 and 3.0 mM of protein in 20 mM Na2HPO4-NaH2PO4 pH 204
7.2, 10% acetic acid solution (final pH of 3) for up to 30 days at 37°C as described 205
previously (Pazzagli et al. 2009). As controls, the proteins were incubated in 20 mM 206
Page 52 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

10
Na2HPO4-NaH2PO4 pH 7.2 at 4°C and 37°C. MpCPs ordered aggregates formation was 207
monitored by fluorescence spectroscopy using Thioflavin T (ThT; Sigma-Aldrich) probe. 208
ThT is known to fluoresce upon contact with amyloid-like aggregates (Biancalana and 209
Koide 2010, Khurana et al. 2005, Marcon et al. 2005). Beyond 0.8 mM of MpCP2, 3 and 5, 210
the protein solution became jellified, thus precluding sample handling and analysis. A fresh 211
0.22 uM filtered stock solution of 1 mg/mL of ThT in 50 mM Glycine-NaOH pH 8.5 was 212
prepared before each measurement. The stock concentration was double checked by 213
absorbance measurement at 475 nm in a Nanodrop 2000C spectrometer (Thermo Scientific) 214
and comparison with a calibration curve, and then diluted to 10 µM in 50 mM Glycine-215
NaOH pH 8.5 before use. Two nanomols of protein were diluted to 20 µL in 50 mM 216
Glycine-NaOH pH 8.5 solution and centrifuged at 20.000xg for 10 minutes. The 217
supernatant was removed and 200 µL of 10 µM ThT solution was added to the pellet (final 218
protein concentration of 10 µM considering complete protein precipitation). The sample 219
was then vortexed for one minute and the fluorescence measurements recorded (excitation 220
wavelength of 435 nm and emission wavelength of 485) on a plate reader 221
spectrofluorimeter (EnVision, Perkin Elmer) using a 96-well all-black-walled plate 222
(Eppendorf) and the default configurations of the equipment at 25oC. Time course of 223
aggregation of MpCP2 and 3 at 0.8 mM was monitored using ThT fluorescence during 3 224
and 30 days, respectively. 225
1,8-ANS and bis-ANS measurements were made accordingly to previously 226
published (Pazzagli et al. 2009), using MpCPs (at 0.8 mM) incubated at 37°C in low pH 227
solution for 5 and 20 days or control proteins incubated at 4°C in 20 mM Na2HPO4-228
NaH2PO4 pH 7.2 for 20 days. The final protein concentration used in the measurements was 229
2 µM, with 10 µM of ANS or 2 µM of bis-ANS. 230
Page 53 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

11
231
Cellulose fragmentation. Fragmentation activity of soluble and aggregated MpCP2 on 232
cellulose was assayed as described elsewhere (Baccelli et al. 2013). Briefly, 0.5 mm 233
diameter whatman n. 1 filter paper disks were incubated at 37 °C in a 96-well plate with 40 234
μM of protein or 20 mM Na2HPO4-NaH2PO4 pH 7.2 solution to a final volume of 100 uL. 235
The pH of the aggregated MpCPs solution was adjusted to 7.2 prior to the experiment. 236
After 72 h, 500 nm absorbance of the supernatant were measured in a plate reader 237
(EnVision; Perkin Elmer). 238
239
Spore germination. Spores obtained as described in the main text (RNA-seq methodology) 240
were gently centrifuged and ressuspended in distilled water and then added to soluble, or 241
aggregated (pH adjusted to 7.2) protein samples (40 μM), as well as 20 mM Na2HPO4-242
NaH2PO4 pH 7.2 and incubated in the dark at 28°C for 6 h. Samples were inspected in an 243
optical microscope for germination rate and germination tube length measurements by 244
using the ImageJ software. 245
246
Circular dichroism spectroscopy. Eight hundred micromolar of protein were incubated at 247
37°C in 20 mM Na2HPO4-NaH2PO4 pH 7.2 or in a low pH solution containing 20 mM 248
Na2HPO4-NaH2PO4 pH 7. 2 and 10% acetic acid for up to 30 days and then diluted to 10 249
µM in Milli-Q water for immediate measurement by far-UV CD spectroscopy (190-260 250
nm) using a JASCO J810 spectropolarimeter. The CD spectra were acquired at 20°C in a 251
0.1 cm optical path cuvette with a band-width of 1 nm, response time of 4 s, data pitch of 252
0.5 nm and scanning speed of 100 nm/min. Each data point was generated by averaging ten 253
Page 54 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

12
accumulations, subtracted from the blank spectrum and converted to mean residue molar 254
ellipticity (MRE [θ]). 255
256
Identification of native MpCPs by mass spectrometry. M. perniciosa FA553 was 257
cultivated in light malt extract culture medium (0.17% malt extract, 0.5% yeast extract) 258
complemented with 5% glycerol under agitation at 25°C. After seven days of growth, the 259
hyphae were removed by filtration through a nylon mesh and the culture medium dialyzed 260
against three changes of distilled water (Spectral/Por, Spectral Laboratories, Inc.; nominal 261
cutoff of membrane of 3.5 kDa). The dialyzed media was filtered through a 0.22 µM PVDF 262
syringe filter (Millipore) and concentrated 100-fold with a Amicon Ultra filter device 263
(Millipore) (MWCO of 10 kDa). The final sample was subjected to Comassie Blue-stained 264
15% SDS-PAGE analysis. The gel lanes were cut into small slices, the protein digested 265
with trypsin (Promega), extracted from the gel and desalted with the trapping column 266
Symmetry C18 (180 µm x 20mm, Waters) at a flow rate of 5 µl/min for 2 min. LC/MS-MS 267
analysis was performed with a C18 1.7 µm BEH 130 (100 µm x 100 mm) RP-UPLC 268
(nanoAcquity UPLC, Waters) chromatography coupled with nano-electrospray tandem 269
mass spectrometry on a Q-Tof Premier API mass spectrometer (MicroMass/Waters) over a 270
0-50% acetonitrile gradient in 0.1% formic acid for 45 min (flow rate of 600 nL/min). The 271
instrument was operated in MS positive mode, with data continuum acquisition from m/z 272
100–2,000Da at a scan rate of 1 s and an interscan delay of 0.1 s. The identification of the 273
peptides detected from LC/MS-MS experiments were done with the Mascot Distiller 274
v.2.3.2.0 (Matrix Science, Boston, MA) program using carbamidomethyl–cys as fixed 275
modification (monoisotopic mass 57.0215Da), methionine oxidation as variable 276
Page 55 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

13
modification (monoisotopic mass 15.9949), 0.1Da MS and MSMS fragment tolerances and 277
using the M. perniciosa database (17,105 sequences). 278
279
Transmission Electron Microscopy. Transmission Electron Microscopy was performed 280
with MpCP2 (0,8 mM) after 20 days of incubation at low pH. For visualization by negative 281
stain, protein samples at a concentration of 1.0 mg/mL were prepared in Tris-HCl buffer 282
(30 mM Tris-HCl pH 8.5, 50 mM NaCl). Holey carbon-coated grids were glow discharged 283
for 25 seconds at 15 mA using an easiGlow system (PELCO). A 3 µL sample was 284
deposited onto the grid for 60 seconds followed by two steps of blotting and staining with 3 285
µL of 2% uranyl acetate (30 seconds). Images were recorded at -3 µm defocus at x50,000 286
magnification using a Jeol JEM-2100 operating at 200 kV. Images were taken using an F-287
416 CMOS camera (TVIPS). 288
289
Studies of protein cleavage associated to self-assembling by mass spectrometry. Protein 290
cleavage studies were performed as decribed elsewhere (Pazzagli et al. 2009). Eight 291
hundred micromolar of MpCP1, 2, 3 and 5 were incubated in low pH solution for 20 days 292
for the self-assembly formation as confirmed by ThT fluorescence. Two hundred 293
micrograms (20 µL) were then fractionated with MicroconYM30 (cut-off of 30 kDa; 294
Milipore) and the flow through collected (FT). The Microcon was then washed with water 295
twice and the retentate dissolved in 20 µL of 0.1% TFA after vigorous vortexation 296
(retentate fraction). Both retentate and FT fractions were assayed for ThT fluorescence. 297
Half of each fraction was treated with 10 mM dithiothreitol for 25 min at 56°C and then 298
with 50 mM iodoacetamide during 30 min at room temperature for disulfide bridge 299
disruption. Samples of retentate fractions were also digested with tripsin (Promega) during 300
Page 56 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

14
16 hours at 37°C. All the fractions were dried in speed vacuum, dissolved in 0.1% formic 301
acid and submitted to LC/MS-MS analysis as described above, with data continuum 302
acquisition from m/z 100–3,000Da. 303
304
Injection of MpCPs in tobacco leaves. MpCP1, 2, 3 and 5, at a final concentration of 0.5 305
mg/mL (approximately 40 uM) in 20 mM Na2HPO4-NaH2PO4 pH 7.2 were individually 306
infiltrated into the central nerves leaves of Nicotiana tabacum var. Petite Havana (4-6 307
weeks old). Right after the infiltration, the leaves were detached and left inside of a dark 308
moist chamber for 24 hours, upon which they were inspected for necrotic symptom and 309
phytoalexin production. Phytoalexin production was measured by the auto-fluorescence 310
property of the phytoalexin compounds and registred by the Gel Logic 2200 Imaging 311
System (Carestream) (excitation at 365 nm and emission at 535 nm). 312
313
SM References 314
Baccelli, I., Luti, S., Bernardi, R., Scala, A., and Pazzagli, L. (2013). Cerato-315
platanin shows expansin-like activity on cellulosic materials. Applied Microbiology and 316
Biotechnology[Epub ahead of publication]. 317
Biancalana, M. & Koide, S. (2010). Molecular mechanism of Thioflavin-T 318
binding to amyloid fibrils. Biochim. Biophys. Acta 1804, 1405–1412. 319
Cavanagh, J., Fairbrother, W., Palmer III, A., Rance, M. & Skelton, N. (2007). 320
Protein NMR Spectroscopy: Principles and Practice, Academic Press, San Diego, CA. 321
Darriba, D., Taboada, G.L., Doallo, R. & Posada, D. (2011). ProtTest 3: fast 322
selection of best-fit models of protein evolution. Bioinformatics 27, 1164–1165. 323
Page 57 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

15
de Laureto, P., Frare, E., Battaglia, F., Mossuto, M., Uversky, V. & Fontana, A. 324
(2005). Protein dissection enhances the amyloidogenic properties of alpha-lactalbumin. 325
FEBS J. 272, 2176–2188. 326
de Oliveira, A., Gallo, M., Pazzagli, L., Benedetti, C., Cappugi, G., Scala, A., 327
Pantera, B., Spisni, A., Pertinhez, T. & Cicero, D. (2011). The structure of the elicitor 328
Cerato-platanin (CP), the first member of the CP fungal protein family, reveals a double 329
ψβ-barrel fold and carbohydrate binding. J. Biol. Chem. 286, 17560–17568. 330
Edgar, R.C. (2004). MUSCLE: multiple sequence alignment with high accuracy 331
and high throughput. Nucleic Acids Res. 32, 1792–1797. 332
Emsley, P., Lohkamp, B., Scott, W.G. & Cowtan, K. (2010). Features and 333
development of Coot. Acta Crystallogr. D 66, 486–501. 334
Frare, E., de Laureto, P., Zurdo, J., Dobson, C. & Fontana, A. (2004). A highly 335
amyloidogenic region of hen lysozyme. J. Molecul. Biol. 340, 1153–1165. 336
Keegan, R.M. & Winn, M.D. (2008). MrBUMP: an automated pipeline for 337
molecular replacement. Acta Crystallogr. D 64, 119–124. 338
Kerff, F., Amoroso, A., Herman, R., Sauvage, E., Petrella, S., Filée, P., Charlier, 339
P., Joris, B., Tabuchi, A., Nikolaidis, N., et al. (2008). Crystal structure and activity of 340
Bacillus subtilis YoaJ (EXLX1), a bacterial expansin that promotes root colonization. 341
P. Natl. Acad. Sci. USA 105, 16876–16881. 342
Khurana, R., Coleman, C., Ionescu-Zanetti, C., Carter, S.A., Krishna, V., 343
Grover, R.K., Roy, R. & Singh, S. (2005). Mechanism of thioflavin T binding to 344
amyloid fibrils. J. Struct. Biol. 151, 229–238. 345
Krieger, E., Joo, K., Lee, J., Lee, J., Raman, S., Thompson, J., Tyka, M., Baker, 346
D. & Karplus, K. (2009). Improving physical realism, stereochemistry, and side-chain 347
Page 58 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

16
accuracy in homology modeling: Four approaches that performed well in CASP8. 348
Proteins 77, 114–122. 349
Krissinel, E. & Henrick, K. (2004). Secondary-structure matching (SSM), a new 350
tool for fast protein structure alignment in three dimensions. Acta Crystallogr D 60, 351
2256–2268. 352
Livak, K., Schmittgen, T. D. (2001). Analysis of Relative Gene Expression 353
Data Using Real-Time Quantitative PCR and the 2-∆∆Ct Method. Methods. 25,402–354
408. 355
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., 356
McWilliam, H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., et al. (2007). Clustal 357
W and Clustal X version 2.0. Bioinformatics 23, 2947–2948. 358
Marcon, G., Plakoutsi, G., Canale, C., Relini, A., Taddei, N., Dobson, C.M., 359
Ramponi, G. & Chiti, F. (2005). Amyloid formation from HypF-N under conditions in 360
which the protein is initially in its native state. J. Molecul. Biol. 347, 323–335. 361
McCoy, A.J., Grosse-Kunstleve, R.W., Adams, P.D., Winn, M.D., Storoni, L.C. 362
& Read, R.J. (2007). Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–363
674. 364
Melton, L.D., & Smith, B.G. (2001). Isolation of plant cell walls and 365
fractionation of cell wall polysaccharides. In Current Protocols in Food Anal Chem, 366
John Wiley & Sons Inc, Hoboken, NJ. 367
Miller, G.L. (1959). Use of Dinitrosalicylic Acid Reagent for Determination of 368
Reducing Sugar. Anal. Chem. 31, 426–428. 369
Mishra, R., Sörgjerd, K., Nyström, S., Nordigården, A., Yu, Y. & 370
Hammarström, P. (2007). Lysozyme amyloidogenesis is accelerated by specific nicking 371
Page 59 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

17
and fragmentation but decelerated by intact protein binding and conversion. J. Mol. 372
Biol. 366, 1029–1044. 373
Mott, H. Nietlispach, D., Hopkins, L., Mirey, G., Camonis, J. & Owen, D. 374
(2003). Structure of the GTPase-binding Domain of Sec5 and Elucidation of its Ral 375
Binding Site. J. Biol. Chem. 278, 17053–17059. 376
Nylander, J.A., Wilgenbusch, J.C., Warren, D.L. & Swofford, D.L. (2008). 377
AWTY (are we there yet?): a system for graphical exploration of MCMC convergence 378
in Bayesian phylogenetics. Bioinformatics 24, 581–583. 379
Pazzagli, L., Zoppi, C., Carresi, L., Tiribilli, B., Sbrana, F., Schiff, S., Pertinhez, 380
T.A., Scala, A. & Cappugi, G. (2009). Characterization of ordered aggregates of cerato-381
platanin and their involvement in fungus-host interactions. Biochem. Biophys. Acta 382
1790, 1334–1344. 383
Perkins, D.N., Pappin, D.J., Creasy, D.M., & Cottrell, J.S. (1999). Probability-384
based protein identification by searching sequence databases using mass spectrometry 385
data. Electrophoresis 20, 3551–3567. 386
Perrakis, A., Morris, R. & Lamzin, V.S. (1999). Automated protein model 387
building combined with iterative structure refinement. Nat. Struct. Biol. 6, 458–463. 388
Ronquist, F. & Huelsenbeck, J.P. (2003). MrBayes 3: Bayesian phylogenetic 389
inference under mixed models. Bioinformatics 19, 1572–1574. 390
Sheldrick, G.M. (2010). Experimental phasing with SHELXC/D/E: combining 391
chain tracing with density modification. Acta Crystallogr. D 66, 479–485. 392
Sukumaran, J. & Holder, M.T. (2010). DendroPy: a Python library for 393
phylogenetic computing. Bioinformatics 26, 1569–1571. 394
Page 60 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

18
Tsiroulnikov, K., Chobert, J. & Haertlé, T. (2006). Copper-dependent 395
degradation of recombinant ovine prion protein. Phosphatidylinositol stimulates 396
aggregation and copper-driven disappearance of prion protein. FEBS J. 273, 1959–397
1965. 398
Vagin, A.A., Steiner, R.A., Lebedev, A.A., Potterton, L., McNicholas, S., Long, 399
F. & Murshudov, G.N. (2004). REFMAC5 dictionary: organization of prior chemical 400
knowledge and guidelines for its use. Acta Crystallogr. D 60, 2184–2195. 401
van Straaten, K.E., Barends, T.R.M., Dijkstra, B.W. & Thunnissen, A. W.H. 402
(2007). Structure of Escherichia coli Lytic transglycosylase MltA with bound 403
chitohexaose: implications for peptidoglycan binding and cleavage. J. Biol. Chem. 282, 404
21197–21205. 405
Winn, M.D., Ballard, C.C., Cowtan, K.D., Dodson, E.J., Emsley, P., Evans, 406
P.R., Keegan, R.M., Krissinel, E.B., Leslie, A.G.W., McCoy, A., et al. (2011). 407
Overview of the CCP4 suite and current developments. Acta Crystallogr. D 67, 235–408
242. 409
Zaparoli, G., Cabrera, O., Medrano, F., Tiburcio, R., Lacerda, G. & Pereira, G. 410
(2009). Identification of a second family of genes in Moniliophthora perniciosa, the 411
causal agent of Witches’ broom disease in cacao, encoding necrosis-inducing proteins 412
similar to cerato-platanins. Mycol. Res. 113, 61–72. 413
414
Author-Recommended Internet Resources 415
416
http://tree.bio.ed.ac.uk/software/tracer/ 417
http://www.bionmr.ualberta.ca/bds/software/xcrvfit/html-v4.0.12/index418
Page 61 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

19
SM Tables 419
Supplemental Table 1A: Peptide sequences identified by LC/MS-MS mass spectrometry of 420
cultivating media of M. perniciosa in vitro- grown necrotrophic mycelium. 421
Supplemental Table 1B: Sequence alignment between the CpCP, MpCPs 1-12 and Sm1 422
highlighting in yellow the N-X-S/T sequence predicted to be glycosylated by the NetNGlyc 423
1.0 online server. NGS sequence on Sm1 was experimentally shown to be glycosylated 424
(Vargas et al. 2008). The colors follow the color pattern stipulated for the clusters Mper1 425
(blue), Mper2 (green) e Epl1/Epl2 (light red) on Figure 2 of the main text. 426
Supplemental Table 2: X-ray crystallography data collection parameters and structure 427
refinement statistics. 428
Supplemental Table 3: Peptides identified by mass spectrometry of MpCP1, 2, 3 and 5 flow 429
through fraction after low pH-induced aggregation and fractionation. 430
431
SM Figures Legends 432
Supplemental Figure S1: MpCP proteins sequence alignment. ClustalW (Larkin et al. 2007) 433
was used to align all identified CPs from M. perniciosa as well as the CP from C. platani 434
(CpCP). In green, residues 100% conserved throughout all sequences. In cyan, MpCP3 435
residues involved on NAG4 binding (as shown by NMR and docking, and confirmed by 436
mutagenesis), as well as equivalent ones found conserved in other sequences. Equivalent 437
residues in CpCP are also among those mostly disturbed upon NAG4 binding (de Oliveira 438
et al 2011). The pairs of cysteines involved in disulfide bridges, which are also fully 439
conserved across the aligned sequences, are depicted in yellow and are connected by 440
Page 62 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

20
straight lines. The NAG4-binding regions in MpCP3, as judged by NMR and the docking 441
model (loops between β2 and β3 and helices α1 and α2) are surrounded by red boxes 442
Supplemental Figure S2: Crystal structure of MpCP1 highlighting the coordinating ions. 443
Cartoon representation of the polypeptide chain. Five strong peaks (over 7σ in height) were 444
observed in the Fourier difference electron density map for MpCP1, which, with respect to 445
their contributions to the dispersive component of the scattering factor, were interpreted as 446
three zinc ions (lying in the proximity of residues Asp69, Glu106 and Asp137), one sodium 447
ion (coordinating with Glu124) and a chloride ion (coordinating with the zinc ion near 448
Asp69). The ions of zinc, chloride and sodium are depicted as grey, green and purple 449
spheres, respectively. Respective coordinating residues are shown as sticks. Omit Fourier 450
difference map (at a sigma level of 3, green mesh), were calculated from the final 451
coordinate file, excluding the ions. 452
Supplemental Figure S3: Binding of NAG and NAG4 to MpCPs as detected by NMR 453
spectroscopy. Overlay of the 15N-HSQC spectra of 15N-labeled MpCP1, 2, 3 and 5 (all 454
proteins at 0.05 mM) without ligand (black) and with 10 mM (pink) of NAG (A) or titrated 455
with 0.1 mM (red), 2.5 mM (green), 5 mM (purple) and 10 mM (pink) of NAG4 (B). All 456
spectra were acquired with the same number of scans. When a protein and its ligand 457
interact, the chemical environment of the backbone amides changes and this usually causes 458
a change in chemical shift. Such changes can be grouped into three regimes: fast, 459
intermediate, and slow. The regime observed for any amide depends on the relationship 460
between the chemical shift measurement and the rate of exchange between the free and 461
bound states. If the exchange rate were higher than the chemical shift, a single peak would 462
appear at a position between the chemical shifts of the free and bound forms. If the 463
Page 63 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

21
exchange rate is lower than the chemical shift, two peaks would be observed, one for the 464
free form and one for the bound. In the intermediate case (the exchange rate is comparable 465
to the chemical shift difference), the peaks become broadened and may be unobservable 466
(Mott et al. 2003). During the titration of MpCP3 with NAG4, we could only observe peak 467
shifting phenomena, a clear indication that some residues of this complex are in the fast 468
exchange limit. On the MpCP5-NAG4 spectra, however, we could observe that, in addition 469
to the peak shifting behavior, the intensities of some peaks gradually decrease indicating 470
that the off-rate of the complex is in intermediate exchange on the NMR time scale. Since 471
the binding constant could not be calculated from the intensity data of the residues in 472
intermediate exchange because their peak intensities dropped sharply when the NAG4 473
concentration increased, the affinity estimated from residues in fast exchange may be 474
underestimate. (C) Scaled chemical shift differences between the 15N-HSQC spectra of the 475
proteins without NAG4 and incubated with 10 mM of NAG4, with affected residues 476
labeled. A cut-off of 0.05 ppm was considered. Scaling was performed according to the 477
equation ∆δ(15N + 1H) = [(∆δ15N/10)2 + (∆δ1H)2]1/2 (upper panel). MpCP5-NAG4 chemical 478
shift graphic depicting the plot of fractional changes in peak intensities for individual 479
backbone amide resonances of MpCP5 upon addition of NAG4. The axis Y presents the 480
relative peak intensity change (I-I0)/I0 of each assigned cross-peak caused by the addition of 481
NAG4, where I is the peak intensity measured in the presence of 10 mM NAG4 and I0 is 482
the peak intensity measured in the free protein. All the residues under either a fast or 483
intermediate off-rate regime are listed on the right followed by the chemical shift difference 484
value (middle panel) or relative peak intensity (lower panel). (D) Surface representation of 485
MpCP5 crystal structure highlighting the regions likely involved in NAG4 binding (in blue, 486
residues presenting chemical shift difference and in red residues displaying chemical shift 487
Page 64 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

22
broadening upon NAG4 addition). (E) ClustalW (Larkin et al. 2007) alignment of MpCP4, 488
5 and 11 showing the conservation of the MpCP5 residues which peaks displayed chemical 489
shift difference (blue) and signal suppression (red) upon NAG4 binding. The MpCP5 490
regions affected by the NAG4 binding involve residues present in β-strand 1 and α-helix 1, 491
as well as the connecting loop (Leu24, Iso25, Lys26, Trp27, Asp28, Asp29, Lys30 and 492
Phe31), the β-hairpin between β-strands 3 and 4 (Tyr88, Lys89, Gly90), part of β-strand 4 493
(Asn91, His92), the loop connecting β4 and β5 (Arg100), part of β5 (Val101, Glu102, 494
Glu103), the loop connecting β5 and α4 (Val108, Gly109 and Gly110), α-helix α4 (Thr111, 495
Asp112, Val114, Lys115, Asn116, Leu117, Thr118), the loop connecting α4 and α5 496
(Thr119, Phe120, Ala123), part of α-helix α5, the connecting loop between α5 and β6 and 497
the β-strand β6 (Tyr127, Trp129, Gly130, Thr131, Ala132, Gln133, Leu134). 498
Supplemental Figure S4: Fluorescence-based thermal shift assay showing MpCP3 and 5 499
residues involved in NAG4 binding. NAG4 binding to MpCP3 wild-type leads to an 500
increase on protein stability (higher melting temperature, Tm). Mutated MpCP3 protein 501
(Ala36Thr/Asp39Gly and Trp68Tyr) are less stable in the presence of NAG4 (left panel). 502
MpCP5 mutations Thr119Ser and Thr119Ala did not disturbed NAG4-induced thermal 503
stability, while Asn116Lys mutation apparently increased protein affinity for NAG4, as 504
judged by an increase on Tm upon NAG4 binding (right panel). These results indicate that 505
the MpCP5:NAG4 binding surface is rather large and likely involves the simultaneous 506
interaction of more than one molecule. 507
Supplemental Figure S5: Structure of MpCP1 and comparison with other proteins that 508
present a double Ψβ-barrel fold and interact with polysaccharides. (A) Cartoon 509
representation of MpCP1 (red) and E. coli expansin EXLX1 (Kerff et al. 2008) (gray; PDB 510
Page 65 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

23
ID: 3D30) superimposed. (B) Cartoon representation of MpCP1 and GH45 endoglucanase 511
MeCel45A (unpublished data) (PDB ID: 1WC2) superimposed. (C) Cartoon representation 512
of MpCP1 and EcMltA(D308A)–chitohexaose (van Straaten et al. 2007) (PDB ID: 2PI8) 513
superimposed. Chitohexaose molecule, co-crystallized with EcMltA, is represented by 514
spheres. Superposition using the secondary structure matching (SSM) algorithm (Krissinel 515
and Henrick 2004) was performed with the program Coot (Emsley et al. 2010). 516
Supplemental Figure S6: RMN spectroscopy of MpCP1 and MpCP5 after 4 days of 517
incubation in low pH solution. Overlay of initial (black) and after 4 days of incubation at 518
37oC in low pH (green) 15N-HSQC spectra of 15N-labeled MpCP1 and 5 (at 0.1 and 0.4 519
mM, respectively) indicating MpCP1 (left panel) unfolding tendency (the majority of the 520
resonance peaks in the 15N-HSQC spectra are clustered between 8.0 ppm and 8.8 ppm in 521
the 1H dimension, characteristic of unfolded structures; Cavanagh et al. 2007) and MpCP5 522
(right panel) aggregating behavior (aggregating protein has a tendency of losing intensity 523
signa; Cavanagh et al. 2007). 524
Supplemental Figure S7: Identification of the aggregating peptides by mass spectrometry. 525
The C. platani CP self-aggregation correlates with a fragmentation process as for some 526
amyloid-like proteins which undergo self-assemblage after cleavage of the polypeptide 527
chain (de Laureto et al. 2005, Frare et al. 2004, Mishra et al. 2007, Pazzagli et al. 2009, 528
Tsiroulnikov et al. 2006). Having shown that the MpCPs also undergo self-assembling 529
under mild-denaturing conditions we set out to learn whether this process was governed by 530
protein fragmentation. (A) Comassie stained SDS-PAGE gel of MpCP1, 2, 3 and 5 531
incubated at 4oC and 37oC under pH 7.2 and at 37oC in low pH solution for 10, 15, 20 and 532
25 days. It was evident in the electrophoresis results that after 10 days all the proteins at 533
Page 66 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

24
low pH had already presented alterations on their migration behavior compatible with a 534
degradation process. In order to identify the fragments, 0.8 mM of MpCP1, 2, 3 and 5 were 535
incubated for 20 days in low pH at 37°C or in pH 7.2 at 4°C and samples were fractionated 536
to separate the aggregated protein from the cleavage products (flow through fraction, FT). 537
(B) MpCPs and CpCP protein sequences aligned with ClustalW (Larkin et al. 2007). 538
Protein segment identified by LC/MS-MS in the flow-through fraction (cleaved-off 539
fragments) are colored according to the structure representation color. Aggregating peptides 540
are underlined. The disulfide bridges are indicated by connecting lines. The FT (which did 541
not present ThT fluorescence, Figure S8) were then analyzed by nanospray LC/MS-MS, 542
with a 3 KDa cut-off for mass fragmentation. FT fraction analysis of protein incubated in 543
pH 7.2 at 4°C did not reveal any peptide (data not shown). On the other hand, the observed 544
fragmentation pattern of the proteins incubated in low pH at 37°C revealed that the major 545
cleavage sites for the isoforms 1, 2, 3 and 5 are in the N-terminus and in the regions 546
peripheral to the two disulphide bridges, in clear accordance with what was described for 547
CpCP (Pazzagli et al. 2009). Several attempts of characterizing the aggregate-forming 548
peptides by LC/MS-MS were made, which included tripsinization of the retentate fraction, 549
but they all proved unsuccessful. By excluding what was identified in the FT fraction, we 550
concluded that MpCP1 presented the shortest aggregating-peptide (Ile70-Asp102), while in 551
the remaining isoforms they are formed by structurally correlated regions (Ser73-Arg91, 552
Ser72-Gly83 and Cys80-Lys89, for MpCP2, 3 and 5, respectively) added to non-553
consecutive segments that are hold together by the two disulphide in MpCP2 and 3. More 554
specifically, MpCP2 aggregating-peptide is covalently joined to helix α2 and part of α6, 555
MpCP3 to α2 (and adjacent loops) and part of α6, MpCP5 to helix α6 and β strands β6 and 556
Page 67 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

25
β6*. (C) Cartoon representation of the MpCP1 (red), 2 (blue), 3 (yellow) and 5 (green) 557
structures with the aggregating peptides colored in gray. See also Table S3. 558
Supplemental Figure S8: ThT fluorescence of retentate and flow through fractions of 559
MpCP1, 2, 3 or 5 aggregates before and after fractionation. Eight hundred micromolar of 560
each protein were incubated for 20 days in low pH solution and ThT fluorescence measured 561
for the total solution and for the flow through (FT) and retentate fractions, after 562
fractionation with MicroconYM30 (cut-off of 30 kDa, Milipore). Most of the ThT 563
fluorescence detected in the initial protein solution is measured in the retentate fraction, 564
clearly showing that it contains the aggregates. Background refers to ThT fluorescence 565
without the protein. MpCP1 aggregate presents low ThT fluorescence at 0.8 mM protein 566
concentration. 567
Supplemental Figure S9: MpCP1, 2, 3 or 5 injection into tobacco leaves did not induce 568
neither phytoalexin production nor necrosis. MpCP1, 2, 3 and 5 at a final concentration of 569
0.5 mg/mL were individually infiltrated into the central nerves leaves of Nicotiana tabacum 570
var. Petite Havana (4-6 weeks old). Right after the infiltration, the leaves were detached, a 571
picture was taken (left panel, 0 hours) and the leaves were kept inside of a dark moist 572
chamber for 24 hours, upon which they were inspected for phytoalexin production (A) and 573
necrotic symptom (B), having tested negative for both outputs (right panels, 24 hours). 574
575
Page 68 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

1
Supplemental Table 1 A 576
Peptide sequences identified by LC/MS-MS mass spectrometry of cultivating media of M. perniciosa in vitro- grown necrotrophic 577
mycelium. Mass (exp), experimental mass; Mass (calc), calculated mass. In yellow, segment identified by the NetNGlyc 1.0 as 578
putatively glycosylated (see Supplemental Table 1 B). 579
580 581 582 583
Protein Peptide sequence identified m/z Charge
Mass
(exp),
Da
Mass
(calc),
Da
Number
of times
identified MpCP2 FDNTYDNASGSMNTVACSTGANGLSQR 1419.5608 +2 2837.1070 2837.1981 5
FDNTYDNASGSMNTVACSTGANGLSQR 946.7344 +3 2837.1815 2837.1981 2
SINLVAIDHAGNGFNVAQAAMDELTNGNAVALGTIDVQSQQVAR 1498.4107 +2 4492.2103 4492.2405 5
MpCP3 TIHVVGVDVAGNGFNVGQR 970.0119 +2 1938.0092 1938.0072 1
TIHVVGVDVAGNGFNVGQR 647.0073 +3 1938.0001 1938.0072 8
GYSTFGSVPSYVGAVDTITGWNSESCGTCYQITWSGTGK 1394.2909 +3 4179.8509 4179.8467 4
MpCP7 LQYDPTYDNR 642.7943 +2 1283.5740 1283.5782 8
GFSTFGSLPNFPR 713.8508 +2 1425.6870 1425.7041 12
DGSLGTTACSDGPNGLITK 932.4243 +2 1862.8340 1862.8680 4
Page 69 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

2
Supplemental Table 1 B
CpCP 1 -----MKFSI------LPMIASAMA------------------VSISYDPIYA-ADLSMGSVACSNGDH--GLMAQ--YPTLGEVPGF-----PNVGGIPDIAGWDSPSCGTCWKVTIPN 81
Sm1 1 -----MQLSNIF--TLALFTAAVSA------------------DTVSYDTGYDNGSRSLNDVSCSDGPN--GLETRYHWSTQGQIPRF-----PYIGGAAAVAGWNSASCGTCWKLQYS- 87
MpCP1 1 -----MKSIAIFTPILILLTISAGA------------------VKLSYDEAYDNPSSSLLSVTCSDGEN--GLYPK--YRTFGDLPGF-----PCIGGSSDIAGYNSPNCGSCYQLTYSS 88
MpCP9 1 -----MKFFALC--STLALVSSTTA------------------VNLAWDDAYNDRAGSLNAVACSDGAN--GLETRFGFAVFGDIPTF-----PFVGGAPNIPGWNSSACGTCWALTFVN 88
MpCP4 1 -----MKSFTTL----LSLGFVASVASALPSPQGDPTPATPAKYRLSHNPVYDNRSMEIGRLACGGGEN--GLVT-KGYRTLGDLPFF-----DNIGAA-HPSSYNSLECGSCWNLTYK- 101
MpCP5 1 -----MKLFVIL----LSLSLSLSA------------SCSTANWLIKWDDKFQNDTLSISEFKCSAALAKLGPDPKHPPTKLGEVLNF-----PHFVAA----PEAQTECGSCWKLRYK- 89
MpCP11 1 -------MKTTL----LSLGFAASVAFALPSSQGGSA---PEQYRVTHNPVYGNSSQRISDLACGGGEN--GIDTKWGYQTLGQIPNF-----DDVGAA-HVSSYNSPECGGCWYLEYK- 97
MpCP2 1 -----MKFTTTI--IALALAASTGA------------------VQLRFDNTYDNASGSMNTVACSTGAN--GLS-Q-RFPTFGSVPTF-----PHIGASSDIGGFNSPACGNCYTISFTF 86
MpCP3 1 -----MKFIAAV----ALLATSAVA------------------VQLQYDPVYDNADQSFGTVACSDGPN--GMLTK-GYSTFGSVPSY-------VGAVDTITGWNSESCGTCYQITWS- 82
MpCP6 1 MKAPALLTSLIL----TLLPLHASA------------------VTLAYDTVYDDRSRSLATVACSDGRN--GLLTR-NFTTFGSLPSF-----PRIGGAQAITGWNSSACGTCWEVTYTN 90
MpCP7 1 -----MKFTAAV----ALLATSAAA------------------VRLQYDPTYDNRDGSLGTTACSDGPN--GLITK-GFSTFGSLPNF-----PRIGAVDAITGWNSPQCGSCWQVTWN- 84
MpCP8 1 -----MKFIVAL----ALLTTSAVA------------------VQLQYDTAYDHADQSLSSVACSNGEN--GLLTK-GYTTFGSVKGTTA--STYVGAAEAITGWNSAACGNCYQIKWS- 87
MpCP10 1 -----MKFVVTI----AILATSAAA---LSVKR---RNTDGQSYKLRYVSEFDNGDYSFNNVACSNGEH--GMPAK-KLNKFGDLPRNANGVNVYGGGGFAVSGYGDEECGSCWEVYYND 102
MpCP12 1 -----MKFIAAI----AILATSAAA---LSIKR---QSADG-TLKLRYINEYDNSDFTLSDHGSC--SN--ILSPK-GINTVGDVNKSVDGVNVYPGGAFPVKEWNSDSCGTCWKVSYDD 99
CpCP 82 ----GNSIFIRGVDSGRG--GFNVNPTAFTKLVGSTE-----AGRVDNV-NYVQVDLSNCINGAN------------------------------------------------------- 134
Sm1 88 ----GHTIYVLAVDHAAS--GFNIALDAMNALTGGQA----VQLGRVSA-TATQVPVKNCGL---------------------------------------------------------- 138
MpCP1 89 AHTTPKSIYMVAIDRSAE--GFTASKQAMDDLTNKRA----EELGTVNV-DVRKVDFSRCERKS-------------------------------------------------------- 145
MpCP9 89 STGT-HTINFTAIDAGGS--DFVTGRVALDLLTNGQA----EQLGVIPV-NATAVNASACGL---------------------------------------------------------- 142
MpCP4 102 ----GNWAYIIAVDNASDEDLFVISDEANKALTRVNGRNEGIEQDIVDLDSAHEVDRACCGFNTGTQCPTALNND-----------------------G--------------------- 173
MpCP5 90 ----GNHAFVTVVDRVEEANLFVGGTDLVKNLTTFNGAPEGYDWGTAQLFSAYQVDGSCCQQNTGKQCGDP------------------------------------------------- 156
MpCP11 98 ----GNSLYVTAVDNAAGENLFVISDGAIRSLTTVNGVNEGIEKGTVDLDSAYEVDHSYCHLPDPTGNTRTGTDK----KSPSTGKKSPT---SSNNYSV-------------------- 186
MpCP2 87 -QGVTRSINLVAIDHAGN--GFNVAQAAMDELTNGNA----VALGTIDV-QSQQVARSVCGL---------------------------------------------------------- 140
MpCP3 83 --GTGKTIHVVGVDVAGN--GFNVGQRAMDDLTNGQA----VALGNIDV-TATLVDKSACGL---------------------------------------------------------- 135
MpCP6 91 SSGIAKSLNITAIDVAGA--GFNLAQVAMDELTNGHA----VEFGRVEV-TSRQVNASACGL---------------------------------------------------------- 145
MpCP7 85 --GN--SINILGMDVAGN--GFNIAQAAMDELTNGQA----VALGNIDV-QATQVDASACGL---------------------------------------------------------- 135
MpCP8 88 --GSDRTINVIAID---------------------------------------------------------------------------------------------------------- 99
MpCP10 103 -----RKVRVIVVDTAND--GFNLPKKGMDELTNGQA----YDLGVIDV-TTQKLKPADCGLNSTEGQTSMNGQTSTQGKPSTQGKTSTQEKTSTNGKALTEGKTSTNEQPPTQENTSTN 210
MpCP12 100 -NGTVRTIRVVAVDTAQE--GFNVAQKAMDELTDGKA----YEKGFVTV-TAEQLSADDCKIGTN------------------------------------------------------- 156
CpCP -----------------------------------------------------------------------------------------------------------------
Sm1 -----------------------------------------------------------------------------------------------------------------
MpCP1 -----------------------------------------------------------------------------------------------------------------
MpCP9 -----------------------------------------------------------------------------------------------------------------
MpCP4 -----------------------------------------------------------------------------------------------------------------
MpCP5 -----------------------------------------------------------------------------------------------------------------
MpCP11 -----------------------------------------------------------------------------------------------------------------
MpCP2 -----------------------------------------------------------------------------------------------------------------
MpCP3 -----------------------------------------------------------------------------------------------------------------
MpCP6 -----------------------------------------------------------------------------------------------------------------
MpCP7 -----------------------------------------------------------------------------------------------------------------
MpCP8 -----------------------------------------------------------------------------------------------------------------
MpCP10 211 KQTSTEGKTSTNGKTLTEEKTSTNEQTSTNGKALTEGKTSTNEQPPTQENASTNEQTSTEGKTLTQEKTSTNEQTPTQENTSTNGKTLTEGKAATEEQTATEGKTSTEGMTAS 323
MpCP12 -----------------------------------------------------------------------------------------------------------------
Page 70 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

3
Supplemental Table 2
Data Collection
MpCP1 (3SUJ) MpCP2 (3SUK) MpCP3 (3SUL) MpCP5 (3SUM)
Beamline, at LNLS - Brazil W01B-MX2 W01B-MX2 D03B-MX1 D03B-MX1
Wavelength (Å) 1.4586 1.4586 1.4299 1.4370
Space group P43212 P1211 P3121 P212121
Cell parameters a, b, c (Å) 45.3, 45.3, 105.74
29.5, 86.0, 43.6, (β=106.8o)
48.3, 48.3, 94.0 49.7, 107.9, 112.7
Resolution range (Å) 14.61-1.34 (1.41-1.34)
14.62-1.34 (1.41-1.34)
14.11-1.63 (1.72-1.63)
23.02-1.87 (1.97-1.87)
Unique reflections 25598 (3654) 43181 (5984) 16287 (2314) 50913 (7306)
Average redundancy 39.3 (36.9)a 8.3 (8.2) 13.2 (13.2) 6.3 (6.0)
Rsymm (%) 8.5 (51.8) 6.5 (27.0) 5.4 (59.0) 8.4 (32.7)
Completeness (%) 99.8 (100)a 92.4 (87.7) 99.2 (98.4) 99.8 (99.6)
I/σ(I) 42.1 (11.7) 22.2 (8.9) 25.5 (4.3) 13.2 (4.4)
Monomers per asymmetric unit 1 2 1 4
Solvent content (%) 37.3 40.33 50.88 51.28
Number of sites found (expected) 8 (7) - - -
Mean map CC (%) 76.7 - - -
Refinement
Resolution range (Å) 14.51-1.34 14.50-1.34 13.98-1.63 20.00-1.87
Reflections (cross-validationb) 24215 (1299) 41002 (2177) 15419 (825) 48195 (2571)
Rfactor/Rfree (%) 18.43/22.00 17.32/20.50 23.77/26.77 16.51/22.33 Average B-factor/r.m.s.d. (Å2)
main chain (no. of residues) side chain (no. of residues) ligands (no. of molecules) solvent (no. of molecules)
11.56/0.86 14.93/2.18
19.96 26.63
10.86/0.79a 12.87/1.59a
- 24.6
20.76/0.99 22.56/1.96
- 30.73
19.25/0.79b 21.46/2.21b
- 31.13
R.m.s.d. from standard geometry bond length (Å) bond angles (°)
0.025 2.254
0.024 1.698
0.023 2.23
0.022 1.887
Ramachandran plot most favored (%) allowed (%) outlier (%)
95.7 3.5 0.8
96.6 2.5 0.9
94.1 4.2 1.7
96.8 2.5 0,7
a Friedel pairs treated separately. b Cross-validation test set size of 5%. Data for outer shell shown in parentheses.
Page 71 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

4
Supplemental Table 3 MpCP1 fragments detected in the flow through fraction (in red):
a Score > 5 was considered significative by Mascot (Perkins et al. 1999) analysis.
Peptide
Number
(MpC1)
Peptide sequence identified m/z Charge
Number of
times
identified
Scorea
1 SAVKLSYD 882,4640 +1 1 14
2 FSRCERKS 535,2494 +2 3 12
3 SAVKLSYDEAY 632,3006 +2 2 32
4 SAVKLSYDEAYD 680,7709 +2 10 45
5 NGLYPKYRTFGD 715,8497 +2 3 40
6 GENGLYPKYRTFGD 808,8441 +2 5 48
7 VRKVDFSRCERKS 833,9015 +2 2 9
8 VRKVDFSRCERKS 556,2705 +3 1 6
9 NVDVRKVDFSRCERKS 665,6593 +3 1 12
10 SAVKLSYDEAYDNPSSSLL 1029,9834 +2 3 53
11 SAVKLSYDEAYDNPSSSLLS 1073,4673 +2 1 21
12 TVNVDVRKVDFSRCERKS 732,3623 +3 1 4
13 LTNKRAEELGTVNVDVRKVD 752,7372 +3 1 27
14 GENGLYPKYRTFGDLPGFPCIGGSSD 935,4387 +3 1 14
15 NPSSSLLSVTCSDGENGLYPKYRTFGD 988,7849 +3 2 54
16 RSAEGFTASKQAMDDLTNKRAEELGTVNVD 1085,1835 +3 1 2
17 EAYDNPSSSLLSVTCSDGENGLYPKYRTFGD 1148,1814 +3 1 13
Page 72 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

5
MpCP2 fragments detected in the flow through fraction (in red):
Peptide
Number
(MpCP2)
Peptide sequence identified m/z Charge
Number of
times
identified
Scorea
1 SDIGGFN 709,3103 +1 4 21
2 SINLVAI 729,4543 +1 1 16
3 PTFPHIG 384,7053 +2 2 24
4 SGAVQLRFD 496,7491 +2 2 61
5 PTFPHIGASSD 564,7704 +2 2 47
6 TIDVQSQQVAR 622,8301 +2 2 75
7 TFGSVPTFPHIG 630,3252 +2 3 13
8 GTIDVQSQQVAR 651,3428 +2 2 92
9 SGAVQLRFDNTY 685,8399 +2 4 46
10 LGTIDVQSQQVAR 707,8842 +2 1 84
11 SGAVQLRFDNTYD 743,3260 +2 14 64
12 SINLVAIDHAGNGFN 771,3869 +2 2 42
13 SGAVQLRFDNTYDNA 835,8828 +2 3 62
14 SINLVAIDHAGNGFNVA 856,4368 +2 4 11
15 NVAQAAMDELTNGNAVAL 901,4390 +2 2 9
16 SGAVQLRFDNTYDNASG 907,9082 +2 3 80
17 GNAVALGTIDVQSQQVAR 913,9869 +2 3 73
18 SINLVAIDHAGNGFNVAQ 920,4665 +2 3 48
Page 73 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.
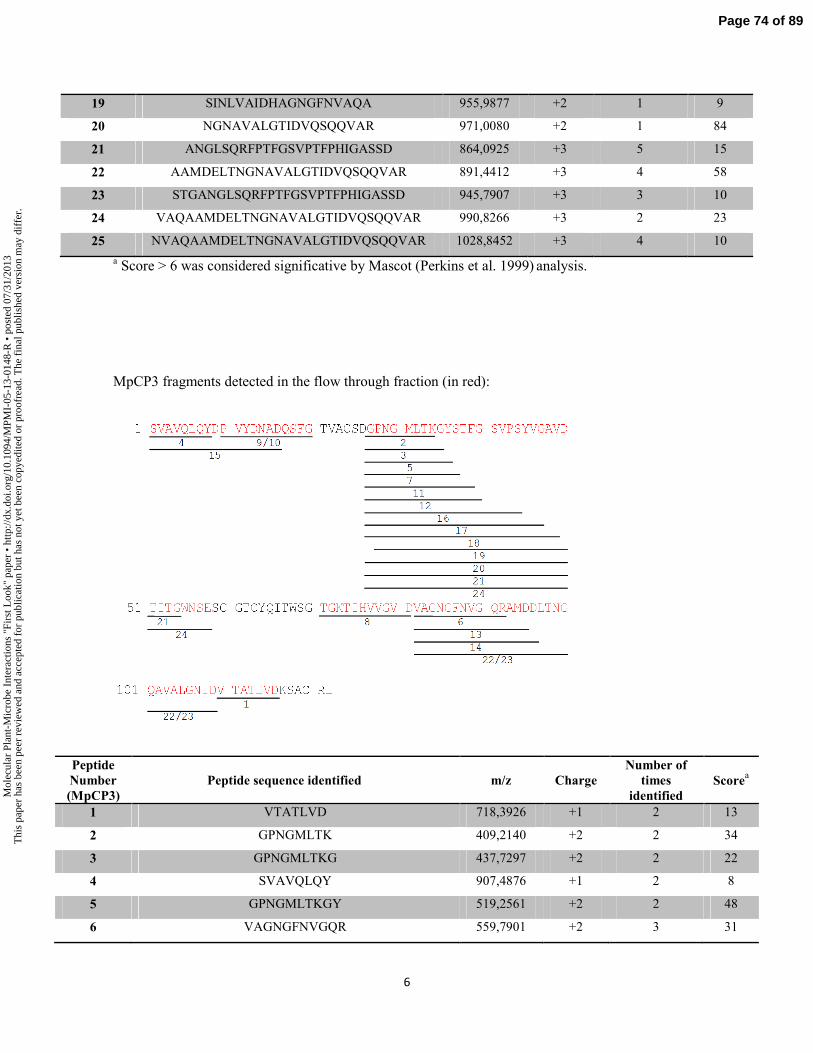
6
19 SINLVAIDHAGNGFNVAQA 955,9877 +2 1 9
20 NGNAVALGTIDVQSQQVAR 971,0080 +2 1 84
21 ANGLSQRFPTFGSVPTFPHIGASSD 864,0925 +3 5 15
22 AAMDELTNGNAVALGTIDVQSQQVAR 891,4412 +3 4 58
23 STGANGLSQRFPTFGSVPTFPHIGASSD 945,7907 +3 3 10
24 VAQAAMDELTNGNAVALGTIDVQSQQVAR 990,8266 +3 2 23
25 NVAQAAMDELTNGNAVALGTIDVQSQQVAR 1028,8452 +3 4 10
a Score > 6 was considered significative by Mascot (Perkins et al. 1999) analysis.
MpCP3 fragments detected in the flow through fraction (in red):
Peptide
Number
(MpCP3)
Peptide sequence identified m/z Charge
Number of
times
identified
Scorea
1 VTATLVD 718,3926 +1 2 13
2 GPNGMLTK 409,2140 +2 2 34
3 GPNGMLTKG 437,7297 +2 2 22
4 SVAVQLQY 907,4876 +1 2 8
5 GPNGMLTKGY 519,2561 +2 2 48
6 VAGNGFNVGQR 559,7901 +2 3 31
Page 74 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

7
7 GPNGMLTKGYS 562,7736 +2 1 38
8 TGKTIHVVGVD 563,3158 +2 2 30
9 PVYDNADQSFG 1212,5112 +1 1 47
10 PVYDNADQSFG 606,7616 +2 1 29
11 GPNGMLTKGYSTF 686,8341 +2 2 56
12 GPNGMLTKGYSTFG 715,3410 +2 4 78
13 VAGNGFNVGQRAMD 718,3234 +2 3 73
14 VAGNGFNVGQRAMDD 775,8395 +2 4 76
15 SVAVQLQYDPVYDNAD 898,9180 +2 19 18
16 GPNGMLTKGYSTFGSVPS 900,4334 +2 1 9
17 GPNGMLTKGYSTFGSVPSYVG 1060,0060 +2 4 33
18 GPNGMLTKGYSTFGSVPSYVGAV 1145,0629 +2 3 7
19 PNGMLTKGYSTFGSVPSYVGAVD 1174,0703 +2 1 10
20 GPNGMLTKGYSTFGSVPSYVGAVD 1202,5554 +2 2 71
21 GPNGMLTKGYSTFGSVPSYVGAVDTITG 1388,6716 +2 3 15
22 VAGNGFNVGQRAMDDLTNGQAVALGNID 939,7775 +3 5 30
23 VAGNGFNVGQRAMDDLTNGQAVALGNID 1409,1759 +2 2 22
24 GPNGMLTKGYSTFGSVPSYVGAVDTITGWNSE 1098,1767 +3 4 32
a Score > 6 was considered significative by Mascot (Perkins et al. 1999) analysis.
MpCP5 fragments detected in the flow through fraction (in red):
Page 75 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

8
Peptide
Number
(MpCP5)
Peptide sequence identified m/z Charge Number of
times identified Score
a
1 SNWLIK 760,4230 +1 1 37
2 WLIKWD 860,4585 +1 1 22
3 PKHPPTKL 459,2786 +2 1 11
4 SAALAKLGPD 942,5171 +1 2 45
5 SNWLIKW 473,7583 +2 2 17
6 SNWLIKW 946,5098 +1 3 16
7 PKHPPTKLG 487,7912 +2 1 13
8 SNWLIKWD 531,2532 +2 4 30
9 SNWLIKWD 1061,5071 +1 2 41
10 PKHPPTKLGE 552,3131 +2 1 11
11 SNWLIKWDD 588,7675 +2 3 35
12 SNWLIKWDD 1176,5611 +1 2 39
13 SNWLIKWDDK 652,8315 +2 2 22
14 PKHPPTKLGEVL 658,3866 +2 2 30
15 LVKNLTTFNGAPEG 730,8922 +2 1 6
16 PKHPPTKLGEVLNF 526,2931 +3 1 2
17 WLIKWDDKFQND 804,3898 +2 3 11
18 LVKNLTTFNGAPEGY 812,4177 +2 3 6
19 LVKNLTTFNGAPEGYD 869,9199 +2 3 20
20 SNWLIKWDDKFQND 603,6128 +3 4 32
21 SNWLIKWDDKFQND 904,9174 +2 6 35
22 PKHPPTKLGEVLNFPH 604,3268 +3 4 25
23 PKHPPTKLGEVLNFPHF 653,3468 +3 3 49
24 WDDKFQNDTLSISEFK 658,3108 +3 1 21
25 TLSISEFKCSAALAKLGPD 670,0140 +3 1 39
26 PKHPPTKLGEVLNFPHFVAAPE 809,0981 +3 1 5
27 PKHPPTKLGEVLNFPHFVAAPEAQ 875,4647 +3 3 17
28 KFQNDTLSISEFKCSAALAKLGPD 880,7803 +3 2 34
29 GNHAFVTVVDRVEEANLFVGGTDLVK 929,4800 +3 5 28
a Score > 6 was considered significative by Mascot (Perkins et al. 1999) analysis.
Page 76 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

9
Supplemental Figure S1
.
Page 77 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

10
Supplemental Figure S2
Page 78 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

11
Supplemental Figure S3A
Page 79 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

12
Supplemental Figure S3B
Page 80 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

13
Supplemental Figure S3C
Page 81 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

14
Supplemental Figure S3D
Page 82 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

15
Supplemental Figure S3E
Page 83 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

16
Supplemental Figure S4
Page 84 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

17
Supplemental Figure S5
Page 85 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

18
Supplemental Figure S6
Page 86 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

19
Supplemental Figure S7
Page 87 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

20
Supplemental Figure S8
Page 88 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.

21
Supplemental Figure S9
Page 89 of 89M
olec
ular
Pla
nt-M
icro
be I
nter
actio
ns "
Firs
t Loo
k" p
aper
• h
ttp://
dx.d
oi.o
rg/1
0.10
94/M
PMI-
05-1
3-01
48-R
• p
oste
d 07
/31/
2013
Thi
s pa
per
has
been
pee
r re
view
ed a
nd a
ccep
ted
for
publ
icat
ion
but h
as n
ot y
et b
een
copy
edite
d or
pro
ofre
ad. T
he f
inal
pub
lishe
d ve
rsio
n m
ay d
iffe
r.