g anirep-4711; article in press animal reproduction...
TRANSCRIPT

GA
Rg
TCa
Gb
c
ARRAA
KSRGS
1
rdfisiw
0h
ARTICLE IN PRESS ModelNIREP-4711; No. of Pages 9
Animal Reproduction Science xxx (2013) xxx– xxx
Contents lists available at SciVerse ScienceDirect
Animal Reproduction Science
jou rn al hom epage : w ww.elsev ier .com/ locate /an i r eprosc i
eproductive impairment and endocrine disruption inoldfish by feeding diets containing soybean meal
ahere Bagheria,∗, Mohamad Reza Imanpoorb, Valiollah Jafarib,atherin Bennetau-Pelisseroc
Young Researchers Club, Gorgan Branch, Islamic Azad University, Gorgan, Iran & Scientific board, fisheris group, Gonbad university,onbad, IranScientific Board, Fisheries Group, Gorgan University of Agricultural Sciences and Natural Resources, IranENITA de Bordeaux, 1, cours du Ge’ne’ral de Gaulle, 33 175 Gradignan cedex, France
a r t i c l e i n f o
rticle history:eceived 22 May 2012eceived in revised form 5 February 2013ccepted 10 February 2013vailable online xxx
eywords:ex steroideproductionoldfishoybean
a b s t r a c t
A long-term feeding experiment was conducted to investigate the inclusion of soybeanmeal in diets for goldfish (Carassius auratus) on fish reproduction. In the present study, 20weeks after hatching, goldfish with an initial average weight of 2 ± 0.03 g (mean ± SD) weredivided into 12 groups (three tanks per dietary treatment) and fed 400 g kg−1 crude pro-tein diets. The four experimental diets were as follows: diet 1, fish meal (FM); diet 2, 35%soybean meal (SBM35%); diet 3, 65% soybean meal (SBM65%); diet 4, 100% soybean meal(SBM100%). After feeding with experimental diets, the impact on reproduction was investi-gated. In both males and females, the plasma testosterone (T) was significantly decreased,while 17�-estradiol (E2) levels were significantly increased. Levels of 17�, hydroxypro-gesterone. (17-OH-P) did not differ as a result of soybean meal feeding in either males orfemales. The average number of eggs spawned and sperm quality were reduced on feed-ing with soybean inclusion. Histological examination showed impact on oocyte maturationprogress and spermatogenesis process in female and male fish, respectively. In addition,
feeding goldfish with soybean meal until maturation caused reduction in fertilization andhatching rates in parallel to increasing soybean meal inclusion. The results demonstratedthat inclusion of soybean meal might cause sex hormone biosynthesis disruption and repro-ductive impairments in fish, ultimately decreased fertilization as well as hatching rates inthe offspring.. Introduction
The aquaculture production has been growing up andapid growth of aquafeed production for satisfying theemand for aquaculture industry is inevitable. Becausesh meal is the most limited, unpredictable and expen-
Please cite this article in press as: Bagheri, T., et al., Reproductfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013
ive constitutes in diets for cultured species in aquaculturendustry, the issue that the aquaculture industry is facing
ith in this case is looking for cheaper, more sustainable
∗ Corresponding author. Tel.: +98 9378322339; fax: +98 1712339721.E-mail address: [email protected] (T. Bagheri).
378-4320/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.anireprosci.2013.02.003
© 2013 Elsevier B.V. All rights reserved.
protein sources and environmentally friendly alternativesto fish meal (Gatlin III et al., 2007). In the meantime, sincesoybean meal is high in crude protein, more digestibile andhas a relatively well-balanced amino acid profile as well aslower price and more sustainable protein source than fishmeal, it has been evaluated as a suitable ingredient for fishfeed (Carter and Hauler, 2000; Ng et al., 2006; Gatlin IIIet al., 2007; Imorou Toko et al., 2008; Yue and Zhou, 2008).
Although it is known to be one of the most nutritious
ive impairment and endocrine disruption in goldfish by), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
plant protein sources (Mateos-Aparicio et al., 2008), theuse of soybean meal has not been completely admittedthroughout the aquaculture industry because there is con-cern about some biologically active compounds within the

ING Model
oduction
ARTICLEANIREP-4711; No. of Pages 9
2 T. Bagheri et al. / Animal Repr
soybean-based diets that have influences on fish health,growth, and reproductive development (Ng et al., 2006).
In fact, one of the reasons for uncertainty to usesoybean meal in fish feeds is it contains high levels ofphenolic compounds which are known as estrogenic com-pounds and called isoflavonoid phytoestrogens (Dixon,2004). The three most abundant isoflavones found in soy-beans are genistein (4′,5,7- trihydroxyisoflavone), daidzein(4′,7-dihydroxyisoflavone), and glycitein (4′,7-dihydroxy-6-methoxyisoflavone) (Ng et al., 2006).
Isoflavones structures are similar to estrogen and areknown to have estrogenic effects by binding to estrogenreceptors (Kuiper et al., 1998; Ng et al., 2006). The possibil-ity of interactions between isoflavones and the endocrinesystem of organisms has been the focus of several stud-ies in both mammals (Kuiper et al., 1997, 1998; Friendmanand Baron, 2001; Barrett, 1996; Woclawek-Potocka et al.,2005), and fish (Van den Ingh and Krogdahl, 1990; Tollefsonet al., 2001; Latonnelle et al., 2002).
In the present study, we investigated the impact oflong-term feeding with inclusion of soybean meal in gold-fish diets on endocrine status and reproduction. Endpointssuch as cumulative egg production, egg diameter, spermdensity, sperm production of adult goldfish as well asgonad histology, and measurement of sex steroid levels,including testosterone (T), 17�-estradiol (E2) and 17�,hydroxyprogesterone. (17-OH-P), were examined to studythe reproductive influence of soybean meal. The hatchingrates of offspring were also examined. Because goldfishendocrine system has been widely studied and charac-terized and because it is a laboratory model, we chose itfor these studies (Trudeau, 1997; Blazquez et al., 1998;Mimeault et al., 2005).
2. Material and methods
2.1. Experimental fish and rearing condition
Eggs obtained from seven female goldfish, Carassiusauratus, were fertilized with sperm taken from five males.After hatching, larvae were fed with freshly hatched brineshrimp nauplii (Artemia salina) for a period of 4 weeks.Fish were fed with the commercial goldfish pellets (Energy,Thailand). The experiment emerged 20 weeks after hatch-ing (June 2011). The fish (the mean body weight 2 g) weredivided evenly among four groups (120 fish per group).Each group was placed in a 400 l fiberglass aquarium, fedand maintained for one year until maturation. The fishwere fed with experimental diets, presented in Table 1.In order to accustom the fish to experimental food, fishwere gradually fed with experimental diet within a weekby 20%, 40%, 60%, 80% and 100% replacement to granulatedfood. The time we completely commenced feeding fishwith experimental diet was designed as zero point. Feedingwith experimental diets lasted from June to December fora period of 28 weeks. The amount of genistein and diadzeinwithin the food is presented in Table 1 showing each treat-
Please cite this article in press as: Bagheri, T., et al., Reproductfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013
ment diet composition.The daily feeding rate was 3% of fish body weight and fed
three times a day during the experimental period (Yanaret al., 2008). Water in fiberglass tanks was changed twice
PRESS Science xxx (2013) xxx– xxx
a week. In order to maintain adequate oxygen through-out the experiment, compressed aeration was used. Duringthe experiment time, fish were monthly weighed to adaptfeeding rations to actual fish body weight.
2.2. Sex hormones analysis
Fish were sampled in the morning before noon. Sixfish were blood sampled from each treatment for analyz-ing sex steroids (T, E2 and 17-OH-P). Blood samples weretaken from the caudal vessel through cutting caudal pedun-cle, centrifuged for 3 min at 13,000 g and the plasma wascollected and stored at −20 ◦C until assayed for steroidswith ELISA (enzyme-linked immunosorbent assay) method(Mimeault et al., 2005).
Body and gonad weights were recorded and the gonado-somatic index [GSI = (gonad weight/body weight) × 100]was determined for evaluating the gonad developmentduring experimental duration (Spanò et al., 2004).
2.3. Proximate analyses of experimental diets
Moisture content was determined by drying the feedsin an oven at 110 ◦C for 24 h. Crude protein of the dietswas determined by Kjeldahl method (Total Kjeldahl Nitro-gen × 6.25) (AOAC, 1990). Crude fat was estimated by thesoxhlet extraction method using petroleum ether (boilingpoint 60 ◦C) in the electro-thermal soxhlet apparatus. Todetermine the ash contents, dried samples were inciner-ated in a muffle furnace at 550 ◦C for 24 h (AOAC, 1990).Genistein and diadzein contents were detected with ELISAmethods previously described by Bennetau-Pelissero et al.(2003).
2.4. Gamete quality evaluation
At the end of the feeding experiment, from each treat-ment 14 females and 9 males were selected and transferredto the breeding tanks. For the induction of natural con-ditions in “winter period”, the water temperature wasdecreased to 10 ◦C and the illumination was turned off.Fish were kept for two weeks in winter condition. Afterthen, water temperature was elevated to 20 ◦C in orderto induce spring condition. Finally, intramuscular injec-tions of Ovaprim (Syndel laboratories, India) at 0.5 �L/g(estimated brood fish weight) were applied for spawning.To measure fecundity, eggs were collected by strippingfrom three females from each treatment, and fifteen ran-domly selected eggs were applied for determining the eggdiameter. The egg diameter was evaluated on an Olym-pus microscope with a graded rule. In the case of males,12 h post-injection, sperms were stripped and collectedwith capillary pipette and the expressive milt volumeswere determined on the basis of the amount of spermswithin the pipettes. Sperm volume was expressed inml. Density was determined within two hours of semencollection. Briefly, semen was diluted 1000 times with
ive impairment and endocrine disruption in goldfish by), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
0.7% NaCl; for determining spermatozoa concentration, ahaemocytometer was used and a drop (10 �l) of dilutedsemen was placed onto a haemocytometer (0.1 mm depthand 0.2 lengths) and left for 10 min to allow sperm

ARTICLE IN PRESSG Model
ANIREP-4711; No. of Pages 9
T. Bagheri et al. / Animal Reproduction Science xxx (2013) xxx– xxx 3
Table 1Proximate composition of the experimental diets as dried weight.
Experimental diets
FM SBM 35% SBM 65% SBM 100%
Ingredients (g kg)Fish meal 500 340 160 0Soybean meal 0 160 340 500Caseina 51 80 143 155Gelatina 70 80 70 80Starch 310 251 183 146Fish oil 40 60 75 90Methioninea 2 2 2 2Lysineb 2 2 2 2Vitamin + mineral premixc 20 20 20 20Antioxidantd 2.5 2.5 2.5 2.5Mold inhibitore 2.5 2.5 2.5 2.5Total 1000 1000 1000 1000
Proximate compositionGenistein (�g/g) 0 24.263 51.566 75.833Diadzein (�g/g) 0 21.702 46.126 67.821Protein (%) 40.3 40.37 40.23 40.46Fat (%) 8.03 9.56 9.06 9.12Carbohydrate (%) 40.15 40.64 40.4 42.64Energy (kJ/g) 18.4 18 17.86 17
a Sigma.b Aldrich.c Supplies per gram A: 5000 I.U., D3: 1000 I.U., E: 3 mg, B1: 2 mg, B2: 1 mg, B6: 0.3 mg, Nicotinamide: 15 mg, calcium pantothenate: 4 mg, K3: 1.5 mg,
C
slcsmNoepasaaFgmFocw
2
iptgmdto
U++: 3 mg, Fe++: 10 mg, Zn++: 15 mg, Mn++: 20 mg.d t-Butyl-4-hydroxyanisole, from Serva Co.e Sodium benzoate, from Sigma Co.
edimentation. Sperm concentration was expressed in bil-ions of sperms per ml. Spermatocrit was evaluated byentrifuging semen samples at 3000 rpm for 10 min Theperm motility was estimated through observing at × 200agnification using dark-field microscopy (Optiphot 2;ikon Inc., Garden City, NY, USA) from the initiation timef activation until complete cessation of movement. Invery stripping, sperm motility was recorded four timeser each male. The motility was expressed as the percent-ge of active sperm that has forward movement; vibratingperms were not counted as motile. For motility duration,s soon as sperms were activated, a stopwatch was startednd when spermatozoa movement ceased it was stopped.or fertilization rate, 5 females and 3 males from eachroup were used and after stripping and mixing eggs andilts, fertilization rate was estimated after 1 day (24оC).
or hatching rate evaluation, one hundred fertilized eggsf each treatment were randomly selected and separatelyultured in glass jars using fresh water and hatching rateas determined.
.5. Histological examination
The gonad tissues from females and males were fixedn Bouin’s solution, dehydrated in ethanol, embedded inaraffin wax, sectioned at 4 �m and stained with hema-oxylin and eosin (H&E). Because Carassius auratus is aroup-synchronous multispawning fish, the ovaries in
Please cite this article in press as: Bagheri, T., et al., Reproductfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013
ature females contain simultaneously oocyte at variousevelopmental stages. Oocyte development was staged inhis study according to the presence of the most advancedocytes present (Spanò et al., 2004). For each section
analyzed, 150 oocytes were scored and the proportion ofoocytes at each stage of development (0, Juvenile; I, pri-mary; II, pre-vitellogenic; III, vitellogenic; IV, mature; V,ovulated) calculated. Atretic oocytes were not measured inoocyte calculation but considered in the ovary analyses.
In males, three different histological sections from sixindividuals were analyzed and for each section, four dif-ferent areas of 1 mm2 were scored for sexual status andthe proportion of the section occupied by spermatogonia(including both spermatogonia a and b), spermatocytes,spermatids and spermatozoa was calculated.
2.6. Statistical analysis
All data are presented as means ± standard deviations.After testing data homogeneity, One-way analysis of vari-ance (ANOVA) followed by Duncan’s multiple range testwere used to determine significant differences betweengroups. The value P < 0.05 was used as the criterion forstatistical significance. All the tests were performed usingstatistics SPSS version 10.0.
3. Results
3.1. Plasma sex hormones
Among males and females, the small decline in plasmatestosterone levels was seen after inclusion of 35%, 65%
ive impairment and endocrine disruption in goldfish by), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
and 100% soybean meal in diet (Fig. 1A and B). In themales, the plasma E2 levels were increased after feedingincreasing concentrations of soybean meal, reaching signif-icance at 100% feeding rate (Fig. 1A). Similarly in females,

ARTICLE IN PRESSG Model
ANIREP-4711; No. of Pages 9
4 T. Bagheri et al. / Animal Reproduction Science xxx (2013) xxx– xxx
rone. (1 represeans at
Fig. 1. Plasma testosterone (T), estradiol (E2) and 17�, hydroxyprogestediets containing (35%, 65% and 100%) soybean meal for 28 weeks. Valuessuperscript letters indicate statistically significant differences between m
replacement of 100% of fish meal with soybean mealincreased the plasma E2 level (Fig. 1B). There were no sig-nificant differences in 17-OH-P levels after feeding dietscontaining soybean meal in either males or females (Fig. 1Aand B).
3.2. Histological examination
Feeding females with diets containing soybean mealsignificantly decreased the gonadosomatic index (GSI)in parallel to increasing soybean inclusion (Fig. 2).Histological examination showed that soybean mealinclusion significantly increased the percentages of pre-ovulatory (PreO) oocytes in a dose-dependent manner(Figs. 3, 9 and 10). Soybean inclusion in diets resulted
Please cite this article in press as: Bagheri, T., et al., Reproductfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013
in significantly increased previtellogenic (PreV) oocytesonly at the highest, 100% replacement rate, whereasthe percentage of vitellogenic (Vit) oocytes was signif-icantly increased after feeding with diets containing all
7-OH-P) levels in males (A) and females goldfish (B) after feeding withent the mean ± SD of three replicate tanks (nine fish per tank). DifferentP ≤ 0.05.
three levels of soybean meal. The percentage of pre-ovulatory oocytes (PreO) in female fish was significantlyand dose-dependently decreased after feeding diets con-taining all three levels of soybean meal (Fig. 3). Someatretic oocytes were also present after feeding withsoybean meal inclusion in diets (Figs. 11 and 12). Inmales, feeding with diets containing soybean meal sig-nificantly decreased the gonadosomatic index (GSI) inparallel to increasing soybean inclusion (Fig. 2). Therewas no significant difference in the percentage of sper-matids (ST) after feeding with experimental diets (Fig. 4),while a significant increase in the percentage of sper-matogonia (SG) and spermatocytes (SC) and a significantdecrease in the percentage of spermatozoa (SZ) wereobserved in parallel to increasing soybean inclusion
ive impairment and endocrine disruption in goldfish by), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
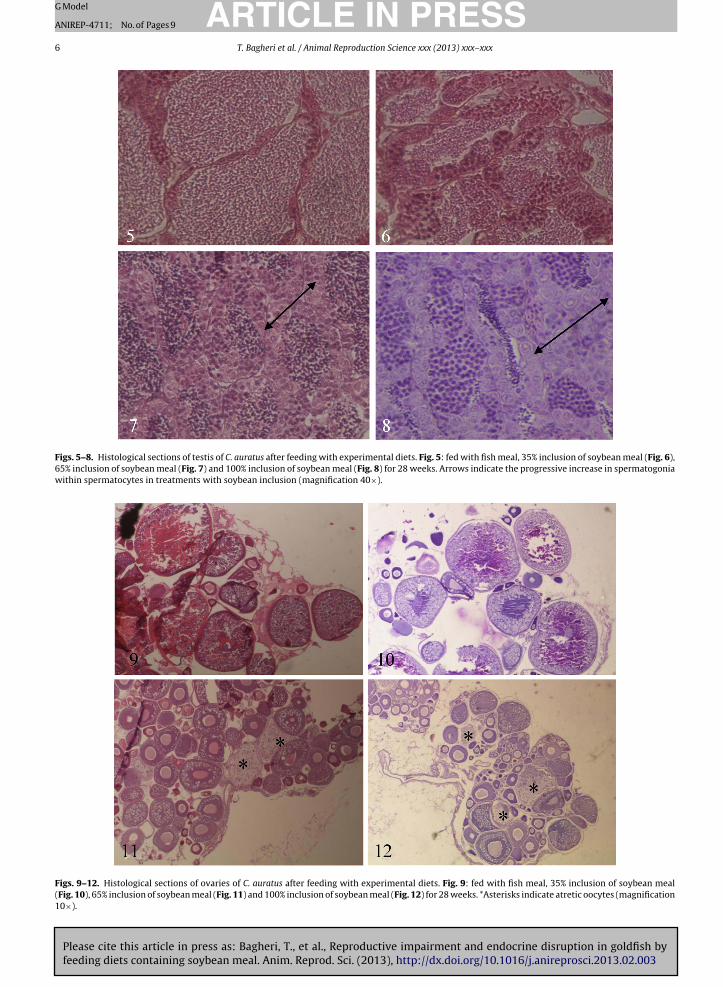
(Figs. 4 and 5–7). Histological observation also indicatesthe progressive increase in spermatogonia within sperma-tocytes in treatments with 65% and 100% soybean inclusion(Figs. 7 and 8).

Please cite this article in press as: Bagheri, T., et al., Reproductive impairment and endocrine disruption in goldfish byfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
ARTICLE IN PRESSG Model
ANIREP-4711; No. of Pages 9
T. Bagheri et al. / Animal Reproduction Science xxx (2013) xxx– xxx 5
Fig. 2. The gonadal–somatic index (GSI) in males and females of experimental diets. Values represent the mean ± SD of three replicate tanks (nine fish pertank). Different superscripts letters indicate statistically significant differences between means at P ≤ 0.05.
Fig. 3. The percentages of primary Oocyte (PO), previtellogenic oocytes (PreV), Vitellogenic (V) and Pre Ovulatory (PreO) in female goldfish. The goldfishwere fed with fish meal (FM), 35% inclusion of soybean meal (SBM35%), 65% inclusion of soybean meal (SBM65%) and 100% inclusion of soybean meal(SBM100%) for 28 weeks. Values represent the mean ± SD of six individual fish from three replicate tanks. Different superscript letters are significantlydifferent (P < 0.05).
Fig. 4. The percentages of spermatogonia (SG), spermatocytes (SC), spermatids (ST) and spermatozoa (SZ) in male goldfish. The goldfish were fed withfish meal (FM), 35% inclusion of soybean meal (SBM35%), 65% inclusion of soybean meal (SBM65%) and 100% inclusion of soybean meal (SBM100%) for 28weeks. Values represent the mean ± SD of six individual fish from three replicate tanks. Different superscript letters are significantly different (P < 0.05).
Please cite this article in press as: Bagheri, T., et al., Reproductive impairment and endocrine disruption in goldfish byfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
ARTICLE IN PRESSG Model
ANIREP-4711; No. of Pages 9
6 T. Bagheri et al. / Animal Reproduction Science xxx (2013) xxx– xxx
Figs. 5–8. Histological sections of testis of C. auratus after feeding with experimental diets. Fig. 5: fed with fish meal, 35% inclusion of soybean meal (Fig. 6),65% inclusion of soybean meal (Fig. 7) and 100% inclusion of soybean meal (Fig. 8) for 28 weeks. Arrows indicate the progressive increase in spermatogoniawithin spermatocytes in treatments with soybean inclusion (magnification 40×).
Figs. 9–12. Histological sections of ovaries of C. auratus after feeding with experimental diets. Fig. 9: fed with fish meal, 35% inclusion of soybean meal(Fig. 10), 65% inclusion of soybean meal (Fig. 11) and 100% inclusion of soybean meal (Fig. 12) for 28 weeks. *Asterisks indicate atretic oocytes (magnification10×).

ARTICLE IN PRESSG Model
ANIREP-4711; No. of Pages 9
T. Bagheri et al. / Animal Reproduction Science xxx (2013) xxx– xxx 7
Table 2Gamete parameters in goldfish fed experimental diets.
Sex Reproductive endpoints FM SBM 35% SBM 65% SBM 100%
Malea Spermatocrit (%) 42.6 ± 6.7a 41 ± 10.4ab 32.9 ± 7.5bc 29.4 ± 9.3c
Movement duration (s) 121.9 ± 0.8a 87.6 ± 1.1b 85.2 ± 1.2c 75.9 ± 3.7d
Spermatozoa motility (%) 81.9 ± 6.9a 80 ± 7.1a 70.6 ± 6.3b 55 ± 4.8 cSemen volume (ml/kg) 20.7 ± 19.2 10.7 ± 3.8 8.3 ± 1.8 4.8 ± 1.6Sperm density (×109/ml) 9 ± 2.3a 8 ± 1.7ab 5 ± 0.9b 4 ± 2.6 b
Femaleb Egg production (no. eggs/broodstock) 3289.78 ± 390.22a 3019.62 ± 844.37a 2628 ± 228a 1341.5 ± 8.5b
Egg diameter (mm) 1.50 ± 0.05 1.49 ± 0.05 1.47 ± 0.07 1.46 ± 0.03Hydrated egg diameter (mm) 1.64 ± 0.07 1.65 ± 0.07 1.64 ± 0.07 1.63 ± 0.04No. eggs/g body weight 202.81 ± 11.03a 204.98 ± 64.58a 163.61 ± 25.43a 74.12 ± 3.66b
V tly differ
3
wiieeifirmfwfcawt
4
tEpeeR
sshoefrb(igat
t
alues in the same row having different superscript letters are significana n = 3.b n = 5.
.3. Reproductive performances and hatching rates
During the feeding experiment period, survival ratesere higher than 90% in all groups. In female fish, feed-
ng with 100% soybean meal caused a significant decreasen cumulative egg production and number of eggs perach gram of broodstock body weight (Table 2); while theggs and hydrated eggs diameter were not significantlynfluenced by the experimental diets (Table 2). In malesh, soybean meal inclusion at 65% and 100% significantlyeduced sperm density, milt volume, sperm motility, spermovement duration and spermatocrit (Table 2). Similarly,
ertilization was significantly decreased by feeding fishith soybean inclusion at all concentrations. Over 89% of
ertilized eggs from the parental fed fish meal hatched suc-essfully at 2 dpf. In soybean meal fed groups, there was
significant decrease in hatching rates after parental fedith soybean inclusion of 65% and 100% compared with
he fish meal fed groups.
. Discussion
The present study revealed that feeding with diets con-aining soybean meal significantly disturbed plasma T and2 levels in both female and male goldfish, which sup-orted the findings that soybean phytoestrogens disruptndocrine homeostasis in previous studies in vitro (Ngt al., 2006) and in vivo (Bennetau-Pelissero et al., 2001;yokkynen et al., 2006).
One probable way of isofavonoids action is influencingex steroid-binding proteins in plasma, which could affectteroidogenesis regulation and the amounts of circulatingormones (Gatlin III et al., 2007). However, in a previ-us study in Arctic char competition in affinity of hepaticstrogen receptor (ER) for genistein with oestradiol wasour-fold lower (Tollefsen et al., 2004), and in human andainbow trout very weak binding affinities of sex steroidinding proteins for daidzein and genistein were shownMilligan et al., 1998). The probable mechanisms of earlynterference with the developing hypothalamo-pituitary-onad (HPG) axis in fish which may result in effects in the
Please cite this article in press as: Bagheri, T., et al., Reproductfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013
dult are not well understood, though it seems to be similaro other laboratory species (Maunder et al., 2007).
Histological examination demonstrated a reduction inhe pre-ovulatory oocytes stage in treatments which were
ent (P < 0.05).
fed with higher inclusion of soybean meal. This reduction isprobably attributed to the significant decrease in the GSI offish fed with diet high in soybean meal. However, a previousstudy with rainbow trout fed with genistein showed no sig-nificantly effect on GSI in the female (Bennetau-Pelisseroet al., 2001).
Although the impact of soybean phytoestrogens on fishreproduction remains largely ambiguous, our results showdecreased egg numbers in females as well as the decrease ofsperm density and production in males as a result of feedingwith soybean meal, whereas no changes on egg diame-ter have been shown. The diminished gamete quality andquantity could impair fish reproduction, thereby affectingspecies population (Au et al., 2001). It should be noted thatthe feeding study was in a long-term period, from larvae toadult, and feeding with soybean meal assessments on fishreproduction to the first spawning.
Since all nutritional requirements for embryonic devel-opment are supplied through fish eggs, a decreased eggsize of the soybean meal fed groups might have affectedthe nutritional content of the eggs, which may in turn,reduce hatching and reproductive success of goldfish,although in the present study no evidence of decreased eggsize was noted. High quality gametes from captive-rearedfish broodstock are critical for production of suitable off-spring for aquaculture (Kjorsvik et al., 1990; Bromage andRoberts, 1995). Not only good quality of eggs, but also thesperm quality of male broodfish is of great importance forproduction of healthy larvae. Therefore, if milt is inade-quate both in quantity and quality, successful fertilizationwill be impaired in the artificial insemination programs(Rurangwa et al., 2004).
One way to improve gamete quality and viable larvaeproduction is improving broodfish nutrition and feed-ing (Izquierdo et al., 2001). Several antinutrients existin plant proteins which could affect sexual developmentand reproduction in fish. Antinutrients such as gossypol, anaturally occurring compound in cotton seeds, when fedto male lamprey (Petromyzon marinus) result in reducedsperm motility (Rinchard et al., 2000) and lead to ele-vated testosterone (T) and 11- ketotestosterone (11-KT)
ive impairment and endocrine disruption in goldfish by), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
in rainbow trout (Dabrowski et al., 2000, 2001). Rain-bow trout fed with genistein, an isoflavone phytoestrogenfound in soybean meal, had weaker sperm motility andconcentration in a dose-dependent manner at spawning

ING Model
oduction
ARTICLEANIREP-4711; No. of Pages 9
8 T. Bagheri et al. / Animal Repr
(Bennetau-Pelissero et al., 2001). Previous studies ofestradiol (1 ng/L) during pre-spawning observed reducedseminal fluid volume, sperm density and fertilization andled to reduced sperm fertility after a 50-day exposure inmaturing rainbow trout (Lahnsteiner et al., 2006). Theseresults suggest that estrogens could affect male reproduc-tive success in fish.
Positive correlations have been found between fertil-izing capacity and defined sperm motility characteristicsin fish (Rurangwa et al., 2004), so it is not surprising thatthe fish in soybean meal fed groups in the present studyshowed a significant reduction in fertility success and lar-vae production.
In the present study some of the biomarkers of spermquality were assessed that directly relate to the fertil-ization capacity of sperm, including spermatocrit, spermdensity, motility, semen volume and movement duration(Rurangwa et al., 2004). All of the mentioned biomarkerswere significantly influenced by experimental diets in themale fish. Reduced assessed sperm quality biomarkerswere observed along with the markedly increased plasmaE2 level. The reduction in sperm quality biomarkers mightbe due to partial disruption in spermatogenesis process.The previous study in channel catfish Ictalurus punctatus(Rafinesque) and walleye Sander vitreus (Mitchill) founda significant negative relationship between genisteinincubation concentration and inactivated spermatozoaATP content in both species (Green and Kelly, 2008). It wasclaimed that genistein is a specific inhibitor of tyrosineprotein kinas due to its ability to bind with ATP (Akiyamaet al., 1987) and reduce available ATP within inactivatedspermatozoa from in vitro incubations. Since ATP contenthas been positively correlated with variables such asmotility time and spermatozoa swimming velocity withinfishes and mammals (Jeulin & Soufir, 1992; Burness et al.,2004), the reduction in sperm motility in soybean mealfed groups might be related to this attribute of soybeanisoflavone phytoestrogens.
Histo-morphological examination of the male gold-fish gonads revealed that spermatogenesis was partiallyinhibited in our study.
These results obviously manifest that the reproduc-tive performance of fish can be adversely influenced byfeeding with food ingredients containing estrogen-likesubstances during a long-term developmental period, frompost hatch to the mature. Furthermore, even if no appar-ent morphological indications of endocrine disruptionin populations exist (e.g., ovotestis absent, normal sexratio) they may still demonstrate abnormal reproductivebiology.
Previous studies on the effects of soybean isoflavonesin fish have indicated great obvious responses, depend-ing upon the dose and the species. In yellow perch, Percaflavescens, induced vitellogenin (egg yolk protein) synthe-sis of liver and a growth promoting effect similar to that ofestradiol-17� (Malison et al., 1985, 1988) with genistein,one of the most abundant isoflavones found in soybeans,
Please cite this article in press as: Bagheri, T., et al., Reproductfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013
has been reported (Ko et al., 1999). Soybean-based dietsalso increased plasma vitellogenin concentrations in maleand female rainbow trout (Pelissero et al., 2001), Siberiansturgeon, Acipenser baeri (Pelissero et al., 1991a,b), and
PRESS Science xxx (2013) xxx– xxx
juvenile striped bass, Morone saxatilis (Pollack and Ottinger,2003).
In conclusion, regarding the disruptive action of phy-toestrons on reproduction, it is better to limit the use ofsoybean meal in diets of fishes which are to be kept as abroodfish.
Acknowledgements
The first author is grateful to the Scientific Boards andstaff of the Aquaculture Department and the FisheriesResearch Center of Gorgan University of Agricultural Sci-ences and Natural Resource for technical and financialsupport. I am also thankful to Fardin Shaluei for his assis-tance in reproduction trials.
References
Akiyama, T., Ishida, J., Nakagawara, S., Ogawara, H., Watanabe, S., Itoh, N.,Shibuya, M., Fukami, Y., 1987. Genistein, a specific inhibitor of tyrosinespecific protein kinases. J. Biol. Chem. 262, 5592–5595.
AOAC, 1990. In: Helrich, W. (Ed.), Official Methods of Analysis of the Asso-ciation of Official Analytical Chemists, vol. 1, 15th ed. Association ofOfficial Analytical Chemists, Washington, DC.
Au, D.W.T, Lee, C.Y., Chan, K.L., Wu, R.S.S., 2001. Reproductive impairmentof sea urchins upon chronic exposure to cadmium. Part I: effects ongamete quality. Environ. Pollut. 111, 1–9.
Barrett, J., 1996. Phytoestrogens: friends or foes? Environ. Health Perspect.104, 478–482.
Bennetau-Pelissero, C., Breton, B., Bennetau, B., Corraze, G., LeMenn, F.,Davail-Cuisset, B., Helou, C., Kaushik, S.J., 2001. Effect of genistein-enriched diets on the endocrine process of gametogenesis and onreproduction effciency of the rainbow trout (Oncorhynchus mykiss).Gen. Comp. Endocr. 121, 173–187.
Bennetau-Pelissero, C., Arnal-Schnebelen, B., Lamothe, V., Sauvant, P.,Sagne, J.L., Verbruggen, M.A., Mathey, J., Lavialle, O., 2003. ELISA asa new method to measure genistein and daidzein in food and humanfluids. Food Chem. 82, 645–658.
Blazquez, M., Bosma, P.T., Fraser, E.J., Van Look, K., Trudeau, V.L., 1998. Fishas models for neuroendocrine regulation of growth and reproduction.Comp. Biochem. Physiol. C 119, 339–364.
Bromage, N.R., Roberts, R.J. (Eds.), 1995. Broodstock Management and Eggand Larval Quality. Blackwell Science Ltd., Oxford, p. 424.
Burness, G., Casselman, S.J., Schulte-Hostedde, A.I., Moyes, C.D., Mont-gomerie, R., 2004. Sperm swimming speed and energetics vary withsperm competition risk in bluegill (Lepomis macrochirus). Behav. Ecol.Sociobiol. 56, 65–70.
Carter, C.G., Hauler, R.C., 2000. Fish meal replacement by plant mealsin extruded feed for Atlantic salmon, Salmo salar L. Aquaculture 185,299–311.
Mateos-Aparicio, I., Redondo Cuenca, A., Villanueva-Suárez, M.J., Zapata-Revilla, M.A., 2008. Soybean, a promising health source. Nutr. Hosp.23, 305–312.
Dabrowski, K., Lee, K.J., Rinchard, J., Blom, J.H., Ciereszko, A., Otto-bre, J., 2000. Effects of diets containing gossypol on reproductivecapacity of rainbow trout (Oncorhynchus mykiss). Biol. Reprod. 62,227–234.
Dabrowski, K., Rinchard, J., Lee, K.J., Ciereszko, A., Blom, J.H., Ottobre, J.,2001. Gossypol isomers bind specifically to blood plasma proteinsand spermatozoa of rainbow trout fed diets containing cotton seedmeal. Biochim. Biophys. Acta 1525, 37–42.
Dixon, R.A., 2004. Phytoestrogens. Annu. Rev. Plant Biol. 55, 225–261.Kuiper, G.G.J.M., Carlsson, B., Grandien, K., Enmark, E., Hågg-blad, J., Nils-
son, S., Gustafsson, J.Å., 1997. Comparison of the ligand bindingspecificity and transcript tissue distribution of estrogen receptorsalpha and beta. Endocrinology 138, 863–870.
Friendman, M., Baron, D.L., 2001. Nutritional and health benefits of soyproteins. J. Agric. Food Chem. 49, 1069–1086.
ive impairment and endocrine disruption in goldfish by), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
Gatlin III, D.M., Barrows, F., Brown, T., Dabrowski, P., Gaylord, K., Hardy,T.G., Herman, R.W., Hu, E., Krogdahl, G., Nelson, A., Overturf, R., Rust,K., Sealey, M., Skonberg, W., Souza, D., Stone, E.J., Wilson, D., Wurtele,R.E., 2007. Expanding the utilization of sustainable plant products inaquafeeds: a review. Aquac. Res. 38, 551–579.

ING Model
A
oduction
G
I
I
J
K
K
K
L
L
M
M
M
M
M
ARTICLENIREP-4711; No. of Pages 9
T. Bagheri et al. / Animal Repr
reen, C.C., Kelly, A.M., 2008. Effect of the exogenous soyabean phyto-oestrogen genistein on sperm quality, ATP content and fertilizationrates in channel catfish Ictalurus punctatus (Rafinesque) and walleyeSander vitreus (Mitchill). J. Fish Biol. 72, 2485–2499.
morou Toko, I., Fiogbe, D.E., Kestemont, P., 2008. Mineral status of Africancatfish (Clarias gariepinus) fed diets containing graded levels of soy-bean or cottonseed meals. Aquaculture 275, 298–305.
zquierdo, M.S., Fernandez-Palacios, H., Tacon, A.G.J., 2001. Effect of brood-stock nutrition on reproductive performance of fish. Aquaculture 197,25–42.
eulin, C., Soufir, J.-C., 1992. Reversible intracellular ATP changes in intactrat spermatozoa and effects on flagellar sperm movement. Cell Motil.Cytoskeleton 21, 210–222.
jorsvik, E., Mangor-Jensen, A., Holmefjord, I., 1990. Egg quality in fishes.In: Blaxter, J.H.S., Southward, A.J. (Eds.), Advance in Marine Biology,26, pp. 71–113.
uiper, G.G., Lemmen, J.G., Carlsson, B., Corton, J.C., Safe, S.H., van derSaag, P.T., van der Burg, B., Gustafsson, J.A., 1998. Interaction ofestrogenic chemicals and phytoestrogens with estrogen receptor beta.Endocrinology 139, 4252–4263.
o, K., Malison, J.A., Reed, J.R., 1999. Effect of genistein on the growthand reproductive function of male and female yellow perch Percaflavescens. J. World Aquacult. Soc. 30, 73–78.
ahnsteiner, F., Berger, B., Kletzl, M., Weismann, T., 2006. Effect of 17@b-estradiol on gamete quality and maturation in two salmonid species.Aquat. Toxicol. 79, 124–131.
atonnelle, K., Fostier, A., LeMenn, F.E., Bennetau-Pelissero, C., 2002. Bind-ing affinities of hepatic nuclear estrogen receptors for phytoestrogensin rainbow trout and Siberian sturgeon. Gen. Comp. Endocrinol. 129,69–79.
alison, J.A., Best, C.D., Kayes, T.B., Amundson, C.H., Wentworth, B.C.,1985. Hormonal growth promotion and evidence for a sizerelated dif-ference in response to estradiol-17� in yellow perch (Perca flavescens).Can. J. Fish Aquat. Sci. 42, 1627–1633.
alison, J.A., Kayes, T.B., Wentworth, B.C., Amundson, C.H., 1988. Growthand feeding responses of male versus female yellow perch (Percaflavescens) treated with estradiol-17�. Can. J. Fish Aquat. Sci. 45,1942–1948.
aunder, R.J., Matthiessen, P., Sumpter, J.P., Pottinger, T.G., 2007.Impaired reproduction in three-spined sticklebacks exposed to
ethinyl estradiol as juveniles. Biol. Reprod. 77, 999–1006.illigan, S.R., Khan, O., Nash, M., 1998. Competitive binding of xenobiotic
oestrogens to rat alpha-fetoprotein and to sex steroid binding pro-teins in human and rainbow trout (Oncorhynchus mykiss) plasma. Gen.
Please cite this article in press as: Bagheri, T., et al., Reproductfeeding diets containing soybean meal. Anim. Reprod. Sci. (2013
Comp. Endocrinol. 112, 89–95.imeault, C., Woodhouse, A.J., Miaob, X.-S., Metcalfe, C.D., Moona, T.W.,
rudeau, V.L., 2005. The human lipid regulator, gemfibrozil biocon-centrates and reduces testosterone in the goldfish, Carassius auratus.Aquat. Toxicol. 73, 44–54.
PRESS Science xxx (2013) xxx– xxx 9
Ng, Y., Hanson, S., Malison, J.A., Wentworth, B., Barry, T.P., 2006. Genisteinand other isoflavones found in soybeans inhibit estrogen metabolismin salmonid fish. Aquaculture 254, 658–665.
Pelissero, C., Le Menn, F., Kaushik, S., 1991a. Estrogenic effect of dietarysoya bean meal on vitellogenesis in cultured Siberian sturgeonAcipenser baeri. General Comparative Endocrinology 83, 447–457.
Pelissero, C., Bennetau, B., Babin, P., Le Menn, F., Dunogues, J., 1991b.Estrogenic activity of certain phytoestrogens in the Siberian sturgeon(Acipenser baeri). J. Steroid Biochem. 38, 293–299.
Pollack, S.J., Ottinger, M.A., 2003. The effects of the soy isoflavone genis-tein on the reproductive development of striped bass. North AmericanJournal of Aquaculture 65, 226–234.
Rinchard, J., Ciereszko, A., Dabrowski, K., Ottobre, J., 2000. Effects of gossy-pol on sperm viability and plasma sex steroid hormones in male sealamprey, Petromyzon marinus. Toxicol. Lett. 111, 189–198.
Rurangwa, E., Kime, D.E., Ollevier, F., Nash, J.P., 2004. The measurementof sperm motility and factors affecting sperm quality in cultured fish:review article. Aquaculture. 234, 1–28.
Ryokkynen, A., Kukkonen, J.V.K., Nieminen, P., 2006. Effects of dietarygenistein on mouse reproduction, postnatal development and weight-regulation. Anim. Reprod. Sci. 93, 337–348.
Spanò, L., Tyler, C.R., Aerle, R.V., Devos, P., Mandiki, S.N.M., Silvestre,F., Thomé, J.P., Kestemont, P., 2004. Effects of atrazine on sexsteroid dynamics, plasma vitellogenin concentration and gonaddevelopment in adult goldfish, Carassius auratus. Aquat. Toxicol. 6,369–379.
Tollefson, K.E., Mathisen, R., Stenersen, J., 2001. Estrogen mimics bindwith similar affinity and specificity to the hepatic estrogen receptorin Atlantic salmon and rainbow trout. Gen. Comp. Endocr. 126, 14–22.
Tollefsen, K.E., Ovrevik, J., Stenersen, J., 2004. Binding of xenoestrogens tothe sex steroid-binding protein in plasma from Arctic charr (Salvelinusalpinus L.). Comp. Biochem. Phys. 139 (C), 127–133.
Trudeau, V.L., 1997. Neuroendocrine regulation of gonadotrophin-IIrelease and gonadal growth in the goldfish, Carassius auratus. Rev.Reprod. 2, 55–68.
Van den Ingh, T.S.G.A.M., Krogdahl, A., 1990. Negative effects of antinutri-tional factors from soya beans in salmonids. Tijdschr. Diergeneeskd.115, 935–938.
Woclawek-Potocka, I., Bah, M.M., Korzekwa, A., Piskula, M.K., Wizkowski,E., Depta, A., Skarzynski, D.J., 2005. Soybean derived phytoestro-gens regulate prostaglandin secretion in endometrium during cattleestrous cycle and early pregnancy. Exp. Biol. Med. 230, 189–199.
Yanar, M., Erc en, Z., Hunt, A.Ö., Büyükc apar, H.M., 2008. The use of alfalfa,Medicago sativa as a natural carotenoid source in diets of goldfish,
ive impairment and endocrine disruption in goldfish by), http://dx.doi.org/10.1016/j.anireprosci.2013.02.003
Carassius auratus. Aquaculture 284, 196–200.Yue, Y.R., Zhou, Q.C., 2008. Effect of replacing soybean meal with cotton-
seed meal on growth, feed utilization, and hematological indexes forjuvenile hybrid tilapia, Oreochromis niloticus × O. aureus. Aquaculture284, 185–189.