gene therapy (1997) 4 Ó ex vivo adenovirus-mediated gene
TRANSCRIPT

Gene Therapy (1997) 4, 639–647 1997 Stockton Press All rights reserved 0969-7128/97 $12.00
Ex vivo adenovirus-mediated gene transfer andimmunomodulatory protein production in humancornea
HB Oral1, DFP Larkin1,2, Z Fehervari1,3, AP Byrnes4,5, AM Rankin6, DO Haskard7, MJA Wood4,MJ Dallman6 and AJT George1
1Department of Immunology, Royal Postgraduate Medical School, Hammersmith Hospital, London; 2Department of Pathology,Institute of Ophthalmology, London; 4Department of Human Anatomy, University of Oxford; 6Infection and Immunity Section,Department of Biology, Imperial College of Science, Technology and Medicine, London; 7Department of Medicine, CardiovascularMedicine, Royal Postgraduate Medical School, Hammersmith Hospital, London, UK
One attractive strategy to prevent or control allograft rejec- transcription after infection. In contrast, adenoviral DNAtion is to genetically modify the donor tissue before trans- persisted for at least 56 days. Subsequently, we examinedplantation. In this study, we have examined the feasibility the expression of a potential therapeutic gene, CTLA-4 Igof gene transfer to human corneal endothelium, using a fusion protein. Following infection of human corneas withnumber of recombinant adenovirus constructs. Ex vivo adenoviral vectors encoding CTLA-4 Ig protein, high levelsinfection of human corneas with adenoviral vectors con- of the fusion protein were detected in corneal culturetaining lacZ, under transcriptional control of either cyto- supernatants for up to 28 days. This protein was func-megalovirus (CMV) or Rous sarcoma virus (RSV) pro- tionally active, as determined by binding to B7.1 (CD80)-moters, provided high-level gene expression, which was expressing transfectants. This study suggests that geneticlargely restricted to endothelium. Expression of the alteration of donor cornea before transplantation is a feas-reporter gene persisted at relatively high levels for up to 7 ible approach for preventing or controlling allograft rejec-days, followed by a decline to indetectable levels by 28 tion. Similar gene-based strategies might also be feasibledays. RT-PCR analysis of lacZ transcription showed a to prevent rejection of other transplanted tissues or organs.similar picture with a short period (3–7 days) of RNA
Keywords: transplantation; CTLA-4 Ig; b-galactosidase; RT-PCR; endothelium
easy accessibility and critical importance in maintenanceIntroductionof corneal transparency, which itself allows direct obser-
Gene therapy offers the exciting possibility of expressing vation of the effects of gene transfer. In addition, modifi-genes in vivo with therapeutic potential in target cells. cation of endothelial cell gene expression with the objec-However, targeting novel genes to specific organs in vivo tive of local modulation of immune response or alterationis problematic. One of the strategies to achieve efficient of cellular physiology could have widespread clinicaltherapeutic gene expression is ex vivo genetic modifi- application, as the negligible replication capacity of cor-cation of the target cells or organs, followed by their neal endothelium is insufficient to compensate for endo-transplantation. Cornea is a particularly suitable candi- thelial damage which may follow surgical trauma,date tissue for such gene-based approaches, because inflammation (such as allograft rejection) or degenerativeunlike other transplanted tissues, it can be maintained in disease. At present, the only available treatment for suchstandard culture conditions for periods of up to 1 month, corneal injury is replacement of the cornea with an allo-allowing time for its genetic alteration before transplan- graft. Primary endothelial disease is the indication fortation. The corneal endothelium has distinct advantages approximately 50% of corneal transplants,1,2 and theas a target for gene therapy. It is a cell monolayer on the endothelium is itself the critical target of allograft rejec-internal surface of the cornea with well-defined anatomy, tion. In addition, short-term and chronic endothelial cell
loss following transplantation, in the absence of overtrejection, is a frequent cause of corneal graft failure.3
We have previously examined gene transfer to wholeCorrespondence: AJT George, Department of Immunology, RPMS, Ham-mersmith Hospital, Du Cane Road, London W12 0NN, UK rabbit corneas using a recombinant adenovirus vectorCurrent addresses: 3Department of Immunology, Glaxo Wellcome Medi- carrying the lacZ gene driven by a human cytomegalo-cines Research Centre, Gunnels Wood Road, Stevenage SG1 2NY, UK; virus (CMV) promoter. In these studies, rabbit corneas5Department of Molecular Microbiology and Immunology, Johns Hopkins
were incubated with adenovirus ex vivo, and theUniversity, School of Public Health, 615 N Wolfe Street, Baltimore, MDefficiency and time course of gene expression following21205-2179, USA
Received 8 January 1997; accepted 7 March 1997 periods of corneal culture were investigated.4 Transgene

Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
640 expression was comparatively restricted to the corneal adenoviral infection is similar to that seen in the rabbitand rat corneas.4,5endothelium with expression in almost 100% of cells at
day 1, diminishing to expression in 10% by day 14. Usingthe same experimental conditions, it has been found that Kinetics of b-gal expression
Using a b-gal colorimetric assay, we examined the timethe lacZ transgene was expressed in rat corneas in up to50% of endothelial cells for 21 days ex vivo,5 suggesting course of the recombinant protein expression in corneal
samples, infected by either AdRL or AdCB2. In thesea disparity in expression between the species.If techniques of ex vivo gene transfer are to be of clinical experiments, b-galactosidase expression was assayed fol-
lowing 1, 3, 7, 14, 21 and 28 days of ex vivo culture. Fol-utility, several key issues require resolution. Studiesusing animal tissues may not accurately predict the effi- lowing infection with AdCB2, highest expression was
seen at day 7, whilst AdRL yielded maximal b-galcacy and safety of gene transfer to human tissues. Onemore fundamental problem is the loss of transgene activity at day 3 (Figure 2a and b). b-Gal activity was not
detectable 28 days after infection with either virus. Theexpression, which has characterised E1-deleted adeno-viruses and other gene delivery systems such as lipofec- RSV promoter-driven expression was quantitatively
lower at all time-points, other than day 3. Control, mock-tion and receptor–DNA conjugate mediated gene deliv-ery.6 Here we describe efficient marker gene expression infected, corneas did not show any b-gal activity. The
variability between samples was greater than that pre-in the endothelium of human corneas, with high levelsof b-gal production for 7 days following ex vivo infection viously reported for rabbit or rat corneas.4,5 This may be
due to the heterogeneity of the human tissue samples,with recombinant adenoviruses under transcriptionalcontrol of either Rous sarcoma virus (RSV) or CMV pro- particularly the differences in the number of endothelial
cells present.moters. It has long been recognised that T cells, bothCD4+ and CD8+, play a critical role in rejection of allo-grafts.7–9 One of the approaches to prevent graft rejection Reverse transcriptase PCR for RNA analysis
Analysis of the lacZ gene transcription was performedis to prevent T cell activation as a result of interactionwith graft antigens. Antigen-specific T cell activation using RT-PCR of corneal samples previously infected
with either AdCB2 or AdRL. RNA was extracted fromrequires not only recognition of TCR by the MHC mol-ecules, but also secondary stimulation provided by other these samples at various times after infection (from day
1 to day 28). Following infection with AdRL, the 1036 bpreceptor–ligand interactions.10 One of the costimulatorypathways is the interaction of B7.1 (CD80) and B7.2 sequence of the lacZ gene was amplified from the
samples at day 1, 3 and 7 by RT-PCR (Figure 3). How-(CD86) molecules on antigen presenting cells with theCD28 molecule on T cells.11,12 The inhibition of CD28 ever, RT-PCR analysis performed in AdCB2-infected cor-
nea samples revealed that transcription of lacZ gene wascostimulation has been shown to cause T cell anergy andhyporesponsiveness.13–15 CTLA-4 is an alternative recep- not detectable after day 3 after 29 cycles of RT-PCR
(Figure 3). No such band was seen in cDNA preparedtor on T cells for B7.1 and B7.2, with high homology toCD28.16,17 In comparison to CD28, it binds B7.1 with a 20- from cornea segments at later time-points and from unin-
fected cornea. In addition, direct PCR of the RNAfold higher avidity.12 Furthermore, CTLA-4 Ig has beenshown to extend graft survival in a number of rat and samples did not lead to appearance of the 1036 bp band,
suggesting that DNA contamination was not responsiblemouse transplant models.18–20 This indicates the possi-bility of gene-based immunomodulation of the allogeneic for the results seen (data not shown).response to a corneal allograft by ex vivo virus-mediatedgene transfer to the donor cornea before transplantation. Persistence of adenoviral DNA and the lacZ gene in
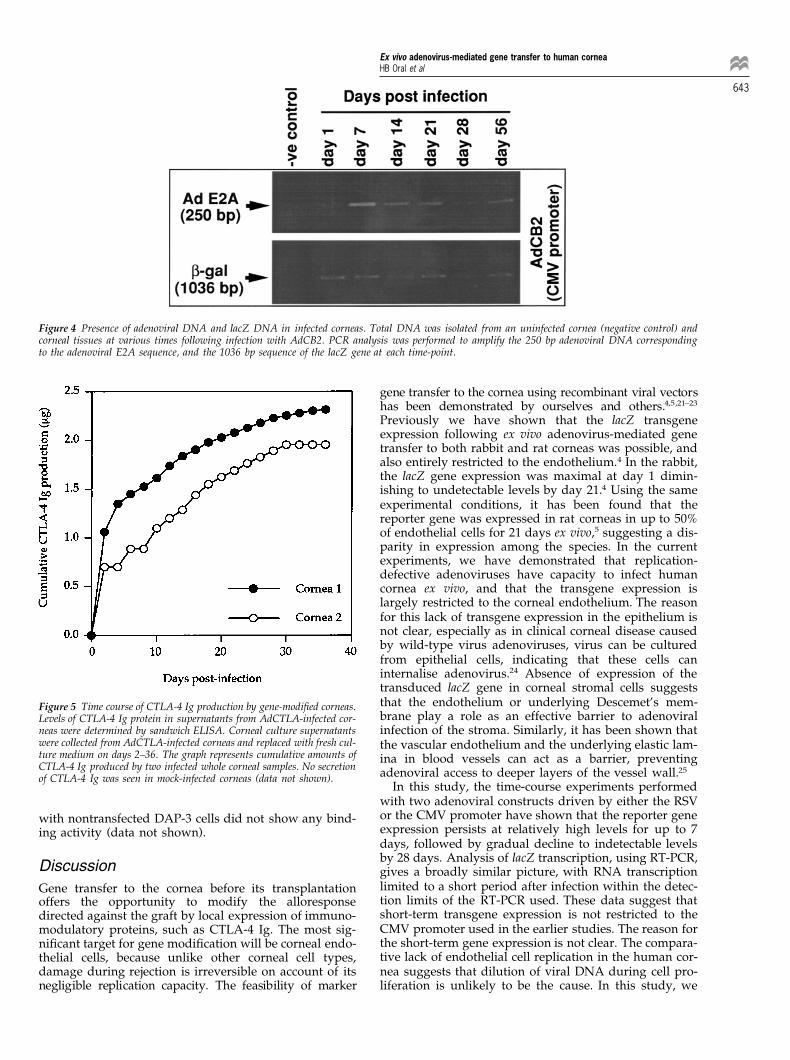
gene-modified human corneasTo determine the presence of adenoviral DNA and the lacZ
Results gene, PCR analysis was performed on the DNA samplesextracted from AdCB2 and mock-infected cornea speci-mens at various time-points, from 1 to 56 days, followingEfficiency and localisation of lacZ expression in corneas
In preliminary experiments conditions were optimised infection. The 250 bp band corresponding to the adenoviralE2A sequence was seen at all time-points. The PCR analysisfor infecting human corneas. Incubation of corneal
samples with various concentrations of the AdCB2 or the amplifying the 1036 bp internal sequence of the lacZ genealso revealed the presence of the reporter gene up to dayAdRL virus constructs allowed us to determine the opti-
mal conditions for efficient gene transfer. Maximal gene 56. No such band was demonstrable in DNA extractedfrom mock-infected corneal specimens (Figure 4). Theseexpression, with 90–100% of endothelial cells expressing
b-gal, was obtained with 4 × 108 p.f.u. per cornea of the results indicate that adenoviral DNA remains present forlong periods after expression of the gene has terminated.AdRL, whereas 1.5 × 108 p.f.u. per cornea of the AdCB2
provided a maximal transgene expression (data not Similar results were obtained with AdRL vector, with DNAseen up to 56 days (data not shown).shown). These viral numbers were used for all further
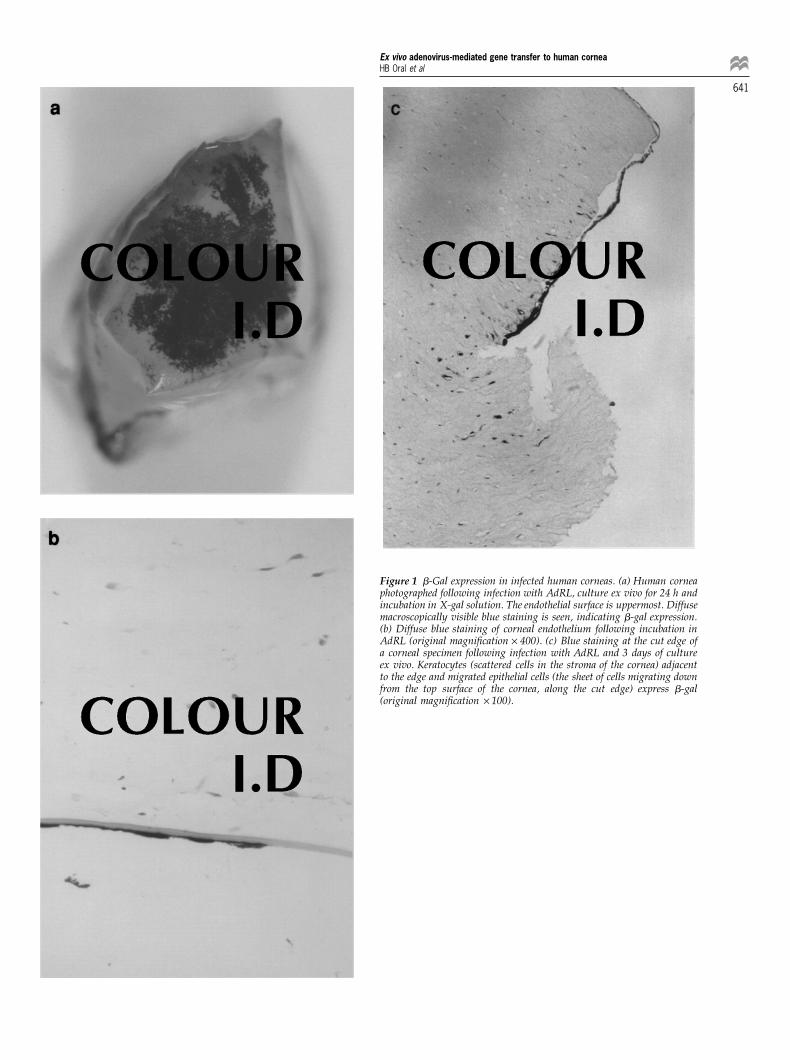
experiments. b-Gal expression was almost entirely local-ised in the corneal endothelium following infection with Time course of CTLA-4 Ig secretion by gene-modified
corneaseither AdRL or AdCB2 (Figure 1a and b). However, someblue staining was observed in both keratocytes and epi- CTLA-4 Ig protein secretion from two whole cornea
samples infected with AdCTLA was determined by sand-thelial cells, especially at the cut edge of the specimens ashas been seen in the rabbit and rat (Figure 1c).4,5 Control, wich ELISA. Both corneas were washed three times after
infection to prevent the possibility of carryover of CTLA-nontransduced corneas did not stain positively with X-gal (data not shown). These results indicate that the gen- 4 Ig from the purified viral preparation. The corneas
showed similar kinetics for the production of CTLA-4 Igeral expression pattern of b-galactosidase following
Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
641
Figure 1 b-Gal expression in infected human corneas. (a) Human corneaphotographed following infection with AdRL, culture ex vivo for 24 h andincubation in X-gal solution. The endothelial surface is uppermost. Diffusemacroscopically visible blue staining is seen, indicating b-gal expression.(b) Diffuse blue staining of corneal endothelium following incubation inAdRL (original magnification × 400). (c) Blue staining at the cut edge ofa corneal specimen following infection with AdRL and 3 days of cultureex vivo. Keratocytes (scattered cells in the stroma of the cornea) adjacentto the edge and migrated epithelial cells (the sheet of cells migrating downfrom the top surface of the cornea, along the cut edge) express b-gal(original magnification × 100).

Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
642
Figure 2 Persistence of b-galactosidase expression in two sets of human cornea samples maintained ex vivo following infection with AdCB2 (a) orAdRL (b). b-Gal activity in cornea lysates was determined by soluble b-galactosidase assay at the indicated days after infection. Activity is expressedas milliunits (mU) per 20 mg of total protein in the cornea extracts. Detection threshold of the assay is 5 mU/20 mg.
Figure 3 RT-PCR analysis of lacZ gene transcription. Total RNA was prepared from an uninfected cornea sample (negative control) and corneal samplesat various times after AdRL or AdCB2 infection. RT-PCR analysis was carried out to amplify the 1056 bp sequence of the lacZ gene and the 540 bpsequence of b-actin gene. Direct PCR product from pCMV/b-gal vector was used as a positive control for the lacZ gene.
fusion protein. Following high-level initial protein pro- its ability to recognise its specific ligand B7.1 (CD80) byflow cytometry. The flow cytometry profiles in Figure 6cduction, the secretion of the CTLA-4 Ig fusion protein
into culture supernatants was continued over 28 days demonstrate specific binding of the fusion protein foundin the supernatant of AdCTLA-infected corneas to B7.1(Figure 5). The cumulative CTLA-4 Ig production reached
approximately 2 mg over 28 days. These data demon- (CD80) transfected DAP-3 cells, in a similar manner tothat seen with the purified CTLA-4 Ig fusion proteinstrate that adenovirus-mediated gene delivery
approaches are feasible for expression of therapeutic (Figure 6d). The small peak at around 10 fluorescent unitswas seen in both samples, and is presumably due togenes in corneal endothelium.lower expression of the transfected gene in the same cells(Figure 6c and d). No binding appeared with either ofFlow cytometric analysis of CTLA-4 Ig binding
The binding activity of the CTLA-4 Ig fusion protein, the negative controls, PBS or human IgG1 (Figure 6a andb, respectively). In addition, incubation of the sampleswhich blocks T cell costimulation, was tested by assessing
Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
643
Figure 4 Presence of adenoviral DNA and lacZ DNA in infected corneas. Total DNA was isolated from an uninfected cornea (negative control) andcorneal tissues at various times following infection with AdCB2. PCR analysis was performed to amplify the 250 bp adenoviral DNA correspondingto the adenoviral E2A sequence, and the 1036 bp sequence of the lacZ gene at each time-point.
gene transfer to the cornea using recombinant viral vectorshas been demonstrated by ourselves and others.4,5,21–23
Previously we have shown that the lacZ transgeneexpression following ex vivo adenovirus-mediated genetransfer to both rabbit and rat corneas was possible, andalso entirely restricted to the endothelium.4 In the rabbit,the lacZ gene expression was maximal at day 1 dimin-ishing to undetectable levels by day 21.4 Using the sameexperimental conditions, it has been found that thereporter gene was expressed in rat corneas in up to 50%of endothelial cells for 21 days ex vivo,5 suggesting a dis-parity in expression among the species. In the currentexperiments, we have demonstrated that replication-defective adenoviruses have capacity to infect humancornea ex vivo, and that the transgene expression islargely restricted to the corneal endothelium. The reasonfor this lack of transgene expression in the epithelium isnot clear, especially as in clinical corneal disease causedby wild-type virus adenoviruses, virus can be culturedfrom epithelial cells, indicating that these cells caninternalise adenovirus.24 Absence of expression of thetransduced lacZ gene in corneal stromal cells suggeststhat the endothelium or underlying Descemet’s mem-Figure 5 Time course of CTLA-4 Ig production by gene-modified corneas.brane play a role as an effective barrier to adenoviralLevels of CTLA-4 Ig protein in supernatants from AdCTLA-infected cor-
neas were determined by sandwich ELISA. Corneal culture supernatants infection of the stroma. Similarly, it has been shown thatwere collected from AdCTLA-infected corneas and replaced with fresh cul- the vascular endothelium and the underlying elastic lam-ture medium on days 2–36. The graph represents cumulative amounts of ina in blood vessels can act as a barrier, preventingCTLA-4 Ig produced by two infected whole corneal samples. No secretion adenoviral access to deeper layers of the vessel wall.25of CTLA-4 Ig was seen in mock-infected corneas (data not shown).
In this study, the time-course experiments performedwith two adenoviral constructs driven by either the RSVor the CMV promoter have shown that the reporter genewith nontransfected DAP-3 cells did not show any bind-expression persists at relatively high levels for up to 7ing activity (data not shown).days, followed by gradual decline to indetectable levelsby 28 days. Analysis of lacZ transcription, using RT-PCR,Discussion gives a broadly similar picture, with RNA transcriptionlimited to a short period after infection within the detec-Gene transfer to the cornea before its transplantationtion limits of the RT-PCR used. These data suggest thatoffers the opportunity to modify the alloresponseshort-term transgene expression is not restricted to thedirected against the graft by local expression of immuno-CMV promoter used in the earlier studies. The reason formodulatory proteins, such as CTLA-4 Ig. The most sig-the short-term gene expression is not clear. The compara-nificant target for gene modification will be corneal endo-tive lack of endothelial cell replication in the human cor-thelial cells, because unlike other corneal cell types,nea suggests that dilution of viral DNA during cell pro-damage during rejection is irreversible on account of its
negligible replication capacity. The feasibility of marker liferation is unlikely to be the cause. In this study, we

Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
644
Figure 6 Demonstration of the binding activity of CTLA-4 Ig protein using flow cytometry. The culture supernatant from AdCTLA-infected humancornea 1 day after infection (c) was removed and incubated with B7.1 (CD80) transfected DAP-3 fibroblasts (DAP3-B7.1). The binding activity wasdetermined by flow cytometry following incubation with FITC-conjugated mouse anti-human IgG1. Purified CTLA-4 Ig protein (d) was employed as apositive control, whilst PBS (a) and a human IgG1 standard (b) were used as negative controls.
demonstrated that adenoviral DNA existed for at least 56 shown that functionally active CTLA-4 Ig can be secretedby human cornea which has been transduced ex vivo withdays after infection, suggesting that loss of viral DNA is
not responsible for the short-term gene expression. How- a recombinant adenovirus vector containing the geneencoding this protein.ever it is possible either that both the CMV and RSV pro-
moter are inactivated over this time, or that the viral orrecombinant DNA is modified or sequestered in some
Materials and methodsway as to prevent gene transcription.Corneal endothelium is a critical cell both in primary
Reagents and antibodiescorneal disease, allograft rejection and allograft fail-RPMI 1640, minimal essential medium (MEM), Dulbec-ure.1,2,26 Therefore, the alteration of the endothelial cellco’s modified Eagle’s medium (DMEM), Hanks’ balancedgenome may have therapeutic potential in diverse cor-salt solution (HBSS), foetal calf serum (FCS), l-glutamine,neal diseases and vector-mediated local expression ofpenicillin and streptomycin were purchased from Gibcoimmunomodulatory molecules may have widespreadBRL (Paisley, UK). Caesium chloride and X-gal wereapplication in clinical transplantation. In this study, wefrom NBL Gene Sciences (Cramlington, UK). Ortho-phen-have examined the feasibility of using the adenoviral con-ylenediamine (OPD) tablets were obtained from Dakostructs for the expression of CTLA-4 Ig, which is a protein(High Wycombe, UK). All other chemicals were pur-that has been shown to block T cell costimulationchased from Sigma Chemical (Poole, UK), unless other-mediated by interactions between CD28 and the B7 fam-wise specified.ily (CD80 and CD86). This protein has been shown in a
Monoclonal antibodies against different epitopes of therange of models to block allograft rejection.18–20 ThisFc portion of human g1 immunoglobulin (SB7E6 andreport also shows that corneas modified by ex vivoSB7H2) were kindly donated by Dr Martin Glennieadenovirus-mediated gene transfer can produce CTLA-4(Tenovus Research Laboratories, Southampton, UK).Ig for up to 30 days, and this potential therapeutic pro-
tein, obtained from the culture supernatants of gene-modified corneas, is capable of binding B7.1 molecules. Cell lines
EA.hy-926, an endothelial cell hybridoma,27 was main-It is of interest that the CTLA-4 Ig is produced for alonger period than b-gal, suggesting that a secreted pro- tained in growth medium composed of RPMI 1640, 5%
FCS, 2 mm l-glutamine, 100 U/ml penicillin and 100tein may have a different kinetic of expression than acytoplasmic protein possibly due to its slow release from mg/ml streptomycin at 37°C and 5% CO2. DAP-3 human
fibroblast cells and DAP-3 transfected with B7.1 (CD80)secretory vesicles.In summary, this study has established a model for (DAP-3-B7.1) were cultured in DMEM supplemented
with 5% FCS, 2 mm l-glutamine, 100 U/ml penicillin andsuccessful reporter and therapeutic gene transfer to cor-neal endothelium. We have also demonstrated that 100 mg/ml streptomycin.28 293 Embryonic human kidney
cells, that contain the E1 region of the adenovirus gen-marker gene mRNA is detectable for only 3–7 days fol-lowing infection with recombinant virus, but the adeno- ome, were used in propagation and titration of adenovi-
ral vectors.29 This cell line was cultured in DMEM con-viral DNA can be detected for 56 days. Finally, we have

Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
645taining 10% FCS, 2 mm l-glutamine, 100 U/ml penicillin Infection and culture conditions for corneasEach human cornea segment was incubated with 1 ml ofand 100 mg/ml streptomycin.2% FCS in MEM containing 4 × 108 p.f.u. AdRL, 1.5 × 108
p.f.u. AdCB2 or 1 × 108 p.f.u. AdCTLA for 3 h at 37°C inCorneal samples5% CO2. Control mock-infected corneas were incubatedCorneas were removed at surgery from patients with ker-in virus-free medium. Following incubation, infected andatoconus, a disease in which corneal shape is abnormalcontrol specimens were washed three times with HBSSbut endothelial and epithelial cells normal. These corneasto avoid prolonged viral exposure and the possiblewere cut into four segments before infection with recom-recombinant protein contamination in the viral prep-binant adenovirus, which commenced in each case withinarations, which might have been produced during culti-4 h of surgical removal.vation in host 293 cells. The cornea samples, were main-tained in MEM supplemented with 10% FCS, 2 mm l-
Viral vectors glutamine, 100 U/ml penicillin and 100 mg/ml strepto-The recombinant E1-deleted type 5 adenoviral vectors mycin at 37°C and 5% CO2 for variable periods. CultureAdCB2, AdRL and AdCTLA were used in this study. medium was changed every 3 days.AdRL and AdCB2 contain the E. coli lacZ gene, encodingb-galactosidase under the transcriptional control of a Histochemical analysis of lacZ expressionRous sarcoma virus (RSV) and a human cytomegalovirus For morphological studies to determine the tissue localis-(CMV) promoter, respectively. The AdRL construct has ation of transgene expression, corneal specimens werepreviously been described.30 AdCB2 was made using a examined by histochemical X-gal staining at day 1 follow-642 bp fragment from pCDNAI (Invitrogen, NV Leek, ing infection by the AdRL or the AdCB2 recombinantThe Netherlands) containing the CMV promoter. The adenovirus constructs.4 Corneal sections were examinedvirus was produced by recombination of the plasmid for blue staining of cells by light microscopy.encoding the CMV promoter and the lacZ gene(pXCXCB2) with pJM17, containing the d1309 genome,31
Assay for b-gal activityin 293 cells as previously described.30 AdCTLA encodes b-Gal activity in corneal extracts was determined usinga mouse CTLA-4 human Fcg1 fusion protein driven by o-nitrophenyl-b-d-galactopyronidase (ONPG) as a sub-the RSV promoter. The extracellular region of mouse strate.4 The total protein content of the corneal lysatesCTLA-4 gene was amplified from BALB/c splenic RNA was also estimated using a modified Lowry method.35 b-by RT-PCR, using the following primers: mCTLA-4E: 5′- Gal activity was presented by normalizing to the totalUACGAATTCCTGAGGACCTCAGGCACATT-3′ (cont- protein content of the corneal sample.aining 5′ untranslated sequence and EcoRI site); andmCTLA-4J: 5′ UACGGATCCACTTACCTGGTAGAA- PCR analysis of lacZ mRNATCCGGGCATGGTTCT-3′ (containing membrane proxi- Total RNA was extracted from infected or mock-infectedmal extracellular sequence, a BamHI site and a splice human corneas using the RNAzol B RNA Isolation Sol-donor sequence). The CTLA-4 PCR product was direc- vent (Cinna/Biotecx Laboratories, Houston, TX, USA)tionally cloned into pBluescript (KS+), then subcloned according to the manufacturer’s instructions. The cDNAinto pIg vector,32 which contains the gene encoding for was prepared using the Pharmacia First-Strand cDNAhuman Fcg1, immediately preceded by a splice acceptor Synthesis kit (Pharmacia, Uppsala, Sweden) and storedsite. This complete CTLA-4 Ig insert was further sub- at −20°C. PCR amplification of the 1036 bp segment ofcloned into the pXCXRSV downstream of the RSV pro- the lacZ gene was performed as described below. Amoter,30 and the virus was produced by recombination primer pair to amplify a 540 bp segment of the house-with pJM17 as above. Viruses from the resulting plaques keeping b-actin gene (5′ primer: 5′-GTGGGGCGCCCCAwere plaque-purified three times and the DNA structure GGCACCA-3′; 3′ primer: 5′-CTCCTTAATGTCACGCACwas verified by restriction analysis. GATTTC-3′ was used for testing the presence of total
Viruses were grown in 293 cells until the cytopathic cDNA in each sample.36 The PCR reactions prepared aseffect occurred. The virus particles were released from described below were run under the following cyclingthe pellet of these cells by four freeze–thaw cycles, and conditions: 29 cycles of denaturation at 94°C for 1 min,purified by two rounds of caesium chloride (CsCl) den- annealing at 65°C for 1 min and extension at 72°C forsity gradient centrifugation, as described previously.33
2 min.After dialysis against PBS to remove CsCl, glycerol wasadded to a final concentration of 10% (v/v). The viral DNA isolation and analysis by polymerase chainpreparations were passed through a 0.2 mm filter and reaction (PCR)stored at −70°C. Viral titers were assessed by plaque Tissue debris remaining from corneal lysate preparationassay on 293 cells as described.33 The viral titres were was used for total DNA extraction and purification.1.9 × 109 p.f.u./ml, 5 × 109 p.f.u./ml, and 7.9 × 109
Briefly, corneal tissues were washed with ice cold PBS,p.f.u./ml for AdCB2, AdRL and AdCTLA4, respectively. incubated in a digestion buffer (100 mm NaCl, 10 mm Tris
Cl, 25 mm EDTA, 0.5% SDS and 0.1 mg/ml proteinase K)for 18 h at 50°C, followed by phenol/chloroform extrac-Determination of optimum conditions for infecting
endothelial cells tion, ethanol precipitation and resuspension in TE buffer(pH 8.0).37 This yielded approximately 200 ng/ml ofOptimum infection conditions for adenoviral vectors
were determined in EA.hy-926 cells as described pre- DNA. A primer pair was designed to amplify a 250 bpsegment of adenoviral DNA corresponding to the E2aviously.4,34 To determine the optimum conditions for
AdCTLA virus infection, the CTLA-4 Ig fusion protein sequence (5′ primer, 5′-ACTGGCAGGGACACGTTG-CGA-3′; 3′ primer, 5′-AGGAGCGTGCTGGCCAGCGT-was detected using an ELISA, as described below.

Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
646 3′). The PCR reaction mixture was prepared in a total styrene round-bottom analysis tubes before analysis witha Flow cytometer (Coulter, Luton, UK).volume of 50 ml containing 8 ml of corneal DNA
(approximately 5 ng/ml), 1 mm of deoxynucleoside tri-phosphate mixture, 10 mm of 5′ and 3′ oligonucleotide Acknowledgementsprimers, 10 × PCR buffer (Bioline, London, UK) and 2.5
The authors thank Professor Robert Lechler for kindlyU Taq polymerase (Bioline). The PCR was performedproviding the DAP-3 and the DAP-3-B.7.1 cells and Drunder the following conditions: 94°C for 1 min; followedJon Friedland for giving the b-actin primers and the pro-by addition of Taq polymerase, 29 cycles of 94°C for 1tocol. We also thank Dr Martin Glennie for providingmin, 55°C for 1 min, and 72°C for 1 min. PCR productsSB7E6 and SB7H2 antibodies. This work was supportedobtained in this manner were run on a 2% agarose gelby the Iris Fund for Prevention of Blindness, the Specialand visualised with UV light.Trustees of Moorfields Eye Hospital (DFPL), Uludag Uni-The oligomeric primers flanking an internal 1036 bpversity, Turkey (HBO), and Medical Research Councilsequence of the lacZ gene have been previously pub-(MJD). DOH is the recipient of a British Heart Foundationlished (primer 1: 5′-GCCGACCGCACGCCGCATCCProfessorial award.AGC-3′, primer 2: 5′-CGCCGCGCCACTGGTGTGGGC-
3′).38 These primers were used for detection of the lacZgene. Fifty microlitres of PCR reaction mixture was pre- Referencespared as described above and subjected to the following
1 Price FW, Whitson WE, Collins KS, Marks RG. Five-year cornealcycling conditions; 30 cycles of denaturation at 94°C forgraft survival. Arch Ophthalmol 1993; 111: 799–805.1 min, annealing and extension at 72°C for 2 min. PCR
2 Vail A et al. Corneal transplantation in the United Kindom andproducts were analyzed by gel electrophoresis as above.Republic of Ireland. Br J Ophthalmol 1993; 77: 650–652.
3 Bourne FW, Hodge DO, Nelson LR. Corneal endothelium fiveyears after transplantation. Am J Ophthalmol 1994; 118: 185–196.Time course of CTLA-4 Ig fusion protein production
4 Larkin DFP et al. Adenovirus-mediated gene delivery to the cor-Levels of CTLA-4 Ig fusion protein in the supernatantsneal endothelium. Transplantation 1996; 61: 363–370.from AdCTLA-infected corneas were measured by sand-
5 Fehervari Z et al. Gene transfer to ex vivo stored corneas. Corneawich ELISA. Corneal culture supernatants were removed(in press).from human corneas incubated with AdCTLA or virus- 6 Schofield JP, Caskey CT. Non-viral approaches to gene therapy.
free medium, and replaced with fresh culture medium Br Med Bull 1995; 51: 56–71.on days 2–36 following infection. 96-Well ELISA plates 7 Khodadoust AA, Silverstein AM. Transplantation and rejectionwere coated with 10 mg/ml of SB7E6 mAb recognising of individual cell layers of the cornea. Invest Ophthalmol 1969; 8:the Fc portion of human IgG1. After incubation for 1 h at 180–195.
8 Williams KA et al. Patterns of corneal graft rejection in the rabbit37°C, the plates were washed with PBS/0.1% Tween-20and reversal of rejection with monoclonal antibodies. Transplan-and blocked with 100 ml of 0.1% milk for 30 min at roomtation 1992; 54: 38–43.temperature. Standard calibration curves were con-
9 Larkin DFP, Calder VL, Lightman SL. Identification and charac-structed using human IgG1. Sixty microlitres of cornealterisation of cells infiltrating the graft and aqueous humour inculture supernatant or standard was added to the appro- rat corneal allograft rejection. Clin Exp Immunol 1997; 107:priate wells in triplicate and incubated for 1 h at 37°C. 381–391.
After the primary incubation, the plates were washed 10 Mueller DL, Jenkins MK, Schwartz RH. Clonal expansion versusfour times with PBS/0.1% Tween-20. Sixty microlitres of functional clonal inactivation: a costimulatury signalling path-HRP-conjugated SB7H2 mAb (anti-human IgG1 Fc) was way determines the outcome of T cell antigen receptor occu-
pancy. Annu Rev Immunol 1989; 7: 445–480.added to each well at a dilution 1:2000 and incubated for11 Linsley PS et al. Binding of the B cell activation antigen B7 to1 h at 37°C. After an additional four washes, 50 ml of
CD28 costimulates T cell proliferation and interleukin 2 mRNAOPD substrate, prepared according to the manufacturer’saccumulation. J Exp Med 1991; 173: 721–730.instructions, was added to the wells and incubated in the
12 Gimmi CD et al. B-cell surface antigen B7 provides a costimu-dark at room temperature for 20–30 min. The reactionlatory signal that induces T cells to proliferate and secretewas stopped by adding 50 ml of 5% H2SO4, and the plates interleukin 2. Proc Natl Acad Sci USA 1991; 88: 6575–6579.
were read at 491 nm using the microplate reader. 13 Harding FA et al. CD28-mediated signalling co-stimulatesmurine T cells and prevents induction of anergy in T cell clones.Nature 1992; 356: 607–609.Flow cytometric analysis of CTLA-4 Ig activity
14 Tan P et al. Induction of alloantigen-specific hyporesponsivenessThe binding activity of the CTLA-4 Ig fusion protein was in human T lymphocytes by blocking interaction of CD28 withtested by flow cytometry. Briefly, CD80 transfected DAP- its natural ligand B7/BB1. J Exp Med 1993; 177: 165–173.3 fibroblast (DAP3-B7.1) cells were adjusted to 1 × 106
15 Boussiotis VA et al. B7 but not intracellular adhesion molecule-cells per ml and aliquoted in ice cold microcentrifuge 1 costimulation prevents the induction of human alloantigen-tubes. One hundred microlitres of supernatant from specific tolerance. J Exp Med 1993; 178: 1753–1763.
16 Linsley PS et al. CTLA-4 is a second receptor for the B cell acti-AdCTLA-infected corneal samples, removed after ex vivovation antigen B7. J Exp Med 1991; 174: 561–569.culture for 24 h following infection, were incubated with
17 Harper K et al. CTLA-4 and CD28 activated lymphocytethese cells on ice for 30 min. PBS and a human IgG1molecules are closely related in both mouse and human as tostandard were employed as negative controls, whilesequence, message expression, gene structure, and chromo-purified CTLA-4 Ig protein was used as a positive con-somal location. J Immunol 1991; 147: 1037–1044.trol. After incubation, cells were washed once with 18 Lenschaw DJ et al. Long-term survival of xenogeneic pancreatic
PBS/azide (0.1% azide in PBS) and then incubated with islet grafts induced by CTLA4Ig. Science 1992; 257: 789–792.100 ml of FITC-conjugated mouse anti-human IgG1 mAb 19 Lin H et al. Long-term acceptance of major histocompatibility(Dako) on ice for 30 min. Finally, the cells were washed complex mismatched cardiac allografts induced by CTLA-4 Ig
plus donor specific transfusion. J Exp Med 1993; 178: 1801–1806.once and resuspended in 100 ml of PBS/azide in poly-

Ex vivo adenovirus-mediated gene transfer to human corneaHB Oral et al
64720 Pearson TC et al. Transplantation tolerance induced by CTLA- 30 Byrnes AP, Rusby JE, Wood MJA, Charlton HM. Adenovirus4 Ig. Transplantation 1994; 57: 1701–1706. gene transfer causes inflammation in the brain. Neuroscience
21 Mashhour B, Couton D, Perricaudet M, Briand P. In vivo 1995; 66: 1015–1024.adenovirus-mediated gene transfer into ocular tissues. Gene 31 McGrory WJ, Bautista DS, Graham FL. A simple technique forTherapy 1994; 1: 122–126. the rescue of early region I mutations into infectious human
22 Feldman ST et al. Gene transfer to the anterior segment of the adenovirus type 5. Virology 1988; 163: 614–617.eye by infection with recombinant viral vector. Invest Opthalmol 32 Simmons DL. Cloning cell surface molecules by transientVis Sci 1994; 35 (Suppl.): 1383 (Abstr.). expression in mammalian cells. In: Hartley D (ed). Cellular Inter-
23 Budenz DL, Bennett J, Alonso L, Maguire A. In vivo gene trans- actions in Development. Oxford University Press: Oxford, 1993,fer into murine corneal endothelial and trabecular meshwork pp 93–127.cells. Invest Opthalmol Vis Sci 1995; 36: 2211–2215. 33 Green M, Wold WSM. Human adenoviruses: growth, purifi-
24 Chodosh J, Miller D, Stoop WG, Pflugfelder SC. Adenovirus epi- cation and transfection assay. Meth Enzymol 1979; 58: 434–435.thelial keratitis. Cornea 1995; 14: 167–174. 34 Sanes JR, Rubinstein JLR, Nicholas J-F. Use of recombinant
25 Guzman RJ et al. Efficient and selective adenovirus-mediated retroviruses to study post-implantation cell image in mousegene transfer into vascular neointima. Circulation 1993; 88: embryos. EMBO J 1986; 5: 3133–3142.2838–2848. 35 Krezse G-B. Methods for protein determination. In: Bergmeyer J,
26 Larkin DFP. Corneal allograft rejection. Br J Ophthalmol 1994; 78: Grassl M (eds). Methods of Enzymatic Analysis: Samples, Reagents,649–652.
Assessment of Results, vol 2. Verlag Chemie: Weinheim, 1983, pp27 Edgell C-JS, McDonald CC, Graham JB. Permanent cell line89–92.expressing human factor VIII-related antigen established by
36 Zhang M et al. Interleukin-12 at the site of disease in tubercu-hybridization. Proc Natl Acad Sci USA 1983; 80: 3734–3737.losis. J Clin Invest 1994; 93: 1733–1739.28 Hargreaves R, Logiou V, Lechler R. The primary alloresponse
37 Moore DD. Preparation of genomic DNA. In: Ausubel FM et alof human CD4+ T cells is dependent on B7 (CD80), augmented(eds). Current Protocols in Molecular Biology, vol 1. John Wiley:by CD58, but relatively uninfluenced by CD54 expression. IntNew York, NY, 1992, pp 2.2.1–2.2.3.Immunol 1995; 7: 1505–1513.
38 Csete ME et al. Efficient gene transfer to pancreatic islets29 Graham FL, Smiley J, Russell WC, Nairn R. Characteristics of amediated by adenoviral vectors. Transplantation 1995; 59: 263–human cell line transformed by DNA from human adenovirus268.type 5. J Gen Virol 1977; 36: 59–74.