genetic technologies for the identification of plant genes (genetic technologies for the...
TRANSCRIPT

Genetic technologies for the identification of plant genescontrolling environmental stress responses
Csaba PapdiA, Mary Prathiba JosephA, Imma Pérez SalamóA, Sabina VidalB
and László SzabadosA,C
AInstitute of Plant Biology, Biological Research Centre, 6726-Szeged, Temesvári krt. 62, Hungary.BFacultad de Ciencias, Universidad de la República, Iguá 4225, CP 11400, Montevideo, Uruguay.CCorresponding author. Email: [email protected]
Abstract. Abiotic conditions such as light, temperature, water availability and soil parameters determine plant growth anddevelopment. The adaptation of plants to extreme environments or to sudden changes in their growth conditions is controlledby awell balanced, genetically determined signalling system,which is still far from being understood. The identification andcharacterisation of plant geneswhich control responses to environmental stresses is an essential step to elucidate the complexregulatory network, which determines stress tolerance. Here, we review the genetic approaches, which have been used withsuccess to identify plant geneswhich control responses to different abiotic stress factors.Wedescribe strategies and conceptsfor forward and reverse genetic screens, conventional and insertion mutagenesis, TILLING, gene tagging, promotertrapping, activation mutagenesis and cDNA library transfer. The utility of the various genetic approaches in plant stressresearch we review is illustrated by several published examples.
Additional keywords: abiotic stress, activation, Arabidopsis, cold, drought, forward genetics, gene, genetic screen, genetagging, heat stress mutagenesis, osmotic stress, oxidative stress, reverse genetics, salinity, variability.
Introduction
Abiotic stresses, especially drought, extreme temperatures andsoil salinity, have increasing agronomic, economic andecologicalimpacts in Europe and worldwide. Recent studies indicate acorrelation between the increased frequency of extremeenvironmental events and global warming, underlining anurgent need for protective measures including the developmentand introduction of new crop cultivarswith enhanced tolerance toenvironmental stresses (Boyer 1982; Etterson and Shaw 2001;Mittler 2006). Most of our knowledge of the control of abioticstress responses has derived from molecular genetic analysisof the model plant Arabidopsis thaliana (L.). Arabidopsis isnot particularly stress tolerant and has limited capacity towithstand harmful effects of extreme environmentalconditions. Nevertheless, the contribution of this model plantto the advance of plant stress biology cannot be underestimated.Information on abiotic stress signal transduction has beengenerated almost exclusively using the Arabidopsis model.Arabidopsis became the model plant for plant biology becauseit has numerous advantageous features, which allow the efficientcombination of genetic and molecular analysis (Redei 1975;Meyerowitz 1989). The publication of the whole Arabidopsisgenome sequence in 2000 (The Arabidopsis Genome Initiative2000a), and the development of genomic tools for Arabidopsisresearch have allowed the adaptation of systems biologyapproaches to this species, accelerating research on differentaspects of plant biology, including abiotic stress responses
(Weigel et al. 2000; Bohnert et al. 2006; Koiwa et al. 2006).Information using Arabidopsis can easily be validated in otherspecies, and technical advances can be adapted to research anddevelopment programs focusing on crop plants.
Plant responses to environmental stresses are controlled byinterconnected signalling networks, which regulate theexpression of large sets of target genes. Receptor moleculesinvolved in primary perception of abiotic stress stimuli are atpresent largely unknown in higher plants, with the exceptionof AtHK1, a putative two-component hybrid histidine kinaseosmosensor, which is implicated in osmosensing under saltstress (Urao et al. 1999). G-protein complexes have beensuggested to function as receptors in sensing stress signals(Misra et al. 2007; Pandey et al. 2009), but the transmission ofprimary stress signals, sensed by receptors to downstreameffector molecules is poorly understood. One of the earliestevents in the process is the release of intercellular calcium thattriggers further stress specific adaptation responses (Knight 1999;Sanders et al. 2002). Numerous further downstream signallingcomponents have been identified including hydrogen peroxide,phospholipid secondary messengers, and cytosolic Ca2+
oscillation, which controls the activity of CDPK, SnRK andMAPK-type protein kinase phosphorylation cascades (Knight1999; Zhu 2002, 2003; Wang 2004; Bartels and Sunkar2005). Abscisic acid (ABA) is a major ‘stress’ hormone thatplays a central role in the control of abiotic stress responses. ABAcontrols gas exchange and photosynthesis by triggering stomatal
CSIRO PUBLISHING Review
www.publish.csiro.au/journals/fpb Functional Plant Biology, 2009, 36, 696–720
� CSIRO 2009 10.1071/FP09047 1445-4408/09/080696

closure, induces the synthesis of seed storage proteins and lipids,promotes seed desiccation tolerance and dormancy, inhibitstransition from embryonic stage to germination, and regulatesnumerous other aspects of plant development. ABA stimulatesthe expression ofmany drought and osmotic stress-induced genesthrough a complex signalling mechanism. ABA perception andsignalling appear to involve a branching pathway, in which Ca2+
release, reactive oxygen species (ROS), and protein kinases andphosphatases are the most important components (Leung andGiraudat 1998; Finkelstein et al. 2002; Wang and Zhang 2008).The ABA dependent and independent signalling cascadescontrol the activity of a large set of transcription factors andregulate the expression of downstream target genes through atleast six different pathways (Zhu 2002; De Smet et al. 2006;Yamaguchi-Shinozaki and Shinozaki 2006; Nakashima et al.2009).
In this paper, we review the genetic and genomic approacheswhich have been used in plants to dissect the regulation ofabiotic stress responses. Although most examples are obtainedfrom Arabidopsis, references to other plants are made whererelevant advances have been made (Table S1 available as anAccessory Publication to this paper).
Mutant isolation and forward genetic approaches
In order to perform genetic analysis, genetic variation has tobe identified by specific screening procedures (Fig. 1). Inforward genetic strategies, genetic variation is generated firstthrough chemical, physical or biological mutagenesis, andthe mutagenised plant population is screened for the phenotypeof interest. Alternatively, natural variation can provide valuableresourcesofgeneticpolymorphism. InArabidopsis, ethylmethane-sulfonate (EMS) is the most frequently used chemical mutagen,which predominantly generates point mutations (Rédei andKoncz 1992; Kim et al. 2006). Physical mutagenesis can beperformed by gamma irradiation and by fast neutrons, whichusually generates small deletions (Rédei and Koncz 1992; Liand Zhang 2002; Shikazono Suzuki et al. 2005). The number ofmutagenised plants to be screened depends on the mutationfrequency, which depends on the mutagenesis treatment. Afterlow mutagen doses, large number of mutagenised plants have tobe screened. High mutagen doses can be more efficient, butundesired mutations need to be removed later by repeatedbackcrossing of the mutant line with parental lines (Rédei andKoncz 1992). To isolate mutations in genes which control stressresponses, mutagenesis is usually followed by genetic screensdesigned to identify mutants with enhanced or reduced toleranceto controlled environmental stress conditions. In order to screenlarge numbers of plants, and increase the chances of identifying thehighest number of mutants, the screening procedure should beas simple as possible (Page and Grossniklaus 2002; Alonso andEcker 2006). For these reasons, in vitro-based screens have oftenbeen developed.
Forward genetic screens in Arabidopsis represent a powerfuland unbiased approach to identify mutants with altered stressresponses. Identification of the corresponding gene requiresmap-based cloning, which is often a tedious and time consumingprocedure. For mapping, mutant plants are crossed with anotherecotype and a small population of F2 plants are genotyped to
determine genetic linkage to molecular markers representingeach chromosomal region. Fine-scale mapping is performed ona larger population of F2 plants narrowing linkage to a 30–50 kbpregion. Localisation of the mutation is done by sequencing this
5′
5′
3′
3′
5′
5′
5′
5′
5′
3′
3′
3′
3′
3′
transcribed region
M
deletion
T-DNA
T-DNA
T-DNA
cDNA
reporter
enhancer
+
(a)
(b)
(c)
(d )
(e )
(f )
(g )
Fig. 1. Schematic representation of several genetic strategies used toidentify plant genes. (a) Simplified structure of a plant gene consisting of50 regulatory sequences, transcribed region with exons and introns, and30 regulatory sequences. (b) Point mutations modify but do not destroygene structure, resulting in modified gene product. (c) Deletions remove agene or part of it, which results in loss of the gene function. (d) Insertionsinterrupt and destroy gene structures, leading to loss-of-function mutations.(e) Gene traps can inactivate genes and simultaneously create in situ genefusions with the reporter gene of the insertion element. ( f ) Activatormutagenesis uses strong constitutive enhancers or promoters in theinsertion element, and usually generates gain-of-function mutations byactivating the transcription of flanking genes. (g) Random cDNA transferuses a cDNAlibrary cloned in a plant expressionvector,where transcriptionofthe inserted cDNA is controlled by the promoter of the expression cassette.The result is dominant gain or loss-of-function phenotype, depending on theintegrity of the inserted cDNA.
Genetic identification of plant genes in response to stress Functional Plant Biology 697

region of the mutant and wild type chromosomal DNA (Jander2006). The efficiency of map-based cloning depends on theavailability of a detailed genetic map, and saturating thegenome with PCR-based markers facilitates mapping andcloning of mutant loci. Thousands of markers, Insertion/Deletion (InDel) polymorphism and Single NucleotidePolymorphism (SNPs) are available for fine mapping inArabidopsis, making map-based cloning feasible within a year(Cho et al. 1999; Peters et al. 2003; Jander 2006). Recently, DNAmicroarray technologies have been adapted for mappingmutations in forward genetic studies. Oligonucleotide arrayswere originally designed for transcript profiling, but have beenadapted for high density genotyping to detect polymorphisms,mutation mapping and mapping and cloning of quantitative traitloci (QTL) (Spiegelman et al. 2000; Borevitz et al. 2003; Hazenet al. 2005). Although the detection of single base substitutionsis limited, deletions, as caused by fast neutrons can easily bedetected. Natural variation or induced mutations with InDelpolymorphism can be recognised with higher precision withspecific genotyping arrays (Salathia et al. 2007). To improvethe efficiency of deletion mapping, a novel tiling array, AtMap1was constructed and used for mapping several deletion mutantsand the detection of natural polymorphism which involvesdeletions (Nagano et al. 2008).
Genome-wide mapping of mutations can be performed inplant species which have a well defined genetic map. Highdensity linkage maps have been established for most of theimportant crops, such as barley (Hordeum vulgare L.; Horiet al. 2003; Wenzl et al. 2006), wheat (Triticum aestivum L.;Dilbirligi et al. 2006; Peleg et al. 2008), rice (Oryza sativa L.;Harushima et al. 1998), Sorghum sp. (Draye et al. 2001; Menzet al.2002), potato (Solanum tuberosumL.;Tanksley et al.1992),tomato (Lycopersicom esculentum L.; Tanksley et al. 1992;Saliba-Colombani et al. 2000), Brassica sp. (Pradhan et al.2003; Sun et al. 2007), soybean (Glycine max L.; Xia et al.2007), papaya (Carica papaya L.; Ma et al. 2004), Lotus sp.(Wang et al. 2008) and Rose sp. (Yan et al. 2005). Althoughgene identification in non-model species is usually a difficulttask due to technical reasons, i.e. large genomes, low markerdensities, lack of genomic sequence information, etc., microarraytechnology offers a powerful strategy to identify deletions andfacilitate cloning of mutant alleles in complex genomes such asCitrus (Ríos et al. 2008).
Recent advances in massively parallel sequencingtechnology offer new and robust strategies to identify geneticpolymorphism and for mutation mapping (Shendure and Ji 2008;Ansorge 2009). Combination of bulked segregant analysisand deep sequencing of F2 generation mutant plants willfacilitate mapping of Arabidopsis mutations in a short time atrelatively low cost (Ossowski et al. 2008; Lister et al. 2009).Withfurther improvement of sequencing technology the mapping ofmutations and polymorphism in plant specieswith large genomeswill also be possible.
Examples for genetic screens and gene identification
In order to identify salt hypersensitive Arabidopsismutants, Zhu(2002) screened mutagenised M2 generation seedlings forimpaired root curling and elongation on inverted vertical plates
containing culture medium with sublethal concentrations ofNaCl (Ishitani et al. 1997). Differences in root growth onthese conditions could be scored easily. Identification andmolecular analysis of the Salt Overly Sensitive (sos) mutantshave confirmed the utility of this screening procedure and leadto the discovery of a regulatory pathway which is essentialto maintain ion homeostasis (Zhu 2002; Chinnusamy et al.2006). The so-called SOS pathway detects and transducesCa2+ signals generated by salt stress. SOS3, a calcineurinB-type calcium binding protein is able to detect changes inintracellular Ca2+ concentrations caused by high salinity(Ishitani et al. 2000). SOS3 interacts with the SnRK3-typeSOS2 protein kinase (Halfter et al. 2000; Liu et al. 2000;Gong et al. 2002a), which activates several membrane iontransporters such as the plasmamembrane Na+/H+ antiporterSOS1 (Wu et al. 1996; Shi et al. 2000), the H+/Ca2+ antiporterCAX1 (Cheng et al. 2004), or the vacuolar H+-ATPase(Batelli et al. 2007). Conservation of the SOS pathway in riceand wheat suggests that these factors are important for salttolerance through maintaining Na+/K+ homeostasis in dicotsand monocots alike (Martínez-Atienza et al. 2007; Mullanet al. 2007). In a similar forward genetic approach, inhibitionof lateral root development by osmotic stress was employed toisolate 30Arabidopsismutants with altered sensitivity to droughtand abscisic acid (Xiong et al. 2006). Analysis of the DroughtInhibition of lateral root Growth (dig) mutants revealedthat responses of root growth to osmotic stress are linked todrought tolerance in the whole plant.
A salt hypersensitive mutant was mapped with a microarray-based strategy and the mutation was identified as a 523 bpgenomic deletion in the AtHKT1 gene (Gong et al. 2004b).Segregation analysis and mutant complementation confirmedthat the deletion and inactivation of AtHKT1 was responsiblefor the salt sensitivity of the mutant.
Genetic screens for deregulated expression of stress-responsive reporter gene constructs provide an alternativeoption to isolate mutations in regulatory genes that eitheractivate or repress the activity of the reporter gene (Xionget al. 1999b). As multiple regulatory networks control theexpression of stress-induced genes (Yamaguchi-Shinozaki andShinozaki 2006; Nakashima et al. 2009) diverse reporter geneconstructs can be designed to dissect each of these regulatorycascades. Combined with non-destructive bioluminescenceimaging, alterations of the cold, drought and ABA-responsiveRD29A-LUC reporter gene activity led to the identification ofcos, los and hos mutants, corresponding to constitutive, low orhigh expression of osmotically responsive genes, respectively(Ishitani et al. 1997; Chinnusamy et al. 2002). Such luciferasescreens were employed to identify important regulatorycomponents of salt, cold and ABA signalling pathways(Ishitani et al. 1998; Xiong et al. 1999a; Xiong et al. 2001;Lee et al. 2002; Zhu et al. 2004). The RD29A-LUC reportersystem was used for example to identify several alleles of thetranscriptional activator FIERY2, which regulates the expressionof genes with DRE/CRT-type cis elements in their promoters(Xiong et al. 2002). The Arabidopsis RCI2A gene is induced byseveral abiotic stresses through an ABA-dependent pathway.Alterations in RCI2A-LUC activity were used to identifymutants with impaired tolerance to freezing, dehydration, salt
698 Functional Plant Biology C. Papdi et al.

stress and ABA sensitivity (Medina et al. 2005). The 2-cysteineperoxiredoxin A (2CPA) gene encodes an important chloroplastantioxidant enzyme. 2CPA transcription is controlled by redoxbalance, and accumulation of ROS. A transgenic Arabidopsisline, expressing the 2CPA-LUC reporter gene was used toidentify the Arabidopsis redox imbalanced (rimb) mutants,where transcriptional regulation of the 2CPA gene, andnumerous other redox-controlled genes were affected (Heiberet al. 2007).
Abscisic acid has a central role in controlling droughtresponses, stomatal closure, root growth, expression ofnumerous stress-induced genes and the accumulation ofprotective compounds such as proline. The isolation andcharacterisation of Arabidopsis mutants defective in ABAsynthesis, perception or signal transduction is thereforeessential for understanding the genetic basis of droughttolerance. The first ABA response mutants were identified byABA insensitive or hypersensitive germination, seedling or rootgrowth. The dominant abi1 and abi2 mutants show ABA-resistant germination and pleiotropic defects in ABA responsesin whole plants (Koornneef et al. 1984). Map-based cloning leadto the molecular characterisation of ABI1 and ABI2 genes, whichencode similar PP2C type serine-threonine phosphatases and areimportant components of whole-plant responses to water deficitand osmotic stress (Leung et al. 1994, 1997; Meyer et al. 1994).Other ABA signalling mutants have been isolated in similargermination screens, including the ABA insensitive abi3, abi4,and abi5 mutants, which lead to the identification of thecorresponding genes encoding different transcription factors,essential for the activation of different classes of ABA-regulated genes (Giraudat et al. 1992; Finkelstein et al. 1998;Finkelstein and Lynch 2000). ABA hypersensitive mutants havebeen identified in several genetic screens, many of them based onthe inhibition of germination by ABA. Examples include theenhanced response toABA (era1) mutant (Cutler et al. 1996), thegrowth control byABA (gca1-gca8) mutants (Himmelbach et al.1998) and the ABA hypersensitivity (abh1) mutant (Hugouvieuxet al. 2001), which displays pleiotropic ABA responses such asenhanced guard cell response and drought tolerance. ERA1encodes the b subunit of farnesyl transferase, a lipid transferenzyme, while ABH1 and other ABA response loci encodeRNA binding proteins, suggesting that they are involved inRNA processing.
Infrared thermal imaging offers an alternative approach toisolate ABA insensitive mutants, in which genetic defectsspecifically affect the ABA sensitivity of stomatal guard cells.Changes in leaf temperature indicate alterations in ABA-dependent stomatal opening and respiration, which can bevisualised by infrared thermography. Differences in leaftemperature led to the isolation of the open stomata mutantsost1 andost2,whichdisplayed lower leaf temperatures andhighertranspirational water loss than wild type plants (Merlot et al.2002). Map-based cloning revealed that the OST1 locus encodesa SnRK2-type protein kinase, which mediates the closureof stomatal aperture triggered by ABA (Mustilli et al. 2002).Advances in the non-invasive infrared thermal imagingtechnology offer an efficient screening tool to identify novelmutants defective in regulation of stomatal opening and droughttolerance (Wang et al. 2004).
Natural variation and quantitative trait loci
Naturally occurring variation is one of themost important geneticresources for plant breeding and is available in practically all plantspecies. Responses to environmental conditions depend onnumerous genes and are typically controlled by QTLs, whichrepresent heritable variability of traits and characters. QTLscorrespond to natural alleles, which otherwise have smalleffects on the tolerance to abiotic constrains. Mapping of suchQTLs may lead to identification and cloning of importantregulatory genes or allelic variants and provide geneticmarkers for molecular breeding or cloned genes for geneticengineering to improve stress tolerance. Thus, improvement ofcrop performance in suboptimal or extreme environmentalconditions will benefit from the identification andcharacterisation of such adaptive QTLs (Collins et al. 2008).Hundreds of reports have described QTLs which can affectabiotic stress tolerance (Flowers et al. 2000; Koornneef et al.2004; Munns et al. 2006; Kliebenstein 2009). However, geneticdissection of QTLs, mapping the genes and alleles thatcontribute to stress adaptation as well as the identification ofpolymorphism at the nucleotide level has been hindered for along time by the lack of adequate tools. With the use ofrecombinant inbred lines (RILs) the dissection and analysis ofQTLs became possible in Arabidopsis as well as in severalimportant crops (Koornneef et al. 2004; Collins et al. 2008;O’Neill et al. 2008). Nevertheless, QTL mapping remains atedious and time-consuming task. Next-generation sequencingof natural accessions can reveal sequence variability in genomescale and will facilitate the large scale identification of SNP-sand InDels in different ecotypes and varieties (Ossowski et al.2008) (http://1001genomes.org/, verified 7 July 2009). Thecomprehensive analysis of natural polymorphism and variationin stress tolerance will facilitate the understanding of geneticbases of adaptation to extreme environmental conditions.
QTLs regulating responses to different abiotic stress factorshave been identified in several plant species. For example, inter-and intra-specific variability in salt tolerance has allowed theidentification of several QTLs that contribute to this trait inplants. Salt tolerance is determined by several physiologicalcomponents such as sodium transport and exclusion, toleranceto osmotic and oxidative stress, and tolerance to ion toxicity attissue and cell level (Munns and Tester 2008). Ion homeostasisis a key element of salt tolerance, and is determined by iontransport, exclusion or accumulation (Serrano and Rodriguez-Navarro 2001). Several major and minor QTLs have beenmapped in different plants, and used to identify importantcomponents controlling Na accumulation. High-throughputionomic profiling, coupled with microarray-based geneticanalysis lead to the identification of novel natural variants of theNa+ transporter AtHKT1, which controls salt tolerance inArabidopsis (Rus et al. 2006). Natural variation of sodiumaccumulation in shoots and salt tolerance was shown to dependon the expression patterns of AtHKT1 alleles in differentArabidopsis accessions. Polymorphism of the AtHKT1 genein salt tolerant and sensitive wild populations of Arabidopsistherefore confirmed the importance of this transporter in salttolerance (Rus et al. 2006). Nax1/TmHKT1;4 and Nax2/TmHKT1;5 salt tolerance loci of wheat have been identified by
Genetic identification of plant genes in response to stress Functional Plant Biology 699

QTL mapping and were shown to control sodium transport(Lindsay et al. 2004; James et al. 2006). Nax1/TmHKT1;4 is aNa+ transporter (HKT7) gene (Huang et al. 2006). Fine mappingwith microsatellite markers suggested that the high-affinity K+
transporter TmHKT1,5 (HKT8) was the candidate gene forNax2 and the homoeologous Kna1/TaHKT1;5 in bread wheat(Byrt et al. 2007). SKC1 is a major QTL for salt tolerance in rice,which affects sodium accumulation and transport. Map-basedcloning of the SKC1 locus lead to the identification ofOsHKT1,5,which is an orthologue of the wheat TmHKT1,5 gene andfunctions as a Na+-selective transporter (Ren et al. 2005).Physiological data confirmed that SKC1 regulates K+/Na+
homeostasis and is important in salt tolerance (Ren et al. 2005).Mapping and analysis of HKT1 alleles in several plant species,confirmed that control of Na+ accumulation and K+ uptakethrough specific transporters is a general mechanism for salttolerance (Platten et al.2006; Rodriguez-Navarro andRubio 2006;Gupta et al. 2008; Huang et al. 2008; Munns and Tester 2008).
Transpiration efficiency is an important component ofwater use and drought tolerance. QTLs have been identifiedfor transpiration efficiency in several plant species, withoutthe contributing genes having been identified. ERECTA wasthe first Arabidopsis gene, which was mapped as a mainQTL, and regulates transpiration efficiency by controlling leafphotosynthesis efficiency and stomatal conductance (Masle et al.2005). ERECTA encodes a LRR-type receptor-like kinase (RLK)and was previously known to control inflorescence development(Torii et al. 1996) and resistance against bacterial wilt (Godiardet al. 2003).
Cold tolerance correlates with geographical distribution ofa species and is an important component of adaptation toclimates with low temperatures. Freezing tolerance iscontrolled by seven QTLs in Arabidopsis. QTL mappingrevealed that the C-repeat Binding Factor (CBF) locus is themost important component in cold acclimation (Alonso-Blancoet al. 2005). CBF genes encode transcriptional activators whichbind to C-repeat or DRE promoter elements and activate a largeset of stress-induced genes. Low freezing tolerance of the Cviecotype was associated with a low expression level of CBF2caused by a promoter deletion (Alonso-Blanco et al. 2005).Nevertheless correlation of freezing tolerance and CBF geneexpression levels in contrasting Arabidopsis ecotypes couldnot be confirmed in more recent studies, indicating that coldtolerance is determined by complex network of genes (Le et al.2008; McKhann et al. 2008). In barley, QTL mapping identifiedthe CBF genes as important determinants of cold and droughttolerance (Tondelli et al. 2006). Frost tolerance in diploid wheat(Triticum monococcum L.) is controlled by multiple QTLs.Several CBF genes were mapped into QTLs which determinesfrost survival, suggesting that wheat CBF genes are importantfor cold acclimation and resistance to freezing temperatures(Miller et al. 2006; Knox et al. 2008). Differences inexpression levels of several CBF genes were shown toinfluence frost tolerance in wheat (Vágújfalvi et al. 2005).
Insertion mutagenesis
Traditional mutagenesis has generated numerous mutants whichare valuable resources for genetic analysis. Identification of the
corresponding gene is however often difficult due to thetedious work of map-based cloning. Linking of molecular andphysiological studies with genetic analysis is easier whenmutations are generated with insertions of DNA elements.Mobile genetic elements can be endogenous or introducedtransposons or the T-DNA of the Ti plasmid of Agrobacteriumtumefaciens, which is covalently integrated into the genome ofthe host cell (Bancroft et al. 1992; Zupan and Zambryski1995; Azpiroz-Leehan and Feldmann 1997; Meissner et al.2000). Integration of a DNA fragment with the size ofseveral thousand base pairs can disrupt the sequence of agene and effectively eliminate or reduce its activity (Fig. 1).Chromosomal loci are not onlymutated by T-DNAor transposoninsertions but also labelled by these molecular tags, which canbe detected by standard molecular methods (Koncz et al. 1992;Azpiroz-Leehan and Feldmann 1997; Martienssen 1998).
The most frequently used insertion elements in Arabidopsisgenetics are the Ac/DS or En/I type transposable elements(Bancroft et al. 1992; Martienssen 1998; Wisman et al. 1998),or theAgrobacteriumT-DNA (Feldmann et al. 1989;Koncz et al.1989; Koncz et al. 1992; Azpiroz-Leehan and Feldmann 1997).A. tumefaciens is a Gram negative soil bacterium, which is able totransfer and covalently insert a fragment of its large Ti plasmid(T-DNA) into the genomic DNA of the infected host cell (Schellet al. 1979; Caplan et al. 1983; Zupan and Zambryski 1995;Zupan et al. 2000). Integration of the T-DNA into the genome ofthe host cell takes place via illegitimate recombination, whichdoes not require sequence homology of the target genomic region(Tinland and Hohn 1995; Hansen and Chilton 1999; Zupan et al.2000). Consequently, distribution of the T-DNA integration sitesin theArabidopsisgenome is nearly random, although integrationfrequency is lower in centromeric regions, andhigher in gene-richchromosomal regions (Szabados et al. 2002; Alonso et al. 2003).Moreover, T-DNA integration in promoter and 50 untranslatedregions of actively transcribed plant genes is more frequentthan in intergenic regions (Szabados et al. 2002; Li et al.2006). Preference for integration in promoter regions can beexplained by the interaction of the TATA binding protein withthe Agrobacterium VirD2 protein, which is phosphorylated byPolII-CTD interacting CDKD-type kinases (Bako et al. 2003).To make use of the Agrobacterium T-DNA transfer mechanism,numerous plant transformation vectors have been generated inthe last two decades, which have been used in several insertionmutagenesis programs (Bevan 1984; McBride and Summerfelt1990; Düring 1994; Koncz et al. 1994; Fuse et al. 1995; Hellenset al. 2000; Szabados and Koncz 2003; Alvarado et al. 2004;Calderon-Villalobos et al. 2006; Komari et al. 2006; Thole et al.2007).
Establishment of efficient transformation proceduresor identification of transposable elements allows theimplementation of large-scale insertion mutagenesis in plantspecies. Arabidopsis can easily be transformed by dipping orby vacuum infiltration of floral buds with the Agrobacteriumsuspension, carrying the appropriate vector, and subsequentlyselecting the progenies for antibiotic or herbicide resistance,determined by the T-DNA marker gene (Bechtold et al. 1993;Bechtold andPelletier 1998;Bent 2000). Inplanta transformationof Arabidopsis can be scaled up easily, and is suitablefor generating large number of transgenic lines for T-DNA
700 Functional Plant Biology C. Papdi et al.

tagging programs (Parinov and Sundaresan 2000; Sussman et al.2000; Weigel et al. 2000; Bouché and Bouchez 2001; Ríos et al.2002; Szabados et al. 2002;Alonso et al. 2003; Rosso et al. 2003;Koiwa et al. 2006). Among the monocotyledonoues plants,the most efficient Agrobacterium-mediated transformation hasbeen developed for rice (Hiei et al. 1994), leading to large-scaleT-DNA insertion mutagenesis programs (Jeong et al. 2002; Anet al. 2005; Hsing et al. 2007; Wan et al. 2009). In the legumemodel plant Medicago truncatula (L.), the Trt1 retrotransposonhas been used for insertion mutagenesis (Tadege et al. 2005;Benlloch et al. 2006; Tadege et al. 2008).
T-DNA insertions usually lead to loss-of-function, recessivemutations, which show Mendelian segregation among T2
generation plants. In order to isolate mutants with alteredstress responses, forward genetic screens can be employed andT2 generation transgenic plants can be screened eitherindividually or in pooled stocks to identify mutants displayingenhanced or reduced stress tolerance (Alonso and Ecker 2006;Koiwa et al. 2006; Schneider and Leister 2006). Progenies ofselected plants are subsequently tested for co-segregation of theobserved phenotype and the T-DNA insert. Multiple T-DNAinsertions may occur during the transformation procedure, thusthe determination of linkage between a T-DNA insert and amutation require special care. Once linkage of the mutationand the T-DNA insertion is confirmed, T-DNA insertion sitescan be recovered by PCR amplification of the T-DNA junctions.PCR fragments can subsequently be sequenced and the insertionsites can be exactly mapped by sequence homology searches(Earp et al. 1990; Liu et al. 1995; Mathur et al. 1998; Szabadosand Koncz 2003).
In order to identify tagged Arabidopsis, mutants with alteredsalt tolerance, T-DNA insertion lines were generated in differentArabidopsis genetic backgrounds, and large forward geneticscreens of T-DNA mutagenised lines were carried out (Koiwaet al. 2006). Employing a root bending assay, number of saltand osmotic stress sensitive mutants were identified, includingnew alleles of sos1. New mutations have been identified inloci encoding proteins with different cellular functions such assignalling (protein phosphatase, transcription factors), vesiculartrafficking (syntaxin), protein glycosylation and solute transport(Koiwa et al. 2006). Using the RD29A-LUC reporter gene andluciferase imaging screens, several mutants with enhancedluciferase activity were identified, including alleles of cpl. TheArabidopsis CPL genes encodes isoforms of Polymerase IICTD phosphatase, suggesting that CTD phosphorylationcontrols stress signal transduction (Koiwa et al. 2002). Inanother study, mutants with salt tolerance phenotypes wereisolated by screening for enhanced capacity for germinationand growth on high salt medium. In the salt tolerant sto1mutant the insertion inactivated the NCED3-like geneencoding an isoform of 9-cis-epoxicarotenoid dioxygenase, akey enzyme inABAbiosynthesis (Ruggiero et al.2004). The sto1mutant was more tolerant to salt in vitro, but was susceptible fordehydration due to its inability to accumulate ABA. Screeningfor stress hypersensitivity in our T-DNA mutant collection(Szabados et al. 2002) led to the identification of a semi-dwarftagged mutant with enhanced sensitivity to ABA, salt, osmoticand oxidative stress. In the ppr40 mutant, the T-DNA waslocalised in the coding region of the PPR40 gene, which
encodes for a mitochondrial pentatricopeptide repeat (PPR)domain protein, controlling electron flow through Complex IIIin the mitochondrial electron transport chain (Zsigmond et al.2008).
Gene and promoter trapping
Promoter and enhancer trap technologies use promoterlessreporter genes facing the boundaries of T-DNA or transposoninsertions. Promoter trap insertions in transcribed chromosomalloci may generate in situ fusions between plant genes andreporter genes, which can be identified by monitoring theactivity of the reporters (Teeri et al. 1986; Koncz et al. 1989;Kertbundit et al. 1991). Enhancer traps use reporter genes fusedto minimal promoter elements to facilitate the activation ofreporter genes by flanking plant enhancer sequences (Klimyuket al. 1995; Campisi et al. 1999; Ito et al. 2004). Plant gene fusiontechnologies were first demonstrated in tobacco and Arabidopsisusing a promoterless kanamycin phosphotransferase (npt II;Teeri et al. 1986; Koncz et al. 1989) and the b-glucuronidase(GUS; uidA) reporter genes (Jefferson et al. 1987; Kertbunditet al. 1991; Fobert et al. 1994). Using T-DNA or transposoninsertions, gene and promoter trapping programs have been usedto identify tagged genes, promoters and enhancers with tissuespecific activity in crop species such as rice (Chin et al. 1999;Acosta-Garcia et al. 2004; Ito et al. 2004), barley (Lazarow andLutticke 2009). The GUS reporter is suitable for sensitivehistochemical detection and analysis of cell and tissue specificexpression patterns, but is unsuitable for nondestructivescreening purposes, which is essential for the identification ofenvironmentally controlled gene activities. The light-emittingbacterial (LUX) or firefly (LUC) luciferase reporter enzymes canbe detected in vivo by bioluminescence imaging with sensitiveCCD cameras and are better suited as real-time reporters forin planta gene expression studies (Riggs and Chrispeels 1987;Jiang et al. 1992; Millar et al. 1992). The luciferase reportersystem has been used in random promoter and gene trappingexperiments in tomato (Meissner et al. 2000), Arabidopsis(Yamamoto et al. 2003; Alvarado et al. 2004; Calderon-Villalobos et al. 2006) and banana (Musa sp.; Remy et al.2005). Recently a dual reporter system has been developed, inwhich a fused GUS and LUC reporter gene is employed for thedetection of the activity of the trapped gene (Koo et al. 2007). Inthis system, the luciferase allows the sensitive and non-invasivedetection of the gene activity, while GUS activity can bevisualised in histochemical assays to analyse tissue and cellspecific expression patterns. A Gal4-mediated LUC/GFP/GUSreporter system has been used for enhancer trapping inArabidopsis, to identify specific enhancers and to study genesilencing (Engineer et al. 2005).
Using gene or promoter trap technologies, characterisation ofthe mutant phenotype and expression analysis of the tagged genecan be performed simultaneously with the same line. Transposontagging with a DS-GUS gene trap construct has been used toidentify 15Arabidopsisgenes, induced by anoxia (Baxter-Burrellet al. 2003). Several tagged genes were shown to control theactivity of the anoxia-responsive alcohol dehydrogenase (ADH)gene. Two mutants were hypersensitive to low-oxygenenvironment, suggesting that the corresponding genes were
Genetic identification of plant genes in response to stress Functional Plant Biology 701
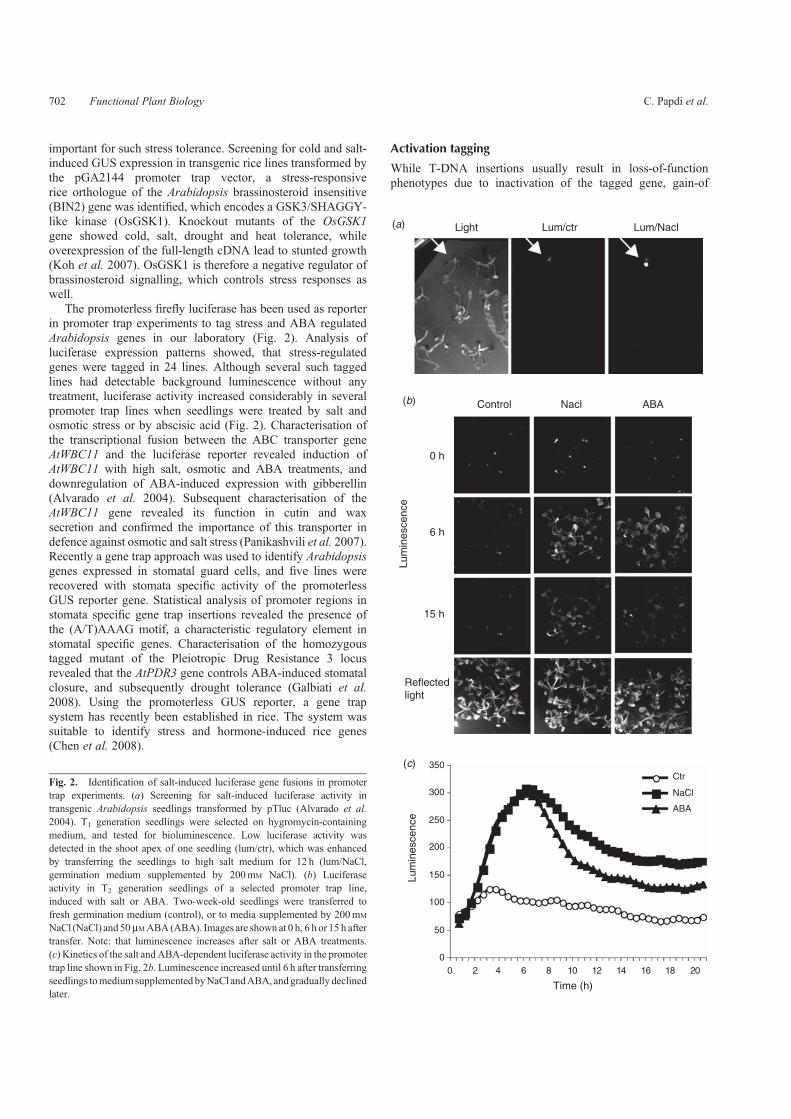
important for such stress tolerance. Screening for cold and salt-induced GUS expression in transgenic rice lines transformed bythe pGA2144 promoter trap vector, a stress-responsiverice orthologue of the Arabidopsis brassinosteroid insensitive(BIN2) gene was identified, which encodes a GSK3/SHAGGY-like kinase (OsGSK1). Knockout mutants of the OsGSK1gene showed cold, salt, drought and heat tolerance, whileoverexpression of the full-length cDNA lead to stunted growth(Koh et al. 2007). OsGSK1 is therefore a negative regulator ofbrassinosteroid signalling, which controls stress responses aswell.
The promoterless firefly luciferase has been used as reporterin promoter trap experiments to tag stress and ABA regulatedArabidopsis genes in our laboratory (Fig. 2). Analysis ofluciferase expression patterns showed, that stress-regulatedgenes were tagged in 24 lines. Although several such taggedlines had detectable background luminescence without anytreatment, luciferase activity increased considerably in severalpromoter trap lines when seedlings were treated by salt andosmotic stress or by abscisic acid (Fig. 2). Characterisation ofthe transcriptional fusion between the ABC transporter geneAtWBC11 and the luciferase reporter revealed induction ofAtWBC11 with high salt, osmotic and ABA treatments, anddownregulation of ABA-induced expression with gibberellin(Alvarado et al. 2004). Subsequent characterisation of theAtWBC11 gene revealed its function in cutin and waxsecretion and confirmed the importance of this transporter indefence against osmotic and salt stress (Panikashvili et al. 2007).Recently a gene trap approach was used to identify Arabidopsisgenes expressed in stomatal guard cells, and five lines wererecovered with stomata specific activity of the promoterlessGUS reporter gene. Statistical analysis of promoter regions instomata specific gene trap insertions revealed the presence ofthe (A/T)AAAG motif, a characteristic regulatory element instomatal specific genes. Characterisation of the homozygoustagged mutant of the Pleiotropic Drug Resistance 3 locusrevealed that the AtPDR3 gene controls ABA-induced stomatalclosure, and subsequently drought tolerance (Galbiati et al.2008). Using the promoterless GUS reporter, a gene trapsystem has recently been established in rice. The system wassuitable to identify stress and hormone-induced rice genes(Chen et al. 2008).
Activation tagging
While T-DNA insertions usually result in loss-of-functionphenotypes due to inactivation of the tagged gene, gain-of
Light
Control
Lum/ctr Lum/Nacl
Nacl ABA
0 h
6 h
15 h
Reflectedlight
350
300
250
200
150
100
50
00 2 4 6 8 10 12 14 16 18 20
Time (h)
Lum
ines
cenc
eLu
min
esce
nce
(a)
(b)
(c)Ctr
NaCl
ABA
Fig. 2. Identification of salt-induced luciferase gene fusions in promotertrap experiments. (a) Screening for salt-induced luciferase activity intransgenic Arabidopsis seedlings transformed by pTluc (Alvarado et al.2004). T1 generation seedlings were selected on hygromycin-containingmedium, and tested for bioluminescence. Low luciferase activity wasdetected in the shoot apex of one seedling (lum/ctr), which was enhancedby transferring the seedlings to high salt medium for 12 h (lum/NaCl,germination medium supplemented by 200mM NaCl). (b) Luciferaseactivity in T2 generation seedlings of a selected promoter trap line,induced with salt or ABA. Two-week-old seedlings were transferred tofresh germination medium (control), or to media supplemented by 200mM
NaCl (NaCl) and50mMABA(ABA). Images are shownat 0 h, 6 h or 15 h aftertransfer. Note: that luminescence increases after salt or ABA treatments.(c) Kinetics of the salt andABA-dependent luciferase activity in the promotertrap line shown in Fig. 2b. Luminescence increased until 6 h after transferringseedlings tomediumsupplemented byNaCl andABA, andgradually declinedlater.
702 Functional Plant Biology C. Papdi et al.

function mutations can be generated by activation of plant geneswith strong transcriptional enhancers or promoters in the insertionelement (Koncz et al. 1994;Weigel et al. 2000; Jeong et al. 2002;Szabados and Koncz 2003; Koiwa et al. 2006; Wan et al. 2009).Activation tagging induces ectopic expression of genes adjacentto the insertion sites, and can be used for generation of dominantmutations that can reveal the function of unique genes as well asindividual members in a gene family (Weigel et al. 2000;Nakazawa et al. 2003). Ectopic expression of stress-inducedregulatory genes such as transcription factors can lead tooverexpression of downstream stress-responsive genes andmay increase tolerance to extreme environmental conditions.Activation tagging with strong promoters is not only capableto activate transcription of flanking genomic sequences but alsocan generate antisense transcripts and therefore can be used forgene suppression, generating loss-of-function type mutations(Ülker et al. 2008). Functional identification of the cytokininreceptor kinase (Kakimoto 1996), and various microRNA genes(Palatnik et al. 2003) illustrates the potential of activationtagging. While most activation tagging has employed T-DNA-derived vectors, transposon-based vector systems have also beencreated for monocots, including cereals (Ayliffe et al. 2007; Quet al. 2008). A large tagged rice population was generated with aT-DNA activation tagging vector, containing double CaMV35Senhancer at the right border (Wan et al. 2009). A combinedpromoter trap/activation tagging T-DNA vector was also used inrice, which contained multiple enhancers of the CaMV35Spromoter, able to activate the genes adjacent to the insertionsite (Jeong et al. 2002). The maize (Zea mays L.) Ac/Dstransposable element system was also employed for activationtagging in rice (Qu et al. 2008) and barley (Ayliffe et al. 2007).
Activation tagging was used to isolate several Arabidopsisgain-of-functionmutantswith enhanced tolerance to high salinityanddrought, leading to the identificationofnumerousnovelgenescontrolling such traits. The Shine (SHN) gene was identified inactivation tagging screens and was shown to control droughttolerance through regulating cuticular wax composition(Aharoni et al. 2004). The SHN gene encodes an AP2/EREBPtype transcription factor, which has an important role inconstructing plant protective layers in cuticle, wounding,abscission and dehiscence. By screening for drought and salttolerance of Arabidopsis activation tagged lines, the sdt1mutantwas isolated and was found to survive lethal levels of salt anddrought stresses (Li et al. 2005). Isolation of dominant mutationsconferring enhanced salt tolerance in direct genetic screensconfirmed the utility of this approach for salt stress research(Koiwa et al. 2006). The edt1 mutant was identified using asimilar approach, andwas shown to exhibit drought tolerance as aresult of the enhanced expression of a homeodomain-STARTtranscription factor (Yu et al. 2008). Similarly, mutants withincreased tolerance to chilling were also identified in tobaccousing activation tagging (Ahad et al. 2003). While isolation ofrecessive mutations in tetraploid species, such as tobacco(Nicotania tabacum L.), is very difficult, activation tagginggenerates dominant mutations, which can be identified easilyin all type of plants regardless of the ploidy level. TheCDT-1geneof Craterostigma plantagineum Hochst. was identified byactivation tagging, screening for desiccation tolerance in calluscultures. Constitutive overexpression of CDT-1 could enhance
desiccation tolerance andwas able to upregulate the expression ofseveral dehydration and ABA-induced transcripts in the absenceof ABA (Furini et al. 1997). CDT-1 was shown to be similar toretroelements and directs the synthesis of 21 bp short interferingRNA (siRNA). Recovery after dehydration in C. plantagineumwas suggested to depend on the capacity to synthesise siRNA(Hilbricht et al. 2008).
Combination of activation tagging with screening forinduction (or repression) of promoter driven luciferasereporters provides a more specific technique to identify and tagregulatory genes. Screening for enhanced luminescence of apathogen responsive PR1-luciferase reporter gene was usedfor tagging of the Arabidopsis ADR1 pathogen resistance gene(Grant et al. 2003). The activated ADR1 gene was later shown toconfer drought tolerance through a salicylic acid-dependentpathway, suggesting that biotic and abiotic signalling canshare common components and have functional overlaps(Chini et al. 2004).
Activation tagging offers a unique strategy for dissection ofstress signalling pathways through mutant suppression screens.The salt hypersensitive sos3–1 mutation inactivates the SOS3gene encoding a CBL-type calcium sensor, which controls theactivity of the SnRK3-type SOS2 protein kinase (Halfter et al.2000; Ishitani et al. 2000; Gong et al. 2004a). Activation taggingwas used to identify suppressor or enhancer mutations of thesos3–1 mutant. Among the 16 lines that suppressed the saltsensitive phenotype of sos3–1, a new mutant allele of theAtHKT1 gene was isolated which is an important Na+
transporter mediating salt influx into plant cells (Koiwa et al.2006).
Complementary DNA library transformation
Transfer and expression of plant cDNA libraries in yeast is afunctional equivalent of multicopy suppressor screens developedto identify yeast genes able to suppress salt sensitivity (Benderand Pringle 1991; Masson and Ramotar 1998).
AnArabidopsis cDNAlibrary in ayeast expressionvectorwasused to identify lithium tolerant yeast transformants and lead tothe identification of SAL1, which participates to maintain salttolerance (Quintero et al. 1996). Suppressor screen of the saltsensitive G19 yeast mutant with a rice full-length cDNAexpression library lead to the identification of Shaker familyK+ channel KAT1 (OsKAT1) gene, which participates inmaintaining cation homeostasis in rice during salt stress(Obata et al. 2007). Several Arabidopsis genes that couldenhance antioxidant defences and improve tolerance tooxidative stress were identified using Arabidopsis cDNAlibrary transfer to yeast and subsequent selection for diamide(Kushnir et al. 1995).
Transfer and expression of cDNA libraries in plants providea novel strategy to screen for gain-of-function phenotypes.An Arabidopsis cDNA library driven by the constitutiveCaMV35S promoter has been used to generate transgenicArabidopsis lines, showing altered developmental traits(LeClere and Bartel 2001). The Full-length cDNA Over-eXpressing gene hunting system (FOX) uses a normalised full-length cDNA collection under the control of the constitutiveCaMV35S promoter, and offers a technique to identify novel
Genetic identification of plant genes in response to stress Functional Plant Biology 703

gain-of-function phenotypes in Arabidopsis (Ichikawa et al.2006). A similar FOX-type cDNA overexpression system isavailable for rice (Nakamura et al. 2007). These cDNAlibraries were used in different genetic screens to identifyseveral important regulatory factors in hormonal and stresssignalling. Large-scale transformation of Arabidopsis rootswith a cDNA library lead to the identification of the ESR1gene, encoding a putative AP2/EREBP transcription factor,overexpression of which stimulates cytokinin-independentplant regeneration (Banno et al. 2001). By screening acollection of cDNA overexpressing plants for ABAindependent germination, Kuhn et al. (2006) identified theAtPP2AC gene, encoding an unknown protein phosphatase,which plays an important regulatory role in ABA-controlledclosure of stomata and thereby controls drought tolerance(Kuhn et al. 2006). A modified version of the FOX system hasbeen used to screen for salt tolerant phenotypes, leading to theidentification of several lines with enhanced tolerance to highsalinity. One of the identified lines overexpressed the bZIP-typetranscription factor AtbZIP60 which was subsequently shown tocontrol the expression of at least 29 Arabidopsis genes andimprove salt tolerance through modulating stress signaltransduction (Fujita et al. 2007).
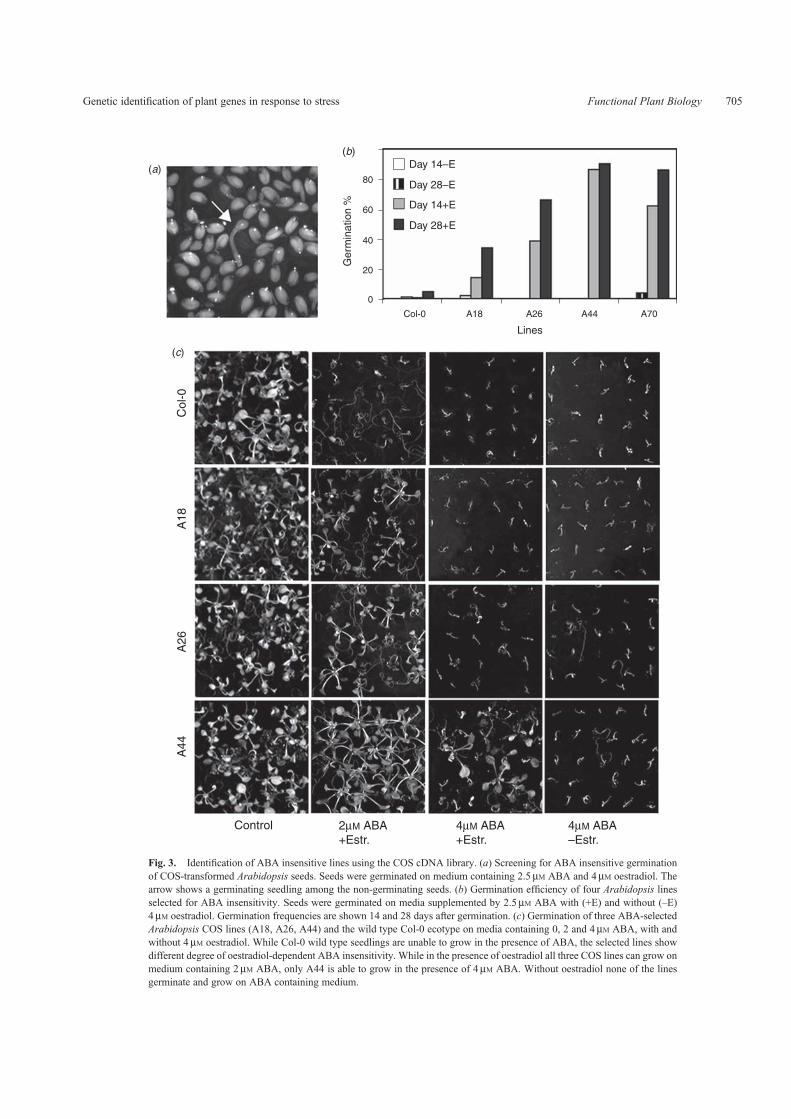
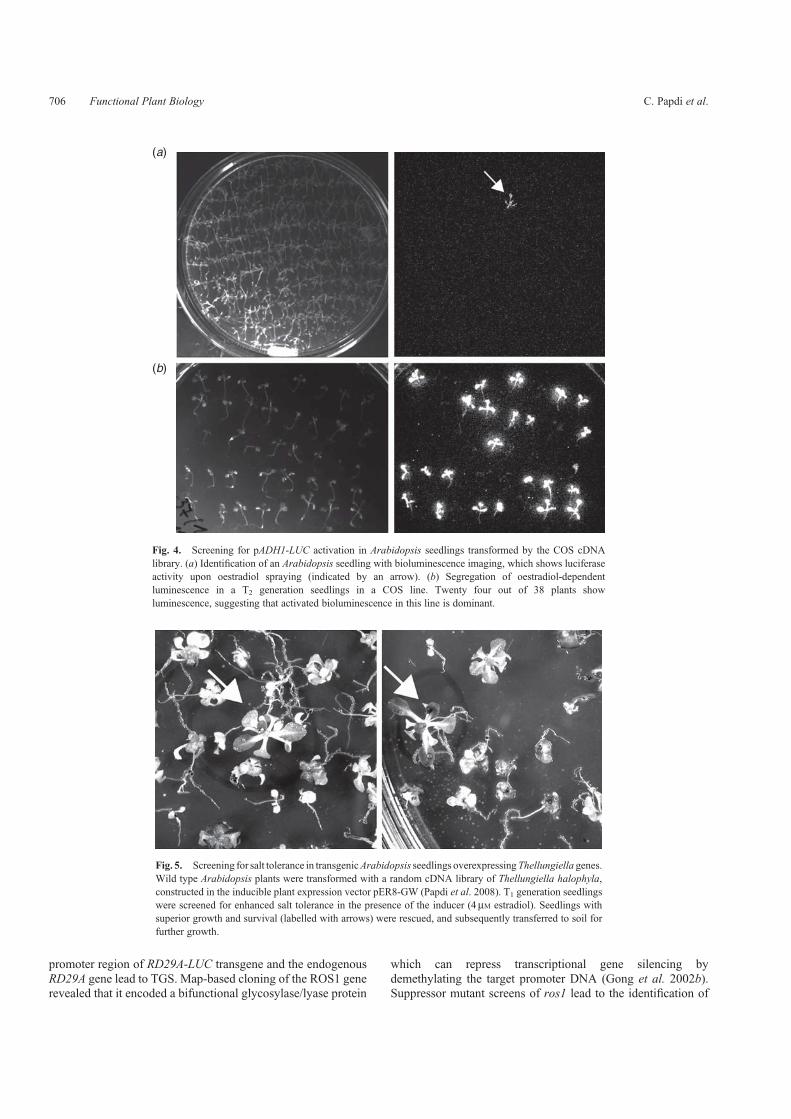
The Controlled cDNA Overexpression System (COS) wascreated in our laboratory to overcome the problems causedby the constitutive gene activation. High level and constitutiveexpression of stress regulatory genes can disturb cell proliferationand development resulting in dwarf and sterile plants(Kasuga et al. 1999; Gilmour et al. 2000). The COS librarywas created in a Gateway version of the oestradiol-induciblepER8 plasmid, in which transcription of the inserted cDNAis strictly controlled by the addition of the inducer (Zuo et al.2000). The COS system was used to identify a set of cDNAsconferring dominant stress-tolerance phenotypes in threedifferent genetic screens: tolerance to salt stress, insensitivitytoABAand activation of the stress-inducedADH1-LUC reportergene (Papdi et al.2008). Screening forABA insensitivity resultedin the identification of 26 transgenic lines, which showedoestradiol-dependent germination and growth on culturemedium supplemented by inhibitory concentration of ABA(Fig. 3). Complementary DNA inserts were recovered andidentified in selected lines. We showed that overexpressionof HSP17.6 cDNA conferred ABA insensitivity whereasactivation of the 2-alkenal reductase 2AER resulted inimproved salt tolerance. The ADH1-LUC reporter gene wasused in combination with the COS activation to identify noveltranscriptional regulators of the ADH1 gene. Bioluminescencewas detected in several Arabidopsis lines when seedlings weretreated with oestradiol in the absence of any stress or hormoneeffect (Fig. 4). In line ADH121, oestradiol-dependent activationof the ERF/AP2-type transcription factor RAP2.12 sustainedhigh-level ADH1-LUC bioluminescence, and increased ADHenzyme activity in the roots (Papdi et al.2008). The cDNA librarytransformation approach usually creates dominant, gain-of-function phenotypes, but may also produce dominant loss-of-function phenotypes, which result from co-suppression ofendogenous genes by overexpression of truncated or antisensecDNAs (LeClere and Bartel 2001). An important advantageof the cDNA library transformation is that gene identification
in a selected line is extremely fast. A cDNA insert can beamplified by PCR from genomic DNA templates usingprimers that anneal to the vector T-DNA sequences flankingthe cDNA cloning site. The isolated cDNA is subsequentlysequenced and the identity of the insert is determined byhomology searches of sequence databases. In the COS systemeach cDNA was flanked by attB1 and attB2 recombination sites,therefore PCR fragments could easily be recloned in aGATEWAY entry vector and subsequently transferred intoany destination vector for recurrent transformation and othermolecular and biochemical analysis (Fig. 6; Papdi et al. 2008).
Complementary DNA library transformation is not restrictedto interspecific experimentation, but can also be used to transforma cDNA to a plant species different from the RNA source. In suchscenario cDNAs from any plant species, including naturalvariants of drought, salt or cold tolerant plant species can betransferred and tested in Arabidopsis or other model species.Ectopic expression of rice full length cDNA library (Rice FOX)in Arabidopsis offers a genetic resource to identify and analyserice genes in a heterologous system (Kondou et al. 2009).Overexpression of the rice cDNA library in Arabidopsis andsubsequent screen for thermotolerance lead to the identificationof the NAC type transcription factor ONAC063, which couldimprove tolerance to high temperature, salinity and osmoticstress (Yokotani et al. 2009). Overexpression of ONAC063 inArabidopsis upregulated several stress-responsive genes,suggesting that NAC-type transcription factors are importantfor stress defences and have conserved function in differentplant species. Complementary DNA expression libraries wererecently created from salt cres (Thellungiella halophylaC.A. Meyer.), a salt tolerant relative of Arabidopsis, and weresubsequently used to generate transgenic Arabidopsis lines.Screening for salt tolerance of transgenic Arabidopsisseedlings expressing Thellungiella cDNAs, resulted in theidentification of several lines showing enhanced salt tolerance(I. Pérez-Salamó and L. Szabados, unpubl. data; Fig. 5).Identification of several novel small protein coding genes,which could confer salt tolerance to Arabidopsis, confirmedthe potential of this approach (Du et al. 2008). Using atargeted interspecific gene transfer strategy, it is possible toidentify natural sequence variations in regulatory genes thatcan confer tolerance to different biotic or abiotic stresses.
Epigenetic regulation of stress responses
Gene silencing is an important regulatory mechanism thatcontrols gene activity at the transcriptional and posttranscriptional level (Baulcombe 2004). Post transcriptionalgene silencing is regulated by micro RNAs (miRNA) andsmall interfering RNA (siRNA), which are generated byenzymes of the Dicer family and processed with the RNA-induced silencing complex (RISC) (Tang 2005). siRNA of the24 nt size class control transcriptional gene silencing throughDNA and histone methylation (Matzke and Birchler 2005).Transcriptional gene silencing (TGS) has recently beenidentified as an important regulator of stress responses. Therepressor of silencing 1 and 3 (ros1 and ros3) mutants wereisolated by screening for suppressed activity of the RD29A-LUCreporter. In the ros1 mutant DNA hypermethylation in the
704 Functional Plant Biology C. Papdi et al.
80
60
40
20
0
Col-0 A18 A26 A44 A70
Lines
Ger
min
atio
n %
Day 14–E
Day 28–E
Day 14+E
Day 28+E
Control
A44
A26
A18
Col
-0
2μM ABA+Estr.
4μM ABA+Estr.
4μM ABA–Estr.
(a)
(b)
(c)
Fig. 3. Identification of ABA insensitive lines using the COS cDNA library. (a) Screening for ABA insensitive germinationof COS-transformed Arabidopsis seeds. Seeds were germinated on medium containing 2.5mM ABA and 4mM oestradiol. Thearrow shows a germinating seedling among the non-germinating seeds. (b) Germination efficiency of four Arabidopsis linesselected for ABA insensitivity. Seeds were germinated on media supplemented by 2.5mM ABA with (+E) and without (–E)4mM oestradiol. Germination frequencies are shown 14 and 28 days after germination. (c) Germination of three ABA-selectedArabidopsis COS lines (A18, A26, A44) and the wild type Col-0 ecotype on media containing 0, 2 and 4mM ABA, with andwithout 4mM oestradiol. While Col-0 wild type seedlings are unable to grow in the presence of ABA, the selected lines showdifferent degree of oestradiol-dependent ABA insensitivity. While in the presence of oestradiol all three COS lines can grow onmedium containing 2mM ABA, only A44 is able to grow in the presence of 4mM ABA. Without oestradiol none of the linesgerminate and grow on ABA containing medium.
Genetic identification of plant genes in response to stress Functional Plant Biology 705
promoter region of RD29A-LUC transgene and the endogenousRD29A gene lead to TGS. Map-based cloning of the ROS1 generevealed that it encoded a bifunctional glycosylase/lyase protein
which can repress transcriptional gene silencing bydemethylating the target promoter DNA (Gong et al. 2002b).Suppressor mutant screens of ros1 lead to the identification of
(a)
(b)
Fig. 4. Screening for pADH1-LUC activation in Arabidopsis seedlings transformed by the COS cDNAlibrary. (a) Identification of an Arabidopsis seedling with bioluminescence imaging, which shows luciferaseactivity upon oestradiol spraying (indicated by an arrow). (b) Segregation of oestradiol-dependentluminescence in a T2 generation seedlings in a COS line. Twenty four out of 38 plants showluminescence, suggesting that activated bioluminescence in this line is dominant.
Fig. 5. Screening for salt tolerance in transgenicArabidopsis seedlings overexpressingThellungiella genes.Wild type Arabidopsis plants were transformed with a random cDNA library of Thellungiella halophyla,constructed in the inducible plant expression vector pER8-GW (Papdi et al. 2008). T1 generation seedlingswere screened for enhanced salt tolerance in the presence of the inducer (4mM estradiol). Seedlings withsuperior growth and survival (labelled with arrows) were rescued, and subsequently transferred to soil forfurther growth.
706 Functional Plant Biology C. Papdi et al.
rpa2 and ago6–1mutants, on base of enhanced bioluminescenceof RD29A-LUC reporter gene. The ros1 ago6–1 double mutantshowed reduced DNA methylation and transcriptional genesilencing of RD29A (Zheng et al. 2007), while in theros1rpa2 double mutant silencing of RD29A-LUC was notaffected (Kapoor et al. 2005). The ros3 mutation causesconsiderable reduction of RD29A-LUC activity by promotingTGS through hypermethylation of the RD29A promoter regions.The ROS3 protein has RNA recognition motif (RRM) and canbind small RNA which may direct demethylation of targetpromoter sequences (Zheng et al. 2008). Genetic analysisshowed, that ROS1 and ROS3 act in the same pathway tocontrol DNA methylation and transcriptional gene silencing.Histone acetylation in chromatin is important to defineeukaryotic gene activity. Genetic screen to identify regulatorsofRD29A-LUC reporter gene lead to the identification ofHOS15,aWD40-type regulator of histone acetylation.HOS15was shown
to interact with and reduce histone H4 acetylation while analysisof the hos15mutant showed that thewild type gene is essential forcold tolerance (Zhu et al. 2008).
Recent studies show that gene silencing, mediated bymiRNAor siRNA can play an important role in controlling abiotic stressresponses. Stress-regulated miRNA and siRNA were identifiedin different plant species including Arabidopsis (Sunkar and Zhu2004; Zhou et al. 2008), rice (Zhao et al. 2007, 2009), maize(Zhang et al. 2008),Phaseolus (Arenas-Huertero et al. 2009) andPopulus spp. (Lu et al. 2005, 2008). miRNA-mediated genesilencing can control cold tolerance (Lu Sun et al. 2008; Zhouet al. 2008), phosphate starvation (Fujii et al. 2005; Bari et al.2006), heat stress (Lu et al. 2008), UV-B responses (Zhou et al.2007), drought and salt tolerance (Arenas-Huertero et al. 2009;Zhao et al. 2009), adaptation to anaerobic conditions (Zhanget al. 2008), and to mechanical stresses (Lu et al. 2008). TheArabidopsis CSD1 andCSD2 genes are encoding the detoxifying
Fig.6. Flowchart of theCOScDNAlibrary transformationandscreening strategies to identify stressregulatory functions in Arabidopsis.
Genetic identification of plant genes in response to stress Functional Plant Biology 707

superoxide dismutase (SOD) enzymes and were shown to beregulated by miR398, which repress the activity of CSD genes inthe absence of stress (Sunkar et al. 2006). Post transcriptionalgene silencingwas shown to control the activity of a NF-YA typetranscription factor which regulates the expression of numerousstress-related genes (Zhao et al. 2009). The involvement ofseveral class of microRNA in control of ABA-mediated stressresponses was recently confirmed in Arabidopsis and rice(Han et al. 2004; Jung and Kang 2007; Reyes and Chua 2007;Zhao et al. 2007), showing that miRNAs and epigeneticregulation is implicated in modulating ABA signalling.Endogenous siRNA was shown to control the expression ofP5C dehydrogenase gene (P5CDH), which regulates prolinedegradation in mitochondria, suggesting that small RNA canregulate metabolic responses to environmental stresses (Borsaniet al. 2005). Genetic screens for mutants deficient in phosphatenutrition or activation of the Pi starvation induced reporter gene(AtIPS1-GUS) lead to the identification of miR399, whichcontrols the activity of Pi-responsive genes and phosphatemetabolism (Fujii et al. 2005; Bari et al. 2006; Chiou et al.2006). Small RNA can therefore function as master regulators ofstress responses by silencing a well defined group of stress-responsive and defence genes. Stress-induced small RNAs canrepress the activity of target genes, while small RNAs which aredownregulated during stress may enhance the activity of a groupof stress-induced genes (Shukla et al. 2008).
Reverse genetics
In forward genetics, a mutant is isolated first on base of aphenotype, and the corresponding gene is assigned when themutated locus is identified either by map-based cloning or bymapping the insertion site. In reverse genetic approachesmutantsare identified or created for selected genes or gene families inorder to understand their function. Reverse genetics is a typicalanalytical tool for the post-genomic era, when information onwhole genome sequence is available and precise gene predictionsallow the systematic analysis of all genes in a genome.
Allele replacement by homologous recombination isconsidered as ideal tool for targeted modification of endogenousgenes (Mortensen 2006). Gene targeting with homologousrecombination is a method of choice to engineer mouse genes(Evans et al. 2001), but has limited success in plants. Lowfrequency of recombination into target sequences, too manyrandom integration events, non-homologous end joining, lack ofgene specific selection system prevent this technique fromwidespread use in higher plants (Puchta 2002; Hanin andPaszkowski 2003; Reiss 2003). Gene targeting via homologousrecombination have been used for engineering Arabidopsis plantsfor herbicide resistance (Hanin et al. 2001). Engineering of theacetolactate synthase (ALS) gene by introducing two amino acidchanges by gene targeting resulted in herbicide tolerant rice(Endo et al. 2007). Modification of the stress-induced ADH2gene by gene targeting have been reported in rice when strongpositive selection was applied (Terada et al. 2007; Johzuka-Hisatomi et al. 2008). However, despite sporadic success,genome modification by homologous recombination is not aroutine technology for higher plants, and further technologicalimprovements are needed for widespread use for reverse
genetics (Iida and Terada 2005; Johzuka-Hisatomi et al. 2008).Physcomitrella patens is a moss which exhibits high frequency ofhomologous recombination and ismore suitable for gene targetingthan higher plants (Schaefer and Zryd 1997; Kamisugi et al. 2005,2006). The haploid dominant stage of the plant life cycle facilitatesthephenotypiccharacterisationof theresultingmutants.Thusallelereplacement and directed mutagenesis is more accessible for thisspecies.Genome sequence ofPhyscomitrella is available (Rensinget al. 2008), making this plant an ideal model for reverse geneticapproaches. Targeted gene disruption in Physcomitrella involvesthe generation of transgenic lines that have replaced a specificgenomic locus by a targeting construct consisting of a selectioncassette flanked by two stretches of the genomic sequence. Theefficiency of allele replacement has been shown to be dependenton the length and the symmetry of the genomic sequencespresent in the construct (Kamisugi et al. 2005, 2006; Quatranoet al. 2007). When these parameters are taken into account, it ispossible to achieve levels of gene targeting comparable withthose shown in Saccharomyces cereviceae (Schaefer 2001).
Physcomitrella is also known for high degree of toleranceto different abiotic stresses such as high salinity, dehydrationand osmotic stress (Kroemer et al. 2004; Frank et al. 2005).This plant survives dehydration to water content as low as 2%(Oldenhof et al. 2006), and is able to rapidly recover fromprolonged periods of high salinity (Saavedra et al. 2006).Mosses often exhibit desiccation tolerance of vegetativetissues, in contrast with higher plants, in which this trait isusually restricted to reproductive structures such as pollen andseeds (Oliver 1996). Desiccation tolerant plants have protectivemechanisms that limit and repair stress-induced tissue damageand allow them to survive and recover from anhydrobiosis(Alpert and Oliver 2002). The functional analysis ofPhyscomitrella genes involved in stress responses may lead tothe identification of candidate genes for engineering stresstolerance in crops, as well as helping to address fundamentalquestions in plant biology, such as the function and conservationof plant molecular mechanisms involved in abiotic stresstolerance. In this context, the study of PP2C-dependentsignalling in Arabidopsis and Physcomitrella suggestedevolutionarily conserved ABA signalling pathways betweenmosses and higher plants (Komatsu et al. 2009). Knockoutmutants of the dehydrin gene were engineered by genetargeting and used to reveal the importance of the dehydrins inosmotic stress tolerance (Saavedra et al. 2006). As dehydrins areencoded by multigene families in higher plants, similargenetic strategies could not be implemented for the functionalanalysis of these genes in Arabidopsis or other plants.
The genome sequence of Arabidopsis has been available since2000 (The Arabidopsis Genome Initiative 2000b). Although genetargeting cannot be routinely used inArabidopsis, alternative toolsare available for reverse genetic analysis. Large-scale T-DNA andtransposon insertion mutagenesis programs lead to the applicationof reverse genetic strategies for different model species such asArabidopsis, rice andM. truncatula. One possibility is the creationof pooled genomic DNA templates from T-DNA and transposoninsertion mutant collections which serve as PCR templates toscreen for insertions in known genes (Martienssen 1998;Sussman et al. 2000; Mahalingam and Fedoroff 2001; Ríos et al.2002). Mutations are identified by PCR screening of the DNA
708 Functional Plant Biology C. Papdi et al.

templates with combination of gene and T-DNA border specificprimers. A collection of 90 000 Arabidopsis lines (consisting of~116000 T-DNA inserts) was shown to be suitable for detectinga mutation in any gene with 77% probability (Ríos et al. 2002).Saturation mutagenesis and large-scale mapping of T-DNAinsertion sites on the genome allows the easy identificationof mutations for any genes. Insertions are localised throughinternet-based databases and mutant lines are obtained frompublic strain collections. Such resources are most abundantfor Arabidopsis, making it possible to identify mutations fornearly all of Arabidopsis genes (Samson et al. 2002; Szabadoset al. 2002; Alonso et al. 2003; Rosso et al. 2003). Among themonocotyledonous plants large collections of indexed T-DNA ortransposon insertion mutants have been established for rice(An et al. 2005; van Enckevort et al. 2005; Chern et al. 2007;Hsing et al. 2007; Piffanelli et al. 2007). The rice reverse geneticsdatabase OryGenesDB (http://orygenesdb.cirad.fr/index.html,verified 7 July 2009) has been established to identify flankingsequence tags (FSTs) for each rice genes and to support ricefunctional genomics programs (Droc et al. 2006, 2009). TheOryza Tag Line mutant database (http://urgi.versailles.inra.fr/OryzaTagLine/, verified 7 July 2009) contains phenotypic dataof30 000T-DNAenhancer trap lines,complementedbyGUS/GFPexpression data (Larmande et al. 2008). Such genetic resourcesallow the application of systematic reverse genetic screens toidentify mutants for specific traits, including stress responses(Alonso and Ecker 2006; Ülker et al. 2008).
Identification of putative stress responsive genes is possible byanalysis of global datasets on stress-dependent gene expression(Kant Gordon et al. 2008). Large scale transcript profilingexperiments showed, that expression of thousands ofArabidopsis genes is modified during different environmentalstresses, suggesting that these genes are involved in stressresponses (Chen et al. 2002; Seki et al. 2002; Oono et al.2003; Hannah et al. 2005; Vanderauwera et al. 2005; Gadjevet al. 2006; Robinson and Parkin 2008). Functional analysis ofstress-regulated Arabidopsis and rice genes is therefore possibleby reverse genetic approaches where identification ofmutants formost genes is feasible.
Asystematic useof a reverse genetic strategyhas recentlybeendescribed identifying and analysing a large set of Arabidopsisgenes, which regulate responses to multiple abiotic stresses.Multiple Stress Regulatory genes (MSTR) have been firstidentified by bioinformatics analysis of microarray data.Insertion mutants were subsequently identified for the MSTRgenes and their stress sensitivity and function in regulatingstress responses was analysed. Among the 16 identified MSTRgenes, the highest score was assigned to the Circadian ClockAssociated 1 (CCA1) transcription factor, which has a structuralhomologue, Late ElongatedHypocotyl (LHY), previously knownto be part of circadian oscillator (Schaffer et al. 1998; Wang andTobin 1998). Functional analysis of cca1 and lhymutants showedthat the double mutant has greater sensitivity to differentenvironmental stresses than single mutants or wild type plants,
Fig.7. Simplifieddepictionofgenetic strategiesused to identifyandcharacterise stressgenes inplants. Important steps are shown ineachprocedure, highlightingthe most critical, time-consuming or bottleneck step. Numerous modifications of the core technologies exist and adaptations to particular plants, conditions orpurposes have been published.
Genetic identification of plant genes in response to stress Functional Plant Biology 709

suggesting that the CCA1 and LHY genes control not onlycircadian clockbut also abiotic stress responses (Kant et al.2008).
Reverse genetic strategies can also be based on chemical orphysical mutagenesis as well as on natural variability. As analternative to sequence-indexed insertion collections, Targeting-induced local lesions in genomes (TILLING) has been adoptedin various plant species to identify point mutations and singlenucleotide polymorphism (SNP) in defined genes (Greene et al.2003; Till et al. 2003; Henikoff et al. 2004; Barkley and Wang2008). For TILLING, gene fragments from mutated plants areamplified with primers labelled by fluorescent dyes, and theproducts are denatured and annealed to form heteroduplexes.These molecules are in turn recognised and cleaved by the CelIendonuclease and the resulting DNA fragments are analysed ondenaturing polyacrylamide genes (Till et al. 2006b). A high-throughput TILLING program has been established for theArabidopsis community which can identify series of pointmutations in any Arabidopsis gene (Till et al. 2003, 2006a).As this technology does not use transgenic plants, it can be veryattractive to screen for point mutations in crop plants. TILLINGhas been used to identify mutants in maize (Henikoff et al. 2004;Till et al. 2004), melon (Cucumis melo L.; Nieto et al. 2007),mungbean (Vigna radiate L.; Barkley et al. 2008) and rice(Till Cooper et al. 2007; Suzuki et al. 2008) among otherspecies. Moreover, the TILLING approach has been used fordetection of natural variation in numerous plant species(ecotilling) (Comai et al. 2004; Henikoff et al. 2004; Gilchristet al. 2006). For example, ecotilling was employed for theidentification of allelic variants of melon elF4E factor, whichcontrols susceptibility to biotic stresses such as virus (Nieto et al.2007). TILLING has also been useful to identify mutant allelesin stress-related Arabidopsis genes. Using this approach,hot5 alleles were isolated and used to study heat acclimationin Arabidopsis (Lee et al. 2008). Similarly, swi3b mutantsshowed ABA insensitive germination and reduced expressionof ABA induced genes, suggesting that SWI3B in the wildtype plant functions as positive regulator of ABA signalling(Saez et al. 2008).
Reverse genetic strategies are particularly useful for theanalysis of multigenic families, where forward genetic screensdo not result in recognisable phenotypes due to functionalredundancy. Mutants can be identified for each member of agene family in sequence indexed insertion line collections or byTILLING. Double and multiple mutant lines can subsequentlybe generated by crossing the single mutants, and tested forphenotypic alterations. Numerous examples show, that doublemutants but not single mutants of a gene family can produce aphenotype. The SNF1-Related Protein Kinase 2 gene familyconsist of 10 member genes in Arabidopsis (Hrabak et al.2003), which are implicated in stress and ABA signaltransduction (Boudsocq et al. 2004). Single knockout mutantsof SnRK2.2 and SnRK2.3 genes had no recognisable phenotype,while the snrk2.2/snrk2.3 double mutant showed strong ABAinsensitivity. Altered seed dormancy, proline accumulation,water loss in detached leaves and reduced expression ofnumerous stress and ABA-induced genes in the double mutantsuggested that the SnRK2.2 and SnRK2.3 genes are majorregulators of ABA signalling and control drought tolerance(Fujii et al. 2007). Due to their functional redundancy, forward
screens for ABA insensitivity could not have recovered singlesnrk2.2 or snrk2.3 mutants.
Conclusion
Dissections of regulatory mechanisms controlling responses toenvironmental stresses or functional analysis of genes which areimplicated in defence mechanisms require genetic analysis.Variability can be generated by chemical, physical, insertionmutagenesis and gene transfer or derived from naturalpopulations. Identification of genetic variability which affectstress responses requires phenotypic screens which are able todistinguish between plants with differences in stress toleranceor reverse genetic technologies, which can identify mutants ingiven genes. All these strategies and technologies have certainadvantages and inconveniences, which have to be taken intoconsideration (Fig. 7).
Chemical and physical mutagenesis is easy and can generatelarge numbers of single nucleotide changes or deletions withminimal effort. Forward genetic screens require larger effortwhich ultimately leads to the isolation of either loss or gain offunction mutants, depending on the type of mutation. Majoreffort is required for the mapping and cloning of the genethat is affected by the mutation. Except for the Arabidopsismodel, map-based cloning is a challenging program formost plant species. Advanced microarray technologies andnew generation sequencing technologies can improve theefficiency of positional cloning in many species.
Natural variability offers a large resource of polymorphism,which is often explored to identify traits with adaptationvalue. Natural variability is often based on minor geneticchanges, generating small quantitative alterations in responsesto environmental conditions. However, QTL mapping andcloning is a time-consuming effort, and identification of thepolymorphism at molecular level is tedious.
Insertion mutagenesis facilitates easy identification oftagged mutants through PCR amplification and mapping ofinsertion sites. Insertion mutagenesis is a real alternative inplants which have suitable transposable elements or efficienttransformation systems. The generation of tens of thousands ofT-DNA insertions requires large-scale genetic transformation,which is feasible for Arabidopsis but is a bottleneck for mostplant species, where in planta transformation methods arenot available. Endogenous transposons or the maize Ac/Dssystem offer an alternative for insertion mutagenesis. Insertionmutagenesis most often generates loss-of function mutants, withthe exception of activation tagging, which can generate gain-offunction phenotypes through the ectopic activation of the genesthat are flanking the insertion element.
Variability can be generated by random cDNAtransformation, which creates mainly gain-of functionphenotypes based on ectopic (or controlled) expression. Wheninterspecific cDNA transfer is used, genetic polymorphism ofspecific plant species can be explored to identify genes withadaptive values. Creation of the cDNA library is critical step,while establishment of the transformed plant population is a time-consuming procedure, making this approach generally lessattractive. On the other hand, gene identification and genecloning from selected plants is easy and straight forward, as
710 Functional Plant Biology C. Papdi et al.

the inserts are flanked by known vector sequences, facilitatingPCR amplification of the cDNA.
Variability for known genes can be identified by reversegenetic approaches. Gene targeting through homologousrecombination is routinely available for Physcomitrella but notfor higher plants. Nevertheless, several reverse genetictechnologies are available for several species using traditionalor insertionmutagenesis. Searching the sequence-tagged T-DNAdatabases is the most common procedure to find Arabidopsismutants. Although services of a similar scale are not availablefor other plants, smaller collections have been established forsome species. TILLING, identification of InDels and insertionsby PCR offer an opportunity to identify mutants in numerousplants, including crops. Most of these strategies rely on theestablishment of large, characterised mutant collections whichrequires manpower and extensive funding. Once the mutantplatforms (T-DNA insertion database, TILLING collection)exist, mutant identification is easy and fast.
Research inplant stress biologyhas used all these technologiesextensively and numerous examples are available from bothmodel and crop species. Genetic analysis has become anessential part of the functional characterisation of genescontrolling stress responses and made it possible to dissectcomplex regulatory pathways. The genetic strategies reviewedhave been used and will be used to identify and characterisemaster genes which can improve tolerance of crop plants toharmful environmental conditions such as salinity, drought orextreme temperatures (see Table S1).
Acknowledgements
We are indebted to Paul Lazzeri, Csaba Koncz and László Kozma Bognár forcareful reading and corrections. This work was supported by OTKA grantno. T-46552 and K-68226, EU grant no. FP6–020232–2, ICGEB-TWASProgram CRP.PB/URU06–01. Csaba Papdi and Mary Prathiba Joseph weresupported by fellowships of the Biological Research Centre.
References
Acosta-GarciaG,AutranD,Vielle-Calzada JP (2004)Enhancer detection andgene trapping as tools for functional genomics in plants. Methods inMolecular Biology (Clifton, N.J.) 267, 397–414.
Ahad A, Wolf J, Nick P (2003) Activation-tagged tobacco mutants thatare tolerant to antimicrotubular herbicides are cross-resistant to chillingstress. Transgenic Research 12, 615–629. doi: 10.1023/A:1025814814823
Aharoni A, Dixit S, Jetter R, Thoenes E, van Arkel G, Pereira A (2004) TheSHINE clade of AP2 domain transcription factors activates waxbiosynthesis, alters cuticle properties, and confers drought tolerancewhen overexpressed in Arabidopsis. The Plant Cell 16, 2463–2480.doi: 10.1105/tpc.104.022897
Alonso JM, Ecker JR (2006)Moving forward in reverse: genetic technologiesto enable genome-wide phenomic screens in Arabidopsis. NatureReviews. Genetics 7, 524–536. doi: 10.1038/nrg1893
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, et al. (2003)Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science301, 653–657. doi: 10.1126/science.1086391
Alonso-Blanco C, Gomez-Mena C, Llorente F, Koornneef M, Salinas J,Martinez-Zapater JM (2005) Genetic and molecular analyses of naturalvariation indicate CBF2 as a candidate gene for underlying a freezingtolerance quantitative trait locus in Arabidopsis. Plant Physiology 139,1304–1312. doi: 10.1104/pp.105.068510
AlpertP,OliverMJ (2002)Dryingwithoutdying. In ‘Desiccationand survivalin plants’. (Eds M Black, HW Pritchard) pp. 3–43. (CABI Publishing:Wallingford, UK)
Alvarado MC, Zsigmond LM, Kovacs I, Cseplo A, Koncz C, Szabados LM(2004) Gene trapping with firefly luciferase in Arabidopsis. Tagging ofstress-responsive genes. Plant Physiology 134, 18–27. doi: 10.1104/pp.103.027151
An G, Jeong DH, Jung KH, Lee S (2005) Reverse genetic approaches forfunctional genomics of rice. Plant Molecular Biology 59, 111–123.doi: 10.1007/s11103-004-4037-y
Ansorge WJ (2009) Next-generation DNA sequencing techniques. NewBiotechnology 25, 195–203. doi: 10.1016/j.nbt.2008.12.009
Arenas-Huertero C, Perez B, Rabanal F, Blanco-Melo D, De la Rosa C,Estrada-Navarrete G, Sanchez F, Covarrubias AA, Reyes JL (2009)Conserved and novel miRNAs in the legume Phaseolus vulgaris inresponse to stress. Plant Molecular Biology 70, 385–401. doi: 10.1007/s11103-009-9480-3
Ayliffe MA, Pallotta M, Langridge P, Pryor AJ (2007) A barley activationtagging system. Plant Molecular Biology 64, 329–347. doi: 10.1007/s11103-007-9157-8
Azpiroz-Leehan R, Feldmann KA (1997) T-DNA insertion mutagenesis inArabidopsis: going back and forth. Trends in Genetics 13, 152–156.doi: 10.1016/S0168-9525(97)01094-9
Bako L, Umeda M, Tiburcio AF, Schell J, Koncz C (2003) The VirD2 pilotprotein ofAgrobacterium-transferred DNA interacts with the TATAbox-binding protein and a nuclear protein kinase in plants. Proceedings of theNational Academy of Sciences of the United States of America 100,10 108–10 113. doi: 10.1073/pnas.1733208100
Bancroft I, Bhatt AM, Sjodin C, Scofield S, Jones JD, Dean C (1992)Development of an efficient two-element transposon tagging system inArabidopsis thaliana. Molecular & General Genetics 233, 449–461.doi: 10.1007/BF00265443
BannoH, IkedaY,NiuQW,ChuaNH (2001)Overexpression ofArabidopsisESR1 induces initiation of shoot regeneration. The Plant Cell 13,2609–2618.
Bari R, Datt Pant B, Stitt M, Scheible WR (2006) PHO2, microRNA399,and PHR1 define a phosphate-signaling pathway in plants. PlantPhysiology 141, 988–999. doi: 10.1104/pp.106.079707
Barkley NA, Wang ML (2008) Application of TILLING and EcoTILLINGas reverse genetic approaches to elucidate the function of genes in plantsand animals. Current Genomics 9, 212–226. doi: 10.2174/138920208784533656
Barkley NA, Wang ML, Gillaspie AG, Dean RE, Pederson GA, Jenkins TM(2008) Discovering and verifying DNA polymorphisms in a mung bean[V. radiata (L.) R. Wilczek] collection by EcoTILLING and sequencing.BMC Research Notes 1, 28. doi: 10.1186/1756-0500-1-28
Bartels D, Sunkar R (2005) Drought and salt tolerance in plants. CriticalReviews in Plant Sciences 24, 23–58. doi: 10.1080/07352680590910410
Batelli G, Verslues PE, Agius F, Qiu Q, Fujii H, Pan S, Schumaker KS,Grillo S, Zhu JK (2007) SOS2 promotes salt tolerance in part byinteracting with the vacuolar H+-ATPase and upregulating itstransport activity. Molecular and Cellular Biology 27, 7781–7790.doi: 10.1128/MCB.00430-07
Baulcombe D (2004) RNA silencing in plants. Nature 431, 356–363.doi: 10.1038/nature02874
Baxter-Burrell A, Chang R, Springer P, Bailey-Serres J (2003) Gene andenhancer trap transposable elements reveal oxygen deprivation-regulatedgenes and their complex patterns of expression in Arabidopsis. Annalsof Botany 91, 129–141. doi: 10.1093/aob/mcf119
Bechtold N, Pelletier G (1998) In planta Agrobacterium-mediatedtransformation of adult Arabidopsis thaliana plants by vacuuminfiltration. Methods in Molecular Biology (Clifton, N.J.) 82, 259–266.
Bechtold N, Ellis J, Pelletier G (1993) In planta Agrobacterium mediatedgene transfer by infiltration of adult Arabidopsis thaliana plants. LesComptes Rendus de L’Académie des Sciences 316, 1194–1199.
Genetic identification of plant genes in response to stress Functional Plant Biology 711

Bender A, Pringle JR (1991) Use of a screen for synthetic lethal andmulticopy suppressee mutants to identify two new genes involved inmorphogenesis in Saccharomyces cerevisiae. Molecular and CellularBiology 11, 1295–1305.
Benlloch R, d’Erfurth I, Ferrandiz C, Cosson V, Beltran JP, Canas LA,Kondorosi A, Madueno F, Ratet P (2006) Isolation of mtpim provesTnt1 a useful reverse genetics tool inMedicago truncatula and uncoversnew aspects of AP1-like functions in legumes. Plant Physiology 142,972–983. doi: 10.1104/pp.106.083543
BentAF (2000)Arabidopsis in planta transformation.Uses,mechanisms, andprospects for transformation of other species. Plant Physiology 124,1540–1547. doi: 10.1104/pp.124.4.1540
Bevan M (1984) Binary Agrobacterium vectors for plant transformation.Nucleic Acids Research 12, 8711–8721. doi: 10.1093/nar/12.22.8711
Bohnert HJ, Gong Q, Li P, Ma S (2006) Unraveling abiotic stress tolerancemechanisms – getting genomics going.Current Opinion in Plant Biology9, 180–188. doi: 10.1016/j.pbi.2006.01.003
Borevitz JO, Liang D, Plouffe D, Chang HS, Zhu T, Weigel D, Berry CC,Winzeler E, Chory J (2003) Large-scale identification of single-featurepolymorphisms in complex genomes. Genome Research 13, 513–523.doi: 10.1101/gr.541303
Borsani O, Zhu J, Verslues PE, Sunkar R, Zhu JK (2005) EndogenoussiRNAs derived from a pair of natural cis-antisense transcripts regulatesalt tolerance in Arabidopsis. Cell 123, 1279–1291. doi: 10.1016/j.cell.2005.11.035
Bouché N, Bouchez D (2001) Arabidopsis gene knockout: phenotypeswanted. Current Opinion in Plant Biology 4, 111–117. doi: 10.1016/S1369-5266(00)00145-X
Boudsocq M, Barbier-Brygoo H, Lauriere C (2004) Identification of ninesucrose nonfermenting 1-related protein kinases 2 activated byhyperosmotic and saline stresses in Arabidopsis thaliana. The Journalof Biological Chemistry 279, 41 758–41 766. doi: 10.1074/jbc.M405259200
Boyer JS (1982) Plant productivity and environment. Science 218, 443–448.doi: 10.1126/science.218.4571.443
Byrt CS, Platten JD, Spielmeyer W, James RA, Lagudah ES, Dennis ES,TesterM,Munns R (2007) HKT1;5-like cation transporters linked to Na+
exclusion loci in wheat, Nax2 and Kna1. Plant Physiology 143,1918–1928. doi: 10.1104/pp.106.093476
Calderon-Villalobos LI, Kuhnle C, Li H, Rosso M, Weisshaar B,Schwechheimer C (2006) LucTrap vectors are tools to generateluciferase fusions for the quantification of transcript and proteinabundance in vivo. Plant Physiology 141, 3–14. doi: 10.1104/pp.106.078097
Campisi L, Yang Y, Yi Y, Heilig E, Herman B, Cassista AJ, Allen DW,Xiang H, Jack T (1999) Generation of enhancer trap lines in Arabidopsisand characterization of expression patterns in the inflorescence.The Plant Journal 17, 699–707. doi: 10.1046/j.1365-313X.1999.00409.x
CaplanA,Herrera-Estrella L, InzeD,VanHaute E,VanMontaguM, Schell J,Zambryski P (1983) Introduction of genetic material into plant cells.Science 222, 815–821. doi: 10.1126/science.222.4625.815
Chen W, Provart NJ, Glazebrooka J, Katagiria F, Changa HS, et al. (2002)Expression profile matrix of Arabidopsis transcription factor genessuggests their putative functions in response to environmental stresses.The Plant Cell 14, 559–574. doi: 10.1105/tpc.010410
Chen SY, Wang AM, Li W, Wang ZY, Cai XL (2008) Establishing a genetrap system mediated by T-DNA(GUS) in rice. Journal of IntegrativePlant Biology 50, 742–751. doi: 10.1111/j.1744-7909.2007.00611.x
ChengNH, Pittman JK, Zhu JK, Hirschi KD (2004) The protein kinase SOS2activates the Arabidopsis H+/Ca2+ antiporter CAX1 to integrate calciumtransport and salt tolerance. The Journal of Biological Chemistry 279,2922–2926. doi: 10.1074/jbc.M309084200
Chern CG, FanMJ, Yu SM, Hour AL, Lu PC, et al. (2007) A rice phenomicsstudy – phenotype scoring and seed propagation of a T-DNA insertion-induced rice mutant population. Plant Molecular Biology 65, 427–438.doi: 10.1007/s11103-007-9218-z
Chin HG, Choe MS, Lee SH, Park SH, Park SH, et al. (1999) Molecularanalysis of rice plants harboring anAc/Ds transposable element-mediatedgene trapping system. The Plant Journal 19, 615–623. doi: 10.1046/j.1365-313X.1999.00561.x
Chini A, Grant JJ, Seki M, Shinozaki K, Loake GJ (2004) Drought toleranceestablished by enhanced expression of the CC-NBS-LRR gene, ADR1,requires salicylic acid, EDS1 and ABI1. The Plant Journal 38, 810–822.doi: 10.1111/j.1365-313X.2004.02086.x
Chinnusamy V, Stevenson B, Lee BH, Zhu JK (2002) Screening for generegulation mutants by bioluminescence imaging. Science’s STKE 2002,1–10. doi: 10.1126/stke.2002.140.pl10
Chinnusamy V, Zhu J, Zhu JK (2006) Salt stress signaling and mechanismsof plant salt tolerance. Genetic Engineering 27, 141–177. doi: 10.1007/0-387-25856-6_9
Chiou TJ, Aung K, Lin SI, Wu CC, Chiang SF, Su CL (2006) Regulation ofphosphate homeostasis by microRNA in Arabidopsis. The Plant Cell 18,412–421. doi: 10.1105/tpc.105.038943
Cho RJ, Mindrinos M, Richards DR, Sapolsky RJ, AndersonM, et al. (1999)Genome-wide mapping with biallelic markers in Arabidopsis thaliana.Nature Genetics 23, 203–207. doi: 10.1038/13833
Collins NC, Tardieu F, Tuberosa R (2008) Quantitative trait loci and cropperformance under abiotic stress: where do we stand? Plant Physiology147, 469–486. doi: 10.1104/pp.108.118117
Comai L, Young K, Till BJ, Reynolds SH, Greene EA, et al. (2004) Efficientdiscovery of DNA polymorphisms in natural populations by Ecotilling.The Plant Journal 37, 778–786. doi: 10.1111/j.0960-7412.2003.01999.x
Cutler S, Ghassemian M, Bonetta D, Cooney S, McCourt P (1996) A proteinfarnesyl transferase involved in abscisic acid signal transduction inArabidopsis. Science 273, 1239–1241. doi: 10.1126/science.273.5279.1239
DeSmet I, ZhangH, InzeD,BeeckmanT (2006)Anovel role for abscisic acidemerges from underground. Trends in Plant Science 11, 434–439.doi: 10.1016/j.tplants.2006.07.003
Dilbirligi M, Erayman M, Campbell BT, Randhawa HS, Baenziger PS,Dweikat I, Gill KS (2006) High-density mapping and comparativeanalysis of agronomically important traits on wheat chromosome 3A.Genomics 88, 74–87. doi: 10.1016/j.ygeno.2006.02.001
Draye X, Lin YR, Qian XY, Bowers JE, Burow GB, et al. (2001)Toward integration of comparative genetic, physical, diversity, andcytomolecular maps for grasses and grains, using the sorghum genomeas a foundation. Plant Physiology 125, 1325–1341. doi: 10.1104/pp.125.3.1325
Droc G, Ruiz M, Larmande P, Pereira A, Piffanelli P, Morel JB, Dievart A,Courtois B, Guiderdoni E, Périn C (2006) OryGenesDB: a database forrice reverse genetics. Nucleic Acids Research 34, D736–D740.doi: 10.1093/nar/gkj012
Droc G, Perin C, Fromentin S, Larmande P (2009) OryGenesDB 2008update: database interoperability for functional genomics of rice.Nucleic Acids Research 37, D992–D995. doi: 10.1093/nar/gkn821
Du J, HuangYP,Xi J, CaoMJ, NiWS, ChenX, Zhu JK,Oliver DJ, XiangCB(2008) Functional gene-mining for salt-tolerance genes with the powerof Arabidopsis. The Plant Journal 56, 653–664. doi: 10.1111/j.1365-313X.2008.03602.x
Düring K (1994) A plant transformation vector with a minimal T-DNA.Transgenic Research 3, 138–140. doi: 10.1007/BF01974093
Earp DJ, Lowe B, Baker B (1990) Amplification of genomic sequencesflanking transposable elements in host and heterologous plants: a tool fortransposon tagging and genome characterization.Nucleic Acids Research18, 3271–3279. doi: 10.1093/nar/18.11.3271
712 Functional Plant Biology C. Papdi et al.

Endo M, Osakabe K, Ono K, Handa H, Shimizu T, Toki S (2007)Molecular breeding of a novel herbicide-tolerant rice by genetargeting. The Plant Journal 52, 157–166. doi: 10.1111/j.1365-313X.2007.03230.x
Engineer CB, Fitzsimmons KC, Schmuke JJ, Dotson SB, Kranz RG (2005)Development and evaluation of a Gal4-mediated LUC/GFP/GUSenhancer trap system in Arabidopsis. BMC Plant Biology 5, 9.doi: 10.1186/1471-2229-5-9
Etterson JR, Shaw RG (2001) Constraint to adaptive evolution in responseto global warming. Science 294, 151–154. doi: 10.1126/science.1063656
Evans MJ, Smithies O, Capecchi MR (2001) Mouse gene targeting. NatureMedicine 7, 1081–1083. doi: 10.1038/nm1001-1081
Feldmann KA, Marks MD, Christianson ML, Quatrano RS (1989) A dwarfmutant of Arabidopsis generated by T-DNA insertion mutagenesis.Science 243, 1351–1354. doi: 10.1126/science.243.4896.1351
Finkelstein RR, Lynch TJ (2000) The Arabidopsis abscisic acid responsegene ABI5 encodes a basic leucine zipper transcription factor. The PlantCell 12, 599–609.
Finkelstein RR, Wang ML, Lynch TJ, Rao S, Goodman HM (1998)The Arabidopsis abscisic acid response locus ABI4 encodes anAPETALA 2 domain protein. The Plant Cell 10, 1043–1054.
Finkelstein RR, Gampala SS, Rock CD (2002) Abscisic acid signaling inseeds and seedlings. The Plant Cell 14(Suppl.), S15–S45.
FlowersTJ,KoyamaML,FlowersSA,SudhakarC,SinghKP,YeoAR(2000)QTL: their place in engineering tolerance of rice to salinity. Journalof Experimental Botany 51, 99–106. doi: 10.1093/jexbot/51.342.99
Fobert PR,LabbeH,Cosmopoulos J,Gottlob-McHughS,Ouellet T,Hattori J,Sunohara G, Iyer VN, Miki BL (1994) T-DNA tagging of a seed coat-specific cryptic promoter in tobacco. The Plant Journal 6, 567–577.doi: 10.1046/j.1365-313X.1994.6040567.x
Frank W, Ratnadewi D, Reski R (2005) Physcomitrella patens is highlytolerant against drought, salt and osmotic stress. Planta 220, 384–394.doi: 10.1007/s00425-004-1351-1
Fujii H, Chiou TJ, Lin SI, Aung K, Zhu JK (2005) A miRNA involved inphosphate-starvation response in Arabidopsis. Current Biology 15,2038–2043. doi: 10.1016/j.cub.2005.10.016
Fujii H, Verslues PE, Zhu JK (2007) Identification of two protein kinasesrequired for abscisic acid regulation of seed germination, root growth,and gene expression in Arabidopsis. The Plant Cell 19, 485–494.doi: 10.1105/tpc.106.048538
FujitaM,MizukadoS, FujitaY, IchikawaT,NakazawaM,SekiM,MatsuiM,Yamaguchi-Shinozaki K, Shinozaki K (2007) Identification of stress-tolerance-related transcription-factor genes via mini-scale Full-lengthcDNA Over-eXpressor (FOX) gene hunting system. Biochemical andBiophysical Research Communications 364, 250–257. doi: 10.1016/j.bbrc.2007.09.124
Furini A, Koncz C, Salamini F, Bartels D (1997) High level transcription ofa member of a repeated gene family confers dehydration tolerance tocallus tissue of Craterostigma plantagineum. The EMBO Journal 16,3599–3608. doi: 10.1093/emboj/16.12.3599
Fuse T, Kodama H, Hayashida N, Shinozaki K, Nishimura M, Iba K (1995)A novel Ti-plasmid-convertible lambda phage vector system suitable forgene isolation by genetic complementation of Arabidopsis thalianamutants. The Plant Journal 7, 849–856. doi: 10.1046/j.1365-313X.1995.07050849.x
Gadjev I, Vanderauwera S, Gechev TS, Laloi C, Minkov IN, Shulaev V,Apel K, Inzé D, Mittler R, Van Breusegem F (2006) Transcriptomicfootprints disclose specificity of reactive oxygen species signaling inArabidopsis. Plant Physiology 141, 436–445. doi: 10.1104/pp.106.078717
Galbiati M, Simoni L, Pavesi G, Cominelli E, Francia P, Vavasseur A,Nelson T, Bevan M, Tonelli C (2008) Gene trap lines identifyArabidopsis genes expressed in stomatal guard cells. The PlantJournal 53, 750–762. doi: 10.1111/j.1365-313X.2007.03371.x
Gilchrist EJ, Haughn GW, Ying CC, Otto SP, Zhuang J, et al. (2006)Use of Ecotilling as an efficient SNP discovery tool to survey geneticvariation in wild populations of Populus trichocarpa.Molecular Ecology15, 1367–1378. doi: 10.1111/j.1365-294X.2006.02885.x
Gilmour SJ, Sebolt AM, Salazar MP, Everard JD, Thomashow MF (2000)Overexpression of theArabidopsisCBF3 transcriptional activatormimicsmultiple biochemical changes associated with cold acclimation. PlantPhysiology 124, 1854–1865. doi: 10.1104/pp.124.4.1854
Giraudat J, Hauge BM, Valon C, Smalle J, Parcy F, Goodman HM (1992)Isolation of the Arabidopsis ABI3 gene by positional cloning. The PlantCell 4, 1251–1261.
GodiardL,SauviacL,ToriiKU,GrenonO,ManginB,GrimsleyNH,MarcoY(2003) ERECTA, an LRR receptor-like kinase protein controllingdevelopment pleiotropically affects resistance to bacterial wilt. ThePlant Journal 36, 353–365. doi: 10.1046/j.1365-313X.2003.01877.x
GongD,GuoY, JagendorfAT, Zhu JK (2002a) Biochemical characterizationof the Arabidopsis protein kinase SOS2 that functions in salt tolerance.Plant Physiology 130, 256–264. doi: 10.1104/pp.004507
Gong Z, Morales-Ruiz T, Ariza RR, Roldan-Arjona T, David L, Zhu JK(2002b) ROS1, a repressor of transcriptional gene silencing inArabidopsis, encodes a DNA glycosylase/lyase. Cell 111, 803–814.doi: 10.1016/S0092-8674(02)01133-9
GongD,GuoY, SchumakerKS,Zhu JK (2004a) TheSOS3 family of calciumsensors and SOS2 family of protein kinases in Arabidopsis. PlantPhysiology 134, 919–926. doi: 10.1104/pp.103.037440
Gong JM,WanerDA,HorieT,Li SL,HorieR,AbidKB,Schroeder JI (2004b)Microarray-based rapid cloning of an ion accumulation deletionmutant in Arabidopsis thaliana. Proceedings of the National Academyof Sciences of the United States of America 101, 15 404–15 409.doi: 10.1073/pnas.0404780101
Grant JJ, Chini A, Basu D, Loake GJ (2003) Targeted activation tagging ofthe Arabidopsis NBS-LRR gene, ADR1, conveys resistance to virulentpathogens. Molecular Plant-Microbe Interactions 16, 669–680.doi: 10.1094/MPMI.2003.16.8.669
Greene EA, Codomo CA, Taylor NE, Henikoff JG, Till BJ, et al. (2003)Spectrum of chemically induced mutations from a large-scale reverse-genetic screen in Arabidopsis. Genetics 164, 731–740.
Gupta M, Qiu X, Wang L, Xie W, Zhang C, Xiong L, Lian X, Zhang Q(2008) KT/HAK/KUP potassium transporters gene family and theirwhole-life cycle expression profile in rice (Oryza sativa). MolecularGenetics and Genomics 280, 437–452. doi: 10.1007/s00438-008-0377-7
Halfter U, Ishitani M, Zhu JK (2000) The Arabidopsis SOS2 protein kinasephysically interacts with and is activated by the calcium-binding proteinSOS3. Proceedings of the National Academy of Sciences of the UnitedStates of America 97, 3735–3740. doi: 10.1073/pnas.040577697
Han MH, Goud S, Song L, Fedoroff N (2004) The Arabidopsis double-strandedRNA-binding proteinHYL1plays a role inmicroRNA-mediatedgene regulation. Proceedings of the National Academy of Sciences ofthe United States of America 101, 1093–1098. doi: 10.1073/pnas.0307969100
Hanin M, Paszkowski J (2003) Plant genome modification by homologousrecombination. Current Opinion in Plant Biology 6, 157–162.doi: 10.1016/S1369-5266(03)00016-5
HaninM,VolrathS,BoguckiA,BrikerM,WardE,Paszkowski J (2001)Genetargeting in Arabidopsis. The Plant Journal 28, 671–677. doi: 10.1046/j.1365-313x.2001.01183.x
Hannah MA, Heyer AG, Hincha DK (2005) A global survey of generegulation during cold acclimation in Arabidopsis thaliana. PLOSGenetics 1, e26. doi: 10.1371/journal.pgen.0010026
Hansen G, Chilton MD (1999) Lessons in gene transfer to plants by a giftedmicrobe. Current Topics in Microbiology and Immunology 240, 21–57.
HarushimaY, YanoM, ShomuraA, SatoM, Shimano T, et al. (1998) A high-density rice genetic linkage map with 2275 markers using a single F2population. Genetics 148, 479–494.
Genetic identification of plant genes in response to stress Functional Plant Biology 713

Hazen SP, Borevitz JO, Harmon FG, Pruneda-Paz JL, Schultz TF,Yanovsky MJ, Liljegren SJ, Ecker JR, Kay SA (2005) Rapid arraymapping of circadian clock and developmental mutations inArabidopsis. Plant Physiology 138, 990–997. doi: 10.1104/pp.105.061408
Heiber I, Stroher E, Raatz B, Busse I, Kahmann U, Bevan MW, Dietz KJ,Baier M (2007) The redox imbalanced mutants of Arabidopsisdifferentiate signaling pathways for redox regulation of chloroplastantioxidant enzymes. Plant Physiology 143, 1774–1788. doi: 10.1104/pp.106.093328
Hellens RP, Edwards EA, Leyland NR, Bean S, Mullineaux PM (2000)pGreen: a versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Molecular Biology 42, 819–832.doi: 10.1023/A:1006496308160
HenikoffS,TillBJ,ComaiL (2004)TILLING.Traditionalmutagenesismeetsfunctional genomics. Plant Physiology 135, 630–636. doi: 10.1104/pp.104.041061
Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformationof rice (Oryza sativa L.) mediated by Agrobacterium and sequenceanalysis of the boundaries of the T-DNA. The Plant Journal 6,271–282. doi: 10.1046/j.1365-313X.1994.6020271.x
Hilbricht T, Varotto S, Sgaramella V, Bartels D, Salamini F, Furini A(2008) Retrotransposons and siRNA have a role in the evolution ofdesiccation tolerance leading to resurrection of the plant Craterostigmaplantagineum. The New Phytologist 179, 877–887. doi: 10.1111/j.1469-8137.2008. 02480.x
Himmelbach A, Iten M, Grill E (1998) Signalling of abscisic acid to regulateplant growth. Philosophical Transactions of the Royal Society ofLondon. Series B, Biological Sciences 353, 1439–1444. doi: 10.1098/rstb.1998.0299
Hori K, Kobayashi T, Shimizu A, Sato K, Takeda K, Kawasaki S (2003)Efficient construction of high-density linkage map and its application toQTL analysis in barley. Theoretical and Applied Genetics 107, 806–813.doi: 10.1007/s00122-003-1342-9
Hrabak EM, Chan CW, Gribskov M, Harper JF, Choi JH, et al. (2003)The Arabidopsis CDPK-SnRK superfamily of protein kinases. PlantPhysiology 132, 666–680. doi: 10.1104/pp.102.011999
Hsing YI, Chern CG, Fan MJ, Lu PC, Chen KT, et al. (2007) A rice geneactivation/knockout mutant resource for high throughput functionalgenomics. Plant Molecular Biology 63, 351–364. doi: 10.1007/s11103-006-9093-z
Huang S, Spielmeyer W, Lagudah ES, James RA, Platten JD, Dennis ES,Munns R (2006) A sodium transporter (HKT7) is a candidate for Nax1,a gene for salt tolerance in durum wheat. Plant Physiology 142,1718–1727. doi: 10.1104/pp.106.088864
HuangS,SpielmeyerW,LagudahES,MunnsR (2008)Comparativemappingof HKT genes in wheat, barley, and rice, key determinants of Na+
transport, and salt tolerance. Journal of Experimental Botany 59,927–937. doi: 10.1093/jxb/ern033
Hugouvieux V, Kwak JM, Schroeder JI (2001) An mRNA cap bindingprotein, ABH1, modulates early abscisic acid signal transduction inArabidopsis. Cell 106, 477–487. doi: 10.1016/S0092-8674(01)00460-3
Ichikawa T, Nakazawa M, Kawashima M, Iizumi H, Kuroda H, et al. (2006)The FOX hunting system: an alternative gain-of-function gene huntingtechnique. The Plant Journal 48, 974–985. doi: 10.1111/j.1365-313X.2006.02924.x
Iida S, Terada R (2005) Modification of endogenous natural genes by genetargeting in rice and other higher plants. Plant Molecular Biology 59,205–219. doi: 10.1007/s11103-005-2162-x
IshitaniM, Xiong L, StevensonB, Zhu JK (1997)Genetic analysis of osmoticand cold stress signal transduction in Arabidopsis: interactions andconvergence of abscisic acid-dependent and abscisic acid-independentpathways. The Plant Cell 9, 1935–1949.
Ishitani M, Xiong L, Lee H, Stevenson B, Zhu JK (1998) HOS1, a geneticlocus involved in cold-responsive gene expression in Arabidopsis.The Plant Cell 10, 1151–1161.
Ishitani M, Liu J, Halfter U, Kim CS, Shi W, Zhu JK (2000) SOS3 functionin plant salt tolerance requires N-myristoylation and calcium binding.The Plant Cell 12, 1667–1678.
Ito Y, Eiguchi M, Kurata N (2004) Establishment of an enhancer trap systemwithDs andGUS for functional genomics in rice.MolecularGenetics andGenomics 271, 639–650. doi: 10.1007/s00438-004-1023-7
James RA, Davenport RJ, Munns R (2006) Physiological characterization oftwo genes for Na+ exclusion in durum wheat, Nax1 and Nax2. PlantPhysiology 142, 1537–1547. doi: 10.1104/pp.106.086538
Jander G (2006) Gene identification and cloning by molecular markermapping. Methods in Molecular Biology (Clifton, N.J.) 323, 115–126.
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higherplants. The EMBO Journal 6, 3901–3907.
Jeong DH, An S, Kang HG, Moon S, Han JJ, Park S, Lee HS, An K, An G(2002) T-DNA insertional mutagenesis for activation tagging in rice.Plant Physiology 130, 1636–1644. doi: 10.1104/pp.014357
Jiang C, Langridge WHR, Szalay AA (1992) Identification of genes in vivoby tagging with T-DNA border linked luciferase genes followed byinverse polymerase chain reaction amplification. Plant MolecularBiology Reporter 10, 345–361. doi: 10.1007/BF02668911
Johzuka-Hisatomi Y, Terada R, Iida S (2008) Efficient transfer of basechanges from a vector to the rice genome by homologousrecombination: involvement of heteroduplex formation and mismatchcorrection. Nucleic Acids Research 36, 4727–4735. doi: 10.1093/nar/gkn451
JungHJ,KangH(2007)Expression and functional analysesofmicroRNA417in Arabidopsis thaliana under stress conditions. Plant Physiology andBiochemistry 45, 805–811. doi: 10.1016/j.plaphy.2007.07.015
Kakimoto T (1996) CKI1, a histidine kinase homolog implicated in cytokininsignal transduction. Science 274, 982–985. doi: 10.1126/science.274.5289.982
Kamisugi Y, Cuming AC, Cove DJ (2005) Parameters determining theefficiency of gene targeting in the moss Physcomitrella patens. NucleicAcids Research 33, e173. doi: 10.1093/nar/gni172
Kamisugi Y, Schlink K, Rensing SA, Schween G, von Stackelberg M,Cuming AC, Reski R, Cove DJ (2006) The mechanism of genetargeting in Physcomitrella patens: homologous recombination,concatenation and multiple integration. Nucleic Acids Research 34,6205–6214. doi: 10.1093/nar/gkl832
Kant P, Gordon M, Kant S, Zolla G, Davydov O, Heimer YM,Chalifa-Caspi V, Shaked R, Barak S (2008) Functional-genomics-based identification of genes that regulate Arabidopsis responses tomultiple abiotic stresses. Plant, Cell & Environment 31, 697–714.doi: 10.1111/j.1365-3040.2008.01779.x
Kapoor A, Agarwal M, Andreucci A, Zheng X, Gong Z, Hasegawa PM,Bressan RA, Zhu JK (2005) Mutations in a conserved replicationprotein suppress transcriptional gene silencing in a DNA-methylation-independent manner in Arabidopsis. Current Biology 15, 1912–1918.doi: 10.1016/j.cub.2005.09.013
Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, Shinozaki K (1999)Improving plant drought, salt, and freezing tolerance by gene transfer ofa single stress-inducible transcription factor. Nature Biotechnology 17,287–291. doi: 10.1038/7036
Kertbundit S, De Greve H, Deboeck F, Van Montagu M, Hernalsteens JP(1991) In vivo random beta-glucuronidase gene fusions in Arabidopsisthaliana. Proceedings of the National Academy of Sciences of the UnitedStates of America 88, 5212–5216. doi: 10.1073/pnas.88.12.5212
Kim Y, Schumaker KS, Zhu JK (2006) EMS mutagenesis of Arabidopsis.Methods in Molecular Biology (Clifton, N.J.) 323, 101–103.
714 Functional Plant Biology C. Papdi et al.

Kliebenstein D (2009) Quantitative genomics: analyzing intraspecificvariation using global gene expression polymorphisms or eQTLs.Annual Review of Plant Biology 60, 93–114. doi: 10.1146/annurev.arplant.043008.092114
Klimyuk VI, Nussaume L, Harrison K, Jones JD (1995) Novel GUSexpression patterns following transposition of an enhancer trap Dselement in Arabidopsis. Molecular & General Genetics 249, 357–365.doi: 10.1007/BF00287097
Knight H (1999) Calcium signaling during abiotic stress in plants.International Review of Cytology 195, 269–324. doi: 10.1016/S0074-7696(08)62707-2
Knox AK, Li C, Vagujfalvi A, Galiba G, Stockinger EJ, Dubcovsky J(2008) Identification of candidate CBF genes for the frost tolerancelocus Fr-Am2 in Triticum monococcum. Plant Molecular Biology 67,257–270. doi: 10.1007/s11103-008-9316-6
Koh S, Lee SC, Kim MK, Koh JH, Lee S, An G, Choe S, Kim SR (2007)T-DNA tagged knockout mutation of rice OsGSK1, an orthologue ofArabidopsis BIN2, with enhanced tolerance to various abiotic stresses.Plant Molecular Biology 65, 453–466. doi: 10.1007/s11103-007-9213-4
Koiwa H, Barb AW, Xiong L, Li F, McCully MG, et al. (2002) C-terminaldomain phosphatase-like family members (AtCPLs) differentiallyregulate Arabidopsis thaliana abiotic stress signaling, growth, anddevelopment. Proceedings of the National Academy of Sciences ofthe United States of America 99, 10 893–10 898. doi: 10.1073/pnas.112276199
Koiwa H, Bressan RA, Hasegawa PM (2006) Identification of plant stress-responsive determinants in Arabidopsis by large-scale forward geneticscreens. Journal of Experimental Botany 57, 1119–1128. doi: 10.1093/jxb/erj093
KomariT,TakakuraY,Ueki J,KatoN, IshidaY,HieiY (2006)Binaryvectorsand super-binary vectors. Methods in Molecular Biology (Clifton, N.J.)343, 15–41.
Komatsu K, Nishikawa Y, Ohtsuka T, Taji T, Quatrano RS, Tanaka S,Sakata Y (2009) Functional analyses of the ABI1-related proteinphosphatase type 2C reveal evolutionarily conserved regulation ofabscisic acid signaling between Arabidopsis and the mossPhyscomitrella patens. Plant Molecular Biology 70, 327–340.doi: 10.1007/s11103-009-9476-z
Koncz C, Martini N, Mayerhofer R, Koncz-Kalman Z, Korber H, Redei GP,Schell J (1989) High-frequency T-DNA-mediated gene tagging in plants.Proceedings of the National Academy of Sciences of the United Statesof America 86, 8467–8471. doi: 10.1073/pnas.86.21.8467
Koncz C, Nemeth K, Redei GP, Schell J (1992) T-DNA insertionalmutagenesis in Arabidopsis. Plant Molecular Biology 20, 963–976.doi: 10.1007/BF00027166
Koncz C, Martini N, Szabados L, Hrouda M, Bachmair A, Schell J (1994)Specialized vectors for gene tagging and expression studies. In ‘Plantmolecular biology manual’. (Ed. SB Gelvin) pp. 1–22.
Kondou Y, Higuchi M, Takahashi S, Sakurai T, Ichikawa T, et al. (2009)Systematic approaches to using the FOX hunting system to identifyuseful rice genes. The Plant Journal 57, 883–894. doi: 10.1111/j.1365-313X.2008.03733.x
Koo J, Kim Y, Kim J, Yeom M, Lee IC, Nam HG (2007) A GUS/luciferasefusion reporter for plant gene trapping and for assay of promoter activitywith luciferin-dependent control of the reporter protein stability. Plant& Cell Physiology 48, 1121–1131. doi: 10.1093/pcp/pcm081
Koornneef M, Reuling G, Karssen CM (1984) The isolation andcharacterization of abscisic acid-insensitive mutants of Arabidopsisthaliana. Physiologia Plantarum 61, 377–383. doi: 10.1111/j.1399-3054.1984.tb06343.x
Koornneef M, Alonso-Blanco C, Vreugdenhil D (2004) Naturally occurringgenetic variation in Arabidopsis thaliana. Annual Review of PlantBiology 55, 141–172. doi: 10.1146/annurev.arplant.55.031903.141605
Kroemer K, Reski R, Frank W (2004) Abiotic stress response in the mossPhyscomitrella patens: evidence for an evolutionary alteration insignaling pathways in land plants. Plant Cell Reports 22, 864–870.doi: 10.1007/s00299-004-0785-z
Kuhn JM, Boisson-Dernier A, Dizon MB, Maktabi MH, Schroeder JI(2006) The protein phosphatase AtPP2CA negatively regulates abscisicacid signal transduction in Arabidopsis, and effects of abh1 onAtPP2CA mRNA. Plant Physiology 140, 127–139. doi: 10.1104/pp.105.070318
Kushnir S, Babiychuk E, Kampfenkel K, Belles-Boix E, Van Montagu M,Inze D (1995) Characterization of Arabidopsis thaliana cDNAs thatrender yeasts tolerant toward the thiol-oxidizing drug diamide.Proceedings of the National Academy of Sciences of the United Statesof America 92, 10 580–10 584. doi: 10.1073/pnas.92.23.10580
Larmande P, Gay C, LorieuxM, Périn C, Bouniol M, et al. (2008)Oryza TagLine, a phenotypic mutant database for the Génoplante rice insertion linelibrary. Nucleic Acids Research 36, D1022–D1027. doi: 10.1093/nar/gkm762
Lazarow K, Lutticke S (2009) An Ac/Ds-mediated gene trap system forfunctional genomics in barley. BMC Genomics 10, 55. doi: 10.1186/1471-2164-10-55
Le MQ, Engelsberger WR, Hincha DK (2008) Natural genetic variation inacclimation capacity at sub-zero temperatures after cold acclimation at4�C in different Arabidopsis thaliana accessions. Cryobiology 57,104–112. doi: 10.1016/j.cryobiol.2008.06.004
LeClere S, Bartel B (2001) A library of Arabidopsis 35S-cDNA lines foridentifying novel mutants. Plant Molecular Biology 46, 695–703.doi: 10.1023/A:1011699722052
LeeH,GuoY,OhtaM,XiongL, StevensonB,Zhu JK (2002)LOS2, a geneticlocus required for cold-responsive gene transcription encodes a bi-functional enolase. The EMBO Journal 21, 2692–2702. doi: 10.1093/emboj/21.11.2692
Lee U, Wie C, Fernandez BO, Feelisch M, Vierling E (2008) Modulationof nitrosative stress by S-nitrosoglutathione reductase is critical forthermotolerance and plant growth in Arabidopsis. The Plant Cell 20,786–802. doi: 10.1105/tpc.107.052647
Leung J, Giraudat J (1998) Abscisic acid signal transduction. Annual Reviewof Plant Physiology and Plant Molecular Biology 49, 199–222.doi: 10.1146/annurev.arplant.49.1.199
Leung J, Bouvier-Durand M, Morris PC, Guerrier D, Chefdor F, Giraudat J(1994) Arabidopsis ABA response gene ABI1: features of a calcium-modulated protein phosphatase. Science 264, 1448–1452. doi: 10.1126/science.7910981
Leung J, Merlot S, Giraudat J (1997) The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and ABI1 genes encode homologous proteinphosphatases 2C involved in abscisic acid signal transduction.The Plant Cell 9, 759–771.
Li X, Zhang Y (2002) Reverse genetics by fast neutronmutagenesis in higherplants. Functional & Integrative Genomics 2, 254–258. doi: 10.1007/s10142-002-0076-0
Li ZM, Yan CS, Wu G, Cao JS, He ZH (2005) [sdt1, a novel Arabidopsismutant tolerant to drought and high-salinity stresses.] Shi Yan Sheng WuXue Bao 38, 461–466.
Li Y, Rosso MG, Ulker B, Weisshaar B (2006) Analysis of T-DNA insertionsite distribution patterns in Arabidopsis thaliana reveals special featuresof genes without insertions. Genomics 87, 645–652. doi: 10.1016/j.ygeno.2005.12.010
Lindsay MP, Lagudah ES, Hare RA, Munns R (2004) A locus for sodiumexclusion (Nax1), a trait for salt tolerance, mapped in durum wheat.Functional Plant Biology 31, 1105–1114. doi: 10.1071/FP04111
Lister R, Gregory BD, Ecker JR (2009) Next is now: new technologiesfor sequencing of genomes, transcriptomes, and beyond. CurrentOpinion in Plant Biology 12, 107–118. doi: 10.1016/j.pbi.2008.11.004
Genetic identification of plant genes in response to stress Functional Plant Biology 715

Liu YG,Mitsukawa N, Oosumi T, Whittier RF (1995) Efficient isolation andmapping of Arabidopsis thaliana T-DNA insert junctions by thermalasymmetric interlaced PCR. The Plant Journal 8, 457–463. doi: 10.1046/j.1365-313X.1995.08030457.x
Liu J, IshitaniM,HalfterU,KimCS,Zhu JK (2000)TheArabidopsis thalianaSOS2 gene encodes a protein kinase that is required for salt tolerance.Proceedings of the National Academy of Sciences of the United Statesof America 97, 3730–3734. doi: 10.1073/pnas.060034197
Lu S, Sun YH, Shi R, Clark C, Li L, Chiang VL (2005) Novel and mechanicalstress-responsive microRNAs in Populus trichocarpa that are absent fromArabidopsis. The Plant Cell 17, 2186–2203. doi: 10.1105/tpc.105.033456
Lu S, Sun YH, Chiang VL (2008) Stress-responsive microRNAs in Populus.ThePlant Journal 55, 131–151. doi: 10.1111/j.1365-313X.2008.03497.x
MaH,Moore PH, Liu Z, KimMS,YuQ, FitchMM, Sekioka T, PatersonAH,Ming R (2004) High-density linkage mapping revealed suppression ofrecombination at the sex determination locus in papaya. Genetics 166,419–436. doi: 10.1534/genetics.166.1.419
MahalingamR, Fedoroff N (2001) Screening insertion libraries for mutationsin many genes simultaneously using DNA microarrays. Proceedings ofthe National Academy of Sciences of the United States of America 98,7420–7425. doi: 10.1073/pnas.121189598
Martienssen RA (1998) Functional genomics: probing plant gene functionand expression with transposons. Proceedings of the National Academyof Sciences of the United States of America 95, 2021–2026. doi: 10.1073/pnas.95.5.2021
Martínez-Atienza J, Jiang X, Garciadeblas B, Mendoza I, Zhu JK, Pardo JM,Quintero FJ (2007) Conservation of the salt overly sensitive pathway inrice. Plant Physiology 143, 1001–1012. doi: 10.1104/pp.106.092635
Masle J, Gilmore SR, Farquhar GD (2005) The ERECTA gene regulatesplant transpiration efficiency in Arabidopsis. Nature 436, 866–870.doi: 10.1038/nature03835
Masson JY, Ramotar D (1998) The transcriptional activator Imp2pmaintainsion homeostasis in Saccharomyces cerevisiae. Genetics 149, 893–901.
Mathur J, SzabadosL,SchaeferS,GrunenbergB,LossowA, Jonas-StraubeE,Schell J, Koncz C, Koncz-Kalman Z (1998) Gene identification withsequenced T-DNA tags generated by transformation of Arabidopsis cellsuspension. The Plant Journal 13, 707–716. doi: 10.1046/j.1365-313X.1998.00059.x
Matzke MA, Birchler JA (2005) RNAi-mediated pathways in the nucleus.Nature Reviews Genetics 6, 24–35. doi: 10.1038/nrg1500
McBride KE, Summerfelt KR (1990) Improved binary vectors forAgrobacterium-mediated plant transformation. Plant MolecularBiology 14, 269–276. doi: 10.1007/BF00018567
McKhannHI,GeryC,BerardA, LevequeS, Zuther E,HinchaDK,DeMita S,Brunel D, Teoule E (2008) Natural variation in CBF gene sequence, geneexpression and freezing tolerance in the Versailles core collection ofArabidopsis thaliana. BMC Plant Biology 8, 105. doi: 10.1186/1471-2229-8-105
Medina J, Rodriguez-Franco M, Penalosa A, Carrascosa MJ, Neuhaus G,Salinas J (2005) Arabidopsis mutants deregulated in RCI2A expressionreveal new signaling pathways in abiotic stress responses. The PlantJournal 42, 586–597. doi: 10.1111/j.1365-313X.2005.02400.x
Meissner R, Chague V, Zhu Q, Emmanuel E, Elkind Y, Levy AA (2000)Technical advance: a high throughput system for transposon tagging andpromoter trapping in tomato. The Plant Journal 22, 265–274.doi: 10.1046/j.1365-313x.2000.00735.x
Menz MA, Klein RR, Mullet JE, Obert JA, Unruh NC, Klein PE (2002)A high-density genetic map of Sorghum bicolor (L.) Moench based on2926 AFLP, RFLP and SSR markers. Plant Molecular Biology 48,483–499. doi: 10.1023/A:1014831302392
Merlot S, Mustilli AC, Genty B, North H, Lefebvre V, Sotta B, Vavasseur A,Giraudat J (2002) Use of infrared thermal imaging to isolate Arabidopsismutants defective in stomatal regulation. The Plant Journal 30, 601–609.doi: 10.1046/j.1365-313X.2002.01322.x
Meyer K, Leube MP, Grill E (1994) A protein phosphatase 2C involvedin ABA signal transduction in Arabidopsis thaliana. Science 264,1452–1455. doi: 10.1126/science.8197457
Meyerowitz EM (1989) Arabidopsis, a useful weed. Cell 56, 263–269.doi: 10.1016/0092-8674(89)90900-8
Millar AJ, Short SR, Chua NH, Kay SA (1992) A novel circadian phenotypebased on firefly luciferase expression in transgenic plants. The PlantCell 4, 1075–1087.
Miller AK, Galiba G, Dubcovsky J (2006) A cluster of 11 CBF transcriptionfactors is located at the frost tolerance locus Fr-Am2 in Triticummonococcum. Molecular Genetics and Genomics 275, 193–203.doi: 10.1007/s00438-005-0076-6
Misra S,WuY,VenkataramanG, Sopory SK,TutejaN (2007)HeterotrimericG-protein complex and G-protein-coupled receptor from a legume(Pisum sativum): role in salinity and heat stress and cross-talk withphospholipase C. The Plant Journal 51, 656–669. doi: 10.1111/j.1365-313X.2007.03169.x
Mittler R (2006) Abiotic stress, the field environment and stress combination.Trends in Plant Science 11, 15–19. doi: 10.1016/j.tplants.2005.11.002
Mortensen R (2006) Overview of gene targeting by homologousrecombination. Current Protocols in Molecular Biology (Suppl. 76),23.1.1–23.1.12. doi: 10.1002/0471142727.mb2301s76
Mullan DJ, Colmer TD, Francki MG (2007) Arabidopsis-rice–wheat geneorthologues for Na+ transport and transcript analysis in wheat-L. elongatum aneuploids under salt stress. Molecular Genetics andGenomics 277, 199–212. doi: 10.1007/s00438-006-0184-y
Munns R, Tester M (2008) Mechanisms of salinity tolerance. AnnualReview of Plant Biology 59, 651–681. doi: 10.1146/annurev.arplant.59.032607.092911
Munns R, James RA, Lauchli A (2006) Approaches to increasing the salttolerance of wheat and other cereals. Journal of Experimental Botany57, 1025–1043. doi: 10.1093/jxb/erj100
Mustilli AC, Merlot S, Vavasseur A, Fenzi F, Giraudat J (2002) ArabidopsisOST1 protein kinase mediates the regulation of stomatal aperture byabscisic acid and acts upstream of reactive oxygen species production.The Plant Cell 14, 3089–3099. doi: 10.1105/tpc.007906
Nagano AJ, FukazawaM, Hayashi M, Ikeuchi M, Tsukaya H, Nishimura M,Hara-Nishimura I (2008) AtMap1: a DNA microarray for genomicdeletion mapping in Arabidopsis thaliana. The Plant Journal 56,1058–1065. doi: 10.1111/j.1365-313X.2008.03656.x
NakamuraH,HakataM,AmanoK,MiyaoA,TokiN, et al. (2007)Agenome-wide gain-of function analysis of rice genes using the FOX-huntingsystem. Plant Molecular Biology 65, 357–371. doi: 10.1007/s11103-007-9243-y
Nakashima K, Ito Y, Yamaguchi-Shinozaki K (2009) Transcriptionalregulatory networks in response to abiotic stresses in Arabidopsis andgrasses. Plant Physiology 149, 88–95. doi: 10.1104/pp.108.129791
Nakazawa M, Ichikawa T, Ishikawa A, Kobayashi H, Tsuhara Y,Kawashima M, Suzuki K, Muto S, Matsui M (2003) Activationtagging, a novel tool to dissect the functions of a gene family.The Plant Journal 34, 741–750. doi: 10.1046/j.1365-313X.2003.01758.x
Nieto C, Piron F, Dalmais M, Marco CF, Moriones E, et al. (2007)EcoTILLING for the identification of allelic variants of melon eIF4E,a factor that controls virus susceptibility. BMC Plant Biology 7, 34.doi: 10.1186/1471-2229-7-34
O’Neill CM,Morgan C, Kirby J, Tschoep H, Deng PX, et al. (2008) Six newrecombinant inbred populations for the study of quantitative traits inArabidopsis thaliana. Theoretical and Applied Genetics 116, 623–634.doi: 10.1007/s00122-007-0696-9
Obata T, Kitamoto HK, Nakamura A, Fukuda A, Tanaka Y (2007) Riceshaker potassium channel OsKAT1 confers tolerance to salinity stress onyeast and rice cells. Plant Physiology 144, 1978–1985. doi: 10.1104/pp.107.101154
716 Functional Plant Biology C. Papdi et al.

Oldenhof H,WolkersWF, Bowman JL, Tablin F, Crowe JH (2006) Freezingand desiccation tolerance in the moss Physcomitrella patens: an in situFourier transform infrared spectroscopic study. Biochimica et BiophysicaActa 1760, 1226–1234.
Oliver MJ (1996) Desiccation tolerance in vegetative plant cells.Physiologia Plantarum 97, 779–787. doi: 10.1111/j.1399-3054.1996.tb00544.x
Oono Y, Seki M, Nanjo T, Narusaka M, Fujita M, et al. (2003) Monitoringexpression profiles of Arabidopsis gene expression during rehydrationprocess after dehydration using ca 7000 full-length cDNA microarray.ThePlant Journal 34, 868–887. doi: 10.1046/j.1365-313X.2003.01774.x
Ossowski S, Schneeberger K, Clark RM, Lanz C, Warthmann N, Weigel D(2008) Sequencing of natural strains of Arabidopsis thaliana with shortreads. Genome Research 18, 2024–2033. doi: 10.1101/gr.080200.108
Page DR, Grossniklaus U (2002) The art and design of genetic screens:Arabidopsis thaliana.NatureReviews.Genetics3, 124–136.doi:10.1038/nrg730
Palatnik JF,AllenE,WuX,SchommerC,SchwabR,Carrington JC,WeigelD(2003) Control of leaf morphogenesis by microRNAs. Nature 425,257–263. doi: 10.1038/nature01958
PandeyS,NelsonDC,AssmannSM(2009)TwonovelGPCR-typeGproteinsare abscisic acid receptors in Arabidopsis. Cell 136, 136–148.doi: 10.1016/j.cell.2008.12.026
Panikashvili D, Savaldi-Goldstein S, Mandel T, Yifhar T, Franke RB,Hofer R, Schreiber L, Chory J, Aharoni A (2007) The ArabidopsisDESPERADO/AtWBC11 transporter is required for cutin and waxsecretion. Plant Physiology 145, 1345–1360. doi: 10.1104/pp.107.105676
Papdi C, Abraham E, Joseph MP, Popescu C, Koncz C, Szabados L (2008)Functional identification of Arabidopsis stress regulatory genes usingthe controlled cDNA overexpression system. Plant Physiology 147,528–542. doi: 10.1104/pp.108.116897
Parinov S, Sundaresan V (2000) Functional genomics in Arabidopsis: large-scale insertional mutagenesis complements the genome sequencingproject. Current Opinion in Biotechnology 11, 157–161. doi: 10.1016/S0958-1669(00)00075-6
Peleg Z, Saranga Y, Suprunova T, Ronin Y, Roder MS, Kilian A, Korol AB,FahimaT (2008)High-density geneticmapof durumwheat�wild emmerwheatbasedonSSRandDArTmarkers.Theoretical andAppliedGenetics117, 103–115. doi: 10.1007/s00122-008-0756-9
Peters JL, Cnudde F, Gerats T (2003) Forward genetics and map-basedcloning approaches. Trends in Plant Science 8, 484–491. doi: 10.1016/j.tplants.2003.09.002
Piffanelli P, Droc G, Mieulet D, Lanau N, Bès M, et al. (2007) Large-scalecharacterization of Tos17 insertion sites in a rice T-DNA mutant library.Plant Molecular Biology 65, 587–601. doi: 10.1007/s11103-007-9222-3
Platten JD, CotsaftisO, Berthomieu P, Bohnert H,Davenport RJ, et al. (2006)Nomenclature for HKT transporters, key determinants of plant salinitytolerance. Trends in Plant Science 11, 372–374. doi: 10.1016/j.tplants.2006.06.001
PradhanAK, Gupta V,MukhopadhyayA, ArumugamN, Sodhi YS, Pental D(2003) A high-density linkage map in Brassica juncea (Indian mustard)using AFLP and RFLP markers. Theoretical and Applied Genetics 106,607–614.
Puchta H (2002) Gene replacement by homologous recombination in plants.Plant Molecular Biology 48, 173–182. doi: 10.1023/A:1013761821763
Qu S, Desai A, Wing R, Sundaresan V (2008) A versatile transposon-basedactivation tag vector system for functional genomics in cereals and othermonocot plants. Plant Physiology 146, 189–199. doi: 10.1104/pp.107.111427
Quatrano RS, McDaniel SF, Khandelwal A, Perroud PF, Cove DJ (2007)Physcomitrella patens: mosses enter the genomic age. Current Opinionin Plant Biology 10, 182–189. doi: 10.1016/j.pbi.2007.01.005
Quintero FJ, Garciadeblas B, Rodriguez-Navarro A (1996) The SAL1 geneof Arabidopsis, encoding an enzyme with 30(20), 50-bisphosphatenucleotidase and inositol polyphosphate 1-phosphatase activities,increases salt tolerance in yeast. The Plant Cell 8, 529–537.
Redei GP (1975) Arabidopsis as a genetic tool. Annual Review of Genetics9, 111–127. doi: 10.1146/annurev.ge.09.120175.000551
RédeiGP,KonczC (1992)Classicalmutagenesis. In ‘Methods inArabidopsisresearch’. (Eds C Koncz, N-H Chua, J Schell) pp. 16–82. (WorldScientific: Singapore)
Reiss B (2003) Homologous recombination and gene targeting in plantcells. International Review of Cytology 228, 85–139. doi: 10.1016/S0074-7696(03)28003-7
Remy S, Thiry E, Coemans B, Windelinckx S, Swennen R, Sagi L (2005)ImprovedT-DNAvector for tagging plant promoters via high-throughputluciferase screening. BioTechniques 38, 763–770. doi: 10.2144/05385RR01
Ren ZH, Gao JP, Li LG, Cai XL, Huang W, Chao DY, Zhu MZ, Wang ZY,Luan S, Lin HX (2005) A rice quantitative trait locus for salt toleranceencodes a sodium transporter. Nature Genetics 37, 1141–1146.doi: 10.1038/ng1643
Rensing SA, Lang D, Zimmer AD, Terry A, Salamov A, et al. (2008)The Physcomitrella genome reveals evolutionary insights into theconquest of land by plants. Science 319, 64–69. doi: 10.1126/science.1150646
Reyes JL, Chua NH (2007) ABA induction of miR159 controls transcriptlevels of two MYB factors during Arabidopsis seed germination.The Plant Journal 49, 592–606. doi: 10.1111/j.1365-313X.2006.02980.x
Riggs CD, Chrispeels MJ (1987) Luciferase reporter gene cassettes for plantgene expression studies. Nucleic Acids Research 15, 8115. doi: 10.1093/nar/15.19.8115
Ríos G, Lossow A, Hertel B, Breuer F, Schaefer S, et al. (2002) Rapididentification of Arabidopsis insertion mutants by non-radioactivedetection of T-DNA tagged genes. The Plant Journal 32, 243–253.doi: 10.1046/j.1365-313X.2002.01416.x
Ríos G, Naranjo MA, Iglesias DJ, Ruiz-Rivero O, Geraud M, Usach A,Talon M (2008) Characterization of hemizygous deletions in citrususing array-comparative genomic hybridization and microsyntenycomparisons with the poplar genome. BMC Genomics 9, 381.doi: 10.1186/1471-2164-9-381
Robinson SJ, Parkin IA (2008) Differential SAGE analysis in Arabidopsisuncovers increased transcriptome complexity in response to lowtemperature. BMC Genomics 9, 434. doi: 10.1186/1471-2164-9-434
Rodriguez-Navarro A, Rubio F (2006) High-affinity potassium and sodiumtransport systems in plants. Journal of Experimental Botany 57,1149–1160. doi: 10.1093/jxb/erj068
Rosso MG, Li Y, Strizhov N, Reiss B, Dekker K, Weisshaar B (2003)An Arabidopsis thaliana T-DNA mutagenized population (GABI-Kat)forflanking sequence tag-based reversegenetics.PlantMolecularBiology53, 247–259. doi: 10.1023/B:PLAN.0000009297.37235.4a
Ruggiero B, Koiwa H, Manabe Y, Quist TM, Inan G, Saccardo F, Joly RJ,Hasegawa PM, Bressan RA, Maggio A (2004) Uncoupling the effects ofabscisic acid on plant growth and water relations. Analysis of sto1/nced3,an abscisic acid-deficient but salt stress-tolerant mutant in Arabidopsis.Plant Physiology 136, 3134–3147. doi: 10.1104/pp.104.046169
Rus A, Baxter I, Muthukumar B, Gustin J, Lahner B, Yakubova E, Salt DE(2006) Natural variants of AtHKT1 enhance Na+ accumulation in twowild populations of Arabidopsis. PLOS Genetics 2, e210. doi: 10.1371/journal.pgen.0020210
Saavedra L, Svensson J, Carballo V, Izmendi D, Welin B, Vidal S (2006)A dehydrin gene in Physcomitrella patens is required for salt andosmotic stress tolerance. The Plant Journal 45, 237–249. doi: 10.1111/j.1365-313X.2005.02603.x
Genetic identification of plant genes in response to stress Functional Plant Biology 717

Saez A, Rodrigues A, Santiago J, Rubio S, Rodriguez PL (2008)HAB1–SWI3B interaction reveals a link between abscisic acidsignaling and putative SWI/SNF chromatin-remodeling complexes inArabidopsis.ThePlantCell20, 2972–2988. doi: 10.1105/tpc.107.056705
Salathia N, Lee HN, Sangster TA, Morneau K, Landry CR, et al. (2007)Indel arrays: an affordable alternative for genotyping. The Plant Journal51, 727–737. doi: 10.1111/j.1365-313X.2007.03194.x
Saliba-Colombani V, Causse M, Gervais L, Philouze J (2000) Efficiencyof RFLP, RAPD, and AFLP markers for the construction of anintraspecific map of the tomato genome. Genome 43, 29–40.doi: 10.1139/gen-43-1-29
Samson F, Brunaud V, Balzergue S, Dubreucq B, Lepiniec L, Pelletier G,Caboche M, Lecharny A (2002) FLAGdb/FST: a database of mappedflanking insertion sites (FSTs) of Arabidopsis thaliana T-DNAtransformants. Nucleic Acids Research 30, 94–97. doi: 10.1093/nar/30.1.94
SandersD,Pelloux J,BrownleeC,Harper JF (2002)Calciumat the crossroadsof signaling. The Plant Cell 14(Suppl.), S401–S417.
Schaefer DG (2001) Gene targeting in Physcomitrella patens. CurrentOpinion in Plant Biology 4, 143–150. doi: 10.1016/S1369-5266(00)00150-3
Schaefer DG, Zryd JP (1997) Efficient gene targeting in the mossPhyscomitrella patens. The Plant Journal 11, 1195–1206. doi: 10.1046/j.1365-313X.1997.11061195.x
SchafferR,RamsayN, SamachA,CordenS, Putterill J, Carre IA,CouplandG(1998) The late elongated hypocotyl mutation of Arabidopsis disruptscircadian rhythms and the photoperiodic control of flowering. Cell 93,1219–1229. doi: 10.1016/S0092-8674(00)81465-8
Schell J, VanMontaguM, De Beuckeleer M, De BlockM, Depicker A, et al.(1979) Interactions and DNA transfer between Agrobacteriumtumefaciens, the Ti-plasmid and the plant host. Proceedings of theRoyal Society London. Series B, Containing papers of a Biologicalcharacter. Royal Society (Great Britain) 204, 251–266. doi: 10.1098/rspb.1979.0026
Schneider A, Leister D (2006) Forward genetic screening of insertionalmutants. Methods in Molecular Biology (Clifton, N.J.) 323, 147–161.
Seki M, Narusaka M, Ishida J, Nanjo T, Fujita M, et al. (2002) Monitoringthe expression profiles of 7000 Arabidopsis genes under drought,cold and high-salinity stresses using a full-length cDNA microarray.The Plant Journal 31, 279–292. doi: 10.1046/j.1365-313X.2002.01359.x
Serrano R, Rodriguez-Navarro A (2001) Ion homeostasis during salt stressin plants. Current Opinion in Cell Biology 13, 399–404. doi: 10.1016/S0955-0674(00)00227-1
Shendure J, Ji H (2008) Next-generation DNA sequencing. NatureBiotechnology 26, 1135–1145. doi: 10.1038/nbt1486
Shi H, Ishitani M, Kim C, Zhu JK (2000) The Arabidopsis thaliana salttolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proceedingsof the National Academy of Sciences of the United States of America97, 6896–6901. doi: 10.1073/pnas.120170197
Shikazono N, Suzuki C, Kitamura S, Watanabe H, Tano S, Tanaka A (2005)Analysis of mutations induced by carbon ions in Arabidopsis thaliana.Journal of Experimental Botany 56, 587–596. doi: 10.1093/jxb/eri047
Shukla LI, Chinnusamy V, Sunkar R (2008) The role of microRNAs andother endogenous small RNAs in plant stress responses. Biochimica etBiophysica Acta 1779, 743–748.
Spiegelman JI, Mindrinos MN, Fankhauser C, Richards D, Lutes J, Chory J,Oefner PJ (2000) Cloning of the Arabidopsis RSF1 gene by using amapping strategy based on high-densityDNAarrays and denaturing high-performance liquid chromatography. The Plant Cell 12, 2485–2498.
Sun Z, Wang Z, Tu J, Zhang J, Yu F, McVetty PB, Li G (2007)An ultradense genetic recombination map for Brassica napus,consisting of 13551 SRAP markers. Theoretical and Applied Genetics114, 1305–1317. doi: 10.1007/s00122-006-0483-z
Sunkar R, Zhu JK (2004) Novel and stress-regulated microRNAs and othersmall RNAs from Arabidopsis. The Plant Cell 16, 2001–2019.doi: 10.1105/tpc.104.022830
Sunkar R, Kapoor A, Zhu JK (2006) Posttranscriptional induction of twoCu/Zn superoxide dismutase genes in Arabidopsis is mediated bydownregulation of miR398 and important for oxidative stresstolerance. The Plant Cell 18, 2051–2065. doi: 10.1105/tpc.106.041673
Sussman MR, Amasino RM, Young JC, Krysan PJ, Austin-Phillips S(2000) The Arabidopsis knockout facility at the University ofWisconsin–Madison. Plant Physiology 124, 1465–1467. doi: 10.1104/pp.124.4.1465
Suzuki T, Eiguchi M, Kumamaru T, Satoh H, Matsusaka H, Moriguchi K,Nagato Y, Kurata N (2008) MNU-induced mutant pools and highperformance TILLING enable finding of any gene mutation in rice.Molecular Genetics and Genomics 279, 213–223. doi: 10.1007/s00438-007-0293-2
Szabados L, Koncz C (2003) Identification of T-DNA insertions inArabidopsis genes. In ‘Genomics of plants and fungi’.(Eds HJ Bohnert, RA Prade) pp. 255–277. (Marcel Dekker Inc.:New York)
Szabados L, Kovács I, Oberschall A, Ábrahám E, Kerekes I, et al. (2002)Distribution of 1000 sequenced T-DNA tags in the Arabidopsis genome.ThePlant Journal 32, 233–242. doi: 10.1046/j.1365-313X.2002.01417.x
Tadege M, Ratet P, Mysore KS (2005) Insertional mutagenesis: a SwissArmy knife for functional genomics of Medicago truncatula. Trends inPlant Science 10, 229–235. doi: 10.1016/j.tplants.2005.03.009
Tadege M, Wen J, He J, Tu H, Kwak Y, et al. (2008) Large-scale insertionalmutagenesis using the Tnt1 retrotransposon in the model legumeMedicago truncatula. The Plant Journal 54, 335–347. doi: 10.1111/j.1365-313X.2008.03418.x
Tang G (2005) siRNA and miRNA: an insight into RISCs. Trends inBiochemical Sciences 30, 106–114. doi: 10.1016/j.tibs.2004.12.007
Tanksley SD, Ganal MW, Prince JP, de-Vicente MC, Bonierbale MW, et al.(1992) High density molecular linkage maps of the tomato and potatogenomes. Genetics 132, 1141–1160.
Teeri TH, Herrera-Estrella L, Depicker A, VanMontaguM, Palva ET (1986)Identification of plant promoters in situ by T-DNA-mediatedtranscriptional fusions to the npt-II gene. The EMBO Journal 5,1755–1760.
Terada R, Johzuka-Hisatomi Y, Saitoh M, Asao H, Iida S (2007) Genetargeting by homologous recombination as a biotechnological tool forrice functional genomics. Plant Physiology 144, 846–856. doi: 10.1104/pp.107.095992
The Arabidopsis Genome Initiative (2000a) Analysis of the genomesequence of the flowering plant Arabidopsis thaliana. Nature 408,796–815. doi: 10.1038/35048692
The Arabidopsis Genome Initiative (2000b) Analysis of the genomesequence of the flowering plant Arabidopsis thaliana. Nature 408,796–815. doi: 10.1038/35048692
Thole V, Worland B, Snape JW, Vain P (2007) The pCLEAN dual binaryvector system for Agrobacterium-mediated plant transformation. PlantPhysiology 145, 1211–1219. doi: 10.1104/pp.107.108563
Till BJ, Colbert T, Tompa R, Enns LC, Codomo CA, et al. (2003) High-throughput TILLING for functional genomics. Methods in MolecularBiology (Clifton, N.J.) 236, 205–220.
Till BJ, Reynolds SH,Weil C, Springer N, Burtner C, et al. (2004) Discoveryof induced point mutations in maize genes by TILLING. BMC PlantBiology 4, 12. doi: 10.1186/1471-2229-4-12
Till BJ, Colbert T, Codomo C, Enns L, Johnson J, et al. (2006a) High-throughput TILLING for Arabidopsis. Methods in Molecular Biology(Clifton, N.J.) 323, 127–135. doi: 10.1385/1-59745-003-0:127
Till BJ, Zerr T, Comai L, Henikoff S (2006b) A protocol for TILLING andEcotilling in plants and animals. Nature Protocols 1, 2465–2477.doi: 10.1038/nprot.2006.329
718 Functional Plant Biology C. Papdi et al.

Till BJ, Cooper J, Tai TH,Colowit P,GreeneEA,Henikoff S, Comai L (2007)Discovery of chemically induced mutations in rice by TILLING. BMCPlant Biology 7, 19. doi: 10.1186/1471-2229-7-19
Tinland B, Hohn B (1995) Recombination between prokaryotic andeukaryotic DNA: integration of Agrobacterium tumefaciens T-DNAinto the plant genome. Genetic Engineering 17, 209–229.
Tondelli A, Francia E, Barabaschi D, Aprile A, Skinner JS, Stockinger EJ,Stanca AM, Pecchioni N (2006) Mapping regulatory genes ascandidates for cold and drought stress tolerance in barley.Theoretical and Applied Genetics 112, 445–454. doi: 10.1007/s00122-005-0144-7
Torii KU, Mitsukawa N, Oosumi T, Matsuura Y, Yokoyama R, Whittier RF,Komeda Y (1996) The Arabidopsis ERECTA gene encodes a putativereceptor protein kinase with extracellular leucine-rich repeats. The PlantCell 8, 735–746.
Ülker B, Peiter E, Dixon DP, Moffat C, Capper R, Bouché N, Edwards R,Sanders D, Knight H, Knight MR (2008) Getting the most out of publiclyavailable T-DNA insertion lines. The Plant Journal 56, 665–677.doi: 10.1111/j.1365-313X.2008.03608.x
Urao T, Yakubov B, Satoh R, Yamaguchi-Shinozaki K, Seki M,Hirayama T, Shinozaki K (1999) A transmembrane hybrid-typehistidine kinase in Arabidopsis functions as an osmosensor. ThePlant Cell 11, 1743–1754.
Vágújfalvi A, Aprile A, Miller A, Dubcovsky J, Delugu G, Galiba G,Cattivelli L (2005) The expression of several Cbf genes at the Fr-A2locus is linked to frost resistance in wheat. Molecular Genetics andGenomics 274, 506–514. doi: 10.1007/s00438-005-0047-y
van Enckevort LJ, Droc G, Piffanelli P, Greco R, Gagneur C, et al. (2005)EU-OSTID: a collection of transposon insertional mutants for functionalgenomics in rice. Plant Molecular Biology 59, 99–110. doi: 10.1007/s11103-005-8532-6
Vanderauwera S, Zimmermann P, Rombauts S, Vandenabeele S,Langebartels C, Gruissem W, Inze D, Van Breusegem F (2005)Genome-wide analysis of hydrogen peroxide-regulated geneexpression in Arabidopsis reveals a high light-induced transcriptionalcluster involved in anthocyanin biosynthesis. Plant Physiology 139,806–821. doi: 10.1104/pp.105.065896
Wan S, Wu J, Zhang Z, Sun X, Lv Y, et al. (2009) Activation tagging, anefficient tool for functional analysis of the rice genome. Plant MolecularBiology 69, 69–80. doi: 10.1007/s11103-008-9406-5
Wang X (2004) Lipid signaling. Current Opinion in Plant Biology 7,329–336. doi: 10.1016/j.pbi.2004.03.012
Wang ZY, Tobin EM (1998) Constitutive expression of the CIRCADIANCLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythmsand suppresses its own expression. Cell 93, 1207–1217. doi: 10.1016/S0092-8674(00)81464-6
Wang XF, Zhang DP (2008) Abscisic acid receptors: multiple signal-perception sites. Annals of Botany 101, 311–317. doi: 10.1093/aob/mcm284
Wang Y, Holroyd G, Hetherington AM, Ng CK (2004) Seeing ‘cool’ and‘hot’ – infrared thermography as a tool for non-invasive, high-throughputscreening of Arabidopsis guard cell signalling mutants. Journal ofExperimental Botany 55, 1187–1193. doi: 10.1093/jxb/erh135
WangX, Sato S, Tabata S, Kawasaki S (2008) A high-density linkagemap ofLotus japonicus based on AFLP and SSR markers. DNA Research 15,323–332. doi: 10.1093/dnares/dsn022
Weigel D, Ahn JH, BlázquezMA, Borevitz JO, Christensen SK, et al. (2000)Activation tagging in Arabidopsis. Plant Physiology 122, 1003–1014.doi: 10.1104/pp.122.4.1003
Wenzl P, Li H, Carling J, Zhou M, Raman H, et al. (2006) A high-densityconsensus map of barley linking DArT markers to SSR, RFLP and STSloci and agricultural traits. BMC Genomics 7, 206. doi: 10.1186/1471-2164-7-206
Wisman E, Hartmann U, Sagasser M, Baumann E, Palme K, Hahlbrock K,Saedler H, Weisshaar B (1998) Knock-out mutants from anEn-1 mutagenized Arabidopsis thaliana population generatephenylpropanoid biosynthesis phenotypes. Proceedings of the NationalAcademy of Sciences of the United States of America 95, 12 432–12 437.doi: 10.1073/pnas.95.21.12432
Wu SJ, Ding L, Zhu JK (1996) SOS1, a genetic locus essential for salttolerance and potassium acquisition. The Plant Cell 8, 617–627.
XiaZ,TsubokuraY,HoshiM,HanawaM,YanoC, et al. (2007)An integratedhigh-density linkage map of soybean with RFLP, SSR, STS, and AFLPmarkers using A single F2 population. DNA Research 14, 257–269.doi: 10.1093/dnares/dsm027
Xiong L, Ishitani M, Lee H, Zhu JK (1999a) HOS5-a negative regulatorof osmotic stress-induced gene expression in Arabidopsis thaliana. ThePlant Journal 19, 569–578. doi: 10.1046/j.1365-313X.1999.00558.x
Xiong L, Ishitani M, Zhu JK (1999b) Interaction of osmotic stress,temperature, and abscisic acid in the regulation of gene expression inArabidopsis. Plant Physiology 119, 205–212. doi: 10.1104/pp.119.1.205
XiongL,LeeB, IshitaniM,LeeH,ZhangC,ZhuJK (2001)FIERY1encodingan inositol polyphosphate 1-phosphatase is a negative regulator ofabscisic acid and stress signaling in Arabidopsis. Genes &Development 15, 1971–1984. doi: 10.1101/gad.891901
Xiong L, Lee H, Ishitani M, Tanaka Y, Stevenson B, Koiwa H, Bressan RA,Hasegawa PM, Zhu JK (2002) Repression of stress-responsive genes byFIERY2, a novel transcriptional regulator in Arabidopsis. Proceedings ofthe National Academy of Sciences of the United States of America 99,10 899–10 904. doi: 10.1073/pnas.162111599
Xiong L, Wang RG, Mao G, Koczan JM (2006) Identification of droughttolerance determinants by genetic analysis of root response to droughtstress and abscisic acid. Plant Physiology 142, 1065–1074. doi: 10.1104/pp.106.084632
Yamaguchi-Shinozaki K, Shinozaki K (2006) Transcriptional regulatorynetworks in cellular responses and tolerance to dehydration and coldstresses. Annual Review of Plant Biology 57, 781–803. doi: 10.1146/annurev.arplant.57.032905.105444
Yamamoto YY, Tsuhara Y, Gohda K, Suzuki K, Matsui M (2003) Genetrapping of the Arabidopsis genome with a firefly luciferase reporter. ThePlant Journal 35, 273–283. doi: 10.1046/j.1365-313X.2003.01797.x
YanZ,DenneboomC,HattendorfA,DolstraO,DebenerT, StamP,Visser PB(2005) Construction of an integrated map of rose with AFLP, SSR, PK,RGA,RFLP,SCARandmorphologicalmarkers.Theoretical andAppliedGenetics 110, 766–777. doi: 10.1007/s00122-004-1903-6
Yokotani N, Ichikawa T, Kondou Y, Matsui M, Hirochika H, Iwabuchi M,Oda K (2009) Tolerance to various environmental stresses conferredby the salt-responsive rice gene ONAC063 in transgenic Arabidopsis.Planta 229, 1065–1075. doi: 10.1007/s00425-009-0895-5
Yu H, Chen X, Hong YY, Wang Y, Xua P, Kea SD, Liu HY, Zhu JK,Oliver DJ, Xiang CB (2008) Activated expression of an ArabidopsisHD-START protein confers drought tolerance with improved rootsystem and reduced stomatal density. The Plant Cell 20, 1134–1151.doi: 10.1105/tpc.108.058263
Zhang Z, Wei L, Zou X, Tao Y, Liu Z, Zheng Y (2008) Submergence-responsive microRNAs are potentially involved in the regulation ofmorphological and metabolic adaptations in maize root cells. Annals ofBotany 102, 509–519. doi: 10.1093/aob/mcn129
Zhao B, Liang R, Ge L, Li W, Xiao H, Lin H, Ruan K, Jin Y (2007)Identification of drought-induced microRNAs in rice. Biochemical andBiophysical Research Communications 354, 585–590. doi: 10.1016/j.bbrc.2007.01.022
Zhao B, Ge L, Liang R, Li W, Ruan K, Lin H, Jin Y (2009) Membersof miR-169 family are induced by high salinity and transiently inhibitthe NF-YA transcription factor. BMC Molecular Biology 10, 29.doi: 10.1186/1471-2199-10-29
Genetic identification of plant genes in response to stress Functional Plant Biology 719

Zheng X, Zhu J, Kapoor A, Zhu JK (2007) Role of Arabidopsis AGO6 insiRNA accumulation, DNA methylation and transcriptional genesilencing. The EMBO Journal 26, 1691–1701. doi: 10.1038/sj.emboj.7601603
Zheng X, Pontes O, Zhu J, Miki D, Zhang F, Li WX, Iida K, Kapoor A,Pikaard CS, Zhu JK (2008) ROS3 is an RNA-binding protein requiredfor DNA demethylation in Arabidopsis. Nature 455, 1259–1262.doi: 10.1038/nature07305
Zhou X, Wang G, Zhang W (2007) UV-B responsive microRNA genes inArabidopsis thaliana. Molecular Systems Biology 3, 103. doi: 10.1038/msb4100143
Zhou X, Wang G, Sutoh K, Zhu JK, Zhang W (2008) Identification of cold-inducible microRNAs in plants by transcriptome analysis. Biochimicaet Biophysica Acta 1779, 780–788.
Zhu JK (2002) Salt and drought stress signal transduction in plants. AnnualReview of Plant Biology 53, 247–273. doi: 10.1146/annurev.arplant.53.091401.143329
Zhu JK (2003) Regulation of ion homeostasis under salt stress. CurrentOpinion in Plant Biology 6, 441–445. doi: 10.1016/S1369-5266(03)00085-2
Zhu J, Shi H, Lee BH, Damsz B, Cheng S, Stirm V, Zhu JK, Hasegawa PM,Bressan RA (2004) An Arabidopsis homeodomain transcription factorgene, HOS9, mediates cold tolerance through a CBF-independentpathway. Proceedings of the National Academy of Sciences of theUnited States of America 101, 9873–9878. doi: 10.1073/pnas.0403166101
Zhu J, Jeong JC, Zhu Y, Sokolchik I, Miyazaki S, et al. (2008) Involvementof Arabidopsis HOS15 in histone deacetylation and cold tolerance.Proceedings of the National Academy of Sciences of the United Statesof America 105, 4945–4950. doi: 10.1073/pnas.0801029105
Zsigmond L, Rigo G, Szarka A, Székely G, Ötvös K, Darula Z,Medzihradszky KF, Koncz C, Koncz Z, Szabados L (2008)Arabidopsis PPR40 connects abiotic stress responses to mitochondrialelectron transport. Plant Physiology 146, 1721–1737. doi: 10.1104/pp.107.111260
Zuo J, Niu QW, Chua NH (2000) Technical advance: an estrogen receptor-based transactivator XVE mediates highly inducible gene expression intransgenic plants. The Plant Journal 24, 265–273. doi: 10.1046/j.1365-313x.2000.00868.x
Zupan JR, Zambryski P (1995) Transfer of T-DNA from Agrobacterium tothe plant cell. Plant Physiology 107, 1041–1047. doi: 10.1104/pp.107.4.1041
Zupan J, Muth TR, Draper O, Zambryski P (2000) The transfer of DNAfrom Agrobacterium tumefaciens into plants: a feast of fundamentalinsights. The Plant Journal 23, 11–28. doi: 10.1046/j.1365-313x.2000.00808.x
Manuscript received 26 February 2009, accepted 11 June 2009
720 Functional Plant Biology C. Papdi et al.
http://www.publish.csiro.au/journals/fpb