genotypic and symbiotic diversity of rhizobium populations associated with cultivated lentil and pea...
TRANSCRIPT

S
Gar
NPa
Ab
a
ARR2A
TG
KLPRAPE
I
cttranaiatco
0h
ARTICLE IN PRESSG ModelYAPM-25606; No. of Pages 8
Systematic and Applied Microbiology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Systematic and Applied Microbiology
j ourna l h omepage: www.elsev ier .de /syapm
enotypic and symbiotic diversity of Rhizobium populationsssociated with cultivated lentil and pea in sub-humid and semi-aridegions of Eastern Algeria
assira Riaha,b, Gilles Bénab,∗, Abdelhamid Djekouna, Karine Heulinb,hilippe de Lajudieb, Gisèle Laguerreb
Laboratoire de Génétique, Biochimie et Biotechnologies Végétalesm, Faculté des Sciences de la Nature, Université Constantine 1, Route Ain El Bey 25000,lgeriaIRD – INRA LSTM, Tropical and Mediterranean Symbiosis Laboratory, Campus International de Baillarguet, TA A-82/J, 34398 Montpellier Cedex 5, France
r t i c l e i n f o
rticle history:eceived 16 October 2013eceived in revised form7 November 2013ccepted 4 December 2013
his paper is dedicated to the memory ofisèle Laguerre.
a b s t r a c t
The genetic structure of rhizobia nodulating pea and lentil in Algeria, Northern Africa was determined.A total of 237 isolates were obtained from root nodules collected on lentil (Lens culinaris), proteaginousand forage pea (Pisum sativum) growing in two eco-climatic zones, sub-humid and semi-arid, in EasternAlgeria. They were characterised by PCR-restriction fragment length polymorphism (RFLP) of the 16S–23SrRNA intergenic region (IGS), and the nodD-F symbiotic region. The combination of these haplotypesallowed the isolates to be clustered into 26 distinct genotypes, and all isolates were classified as Rhizobiumleguminosarum. Symbiotic marker variation (nodD-F) was low but with the predominance of one nodhaplotype (g), which had been recovered previously at a high frequency in Europe. Sequence analysis of
eywords:entileahizobiumlgeriaopulation genetic structureco-climatic zones
the IGS further confirmed its high variability in the studied strains. An AMOVA analysis showed highlysignificant differentiation in the IGS haplotype distribution between populations from both eco-climaticzones. This differentiation was reflected by differences in dominant genotype frequencies. Conversely,no host plant effect was detected. The nodD gene sequence-based phylogeny suggested that symbioticgene diversity in pea and lentil nodulating rhizobial populations in Algeria was low compared to thatreported elsewhere in the world.
© 2014 Elsevier GmbH. All rights reserved.
ntroduction
In Algeria, food and forage legume productions are, along withereals, the major challenges for agriculture. In a poor soil con-ext, legume plants are interesting since they have the capacityo enter into nitrogen fixing symbiosis with rhizobial soil bacte-ia. They develop nodules on their roots, where bacteria reducetmospheric nitrogen to ammonia that is then available for plantitrogen nutrition. This symbiosis may have environmental andgronomic impacts due to the reduced needs for nitrogen chem-cal fertilizers. Legumes are frequently cultivated in rotation or inssociation with cereals in order to improve their nitrogen nutri-
Please cite this article in press as: N. Riah, et al., Genotypicated with cultivated lentil and pea in sub-humid and semi-arihttp://dx.doi.org/10.1016/j.syapm.2013.12.008
ion, crop yields and soil fertility. Among food legumes, lentil (Lensulinaris) cultivation has been promoted by the Algerian Ministryf Agriculture since 2007–2008 (http://www.minagri.dz). Lentil
∗ Corresponding author. Tel.: +33 467593862.E-mail address: [email protected] (G. Béna).
723-2020/$ – see front matter © 2014 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.syapm.2013.12.008
is one of the most ancient legume crops cultivated in occiden-tal Asia, Egypt and South Europe, and was probably spread fromWestern Asia [10]. Many ancient and long cultivated lentil vari-eties in Algeria have been lost and, nowadays, cultivated lentil areeither local – heterogenous mixtures – or of European origin. Sev-eral varieties have been introduced, and several new ones havebeen selected and bred for their adaptation to the various agro-climatic conditions found in this country [17]. Diversification offorage cultures is limited due to only a few research efforts carriedout on variety creation and introduction. Proteaginous pea (Pisumsativum) was introduced recently to Algeria, but available areasdevoted to its culture remain limited, despite its strategic impor-tance. Forage pea (P. sativum subsp. arvense) is well adapted andproductive in the whole northern part of Algeria. Although Pisumis a common genus in the Algerian flora, all cultivated peas are
and symbiotic diversity of Rhizobium populations associ-d regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),
introduced varieties. Areas cultivated with forage pea increased alot during the 1980s but decreased afterwards, despite the plantbreeding program for productive varieties conducted locally bythe ITGC (Technical Institute for Large Scale Farming). The ITGC

ARTICLE IN PRESSG ModelSYAPM-25606; No. of Pages 8
2 N. Riah et al. / Systematic and Applied Microbiology xxx (2014) xxx–xxx
ria wi
dom
riffrcitRPLlRnSa[
aLRRCsnvinf
[bsslsbi
oa
Fig. 1. Bioclimatic zones of Alge
emonstrated that forage pea associations with several crops (pea-at, -wheat, -barley) were superior to vetch associations due to aore balanced legume/cereal growth [17].Specificity is one of the main characteristics of the legume-
hizobium symbiotic associations. Indeed a rhizobial strain usuallynfects only a limited number of host plant species, which is aeature defined as its host range. The symbiovar concept ([41],ollowing the classical pathovar concept for plant pathogenic bacte-ia) was defined in order to distinguish rhizobia by their symbioticapabilities (i.e. host range), which is supported by symbiotic genenformation irrespective of their species affiliation. For instance, thehree symbiovars phaseoli, trifolii and viciae were created within thehizobium leguminosarum species for strains nodulating the tribeshaseolus, Trifolium and Viciae (including the genera Pisum, Vicia,athyrus and Lens), respectively [41]. Symbiovar viciae strains wereater reported in Rhizobium fabae [49] and Rhizobium pisi [37]. In. leguminosarum bv. viciae (Rlv), the host range is controlled byodulation (nod) genes carried by a symbiotic plasmid (pSym) [23].everal studies have shown genetic heterogeneity of the nodD genemong Rlv strains, and symbiotic specificity towards host plants13,24,25,28,33,34,48,55].
A large amount of literature has reported genetic diversitymong natural rhizobial populations nodulating Pisum, Vicia andathyrus, especially R. leguminosarum [4,12,13,24,32,34,35,50,56].. fabae [49] and R. pisi [37] were described based on ribosomalNA, atpD and recA phylogenies. R. fabae, isolated from Vicia faba inhina, also nodulates pea [49] and its nodC gene is similar to that ofymbiovar viciae [41]. R. pisi was created to reclassify a R. legumi-osarum strain nodulating P. sativum, Trifolium repens and Phaseolusulgaris [37]. Strains isolated from Pisum in Peru are phylogenet-cally close to R. leguminosarum and to Rhizobium etli, a speciesodulating beans in Mexico [43]. R. etli strains were also recovered
rom pea nodules in China [52].Several molecular studies on Rlv nodulating lentil were reported
20,26]. Recently Rashid et al. [38] published a phylogenetic studyased on 16S rRNA, recA, atpD and glnII genes from Rhizobiumtrains nodulating lentil cultivated in several regions in Bangladesh,howing that the majority of them may represent three separateineages in the R. etli – R. leguminosarum clade. Rashid et al. [39]howed that R. leguminosarum was the original symbiont of lentil,ut that it was associated with new lineages in Bangladesh where
Please cite this article in press as: N. Riah, et al., Genotypicated with cultivated lentil and pea in sub-humid and semi-arihttp://dx.doi.org/10.1016/j.syapm.2013.12.008
t was introduced.The agronomic and ecological impacts of rhizobia both rely
n their symbiotic properties (nodulation, efficiency, specificitynd host range) and their adaptation to environmental constraints
th the sampling sites indicated.
affecting nitrogen fixation, such as high temperatures, waterstress, salinity or soil acidity [1,53]. The plant genotype itselfinfluences rhizobial population structure in pea nodules [13].R. leguminosarum is sensitive to environmental perturbations,such as agronomic practices and heavy metal soil contamination[12,22,35]. Water deficit, rain irregularity and soil salinity alsorepresent major limitations for plant growth and agronomic pro-duction in the Mediterranean zone, particularly in Algeria [29],and rhizobial adaptation to drought stress is variable [5,8,40]. Inchickpea, lack of water influences rhizobium diversity, as well asreducing nodule number and aerial biomass [8]. Isolates tolerant tosalt stress are often also tolerant to water stress [47]. Large pheno-typic and genotypic diversities are often recovered among rhizobialisolates sampled in arid and semi-arid zones (i.e. in Morocco [15],Algeria [9], and Tunisia [30,54]).
Until 2008, legumes of the Viciae tribe, including pea and lentil,were considered to nodulate only with Rlv but, since then, R. pisiand R. fabae have been isolated from pea and faba bean. Most of thestudies on Rlv diversity and ecology were conducted in temperateclimate zones in Europe. To our knowledge, no study on rhizo-bia nodulating lentil and pea in Algeria has been reported in theliterature. Therefore, in this study, natural Rhizobium populationsnodulating pea and lentil varieties adapted to environmental con-straints were studied in Algeria. The structure and genetic diversityof natural rhizobial populations were examined at several sites rep-resenting two contrasting eco-climatic conditions – sub-humid andsemi-arid – in Eastern Algeria. Rhizobial isolates were characterisedby PCR/RFLP of the 16S–23S ribosomal intergenic region (16S–23SIGS) and the nodD-F symbiotic region, as well as by sequence anal-yses of 16S–23S IGS, 16S ribosomal RNA and nodD genes.
Materials and methods
Experimental sites, soil composition and cultivars
The study was carried out on three legume plants, proteaginouspea (P. sativum L. subsp. sativum var. sativum cv. messire), foragepea (P. sativum L. subsp. sativum var. arvense cv. Sefrou, see Smykalet al. [44] for new classification of Pisum) and lentil (L. culinaris L.cv. P.B. Dahra). The two pea cultivars cv. messire and cv. Sefrou wereintroduced varieties, selected in France and Morocco, respectively.
and symbiotic diversity of Rhizobium populations associ-d regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),
L. culinaris L. cv. P.B. Dahra was a local cultivar.The plants were grown in six experimental fields in East-
ern Algeria, representing sub-humid to semi-arid climatic zones(Fig. 1). Four well-separated soil samples were collected at the time

IN PRESSG ModelS
plied Microbiology xxx (2014) xxx–xxx 3
op4Ttl1Tswscfibo
E
ad
I
b4TaUntawrwms
N
ptwoAmsogwtcaiw[po1ipraw
f th
e
sam
pli
ng
site
s.
Zon
eaG
PS
coor
din
ates
Soil
typ
ebC
lay
(%)
Fin
e
silt
(%)
Coa
rse
silt
(%)
Fin
e
san
d(%
)C
oars
esa
nd
(%)
Org
anic
mat
ter
(%)
Hu
mid
ity
tosa
tura
tion
(%)
pH
EC
(mil
-li
ohm
s
cm−1
)C
arbo
n(‰
)N
itro
gen
(‰)
C/N
MPN
c×
104
(g−1
soil
)
SH
36◦ 4
6′ 08′′ N
–7◦ 5
1′ 24′′ E
LSA
25
37
12
17
7
1.29
55
7.9
0.6
7.5
0.96
8
1.2
(0.3
–4.5
)z
SH
36◦ 5
2′ 45′′ N
–7◦ 1
8′ 31′′ E
LSA
17
22
20
29
7
1.22
1
50
7.7
0.8
7.1
0.5
14
3.5
(0.9
–13.
3)h
SH
36◦ 4
2′ 27′′ N
–6◦ 4
7′ 09′′ E
L
20
45
16
6
8
2.42
5
62
8.1
1.1
14.1
1.14
12
2.1
(0.5
–7.9
)e
SA
36◦ 2
0′ 27′′ N
–6◦ 4
4′ 42′′ E
LSA
20
20
27
7
29
3.30
2
75
8.1
1.8
19.2
1.38
14
3.9
(0.1
–14.
8)an
eSA
35◦ 5
3′ 42′′ N
–6◦ 2
4′ 19′′ E
LS
14
12
7 35
27
1.54
8
50
8.1
2.1
9.0
1.14
8
2.6
(0.7
–10.
0)ag
hi
SA
35◦ 5
0′ 04′′ N
–7◦ 1
5′ 57′′ E
L
12
40
15
22
6
1.56
5
55
8.1
1.8
9.1
0.8
11
0.2
(0.0
5–0.
7)
um
id;
SA, s
emi-
arid
.sa
nd
-cla
y;
L,
silt
;
LS, s
ilt-
san
dy.
mos
t
pro
babl
e
nu
mbe
r
wit
h
the
95%
con
fid
ence
inte
rval
give
n
in
brac
kets
.
ARTICLEYAPM-25606; No. of Pages 8
N. Riah et al. / Systematic and Ap
f sowing from each site at depths of 0–30 cm. They were thenooled together, sieved through a 2 mm sieve and maintained at◦C before being used in most probable numbers (MPN) testing.able 1 shows the geographical coordinates of the sites, as well asheir physical and chemical properties. In every experimental site,entil, forage pea and proteaginous pea were each cultivated on a
m2 wide plot, at a sowing density of 100 untreated seeds per m2.he cultures were maintained without any watering. The sampledites in Oum El Bouaghi and Souk Naamane had not been cultivatedith legumes for at least 30 years. Lentil was cultivated in the Con-
tantine site two years before being sampled in the current study. Aombination of vetch with barley and wheat was grown in El-Tarefve years earlier, whereas the El-Harrouch site was cultivated withean and pea 20 years before our sampling. The cultivation historyf Ben-Azzouz was unknown.
stimation of rhizobial populations
Most probable numbers (MPN) of compatible rhizobia with peand lentil in soils were estimated by plant tests, as previouslyescribed [27].
solation of bacteria
The harvest of 12 plants per plot was performed at the earlylooming stage (12 weeks). A total of four nodules per plant (i.e.8 per plot) were collected at random and dried on filter paper.he nodules were laid on a cotton layer in glass bottles filled to
quarter of their volume capacity with CaCl2 and stored at 4 ◦C.pon use, the nodules were rehydrated in distilled water for oneight, sterilised by immersion in 95% ethanol for 5 min, and thenransferred to 0.1% acidic mercuric chloride solution for 2–4 minccording to their size. The nodules were then washed ten timesith sterile distilled water. Rhizobial strains were isolated from
oot nodules by crushing, according to Vincent [51]. The isolatesere grown on Yeast Mannitol (YM) agar medium at 28 ◦C, and thenaintained at 4 ◦C in 20% (w/v) glycerol at −80 ◦C for long-term
torage.
odulation test
The isolates were tested for nodulation on their original hostlant. Pea seeds were surface sterilised in 95% ethanol for 5 min, andhen in concentrated sulphuric acid for 5 min. The seeds were thenashed ten times and left for 2 hours in sterile water, then placed
n sterile 1% agar–water in Petri dishes at 24–25 ◦C for germination.fter 2–3 days, two seedlings were transferred to vermiculite–sandixture (4:1) in a Leonard jar moistened with Fahraeus nutrient
olution at pH 6.8 [51]. Each seedling was inoculated with 1 mLf rhizobial suspension (∼109 cells mL−1). Plants were grown inrowth chambers under conditions of 25 ◦C day and 16 ◦C nightith a 10–14 h photoperiod. Lentil culture was carried out in test
ubes (22 mm × 20 mm). The seeds were sterilised in 3% (w/v) cal-ium hypochlorite for 5 min, rinsed several times with sterile waternd finally left in water for 1 h then placed on sterile 1% agar–watern Petri dishes for germination at 25 ◦C for 2–3 days. The seedlings
ere transferred to tubes filled with Fahraeus nutrient solution51] with the addition of 1% (w/v) CaCO3 and closed with 2-hole-erforated plastic caps, one for inserting the sprout rootlet and thether for inoculation. Two days old seedlings were inoculated with
mL of a rhizobial suspension (∼107 cells mL−1). Plants were grownn growth chambers (22 ◦C day/18 ◦C night temperature, 10–16 h
Please cite this article in press as: N. Riah, et al., Genotypic and symbiotic diversity of Rhizobium populations associ-ated with cultivated lentil and pea in sub-humid and semi-arid regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),http://dx.doi.org/10.1016/j.syapm.2013.12.008
hotoperiod, 60% day/70% night humidity). Each treatment wasepeated twice. Two non-inoculated treatments were included as
control. Nodule development was followed during the first foureeks after inoculation. Ta
ble
1D
escr
ipti
on
o
Site
s
El-T
aref
Ben
-Azz
ouEl
-Har
rou
cC
onst
anti
nSo
uk
Naâ
mO
um
El
Bou
aSH
, su
b-h
bLS
A, s
ilt-
cR
hiz
obia

ING ModelS
4 plied M
P
dwsl
S
Rr(iptfp
Csn[lSa
S
tpmil(Tt(bf[
R
osfien
AR
rrIcSsts
ARTICLEYAPM-25606; No. of Pages 8
N. Riah et al. / Systematic and Ap
CR-RFLP characterisation of rhizobia
The isolates were first characterised by PCR-RFLP, as previouslyescribed [24]. Two genomic regions were amplified and digestedith the restriction enzyme HaeIII: the chromosomal intergenic
pacer region 16S–23S rRNA genes (IGS) and the plasmidic nodu-ation gene region nodD-F.
equencing and phylogenetic analysis
One representative of each haplotype detected in the PCR-FLP approach was sequenced. PCR products (IGS and nodD-Fegions) were purified using the PureLinkTM PCR Purification KitInvitrogenTM) and sent to Genoscreen© (Lille, France) for sequenc-ng. The nodD-F region was sequenced using the NODDRL2′ innerrimer [25] in addition to the two primers used for PCR amplifica-ion [24]. The 16S rRNA gene was amplified using universal primersD1 and rD1 as described by [21]. 16S rRNA gene PCR products wereartly sequenced using primer 16S-1080r [54].
The sequences obtained were checked and assembled withhromas Pro software (version 1.5), then analysed for sequenceimilarities in GenBank using the BLASTN program. The phyloge-etic analyses were performed on the www.phylogeny.fr platform14]. The phylogenetic trees were constructed by the maximumikelihood method, and indels were removed from the alignments.equences were deposited in the GenBank database under theccession numbers given in Table S1.
tatistical analyses
Tests for a significant genetic structure of Rhizobium popula-ions between the two climatic zones and among host plants wereerformed with an analysis of molecular variance (AMOVA) imple-ented in Arlequin version 2.0 software [42]. Rarefaction analysis
ndicated that the size of the sampling (number of isolates) wasarge enough to allow direct comparisons of genotype richnessnumber of types) between populations using EstimateS 8.2 [11].he number of dominant types was estimated by calculation ofhe Simpson inverse diversity index, 1/D, with D =
∑[ni (ni − 1)]/(N
N − 1)), where ni is the number of the ith type and N is the num-er of individuals in the population [23]. Significant differencesor diversity indices were tested between populations using PAST18,19].
esults and discussion
This study represents the first report on the characterisationf rhizobia nodulating pea and lentil across a climatic gradient ateveral sites in Algeria. A total number of 237 bacteria were isolatedrom root nodules of three crops that were comprised of 76–82solates from each host plant and 31–42 isolates per site. All isolatesxhibited rapid growth on YMA at 28 ◦C and were able to induceodulation on their original host plant.
ll rhizobia isolated from lentil and pea were affiliated to. leguminosarum
The PCR amplifications of the ribosomal IGS and the nodD-Fegion produced unique bands of 1000–1200 and 1100–1400 pb,espectively, and RFLP analysis distributed the 237 isolates into 14GS and 10 nod haplotypes. The combination of these haplotypeslustered the isolates into 26 different IGS/nod genotypes (Table
Please cite this article in press as: N. Riah, et al., Genotypicated with cultivated lentil and pea in sub-humid and semi-arihttp://dx.doi.org/10.1016/j.syapm.2013.12.008
2). The partial (907 bp) sequences of the 16S rRNA gene from 24trains, each representing one IGS/nod genotype, showed very lit-le variation and were identified as R. leguminosarum. Twenty twoequences were identical and the remaining two only differed by
PRESSicrobiology xxx (2014) xxx–xxx
one and two mutations, respectively (strain SL15/genotype 13 g;strain CL4/genotype 3 g).
Several studies had previously focused on the genetic diversityof rhizobia associated with legumes in the Viciae tribe across allcontinents. Most of them only isolated R. leguminosarum strains,but several could also recover other Rhizobium species, such as R.etli from V. faba and P. sativum in Peru and from P. sativum in China[52], as well as R. pisi from P. sativum [37] and several putativenew species or lineages associated with L. culinaris in Bangladesh[38]. No species other than R. leguminosarum were recovered inthe current study. This lack of diversity did not result from anabsence of other Rhizobium species in soils, since several have beenpreviously isolated in Algeria, or at a wider scale across NorthernAfrica (Morocco, Tunisia and Algeria): R. gallicum (in Algeria fromAcacia saligna [3]), R. gallicum bv. gallicum, R. etli bv. phaseoli, R.giardinii bv. giardinii (in Tunisia from P. vulgaris [31]), R. sullae fromHedysarum coronarium in Algeria [45], and R. tropici from P. vulgarisin Morocco [16]. This suggested that the recovery of the single R.leguminosarum species was rather due to true symbiotic host plantspecificity. Although some exceptions have been noticed, membersof the genera Vicia and Pisum must still be considered as highlyspecific legumes.
The genetic diversity levels were contrasted betweenchromosomal and symbiotic loci
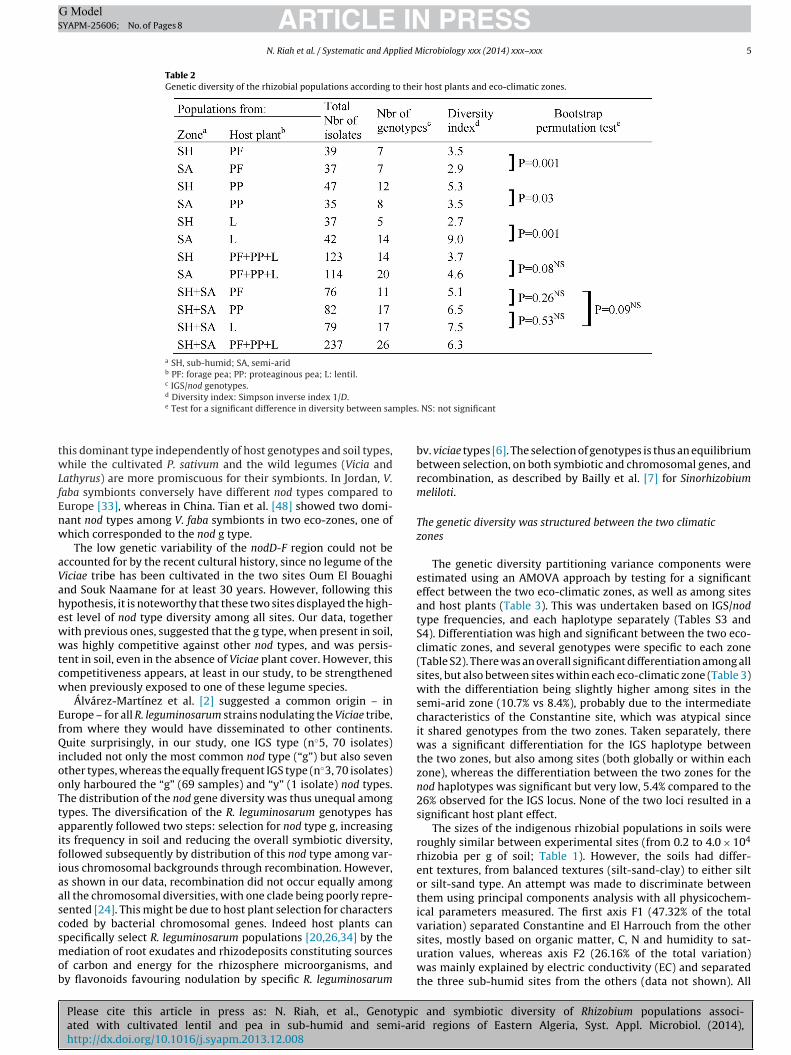
Among the 26 IGS/nod genotypes detected, five genotypes rep-resenting 5.9–29.1% of the 237 isolates (77.7% altogether, Table 2)were dominant (1/D diversity index of 6.3, Table 2). No significantdifference (p > 0.05) in diversity could be detected either betweeneach host plant, or between subpopulations of sub-humid andsemi-arid zones. Significant differences of diversity were detectedbetween semi-arid and sub-humid zones for each plant separately,with a higher diversity value in the sub-humid zone for lentil andforage pea, and the reverse for proteaginous pea.
IGS rDNA PCR-RFLP has been widely used to explore the diver-sity within R. leguminosarum bv. viciae. Palmer et al. [35] detected25 genotypes in 285 isolates using HaeIII, and considered this to bea normal range of variation for this species. Depret and Laguerre[13] detected 28 haplotypes among 1100 isolates (from a singlesite in France using HaeIII). Conversely, Mutch and Young [34] onlydetected nine different types among 625 isolates (with TaqI). Atthe other extreme, Aoki et al. [4], with only 15 isolates, detectedeight different haplotypes, but at a very large geographical scale.Our study revealed an intermediate level of chromosomal diver-sity, with 14 different genotypes among 237 isolates. However, thisdiversity was strongly unbalanced with three types (1, 3, 5) repre-senting 73.8% of the isolates. In the 16S–23S IGS rDNA phylogenyperformed for 13 strains representing the various IGS types, 12 fellinto two closely related clades (Fig. 2).
The nod type diversity was conversely rather low comparedto IGS (1/D = 1.3) with the nod “g” type representing 88% of thetotal (up to 97.5% at the El-Harrouche site) regardless of the hostisolation plant. The other nod haplotypes were only marginally rep-resented in our sampling, with one to six individuals. The nodD-Ffragment sequencing was performed for 16 strains, ten represent-ing the different nod types and six representing the main nod g type.The phylogenetic tree (Fig. S1) corroborated the results obtainedby PCR-RFLP analysis, with groupings of the strains regardless oftheir origins. The six type g samples fell in a single clade togetherwith the two closely related types v and y. The main nod “g” hap-lotype had previously been isolated elsewhere, and was similarly
and symbiotic diversity of Rhizobium populations associ-d regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),
described as a highly frequent type. In France, Laguerre et al. [24]noted that the nod g type was mainly found with V. faba, and Mutchand Young [34] reported it in the United Kingdom on P. sativum,Lathyrus and V. faba. In Europe, cultivated V. faba strongly selects
ARTICLE IN PRESSG ModelSYAPM-25606; No. of Pages 8
N. Riah et al. / Systematic and Applied Microbiology xxx (2014) xxx–xxx 5
Table 2Genetic diversity of the rhizobial populations according to their host plants and eco-climatic zones.
a SH, sub-humid; SA, semi-aridb PF: forage pea; PP: proteaginous pea; L: lentil.c IGS/nod genotypes.
ples
twLfEnw
aVahewwtcw
EfQiooTtaifiaascsmob
d Diversity index: Simpson inverse index 1/D.e Test for a significant difference in diversity between sam
his dominant type independently of host genotypes and soil types,hile the cultivated P. sativum and the wild legumes (Vicia and
athyrus) are more promiscuous for their symbionts. In Jordan, V.aba symbionts conversely have different nod types compared tourope [33], whereas in China. Tian et al. [48] showed two domi-ant nod types among V. faba symbionts in two eco-zones, one ofhich corresponded to the nod g type.
The low genetic variability of the nodD-F region could not beccounted for by the recent cultural history, since no legume of theiciae tribe has been cultivated in the two sites Oum El Bouaghind Souk Naamane for at least 30 years. However, following thisypothesis, it is noteworthy that these two sites displayed the high-st level of nod type diversity among all sites. Our data, togetherith previous ones, suggested that the g type, when present in soil,as highly competitive against other nod types, and was persis-
ent in soil, even in the absence of Viciae plant cover. However, thisompetitiveness appears, at least in our study, to be strengthenedhen previously exposed to one of these legume species.
Álvárez-Martínez et al. [2] suggested a common origin – inurope – for all R. leguminosarum strains nodulating the Viciae tribe,rom where they would have disseminated to other continents.uite surprisingly, in our study, one IGS type (n◦5, 70 isolates)
ncluded not only the most common nod type (“g”) but also seventher types, whereas the equally frequent IGS type (n◦3, 70 isolates)nly harboured the “g” (69 samples) and “y” (1 isolate) nod types.he distribution of the nod gene diversity was thus unequal amongypes. The diversification of the R. leguminosarum genotypes haspparently followed two steps: selection for nod type g, increasingts frequency in soil and reducing the overall symbiotic diversity,ollowed subsequently by distribution of this nod type among var-ous chromosomal backgrounds through recombination. However,s shown in our data, recombination did not occur equally amongll the chromosomal diversities, with one clade being poorly repre-ented [24]. This might be due to host plant selection for charactersoded by bacterial chromosomal genes. Indeed host plants can
Please cite this article in press as: N. Riah, et al., Genotypicated with cultivated lentil and pea in sub-humid and semi-arihttp://dx.doi.org/10.1016/j.syapm.2013.12.008
pecifically select R. leguminosarum populations [20,26,34] by theediation of root exudates and rhizodeposits constituting sources
f carbon and energy for the rhizosphere microorganisms, andy flavonoids favouring nodulation by specific R. leguminosarum
. NS: not significant
bv. viciae types [6]. The selection of genotypes is thus an equilibriumbetween selection, on both symbiotic and chromosomal genes, andrecombination, as described by Bailly et al. [7] for Sinorhizobiummeliloti.
The genetic diversity was structured between the two climaticzones
The genetic diversity partitioning variance components wereestimated using an AMOVA approach by testing for a significanteffect between the two eco-climatic zones, as well as among sitesand host plants (Table 3). This was undertaken based on IGS/nodtype frequencies, and each haplotype separately (Tables S3 andS4). Differentiation was high and significant between the two eco-climatic zones, and several genotypes were specific to each zone(Table S2). There was an overall significant differentiation among allsites, but also between sites within each eco-climatic zone (Table 3)with the differentiation being slightly higher among sites in thesemi-arid zone (10.7% vs 8.4%), probably due to the intermediatecharacteristics of the Constantine site, which was atypical sinceit shared genotypes from the two zones. Taken separately, therewas a significant differentiation for the IGS haplotype betweenthe two zones, but also among sites (both globally or within eachzone), whereas the differentiation between the two zones for thenod haplotypes was significant but very low, 5.4% compared to the26% observed for the IGS locus. None of the two loci resulted in asignificant host plant effect.
The sizes of the indigenous rhizobial populations in soils wereroughly similar between experimental sites (from 0.2 to 4.0 × 104
rhizobia per g of soil; Table 1). However, the soils had differ-ent textures, from balanced textures (silt-sand-clay) to either siltor silt-sand type. An attempt was made to discriminate betweenthem using principal components analysis with all physicochem-ical parameters measured. The first axis F1 (47.32% of the totalvariation) separated Constantine and El Harrouch from the other
and symbiotic diversity of Rhizobium populations associ-d regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),
sites, mostly based on organic matter, C, N and humidity to sat-uration values, whereas axis F2 (26.16% of the total variation)was mainly explained by electric conductivity (EC) and separatedthe three sub-humid sites from the others (data not shown). All

ARTICLE IN PRESSG ModelSYAPM-25606; No. of Pages 8
6 N. Riah et al. / Systematic and Applied Microbiology xxx (2014) xxx–xxx
Fig. 2. Phylogenetic tree based on a 470 bp alignment of nucleotide sequences of the rDNA 16S–23S IGS. Only bootstrap probability values ≥70% (over 100 replicates)are indicated. The IGS types isolated in this study are shown in bold, with their frequency in the two climatic zones being indicated in brackets. SH: sub-humid zone;SA: semi-arid zone. The host plant of origin and geographic origins of several strains are indicated in brackets. Rlp: Rhizobium leguminosarum symbiovar phaseoli;Rlt: Rhizobium leguminosarum symbiovar trifolii; Rlv: Rhizobium leguminosarum symbiovar viciae. The scale bar indicates the number of substitutions per site.
Table 3Genetic differentiation of the rhizobial populations according to their geographical origins and their host plants of isolation by AMOVA based on multilocus IGS/nod typefrequencies.
Source of variationa d.f. % variationb Source of variation d.f. % variationb
Among eco-climatic zones 1 17.6*** Among host plant 2 −3.35NSAmong sites 5 18.6*** Among host plants within sites 12 2.72***Among sites within SA zone 2 10.7* Among host plants within SA 6 −1.07NSAmong sites within SH zone 2 8.4** Among host plants within SH 6 −1.04NS
a
robabiv t).
sreo
ieprt
SH, sub-humid zone; SA, semi-arid zone.b Variance among populations is given as a percentage of the total variance. The p
alues by chance alone is given (*p < 0.05; **p < 0.01; ***p < 0.001; NS: not significan
oils studied exhibited similar physicochemical characteristics andhizobial population sizes, and the hypothesis of a soil effect forxplaining the genetic structure of rhizobial populations was ruledut.
Due to the small number of sites, and their non-independencen terms of relative distance (sites from each zone were closer to
Please cite this article in press as: N. Riah, et al., Genotypicated with cultivated lentil and pea in sub-humid and semi-arihttp://dx.doi.org/10.1016/j.syapm.2013.12.008
ach other than to sites of the other zone), the hypothesis of a sim-le differentiation by distance between the two zones could not beejected. The Constantine site, which was intermediate betweenhe three northern and two southern sites, would reflect such a
lity of having a more extreme variance component and F statistic than the observed
differentiation linked to the geographical distance. A similar situa-tion was clearly described by Rahi et al. [36] who reported a geneticdiscontinuity and limited gene flow in the R. leguminosarum bv.viciae pea-nodulating populations cultivated on the Indian side ofWest Himalayan.
This differentiation may finally be explained by the contrasting
and symbiotic diversity of Rhizobium populations associ-d regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),
climatic environmental constraints, mainly through low rainfallin the semi-arid climate resulting in drought stress. Resistanceto drought is a selective factor structuring rhizobial diversity andenvironmental variations impacting the structuring patterns of

ING ModelS
plied M
sMspcawbgba
Hg
wnoafoep
daaaeaTctapvosrsvtp
A
sIB
A
i1
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[isolation of Rhizobium leguminosarum from soils. Soil Biol. Biochem. 27,
ARTICLEYAPM-25606; No. of Pages 8
N. Riah et al. / Systematic and Ap
oil bacterial communities (i.e. latitude and temperature [46]).any studies have reported on the capacity of rhizobia to adapt to
everal environmental constraints, such as water stress, high tem-eratures, farming practices, soil acidity, salinity, and heavy metalontamination. Elboutahiri et al. [15] showed the large phenotypicnd genotypic diversity of S. meliloti isolates tolerant to saline andater stress sampled in arid and semi-arid zones in Morocco. Com-
ined soil and climatic effects were reported for pea in differenteographical subtropical regions in China where rhizobial distri-ution and diversity structure are directed by pH, water contentnd soil temperature [52].
ost plant had no significant influence on the recovered bacterialenetic diversity
Based on the AMOVA approach, no significant host plant effectas detected when considering the entire data set, but a sig-ificant differentiation among host plants within each site wasbserved. Differentiation tests were also performed by pairs amongll sites*host–plant combinations (data not shown). Even after Bon-erroni correction, two sites were highly differentiated from allthers, since Souk Naâmane and Oum El Bouaghi showed the high-st Fct values (p < 0.0001 for most pairs) irrespective of the hostlant species.
This lack of significant host plant effect on the bacterial geneticiversity recovered, either when considering all sites or when sep-rating the two climatic zones, was quite surprising, since severaluthors previously suggested that pea, lentil, and faba bean oper-ted some rhizobial genotype selection [20,26]. The only significantffect was detected between host plants within sites of the semi-rid zone (p < 0.0001). Two reasons might explain this lack of effect.he first one, somewhat trivial, could be the power of our statisti-al test, although the percentage variation estimated in the AMOVAests was either low or even negative, which apparently reflected
true lack of differentiation. Even the significant effect betweenlants within semi-arid sites was supported by a low percentageariation value (7.4%). The second explanation could be a true lackf any host plant effect. Considering previous studies that suggesteduch an effect, this might be due to a low local bacterial diversity,educing the possibility for the plants to select their most adaptedymbiont and thus reflecting their real specificity. Supporting thisiew, a very low nucleotidic divergence was detected among nodDypes, which contrasted with other studies where high nodD genehylogenetic diversity was highlighted.
cknowledgements
This work was supported by the French-Algerian program forcience (PROFAS). The authors are grateful to Naima Rezkallah andsabelle Babic for technical assistance, and thank Lionel Moulin andrigitte Brunel for helpful comments on the manuscript.
ppendix A. Supplementary data
Supplementary data associated with this article can be found,n the online version, at http://dx.doi.org/10.1016/j.syapm.2013.2.008.
eferences
[1] Abdelmoumen, H., Filali-Maltouf, A., Neyra, M., Belabed, A., Missbah El Idrissi,M. (1999) Effect of high salts concentrations on the growth of rhizobia andresponses to added osmotica. J. Appl. Microbiol. 86, 889–898.
Please cite this article in press as: N. Riah, et al., Genotypicated with cultivated lentil and pea in sub-humid and semi-arihttp://dx.doi.org/10.1016/j.syapm.2013.12.008
[2] Álvarez-Martínez, E.R., Valverde, A., Ramirez-Bahena, M.H., Garcia-Fraile, P.,Tejedor, C., Mateos, P.F., Santillana, N., Zuniga, D., Peix, A., Velazquez, E. (2009)The analysis of core and symbiotic genes of rhizobia nodulating Vicia fromdifferent continents reveals their common phylogenetic origin and suggests
[
PRESSicrobiology xxx (2014) xxx–xxx 7
the distribution of Rhizobium leguminosarum strains together with Vicia seeds.Arch. Microbiol. 191, 659–668.
[3] Amrani, S., Noureddine, N.E., Bhatnagar, T., Argandona, M., Nieto, J.J., Vargas, C.(2010) Phenotypic and genotypic characterization of rhizobia associated withAcacia saligna (Labill.) Wendl. in nurseries from Algeria. Syst. Appl. Microbiol.33, 44–51.
[4] Aoki, S., Kondo, T., Prévost, D., Nakata, S., Kajita, T., Ito, M. (2010) Genotypic andphenotypic diversity of rhizobia isolated from Lathyrus japonicus indigenous toJapan. Syst. Appl. Microbiol. 33, 383–397.
[5] Athar, M., Johnson, D.A. (1997) Effect of drought on the growth and survivalof Rhizobium meliloti strains from Pakistan and Nepal. J. Arid. Environ. 35,335–340.
[6] Badri, D.V., Vivanco, J.M. (2009) Regulation and function of root exudates. PlantCell Environ. 32, 666–681.
[7] Bailly, X., Olivieri, I., De Mita, S., Cleyet-Marel, J.C., Béna, G. (2006) Recombi-nation and selection shape the molecular diversity pattern of nitrogen fixingSinorhizobium sp associated to Medicago. Mol. Ecol. 15, 2719–2734.
[8] Ben Romdhane, S., Trabelsi, M., Aouani, M.E., de Lajudie, P., Mhamdi, R. (2009)The diversity of rhizobia nodulating chickpea (Cicer arietinum) under waterdeficiency as a source of more efficient inoculants. Soil Biol. Biochem. 41,2568–2572.
[9] Boukhatem, Z.F., Domergue, O., Bekki, A., Merabet, C., Sekkour, S., Bouazza, F.,Duponnois, R., de Lajudie, P., Galiana, A. (2012) Symbiotic characterization anddiversity of rhizobia associated with native and introduced acacias in arid andsemi-arid regions in Algeria. FEMS Microbiol. Ecol. 80, 534–557.
10] Brink, M., Belay, G. 2006 Céréales et légumes secs, ressources végétales del’Afrique tropicale, Fondation Prota, Wageningen Pays-Bas, pp. , 102.
11] Colwell, R.K. 2013 EstimateS: Statistical estimation of species richnessand shared species from samples. Version 8.2 http://viceroy.eeb.uconn.edu/estimateS
12] Depret, G., Houot, S., Allard, M.R., Breuil, M.C., Nouaïm, R., Laguerre, G. (2004)Long-term effects of crop management on Rhizobium leguminosarum biovarviciae populations. FEMS Microbiol. Ecol. 51, 87–97.
13] Depret, G., Laguerre, G. (2008) Plant phenology and genetic variability in rootand nodule development strongly influence genetic structuring of Rhizobiumleguminosarum biovar viciae populations nodulating pea. New Phytol. 179,224–235.
14] Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., Dufayard,J.F., Guindon, S., Lefort, V., Lescot, M., Claverie, J.M., Gascuel, O. (2008) Phy-logeny. fr: robust phylogenetic analysis for the non-specialist. Nucleic AcidsRes. 36, 465–469.
15] Elboutahiri, N., Thami-Alami, I., Udupa, S.M. (2010) Phenotypic and geneticdiversity in Sinorhizobium meliloti and S. medicae from drought and salt affectedregions of Morocco. BMC Microbiol. 10, 15.
16] Faghire, M., Mandri, B., Oufdou, K., Bargaz, A., Ghoulam, C., Ramírez-Bahena,M.H., Velázquez, E., Peix, A. (2012) Identification at the species and symbiovarlevels of strains nodulating Phaseolus vulgaris in saline soils of the Marrakechregion (Morocco) and analysis of the otsA gene putatively involved in osmotol-erance. Syst. Appl. Microbiol. 35, 156–164.
17] FAO, 2006 Deuxième rapport national sur l’état des ressources phytogéné-tiques, INRAA. FAO/ONU FA.
18] Hammer, Ø., Harper, D.A.T., Ryan, P.D. (2001) PAST: paleontological statisticssoftware package for education and data analysis. Palaeontologia Electronica 4(1), 9.
19] Hill, M.O. (1973) Diversity and evenness: a unifying notation and its conse-quences. Ecology. 54, 427–432.
20] Hynes, M.F., O’Connell, M.P. (1990) Host plant effect on competition amongstrains of Rhizobium leguminosarum. Can. J. Microbiol. 36, 864–869.
21] Laguerre, G., Allard, M.R., Revoy, F., Amarger, N. (1994) Rapid identificationof rhizobia by restriction fragment length poly-morphism analysis of PCR-amplified 16S rRNA genes. Appl. Environ. Microbiol. 60, 56–63.
22] Laguerre, G., Courde, L., Nouaïm, R., Lamy, I., Revellin, C., Breuil, M.C., Chaussod,R. (2006) Response of rhizobial populations to moderate copper stress appliedto an agricultural soil. Microb. Ecol. 52, 426–435.
23] Laguerre, G., Geniaux, E., Mazurier, S.I., Rodriguez-Casartelli, R., Amarger,N. (1993) Conformity and diversity among field isolates of Rhizobiumleguminosarum bv. viciae, bv. trifolii, and bv. phaseoli revealed by DNAhybridization using chromosome and plasmid probes. Can. J. Microbiol. 39,412–419.
24] Laguerre, G., Louvrier, P., Amarger, N. (2003) Compatibility of rhizobial geno-types within natural populations of Rhizobium leguminosarum biovar viciae fornodulation of host legumes. Appl. Environ. Microbiol. 69, 2276–2283.
25] Laguerre, G., Mavingui, P., Allard, M.R., Charnay, M.P., Louvrier, P., Mazurier, S.I.,Rigottier-Gois, L., Amarger, N. (1996) Typing of rhizobia by PCR DNA finger-printing and PCR-RFLP analysis of chromosomal and symbiotic gene regions:application to Rhizobium leguminosarum and its different biovars. Appl. Environ.Microbiol. 62, 2029–2036.
26] Laguerre, G., Mazurier, S.I., Amarger, N. (1992) Plasmid profiles and restrictionfragment length polymorphism of Rhizobium leguminosarum bv. viciae in fieldpopulations. FEMS Microbiol. Ecol. 101, 17–26.
27] Louvrier, P., Laguerre, G., Amarger, N. (1995) Semiselective medium for
and symbiotic diversity of Rhizobium populations associ-d regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),
919–924.28] Louvrier, P., Laguerre, G., Amarger, N. (1996) Distribution of symbiotic geno-
types in Rhizobium leguminosarum biovar viciae populations isolated directlyfrom soils. Appl. Environ. Microbiol. 62, 4202–4205.

ING ModelS
8 plied M
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[community DNA reveals the genetic diversity of Rhizobium leguminosarum in
ARTICLEYAPM-25606; No. of Pages 8
N. Riah et al. / Systematic and Ap
29] L’Taief, B., Sifi, B., Zaman-Allah, M., Hajji, M., Lachaâl, M. (2009) Effets de lafertilisation azotée, de l’inoculation par Rhizobium sp. et du régime des pluiessur la production de la biomasse et la teneur en azote du pois chiche. Biotechnol.Agron. Soc. Environ. 13, 537–544.
30] Merabet, C., Martens, M., Mosbah, M., Zakhia, F., Sy, A., Le Roux, C., Domergue,O., Coopman, R., Bekki, A., Mars, M., Willems, A., de Lajudie, P. (2010) Multilocussequence analysis of root nodule isolates from Lotus arabicus (Senegal), Lotuscreticus, Argyrolobium uniflorum and Medicago sativa (Tunisia) and descriptionof Ensifer numidicus sp. nov. and Ensifer garamanticus sp. nov. Int. J. Syst. Evol.Microbiol. 60, 664–674.
31] Mhamdi, R., Laguerre, G., Aouani, M.E., Mars, M., Amarger, N. (2002) Differ-ent species and symbiotic genotypes of field rhizobia can nodulate Phaseolusvulgaris in Tunisian soils. FEMS Microbiol. 41, 77–84.
32] Moschetti, G., Peluso, A.L., Protopapa, A., Anastasio, M., Pepe, O., Defez, R. (2005)Use of nodulation pattern, stress tolerance, nodC gene amplification, RAPD-PCR and RFLP – 16S rDNA analysis to discriminate genotypes of Rhizobiumleguminosarum biovar viciae. Syst. Appl. Microbiol. 28, 619–631.
33] Mutch, L.A., Tamimi, S.M., Young, J.P.W. (2003) Genotypic characterisation ofrhizobia nodulating Vicia faba from the soils of Jordan: a comparison with UKisolates. Soil Biol. Biochem. 35, 709–714.
34] Mutch, L.A., Young, J.P.W. (2004) Diversity and specificity of Rhizobium legumi-nosarum biovar viciae on wild and cultivated legumes. Mol. Ecol. 13, 2335–2344.
35] Palmer, K.M., Young, J.P.W. (2000) Higher diversity of Rhizobium leguminosarumbv. viciae populations in arable soils than in grass soils. Appl. Environ. Microbiol.66, 2445–2450.
36] Rahi, P., Kapoor, R., Young, J.P.W., Gulati, A. (2012) A genetic discontinuity inroot-nodulating bacteria of cultivated pea in the Indian trans-Himalayas. Mol.Ecol. 21, 145–159.
37] Ramírez-Bahena, M.H., García-Fraile, P., Peix, A., Valverde, A., Rivas, R., Igual,J.M., Mateos, P.F., Martínez-Molina, E., Velázquez, E. (2008) Revision of thetaxonomic status of the species Rhizobium leguminosarum (Frank 1879) Frank1889AL, Rhizobium phaseoli Dangeard 1926AL and Rhizobium trifolii Dangeard1926AL. R. trifolii is a later synonym of R. leguminosarum. Reclassification of thestrain R. leguminosarum DSM 30132 (=NCIMB 11478) as Rhizobium pisi sp. nov.Int. J. Syst. Evol. Microbiol. 58, 2484–2490.
38] Rashid, M.H., Schäfer, H., Gonzalez, J., Wink, M. (2012) Genetic diversity of rhi-zobia nodulating lentil (Lens culinaris) in Bangladesh. Syst. Appl. Microbiol. 35,98–109.
39] Rashid, M.H., Gonzalez, J., Young, J.P.W., Wink, M. (2014) Rhizobium legumi-nosarum is the symbiont of lentils in the Middle East and Europe but not inBangladesh. FEMS Microbiol. Ecol. 87 (1), 64–77.
40] Rehman, A., Nautiyal, C.S. (2002) Effect of drought on the growth and survival ofthe stress-tolerant bacterium Rhizobium sp. NBRI2505 sesbania and its drought-sensitive transposon Tn5 mutant. Curr. Microbiol. 45, 368–377.
Please cite this article in press as: N. Riah, et al., Genotypicated with cultivated lentil and pea in sub-humid and semi-arihttp://dx.doi.org/10.1016/j.syapm.2013.12.008
41] Rogel, M.A., Ormeno-Orrillo, E., Romero, E.M. (2011) Symbiovars in rhizobiareflect bacterial adaptation to legumes. Syst. Appl. Microbiol. 34, 96–104.
42] Schneider, S., Roessli, D., Excoffier, L. 2000 Arlequin ver. 2.000: a Softwarefor Population Data Analysis, Genetics and Biometry Laboratory, University ofGeneva, Switzerland.
[
PRESSicrobiology xxx (2014) xxx–xxx
43] Segovia, L., Young, J.P.W., Martínez-Romero, E. (1993) Reclassification of Amer-ican Rhizobium leguminosarum biovar phaseoli type I strains as Rhizobium etlisp. nov. Int. J. Syst. Bacteriol 43, 374–377.
44] Smykal, P., Kenicer, G., Flavell, A.J., Corander, J., Kosterin, O., Redden, R.J., Ford,R., Coyne, C.J., Maxted, N., Ambrose, M.J., Ellis, N.T.H. (2011) Phylogeny, phy-logeography and genetic diversity of the Pisum genus. Plant Genet. Resour. 9,4–18.
45] Squartini, A., Struffi, P., Döring, H., Selenska-Pobell, S., Tola, E., Giacomini, A.,Vendramin, E., Velázquez, E., Mateos, P.F., Martínez-Molina, E., Dazzo, F.B.,Casella, S., Nuti, M.P. (2002) Rhizobium sullae sp. nov. (formerly Rhizobium‘hedysari’): the root-nodule microsymbiont of Hedysarum coronarium L. Int. J.Syst. Evol. Microbiol. 52, 1267–1276.
46] Staddon, W.J., Trevors, J.T., Duchesne, L.C., Colombo, C. (1998) Soil micro-bial diversity and community structure across a climatic gradient in westernCanada. Biodivers. Conserv. 7, 1081–1092.
47] Thami-Alami, I., Elboutahiri, N., Udupa, S.M. (2010) Variability in natural popu-lations of Sinorhizobium meliloti in Morocco. In: Porqueddu, C., Ríos, S. (Eds.),The contributions of grasslands to the conservation of Mediterranean biodiver-sity (Options Méditerranéennes: Série A. Séminaires Méditerranéens; n. 92),CIHEAM/CIBIO/FAO/SEE, Zaragoza, pp. P265–P269.
48] Tian, C.F., Wang, E.T., Han, T.X., Sui, X.H., Chen, W.X. (2007) Genetic diversity ofrhizobia associated with Vicia faba in three ecological regions of China. Arch.Microbiol. 188, 273–282.
49] Tian, C.F., Wang, E.T., Wu, L.J., Han, T.X., Chen, W.F., Gu, C.T., Gu, J.G., Chen, W.X.(2008) Rhizobium fabae sp. nov., a bacterium that nodulates Vicia faba. Int. J.Syst. Evol. Microbiol. 58, 2871–2875.
50] Vessey, J.K., Chemining’wa, G.N. (2006) The genetic diversity of Rhizobium legu-minosarum bv. viciae in cultivated soils of the eastern Canadian prairie. Soil Biol.Biochem. 38, 153–163.
51] Vincent, J.M. 1970 A Manual for the Practical Study of Root-Nodule Bacteria.IBP Handbook No. 15, Blackwell Scientific Publications Ltd., Oxford, UnitedKingdom.
52] Yang, C., Yang, J., Li, Y., Zhou, J. (2008) Genetic diversity of root-nodulatingbacteria isolated from pea (Pisum sativum) in subtropical regions of China. Sci.China C Life Sci. 51, 854–862.
53] Zahran, H.H. (1999) Rhizobium-legume symbiosis and nitrogen fixation undersevere conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 63,968–989.
54] Zakhia, F., Jeder, H., Willems, A., Gillis, M., Dreyfus, B., de Lajudie, P. (2006)Diverse bacteria associated with root nodules of spontaneous legumes inTunisia and first report for nifH-like gene within the genera Microbacteriumand Starkeya. Microb. Ecol. 5, 375–393.
55] Zézé, A., Mutch, L.A., Young, J.P.W. (2001) Direct amplification of nodD from
and symbiotic diversity of Rhizobium populations associ-d regions of Eastern Algeria, Syst. Appl. Microbiol. (2014),
soil. Environ. Microbiol. 3, 363–370.56] Zhang, X.X., Kosier, B., Priefer, U.B. (2001) Genetic diversity of indigenous Rhi-
zobium leguminosarum bv. viciae isolates nodulating two different host plantsduring soil restoration with alfalfa. Mol. Ecol. 10, 2297–2305.