‘ghost’ experiments and the dissection of social learning ... (2010).pdf · of the...
TRANSCRIPT

Biol. Rev. (2010), pp. 000–000. 1doi: 10.1111/j.1469-185X.2010.00120.x
‘Ghost’ experiments and the dissectionof social learning in humans and animals
Lydia M. Hopper∗Department of Psychology, Georgia State University, Atlanta, GA, USA
(Received 22 June 2009; revised 17 December 2009; accepted 3 January 2010)
ABSTRACT
The focus of this review is the experimental techniques used to identify forms of social learning shown by humans andnonhuman animals. Specifically, the ‘ghost display’ and ‘end-state’ conditions, which have been used to tease apartimitative and emulative learning are evaluated. In a ghost display, the movements of an apparatus are demonstrated,often through the discrete use of fishing-line or hidden mechanisms, without a live model acting directly upon theapparatus so that the apparatus appears to be operated as if by a ‘ghostly’ agent. In an end-state condition, an observingindividual is shown the initial state of the test apparatus, the apparatus is then manipulated out-of-sight and thenrepresented to the individual in its final state. The aim of the ghost display condition is to determine whether individualsare able to emulate by replicating the movements of an apparatus, or perform a task, without requiring informationabout the bodily movements required to do so (imitation). The end-state condition is used to identify goal-emulation byassessing whether the observer can replicate the steps required to solve the task without having been shown the requiredbody actions or task movements. The responses of individuals tested with either the ghost display and/or end-stateconditions are compared to those of further individuals who have observed a full demonstration by either a humanexperimenter or a conspecific. The responses of a control group, to whom no information has been provided aboutthe test apparatus or required actions, are also compared and evaluated. The efficacy of these experimental techniquesemployed with humans, nonhuman primates, dogs, rats and birds are discussed and evaluated. The experimentsreviewed herein emphasise the need to provide ghost displays and end-state conditions in combination, along with fulllive demonstrations and a no-information control. Future research directions are proposed.
Key words: ghost condition, end-state condition, imitation, emulation, social learning, experimental technique,observational learning
CONTENTS
I. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2II. Forms of social learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
(1) Imitation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3(2) Emulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
(a) Goal-emulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3(b) Affordance learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3(c) Object movement reenactment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
(3) Simpler forms of social learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3(a) Contagion, mere presence effect and response facilitation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3(b) Local and stimulus enhancement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
III. Methods for identifying observational learning mechanisms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4(1) The two-action versus the bidirectional methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4(2) Ghost display conditions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
(a) Ghost display conditions employed with humans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6(b) Ghost display conditions employed with nonhuman primates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
* Address for correspondence: E-mail: [email protected]
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

2 Lydia M. Hopper
(c) Ghost display conditions employed with dogs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9(d) Ghost display conditions employed with rats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10(e) Ghost display conditions employed with birds . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
(3) End-state conditions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12IV. Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
(1) Ghost display condition evaluation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13(2) End-state condition evaluation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14(3) What forms of learning have been identified through the use of ghost display and end-state conditions? 14
V. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15VI. Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
VII. References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
I. INTRODUCTION
There is a growing interest in the use of experimentaltechniques to identify social learning mechanisms evidencedby humans and nonhuman animals (hereafter termed‘animals’). This review will evaluate the merit and efficacyof methodological techniques used for this purpose. Of anumber of approaches that have been used by developmentaland comparative psychologists, two are the foci of this presentreview – ‘ghost’ display and ‘end-state’ conditions – both ofwhich remove the agent from the demonstration. Thesemethods have been selected for discussion because not only istheir use for dissecting social learning mechanisms becomingincreasingly popular but they have also been used with anumber of species allowing for a more in-depth and wide-reaching evaluation. However before these methods can bedescribed, the social learning mechanisms they are used toidentify will first be defined.
Humans and animals have been shown to be capableof social learning. This describes a process through whichinformation is transmitted between individuals, and downgenerations, by a process of learning, rather than inheritedgenetically (Mesoudi & Whiten, 2004, 2008; for reviews of theefficiency of social learning see Laland, 2004; Kendal et al.,2005). Boyd & Richerson (1985) proposed that social learningwould be beneficial when the costs accrued from individuallearning outweigh those of social learning (Templeton &Giraldeau, 1996) and/or when the environment is relativelystable (Galef & Whiskin, 2004), both of which have beensupported empirically (see Kendal et al., 2005 for a review;but see Eriksson, Enquist & Ghirlanda, 2007).
Social interactions that facilitate observational learning arecommon within the daily experiences of humans and othersocial animals. Even at an early age, both humans (Meltzoff& Moore, 1977; Meltzoff, 1988; Uzgiris, 1991; Want &Harris, 2002) and primates (Maestripieri, Ross & Megna,2002; Biro et al., 2003; Matsuzawa, Tomonaga & Tanaka,2006) have shown evidence of social learning. It has beenproposed that social learning by humans has evolved becauseit allows for the development of ‘cultures’, an advantageousadaptation (Uzgiris, 1991, 1993; Boyd & Richerson, 1996).Whether animals also show evidence for culture is an oftdisputed argument (Laland & Galef, 2009). If ‘culture’ isdefined as ‘‘multiple traditions, spanning different domains
of behaviour, such as foraging techniques and social customs’’(Whiten & van Schaik, 2007, p. 605), however, then it hasbeen described, not just among humans, but also nonhumanprimates including wild chimpanzees (Pan troglodytes, Whitenet al., 1999, 2001), orangutans (Pongo pygmaeus, van Schaiket al., 2003; van Schaik & Pradham, 2003; Russon et al.,2008), capuchins (Cebus capucinus, Perry, 2008) and Japanesemacaques (Macaca fuscata, Leca, Gunst & Huffman, 2007,2008) as well as other nonprimate species (see Laland &Galef, 2009).
In each of these species, when an individual observesanother perform a novel behaviour there are numerous waysin which the information can be acquired. The fidelity oftransmission depends upon the observers’ understanding ofwhat they see and to what they attend. The critical ques-tion is to ask how this learning occurs (Want & Harris,2002; Whiten et al., 2004). The focus of this review is todiscuss methods used to identify underlying mechanisms ofsocial learning, allowing for distinctions between imitationand emulation in particular. These forms of social learn-ing are defined and discussed in Section II and the ghostdisplay and end-state conditions employed to identify themare described and evaluated in Section III. The Discus-sion, Section IV, will consider the efficacy of these methodsfor researching observational learning mechanisms. Futureconsiderations and research directions are proposed inSection V.
II. FORMS OF SOCIAL LEARNING
Many classifications have been proposed to describe theforms of social learning evidenced by humans and animalsand, for clarity, an overview of the major definitions will bepresented. This overview does not aim be exhaustive, butmerely to note those definitions that will be later referredto throughout this review (for more detailed definitions anddiscussion see Whiten & Ham, 1992; Heyes, 1993; Nicol,1995; Byrne & Russon, 1998; Call & Carpenter, 2002, 2003;Zentall, 2003, 2006; Hopper et al., 2008; Whiten et al., 2009).Put simply, individuals can either replicate the bodily actionsof others (imitation) or the outcomes and goals of their actions(emulation).
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

Identifying social learning mechanisms 3
(1) Imitation
Whiten & Ham (1992) defined imitation simply as a processin which ‘‘B learns some aspect(s) of the intrinsic form ofan act from A’’ (p. 250). Others consider that imitationincorporates an understanding of the model’s intentions(Tomasello, 1999; Horowitz, 2003; Bellagamba, Camaioni& Calonnesi, 2006; Huang, Heyes & Charman, 2006).Heyes (1998) proposed that the act of imitating another’sphysical actions is potential evidence of higher intelligence,as along with Boyd & Richerson (1996), Heyes (1998)suggests that perspective-taking is an essential element oftrue imitation. Perspective-taking implies an understandingof the demonstrator’s goals (Gattis, 2002) and Bekkering.Wohlschlager & Gattis (2000) suggested that imitation bychildren is mediated by perceived goals (see also Bekkering& Prinz, 2002). Following Bekkering et al. (2000) andGleissner, Meltzoff & Bekkering (2000), Wohlschlager, Gattis& Bekkering (2003) developed a theory of goal-directedimitation (GOADI). According to the GOADI theory a‘‘similarity between the movement of the model and theimitator is only superficial and incidental: the underlyingsimilarity is a similarity of action goals’’ (Wohlschlager et al.,2003, p. 512). Zentall (2001), however, argued that becauseimitation has been shown in ‘‘species as varied as rats,pigeons, and Japanese quail. . . the responsible mechanism isnot likely to be theory of mind or perspective taking. But incases in which stimulus matching is inadequate to account forimitation, some precursor of perspective taking is likely to beinvolved’’ (p. 85). Following Zentall (2001), for the purposeof this review, imitation will be defined following Whiten &Ham (1992).
(2) Emulation
Emulation was first defined by Wood (1989) to describe theresponses of children who achieved the same end-state of ademonstrated action but used a novel way to reach that goal.Emulative learning can be subdivided into three specificforms: ‘goal emulation’, ‘affordance learning’ and ‘objectmovement reenactment’ (Byrne, 2002a; Custance, Whiten& Fredman, 1999; Huang & Charman, 2005).
(a) Goal-emulation
An example of goal-emulation was provided by Tomasello(1990, referring to Tomasello et al., 1987). Young chimpan-zees saw either another chimpanzee use a tool in one oftwo specific ways to retrieve an out-of-reach food item or aninactive chimpanzee not using the tool. When presented withthe same task, the chimpanzees (aged four to six years old)that had observed a successful conspecific also used the toolto access the food. However they did not replicate the exactbehavioural strategies demonstrated. Instead they learnt thatthe tool could be used to retrieve the food, but not necessarilythe precise manner with which the model chimpanzees hadused it. Specifically, this form of emulation can be classed asgoal-emulation because the chimpanzees attained the samegoal through a novel method (Tomasello, 1990, 1999).
(b) Affordance learning
Affordance learning describes what an individual learnsabout the physical properties of the environment andrelations among objects (Byrne, 1998). An example ofaffordance learning was proposed by Zentall & Akins(2001) to describe the findings of Bugnyar & Huber (1997).Marmosets (Callithrix jacchus) observed cage mates entering abox, to retrieve food, from behind a door which could eitherbe pulled or pushed. Observers showed a tendency to usethe same method as that which they observed. Rather thanrevealing imitation, as Bugnyar & Huber (1997) proposed,Zentall & Akins (2001) suggested that the marmosets mighthave learnt about the affordances of the door (‘it can bepulled’ or ‘it can be pushed’).
(c) Object movement reenactment
In their review of social learning mechanisms, Whiten et al.(2004) were careful to distinguish imitation from objectmovement re-enactment (OMR; Custance et al., 1999).Whiten et al. (2004) defined OMR as ‘‘copying what theobject does’’ (p. 39), distinct from imitation which requiresthe observer to replicate the bodily movements of thedemonstrator. Caution should be maintained when reportingevidence of imitation, as OMR may often be a moreparsimonious explanation.
(3) Simpler forms of social learning
Although this review will focus on methods used to distinguishimitation and emulation, acknowledgement should also bemade of other forms of social learning.
(a) Contagion, mere presence effect and response facilitation
Contagion describes the phenomenon by which species-typical behaviours are elicited from an animal when itobserves another (Thorpe, 1963), for example when ananimal eats after observing a conspecific feeding, even if theobserver is satiated (Tolman, 1964).
Mere presence effect describes how the presence of aconspecific can affect the ability and speed with which anobserver can acquire a new behaviour. The presence of aconspecific may not necessarily facilitate learning, and indeedZajonc (1965) believed that the presence of a conspecific wasdetrimental to the observer. However, Caldwell & Whiten(2003) found that the presence of scrounging conspecificsfacilitated observational learning by marmosets.
Response facilitation (Byrne, 1994), describes theincreased probability of an animal to perform a familiar(not novel) act that it has observed a conspecific perform.Crucially, Byrne (1994) noted, response facilitation differsfrom imitation because the actions done are already inthe repertoire of the observing animal (but see Hoppitt,Blackburn & Laland, 2007). As has often been pointed out,defining what is a ‘novel’ action for a particular animal orindividual is a notoriously difficult task.
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

4 Lydia M. Hopper
(b) Local and stimulus enhancement
Local enhancement and stimulus enhancement occur whenan individual’s attention is drawn to a particular localeor object merely by the presence of another animal, or thechange in the environment resulting from such an interaction(Whiten & Ham, 1992; see also Zentall, 2001 for a review).Local enhancement refers to when an animal’s attention isdrawn to a particular location. Stimulus enhancement refersto when the observer’s attention is focused on a specific partof an apparatus or object.
III. METHODS FOR IDENTIFYINGOBSERVATIONAL LEARNING MECHANISMS
As highlighted above, there are a number of ways anindividual can gain new information by observing another,and experimental techniques have been developed to helpidentify and distinguish these. Specifically, this review willprovide an overview of experiments that have employedghost displays (Fawcett, Skinner & Goldsmith, 2002) and/orend-state (Meltzoff, 1985) conditions with both humans andanimals (Table 1). These methods have been used with theaim to distinguish forms of social learning, in particularseparating imitative from emulative learning. Further tothis, it is useful first to describe and define the two-action(Dawson & Foss, 1965) and bidirectional (Heyes & Dawson,1990) methods that are commonly used in such experiments.
(1) The two-action versus the bidirectional methods
To test for imitation, Dawson & Foss (1965) developeda two-action method in which the goal of the task wasalways the same, but the behaviour required to achieve itcould be demonstrated in more than one way. Imitationcould then be determined if the observer used the samebody movement as the model observed, to complete thetask. In Dawson & Foss’s (1965) original study, budgerigars(Melopsittacus undulatus) observed conspecifics remove a squareof card (lid) from a dish containing food using one of threemethods; ‘edged off by the beak’, ‘lifted off by the beak’ or‘used foot to dislodge the square’. The budgerigars tendedto use the same method as that demonstrated, but due to thepaucity of detail provided in the report, the conclusion shouldbe regarded with caution (Galef, Manzig & Field, 1986).
However Zentall & Akins (2001) criticised the two-actionmethod, focusing on the conclusions drawn by Galef et al.(1986) who replicated Dawson & Foss’ design. Zentall &Akins (2001) stated that as the lid moved in different ways(slide off versus lift off) the observing birds may actually havelearnt about the affordances of the lid rather than matchedthe model’s behaviour. To overcome such limitations, Zentalland colleagues designed experiments in which a pedal couldbe depressed by either a bird’s beak or foot. Thus, theresultant movement of the target object was the same forboth actions (Akins & Zentall, 1996; Zentall, Sutton &
Sherbourne, 1996; Kaiser, Zentall & Galef, 1997, but seeHoppitt et al., 2007).
A bidirectional task, by contrast, is composed of anapparatus with a manipulandum that can be moved in one oftwo directions; left/right or up/down. This has the advantageof being able to control against stimulus enhancement insofaras the demonstrator always interacts with the same part ofthe device. If a subject replicates the movement of this onetarget, without the presence of a demonstrator, it is likelythat they have relied on object movement reenactment.
(2) Ghost display conditions
In a ghost display (coined by Fawcett et al., 2002) the pertinentparts of an apparatus or tool are moved discretely andsurreptitiously by the experimenter; the apparatus appears tomove as if operated by a ‘ghostly’ agent. This is often achievedthrough the use of hidden mechanisms or fishing line. Whendescribing emulation, Byrne (2002b) stated ‘‘the importantdistinction from other kinds of imitation is that motorbehavior per se is not copied. Thus, in principle, emulationcould be as effective if the behavior were not seen’’ (p. 90).It is following this rationale that ghost display conditionswere developed, as they allow for the demonstration of themovements of an apparatus without a live demonstratoracting on it.
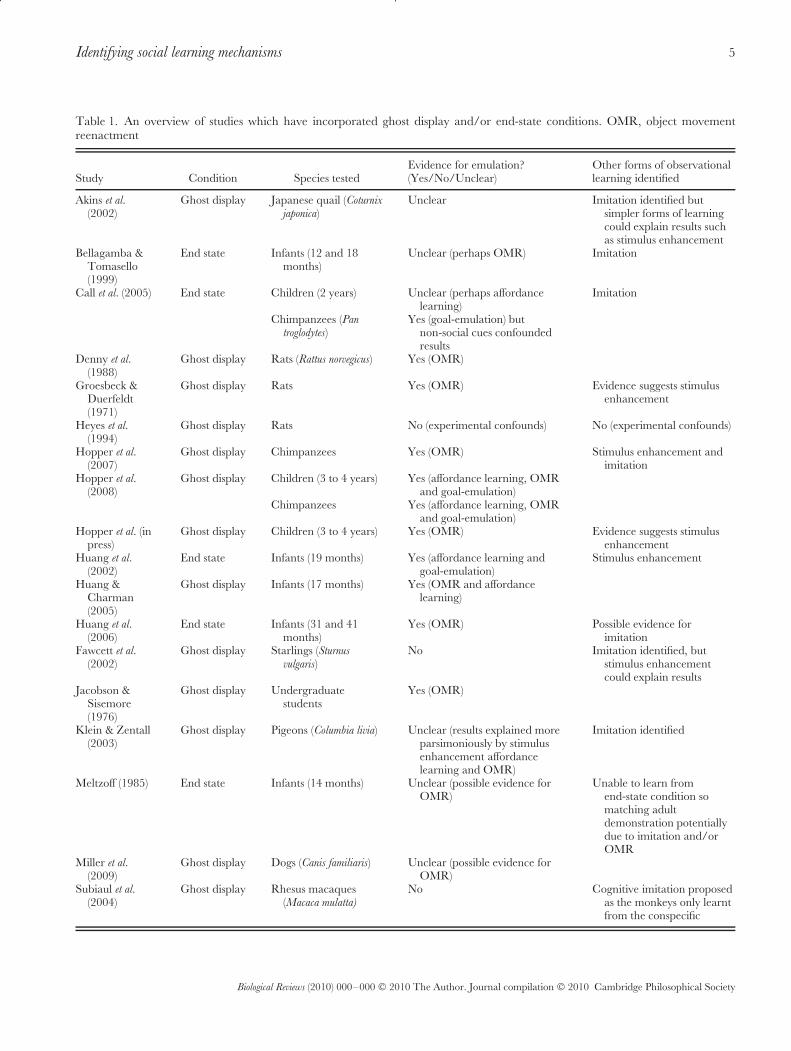
To date, ghost display conditions have been employed withhumans (Jacobson & Sisemore, 1976; Thompson & Russell,2004; Tennie, Call & Tomasello, 2006; Subiaul et al., 2007;Hopper et al., 2008, in press), nonhuman primates (Subiaulet al., 2004; Tennie et al., 2006; Hopper et al., 2007, 2008),dogs (Canis familiaris, Miller, Rayburn-Reeves & Zentall,2009), rats (Rattus norvegicus, Groesbeck & Duerfeldt, 1971;Denny, Clos & Bell, 1988; Heyes et al., 1994) and birds(Akins, Klein & Zentall, 2002; Fawcett et al., 2002; Klein &Zentall, 2003). These provide a growing corpus of literatureon the abilities of these species to learn emulatively (Table 1provides an overview). The fact that this method has nowbeen used with such a diverse number of species allows areview of the ghost condition to be particularly informative.
Before a discussion of the ghost display condition can bemade, the distinction between goal-emulation and objectmovement reenactment (OMR) should again be defined asthe conclusions drawn from studies that have presented ghostdisplays have often failed to acknowledge this distinction. Asdescribed in Section II, goal-emulation is a form of emulativebehaviour in which the individual replicates the outcomeof an action regardless of whether he/she use the samephysical behaviour or not. Indeed, Wood’s (1989) originaldefinition of emulation was that children try to emulate others‘‘by achieving similar ends or objectives’’ (p.71). OMR, bycontrast, refers to an individual replicating the movementsof the apparatus to achieve the same end state. Through theuse of ghost and end-state conditions in combination stepscan be made to distinguish goal-emulation and OMR withgreater confidence as will be discussed below.
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society
Identifying social learning mechanisms 5
Table 1. An overview of studies which have incorporated ghost display and/or end-state conditions. OMR, object movementreenactment
Study Condition Species testedEvidence for emulation?(Yes/No/Unclear)
Other forms of observationallearning identified
Akins et al.(2002)
Ghost display Japanese quail (Coturnixjaponica)
Unclear Imitation identified butsimpler forms of learningcould explain results suchas stimulus enhancement
Bellagamba &Tomasello(1999)
End state Infants (12 and 18months)
Unclear (perhaps OMR) Imitation
Call et al. (2005) End state Children (2 years) Unclear (perhaps affordancelearning)
Imitation
Chimpanzees (Pantroglodytes)
Yes (goal-emulation) butnon-social cues confoundedresults
Denny et al.(1988)
Ghost display Rats (Rattus norvegicus) Yes (OMR)
Groesbeck &Duerfeldt(1971)
Ghost display Rats Yes (OMR) Evidence suggests stimulusenhancement
Heyes et al.(1994)
Ghost display Rats No (experimental confounds) No (experimental confounds)
Hopper et al.(2007)
Ghost display Chimpanzees Yes (OMR) Stimulus enhancement andimitation
Hopper et al.(2008)
Ghost display Children (3 to 4 years) Yes (affordance learning, OMRand goal-emulation)
Chimpanzees Yes (affordance learning, OMRand goal-emulation)
Hopper et al. (inpress)
Ghost display Children (3 to 4 years) Yes (OMR) Evidence suggests stimulusenhancement
Huang et al.(2002)
End state Infants (19 months) Yes (affordance learning andgoal-emulation)
Stimulus enhancement
Huang &Charman(2005)
Ghost display Infants (17 months) Yes (OMR and affordancelearning)
Huang et al.(2006)
End state Infants (31 and 41months)
Yes (OMR) Possible evidence forimitation
Fawcett et al.(2002)
Ghost display Starlings (Sturnusvulgaris)
No Imitation identified, butstimulus enhancementcould explain results
Jacobson &Sisemore(1976)
Ghost display Undergraduatestudents
Yes (OMR)
Klein & Zentall(2003)
Ghost display Pigeons (Columbia livia) Unclear (results explained moreparsimoniously by stimulusenhancement affordancelearning and OMR)
Imitation identified
Meltzoff (1985) End state Infants (14 months) Unclear (possible evidence forOMR)
Unable to learn fromend-state condition somatching adultdemonstration potentiallydue to imitation and/orOMR
Miller et al.(2009)
Ghost display Dogs (Canis familiaris) Unclear (possible evidence forOMR)
Subiaul et al.(2004)
Ghost display Rhesus macaques(Macaca mulatta)
No Cognitive imitation proposedas the monkeys only learntfrom the conspecific
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

6 Lydia M. Hopper
Table 1. (cont.)
Study Condition Species testedEvidence for emulation?(Yes/No/Unclear)
Other forms of observationallearning identified
Subiaul et al.(2007)
Ghost display Children (3 and 4years)
Yes Cognitive imitation
Autistic individuals Yes Cognitive imitationTennie et al.
(2006)Ghost display Infants (12, 18, 24
months)No Asocial learning
Chimpanzees Bonobos(Pan paniscus)Gorillas (Gorillagorilla) Orangutans(Pongo pygmaeus)
No Asocial learning
Thompson &Russell (2004)
Ghost display Infants (14 to 26months)
Yes (single-mat task only) Possible imitation
(a) Ghost display conditions employed with humans
Although the majority of ghost studies run with humans havebeen with infants and children, the first was conducted withyoung adults (undergraduate students, Jacobson & Sisemore,1976). Students in a ‘model’ condition saw a fellow studentmove a lever, after receiving a cue from a sequence of lights,to gain a reward (a USD five cent coin). In a ghost displaycondition (‘model-absent condition-apparatus operative’) thelever moved automatically, with no human present. Studentsin a third, no-information, control group were just shown theapparatus in a static state (‘apparatus inoperative condition’).Unlike more recent ghost studies interested in spontaneousresponses, the students in this study were instructed to copywhat they had seen.
Not only did the students in the model condition makemore responses than those in the control, but they also mademore correct responses. Interestingly, Jacobson & Sisemore(1976) reported that there was no significant difference in thelevel of success or number of responses made by the studentsin the ghost display condition compared to those that sawa model. They too performed better than those students inthe control. This early study provides no breakdown of thespecific observational learning mechanisms involved. Thesuccess of the students in the ghost display condition suggestsemulation, specifically OMR. However, this apparatus didnot allow for the identification of goal-emulation as there wasno alternative way for the students to manipulate the lever togain the reward. Furthermore, unlike the classic two-actionstudy conducted with budgerigars by Dawson & Foss (1965),the demonstrator in the model condition only moved thelever using one method (their hand) and so bodily imitationcould not be identified from the success of the students inthis condition.
Nearly 30 years after Jacobson & Sisemore (1976),Thompson & Russell (2004) conducted the first study toinvestigate the ability of children to learn from ghost displays.Fourteen to 26 month old children saw a toy on eitheron one or two mats (the ‘single-mat’ and ‘double-mat’conditions, respectively). The mats were moved either byan adult demonstrator or by a hidden pulley system (the
ghost display). In the single-mat condition, when the matwas pushed backwards the toy would move forwards. Inthe double-mat condition, pulling the mat that did not havethe toy placed on it caused the second, with the toy onit, to move forward. Thus, the actions needed to retrievethe toys were counterintuitive. The adult demonstrations forthe single- and double-mat conditions resulted in a greaternumber of retrieval acts by the children than by those in a no-information baseline. In the single-mat condition, childrenreplicated the actions after both the adult demonstration andghost display. In fact, in this condition children were moresuccessful in the ghost condition, which Thompson & Russell(2004) concluded was evidence of emulative learning. Thesame was not true for the double-mat condition. Thompson& Russell (2004) suggested that the ghost display conditiondid not enhance children’s responses for the double-mat taskbecause it did not result in a single, unambiguous, movement(both mats moved simultaneously).
The failure of the children to replicate the movementsof apparatus in the double-mat, but not the single-mat,condition in the ghost display condition would indicate thatthey were not relying on OMR. If they were using OMR theywould have moved both mats in the double-mat condition.The results suggest that the emulative learning in the single-mat condition was goal-emulation. If the children were tryingto replicate the outcome in the ghost display of the double-mat condition, the ambiguity of task may have preventedthem from doing so.
In a more rigorous study, Huang & Charman (2005)employed a series of experimental controls to tease apartimitation, affordance learning and OMR with infants (agedaround 17 months old). Videos of five object-manipulationtasks, (i) a dumbbell, (ii) box and stick tool, (iii) prongand loop, (iv) cylinder and beads and (v) square and post,similar to those used by Meltzoff (1995), were presented tothe infants in a series of conditions. The responses of theinfants, after seeing a video in which an adult manipulatedthe objects successfully (full demonstration), were recorded.These responses were compared to those of infants who hadseen either (i) a ghost display (‘object movement video’) in
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

Identifying social learning mechanisms 7
which the objects were manipulated but the human operatorwas digitally removed or (ii) a ‘body movement video’ inwhich the objects were removed showing only the adult’sactions. Finally, a control (‘baseline’) video showed only theinitial state of the objects. Huang & Charman (2005) aimedto determine whether OMR or body mimicry (imitation)would explain the infants’s copying.
After seeing the full demonstration and object movement(ghost display) videos, the infants were more likely toproduce the target actions than were those infants in thebody movement condition or control. OMR was proposedto explain the children’s responses because the end statewas shown for both conditions. Huang & Charman (2005)theorised that showing the object movements may haveprovided the infants with more salient information as to howto reach the end state, and thus the affordances of the testobjects (see also Goldenberg & Hagman, 1998, who discussthe ability of children to infer a tool’s affordances from itsphysical properties).
Huang & Charman (2005) concluded that the success ofthe children in the full demonstration condition could be dueto combined effects of emulation, stimulus enhancement,mimicry and goal-directed intention reading and that theirdata ‘‘. . .do not confirm that object-based emulation wasthe only responsible mechanism in the full demonstrationcondition’’ (p. 297). Although Huang & Charman (2005)acknowledge that from the conditions they ran, no singleform of learning could be identified, they should becommended. They ran a series of experimental conditions(including a failed-attempt condition not discussed here) andconsidered a range of social learning mechanisms to explaintheir findings to elucidate a better understanding of thelearning observed.
The first study to use a ghost display condition withchildren and to compare their responses with those ofnonhuman primates was conducted by Tennie et al. (2006).Infants aged 12, 18 and 24 months old were presented witha box which had a hinged door that could either be pushedor pulled to retrieve a reward. Four great ape species werealso tested in the same experimental conditions and theirresponses are discussed below. The infants were shown howto open the box (push or pull) in one of two conditions:(i) a full demonstration by an adult, or (ii) a ghost displayin which the door was moved discretely with fishing-lineby the experimenter. The apparatus used by Tennie et al.(2006) replicated that used by Bugnyar & Huber (1997,described in Section II) and represents an improvement ofthat used by Thompson & Russell (2004) because the actionsdemonstrated were not ambiguous or counterintuitive.
An interesting age-difference in the responses of theinfants was identified. Tennie et al. (2006) found that24 months old infants matched the action demonstratedin both conditions, those aged 18 months old only matchedthe adult demonstration while the 12-month-olds failed tomatch in either condition. Tennie et al. (2006) concludedthat their results ‘‘with 24-month-old children question theidea that emulation tasks may be too complex for human
infants’’ (p. 1166). However, the infants were also able toopen the box successfully in a no-information baseline.Accordingly, Tennie et al. concluded that the informationfrom the demonstrations was somewhat redundant and couldnot infer which social learning mechanism was used.
All of the above studies describe ghost display conditionsthat have tested typically developing children, by contrastSubiaul et al. (2007) compared the responses of typicallydeveloping children and individuals with autism. Individualswith autism aged eight to 20 years old (mean ‘socialization’age was 3.94 years and mean ‘communication’ age was4.79 years), and two groups of children (aged three andfour years old) were presented with a ‘simultaneous chainingparadigm’ in one of three experimental conditions or a no-information baseline. The task consisted of a series of fourimages presented concurrently, at random locations, on atouch-screen monitor. The participants were required toselect the images in a specific order. The three experimentalconditions were: (i) a ‘social human demonstration’, (ii)a ‘nonsocial computer-only condition’ with each correctresponse paired with a sound (comparable to a ghost display),and (iii) a ‘social-plus-computer condition’ in which theparticipant observed an adult select the images in the correctorder along with sound cues from the computer. The aimwas to test for cognitive imitation as this design removedthe confound of physical ability which may impair a child’sability to imitate (see Barr, 2002).
The individuals with autism and children of both agegroups were significantly more successful in the threeexperimental conditions compared to those in the no-information baseline. Although the children and individualswith autism responded successfully in the computer-onlycondition, Subiaul et al. (2007) concluded that (goal-)emulation could not explain what they termed ‘cognitiveimitation’ because the images had to be selected in thecorrect order and there was not an alternative way toreach the same end state. However, the participants didrepeat the actions shown in the absence of a demonstrator,indicative of Byrne’s (2002b) definition of emulation. Subiaulet al. (2007) reasoned that the participants learnt from theghost display condition because, despite being an inanimateobject, it ‘‘behaved as if it were animate and displayed thecharacteristics of social agents including, agency and goal-directedness [and]. . . children, from infancy, are sensitive tocues that index animacy and goal-directedness’’ (p. 240).
More recently, and following from Tennie et al. (2006),Hopper et al. (2008) presented three to four year old childrenwith a bidirectional task (the ‘Slide-box’) from which areward could be gained by sliding a door to either the left orright. The children were presented with one of three formsof demonstration for the Slide-box which were: (i) a fellowchild moving the door, (ii) a ghost display in which the doorwas moved discretely by the experimenter using fishing-line,or (iii) an ‘enhanced ghost’ display condition in which thechildren saw a ghost display but a second child sat in frontof the Slide-box, collecting the rewards, acting as a passivemodel. This third experimental condition was run to rule out
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

8 Lydia M. Hopper
a mere presence effect as a possible cause for why individualsmay perform better in a full demonstration, compared toa ghost display condition (Klein & Zentall, 2003). A fourthgroup of children was presented with the Slide-box in ano-information control.
Hopper et al. (2008) found that children copied thedirection that the door was pushed with their first responseafter seeing an active model, ghost and enhanced ghostdisplays. When all their responses are considered, thechildren continued to match the direction shown by the activemodel and the enhanced ghost display (passive model), whencompared to chance, but not after seeing a ghost display.These findings suggest that children copied by OMR, as notonly did they learn that the door could be moved (affordancelearning), but they also replicated the specific direction ofdoor movement (OMR).
All the ghost display conditions reviewed thus farhave involved the use of simple bidirectional or objectmanipulation tasks, none have required the use of a tool.This is surprising given the number of social learning studieswith children which have involved problem-solving and/ortool-use tasks (e.g. Want & Harris, 2001; Horner & Whiten,2005; McGuigan et al., 2007; Flynn & Whiten, 2008a, but seeNielsen, 2006). It has been proposed that children are ableto ascertain the purpose of a tool from its physical properties(Goldenberg & Hagman, 1998) suggesting that childrencould learn new tool-use tasks through emulative (affordance)learning rather than relying on imitation. However it hascommonly been reported that children learn imitatively(Nagell, Olguin & Tomasello, 1993; Horner & Whiten,2005) but this could be due to the unfamiliar nature of thetasks children are presented with in experimental situationswhich are often designed specifically so that they are novelto preclude the child employing previously held knowledge.
Potentially, both imitation and emulation could allowfor the acquisition of tool-use by children and to answerthis, Hopper et al. (in press) showed three to four year oldchildren ghost displays of a tool-use task, the Pan-pipes(Whiten, Horner & de Waal, 2005; Hopper et al., 2007).Two methods for operating the Pan-pipes with a stick-tool(lift or poke) were shown to the children in one of three ways:(i) a demonstration by an adult, (ii) a ghost display showingthe pertinent moving parts of the task (moved discretely bythe experimenter using fishing-line) or (iii) a ghost-with-toolcondition, in which the tool was incorporated into the ghostdisplay. A fourth group of children, in a no-informationcontrol, were given no form of demonstration and only threeof these 16 control children were able to solve how to operatethe Pan-pipes by trial-and-error.
Despite the apparent complexity of the task, the childrenthat saw an adult demonstration were significantly moresuccessful than those in the control. Over three-quarters(77%) of the children that saw an adult demonstration (ofeither lift or poke) were able to operate the Pan-pipes andall did so using the same method as that shown to them.As might be predicted, the contrast between the responsesof the children in the adult demonstration and two ghost
conditions was greatest for the basic ghost condition, whichprovided the least information to the observing children. Inthe basic ghost condition, not only did the children showreduced success, but they were also less likely to use thesame method as that shown. This suggests that the success ofthose children in the adult demonstration condition was dueto more than emulation of the movements of the Pan-pipesalone (OMR).
The ghost-with-tool condition yielded an intermediatepicture. Hopper et al. (in press) noted that the children wereless successful than those that saw an adult demonstrationand were not significantly more successful than children inthe basic ghost conditions or control. However, with theirfirst attempts at the task they revealed a significant tendencyto match what they had seen. They tried to make the tooldo what it had done in the ghost display. Overall, just undera third of these children succeeded and all employed themethod displayed in the ghost display, just as children inthe demonstration conditions were highly faithful to themethod they had seen, suggesting OMR of the tool, perhapscombined with affordance learning.
From the use of ghost display conditions with humans,evidence for emulative learning has been provided forchildren (Thompson & Russell, 2004; Huang & Charman,2005; Tennie et al., 2006; Hopper et al., 2007, in press;Subiaul et al. 2007), adults (Jacobson & Sisemore, 1976), andindividuals with autism (Subiaul et al., 2007). These reportsof emulative learning provide further information about thevariety of social learning mechanisms that humans employ.This highlights the importance of providing participantswith more than one form of demonstration to determinewhich forms of social learning are at play. In the future,the challenge will be not only to identify the form of sociallearning employed, but also to determine which contextspredict their use.
(b) Ghost display conditions employed with nonhuman primates
Ghost display conditions have been conducted with avariety of nonhuman primate species, often as part ofcomparative studies with children. The first such studywas conducted with rhesus macaques (Macaca mulatta) usinga simultaneous chaining paradigm to test for cognitiveimitation (Subiaul et al., 2004). Like the methodologydescribed for Subiaul et al. (2007), rhesus macaques observedeither a conspecific or computer-generated responses(‘computer-feedback condition’) for selecting a series of fourimages presented on a computer screen in a specific order.
The rhesus macaques that observed a fellow monkeyperform the task took fewer attempts to complete the trialaccurately compared to those in a no-information baseline(Subiaul et al., 2004). However those in the computer-feedback (ghost display) condition showed no greater successthan those in the baseline and nor did a third group whowere merely tested in the presence of a passive conspecific(‘social-facilitation condition’) to rule out a mere presenceeffect. This study was comprehensive, including controls forasocial learning (computer-feedback condition) and social
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

Identifying social learning mechanisms 9
motivation (social-facilitation condition), and the authorswere careful to consider, and rule out, different social learningmechanisms including local and stimulus enhancement andaffordance learning. However, the study only included twosubjects (both male) and they both acted as the ‘expert’(model) and ‘student’ (observer) on separate trials in the sociallearning condition. Furthermore, there was no alternativeway to reach the goal because a successful response wasdeemed when the macaque selected four images in a specificorder rather than attaining a final end state. Due to thedesign of this experiment, therefore, goal-emulation couldnot be identified.
Tennie et al. (2006, described above), in addition to testingchildren, conducted a series of comparative experimentswith chimpanzees, bonobos (Pan paniscus), gorillas (Gorilla
gorilla) and orangutans. Through the use of the ‘push/pull’bidirectional task, Tennie et al. (2006) aimed to testspecifically for OMR. The apes observed either a livedemonstration by a conspecific, a ghost display or werepresented with the task in a no-information control. Tennieet al. (2006) concluded that they ‘‘. . . found no evidence thatapes copied the information that they had observed; neitherthe observation of results nor results with actions changed theapes’ behavior. This is consistent with past studies showing[that] children are more adept than apes at matching theactions of others’’ (p. 1165–1166). Due to these negativeresults by the great ape species Tennie et al. (2006) wereunable to draw conclusions as to the learning mechanisminvolved and whether it be OMR or not, and no comparisonscould be made with the children who potentially learnt via
asocial learning.Following an ‘open diffusion’ experiment which found
that chimpanzees were able to learn a tool-use task (thePan-pipes) from conspecifics in a group setting, Hopper et al.(2007) ran a series of ghost display conditions to determinewhat form of learning would explain this faithful spread ofthe introduced technique. In the open diffusion experimenta chimpanzee, trained how to operate the Pan-pipes usingeither the lift or poke methods, was allowed to perform thetask in the presence of their cage mates. Hopper et al. (2007)reported that the group seeded with a chimpanzee proficientin the lift method was more likely to use the lift than the pokemethod. To investigate the forms of observational learningat play, chimpanzees were shown either a ghost lift displayor a ghost-with-tool display as described above with childrenfor Hopper et al. (in press).
Unlike children presented with ghost displays of the Pan-pipes (Hopper et al., in press), Hopper et al. (2007) reportedthat providing chimpanzees with either form of ghost displaydid not affect their level of success; chimpanzees in theghost display conditions were no more successful than whentested in a no-information baseline (Hopper et al., in press,provides a detailed comparison of the responses of thechimpanzees and children to ghost displays of the Pan-pipes).One proposed explanation for the lack of response by thechimpanzees was because they were tested in isolation, whichmay have made them nervous and unresponsive. However,
further investigation found that even when presented withcomparable ghost displays when in groups with their familiarcage mates, chimpanzees were still unable to learn how tooperate the Pan-pipes (Hopper, 2008).
The studies which have employed ghost display conditionswith nonhuman primates have all reported negative resultsi.e. the primates were unable to learn from a ghost display,even for motorically simple tasks like a bidirectional task(Tennie et al., 2006). The major confound of Tennie et al.’s(2006) study, however, was the inherent bias of the taskso that the apes were more likely to pull the door thanpush it regardless of which demonstration they had seen.Therefore Hopper et al. (2008) presented chimpanzees withthe Slide-box (described above). Although also a bidirectionaltask, because the two actions (left and right) were moremotorically similar (than push and pull) it was predictedthat the responses of the chimpanzees were less likely to beaffected by a personal bias.
Like children, chimpanzees matched the direction that thedoor was moved in the full demonstration and ghost displays(Hopper et al., 2008). This provided the first evidence ofemulative learning by chimpanzees from a ghost display.With their first responses, the chimpanzees matched thedirection in which the door was moved in the enhancedghost and ghost display conditions suggesting OMR. Thispattern of responses was not maintained however. Acrossall their responses, the chimpanzees ‘explored’ more thanthe children, showing an increased likelihood of moving thedoor in both directions, which could possibly be evidence foraffordance learning. Rather than concluding that the successof chimpanzees in the full demonstration and ghost displayconditions was purely due to OMR, it seems more likelythat it was an interplay between all three forms of emulativelearning (OMR, affordance learning and goal-emulation).
The failure of the chimpanzees to learn how to operatethe Pan-pipes from seeing a ghost display (Hopper et al.,2007) could have been because the chimpanzees learnt tobe ‘passive observers’; that the food would be delivered tothem whether they acted upon the device or not. Howeverthe success of chimpanzees presented with the ghost displayof the Slide-box (Hopper et al., 2008) disputes this. It seemsa more parsimonious suggestion that the reduced successof the chimpanzees shown a ghost display of the Pan-pipes,whether a tool was involved or not, is due to the complexity ofthe task, especially in comparison to the simpler bidirectionalSlide-box. The interplay between task complexity and sociallearning mechanism may also explain the differing success ofchildren to learn these same two tasks from ghost displays.This may also be applied to the varying success of childrenpresented with other complex (Thompson & Russell, 2004)and simple (Tennie et al., 2006) tasks.
(c) Ghost display conditions employed with dogs
To date, only one study has used ghost displays to test theobservation learning mechanisms employed by dogs (Milleret al., 2009). Following Klein & Zentall (2003), Miller et al.(2009) presented dogs with a bidirectional task in one of
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

10 Lydia M. Hopper
four conditions: (i) dog imitation (a full demonstration bya dog); (ii) social facilitation/emulation (an enhanced ghostdisplay with a dog present); (iii) human imitation group (ademonstration by an unfamiliar human; and (iv) emulationgroup (a basic ghost display). In all conditions, a door slidto either the left or right, revealing a bowl of food. The firstresponses of the observing dogs showed significant levels ofmatching to the demonstration in only the dog imitationcondition, however when all of the dogs’ responses areconsidered, significant matching was also shown by thosedogs in the emulation (basic ghost) condition. Miller et al.(2009) concluded from this that the high levels of matchingby those dogs in the dog imitation condition was evidence ofimitation but that the dogs were also capable of emulating.
Unlike in other ghost displays discussed in this review, inall test conditions after the door was moved an experimenterbehind the screen door gave praise (verbal reinforcement)and moved the food bowl (food reinforcement) within reachof the ‘demonstrators’ and observers. Miller et al. (2009)acknowledge that this could lead to social conditioning (anassociation developed between the door moving and the foodreinforcement) however, they discredit it as an explanationfor the matching behaviour shown by the dogs. Althoughthis may not be the sole explanation for the matching shownby the dogs, it is likely to be a component in shaping theirresponses and Miller et al. (2009) provide no satisfactoryexplanation why such verbal reinforcement was included.Further, having an experimenter move the food rewardtowards the ‘demonstrator’ or observer could have provideda cue of reinforcement to the dogs; created a ‘clever Hans’effect. This study, however, provides an insight into thelearning mechanisms employed by dogs, a species currentlyreceiving greater attention from those interested in sociallearning (see, for example, Range, Viranyi & Huber, 2007;Pongracz et al., 2008).
(d) Ghost display conditions employed with rats
The earliest study that employed what could be considered aghost display condition was conducted with rats (Groesbeck& Duerfeldt, 1971). Rats were presented with a Y-maze inone of a number of conditions (six in total), three of whichare of interest here. At the entrance to each of the twobranches of the maze was a flap-door that the rat had topush to walk along the passageway. The flap-door at thebeginning to the arm with the reward (a water bottle) wasblack and white striped while the flap-door at the entrance ofthe incorrect arm (no reward) was solid black. Cards with thesame design as the flap-doors were placed at the ends of thecorresponding arms of the maze. In the full-demonstrationcondition the rats observed a conspecific select the correctarm of the maze, push the flap-door out of the way and walkalong the arm to gain the reward. For the ghost display,the experimenter knocked down the correct flap-door witha pointer and tapped the water bottle, while a third groupof rats were presented with the Y-maze in a no-informationcontrol.
Groesbeck & Duerfeldt (1971) reported that the ratsin the full-demonstration and ghost display conditionslearnt the maze quicker than those in the control groupand performed fewer errors, but that those in the full-demonstration condition were the most successful. Althoughthe authors did not define the form of learning responsible,they did conclude that the success of the rats in the ghostdisplay condition could be explained by a ‘‘simpler constructsuch as stimulus enhancement’’ (p. 42). A likely explanationcould be a combination of affordance learning and stimulusenhancement. The rats saw the movement of the flap-doorand their attention was drawn to the specific branch ofthe maze and the water bottle by the experimenter withthe pointer. Furthermore, although the rats were able tocomplete the maze successfully in the ghost display condition,there was no way they could attain the goal using a novelmethod, so goal-emulation could not explain the responsesof the rats for this particular task.
Although not the focus of their study, Denny et al. (1988)used a ghost display to determine whether rats could learnto discriminate between a positive stimulus (a lever, whichwhen pressed delivered a food pellet) and a negative stimulus(a lever, which when pressed delivered no pellet). Rats weretrained by seeing the bars moved by a ‘hidden experimenter’(a ghost display). This revealed that the rats were able tolearn to press the bar which gave them a food reward fromsuch a demonstration and Denny et al. (1988) concludedthat ‘‘the necessary condition for the appropriate barpressappears to be the actual bar movement, that is, the responsedemonstration. In other words, the bar during training is notjust spurious’’ (p. 221). These findings suggest that rats arecapable of learning from a ghost display and are capable ofemulative learning, specifically OMR.
More recently, Heyes et al. (1994) conducted a study usingrats to test their ability to manipulate a rod from having seena ghost display. Rats observed a rod move to either the left orright to release a food reward. In one condition rats observeda conspecific move the rod whilst in an alternative conditionthe rod was moved automatically with no demonstratorpresent (ghost display). Only those rats that had observed aconspecific move the rod also moved the rod, and did so inthe same direction as that demonstrated, from which Heyeset al. (1994) concluded that the rats had imitated. However,Mitchell et al. (1999) later determined that the observing ratsmight have been affected by odour cues left on the bar bythe demonstrating rats, casting doubt on the conclusion ofimitation drawn by Heyes et al. (1994).
(e) Ghost display conditions employed with birds
Ghost display conditions have been conducted with threebird species, all including enhanced ghost display conditionsto control for a mere presence effect (Akins et al., 2002;Fawcett et al., 2002; Klein & Zentall, 2003). Although theapparatus and test species used by Akins et al. (2002) andFawcett et al. (2002) are different, the overall methodologyfor both experiments is the same because the birds either sawa full demonstration by a conspecific or an enhanced ghost
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

Identifying social learning mechanisms 11
display condition (see Hopper et al., 2008; Miller et al., 2009,described above).
In the study conducted by Akins et al. (2002), Japanesequail (Coturnix japonica) observed a door move to either theleft or right (to reveal food pellets) by either a conspecific(‘imitation condition’) or an enhanced ghost display (termedthe ‘control’). Akins et al. (2002) reported that the firstresponse of quail in the imitation condition significantlymatched the direction in which the door had been movedduring the demonstration, but the same was not true forthose in the enhanced ghost display condition. Akins et al.(2002) concluded that Japanese quail are capable of imitativelearning. A limitation of this experimental design was that thecontrol condition was run to guard for both a mere presenceeffect and for OMR. Using just this one condition does notprovide sufficient control and running further conditions,including a basic ghost display, would be ideal. Due to thepaucity of experimental control, no conclusion of imitationshould be drawn because the matching responses of the birdscould be due to other, unidentified, learning mechanisms.
In contrast to Akins et al. (2002), Fawcett et al. (2002)tested starlings (Sturnus vulgaris) with a two-action task. Theadvantage of the two-action task, in combination with a ghostcondition, is that it offers potentially the most convincing wayto tease apart imitation from emulation as the two-actionmethod allows for imitation to be identified with greaterconfidence than in other experimental settings, especiallywhen used in combination with a control group (Kaiser et al.,1997, see also Voelkl & Huber, 2000). The starlings sawa plug either being pulled out of or pushed down into abox containing mealworms, to allow access to the food. Theaction was done by either a conspecific demonstrator or withan enhanced ghost display, in which the plug was moveddiscretely by the experimenter with a passive conspecificpresent. Despite a tendency of the observers to push, ratherthan pull, the plug regardless of condition, of those thatdid respond there was a significant correlation betweenthe action demonstrated and the action performed by theobserver. By contrast, those starlings in the enhanced ghostdisplay condition showed no significant matching.
Like Akins et al. (2002), Fawcett et al. (2002) concludedthat ‘‘. . .observation of the demonstrator’s behaviour wasa crucial component in acquisition of the response, andthat the observers were not simply paying attention to themovement of the plug. Our results provide strong evidenceof imitation in starlings’’ (p. 554). Such a claim for imitationmay be overstated. Like Akins et al. (2002), perhaps it wassome combination of the movement of the apparatus inconjunction with the bird acting upon it which made itmore salient to the observing bird in the demonstrationcondition, for which stimulus enhancement would be a moreparsimonious explanation. To allow the experimenter totease apart the interplay of OMR, a mere presence effectand imitation more fully, birds should also be exposed to abasic ghost display.
Although claims of imitative learning by birds made byAkins et al. (2002) and Fawcett et al. (2002) may be premature,
the responses of the birds in the enhanced ghost displaycondition are suggestive of goal-emulation. In the study runby Fawcett et al. (2002), although the birds did not matchthe specific method shown in the enhanced ghost display,67% of subjects responded and were able to retrieve thefood reward regardless of the method used. This wouldsuggest that the birds were capable of goal-emulation andpossibly affordance learning. The birds were able to learnthe properties of the plug (that it could be moved to reveal afood reward) and of the task (that there was food inside) butnot necessarily the specific way it should be acted upon. Thesame is true for the birds tested by Akins et al. (2002) All butone bird responded in the enhanced ghost display condition,regardless of the direction in which they moved the screendoor. This is comparable to the responses of chimpanzees inthe enhanced condition reported by Hopper et al. (2008).
Extending upon Akins et al. (2002), Klein & Zentall (2003)conducted a study with pigeons (Columbia livia) with fourexperimental conditions. The subjects saw a screen doormove to either the left or right to reveal food pellets,in one of the following ways: (i) a full demonstration bya conspecific; (ii) an enhanced ghost display; (iii) a ghostdisplay; or (iv) a vision-blocked condition. The first threeconditions listed were those replicated with children andchimpanzees by Hopper et al. (2008) and the rationale forall has been described above. The fourth condition was runlike a full demonstration but a piece of cardboard was placedbetween the observer and demonstrator, blocking visualaccess between the two cages to control for possible odourcues (a potential limitation identified by Mitchell et al., 1999).
Klein & Zentall (2003) claimed evidence for imitationbecause, like the Japanese quail (Akins et al., 2002), thepigeons made significantly more matching responses inthe full demonstration condition compared to those in theenhanced ghost display condition. Klein & Zentall (2003)concluded further that, despite being a non-significant trend,because the pigeons made more responses in the enhancedghost display compared to the ghost display condition,social facilitation could be responsible (they used this terminterchangeably with a mere presence effect despite thedifferences highlighted in Section II).
Unfortunately, no direct comparison was drawn betweenthe responses of the pigeons in the ghost display condition tothose in the full demonstration group; rather, the responseswere compared to chance. The pigeons made significantlymore matching responses in the ghost display condition thanwould have been expected by chance from which Klein &Zentall (2003) concluded evidence of affordance learning.However, it seems more appropriate that such responsesare evidence of OMR because the pigeons did not merelylearn the affordances of the door (that it could slide) but alsowhich direction to move it. The success of the pigeons inthe ghost display condition undermines the conclusion thatthey showed evidence of imitation. A more comprehensiveconclusion could be that those pigeons that saw a livemodel may have been learning though a combination of
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

12 Lydia M. Hopper
OMR enhanced by a mere presence effect and stimulusenhancement.
(3) End-state conditions
In the end-state condition, also termed the ‘emulationcondition’ (Huang, Heyes & Charman, 2002), an individualis shown the initial state of the test object, which is thenmanipulated out-of-sight; the individual is then shown theresultant form of the object (Meltzoff, 1985). If the observeris able to complete the task from such information thengoal-emulation is inferred. Unlike those studies that haveemployed ghost display conditions, the end-state techniquehas only been used with humans (infants and children) andnonhuman primates (Table 1); therefore the studies will bediscussed collectively and not subdivided by subject species.
The end-state technique was pioneered by Meltzoff (1985)who showed 14-month-old infants a dumbbell-shaped toy,the ends of which could be removed. The initial-state wasthe intact dumbbell and the end state was the central shaftseparated from the two end pieces. When shown a fulldemonstration of this action, infants were able to reproduceit, even after a 24 h delay. However when infants were shownonly the initial and resultant end state, a non-significantnumber of children could re-enact the action to create thesame end state when presented with the intact dumbbell.Meltzoff (1985) concluded that the failure of the infants inthe end-state condition suggests that they were not capableof emulating.
To determine whether the failure of the infants tested byMeltzoff (1985) was due to their age Bellagamba & Tomasello(1999) replicated and extended Meltzoff’s (1985) study with12and 18-month-old infants. The infants were shown an objectcomparable to that used by Meltzoff (1985) and saw either anadult demonstrate the actions or only the start and end states.Across both conditions, the 18-month-old infants averagedmore than twice as many target acts as the 12-month-olds.Furthermore, the infants performed more target acts afterhaving seen the full demonstration than those in the end-state condition. Bellagamba & Tomasello (1999) concludedthat ‘‘infants of this age also have a stronger tendencyto imitate human intentional actions (imitative learning)than to reproduce interesting changes of states in objectsin the absence of human behavior (emulation learning)’’(p. 281). However, the success of the children who saw a fulldemonstration could be due to OMR, rather than imitation,and replicating this experiment with the inclusion of a ghostdisplay condition would help address this.
Huang et al. (2002) employed the end-state test with agroup of 19-month-old infants, demonstrating actions onfive objects, which resembled those used by Meltzoff (1995),described for Huang & Charman (2005) above. As part ofhis original experiment, Meltzoff (1995) showed children anadult try, but fail, to complete an action with an object,a demonstration that was also replicated by a mechanicalarm in order to assess the children’s sense of intentionality.Huang et al. (2002) ran their experiment specifically totest whether a simpler form of social learning could
explain the success of children in Meltzoff’s (1995) ‘failedattempt’ condition, rather than, as Meltoff (1995) posited,the children’s understanding of the model’s intentions. Toanswer this, Huang et al. (2002) tested children in one of fourconditions, one of which was the end-state. The infants saweither a full-demonstration by an adult, an emulation (end-state) condition or an adult-manipulation condition in whichthe demonstrator interacted with the object in a non-directedmanner. This third condition allowed the experimenter tocontrol for the effect of stimulus enhancement created whenthe experimenter interacted with the test objects in thedemonstration condition. Unlike the younger infants testedby Bellagamba & Tomasello (1999), Huang et al. (2002)reported that 19-month-old infants made just as manytarget acts in the end-state condition as those in the full-demonstration condition. Moreover, infants in both theseconditions performed more target acts than did those in theadult manipulation control. Huang et al. (2002) concludedthat infants were able to learn via stimulus enhancementand emulation. Specifically, I suggest, affordance learningand goal-emulation. Huang et al. (2002) also replicated thisexperiment with 17-month-old infants but unfortunately,none of the responses by the infants were at a significant leveland so no age comparisons could be made.
Most recently, Huang et al. (2006) replicated their 2002study with older children, aged 31 and 41 months. Noeffect of age was reported between the two groups.Unlike the younger children tested previously, Huang et al.(2006) reported that the children performed more targetacts in the full-demonstration compared to the end-statecondition. Such findings are in agreement with Bellagamba& Tomasello (1999), suggesting that age is not the onlydeterminant in a child’s ability to understand and learn froman end-state display. Indeed, as Bauer & Kleinknecht (2002)stated, ‘‘. . .age alone does not determine whether childrenwill evidence emulation. Rather, children can emulate novelcausal sequences well before the age of 4, just so long as thetask demands are manageable. In short, it appears that thelearning strategy a young child is capable of using varies as afunction of task demands, not age’’ (p. 19).
Employing a comparative approach, Call, Carpenter &Tomasello (2005) gave two year old children and chim-panzees one of four demonstrations: (i) a full demonstration;(ii) the actions only; (iii) an end state; or (iv) a no-informationbaseline control. The apparatus used was a tube, eithertransparent or opaque, with a reward inside. The rewardcould be retrieved by either twisting the ends off or pullingthe tube apart, breaking it in the middle. Both methodsresulted in distinct end states. Regardless of the methoddemonstrated, there was no significant difference in thenumber of tubes opened by the chimpanzees across the threeexperimental conditions, or between those and the baseline.However, more clear, than opaque, tubes were opened sug-gesting possible nonsocial enhancement; the chimpanzeeswere encouraged by seeing the reward inside the tube.
Chimpanzees in the end-state condition were morelikely to match the method demonstrated than those in
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

Identifying social learning mechanisms 13
the action-only condition. In fact, the chimpanzees oftenattempted the alternative method when having seen onlythe action demonstrated, possibly because they inferredthat the model had tried, but failed, with one methodand so they used the other. It is interesting to comparethis with the responses of children presented with ‘failedattempt’ demonstrations (see for example Carpenter, Akhtar& Tomasello, 1998; Carpenter, Call & Tomasello, 2002;Heyes, 1998; Bellagamba & Tomasello, 1999; Myowa-Yamakoshi & Matsuzawa, 2000; Charman & Huang, 2002;Huang et al., 2002; Wood et al., 2002; Bellagamba et al., 2006;Biro, Csibra & Gergely, 2007).
Overall, for chimpanzees, the end state, rather than theaction itself, appears to be of more importance. Call et al.(2005) concluded that this provided further evidence thatchimpanzees are more likely to emulate (goal-emulation)than imitate. By contrast, the children performed better inthe social (full-demonstration and action-only conditions)than in the nonsocial (end-state and baseline) conditions.Additionally, there was no significant difference between thetwo social conditions with regards to the level of matchingthe method showed. Like the chimpanzees, the children weremore likely to open the transparent, rather than the opaquetubes. Call et al. (2005) concluded that, unlike chimpanzees,children are more likely to take advantage of the socialinformation and thus imitate rather than emulate. I suggestthat, as that more children were more likely to open theclear than opaque tubes affordance learning should also beconsidered.
IV. DISCUSSION
(1) Ghost display condition evaluation
Since the first study run in the 1970s (Groesbeck & Duerfeldt,1971), experiments that have incorporated ghost displayconditions (Section III) have differed in a number of ways,allowing for the identification of the strengths and weaknessesof this experimental technique. The aim of employing aghost display is to determine whether the observer is able toreach a predetermined goal from seeing only the pertinentparts of the task/apparatus move without an active modeloperating it.
Children appear better able to learn from seeing a ghostdisplay when the action shown has an obvious cause-and-effect relationship. This is highlighted by the failure ofchildren to learn how to solve the ambiguous double-matcondition used by Thompson & Russell (2004) and from thebasic ghost display of the tool-use Pan-pipes task (Hopperet al., in press) compared to the success of children learninga bidirectional task (Hopper et al., 2008). The role of taskcomplexity applies not just to children but also to animals,which were able to learn from ghost displays of simple (Klein& Zentall, 2003; Hopper et al., 2008; Miller et al., 2009) butnot complex tasks (Hopper et al. 2007). However, the taskshould not be so simplistic that subjects are able to solve it
via asocial learning (e.g. Tennie et al., 2006) and nor should ithave an inherent bias so that one of the methods for solutionis favoured over the other(s) (e.g. Call et al., 2005) as noconclusions as to the social learning mechanisms employedcan be drawn.
Two common forms of ghost display condition have beenused: the basic and the enhanced ghost. The basic ghostdisplay condition has been employed with a variety of speciesincluding humans, nonhuman primates, rats and birds whilefewer have included an enhanced ghost display condition(Akins et al., 2002; Fawcett et al., 2002; Klein & Zentall, 2003;Hopper et al., 2008). The enhanced ghost display conditionis a useful extension as it helps to control for a mere presenceeffect. However, when the enhanced ghost display conditionis used in isolation, as by Akins et al. (2002) and Fawcett et al.(2002), it loses much of its usefulness. Researchers shouldaim always to employ a basic ghost display if including anenhanced ghost display.
Three studies that used the ghost display condition pre-sented the demonstrations on television screens or computermonitors (Subiaul et al., 2004, 2007; Huang & Charman,2005). Huang & Charman (2005) used video footage ofdemonstrations in order to allow for the manipulationsof the images – to create a ghost and an action-only dis-play – which were presented to infants. This was an elegantway to create a ghost display because it removed the needfor using potentially obvious fishing-line. A second benefit ofvideo is that all participants observe exactly the same demon-stration (McGuigan et al., 2007; Flynn & Whiten, 2008b;Nielsen, Simcock & Jenkins, 2008; Strouse & Troseth, 2008,Price et al., 2009; see D’eath, 1998 for a review).
Despite the potential benefits of video demonstrations,their integrity compared to a live demonstration is limitedas the images are not three-dimensional, nor life-sized andlack many of the features of a live interaction includingolfactory cues and physical contact (Flynn & Whiten,2008b). Furthermore, there is evidence that children haveto learn how to interpret video images (see Troseth &DeLoache, 1998, for a review) and in certain circumstanceschildren show a ‘video deficit’ (Anderson & Pempek, 2005;Suddendorf, Simcock & Nielsen, 2007). Researchers workingwith nonhuman subjects should note that television screenshave been developed for human vision and are not necessarilysuitable for all species but this can be overcome using LCDflat-screens (D’eath, 1998). These potential disadvantageousshould be acknowledged and, if possible, attended to whenemploying video demonstrations.
The two studies conducted by Subiaul et al. (2004, 2007),although with different species, were both designed to test forcognitive imitation using a simultaneous chaining paradigmpresented on a computer monitor. This form of presentationwas selected because it removed potential physical limitationsof the participants, which may have hindered their abilityto replicate the actions. However, this form of presentationhas its own inherent limitations. The test subjects were eitherhumans, who are familiar with computer monitors, or apair of rhesus macaques, also familiar with interacting with
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

14 Lydia M. Hopper
touch-screen monitors. Caution should be observed whenusing such a method with subjects unfamiliar with computermonitors. As with televisions, the visual capacity of the testspecies should also be considered to ensure that they are ableto perceive, and understand, all presented stimuli.
Although the ghost, and enhanced ghost, displayconditions allow for the identification of forms of emulativelearning they are also inherently unnatural. It is rare thatobjects move by themselves. The use of video could be used toovercome this limitation. A more naturalistic way to presenta ghost display, rather than to remove the demonstrator,may be to conceal them. In the wild, a chimpanzee mayobserve a group member fishing for termites with the bodyof the fishing chimpanzee occluded (for example by a tree)so that all the observer is able to see is the movement of thetool. Manipulated videos may provide a new direction forthe employment of ghost display conditions; creating morenaturalistic displays.
(2) End-state condition evaluation
The end-state condition, unlike a ghost display which revealsthe required movements of a task, just shows the start andend states of an object pre- and post-manipulation. If theobserver is able to reach the same end state then goal-emulation and, following Goldenberg & Hagman (1998),affordance learning can be identified. All but one of the end-state studies published have tested infants and children, andthe majority of these studies have represented replications ofthe first end-state study run by Meltzoff (1985).
Meltzoff’s (1985) original experiment with 14-month-oldinfants revealed no significant evidence that infants coulddeconstruct the dumbbell after having only seen the startand end states. Many of the end-state studies subsequentlyconducted replicated and extended upon Meltzoff’s (1985)first study, using the same dumbbells, sometimes in conjunc-tion with other test objects (Bellagamba & Tomasello, 1999;Huang et al., 2002, 2006). This allows for comparisons to bedrawn between the abilities of the participants tested to agreater degree than can be done between the different ghostdisplay studies. To date, the end-state condition using thedumbbells, has tested 12-, 14-, 18-, 19-, 31- and 41-months-old infants. Only one of these studies found that infants (thoseaged 19 months) in the end-state condition performed withequal success to infants that saw a demonstration by an adult(Huang et al., 2002).
The majority of the published studies have concluded thatchildren are more likely to learn from observing an adult’sdemonstration (imitative learning) than from an end-statecondition (emulative learning). However the lack of responseby children in end-state conditions may highlight limitationsof the method itself. Furthermore, as the majority of theend-state studies have employed the dumbbells the failureof children may be due to the particular test objects used.The ghost display studies have employed a wide rangeof apparatus, reporting mixed levels of success by theindividuals tested; possibly dependent on task complexity,
as discussed (see also Nielsen, 2009 who discusses 12-month-olds’ understanding of intentions and ability to emulate asdependent on the clarity of the affordances of a task). It wouldtherefore be interesting to see test how children respond toend-state conditions with a wider variety of test apparatus(Call et al., 2005).
Only one study has implemented the end-state conditionwith animals (Call et al., 2005). Call et al. (2005) comparedthe responses of chimpanzees and children. Although theydid not use the dumbbell, the test object had similarphysical properties as it was a single object that had tobe deconstructed (broken in half). Call et al. (2005) reportedthat the responses of the chimpanzees provided evidence foremulative learning, however the chimpanzees made no moretarget actions in the end-state condition or full-demonstrationcondition compared to the no-information control and soasocial learning may have been at play.
(3) What forms of learning have been identifiedthrough the use of ghost display and end-stateconditions?
The usually stated rationale for employing ghost display orend-state conditions is to tease apart emulative from imitativelearning. Through the use of ghost display conditions,emulation have been identified in infants aged 14 to26 months (Thompson & Russell, 2004), children aged threeto four years (Hopper et al., 2008, in press), chimpanzees(Hopper et al. 2008), dogs (Miller et al., 2009), rats (Groesbeck& Duerfeldt, 1971; Denny et al., 1988) and pigeons (Klein &Zentall, 2003).
Ghost display conditions have been conducted with anumber of species and with a range of apparatus. Thisdiversity does not preclude comparisons between studiesand species. Indeed, two studies were run specifically tocompare the responses of apes and children tested with thesame apparatus (Tennie et al., 2006; Hopper et al., 2008).The methodology used by Subiaul et al. (2004, 2007) allowsfor comparisons to be drawn between typically developingchildren, individuals with autism and rhesus macaques.Further, direct comparisons can be made between two studiesthat measured the responses of different bird species testedwith the same apparatus (Akins et al. 2002; Klein & Zentall,2003). Lastly, Hopper et al. (2008) and Miller et al. (2009)replicated the methodology used by Klein & Zentall (2003)allowing for a comprehensive comparison with the responsesof pigeons with those of children, chimpanzees, dogs andpigeons.
In comparison to the breadth of successful responsesrecorded of humans and animals to ghost displays, only onestudy has reported children learning how to perform an actfrom having seen an end-state demonstration (Huang et al.,2002). Observing only the start and end states of a test objectprovides the observer with no information about the physicalmovements required to complete the task, successful childrenmust have relied on goal-emulation or affordance learning.
It is interesting to note that through the use of the ghost orend-state conditions emulative learning has been identified in
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

Identifying social learning mechanisms 15
children ranging from as young as 14 months old (Thompson& Russell, 2004) to children aged four years (Hopper et al.,2008). The social dynamics that underlie the motivation ofinfants and children to imitate has been discussed in greatdetail (Uzgiris, 1981; Nadel et al., 1999; Carpenter, 2006;Nielsen, 2008). In contrast to Tennie et al. (2006), Nielsen(2006) demonstrated that 12-month-old infants showed apropensity to emulate while 18- and 24-month-olds weremore likely to imitate with increased age and their likelihoodto do so was also influenced by the ostensive cues providedby the model (see also Topal et al., 2008). Referring tohis earlier study (Nielsen, 2006), Nielsen (2008) stated that‘‘young infants emulate out of motivation to learn aboutthe world, whereas toddlers show an increasing proclivityfor imitation based on a desire to interact with, and to belike, others’’ (p. 33). Although the findings of the ghost andend-state conditions reviewed herein support this partially,it is interesting to note that those older children testedby Tennie et al. (2006) and Hopper et al. (2008, in press)showed evidence for emulative learning suggesting thatperhaps not just age, but task complexity, influence a child’ssocial learning strategy (see also Bauer & Kleinknecht, 2002;Nielsen, 2009).
V. CONCLUSIONS
(1) To date, few studies have employed ghost display andend-state conditions in combination. Further work toidentify the underlying social learning mechanisms ofboth human and animals is welcomed. The currentcorpus of literature has thus far provided evidence forforms of emulative learning in humans and animalsbut further research is required to identify, with greateraccuracy, the specific forms of observational learning.Both ghost display and end-state conditions haveproved their worth but are even more effective ifused in combination, especially when both ghost andenhanced ghost conditions are employed.
(2) It is recommended that, in future studies, authorsshould be more precise when describing the sociallearning mechanisms they have identified (see Huang& Charman, 2005). This call for clarity is not a newone, but was one of the main conclusions drawnby Want & Harris (2002) and Whiten et al. (2004).Not only should researchers show greater detail whendrawing their conclusions, but should also recognisethe possibility that more than one social learningmechanism could be operating simultaneously.
(3) To enable researchers to be more specific in theidentification of different social learning mechanisms,studies should be designed to incorporate a varietyof experimental techniques in combination (e.g.Groesbeck & Duerfeldt, 1971; Klein & Zentall,2003) and more than one test apparatus (e.g. Huanget al., 2002).
(4) The final recommendation is that future studies shouldinvestigate the role of the participants’ motivation tocopy and how this differs between seeing a live modeland a ghost or end-state display. To expand upon this,studies could incorporate the role of ostensive cues,task complexity and the status of the demonstratoron a participants’ motivation and understanding ofthe task.
VI. ACKNOWLEDGEMENTS
This review was written during an ESRC-fundedpostdoctoral fellowship whilst at Durham University, UK.Thanks are due to my postdoctoral mentor Emma Flynn.Appreciation is also felt to Andrew Whiten, NicolaMcGuigan and Claudio Tennie for discussion and commentson earlier drafts of this manuscript.
VII. REFERENCES
Akins, C. K., Klein, E. D. & Zentall, T. R. (2002). Imitativelearning in Japanese quail (Coturnix japonica) using the bidirectionalcontrol procedure. Animal Learning and Behavior 30, 275–281.
Akins, C. K. & Zentall, T. R. (1996). Imitative learning in maleJapanese quail(Coturnix japonica) using the two-action method.Journal of Comparative Psychology 110, 316–320.
Anderson, D. R. & Pempek, T. A. (2005). Television and veryyoung children. American Behavioral Scientist, 48, 505–532.
Barr, R. (2002). Imitation as a learning mechanisms and researchtool: how does imitation interact with other cognitive functions?Developmental Science 5, 16–18.
Bauer, P. J. & Kleinknecht, E. E. (2002). To ‘ape’ or to emulate?Young children’s use of both strategies in a single study.Developmental Science 5, 18–20.
Bekkering, H. & Prinz, W. (2002). Goal representations inimitative actions. In Imitation in Animals and Artifacts (ed.K. Dautenhahn & C. L. Nehaniv), pp. 555–572. A BradfordBook: MIT Press, Cambridge, MA.
Bekkering, H., Wohlschlager, A. & Gattis, M. (2000).Imitation of gestures in children is goal-directed. The Quarterly
Journal of Experimental Psychology 53, 153–164.Bellagamba, F., Camaioni, L. & Calonnesi, C. (2006). Change
in children’s understanding of others’ intentional actions.Developmental Science 9, 182–188.
Bellagamba, F. & Tomasello, M. (1999). Re-enacting intendedacts: comparing 12- and 18-month-olds. Infant Behavior and
Development 22, 277–282.Biro, S., Csibra, G. & Gergely, G. (2007). The role of behavioral
cues in understanding goal-directed actions in infancy. Progress in
Brain Research 164, 303–322.Biro, D., Inoue-Nakamura, N., Tanooka, R., Yamakoshi, G.,
Sousa, C. & Matsuzawa, T. (2003). Cultural innovation andtransmission of tool use in wild chimpanzees: evidence from fieldexperiments. Animal Cognition 6, 213–223
Boyd, R. & Richerson, P. J. (1985). Culture and the Evolutionary
Process. The University of Chicago Press, London, UK.
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

16 Lydia M. Hopper
Boyd, R. & Richerson, P. J. (1996). Why culture is common,but cultural evolution is rare. Proceedings of the British Academy 88,77–93.
Bugnyar, T. & Huber, L. (1997). Push or pull: an experimentalstudy on imitation in marmosets. Animal Behaviour 54, 817–831.
Byrne, R. W. (1994). The evolution of intelligence. In Behavior and
Evolution (ed. P. J. B. Slater & T. R. Halliday), pp. 223–265/Cambridge University Press, Cambridge UK.
Byrne, R. W. (1998). A comment on Boesch, C and Tomasello, Mchimpanzee and human culture. Current Anthropology 39, 604–605.
Byrne, R. W. (2002a). Emulation in apes: verdict ‘not proven’.Developmental Science 5, 20–22.
Byrne, R. W. (2002b). Imitation of novel complex actions: whatdoes the evidence from animals mean? Advances in the Study of
Behavior 31, 77–105.Byrne, R. W. & Russon, A. E. (1998). Learning by imitation: a
hierarchical approach. Behavioural and Brain Sciences 21, 667–721.Caldwell, C. A. & Whiten, A. (2003). Scrounging facilitates
social learning in common marmosets, Callithrix jacchus. Animal
Behaviour 65, 1085–1092.Call, J. & Carpenter, M. (2002). Three sources of information
in social learning. In Imitation in Animals and Artifacts (ed.K. Dautenhahn & C. L. Nehaniv), pp. 211–228. A BradfordBook: Massachusetts Institute of Technology, Cambridge MA.
Call, J. & Carpenter, M. (2003). On imitation in apes andchildren. Infancia y Aprendizaje 26, 325–349.
Call, J., Carpenter, M. & Tomasello, M. (2005). Copyingresults and copying actions in the process of social learning:chimpanzees (Pan troglodytes) and human children (Homo sapiens).Animal Cognition 8, 151–163.
Carpenter, M. (2006). Instrumental, social, and shared goals andintentions in imitation. In Imitation and the Social Mind: Autism
and Typical Development (ED. S. J. Rogers & J. H. G. Williams),pp. 48–70. Guilford Press: New York NY.
Carpenter, M., Akhtar, N. & Tomasello, M. (1998).Fourteen- through 18-month-old infants differentially imitateintentional and accidental actions. Infant Behavior and Development
21, 315–330.Carpenter, M., Call, J. & Tomasello, M. (2002). Understand-
ing ‘‘prior intentions’’ enables two-year-olds to imitatively learna complex task. Child Development 73, 1431–1441.
Charman, T. & Huang, C-T. (2002). Delineating the role ofstimulus enhancement and emulation learning in the behaviouralre-enactment paradigm. Developmental Science 5, 25–27.
Custance, D., Whiten, A. & Fredman, T. (1999). Sociallearning of an artificial fruit task in capuchin monkeys (Cebus
appella). Journal of Comparative Psychology 113, 13–25.Dawson, B. V. & Foss, B. M. (1965). Observational learning in
budgerigars. Animal Behaviour 13, 47–474.D’Eath, R. B. (1998). Can video images imitate real stimuli in
animal behaviour experiments? Biological Reviews 73, 267–292.Denny, M. R., Clos, C. F. & Bell, R. C. (1988). Learning in
the rat of a choice response by observation of S-S contingencies.In Social Learning: Psychological and Biological Perspectives (ed. T. R.Zentall & B. G. Galef), pp. 207–223. Lawrence ErlbaumAssociates, Publishers, Hillsdale, NJ.
Eriksson, K., Enquist, M. & Ghirlanda, S. (2007). Criticalpoints in current theory of conformist social learning. Journal of
Evolutionary Psychology, 5, 67–87.Fawcett, T. W., Skinner, A. M. J. & Goldsmith A. R. (2002).
A test of imitative learning in starling using two-action methodwith an enhanced ghost control. Animal Behaviour 64, 547–556.
Flynn, E. & Whiten, A. (2008a). Cultural transmission of tool-usein young children: a diffusion chain study. Social Development 17,699–718.
Flynn, E. & Whiten, A. (2008b). Imitation of hierarchicalstructure versus component details of complex actions by 3- and5-year-olds. Journal of Experimental Child Psychology 101, 228–240.
Galef, B. G., Manzig, L. A. & Field, R. M. (1986). Imitationlearning in budgerigars: Dawson and Foss (1965) revisited.Behavioural Processes 13, 191–202.
Galef, B. G. & Whiskin, E. E. (2004). Effects of environmentalstability and demonstrator age on social learning of foodpreferences by young Norway rats. Animal Behaviour 68, 897–902.
Gattis, M. (2002). Imitation is mediated by many goals, not justone. Developmental Science 5, 27–29.
Gleissner, B., Meltzoff, A. N. & Bekkering, H. (2000).Children’s coding of human action: cognitive factors influencingimitation in 3-year-olds. Developmental Science 3, 405–414.
Goldenberg, G. & Hagman, S. (1998). Tool use and mechanicalproblem solving in apraxia. Neuropsychologica 36, 581–589.
Groesbeck, R. W. & Duerfeldt, P. H. (1971). Some relevantvariables in observational learning of the rat. Psychonomic Science
22, 41–43.Heyes, C. M. (1993). Imitation, culture and cognition. Animal
Behaviour 46, 999–1010.Heyes, C. M. (1998). Theory of mind in nonhuman primates.
Behavioral and Brain Sciences 21, 101–148.Heyes, C. M. & Dawson, G. R. (1990). A demonstration of
observational learning in rats using a bidirectional control. The
Quarterly Journal of Experimental Psychology B 42, 59–71.Heyes, C. M., Jaldow, E., Nokes, T. & Dawson, G. R. (1994).
Imitation in rats (Rattus norvegicus): the role of demonstrator action.Behavioural Processes 32, 173–182.
Hopper L. M. (2008). Dissection of observational learning
among chimpanzees (Pan troglodytes) and children (Homosapiens). Ph.D. Thesis, University of St Andrews, UK.[http://hdl.handle.net/10023/ 436].
Hopper, L. M., Lambeth, S. P., Schapiro, S. J. & Whiten, A.(2008). Observational learning in chimpanzees and childrenstudied through ‘ghost’ conditions. Proceedings of the Royal Societies
B 275, 835–840.Hopper, L. M., Spiteri, A., Lambeth, S. P., Schapiro, S. J.
& Whiten, A. (2007). Experimental studies of traditionsand underlying transmission processes in chimpanzees. Animal
Behaviour 73, 1021–1032.Hopper, L. M., Flynn, E. G., Wood, L. A. N. & Whiten, A.
(in press). Observational learning of tool use in children:investigating cultural spread through diffusion chains andlearning mechanisms through ghost displays. Journal of
Experimental Child Psychology.Hoppitt, W., Blackburn, L. & Laland, K. N. (2007). Response
facilitation in the domestic fowl. Animal Behaviour 73, 229–238.Horner, V. & Whiten, A. (2005). Causal knowledge and
imitation/emulation switching in chimpanzees (Pan troglodytes)and children (Homo sapiens). Animal Cognition 8, 164–181.
Horowitz, A. C. (2003). Do humans ape? Or do apes human?Imitation and intention in humans (Homo sapiens) and otheranimals. Journal of Comparative Psychology 117, 325–336.
Huang, C-T. & Charman, T. (2005). Gradations of emulationlearning in infants’ imitation of actions on objects. Journal of
Experimental Child Psychology 92, 276–302.Huang, C-T., Heyes, C. & Charman, T. (2002). Infants’
behavioral reenactment of ‘‘failed attempts’’: Exploring the roles
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

Identifying social learning mechanisms 17
of emulation learning, stimulus enhancement, and understandingof intentions. Developmental Science 38, 840–855.
Huang, C-T., Heyes, C. & Charman, T. (2006). Preschoolers’behavioural reenactment of ‘‘failed attempts’’: the roles ofintention-reading, emulation and mimicry. Cognitive Development
21, 36–45.Jacobson, J. M. & Sisemore, D. A. (1976). Observational learning
of a lever pressing task. Southern Journal of Educational Research 10,59–73.
Kaiser, D. H., Zentall, T. R. & Galef, B. G. (1997). Canimitation in pigeons be explained by local enhancement togetherwith trial-and-error learning? Psychological Science, 8, 459–460.
Kendal, R. L., Coolen, I., Van Bergen, Y. & Laland, K. N.(2005). Trade-offs in the adaptive use of social and asociallearning. Advances in the Study of Behavior 35, 333–379.
Klein, E. D. & Zentall, T. R. (2003). Imitation and affordancelearning by pigeons (Columba livia). Journal of Comparative Psychology
117, 414–419.Laland, K. N. (2004). Social learning strategies. Learning and
Behavior 32, 4–14.Laland, K. N. & Galef, B. G. 2009. The Question of Animal Culture.
Harvard University Press: Cambridge, MA.Leca, J-B., Gunst, N. & Huffman, M. A. (2007). Food
provisioning and stone handling tradition in Japanese macaques:a comparative study of ten troops. American Journal of Primatology
70, 803–813.Leca, J-B., Gunst, N. & Huffman, M. A. (2008). Japanese
macaque cultures: inter- and intra-troop behavioural variabilityof stone handling patterns across 10 troops. Behaviour 114,251–281.
Maestripieri, D., Ross, S. K. & Megna, N. L. (2002). Mother-infant interactions in western lowland gorillas (Gorilla gorilla gorilla):spatial relationships, communication and opportunities for sociallearning. Journal of Comparative Psychology 116, 219–227.
Matsuzawa, T., Tomonaga, M. & Tanaka, M. (2006). Cognitive
Development in Chimpanzees Springer: New YorkMcguigan, N., Whiten, A., Flynn, E. & Horner, V. (2007).
Imitation of causally opaque versus causally transparent tool useby 3- and 5-year-old children. Cognitive Development 22, 353–364.
Meltzoff, A. N. (1985). Immediate and deferred imitation infourteen- and twenty-four-month-old infants. Child Development
56, 62–72.Meltzoff, A. N. (1988). The human infant as Homo Imitans.
In Social Learning: Psychological and Biological Perspectives (ed.T. R. Zentall & B. G. Galef), pp. 319–341, LawrenceErlbaum Associates, Publishers, Hillsdale, NJ.
Meltfoff, A. N. (1995). Understanding the intentions of others:re-enactment of intended acts by 18-month-old children.Developmental Psychology 31, 838–850.
Meltzoff, A. N. & Moore, M. K. (1977). Imitation of facial andmanual gestures by human neonates. Science 198, 75–78.
Mesoudi, A. & Whiten, A. (2004). The hierarchical transforma-tion of event knowledge in human cultural transmission. Journal
of Cognition and Culture 4, 1–24.Mesoudi, A. & Whiten, A. (2008). The multiple roles of cultural
transmission experiments in understanding human culturalevolution. Philosophical Transactions of the Royal Society B 363,3489–3501.
Miller, H. C., Rayburn-Reeves, R. & Zentall, T. R. (2009).Imitation and emulation by dogs using a bidirectional controlprocedure. Behavioural Processes, 80, 109–114.
Mitchell, C. J., Heyes, C. M., Gardener, M. R. & Daw-son, G. R. (1999). Limitations of a bidirectional control pro-cedure for the investigation of imitation in rats: odour cues onthe manipulandum. The Quarterly Journal of Experimental Psychology:
B 52, 193–202.Myowa-Yamakoshi, M. & Matsuzawa, T. (2000). Imitation of
intentional manipulatory actions in chimpanzees (Pan troglodytes).Journal of Comparative Psychology 114, 381–391.
Nadel, J., Guerini, C., Peze, A. & Rivet, C. (1999). Theevolving nature of imitation as a format for communication.In Imitation in Infancy (ed. J. Nadel & G. Butterworth),pp. 209–234. Cambridge University Press: Cambridge, UK.
Nagell, K., Olguin, R. S. & Tomasello, M. (1993). Processesof social learning in the tool use of chimpanzees (Pan troglodytes)and human children (Homo sapiens). Journal of Comparative Psychology
107, 174–186.Nicol, C. J. (1995). The social transmission of information and
behaviour. Applied Animal Behaviour Science 44, 79–98.Nielsen, M. (2006). Copying actions and copying outcomes: Social
learning through the second year. Developmental Psychology 42,555–565.
Nielsen, M. (2008). The social motivation for social learning.Behavioral and Brain Sciences, 31, 33.
Nielsen, M. (2009). 12-month-olds produce others’ intended butunfulfilled acts. Infancy, 14, 377–389.
Nielsen, M., Simcock, G. & Jenkins, L. (2008). The effect ofsocial engagement on 24-month-olds’ imitation from live andtelevised models. Developmental Science 11, 722–731.
Perry, S. (2008). Manipulative Monkeys: The Capuchins of Lomas
Barbudal. Harvard University Press: Cambridge, MA.Pongracz, P., Vida, V., Banhegyi, P. & Miklosi, A. (2008).
How does dominance rank status affect individual and sociallearning performance in the dog (Canis familiaris)? Animal
Cognition 11, 75–82.Price, E. E., Lambeth, S.P., Schapiro, S. J. & Whiten, A.
(2009). A potent effect of observational learning in chimpanzeeconstruction. Proceedings of the Royal Society: B 276, 3377–3383.
Range, F., Viranyi, Z. & Huber, L. (2007). Selective imitationin domestic dogs. Current Biology, 17, 868–872.
Russon, A. E., Kuncoro, P., Ferisa, A. & Handayani, D. P.(2008). How Orangutans Produce Innovations. Proffered paperpresented at the XXII Congress of the InternationalPrimatological Society, Edinburgh, UK.
Strouse, G. A. & Troseth, G. L. (2008). ‘‘Don’t try this athome’’: Toddlers’ imitation of new skills from people on video.Journal of Experimental Child Psychology 101, 262–280.
Subiaul, F., Cantlon, J. F., Holloway, R. L. & Terrace,H. S. (2004). Cognitive imitation in rhesus macaques. Science 305,407–410.
Subiaul, F., Lurie, H., Romansky, K., Klein, T., Holmes, D.& Terrace, H. (2007). Cognitive imitation in typically-developing 3- and 4-year olds and individuals with autism.Cognitive Development 22, 230–243.
Suddendorf, T., Simcock, G. & Nielsen, M. (2007). Visualself-recognition in mirrors and live videos: evidence for adevelopmental asynchrony. Cognitive Development 22, 185–196
Templeton, J. J. & Giraldeau, L-A. (1996). Vicarious sampling:the use of personal and public information by starlings foragingin a simple patchy environment. Behavioral Ecology and Sociobiology
38, 105–114.Tennie, C., Call, J. & Tomasello, M. (2006). Push or pull:
imitation vs. emulation in great apes and human children. Ethology
112, 1159–1169.
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society

18 Lydia M. Hopper
Thompson, D. E. & Russell, J. (2004). The ghost condition:imitation versus emulation in young children’s observationallearning. Developmental Psychology 40, 882–889.
Thorpe, W. H. (1963). Learning and Instinct in Animals. Methuen,London, UK.
Tolman, C. W. (1964). Social facilitation of feeding behaviour inthe domestic chick. Animal Behaviour 12, 245–251.
Tomasello, M. (1990). Cultural transmission in the tool use andcommunicatory signaling of chimpanzees. In ‘‘Language’’ and
Intelligence in Monkeys and Apes: Comparative Developmental Perspectives
(ed. S. T. Parker & K. R. Gibson), pp. 274–311. CambridgeUniversity Press, Cambridge, UK.
Tomasello, M. (1999). The Cultural Origins of Human Cognition.Harvard University Press, Cambridge, MA.
Tomasello, M., Davis-Dasilva, M., Camak, L. & Bard, K.(1987). Observational learning of tool-use by young chimpanzees.Human Evolution 2, 175–183.
Topal, J., Gergely, G., Miklosi, A., Erdohegyi, A.& Csibra, G. (2008). Infants’ perseverative search errorsare induced by pragmatic misinterpretation. Science, 321,1831–1834.
Troseth, G. L. & Deloache, J. S. (1998). The medium canobscure the message: young children’s understanding of video.Child Development 69, 950–965
Uzgiris, I. (1981). Two functions of imitation during infancy.International Journal of Behavioral Development, 4, 1–12.
Uzgiris, I. C. (1991). The social context of infant imitation.In Social Influences and Socialization in Infancy (ed. M. Lewis &S. Feinman), pp. 215–251. Plenum Press, London.
Uzgiris, I. C. (1993). Interpersonal interaction as foundation forcultural learning. Behavioral and Brain Sciences 16, 535–536.
Van Schaik, C. P., Ancrenaz, M., Borgen, G., Galdikas, B.,Knott, C. D., Singleton, I., Suzuki, A., Utami, S. S. &Merrill, M. (2003). Orangutan cultures and the evolution ofmaterial culture. Science 299, 102–105.
Van Schaik, C. P. & Pradham, G. R. (2003). A model for tool-use in primates: implications for the coevolution of culture andcognition. Journal of Human Evolution 44, 645–664.
Voelkl, B. & Huber, L. (2000). True imitation in marmosets.Animal Behaviour, 60, 195–202.
Want, S. C. & Harris, P. L. (2001). Learning from other people’smistakes: causal understanding in learning to use a tool. Child
Development 7, 431–443.Want, S. C. & Harris, P. L. (2002). How do children ape?
Applying concepts from the study of non-human primates tothe developmental study of ‘imitation’ in children. Developmental
Science 5, 1–13.
Whiten, A., Goodall, J., Mcgrew, W. C., Nishida, T.,Reynolds, V., Sugiyama, Y., Tutin, C. E. G., Wrangham,R. W. & Boesch, C. (2001). Charting cultural variation inchimpanzees. Behaviour 138, 1481–1516.
Whiten, A., Goodall, J., Mcgrew, W. C., Nishida, T.,Reynolds, V., Sugiyama, Y., Tutin, C. E. G., Wrangham,R. W. & Boesch, C. (1999). Cultures in chimpanzees. Nature
399, 682–685.Whiten, A. & Ham, R. (1992). On the nature and evolution of
imitation in the animal kingdom: reappraisal of a century ofresearch. Advances in the Study of Behavior 21, 239–283.
Whiten, A., Mcguigan, N., Marhsall-Pescini, S. & Hopper,L. M. (2009). Emulation, Imitation, Over-imitation and theScope of Culture for Child and Chimpanzee. Philosophical
Transaction of the Royal Society B 364, 2417–2428.Whiten, A., Horner, V. & De Waal, F. B. M. (2005). Conformity
to cultural norms of tool use in chimpanzees. Nature, 437, 737–40.Whiten, A., Horner, V., Litchfield, C. A. & Marshall-
Pescini, S. (2004). How do apes ape? Learning and Behavior 32,36–52.
Whiten, A. & Van Schaik, C. P. (2007). The evolution of animal‘cultures’ and social intelligence. Philosophical Transactions of the
Royal Society of London, B 363, 603–620.Wohlschlager, A., Gattis, M. & Bekkering, H. (2003).
Action generation and action perception in imitation: an instanceof the ideomotor principle. Philosophical Transactions of the Royal
Society of London B 358, 501–515.Wood, D. (1989). Social interaction as tutoring. In Interaction
in Human Development (ed. M. H. Bornstein & J. S. Bruner),pp. 59–80. Lawrence Erlbaum Associates, Publishers, Hills-dale, NJ.
Wood, J. N., Glynn, D. D., Phillips, B. C. & Hauser, M. D.(2002). The perception of rational, goal-directed action innonhuman primates. Science 317, 1402–1405.
Zajonc, R. B. (1965). Social facilitation. Science 149, 269–274.Zentall, T. R. (2001). Imitation in animals: evidence, function,
and mechanisms. Cybernetics and Systems 32, 53–96.Zentall, T. R. (2003). Imitation by animals: how do they do it?
Current Directions in Psychological Science 12, 91–95.Zentall, T. R. (2006). Imitation: definitions, evidence, and
mechanisms. Animal Cognition 9, 335–353.Zentall, T. & Akins, C. (2001). Imitation in animals: evidence,
function, and mechanisms. In Avian Visual Cognition [On-line] (ed.R. G. Cook). Available: www.pigeon.psy.tufts.edu/avc/zentall
Zentall, T. R., Sutton, J. E. & Sherbourne, L. M. (1996).True imitative learning in pigeons. Psychological Scienc,e 7,343–236.
Biological Reviews (2010) 000–000 © 2010 The Author. Journal compilation © 2010 Cambridge Philosophical Society