golgi and tgn - scitech connectscitechconnect.elsevier.com/wp-content/uploads/2015/07/...golgi...
TRANSCRIPT

Golgi and TGNA Luini and S Parashuraman, Institute of Protein Biochemistry (IBP), Naples, Italy
r 2016 Elsevier Inc. All rights reserved.
The Golgi Apparatus – An Introduction
The Golgi apparatus and the closely associated trans-Golginetwork (TGN) are the central transport stations of the eu-karyotic secretory pathway, a fundamental positioning systemthrough which a large fraction of the eukaryotic proteins(roughly 30% in mammals) are biochemically processed andconveyed from the site of synthesis, the endoplasmic reticu-lum (ER), toward their cellular destinations. The secretorypathway is, together with the endocytic apparatus, a majorcellular organizer responsible for the location of every trans-ported protein to the right cellular membrane or to theextracellular space, where these proteins must operate, asrequired for cell functionality.
Secretory traffic begins at the ER where cargo proteinsare synthesized and inserted into the lumen or the membraneof this organelle. Cargo is then transferred, via the intermedi-ate compartment (Appenzeller-Herzog and Hauri, 2006;Saraste and Kuismanen, 1992), to the Golgi complex, mor-phologically a stack of flat membranous cisternae enrichedin glycosylating enzymes, and then through the cis-, medial-and trans-Golgi cisternae to the TGN, where it is sorted intomembranous carriers for delivery to successive cellular des-tinations (De Matteis and Luini, 2008; Gu et al., 2001). Alongthe way, and especially in the Golgi stack, cargo proteins(and lipids) are biochemically modified by glycosylation, themost abundant and complex eukaryotic posttranslationalmodification (Stanley, 2011); or they are broken down bydegradative systems located in the ER (Vembar and Brodsky,2008) and in the lysosomes (Luzio et al., 2007). Moreover,at each transport stage, anterograde transport is balanced byregulated retrograde compensatory fluxes of membrane andproteins to maintain the morphological and compositionalhomeostasis of the system (Cancino et al., 2014; Lorente-Rodriguez and Barlowe, 2011; Luini et al., 2014). To carry outthis enormous task, the transport apparatus relies on under-lying molecular machinery that is estimated to comprisemore than 2000 proteins, several hundreds of which reside inthe Golgi/TGN (Gannon et al., 2011; Gilchrist et al., 2006).
The following description refers mostly to the mammalianGolgi complex, which has been extensively studied. The Golgi,however, is evolutionarily very ancient, most probably as an-cient as the last common eukaryotic ancestor (LCEA), whichappeared roughly 1.5 billion years ago (Klute et al., 2011).Throughout evolution, the Golgi has varied significantly,especially in morphology, although its main functions,namely, glycosylation and transport and sorting, are likely tohave been substantially preserved (Klute et al., 2011).
The Structure of the Golgi Apparatus
The mammalian Golgi apparatus consists of hundreds ofGolgi stacks that are typically made up of 3–6 flat cisternae of
about 1 mm in diameter (Klumperman, 2011; Figure 1). Cis-ternae are swollen at the rims, where transport takes place, andare decorated with perforations in the periphery, which mayalign across successive cisternae to form ‘wells’ (Rambourg andClermont, 1990). Each Golgi stack is surrounded by numeroustransport vesicles roughly 50–70 nm in diameter. The vesiclesare especially concentrated near the rims and inside the wells(Ladinsky et al., 1994). The Golgi stacks are linked togetherside by side to form a ribbon that is localized near the cen-trosome of the cell, by virtue of microtubule-based motors(Yadav and Linstedt, 2011). The links between stacks areconstituted of tubules and perforated membrane areas (‘non-compact zones’) that typically join cisternae at the same level(i.e., cis-cisternae with cis and medial with medial) (Clermontet al., 1994; Klumperman, 2011). In addition, connections maysometimes join different levels in adjacent stacks (Clermontet al., 1994; Trucco et al., 2004; Vivero-Salmeron et al., 2008).These ‘heterologous’ connections, albeit infrequent, might befunctionally important for transport through the Golgi stack(Beznoussenko et al., 2014; Trucco et al., 2004).
The trans-most and sometimes the penultimate cisternaeextend tubules to form an interconnected tubular networkat the trans-Golgi face, called Trans-Golgi Network or TGN(Gu et al., 2001; Ladinsky et al., 1994). The size of the TGNvaries greatly with the secretory flux (Klumperman, 2011)and with transport conditions (Griffiths et al., 1989). Some-times, the trans-most cisterna can be seen to be ‘peeling off’from the rest of the stack, again possibly as part of the trans-port process (Ladinsky et al., 1994; Mogelsvang et al., 2003;Rambourg and Clermont, 1990). Notably, though the TGN isa well-developed structure closely associated with the Golgistack in mammalian cells, it appears to be an independentorganelle in plants. The plant TGN receives endocytosed ma-terial from the plasma membrane, and acts as an early endo-some. Indeed, TGN and early endosomes are generallyconsidered a single organelle in plants, and distinctions, if any,between them have not been explored (Richter et al., 2009).These and other observations (Lippincott-Schwartz et al.,1991) suggest that the TGN might have evolved from endo-somes early in evolution.
Transport through the Golgi
The Golgi is an obligate intracellular station for most of thesecreted cargoes that move from the ER to other intracellulardestinations. An exception is represented by a few cargomolecules that appear to bypass the entire secretory route andto follow an unconventional secretory pathway (Malhotra,2013; Rabouille et al., 2012). In the conventional route,secretory cargoes leave the ER via exit sites and reach theintermediate compartment before being transported to theGolgi via pleomorphic carriers (Appenzeller-Herzog andHauri, 2006; Barlowe and Miller, 2013). The arrival at
Encyclopedia of Cell Biology, Volume 2 doi:10.1016/B978-0-12-394447-4.20014-X 183

cisGolginetwork(CGN)
cis Cisterna
Medial cisterna
trans Cisterna
Secretory vesicle
Golgi vesicle
transGolginetwork(TGN)
trans Face
cis Face
Golgi complex transcis TGN
ER
COPII
ER exitsite
ERGIC
Nuclearenvelope
Nucleus
Recyclingendosome
Immaturesecretorygranule
Secretorygranule
Earlyendosome
LysosomePlasma
membrane
Clathrin
Late endosome/ multivesicular body
COPI
Nuclearenvelope
Rough ER
Vesiculartubularclusters
cis Golginetwork
1 µm
(a)
(b)
(c)
Figure 1 A representation of the Golgi stack depicting the transport pathways. (a) A cartoon depicting the 3D organization of the stack. (b) Anelectron micrograph showing the cross-section of a Golgi stack. (c) A schematic representation of the intracellular transport pathways and the centralrole played by the Golgi at the intersection of exocytic and endocytic pathways. Reprinted with permission from Alberts, B., 2002. Molecular Biologyof the Cell, fourth ed. New York, NY: Garland Science; Adapted from Bonifacino, J.S., Glick, B.S., 2004. The mechanisms of vesicle budding andfusion. Cell 116, 153–166.
184 Organelles: Structure and Function: Golgi and TGN
the Golgi of cargo-laden carriers stimulates a compensatoryretrograde transport from the Golgi to the ER that maintainsmembrane homeostasis at the Golgi (Cancino et al., 2014;Lorente-Rodriguez and Barlowe, 2011; Luini et al., 2014).
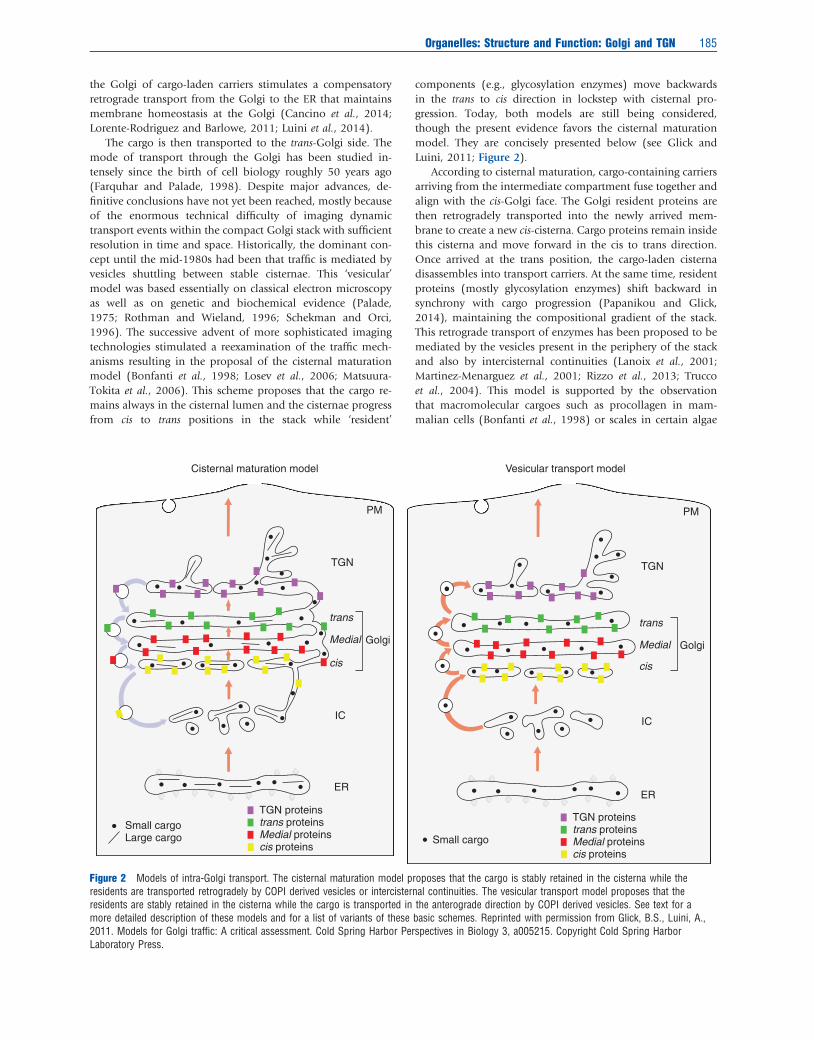
The cargo is then transported to the trans-Golgi side. Themode of transport through the Golgi has been studied in-tensely since the birth of cell biology roughly 50 years ago(Farquhar and Palade, 1998). Despite major advances, de-finitive conclusions have not yet been reached, mostly becauseof the enormous technical difficulty of imaging dynamictransport events within the compact Golgi stack with sufficientresolution in time and space. Historically, the dominant con-cept until the mid-1980s had been that traffic is mediated byvesicles shuttling between stable cisternae. This ‘vesicular’model was based essentially on classical electron microscopyas well as on genetic and biochemical evidence (Palade,1975; Rothman and Wieland, 1996; Schekman and Orci,1996). The successive advent of more sophisticated imagingtechnologies stimulated a reexamination of the traffic mech-anisms resulting in the proposal of the cisternal maturationmodel (Bonfanti et al., 1998; Losev et al., 2006; Matsuura-Tokita et al., 2006). This scheme proposes that the cargo re-mains always in the cisternal lumen and the cisternae progressfrom cis to trans positions in the stack while ‘resident’
components (e.g., glycosylation enzymes) move backwardsin the trans to cis direction in lockstep with cisternal pro-gression. Today, both models are still being considered,though the present evidence favors the cisternal maturationmodel. They are concisely presented below (see Glick andLuini, 2011; Figure 2).
According to cisternal maturation, cargo-containing carriersarriving from the intermediate compartment fuse together andalign with the cis-Golgi face. The Golgi resident proteins arethen retrogradely transported into the newly arrived mem-brane to create a new cis-cisterna. Cargo proteins remain insidethis cisterna and move forward in the cis to trans direction.Once arrived at the trans position, the cargo-laden cisternadisassembles into transport carriers. At the same time, residentproteins (mostly glycosylation enzymes) shift backward insynchrony with cargo progression (Papanikou and Glick,2014), maintaining the compositional gradient of the stack.This retrograde transport of enzymes has been proposed to bemediated by the vesicles present in the periphery of the stackand also by intercisternal continuities (Lanoix et al., 2001;Martinez-Menarguez et al., 2001; Rizzo et al., 2013; Truccoet al., 2004). This model is supported by the observationthat macromolecular cargoes such as procollagen in mam-malian cells (Bonfanti et al., 1998) or scales in certain algae
Cisternal maturation model Vesicular transport model
PM
TGN
trans
Medial
cis
Golgi
IC
ER
TGN proteinstrans proteinsMedial proteinscis proteins
Small cargoLarge cargo Small cargo
TGN proteinstrans proteinsMedial proteinscis proteins
PM
TGN
Golgi
trans
Medial
cis
IC
ER
Figure 2 Models of intra-Golgi transport. The cisternal maturation model proposes that the cargo is stably retained in the cisterna while theresidents are transported retrogradely by COPI derived vesicles or intercisternal continuities. The vesicular transport model proposes that theresidents are stably retained in the cisterna while the cargo is transported in the anterograde direction by COPI derived vesicles. See text for amore detailed description of these models and for a list of variants of these basic schemes. Reprinted with permission from Glick, B.S., Luini, A.,2011. Models for Golgi traffic: A critical assessment. Cold Spring Harbor Perspectives in Biology 3, a005215. Copyright Cold Spring HarborLaboratory Press.
Organelles: Structure and Function: Golgi and TGN 185

(Becker et al., 1995) move efficiently through the stack despitebeing too large to enter the vesicles and while remaining al-ways inside the lumen of cisternae (Bonfanti et al., 1998); andby direct visualization of Golgi components moving back-wards from distal to proximal cisternae, in synchrony withcisternal progression. This observation was made in Sacchar-omyces cereviseae, where the cisterna are unstacked, allowing forbetter microscopic resolution (Losev et al., 2006; Matsuura-Tokita et al., 2006).
The vesicular transport model instead proposes that cis-ternae are stable and the resident enzymes remain perman-ently in their lumen, while the cargo traverses the stack invesicular carriers. This scheme was proposed by Palade andcolleagues (Palade, 1975) and substantiated by the in vitrostudies of Rothman and colleagues, who identified the mo-lecular machinery of transport and proposed that the featuresof such machinery are consistent with vesicular transport(Rothman, 2002). Yeast genetic studies conducted in parallelby Schekman and colleagues reached the same conclusions(Schekman, 2002). The main evidence in support of thismodel is that cargo can be visualized by super-resolution mi-croscopy inside transport vesicles, at least under certaintransport conditions (Jamieson and Palade, 1967; Malhotraet al., 1989; Pellett et al., 2013). In addition, several variants ofthis stable cisternae model have been proposed and are inconsideration. These include the rapid mixing-partitioningmodel (Patterson et al., 2008), the cisternal progenitor model(Pfeffer, 2010), the kiss-and-run transport model (Beznous-senko and Mironov, 2002), and the rimmal maturation model(Lavieu et al., 2013). Finally, recent studies on the transport ofsoluble cargoes across the cisterna have shown that this type ofsecretory cargoes can diffuse across the stack via inter-cisternalcontinuities while other cargoes like procollagen move bycisternal maturation (Beznoussenko et al., 2014). Altogether,the available data suggest that multiple mechanisms of trans-port may coexist in the Golgi stack.
The cargoes arriving at the TGN are transported to multipledestinations including the basolateral domain, apical domain,endosomes, ER, and the Golgi stack itself (Anitei and Hoflack,2011; De Matteis and Luini, 2008). The first step in thetransport of cargoes to their appropriate destinations is theirsorting into cargo domains at the TGN from where theydepart. The segregation of these domains is due in most caseson sorting signals present on the cargo proteins that bind toappropriate adaptors leading to their concentration and seg-regation (Bonifacino, 2014), although other mechanismshave also been demonstrated (De Matteis and Luini, 2008).This step is followed by the formation of cargo-laden transportcarriers. Irrespective of their destination, most carriers thattransport cargo out of the TGN are large pleomorphic struc-tures (Polishchuk et al., 2000, 2006). They form by the ex-trusion of the cargo domains along cytoskeletal tracks bymotor proteins. The tubules connecting the carriers to the TGNare then severed to release the carriers that follow their courseto their appropriate destination using cytoskeletal tracks (Luiniet al., 2008).
In addition to playing a role in the sorting of cargoes out ofthe Golgi, TGN also acts as recipient for incoming traffic fromthe endosomes that allow for the recycling of receptors such asM6PR (Pfeffer, 2011).
Molecular Mechanism of Transport
Membrane traffic in eukaryotic cells uses a ‘toolbox’ of mo-lecular processes that are involved in the transport of cargofrom one compartment to another (De Matteis and Luini,2011; Figure 3). These ‘elementary’ processes include cargosorting and membrane bending and budding mediated by‘coat’ proteins, targeting of carrier vesicles to the target mem-brane by Rab and tethering proteins, and finally membranefusion mediated by soluble NSF attachment receptors(SNAREs), where NSF stands for N-ethylmaleimide sensitivefusion proteins. Each of these processes is mediated by a classof machinery proteins specialized for each of these ‘elem-entary’ events such as fission, fusion, etc. Here, we brieflymention the Golgi-specific variants of the proteins involved inthese processes, thus supporting transport across the stack. Themembers of these families of proteins that act at the Golgiinclude the COPI coat involved in cargo sorting and carrierformation (Popoff et al., 2011), adaptors for the coat for-mation like GOLPH3 in COPI vesicle formation (Banfield,2011) and AP-1 in clathrin-mediated vesicle formation(Bonifacino, 2014), phospholipase D2 involved in vesiclescission (Hsu and Yang, 2009), the COG family of membranetethers (Willett et al., 2013) and SNARE proteins membrin,syntaxin 5, GOS 28 etc. (Malsam and Sollner, 2011) involvedin vesicle fusion. In addition, the Golgi matrix proteins(Munro, 2011) maintain the architecture of the Golgi and arealso involved in vesicle tethering (Shorter and Warren, 2002).
Glycosylation at the Golgi
The proteins and lipids passing through the Golgi are pro-cessed by the Golgi-localized enzymes. The Golgi-localizedglycan-modifying enzymes are type II membrane proteins witha short cytoplasmic tail and the catalytic portion of the enzymelocalized in the lumen of the organelle (Hirschberg and Sni-der, 1987; Rini et al., 2009). Usually, the localization of theenzymes reflects their order of action in the glycosylationpathway, with the early acting enzymes present in the cis-sideof the Golgi and the later acting enzymes in the trans-side(Dunphy and Rothman, 1985).
The Golgi glycosylation enzymes are involved in theprocessing of N-glycans, O-glycan biosynthesis, proteoglycanbiosynthesis, and glycolipid biosynthesis. The protein N-glycosylation starts with the transfer of precursor poly-saccharides en bloc to a conserved Asn residue of the proteinby oligosaccharyltransferase in the ER. The mannose residues arethen trimmed in the ER before reaching the cis-Golgi where α-mannosidases further trim the glycan. This glycan then becomesthe substrate for medial-Golgi localized N-acetylglucosaminyltransferase that transfers a N-acetylglucosamine residue onto theN-glycan. The product of these reactions becomes the substratefor galactosyltransferases and sialyltransferases in the trans-Golgi/TGN, thus leading to the formation of complex N-glycan(for further details see Stanley et al., 2009). In contrast toN-glycosylation, O-glycosylation starts in the Golgi with theaddition of N-acetylgalactosamine on serine/threonine residueof the proteins by galactosamine transferases. The galactosa-mine residue is further modified in the trans-Golgi/TGN to
186 Organelles: Structure and Function: Golgi and TGN

create complex O-glycan forms (Brockhausen et al., 2009).Similar to O-glycosylation, proteoglycan synthesis startswith the addition of a monosaccharide, namely xylose, to the
serine/threonine residue. That is then extended to a tetra-saccharide containing one xylose, two galactoses, and a singleglucuronic acid residue. This tetrasaccharide is then extendeddepending on the type of proteoglycan to be synthesized.The glycan residues can also get other modifications such assulfation as in the case of proteoglycans (Esko et al., 2009) orphosphorylation as in the case of mannose-6-phosphatesynthesis.
Glycolipid biosynthesis starts in the ER with the productionof ceramide (Schnaar et al., 2009). A peculiar feature ofthe lipid biosynthesis is the use of lipid transfer proteins thattransfer lipids to the TGN where most of the glycolipid bio-synthesis takes place. The ceramide produced in the ER istransferred by CERT (ceramide transfer protein) to the TGNwhere it is converted into sphingomyelin by the action ofsphingomyelin synthases 1 and 2 (Hanada et al., 2009). Therest of the ceramide is transported to the cis-Golgi where it isfirst converted into glucosylceramide by glucosylceramidesynthase and then processed in the TGN to produce complexglycosphingolipids (D’Angelo et al., 2013).
The sugar residues needed for glycosylation reactions aretransported into the lumen as nucleotide sugar units bytransporters. While sialic acid is imported as CMP-sialic acidand fucose as GDP-fucose, most other sugars are importedas UDP-sugar conjugates (for further details, see Freeze andElbein, 2009). The vacuolar-ATPase localized to the trans-sidemaintains an acidic pH required for the glycosylation reactionsand the neutralization of pH impairs glycosylation (Rivinojaet al., 2012).
Regulation of the Golgi Apparatus Structure andFunction
Given the complex structural and functional features of theGolgi, it is not surprising that these features are controlledby specialized regulatory mechanisms. Screening studies haveshown that the secretory pathway is under the regulation ofmany kinases and phosphatases (Chia et al., 2012; Farhanet al., 2010; Simpson et al., 2012; Zacharogianni et al., 2011).Here we highlight a few such processes where the significanceof this kinase-mediated regulation is understood.
Some of these processes are involved in the maintenance ofthe homeostasis of the transport apparatus. Transport involveslarge membrane and protein fluxes across the transport sta-tions, that are subjected to physiological and pathologicalperturbations as well as to spontaneous drifts away fromequilibrium (Hirschberg et al., 1998; Mironov et al., 2001;Pulvirenti et al., 2008; Trucco et al., 2004). Such perturbationscan be a serious threat to the homeostasis, as they might alterthe composition and morphology of the transport compart-ments disrupting their function. An example of this possibilityis transport at the interface between ER and the Golgi complex.The ER is roughly 10-fold that of the Golgi in size (Griffithset al., 1984) and, its membrane output toward the Golgi canrepresent a significant fraction of the Golgi volume/surface(Griffiths et al., 1984; Klumperman, 2000; Martinez-Menar-guez et al., 1999; Thor et al., 2009). Thus, even minor changesin export from the ER would cause major alterations in Golgimorphology and composition, if not compensated for by
Cargo sorting and membrane bending
Membrane fission
GTPase dynamin
Vesicle
Vesicle
VesicleVesicle
Vesicle fusion
Acceptor compartment
Acceptor compartment
VesicleSNARE
Tether
TetherCompartment
SNARE
Motor protein
Microtubule
Vesicle translocation
Vesicle docking and tethering
Cargo receptor
Cargo receptor
Cargo A Cargo B
Coat
(a)
(b)
(c)
(d)
(e)
Figure 3 A schematic representation of the ‘toolbox’ of processesused in the intracellular transport. The vesicle formation involvescargo sorting followed by membrane bending initiated by coatproteins. (a) The mature vesicle is then scissioned by dedicatedfactors. (b) The vesicle then translocates via microtubule-basedmotors to the appropriate target compartment (c), where it is tetheredby a Rab and a molecular tether-dependent process (d) that alsoregulates the specificity of this vesicle tethering followed by dockingthrough specific SNARES. (d) SNARE pairing results in membranefusion (e) delivering the cargo to the target compartment. A specifictemporal combination of these elementary processes is used in all thetransport pathways and see text for the Golgi-specific moleculesinvolved in these processes at the Golgi apparatus. Reprinted withpermission from De Matteis, M.A., Luini, A., 2011. Mendeliandisorders of membrane trafficking. New England Journal of Medicine365, 927–938. Copyright 2014, Massachusetts Medical Society.
Organelles: Structure and Function: Golgi and TGN 187

corresponding changes in retrograde transport from the Golgito the ER (Klumperman, 2000; Martinez-Menarguez et al.,1999; Thor et al., 2009). However, the Golgi always maintainsor recovers its normal composition and morphology (Mironovet al., 2001; Trucco et al., 2004). At least some of the under-lying compensatory mechanisms use control devices that sensethe changes/perturbations and respond quickly, precisely, andappropriately to restore homeostasis.
One regulatory device that operates at the interface betweenER and Golgi has been investigated fairly extensively, andconcepts on the molecular composition and design of thesedevices have begun to emerge (Cancino et al., 2014). Mem-brane fluxes leaving the ER for the Golgi result in the activationof a traffic sensor, or receptor (the KDELR), at the cis-Golgi.This acts as a signaling protein that activates transductionpathways that are similar in composition to canonical PMsignaling pathways and kinase cascades. These kinases phos-phorylate a large number of Golgi and cytosolic proteins,some of which are involved in anterograde and retrogradetraffic. As a result, anterograde and retrograde traffic becomeactivated, counteracting the accumulation of membrane/proteins that would result from a non-compensated continu-ous arrival of traffic to the Golgi and hence helping to main-tain Golgi homeostasis. Whether this scheme applies at otherstages of the trafficking pathways remains to be investigated.
Simpler molecular processes involved in the transport arealso subjected to regulation. One particularly clear exampleis the regulation of uncoating of COPII-coated carriers. TheCOPII-coated carriers derived from the ER fuse with the Golgiapparatus to deliver the cargo and the regulation of uncoating ofthe carrier only at the appropriate compartment (Golgi) preventsthe possibility of engagement with an inappropriate compart-ment. This is accomplished by the specific localization of caseinkinase 1d at the Golgi that can phosphorylate the COPII coatcomponents upon arrival of the anterograde carrier at the Golgi,leading to its uncoating and subsequent fusion with the Golgicisternae (Lord et al., 2011). Finally, the glycosylation processesat the Golgi are also subjected to regulation. Examples includethe Src kinase-mediated regulation of the localization ofN-aceteylglucosamine transferases that initiate o-glycosylation(Chia et al., 2012; Gill et al., 2010) and the regulation of importof N-acetylgalactosamine into the Golgi lumen that determinesbranching of N-glycans (Lau et al., 2007).
Role of Golgi in Disease
Given the central role played by the Golgi apparatus in theprocessing and secretion of proteins, a defect in Golgi functionresults in serious disorders/diseases (for a detailed discussionsee Freeze and Ng, 2011). The disorders affecting specific gly-cosylation pathways lead to similar symptoms. For example,defects in O-mannose biosynthetic pathways lead to diseasesinvolving muscular dystrophy probably due to the high levelsof O-mannose present in α-dystroglycan while defects inproteoglycan biosynthesis mainly causes skeletal disorderssince extracellular matrix abundant in proteoglycans plays amajor role in bone development (Freeze and Ng, 2011). Golgi-associated diseases also occur due to mutations in the genesencoding proteins that form part of the structural/mechanistic
processes of the Golgi. Since a defect in structural or mech-anistic processes affect many aspects of Golgi function, thedisorders result in a plethora of symptoms. A classic exampleof such disorders is the congenital glycosylation disorders(CGD) caused due to mutations affecting the COG subunitsinvolved in the recycling of Golgi residents, a process requiredfor their appropriate localization (Zeevaert et al., 2008). CDGdisorders involving almost all the subunits of COG complexhave been documented and the patients with these disordersshow symptoms of impaired development, mental retard-ation, and hypotonia in many cases along with other subunit-specific symptoms (Freeze and Ng, 2011). In addition, mu-tations in a subunit of V-ATPase that is required for themaintenance of the pH inside the Golgi lumen essential forglycosylation result in connective tissue disorders with wrinklyskin (Cutis laxa) (Rosnoblet et al., 2013).
Apart from the genetic diseases mentioned above, Golgiproteins have also been linked with various other diseases in-cluding cancer, infection, etc. (Hu et al., 2007; Varki and Freeze,2009; Varki et al., 2009b). For instance, the Golgi-localized pro-tein GOLPH3 has been recently shown to be overexpressed insolid tumors and to act as an oncogene (Scott et al., 2009).
Evolution of the Golgi Apparatus
The secretory pathway is a central component of the eukaryoticendomembrane system and the Golgi apparatus plays an es-sential role in it. Nevertheless, and rather surprisingly, there areorganisms that do not seem to have a recognizable Golgi ap-paratus and that led to the suggestion these organisms mightrepresent the earliest branches of the eukaryotic tree before theevolution of a functional Golgi apparatus (Cavalier-Smith,1987). Later phylogenetic studies indicated that this was not thecase (reviewed in Mowbrey and Dacks, 2009) and suggested thatthe Golgi apparatus is a universal feature of the eukaryotic cellsand probably the LCEA did have a functional Golgi apparatus.
The Golgi apparatus varies in shape from a ribbon-likeGolgi in mammalian cells to scattered stacks in the plantsor insect cells to unstacked fragmented Golgi cisterna inS.cerevisiae. This led to the question about the nature of Golgiapparatus present in the LCEA. Phylogenetic studies haveshown that organisms that lacked Golgi stacks are embeddedin clades that had stacked Golgi apparatus suggesting that theabsence of a stack in certain organisms could be a secondaryloss and there were at least eight such independent instancesof Golgi unstacking during the eukaryotic evolutionary history(Mowbrey and Dacks, 2009). In addition, a recent phyloge-nomic study has shown that almost all the categories of Golgiproteins including SNAREs, Rabs, and matrix proteins areconserved across all the eukaryotes with secondary losses seenin some branches (Klute et al., 2011). Given the importance ofmatrix proteins in the stacking of the Golgi (Seemann et al.,2000; Shorter and Warren, 2002), this supports the idea thatearly eukaryotes probably had a stacked Golgi.
Apart from this conserved properties, evolutionary innov-ations have played a part in modifying and adapting the Golgito the need of the organisms. A clear example is the evolution ofribbon-like organization in the vertebrates (Wei and Seemann,2010). This structure is also present in sea urchin (Terasaki,
188 Organelles: Structure and Function: Golgi and TGN

2000), suggesting that probably it is a deuterostome innovation.Cell-biological studies have pointed to a role for the Golgiribbon in the efficiency of glycosylation (Puthenveedu et al.,2006; Xiang et al., 2013) and transition to deuterostomes isaccompanied by huge increase in the diversity of glycosylationproducts made by the Golgi apparatus with a pronounced in-crease in sialylated products (Varki et al., 2009a). Thus, theformation of the ribbon-like Golgi apparatus may have ac-companied or contributed to the evolution of deuterostomes.
Another innovation during the course of evolution was thedevelopment of the TGN. While the TGN is rudimentary in mostorganisms, it is a well-developed structure in the mammaliancells (probably also in other vertebrates). Moreover, in someorganisms the TGN and endosomes are identical (e.g., in plants)(Richter et al., 2009) while the mammalian TGN is distinctfrom the endosomes, with extensive membrane traffic betweenthem. The expansion of the TGN and development of the rib-bon-like structure was also accompanied by expansion in therepertoire of membrane traffic regulators with the human gen-ome encoding nearly 30 known Rab proteins (Diekmann et al.,2011). Whether the evolution of TGN also parallels an increasein the number of sorting pathways exiting the TGN and how thismay have contributed to the evolution of mammals remain openquestions.
Perspectives
After more than a century after its discovery, our understand-ing of the functioning of the Golgi apparatus remains uneven.While there is as yet no consensus on aspects as basic as themode of intra-Golgi transport or a definitive list of TGN exitpathways, the genomic and proteomic studies have led to arather extensive catalog of proteins localized to the Golgi ap-paratus and to a satisfactory understanding of the molecularprocesses they execute. The progress in addressing the un-resolved issues is intimately connected with advances in im-aging technology and with the use of novel reconstitutionapproaches (Wakana et al., 2012). Some of these aspects, in-cluding glycosylation, regulation of Golgi homeostasis,and cell-type specific variations of Golgi structure/function areyet to explored in detail. The resolution of these basic ques-tions of transport along with advances in the analysis offunctional aspects of the Golgi at a molecular level is necessaryto build a satisfactory systems understanding of the Golgiapparatus.
See also: Interorganellar Communication: Interplay andProcesses: Post-Golgi Transport − Cargo, Carriers, and Pathways.ER−Golgi Transport. Organelles: Structure and Function: EarlyEndosomal Compartments; Intermediate Compartment: A SortingStation between the Endoplasmic Reticulum and the GolgiApparatus; Signaling from Endosomes; The Late Endosome
References
Anitei, M., Hoflack, B., 2011. Exit from the trans-Golgi network: From molecules tomechanisms. Current Opinion in Cell Biology 23, 443–451.
Appenzeller-Herzog, C., Hauri, H.P., 2006. The ER-Golgi intermediate compartment(ERGIC): In search of its identity and function. Journal of Cell Science 119,2173–2183.
Banfield, D.K., 2011. Mechanisms of protein retention in the Golgi. Cold SpringHarbor Perspectives in Biology 3, a005264.
Barlowe, C.K., Miller, E.A., 2013. Secretory protein biogenesis and traffic in theearly secretory pathway. Genetics 193, 383–410.
Becker, B., Bolinger, B., Melkonian, M., 1995. Anterograde transport of algal scalesthrough the Golgi complex is not mediated by vesicles. Trends in Cell Biology 5,305–307.
Beznoussenko, G.V., Mironov, A.A., 2002. Models of intracellular transport andevolution of the Golgi complex. Anatomical Record 268, 226–238.
Beznoussenko, G.V., Parashuraman, S., Rizzo, R., et al., 2014. Transport of solubleproteins through the Golgi occurs by diffusion via continuities across cisternae.Elife 3, e02009.
Bonfanti, L., Mironov Jr., A.A., Martinez-Menarguez, J.A., et al., 1998. Procollagentraverses the Golgi stack without leaving the lumen of cisternae: evidence forcisternal maturation. Cell 95, 993–1003.
Bonifacino, J.S., 2014. Adaptor proteins involved in polarized sorting. Journal ofCell Biology 204, 7–17.
Brockhausen, I., Schachter, H., Stanley, P., 2009. O-GalNAc glycans. In: Varki, A.,Cummings, R.D., Esko, J.D. et al. (Eds.), Essentials of Glycobiology. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press, pp. 115–128.
Cancino, J., Capalbo, A., Di Campli, A., et al., 2014. Control systems of membranetransport at the interface between the endoplasmic reticulum and the Golgi.Developmental Cell 30, 280–294.
Cavalier-Smith, T., 1987. Eukaryotes with no mitochondria. Nature 326, 332–333.Chia, J., Goh, G., Racine, V., et al., 2012. RNAi screening reveals a large signaling
network controlling the Golgi apparatus in human cells. Molecular SystemsBiology 8, 629.
Clermont, Y., Rambourg, A., Hermo, L., 1994. Connections between the variouselements of the cis- and mid-compartments of the Golgi apparatus of early ratspermatids. Anatomical Record 240, 469–480.
D’Angelo, G., Capasso, S., Sticco, L., Russo, D., 2013. Glycosphingolipids:Synthesis and functions. The FEBS Journal 280, 6338–6353.
De Matteis, M.A., Luini, A., 2008. Exiting the Golgi complex. Nature ReviewsMolecular Cell Biology 9, 273–284.
De Matteis, M.A., Luini, A., 2011. Mendelian disorders of membrane trafficking.New England Journal of Medicine 365, 927–938.
Diekmann, Y., Seixas, E., Gouw, M., et al., 2011. Thousands of rab GTPases for thecell biologist. PLoS Computational Biology 7, e1002217.
Dunphy, W.G., Rothman, J.E., 1985. Compartmental organization of the Golgi stack.Cell 42, 13–21.
Esko, J.D., Kimata, K., Lindahl, U., 2009. Proteoglycans and sulfatedglycosaminoglycans. In: Varki, A., Cummings, R.D., Esko, J.D. et al. (Eds.),Essentials of Glycobiology. Cold Spring Harbor, NY: Cold Spring HarborLaboratory Press (Chapter 16). Available at: http://www.ncbi.nlm.nih.gov/books/NBK1900/#top (accessed 11.12.14).
Farhan, H., Wendeler, M.W., Mitrovic, S., et al., 2010. MAPK signaling to the earlysecretory pathway revealed by kinase/phosphatase functional screening. Journalof Cell Biology 189, 997–1011.
Farquhar, M.G., Palade, G.E., 1998. The Golgi apparatus: 100 years of progress andcontroversy. Trends in Cell Biology 8, 2–10.
Freeze, H.H., Elbein, A.D., 2009. Glycosylation precursors. In: Varki, A., Cummings,R.D., Esko, J.D. et al. (Eds.), Essentials of Glycobiology. Cold Spring Harbor,NY: Cold Spring Harbor Laboratory Press (Chapter 4). Available at: http://www.ncbi.nlm.nih.gov/books/NBK1929/ (accessed 11.12.14).
Freeze, H.H., Ng, B.G., 2011. Golgi glycosylation and human inherited diseases.Cold Spring Harbor Perspectives in Biology 3, a005371.
Gannon, J., Bergeron, J.J., Nilsson, T., 2011. Golgi and related vesicle proteomics:Simplify to identify. Cold Spring Harbor Perspectives in Biology 3, a005421.
Gilchrist, A., Au, C.E., Hiding, J., et al., 2006. Quantitative proteomics analysis ofthe secretory pathway. Cell 127, 1265–1281.
Gill, D.J., Chia, J., Senewiratne, J., Bard, F., 2010. Regulation of O-glycosylationthrough Golgi-to-ER relocation of initiation enzymes. Journal of Cell Biology 189,843–858.
Glick, B.S., Luini, A., 2011. Models for Golgi traffic: A critical assessment. ColdSpring Harbor Perspectives in Biology 3, a005215.
Griffiths, G., Fuller, S.D., Back, R., et al., 1989. The dynamic nature of the Golgicomplex. Journal of Cell Biology 108, 277–297.
Griffiths, G., Warren, G., Quinn, P., Mathieu-Costello, O., Hoppeler, H., 1984.Density of newly synthesized plasma membrane proteins in intracellularmembranes. I. Stereological studies. Journal of Cell Biology 98, 2133–2141.
Organelles: Structure and Function: Golgi and TGN 189

Gu, F., Crump, C.M., Thomas, G., 2001. Trans-Golgi network sorting. Cellular andMolecular Life Sciences: CMLS 58, 1067–1084.
Hanada, K., Kumagai, K., Tomishige, N., Yamaji, T., 2009. CERT-mediated traffickingof ceramide. Biochimica et Biophysica Acta 1791, 684–691.
Hirschberg, C.B., Snider, M.D., 1987. Topography of glycosylation in the rough endo-plasmic reticulum and Golgi apparatus. Annual Review of Biochemistry 56, 63–87.
Hirschberg, K., Miller, C.M., Ellenberg, J., et al., 1998. Kinetic analysis of secretoryprotein traffic and characterization of golgi to plasma membrane transportintermediates in living cells. Journal of Cell Biology 143, 1485–1503.
Hsu, V.W., Yang, J.S., 2009. Mechanisms of COPI vesicle formation. FEBS Letters583, 3758–3763.
Hu, Z., Zeng, L., Huang, Z., Zhang, J., Li, T., 2007. The study of Golgi apparatus inAlzheimer’s disease. Neurochemical Research 32, 1265–1277.
Jamieson, J.D., Palade, G.E., 1967. Intracellular transport of secretory proteins in thepancreatic exocrine cell. I. Role of the peripheral elements of the Golgi complex.Journal of Cell Biology 34, 577–596.
Klumperman, J., 2000. Transport between ER and Golgi. Current Opinion in CellBiology 12, 445–449.
Klumperman, J., 2011. Architecture of the mammalian Golgi. Cold Spring HarborPerspectives in Biology 3, a005181.
Klute, M.J., Melancon, P., Dacks, J.B., 2011. Evolution and diversity of the Golgi.Cold Spring Harbor Perspectives in Biology 3, a007849.
Ladinsky, M.S., Kremer, J.R., Furcinitti, P.S., McIntosh, J.R., Howell, K.E., 1994.HVEM tomography of the trans-Golgi network: Structural insights andidentification of a lace-like vesicle coat. Journal of Cell Biology 127, 29–38.
Lanoix, J., Ouwendijk, J., Stark, A., et al., 2001. Sorting of Golgi resident proteinsinto different subpopulations of COPI vesicles: A role for ArfGAP1. Journal ofCell Biology 155, 1199–1212.
Lau, K.S., Partridge, E.A., Grigorian, A., et al., 2007. Complex N-glycan number anddegree of branching cooperate to regulate cell proliferation and differentiation.Cell 129, 123–134.
Lavieu, G., Zheng, H., Rothman, J.E., 2013. Stapled Golgi cisternae remain in placeas cargo passes through the stack. Elife 2, e00558.
Lippincott-Schwartz, J., Yuan, L., Tipper, C., et al., 1991. Brefeldin A’s effects onendosomes, lysosomes, and the TGN suggest a general mechanism forregulating organelle structure and membrane traffic. Cell 67, 601–616.
Lord, C., Bhandari, D., Menon, S., et al., 2011. Sequential interactions with Sec23control the direction of vesicle traffic. Nature 473, 181–186.
Lorente-Rodriguez, A., Barlowe, C., 2011. Entry and exit mechanisms at thecis-face of the Golgi complex. Cold Spring Harbor Perspectives in Biology 3,a005207.
Losev, E., Reinke, C.A., Jellen, J., et al., 2006. Golgi maturation visualized in livingyeast. Nature 441, 1002–1006.
Luini, A., Mavelli, G., Jung, J., Cancino, J., 2014. Control systems and coordinationprotocols of the secretory pathway. F1000Prime Reports 6, 88.
Luini, A., Mironov, A.A., Polishchuk, E.V., Polishchuk, R.S., 2008. Morphogenesisof post-Golgi transport carriers. Histochemistry and Cell Biology 129, 153–161.
Luzio, J.P., Pryor, P.R., Bright, N.A., 2007. Lysosomes: Fusion and function. NatureReviews Molecular Cell Biology 8, 622–632.
Malhotra, V., 2013. Unconventional protein secretion: An evolving mechanism.EMBO Journal 32, 1660–1664.
Malhotra, V., Serafini, T., Orci, L., Shepherd, J.C., Rothman, J.E., 1989. Purificationof a novel class of coated vesicles mediating biosynthetic protein transportthrough the Golgi stack. Cell 58, 329–336.
Malsam, J., Sollner, T.H., 2011. Organization of SNAREs within the Golgi stack.Cold Spring Harbor Perspectives in Biology 3, a005249.
Martinez-Menarguez, J.A., Geuze, H.J., Slot, J.W., Klumperman, J., 1999. Vesiculartubular clusters between the ER and Golgi mediate concentration of solublesecretory proteins by exclusion from COPI-coated vesicles. Cell 98, 81–90.
Martinez-Menarguez, J.A., Prekeris, R., Oorschot, V.M., et al., 2001. Peri-Golgivesicles contain retrograde but not anterograde proteins consistent with thecisternal progression model of intra-Golgi transport. Journal of Cell Biology 155,1213–1224.
Matsuura-Tokita, K., Takeuchi, M., Ichihara, A., Mikuriya, K., Nakano, A., 2006. Liveimaging of yeast Golgi cisternal maturation. Nature 441, 1007–1010.
Mironov, A.A., Beznoussenko, G.V., Nicoziani, P., et al., 2001. Small cargo proteinsand large aggregates can traverse the Golgi by a common mechanism withoutleaving the lumen of cisternae. Journal of Cell Biology 155, 1225–1238.
Mogelsvang, S., Gomez-Ospina, N., Soderholm, J., Glick, B.S., Staehelin, L.A.,2003. Tomographic evidence for continuous turnover of Golgi cisternae in Pichiapastoris. Molecular Biology of the Cell 14, 2277–2291.
Mowbrey, K., Dacks, J.B., 2009. Evolution and diversity of the Golgi body. FEBSLetters 583, 3738–3745.
Munro, S., 2011. The golgin coiled-coil proteins of the Golgi apparatus. ColdSpring Harbor Perspectives in Biology 3, a005256.
Palade, G., 1975. Intracellular aspects of the process of protein synthesis. Science189, 347–358.
Papanikou, E., Glick, B.S., 2014. Golgi compartmentation and identity. CurrentOpinion in Cell Biology 29C, 74–81.
Patterson, G.H., Hirschberg, K., Polishchuk, R.S., et al., 2008. Transport through theGolgi apparatus by rapid partitioning within a two-phase membrane system. Cell133, 1055–1067.
Pellett, P.A., Dietrich, F., Bewersdorf, J., Rothman, J.E., Lavieu, G., 2013. Inter-Golgitransport mediated by COPI-containing vesicles carrying small cargoes. Elife 2,e01296.
Pfeffer, S.R., 2010. How the Golgi works: A cisternal progenitor model. Proceedingsof the National Academy of Sciences of the United States of America 107,19614–19618.
Pfeffer, S.R., 2011. Entry at the trans-face of the Golgi. Cold Spring HarborPerspectives in Biology 3, a005272.
Polishchuk, R.S., Polishchuk, E.V., Marra, P., et al., 2000. Correlative light-electronmicroscopy reveals the tubular-saccular ultrastructure of carriers operatingbetween Golgi apparatus and plasma membrane. Journal of Cell Biology 148,45–58.
Polishchuk, R.S., San Pietro, E., Di Pentima, A., Tete, S., Bonifacino, J.S., 2006.Ultrastructure of long-range transport carriers moving from the trans Golginetwork to peripheral endosomes. Traffic 7, 1092–1103.
Popoff, V., Adolf, F., Brugger, B., Wieland, F., 2011. COPI budding within the Golgistack. Cold Spring Harbor Perspectives in Biology 3, a005231.
Pulvirenti, T., Giannotta, M., Capestrano, M., et al., 2008. A traffic-activated Golgi-based signaling circuit coordinates the secretory pathway. Nature Cell Biology10, 912–922.
Puthenveedu, M.A., Bachert, C., Puri, S., Lanni, F., Linstedt, A.D., 2006. GM130and GRASP65-dependent lateral cisternal fusion allows uniform Golgi-enzymedistribution. Nature Cell Biology 8, 238–248.
Rabouille, C., Malhotra, V., Nickel, W., 2012. Diversity in unconventional proteinsecretion. Journal of Cell Science 125, 5251–5255.
Rambourg, A., Clermont, Y., 1990. Three-dimensional electron microscopy: Structureof the Golgi apparatus. European Journal of Cell Biology 51, 189–200.
Richter, S., Voss, U., Jurgens, G., 2009. Post-Golgi traffic in plants. Traffic 10,819–828.
Rini, J., Esko, J., Varki, A., 2009. Glycosyltransferases and Glycan-processingenzymes. In: Varki, A., Cummings, R.D., Esko, J.D. et al. (Eds.), Essentials ofGlycobiology. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press(Chapter 5). Available at: http://www.ncbi.nlm.nih.gov/books/NBK1921/ (accessed11.12.14).
Rivinoja, A., Pujol, F.M., Hassinen, A., Kellokumpu, S., 2012. Golgi pH, itsregulation and roles in human disease. Annals of Medicine 44, 542–554.
Rizzo, R., Parashuraman, S., Mirabelli, P., et al., 2013. The dynamics of engineeredresident proteins in the mammalian Golgi complex relies on cisternal maturation.Journal of Cell Biology 201, 1027–1036.
Rosnoblet, C., Peanne, R., Legrand, D., Foulquier, F., 2013. Glycosylation disordersof membrane trafficking. Glycoconjugate Journal 30, 23–31.
Rothman, J.E., 2002. Lasker Basic Medical Research Award. The machinery andprinciples of vesicle transport in the cell. Nature Medicine 8, 1059–1062.
Rothman, J.E., Wieland, F.T., 1996. Protein sorting by transport vesicles. Science272, 227–234.
Saraste, J., Kuismanen, E., 1992. Pathways of protein sorting and membrane trafficbetween the rough endoplasmic reticulum and the Golgi complex. Seminars inCell Biology 3, 343–355.
Schekman, R., 2002. Lasker Basic Medical Research Award. SEC mutants and thesecretory apparatus. Nature Medicine 8, 1055–1058.
Schekman, R., Orci, L., 1996. Coat proteins and vesicle budding. Science 271,1526–1533.
Schnaar, R.L., Suzuki, A., Stanley, P., 2009. Glycosphingolipids. In: Varki, A.,Cummings, R.D., Esko, J.D. et al. (Eds.), Essentials of Glycobiology. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press (Chapter 10). Available at:http://www.ncbi.nlm.nih.gov/books/NBK1909/ (accessed 11.12.14).
Scott, K.L., Kabbarah, O., Liang, M.C., et al., 2009. GOLPH3 modulatesmTOR signaling and rapamycin sensitivity in cancer. Nature 459,1085–1090.
Seemann, J., Jokitalo, E., Pypaert, M., Warren, G., 2000. Matrix proteins cangenerate the higher order architecture of the Golgi apparatus. Nature 407,1022–1026.
Shorter, J., Warren, G., 2002. Golgi architecture and inheritance. Annual Review ofCell and Developmental Biology 18, 379–420.
190 Organelles: Structure and Function: Golgi and TGN

Simpson, J.C., Joggerst, B., Laketa, V., et al., 2012. Genome-wide RNAi screeningidentifies human proteins with a regulatory function in the early secretorypathway. Nature Cell Biology 14, 764–774.
Stanley, P., 2011. Golgi glycosylation. Cold Spring Harbor Perspectives in Biology3, a005199.
Stanley, P., Schachter, H., Taniguchi, N., 2009. N-Glycans. In: Varki, A., Cummings,R.D., Esko, J.D. et al. (Eds.), Essentials of Glycobiology. Cold Spring Harbor,NY: Cold Spring Harbor Laboratory Press (Chapter 8). Available at: http://www.ncbi.nlm.nih.gov/books/NBK1917/ (accessed 11.12.14).
Terasaki, M., 2000. Dynamics of the endoplasmic reticulum and golgi apparatusduring early sea urchin development. Molecular Biology of the Cell 11, 897–914.
Thor, F., Gautschi, M., Geiger, R., Helenius, A., 2009. Bulk flow revisited: Transportof a soluble protein in the secretory pathway. Traffic 10, 1819–1830.
Trucco, A., Polishchuk, R.S., Martella, O., et al., 2004. Secretory traffic triggers theformation of tubular continuities across Golgi sub-compartments. Nature CellBiology 6, 1071–1081.
Varki, A., Freeze, H.H., 2009. Glycans in acquired human diseases. In: Varki, A.,Cummings, R.D., Esko, J.D. et al. (Eds.), Essentials of Glycobiology. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press (Chapter 43). Available at:http://www.ncbi.nlm.nih.gov/books/NBK1946/ (accessed 11.12.14).
Varki, A., Freeze, H.H., Gagneux, P., 2009a. Evolution of glycan diversity. In: Varki,A., Cummings, R.D., Esko, J.D. et al. (Eds.), Essentials of Glycobiology. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press (Chapter 19). Availableat: http://www.ncbi.nlm.nih.gov/books/NBK1942/ (accessed 11.12.14).
Varki, A., Kannagi, R., Toole, B.P., 2009b. Glycosylation changes in cancer. In:Varki, A., Cummings, R.D., Esko, J.D. et al. (Eds.), Essentials of
Glycobiology. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.(Chapter 44). Available at: http://www.ncbi.nlm.nih.gov/books/NBK1963/(accessed 11.12.14).
Vembar, S.S., Brodsky, J.L., 2008. One step at a time: Endoplasmic reticulum-associated degradation. Nature Reviews Molecular Cell Biology 9, 944–957.
Vivero-Salmeron, G., Ballesta, J., Martinez-Menarguez, J.A., 2008. Heterotypictubular connections at the endoplasmic reticulum-Golgi complex interface.Histochemistry and Cell Biology 130, 709–717.
Wakana, Y., van Galen, J., Meissner, F., et al., 2012. A new class of carriers thattransport selective cargo from the trans Golgi network to the cell surface. EMBOJournal 31, 3976–3990.
Wei, J.H., Seemann, J., 2010. Unraveling the Golgi ribbon. Traffic 11, 1391–1400.Willett, R., Ungar, D., Lupashin, V., 2013. The Golgi puppet master: COG complex
at center stage of membrane trafficking interactions. Histochemistry and CellBiology 140, 271–283.
Xiang, Y., Zhang, X., Nix, D.B., et al., 2013. Regulation of protein glycosylation andsorting by the Golgi matrix proteins GRASP55/65. Nature Communications 4, 1659.
Yadav, S., Linstedt, A.D., 2011. Golgi positioning. Cold Spring Harbor Perspectivesin Biology 3, a005322.
Zacharogianni, M., Kondylis, V., Tang, Y., et al., 2011. ERK7 is a negative regulatorof protein secretion in response to amino-acid starvation by modulating Sec16membrane association. EMBO Journal 30, 3684–3700.
Zeevaert, R., Foulquier, F., Jaeken, J., Matthijs, G., 2008. Deficiencies in subunits ofthe Conserved Oligomeric Golgi (COG) complex define a novel group of CongenitalDisorders of Glycosylation. Molecular Genetics and Metabolism 93, 15–21.
Organelles: Structure and Function: Golgi and TGN 191