herpetologists' league -...
TRANSCRIPT

Herpetologists' League
Phylogeographic Patterns in Kinosternon subrubrum and K. baurii Based on MitochondrialDNA Restriction AnalysesAuthor(s): DeEtte Walker, Paul E. Moler, Kurt A. Buhlmann, John C. AviseSource: Herpetologica, Vol. 54, No. 2 (Jun., 1998), pp. 174-184Published by: Herpetologists' LeagueStable URL: http://www.jstor.org/stable/3893425Accessed: 25/11/2008 13:21
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/action/showPublisher?publisherCode=herpetologists.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit organization founded in 1995 to build trusted digital archives for scholarship. We work with thescholarly community to preserve their work and the materials they rely upon, and to build a common research platform thatpromotes the discovery and use of these resources. For more information about JSTOR, please contact [email protected].
Herpetologists' League is collaborating with JSTOR to digitize, preserve and extend access to Herpetologica.
http://www.jstor.org

174 HERPETOLOGICA [Vol. 54, No. 2
trial Vertebrates in the Neotropical Realm. Dr. W Junk, The Hague, The Netherlands.
PARKER, H. W 1935. The lizards of Trinidad. Trop. Agric. 12:65-70.
PETERS, J. A., AND R. DONOSO-BARROS. 1986. Cat- alogue of the Neotropical Squamata. Part II, Liz- ards and Amphisbaenians (Revised ed.). Smithson- ian Institution Press, Washington, D.C., U.S.A.
PINTO, M. N. 1994. Cerrado: CaracterizaVao, Ocu- pa,Vo e Perspectivas (2nd ed.). Editora Universi- dade de Brasilia, Brasilia, Brasil.
PROJETO RADAMBRASIL. 1982a. Folha SD. 21 Cu- iaba; Geologia, Geomorfologia, Pedologia, Vegeta- Vao e Uso Potencial da Terra. Minist6rio das Minas e Energia, Rio de Janeiro, Brasil.
. 1982b. Folha SF. 21 Campo Grande; Geo- logia, Geomorfologia, Pedologia, VegetaVyo e Uso Potencial da Terra. Ministerio das Minas e Energia, Rio de Janeiro, Brasil.
SHERBROOKE, W C. 1975. Reproductive cycle of a tropical lizard, Neusticurus ecpleopus Cope. in Peru. Biotropica 7:194-207.
SILVA, J. M. C. 1995. Avian inventory of the cerrado region, South America: implications for biological conservation. Bird Cons. Int. 5:291-304.
VANZOLINI, P. E. 1961. Bachia: especies brasileiras e conceito generico (Sauria, Teiidae). Pap. Av. Dep. Zool., S. Paulo 14:193-209.
. 1966. Sobre o segundo exemplar de Bachia bresslaui (Sauria, Teiidae). Pap. Av. Zool., S. Paulo 19: 189-192.
VITT, L. J. 1982. Sexual dimorphism and reproduc- tion in the microteiid lizard, Gymnophthalmus multiscutatus. J. Herpetol. 16:325-329.
Accepted: 29 June 1997 Associate Editor: Daniel Formanowicz, Jr.
Herpetologica, 54(2), 1998, 174-184 ? 1998 by The Herpetologists' League, Inc.
PHYLOGEOGRAPHIC PATTERNS IN KINOSTERNON SUBRUBRUM AND K. BAURII BASED ON MITOCHONDRIAL
DNA RESTRICTION ANALYSES
DEETTE WALKER,' PAUL E. MOLER,2 KURT A. BUHLMANN,3 AND JOHN C. AVISE'
'Department of Genetics, University of Georgia, Athens, GA 30602, USA 2Wildlife Research Laboratory, Florida Game and Fresh Water Fish Commission, 4005 South Main Street,
Gainesville, FL 32601, USA 3Savannah River Ecology Laboratory, Drawer E, Aiken, SC 29801, USA
ABSTRACT: We used restriction assays of mitochondrial (mt) DNA to estimate phylogeographic variation in two sister taxa of muid turtles in the southeastern United States. Extensive mtDNA variation characterized Kinosternon subrubrum and, to a lesser degree, K. baurii. Each of 26 mtDNA haplotypes from the 83 assayed specimens was localized spatially. Collectively, these mtDNA haplotypes demarcated four major matrilineal assemblages, each with a well defined re- gional distribution: a western group (A) in Missouri and Louisiana, a central group (B) throughout the Gulf coastal states, an eastern group (C) along the Atlantic coastal states north of Florida, and a southern group (D) in peninsular Florida. All assayed samples of K. baurii belonged to the mtDNA C assemblage. The two species in Florida are thus highly distinct in mtDNA genotype, but they exhibit minimal mtDNA divergence along the Atlantic coastal states. These findings raise questions concerning the evolutionary history and taxonomy of these two recognized species. MtDNA phy- logeographic patterns in the baurii/subrubrum complex are remarkably similar to those reported previously for two other southeastern kinosternids, Sternotherus minor and S. odoratus.
Key words: Mud turtles; Phylogeography; Gene flow; Population struicture; Southeastern United States; Kinosternon.
MUD turtles (Kinosternon) are semi- aquatic organisms typically associated with slow-moving, often ephemeral waters such as shallow bayous, swamps, and ditches. These turtles commonly are observed tra- versing land (Ernst and Barbour, 1989;
Ernst et al., 1994), a habit that may influ- ence patterns of inter-drainage gene flow (Gibbons, 1983) and geographic popula- tion structure. Sixteen species of mud tur- tles are recognized in North, Central, and South America (Ernst et al., 1994), two of
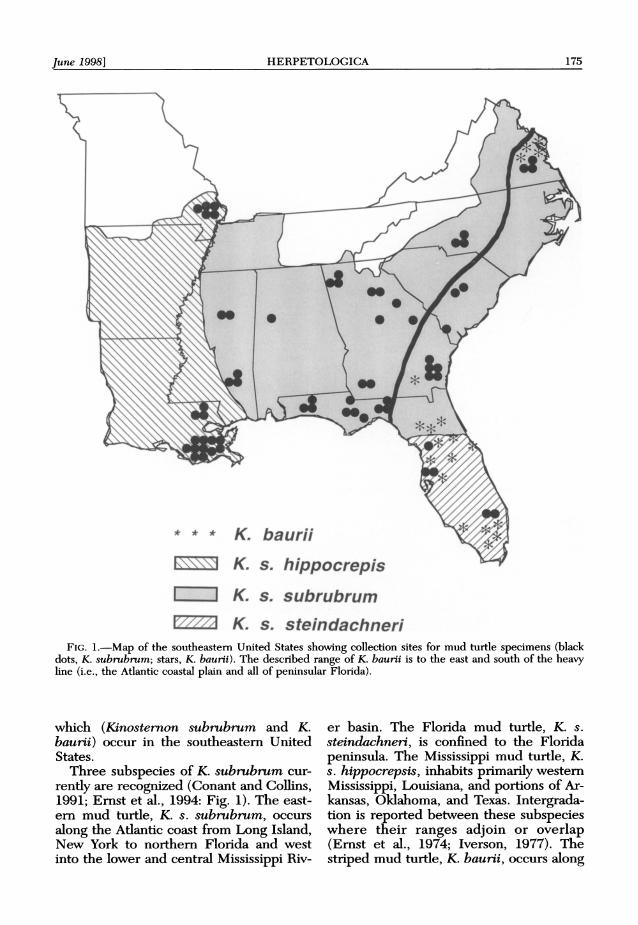
June 1998] HERPETOLOGICA 175
***K. b auriit
K. s. hippocrepis
K. s. subrubrum K. s. steindachneri
FIG. 1.-Map of the southeastern United States showing collection sites for mud turtle specimens (black dots, K. subrubrum; stars, K. baurii). The described range of K. baurii is to the east and south of the heavy line (i.e., the Atlantic coastal plain and all of peninsular Florida).
which (Kinosternon subrubrum and K. baurii) occur in the southeastern United States.
Three subspecies of K. subrubrum cur- rently are recognized (Conant and Collins, 1991; Ernst et al., 1994: Fig. 1). The east- ern mud turtle, K. s. subrubrum, occurs along the Atlantic coast from Long Island, New York to northern Florida and west into the lower and central Mississippi Riv-
er basin. The Florida mud turtle, K. s. steindachneri, is confined to the Florida peninsula. The Mississippi mud turtle, K. s. hippocrepsis, inhabits primarily western Mississippi, Louisiana, and portions of Ar- kansas, Oklahoma, and Texas. Intergrada- tion is reported between these subspecies where their ranges adjoin or overlap (Ernst et al., 1974; Iverson, 1977). The striped mud turtle, K. baurii, occurs along

176 HERPETOLOGICA [Vol. 54, No. 2
the Atlantic coast from southern Virginia to the Florida Keys (Ernst et al., 1994; Lamb and Lovich, 1990; Mitchell, 1994).
Most southern specimens of K. baurii display pronounced stripes on the cara- pace and head, but these stripes ebb in northern specimens, which causes identi- fication difficulties with K. s. subrubrum, a subspecies lacking such markings (Lamb, 1983a,b; Lamb and Lovich, 1990). Prior phylogenetic analyses based on allozymes (Seidel et al., 1986), karyology (Sites et al., 1979), and morphology (Iverson, 1991) suggested that K. baurii and K. subrubrum are closely related sister taxa within the Ki- nosternidae, but these studies were not designed to assess geographic variability within either species. Here we examine mitochondrial (mt) DNA variation within and between geographic populations of K. baurii and K. subrubrum.
MATERIALS AND METHODS
Samples and Laboratory Procedures
We collected 64 specimens of K. subru- brum from 32 locales and 19 specimens of K. baurii from 11 locales (Fig. 1, Appendix I). The specimens of K. baurii from Florida were easily distinguished morphologically from K. subrubrum because they displayed the characteristic stripes on the carapace and head. Specimens of K. baurii from At- lantic coast drainages had head stripes; their identification to species by Joseph Mitchell was based on these morphological criteria. In addition, morphological species assignments were confirmed by application of the discriminant function analyses de- fined in Lamb (1983b), as applied to five shell characteristics measured in all speci- mens of mud turtles collected along the At- lantic coast from the Carolinas through Florida. All specimens are deposited in The University of Georgia Museum of Natural History (UGAMNH 28567-28648) except those from Cohoke Mill Creek (Virginia) which were donated to the Smithsonian Museum (USNM numbers 515120,515121, 515124-515127, 515212, and 515213).
We extracted mtDNA from heart, liver, and muscle tissues following Lansman et al. (1981). Closed-circular mtDNA was di-
TABLE 1.-MtDNA haplotypes observed in Kinoster- non subrubrum and K baurii. Letters from left to right in the descriptions represent digestion profiles for the restriction enzymes BanI, Bcll, BglI, BglII, DraII, EcoRI, Hindll, HindIII, KpnI, NciI, NsiI,
PvuII, StuI, and XbaI.
Haplotype No. of code individuals Description
K subrl 3 CCCDCCCCCCCCCD K subr2 3 CBCDCCCCCCCCCD K subr3 6 CBCDCCCCCDCCCC K subr4 1 BBCDDCCCBDCCCC K subr5 1 CBCDCCCFCDCCCC K subr6 3 CBCDCCCCCCBCCC K subr7 2 CBCDBCCCCDCCCC K subr8 1 CBCDBCCCCDCCDC K subr9 9 BACCFCABBBEDCC K subrlO 2 AACCFCABBBEDCB K subrll 1 BACCFCBBBBEDCC K subrl2 3 BACCFCABBBEECC K subrl3 1 DACDECGACAFCBC K subrl4 1 DACDECGADCFCCC K subrl5 1 DACDECDACCFCCC K subrl6 2 DACEECDACBFCCC K subrl7 2 BACBFCABBFEDCC K subrl8 5 BACCFCABBFEDCC K subrl9 16 CEDCEBEECCHCFC K subr20 1 BACGFCABBBEDCB
K baur2l 4 CBCDCCCCCCCACC K baur22 1 CBCDCCCCCCCCCC K baur23 10 CBCDCCFCCCCBCC K baur24 1 CBCDCCFECCCBCC K baur25 2 CBCDCCICCCCBCC K baur26 1 CACDCCFCCCCBCC
gested by 14 restriction enzymes (Table 1) following recommendations of the manu- facturer (Boehringer Mannheim). Frag- ments were radioactively end-labeled us- ing Klenow and 32P-labeled nucleotides, size-separated by electrophoresis through 1.2-1.5% agarose gels, and visualized by autoradiography (Lansman et al., 1981). The digestion profiles proved informative in the sense that they yielded restriction fragment length polymorphisms (RFLP's) whose differences within and between K. baurii and K. subrubrum provisionally could be interpreted as restriction site gains or losses.
Data Analyses Each mtDNA digestion profile was as-
signed a letter code (Table 1). These let- ters were compiled for each individual into a composite mtDNA haplotype. From the

June 1998] HERPETOLOGICA 177
presence/absence matrix of restriction sites summarizing these haplotypes, sequence divergences (Nei and Li, 1979) and ge- notypic and nucleotide diversities (Nei, 1987) were calculated.
Phenetic relationships among haplo- types were inferred from the genetic dis- tance matrix using the neighbor-joining (N-J) method (Saitou and Nei, 1987) as implemented in PHYLIP (Felsenstein, 1991) and rooted by the mid-point crite- rion. We conducted parsimony analyses from the presence/absence matrix of re- striction sites using the heuristic search option in PAUP (Swofford, 1990). Statis- tical support was based on 1000 bootstrap pseudoreplicates. A parsimony network was hand-generated using observed num- bers of restriction site differences between the mtDNA haplotypes. Outgroup taxa were not employed to root the parsimony networks, because other assayed species of Kinosternidae (Walker et al., 1995, 1997) proved too divergent in most of the mtDNA digestion profiles to permit secure scoring of restriction site changes.
RESULTS
The mtDNA restriction site differences between K. baurii and some specimens of K. subrubrum were minimal, so the data were analyzed collectively. In total, the 14 informative restriction enzymes revealed 26 different mtDNA haplotypes: 20 for K. subrubrum and six for K. baurii (Table 1). The mtDNA molecule was approximately 16.3 kilobases in length in both species, with no evident size differences among in- dividuals. A mean of 43 restriction sites per individual was scored, reflecting 451 base pairs of recognition sequence or about 2.8% of the mtDNA genome.
The gel digestion profiles were inter- preted with respect to restriction site changes by methods described in Avise (1994). Fifty-nine of the 81 scored restric- tion sites were variable (Table 2), and 31 were informative phylogenetically (i.e., not confined to a single individual). For the pooled collection of samples, estimated genotypic diversity was 0.927 and nucleo- tide diversity was 0.041. Most of this di- versity stemmed from large differences be-
tween mtDNA genotypes in separate geo- graphic regions.
Parsimony networks (Fig. 2; see also legend to Fig. 3) and a neighbor-joining tree (Fig. 3) for the mtDNA haplotypes are based strictly on genotypic considera- tions and essentially agree in all major fea- tures. Four fundamental phylogenetic groups (A-D), each showing a strong geo- graphic orientation (Fig. 4), are evident. Group A occurs in the western-most por- tion of the range of K. subrubrum; group B occurs in the central portion of the range of K. subrubrum in the Gulf coastal states; group C includes K. subrubrum from Atlantic coastal states north of Flor- ida, plus all individuals of K. baurii (in- cluding those from the Florida peninsula); and group D consists of all specimens of K. subrubrum from the Florida peninsula. The mean levels of genetic divergence es- timated among haplotypes within each as- semblage (0.000, 0.003, 0.006, and 0.008 for groups A, B, C, and D, respectively) typically are much smaller than those be- tween assemblages (0.071, 0.068, 0.057, 0.054, 0.032, and 0.038, respectively, for the paired combinations A-B, A-C, A-D, B-C, B-D, and C-D).
Intra-assemblage genetic variation dif- fered considerably among the four genetic groups. The largest numbers of mtDNA haplotypes (seven and eight, respectively) were observed in genetic groups B and C. However, the highest values for genotypic diversity (0.918) and nucleotide diversity (0.013) occurred in the Florida peninsula with turtles representing both the C and D groups co-occurring there.
In peninsular Florida, all specimens as- signed by morphology to K. subrubrum belonged to mtDNA group D, and all specimens morphologically referable to K. baurii belonged to group C. However, group C (with bootstrap support 99%: Fig. 3) also included all sampled turtles from the Atlantic coastal states, regardless of taxonomic status (K. baurii or K. subru- brum). Within the C group, specimens of K. baurii from Florida differed consistent- ly from those in Georgia and Virginia by at least two restriction site changes (Fig. 2).
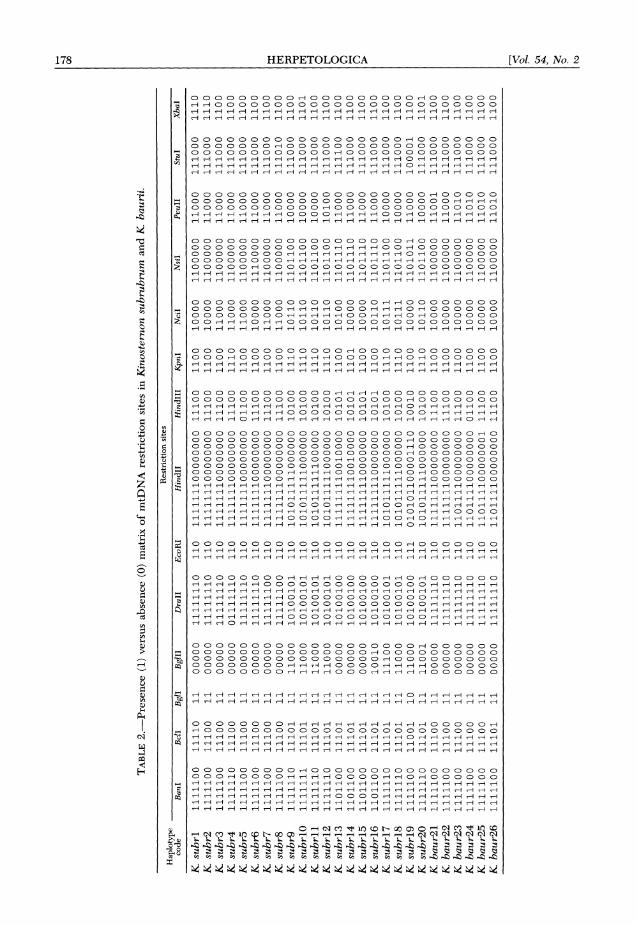
TABLE 2.-Presence (1) versus absence (0) matrix of mtDNA restriction sites in Kinostemnon subrubrum and K, baurii.
tRestriction sites Haplotype
code BanI BclI BglI BglII Drall EcoRI HindIl Hindill KpnI NciI NsiI PvuII StuI XbaI
K subrl 1111100 11110 11 00000 11111110 110 111111100000000 11100 1100 10000 1100000 11000 111000 1110 K subr2 1111100 11100 11 00000 11111110 110 111111100000000 11100 1100 100000 1100000 11000 111000 1110 K subr3 1111100 11100 11 00000 11111110 110 111111100000000 11100 1100 11000 1100000 11000 111000 1100 K subr4 1111110 11100 11 00000 01111110 110 111111100000000 11100 1110 11000 1100000 11000 111000 1100 K subr5 1111100 11100 11 00000 11111110 110 111111100000000 01100 1100 11000 1100000 11000 111000 1100 K subr6 1111100 11100 11 00000 11111110 110 111111100000000 11100 1100 10000 1110000 11000 111000 1100 K subr7 1111100 11100 11 00000 11111100 110 111111100000000 11100 1100 11000 1100000 11000 111000 1100 K subr8 1111100 11100 11 00000 11111100 110 111111100000000 11100 1100 11000 1100000 11000 111010 1100 1 K subr9 1111110 11101 11 11000 10100101 110 101011111000000 10100 1110 10110 1101100 10000 111000 1100 >
K subrlO 1111111 11101 11 11000 10100101 110 101011111000000 10100 1110 10110 1101100 10000 111000 1101 t K subrll 1111110 11101 11 11000 10100101 110 101011111100000 10100 1110 10110 1101100 10000 111000 1100 O
K subrl2 1111110 11101 11 110 00 10100101 110 10101111000000 10 100 1110 10110 1101100 1010 0 111000 1100 t K subrl3 1101100 11101 11 00000 10100100 110 111111110010000 10101 1100 1o0o1o00 1101110 110 00 111100 1100 O K subrl4 1101100 11101 11 00000 101 00100 110 111111110010000 10101 1101 10000 1101110 11000 111000 1100 2 K subrl5 1101100 11101 11 00000 1010 0100 110 111111110000000 10101 1100 1000 0oo0 1101110 1100 0 111000 1100 K subrl6 1101100 11101 11 10010 10100100 110 111111110000000 10101 1100 10110 1101110 11000 111000 1100 K subrl7 1111110 11101 11 11100 10100101 110 101011111000000 10100 1110 10111 1101100 10000 111000 1100 K subrl8 1111110 11101 11 11000 10100101 110 10101111000000 10100 1110 10111 1101100 10 000 111000 1100 K subrl9 1111100 11001 10 11000 10100100 1 010101100001110 100 10 1100 10000 1101011 11000 10 0001 1100 K subr20 1111110 11101 11 11001 10100101 110 101011111000000 10100 1110 10110 1101100 10000 111000 1101 K baur2l 1111100 11100 11 00000 11111110 110 111111100000000 11100 1100 10000 1100000 11001 111000 1100 K baur22 1111100 11100 11 00000 11111110 110 111111100000000 11100 1100 1000 0o0 110 0 0 0 0 1100 0 111000 1100 K baur23 1111100 11100 11 00000 11111110 110 110111100000000 11100 1100 10000 1100000 11010 111000 1100 K baur24 1111100 11100 11 00000 11111110 110 110111100000000 01100 1100 10000 1100000 11010 111000 1100 K baur25 1111100 11100 11 000o00 11111110 110 110111100000001 11100 1100 10000 100000 11010 111000 1100 K baur26 1111100 11101 11 00000 11111110 110 110111100000000 11100 1100 10000 1100000 11010 111000 1100
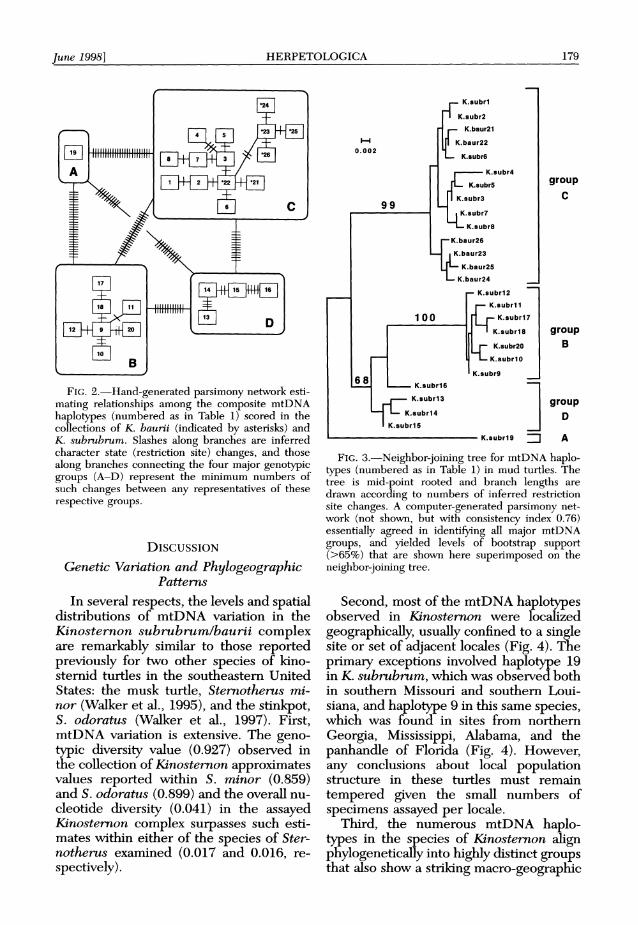
June 1998] HERPETOLOGICA 179
A
6C l / ~~~~~~~~~~~~2
17
18 1
1 3 D
B
FIG. 2.-Hand-generated parsimony network esti- mating relationships among the composite mtDNA haplotypes (numbered as in Table 1) scored in the collections of K. baurii (indicated by asterisks) and K. subrubrum. Slashes along branches are inferred character state (restriction site) changes, and those along branches connecting the four major genotypic groups (A-D) represent the minimum numbers of such changes between any representatives of these respective groups.
DISCUSSION
Genetic Variation and Phylogeographic Patterns
In several respects, the levels and spatial distributions of mtDNA variation in the Kinosternon subrubrum/baurii complex are remarkably similar to those reported previously for two other species of kino- sternid turtles in the southeastern United States: the musk turtle, Sternotherus mi- nor (Walker et al., 1995), and the stinkpot, S. odoratus (Walker et al., 1997). First, mtDNA variation is extensive. The geno- typic diversity value (0.927) observed in the collection of Kinosternon approximates values reported within S. minor (0.859) and S. odoratus (0.899) and the overall nu- cleotide diversity (0.041) in the assayed Kinosternon complex surpasses such esti- mates within either of the species of Ster- notherus examined (0.017 and 0.016, re- spectively).
K.subrl
K.subr2
K.baur2l
HI 4K.baur22 0.002
K.subr6
K.subr4
.subr5 group K.subr3 C
9 9 K.subr7
K.subr8
K.baur26
K.baur23
K.baur25
K.baur24
K.subrI2
K.subrl 1
1 00 K.subrl7
K.subr18 group K.subr20 B
Kasubri 0
K.subrg
16 8 K.subr16
K.subr3 group K.subri4 D
K.subrl 5
K.subrl9 J A
FIG. 3.-Neighbor-joining tree for mtDNA haplo- types (numbered as in Table 1) in mud turtles. The tree is mid-point rooted and branch lengths are drawn according to numbers of inferred restriction site changes. A computer-generated parsimony net- work (not shown, but with consistency index 0.76) essentially agreed in identifying all major mtDNA groups, and yielded levels of bootstrap support (>65%) that are shown here superimposed on the neighbor-joining tree.
Second, most of the mtDNA haplotypes observed in Kinosternon were localized geographically, usually confined to a single site or set of adjacent locales (Fig. 4). The primary exceptions involved haplotype 19 in K. subrubrum, which was observed both in southern Missouri and southern Loui- siana, and haplotype 9 in this same species, which was found in sites from northern Georgia, Mississippi, Alabama, and the panhandle of Florida (Fig. 4). However, any conclusions about local population structure in these turtles must remain tempered given the small numbers of specimens assayed per locale.
Third, the numerous mtDNA haplo- types in the species of Kinosternon align phylogenetically into highly distinct groups that also show a striking macro-geographic
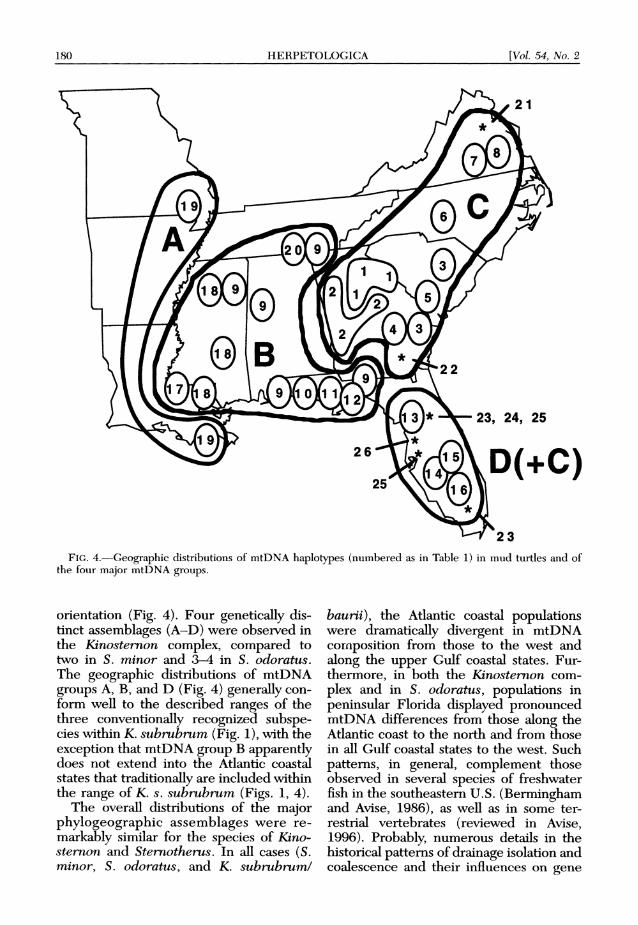
180 HERPETOLOGICA [Vol. 54, No. 2
E FAZ~~~~~~~~~~~~~~~~~~~~2 1
1 ~~~~~~~~~~~~O
2~~~~~~~~~
2 23
25D(C
FIG. 4.-Geographic distributions of mtDNA haplotypes (numbered as in Table 1) in mud turtles and of the four major mtDNA groups.
orientation (Fig. 4). Four genetically dis- tinct assemblages (A-D) were observed in the Kinosternon complex, compared to two in S. minor and 3-4 in S. odoratus. The geographic distributions of mtDNA groups A, B, and D (Fig. 4) generally con- form well to the described ranges of the three conventionally recognized subspe- cies within K. subrubrum (Fig. 1), with the exception that mtDNA group B apparently does not extend into the Atlantic coastal states that traditionally are included within the range of K. s. subrubrum (Figs. 1, 4).
The overall distributions of the major phylogeographic assemblages were re- markably similar for the species of Kino- sternon and Sternotherus. In all cases (S. minor, S. odoratus, and K. subrubrumi
baurii), the Atlantic coastal populations were dramatically divergent in mtDNA cornposition from those to the west and along the upper Gulf coastal states. Fur- thermore, in both the Kinosternon com- plex and in S. odoratus, populations in peninsular Florida displayed pronounced mtDNA differences from those along the Atlantic coast to the north and from those in all Gulf coastal states to the west. Such patterns, in general, complement those observed in several species of freshwater fish in the southeastern U.S. (Bermingham and Avise, 1986), as well as in some ter- restrial vertebrates (reviewed in Avise, 1996). Probably, numerous details in the historical patterns of drainage isolation and coalescence and their influences on gene

June 1998] HERPETOLOGICA 181
flow (Bermingham and Avise, 1986; Swift et al., 1985) have contributed to these ob- served genetic patterns in freshwater tur- tles.
Relationships between K. subrubrum and K. baurii
Considerable discussion has centered on the topic of morphological and taxonomic distinction between K. subrubrum and K. baurii. In an investigation of the subspe- cific status of a lower Florida Keys popu- lation of K. baurii, Iverson (1978) con- cluded that the highly variable color pat- terns on the head and carapace were un- reliable in distinguishing populations of K. baurii. According to Lamb (1983a,b), these patterns also cause occasional misi- dentifications with sympatric K. s. subru- brum. Lamb (1983a,b), using multivariate discriminant function analyses of morpho- metric characters, concluded that the two species in Florida and along the Atlantic Coast could be separated reliably. These analyses also demonstrated that the range of K. baurii extends into Georgia and South Carolina. A broader geographic sur- vey by Lamb and Lovich (1990) again in- dicated that the two species were distin- guishable along the Atlantic coast, and ex- tended the described range of K. baurii into southeastern Virginia. However, a lat- er assessment of samples of K. baurii against the western subspecies (hippocre- pis) of K. subrubrum failed to distinguish these taxa by the same morphometric cri- teria (Lovich and Lamb, 1995).
The current mtDNA restriction site data place all assayed samples morphologically referable to K. baurii in the C matrilineal group, which extends from southern Vir- ginia to southern peninsular Florida. All individuals of K. subrubrum collected from the Atlantic coastal states north of Florida also belong to this mtDNA C group. However, all peninsular Florida specimens referable by morphology to K. subrubrum belong to the sharply differ- entiable mtDNA D assemblage. Further- more, K. subrubrum across its broader geographic distribution to the west dis- plays at least two other matrilineal assem- blages that are at least as distinct from the
C and D groups as these latter are from one another. Finally, in contradistinction to morphological patterns mentioned above (Lovich and Lamb, 1995), samples of K. s. hippocrepis (assemblage A) proved highly divergent in mtDNA composition from K. baurii (assemblage C). Thus, with respect to matrilineal ancestry, K. subru- brum appears to be genealogically para- phyletic (Neigel and Avise, 1986) in rela- tion to K. baurii (Fig. 3).
One conceivable explanation for the ap- parent paraphyly is that the current mt- DNA restriction site data provide grossly inadequate descriptions of matrilineal re- lationships within the Kinosternon com- plex, perhaps because of scoring difficul- ties associated with inferences from diges- tion profiles alone. However, this is un- likely because we also have sequenced portions of the mtDNA control region from representative samples of K. subru- brum and K. baurii, and all conclusions about the major mtDNA phylogeographic groups within the complex are supported fully (Walker et al., 1998). Alternatively, several competing evolutionary explana- tions might account for the paraphyletic pattern observed. Perhaps K. baurtii and K. subrubrum are "good" biological species, as is suggested by partial sympatry and the morphometric differences, but K. baurii is a recent phylogenetic derivative of K. sub- rubrum. Consistent with this possibility is that K. baurii may have split recently from Atlantic-like populations of K. subrubrum, accounting for its overall mtDNA similar- ity to K. subrubrum in Atlantic coast drain- ages, and that the species secondarily in- vaded the Florida peninsula, thus account- ing for its strong mtDNA divergence from K. subrubrum in that area. Such a history also might account for the carapace and facial patterns wherein specimens of K. baurii tend to be easier to differentiate from K. subrubrum in Florida than along the Atlantic coast. On the other hand, his- torical introgressive hybridization in some other plant and animal taxa is known to produce occasional gene tree/species tree discordances (Avise, 1994). Perhaps sam- ples of K. subrubrum from the Atlantic coastal states are similar in mtDNA com-

182 HERPETOLOGICA [Vol. 54, No. 2
position to samples of K. baurii because gender-asymmetric hybridization has moved mtDNA of K. baurii into K. sub- rubrum, or vice versa, in this geographic area. Further evaluation of possibilities in- volving hybridization will require evidence from nuclear genes. In any event, the mtDNA data strongly indicate that any such hybridization has not genetically merged K. subrubrum and K. baurii in peninsular Florida.
It is possible that populations of K. bau- rii from the Atlantic coastal states always have been classified improperly as "K. sub- rubrum". If so, a revised range for K. sub- rubrum would include the Florida penin- sula and all relevant Gulf coastal and in- terior states to the west, but would not ex- tend northward along the Atlantic coast which instead is occupied solely by K. bau- rii. The two species thus overlap only in the Florida peninsula. Arguing against this possibility is the morphological separation between these two species in Atlantic coastal regions (Lamb 1983a,b; Lamb and Lovich, 1990; current study).
Another taxonomic revision imaginable would be to consider the highly divergent mtDNA groups A, B, and D within K. sub- rubrum to reflect the presence of distinct phylogenetic (and, or, biological) species. However, none of these or other such tax- onomic alterations can as yet be recom- mended with great certitude, because the genetic assays thus far are confined to a single "gene" (mtDNA). In principle, "gene trees" can differ from "species trees" for several plausible historical- demographic reasons, such as idiosyncratic sorting of gene-tree lineages in transitional populations that are large relative to inter- nodal times as measured in organismal generations (Maddison, 1995; Neigel and Avise, 1986; Pamilo and Nei, 1988). As emphasized by Avise and Ball (1990) and Avise and Wollenberg (1997), a firm de- marcation of taxonomic entities under ei- ther a biological species concept or a prop- erly formulated phylogenetic species con- cept ideally requires concordant historical inferences from multiple independent genes or the traits that they encode.
Acknowledgmnents.-We thank the following for help with the collections: R. Babb, I. Barak, V. Burke, M. Case, A. Davis, S. Doody, S. Emms, M. Goodis- man, M. Hare, C. Hobson, C. Holod, T. Ingstrom, D. Jansen, T. Johnson, A. Jones, B. Mansell, P. May- ne, J. Mitchell, B. Nelson, G. Ortf, P. Prodohl, F. Rose, C. Starlin, D. Stevenson, R. Vandevender, D. Wilson, K. Wood, and personnel from the Avise lab- oratory and The University of Georgia Golf Course. We also thank personnel from the National Fish Hatcheries of Bo Ginn, Carbon Hill, Tupelo, McKin- ney Lake, Orangeburg, Warm Springs, and Welaka. All specimens were collected under relevant state permits. Work was supported by a National Institutes of Health training grant to DeEtte Walker, by De- partment of Energy contract DE-FC09-96SR18546 between the U.S. Department of Energy and The University of Georgia's Savannah River Ecology Lab- oratory, and by a National Science Foundation grant to John Avise.
LITERATURE CITED
AVISE, J. C. 1994. Molecular Markers, Natural His- tory and Evolution. Chapman & Hall, New York, New York, U.S.A.
. 1996. Toward a regional conservation genet- ics perspective: phylogeography of faunas on the southeastern United States. Pp. 431-470. In J. C. Avise and J. L. Hamrick (Eds.), Conservation Ge- netics: Case Histories from Nature. Chapman & Hall, New York, New York, U.S.A.
AVISE, J. C., AND R. M. BALL, JR. 1990. Principles of genealogical concordance in species concepts and biological taxonomy. Oxford Surv. Evol. Biol. 7:45- 67.
AVISE, J. C., AND K. WOLLENBERG. 1997. Phyloge- netics and the origin of species. Proc. Natl. Acad. Sci. USA 94:7748-7755.
BERMINGHAM, E., AND J. C. AVISE. 1986. Molecular zoogeography of freshwater fishes in the south- eastern United States. Genetics 113:939-965.
CONANT, R., AND J. F. COLLINS. 1991. A Field Guide to Reptiles and Amphibians, Eastern and Central North America. Houghton Mifflin, Boston, Mas- sachusetts, U.S.A.
ERNST, C. H., AND R. W BARBOUR. 1989. Turtles of the World. Smithsonian Institution Press, Washing- ton, D.C., U.S.A.
ERNST, C. H., R. W BARBOUR, E. M. ERNST, AND J. R. BUTLER. 1974. Subspecific variation and inter- gradation in Florida Kinosternon subrubrum. Her- petologica 30:317-320.
ERNST, C. H., J. E. LovICH, AND R. W BARBOUR. 1994. Turtles of the United States and Canada. Smithsonian Institution Press, Washington D.C., U.S.A.
FELSENSTEIN, J. 1991. PHYLIP: Phylogeny Inference Package, Version 3.56. Department of Genetics, SK-50, University of Washington, Seattle, Washing- ton, U.S.A.
GIBBONS, J. W. 1983. Reproductive characteristics and ecology of the mud turtle, Kinosternon sub- rubrum (Lacepede). Herpetologica 39:254-271.

June 1998] HERPETOLOGICA 183
IVERSON, J. B. 1977. Kinosternon subrubrum. Cat. Am. Amphib. Rept. 193:1-4.
. 1978. Variation in striped mud turtles, Kin- osternon baurii (Reptilia, Testudines, Kinosterni- dae). J. Herpetol. 12:135-142.
. 1991. Phylogenetic hypotheses for the evo- lution of modern kinosternine turtles. Herpetol. Monogr. 5:1-27.
LAMB, T. 1983a. On the problematic identification of Kinosternon (Testudines: Kinosternidae) in Geor- gia, with new state localities for Kinosternon bauri. Georgia J. Sci. 41:115-120.
. 1983b. The striped mud turtle (Kinosternon bauri) in South Carolina, a confirmation through multivariate character analysis. Herpetologica 39: 383-390.
LAMB, T., AND J. LOvICH. 1990. Morphometric vali- dation of the striped mud turtle (Kinosternon bau- rni) in the Carolinas and Virginia. Copeia 1990:613- 618.
LANSMAN, R. A., R. 0. SHADE, J. F. SHAPIRA, AND J. C. AVISE. 1981. The use of restriction endonucleas- es to measure mitochondrial DNA sequence relat- edness in natural populations. III. Techniques and potential applications. J. Mol. Evol. 17:214-226.
LovICH, J. E., AND T. LAMB. 1995. Morphometric similarity between the turtles Kinosternon subru- brum hippocrepis and K. baurii. J. Herpetol. 29: 621-624.
MADDISON, W. 1995. Phylogenetic histories within and among species. Pp. 273-287. In P. C. Hoch and A. G. Stephenson (Eds.), Experimental and Molecular Approaches to Plant Biosystematics. Missouri Botanical Garden, St. Louis, Missouri, U.S.A.
MITCHELL, J. C. 1994. The Reptiles of Virginia. Smithsonian Institution Press, Washington, D.C., U.S.A.
NEI, M. 1987. Molecular Evolutionary Genetics. Co- lumbia University Press, New York, New York, U.S.A.
NEI, M., AND W-H. Li. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 76: 5269-5273.
NEIGEL, J. E., AND J. C. AVISE. 1986. Phylogenetic relationships of mitochondrial DNA under various demographic models of speciation. Pp. 515-534. In E. Nevo and S. Karlin (Eds.), Evolutionary Pro- cesses and Theory. Academic Press, New York, New York, U.S.A.
PAMILO, P., AND M. NEI. 1988. Relationships be- tween gene trees and species trees. Mol. Biol. Evol. 5:568-583.
SAITOU, N., AND M. NEI. 1987. The neighbor-joining method: a new method for reconstructing phylo- genetic trees. Mol. Biol. Evol. 4:406-425.
SEIDEL, M. E., J. B. IVERSON, AND M. D. ADKINS. 1986. Biochemical comparisons and phylogenetic relationships in the family Kinosternidae (Testudi- nes). Copeia 1986:285-294.
SITES, J. W, JR., J. W BICKHAM, M. W HAIDUK, AND J. B. IVERSON. 1979. Banded karyotypes of six taxa of kinostemid turtles. Copeia 1979:692-698.
SwiFT, C. C., C. R. GILBERT, S. A. BORTONE, G. H. BURGESS, AND R. W YERGER. 1985. Zoogeography of the southeastern United States: Savannah River to Lake Ponchartrain. Pp. 213-265. In C. H. Ho- cutt and E. 0. Wiley (Eds.), Zoogeography of the North American Freshwater Fishes. John Wiley & Sons, New York, New York, U.S.A.
SWOFFORD, D. L. 1990. PAUP: Phylogenetic Analysis Using Parsimony, Version 3.1.1. Illinois Natural History Survey, Champaign, Illinois, U.S.A.
WALKER, D., V. J. BURKE, I. BARAK, AND J. C. AVISE. 1995. A comparison of mtDNA restriction sites vs. control region sequences in phylogeographic as- sessment of the musk turtle (Sternotherus minor). Mol. Ecol. 4:365-373.
WALKER, D., W S. NELSON, K. A. BUHLMANN, AND J. C. AvISE. 1997. Mitochondrial DNA phylogeog- raphy and subspecies issues in the monotypic fresh- water turtle Sternotherus odoratus. Copeia 1997: 16-21.
WALKER, D., G. ORTI, AND J. C. AvISE. 1998. Phy- logenetic distinctiveness of a threatened aquatic turtle (Sternotherus depressus). Conserv. Biol.:In press.
Accepted: 17 July 1997 Associate Editor: David Green
APPENDIX I
Collection Sites
K. subrubrum: Altamaha River basin-Oconee River, Oconee Co., GA (n = 1); University of Georgia Golf Course, Clarke Co., GA (n = 2); Apalachicola River basin-Warm Springs National Fish Hatchery ponds, Meriwether Co., GA (n = 2); Camel Lake Road, east of SR 12, Liberty Co., FL (n = 1); Bayou Lafourche drainage-Laurel Plantation Road near Thibodaux, LaFourche Parish, LA (n = 1); Bayou Boeuf near Kraemer, LaFourche Parish, LA (n = 2); Bayou L'Ourse drainage-bayou on SR 70, Assump- tion Parish, LA (n = 6); Bayou L'Ourse, Assumption Parish, LA (n = 2); Coosa (Mobile) River basin- James Floyd State Park, Chattooga Co., GA (n = 3); Withlacoochee River basin-Hwy 19, 10 km. north of Crystal River, Citrus Co., FL (n = 1); Edisto River basin-Orangeburg National Fish Hatchery ponds, Orangeburg Co., SC (n = 2); Mississippi River ba- sin-cypress swamp, Big Cane Conservation Area, Butler Co., MO (n = 5); Ochlockonee River basin- County Road 268 near intersection with County Road 65B, Gadsden Co., FL (n = 1); Apalachicola National Forest Rd. 13, Liberty Co., FL (n = 1); Ogeechee River basin-ponds on Ft. Stewart Military Base, Bryan Co., GA (n = 5); Pascagoula River basin- Lake Ivy, Clarkco State Park, Clark Co., MS (n = 3); Pee Dee River basin-McKinney Lake National Fish Hatchery ponds, Richmond Co., NC (n = 3); St. Mark's River basin-Hwy 319 north of Tallahassee, Leon Co., FL (n = 1); pond south of Tallahassee near junction of Hwys. 319/263 and Hwy 363, Leon Co., FL (n = 2); Savannah River basin-Long Creek, Oglethorpe Co., GA (n = 1); Hwy 22 near Philomath, Oglethorpe Co., GA (n = 1); ditch near Tillman, Jas- per Co., SC (n = 1); Sopchoppy River basin-Apa-

184 HERPETOLOGICA [Vol. 54, No. 2
lachicola National Forest Rd 13, Wakulla Co., FL (n = 1); Tickfaw River basin-swamp on Hwy 15 near Pontchatook, Tangipohoa Parish, LA (n = 3); Tom- bigbee (Mobile) River basin-Carbon Hill National Fish Hatchery ponds, Walker Co., AL (n = 1); Tupelo National Fish Hatchery, Lee Co., MS (n = 2); Yellow River basin-Yellow River Flood Plain, Hwy 90, Oka- loosa Co., FL (n = 2); Shoal River at Hwy 90, Oka- loosa Co., FL (n = 1); York River basin-Cohoke Mill Creek, King William Co., VA (n = 3); Florida coastal integrated drainages-Chassahowitzka Wild- life Management Area, Hernando Co., FL (n = 2); Florida disjointed drainages-West Palm Beach, Palm Beach Co., FL (n = 2).
K. baurii: Indian River basin-Maytown Road
near Oak Hill, Volusia Co., FL (n = 1); Ogeechee River basin-Bo Ginn National Fish Hatchery, Jen- kins Co., GA (n = 1); St. John's River basin-Welaka National Fish Hatchery, Putnam Co., FL (n = 1); Hwy 441, Payne's Prairie, Alachua Co., FL (n = 1); Co. Rd 346 near Hwy 121, Alachua Co., FL (n = 2); Withlacoochee River basin-Hwy 471 south of Tar- rytown, Sumter Co., FL (n = 1); Hwy 19, 10 km. north of Crystal River, Citrus Co., FL (n = 1); Hwy 50, approximately 4 km west Sumter Co. line, Her- nando Co., FL (n = 1); York River basin-Cohoke Mill Creek, King William Co., VA (n = 4); Florida coastal integrated drainages-Snapper Creek Canal, Dade Co., FL (n = 4); Florida disjointed drainages- Dade/Collier Training Airport, Collier Co., FL (n = 1).
Herpetologica, 54(2), 1998, 184-206 ? 1998 by The Herpetologists' League, Inc.
THE PHYLOGENETIC POSITION OF THE MEXICAN BLACK- TAILED PITVIPER (SQUAMATA: VIPERIDAE: CROTALINAE)
RONALD L. GUTBERLET, JR.
Department of Biology, Box 19498, The University of Texas at Arlington, Arlington, TX 76019-0498, USA
ABSTRACT: Phylogenetic analyses of 52 morphological characters from Agkistrodon contortrix, Atropoides nummifer, Bothriechis bicolor, B. lateralis, B. nigroviridis, B. schlegelii, Bothrops asper, Cerrophidion godmani, Gloydius blomhoffii, Ophryacus undulatus, Porthidium melanurum, P. na- sutum, and P. ophryomegas indicate that the Mexican black-tailed pitviper (Porthidium melanurum) is more closely related to Ophryacus undulatus than it is to its congeners P. nasutum and P. ophryomegas. To achieve a monophyletic classification, P. melanurum is placed in the genus Ophry- acus.
Key words: Crotalinae; Morphology; Ophryacus; Phylogenetic systematics; Porthidium; Porthi- dium melanurum; Viperidae
RECENT investigations into relationships among Neotropical pitvipers have led to the generic recognition of several putative- ly monophyletic groups. In an unpublished doctoral dissertation, Burger (1971) divid- ed the morphologically diverse and poten- tially polyphyletic genus Bothrops into five genera: Bothriechis, Bothriopsis, Bothrops, Ophryacus, and Porthidium. Subsequent- ly, Perez-Higareda et al. (1985) published this taxonomic arrangement. Campbell and Lamar (1989) adopted this classifica- tion but suggested that additional studies into the relationships and generic limits of these groups, especially Porthidium, were needed.
Further study revealed that Porthidium
(sensu Burger, in Perez-Higareda et al., 1985) is polyphyletic, and new genera were proposed to rectify this unnatural grouping. Werman (1992) erected the ge- nus Atropoides, the jumping pitvipers, to contain three of the 14 species formerly included in Porthidium: A. nummifer, A. olmec, and A. picadoi. Campbell and La- mar (1992) segregated an additional three species of Porthidium into Cerrophidion, the montane pitvipers: C. barbouri, C. godmani, and C. tzotzilorum.
These revisions reduced the content of Porthidium to eight species. Solorzano (1994) later increased the number of spe- cies in Porthidium to nine with the de- scription of P. volcanicum. Eight of these