high-molecular-weight linear multimer formation by single ... · linearreplication products of...
TRANSCRIPT

Vol. 174, No. 1JOURNAL OF BACTERIOLOGY, Jan. 1992, p. 173-1780021-9193/92/010173-06$02.00/0Copyright © 1992, American Society for Microbiology
High-Molecular-Weight Linear Multimer Formation by Single-Stranded DNA Plasmids in Escherichia coliPATRICK DABERT,* S. DUSKO EHRLICH, AND ALEXANDRA GRUSS
Laboratoire de Genetique Microbienne, Institut de Biotechnologie, Institut National de la Recherche AgronomiqueDomaine de Vilvert, 78352 Jouy en Josas Cedex, France
Received 19 July 1991/Accepted 29 October 1991
We inserted foreign DNA segments into plasmids which replicate by a rolling-circle mechanism inEscherichia coli and observed the appearance of high-molecular-weight plasmid multimers (HMW). Thisphenomenon, which occurs more frequently with GC-rich segments, depends on the mode of replication of theplasmid and on host homologous recombination functions. We found that (i) HMW are formed upon insertionof a foreign DNA segment into a single-stranded DNA plasmid, whereas the same DNA insert has no such effecton a theta replicon, and (ii) HMW are not present in a recA mutant strain but are found in a lexA(Ind-)mutant. Enzymatic studies allowed us to define the HMW structure as linear double-stranded tandemhead-to-tail plasmid repeats. Use of heteroplasmid strains showed that HMW production by one plasmid doesnot affect another resident plasmid, indicating that no host functions are phenotypically inactivated. Thisdistinguishes our system from the HMW observed with various replicons in the absence of RecBCD enzymeactivity. We propose that the role of the foreign insert is to protect the DNA from RecBCD exonuclease attack.
Extrachromosomal elements have been shown to undergoone of two basic types of replication, theta and rolling-circlereplication (RCR). Most plasmids found in gram-negativebacteria replicate by a theta mechanism, whereas numerousplasmids of gram-positive bacteria replicate by RCR (25; fora review, see references 8 and 17). Intermediates of RCR arepresumably u-shaped molecules and circular single-strandedDNA (ssDNA) (24, 25); these replicons are referred to hereas ssDNA plasmids. For both types of plasmids, the repli-cation products are usually covalently closed circular mono-mers (12, 24). However, two cases have been described inwhich major replicative products are high-molecular-weightplasmid linear multimers (HMW). In one case, this drasticchange of plasmid distribution is due to a cell phenotypemodification; in the other, it is due to a structural modifica-tion of the plasmid.E1MW produced by cell phenotype modification. In Esche-
richia coli, genotypic or phenotypic inactivation of RecBCDexonuclease was reported to induce HMW formation withtheta-replicating plasmids, such as pl5A or ColEl deriva-tives (4). This phenomenon is greatly enhanced by hostmutations such as sbcA or sbcB sbcC. In the latter context,HMW production necessitates de novo DNA synthesis (21)which does not initiate by the normal plasmid mode ofreplication (20). It has been proposed that random nicks inthe plasmids serve as priming sites for nonterminating RCR,resulting in a displaced ssDNA which is converted to double-stranded DNA (dsDNA). In a recBCD+ background, thelinear dsDNA ends produced by RCR would be degradedand no HMW would be formed.HMW produced by structural modification of the plasmid.
HMW was shown to be induced in wild-type Bacillus subtilisstrains by the insertion of certain foreign DNA segments intossDNA plasmids (7). HMW production depends on the RCRplasmid mode of replication since it is not produced by atheta replicon containing the same inserts. Although dimin-ished amounts ofHMW were observed in a recE4 B. subtilis
* Corresponding author.
strain (deficient in homologous recombination; see reference29 for a review), it was found that interplasmidic recombi-nation is not a major mechanism by which HMW aregenerated (7).To ascertain that the plasmid mode of replication rather
than host-specific factors determine HMW production, weintroduced ssDNA plasmids into recombination-proficientE. coli. It has been previously shown that ssDNA plasmidscan be established in E. coli and replicate by the same RCRmechanism (25). We found that insertion of foreign DNAfragments into these plasmids provokes HMW formation.Theta replicons do not make HMW under the same condi-tions. This observation for E. coli allows us to make a directcomparison with the type ofHMW formed in a recBC sbcBCmutant (4). We show that HMW formation is not a conse-quence of a phenotypic inactivation of RecBCD or of otherloci, indicating that the foreign insert has a specific cis effecton the carrier plasmid.
MATERIALS AND METHODS
Bacterial strains and plasmids. E. coli strains used in thisstudy are presented in Table 1. Plasmids pHV903 andpHV950 contain the replicon of ssDNA plasmid pC194 (10).They were constructed in this laboratory by deletions withinplasmid pHV33 (18): deletion of the PvuII-tetracycline frag-ment gave rise to plasmid pHV903 (pHV33APvu in reference25), and subsequent deletion of the HaeII-pBR322 replica-tion origin fragment led to pHV950 (Fig. 1). These plasmidscontain the rriB conversion site of pBR322, which facilitatesthe conversion of the monomeric circular ssDNA interme-diate to dsDNA in E. coli (25). pHV903 and pHV950 wereobtained from M. F. Gros. pNZ12 and pNZ18 are ssDNAplasmids with cloned antibiotic resistance markers (kanamy-cin and chloramphenicol resistance) and were kindly pro-vided by W. M. de Vos.
Culture media and cultural conditions. Cultures weregrown in L broth medium; when appropriate, chloramphen-icol (10 ,ug/ml for pNZ12, pNZ18, and pHV950; 25 ,ug/ml forpACYC184 [3]) and ampicillin (100 ,ug/ml) were added.
173
on Novem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
174 DABERT ET AL.
TABLE 1. Bacterial strains
Strain Description Source
HVC45 leu-6 thi-J pro lacYI tonA supE44 This laboratoryhsdR str
JM1o1 A(lac pro) thi supE F'traD36 J. MessingproAB lacPZ AMJ5
AB1157 thr-J leu-6 proA2 his4 thi-I A. J. ClarkargE3 lacYl galK2 ara-14 xyl-5mtl-l tsx-33 supEE44 rpsL31kdgK5J
HVC3256 Like AB1157, but srl::TnlO A. J. ClarkrecA306
GY6781 AsfiA::lacZ A(lac pro) gal' R. Devoretmal::Tn9 lexAl rpsL
JC7623 Like AB1157, but recB21 recC22 A. J. ClarksbcBJ5 sbcC201
Bacterial transformation. Plasmids were introduced bystandard transformation methods into wild-type E. colistrains (15) and transferred into mutant strains by electropo-ration (5). We observed that certain plasmids were notreadily established in the recA strains by using the standardtechnique (see Results).DNA preparation and enzymatic treatment. Whole-cell
DNA minilysates (19), plasmid DNA (1), and total DNA (7)were prepared by published methods. Exonuclease andendonuclease treatments were performed according to thesupplier's instructions.
Detection ofHMW by gel electrophoresis. Whole-cell DNAminilysates were run on 0.5% agarose gels prepared inTris-borate-EDTA buffer (15) containing ethidium bromide(0.4 ,uglml) and were electrophoresed at 2.4 to 2.8 V/cmovernight. Supercoiled Ladder (Bethesda Research Labora-tories) was used for plasmid size standards. HMW weredetected by Southern blot hybridization of gels (23) withnick-translated or random-primed plasmid DNA-labelledprobes (kits from Boehringer Mannheim).
pC194
rri %U'pvunFIG. 1. Structure of pHV950. pHV950 is an ssDNA plasmid
derived from staphylococcal plasmid pC194 (heavy line) carryingthe ampicillin (Ap) resistance gene and the rriB conversion site ofpBR322 (thin line). This plasmid replicates by an RCR mechanismand expresses chloramphenicol (Cm) and ampicillin resistance in E.coli. Foreign DNA fragments were cloned between the EcoRI andHindIll sites. ori pC194 and RepA refer to the origin and protein ofreplication of ssDNA plasmid pC194, respectively.
HMW
c.c.c. monomer
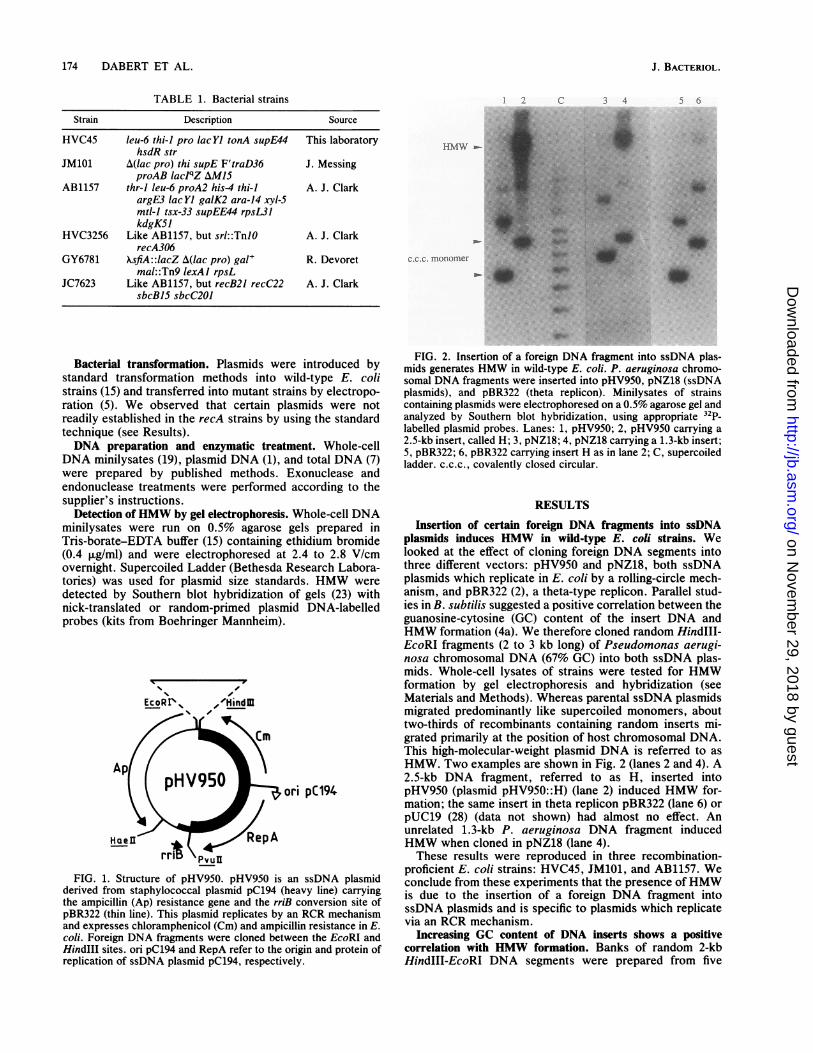
FIG. 2. Insertion of a foreign DNA fragment into ssDNA plas-mids generates HMW in wild-type E. coli. P. aeruginosa chromo-somal DNA fragments were inserted into pHV950, pNZ18 (ssDNAplasmids), and pBR322 (theta replicon). Minilysates of strainscontaining plasmids were electrophoresed on a 0.5% agarose gel andanalyzed by Southern blot hybridization, using appropriate 32p_labelled plasmid probes. Lanes: 1, pHV950; 2, pHV950 carrying a2.5-kb insert, called H; 3, pNZ18; 4, pNZ18 carrying a 1.3-kb insert;5, pBR322; 6, pBR322 carrying insert H as in lane 2; C, supercoiledladder. c.c.c., covalently closed circular.
RESULTS
Insertion of certain foreign DNA fragments into ssDNAplasmids induces HMW in wild-type E. coli strains. Welooked at the effect of cloning foreign DNA segments intothree different vectors: pHV950 and pNZ18, both ssDNAplasmids which replicate in E. coli by a rolling-circle mech-anism, and pBR322 (2), a theta-type replicon. Parallel stud-ies in B. subtilis suggested a positive correlation between theguanosine-cytosine (GC) content of the insert DNA andHMW formation (4a). We therefore cloned random HindIll-EcoRI fragments (2 to 3 kb long) of Pseudomonas aerugi-nosa chromosomal DNA (67% GC) into both ssDNA plas-mids. Whole-cell lysates of strains were tested for HMWformation by gel electrophoresis and hybridization (seeMaterials and Methods). Whereas parental ssDNA plasmidsmigrated predominantly like supercoiled monomers, abouttwo-thirds of recombinants containing random inserts mi-grated primarily at the position of host chromosomal DNA.This high-molecular-weight plasmid DNA is referred to asHMW. Two examples are shown in Fig. 2 (lanes 2 and 4). A2.5-kb DNA fragment, referred to as H, inserted intopHV950 (plasmid pHV950::H) (lane 2) induced HMW for-mation; the same insert in theta replicon pBR322 (lane 6) orpUC19 (28) (data not shown) had almost no effect. Anunrelated 1.3-kb P. aeruginosa DNA fragment inducedHMW when cloned in pNZ18 (lane 4).These results were reproduced in three recombination-
proficient E. coli strains: HVC45, JM101, and AB1157. Weconclude from these experiments that the presence ofHMWis due to the insertion of a foreign DNA fragment intossDNA plasmids and is specific to plasmids which replicatevia an RCR mechanism.
Increasing GC content of DNA inserts shows a positivecorrelation with HMW formation. Banks of random 2-kbHindIII-EcoRI DNA segments were prepared from five
J. BACTERIOL.
on Novem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

LINEAR REPLICATION PRODUCTS OF ssDNA PLASMIDS 175
TABLE 2. Correlation of HMV production with GC content ofinserts
%GCof No. of HMWSource of DNA inserts inseota producers/totalinsert" inserts testedb
Staphylococcus aureus 34 0/6Lactococcus lactis 38 0/7Bacillus subtilis 42 1/10Salmonella typhimurium 51 3/6Pseudomonas aeruginosa 67 7/11
aAverage values taken from published data (17a).b HMW production was tested in recombination-proficient E. coli JM101 as
described in Materials and Methods. The ssDNA plasmid vector used waspHV903.
bacterial species varying greatly in their average GC con-tents. These segments were cloned into plasmid pHV903,which contains an active ssDNA replicon. The resultantplasmids were tested for HMW formation (Table 2). Weobserved that the proportion of HMW producers increaseswith increasing percent GC of the insert DNA. The preciseGC contents of several of the cloned segments were deter-mined by a novel polymerase chain reaction method devel-oped in this laboratory, and they were found to be near theaverage for the strain (9). These results suggest that thequality of the DNA insert is important in HMW formation. Itshould be noted that HMW formation is not induced by allGC-rich DNA insertions, suggesting that some feature otherthan percent GC is involved. Several of these inserts werealso tested for their abilities to provoke HMW formation inB. subtilis. We observed that the DNA inserts provokingHMW formation in E. coli and in B. subtilis are not the same.These experiments indicate that an important role of the hostis to discriminate between sequences.HMW are composed of linear head-to-tail plasmid multim-
ers. The structure of HMW generated by a representativeplasmid, pHV950::H, was determined by different enzy-matic approaches. Whole-cell lysates of strains containingpHV950::H were treated with lambda exonuclease or exo-nuclease III (Fig. 3A). Lambda exonuclease degrades lineardsDNA from the 5' end, whereas exonuclease III degrades itfrom the 3' end. The amount ofHMW was greatly decreasedafter each exonuclease treatment. The reaction was specific,as supercoiled plasmid DNA remained intact. We concludethat the majority ofHMW is in linear form, with at least onedsDNA end accessible to exonucleases. The residual hybrid-ization observed after exonuclease treatment may corre-spond to ssDNA digestion products or may indicate that theends of a small proportion of HMW are protected (13, 16).These results eliminate the possibility that HMW are com-posed of catenated plasmids. DNA samples were alsotreated with restriction endonucleases which do not cut(BglII) or cut once in the plasmid (HindIII) (Fig. 3B).Treatment with BgII digests chromosomal DNA but has noeffect on HMW migration. However, after HindIlI treat-ment, all the hybridization shifts to the size of linear double-stranded plasmid monomeric DNA. Furthermore, partialHindIII digestion yielded a ladder of pHV950::H linearoligomers extending up to at least a 12-mer, as revealed bypulsed-field electrophoresis (data not shown). This indicatesthat HMW are organized in a linear head-to-tail fashion; theycould be present as a-shaped molecules or large plasmidconcatemers that are broken during manipulation.EMW formation is greatly reduced in a recA mutant strain.
A B1 2 3
A NHMW ).
linear monomer )P-
(c.c.c. monomer SII
FIG. 3. HMW are composed of linear double-stranded head-to-tail plasmid multimers. Whole-cell lysates of strains containingpHV950: :H were submitted to exonuclease or endonuclease diges-tion. Samples were run on a 0.5% agarose gel and analyzed bySouthern blot hybridization, using a 32P-labelled plasmid as probe.(A) Lanes 1 and 3, untreated pHV950::H; lane 2, pHV950::H treatedwith lambda exonuclease; lane 4, pHV950: :H treated with exonu-clease III. (B) Lane 1, untreated pHV950::H; lane 2, pHV950::Htreated with Bglll (no site on the plasmid); lane 3, pHV950::Htreated with HindIlI (single site on the plasmid). c.c.c., covalentlyclosed circular.
To see whether homologous recombination functions areinvolved in HMW formation, we transformed a recA mutantstrain (HVC3256) with pHV950::H. Compared with theparental plasmid with no insert, pHV950::H transformedinefficiently, and electroporation had to be used to establishthe plasmid. Gel hybridization (Fig. 4) shows that the level ofHMW is greatly reduced in the recA mutant (lane 2) com-pared with that of the wild-type strain (lane 1). A proportionofHMW remains in the well in lanes 1, 3, and 4; this may bedue to the large size or complex structure of this DNA (seeprevious section).RecA is multifunctional and may exert its effects onHMW
by its role in induction of the SOS repair pathway (26),homologous recombination (22), and/or its protection fromnuclease degradation (reckless degradation phenotype [27]).Furthermore, it may intervene in the formation ofHMW orin its stabilization. Results of two experiments indicate thatthe SOS pathway is not involved. First, we found that HMWare produced in high amounts in a lexA(Ind-) (SOS nonin-ducible) strain (GY6781; Fig. 4, lane 3). Second, we studiedHMW in a recA sbcA double mutant; a recA mutation issuppressed by sbcA for intramolecular recombination (6, 14)but not for SOS pathway induction. HMW are formed in therecA sbcA mutant (Fig. 4, lane 4) at nearly the same level asin the wild-type strain (Fig. 4, lane 1). These results indicate
VOL. 174, 1992
on Novem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

176 DABERT ET AL.
1 2 3 4
HMW
PROBE A PROBE B
1 2 3
_ HMW _
" A
B
c.c.c. monomer -
FIG. 4. HMW formation is greatly reduced in a recA mutantstrain. Minilysates of wild-type (lane 1), recA (lane 2), lexA(Ind-)(lane 3), and recA sbcA (lane 4) strains containing pHV950::H were
run on an agarose gel and analyzed by Southern blot hybridization,using a 32P-labelled plasmid probe. c.c.c., covalently closed circu-lar.
that a step in recombination is necessary, but that SOSinduction is not necessary, for HMW synthesis.
Production ofHMW is not due to a phenotypic alteration inthe cell. The formation of HMW in a recBC sbcB strain iswell documented (4, 16). It is explained by the inactivationof a trans-acting factor, the RecBCD exonuclease, and/oraltered expression of other factors, which affects all plas-mids present in a given cell. We tested the possibility thatan ssDNA plasmid containing a foreign insert may inacti-vate or titrate RecBCD or another product; this would im-ply that any other plasmid in the same cell would makeHMW.A heteroplasmid strain was constructed in a wild-type
background containing pHV950::H and an ssDNA plasmid,pNZ12, which does not generate HMW. If the presence ofpHV950::H provokes phenotypic inactivation of RecBCD,then both plasmids present in the heteroplasmid cell shouldmake HMW. Plasmid pHV950::H generates HMW in awild-type context either alone or in the presence of pNZ12(Fig. 5, probe A, lanes 1 and 2). However, there are nodetectable HMW produced by pNZ12 in any wild-typecontext even if an HMW-producing plasmid is present(Fig. 5, probe B, lanes 2 and 3). Similar results were ob-tained with the heteroplasmid strain carrying pHV950::H andthe theta replicon pACYC184 (data not shown). These re-
sults show that in a wild-type host, production of HMWby one plasmid does not induce production of HMW byanother plasmid present in the same cell. We conclude thatthe insert has a cis effect on the carrier plasmid whichproduces HMW.
FIG. 5. Production of HMW is not a consequence of RecBCDinactivation. Minilysates of wild-type strains containing one or twoplasmids were run in duplicate on an agarose gel (left panel) and thentested separately by Southern blot hybridization with 32P-labelledprobes corresponding to each plasmid (middle and right panels).Since a decrease in pHV950::H copy number was observed whenpNZ12 was present in the same strain, the exposures of the probe Aautoradiographs were adjusted to give comparable intensities ofplasmid monomers. Probes A and B correspsond to plasmidspHV950::H and pNZ12, respectively. Lanes: 1, plasmid pHV950::Halone; 2, plasmids pHV950::H and pNZ12; 3, plasmid pNZ12 alone.
DISCUSSION
Plasmids which replicate by RCR have been found in awide variety of organisms (8) and often show a broad hostrange that includes E. coli (25). If such plasmids acquireforeign DNA (either naturally or artificially), large amountsof HMW may accumulate in the cells. An analysis of themechanism by which ssDNA plasmids containing foreignDNA inserts produce HMW has been undertaken withwild-type E. coli.We considered three parameters which may affect HMW
production: the plasmid vector, the foreign insert, and hostfunctions. We found the following. (i) HMW production isdependent on the rolling circle mode of replication of thevector and is not species specific. Furthermore, the produc-tion of HMW by one plasmid does not provoke its produc-tion by another plasmid in the same cell. (ii) Amounts ofHMW vary according to the nature of the insert, showing apositive correlation with increasing GC content. However,not all GC-rich inserts induce HMW production. (iii) Certainhost homologous recombination functions are important forHMW production: recA-deficient strains do not produceHMW, except when complemented by sbcA, and lexA(Ind-)strains produce nearly the same amounts of HMW as wild-type strains. Both results indicate that SOS functions are notrequired.How is HMW formed in a wild-type cell? The ssDNA
plasmids produce cr-shaped molecules as replication inter-mediates, while theta replicons do not. This difference isimplicated in HMW formation, since plasmids which repli-
J. BACTERIOL.
on Novem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

LINEAR REPLICATION PRODUCTS OF ssDNA PLASMIDS 177
cate by RCR, and not by a theta mechanism, can form HMWin a wild-type cell. We propose that HMW could be formedif replication initiated properly but termination did notoccur. This would generate a a-shaped molecule with a longdsDNA tail. Such a molecule would be expected to be asubstrate for RecBCD exonuclease. Why, then, is HMWproduced by ssDNA plasmids in the wild-type backgroundnot degraded by RecBCD enzyme? Two possible reasons areconsidered. First, the ends of RCR intermediates may beprotected from exonucleolytic attack by the binding of thereplication protein to the 5' end of the displaced strand (11).Such a hypothesis would predict that all ssDNA plasmidsproduce HMW regardless of the presence of an insert, whichis not the case. Second, the insert DNA is resistant toexonucleolytic attack by RecBCD and thus blocks degrada-tion. The latter hypothesis is consistent with our results, asonly the ssDNA plasmids containing inserts make HMW.We therefore propose that the foreign inserts provokingHMW formation actually protect the structure from exonu-cleolytic degradation. If this is the case, the DNA sequenceswhich protect against degradation in E. coli or in B. subtilisare not the same, since we observed that certain insertsmake HMW in one organism but not in the other. Themechanism by which these inserts induce HMW formationmay issue from the observation that increases in insert sizeand GC content also increase the probability of HMWproduction in E. coli.We have demonstrated that recombination is needed for
HMW production. One role of recombination, as proposedfor HMW formation in recBC sbcBC strains, is to amplifythe complexity of multimers (4). However, we showedpreviously for B. subtilis (7) that two homologous HMW-producing plasmids in the same strain do not produce"patchwork" HMW molecules. This result ruled out multi-ple rounds of intermolecular recombination as a mechanismfor the formation of HMW in our system. In two othermodels, the RCR mechanism itself allows recombination:the tails of a-shaped molecules, which are normally suscep-tible to exonuclease degradation, may become frayed (4, 16).Niki et al. (16) proposed that free ssDNA ends are protectedfrom exonuclease degradation by RecA through the forma-tion of synaptic complexes with other molecules. In a secondmodel, recombination could be required in an initial step ofHMW formation to displace the replication fork onto asecond template; in this case, RCR would continue unim-peded. Either role for recombination is consistent with ourpresent data.
ACKNOWLEDGMENTSWe thank A. Cohen, W. de Vos, R. Devoret, and M. F. Gros for
their gifts of bacterial strains or plasmids. We express our appreci-ation to C. Agnanostopoulos, E. Maguin, and P. Langella for criticalreading of the manuscript, E. Dervyn and P. Duwat for frequentdiscussions of the work, and V. Moretto for technical assistance.Artwork was kindly prepared by V. Akueson.
REFERENCES1. Birnboin, H., and J. Doly. 1979. A rapid alkaline extraction
procedure for screening recombinant plasmid DNA. NucleicAcids Res. 7:1513-1523.
2. Bolivar, F., R. Rodriguez, P. Greene, M. Betlach, H. Heynecker,H. Boyer, J. Crosa, and S. Falkow. 1977. Construction andcharacterization of new cloning vehicles. II. Multipurpose clon-ing system. Gene 2:95-113.
3. Chang, A. C. Y., and S. N. Cohen. 1978. Construction andcharacterization of amplifiable multicopy DNA cloning vehiclesderived from p1SA, the cryptic miniplasmid. J. Bacteriol. 134:
1141-1156.4. Cohen, A., and A. J. Clark. 1986. Synthesis of linear plasmid
multimers in Escherichia coli K-12. J. Bacteriol. 167:327-335.4a.Dabert, P., and A. Gruss. Unpublished data.5. Dower, W. J., J. F. Miller, and C. W. Ragsdale. 1988. High
efficiency transformation of Escherichia coli by high voltageelectroporation. Nucleic Acids Res. 16:6127-6145.
6. Fishel, R. A., A. A. James, and R. Kolodner. 1981. RecAindependent general recombination of plasmids. Nature (Lon-don) 294:184-186.
7. Gruss, A., and S. D. Ehrlich. 1988. Insertion of foreign DNAinto plasmids from gram-positive bacteria induces formation ofhigh-molecular-weight plasmid multimers. J. Bacteriol. 170:1183-1190.
8. Gruss, A., and S. D. Ehrlich. 1989. The family of highlyinterrelated single-stranded deoxyribonucleic acid plasmids.Microbiol. Rev. 53:231-241.
9. Gruss, A., V. Moretto, S. D. Ehrlich, P. Duwat, and P. Dabert.1991. GC-rich DNA sequences block homologous recombina-tion in vitro. J. Biol. Chem. 266:6667-6669.
10. Horinouchi, S., and B. Weisblum. 1982. Nucleotide sequenceand functional map of pC194, a plasmid that specifies chloram-phenicol resistance. J. Bacteriol. 150:815-825.
11. Koepsel, R. R., R. W. Murray, W. D. Rosenblum, and S. A.Khan. 1985. The replication initiator protein of plasmid pT181has sequence-specific endonuclease and topoisomerase-like ac-tivities. Proc. Natl. Acad. Sci. USA 82:6845-6849.
12. Kornberg, A. 1980. DNA replication. W. H. Freeman and Co.,San Francisco.
13. Kusano, K., K. Nakayama, and H. Nakayama. 1989. Plasmid-mediated lethality and plasmid multimer formation in an Esch-erichia coli recBC sbcBC mutant: involvement of RecF recom-bination pathway genes. J. Mol. Biol. 209:623-634.
14. Laban, A., and A. Cohen. 1981. Interplasmidic and intraplas-midic recombination in Escherichia coli K-12. Mol. Gen. Genet.184:200-207.
15. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
16. Niki, H., T. Ogura, and S. Hiraga. 1990. Linear multimerformation of plasmid DNA in Escherichia coli hopE recDmutants. Mol. Gen. Genet. 224:1-9.
17. Novick, R. P. 1989. Staphylococcal plasmids and their replica-tion. Annu. Rev. Microbiol. 43:537-565.
17a.Owen, R. J. 1982. National Collection of Type Cultures, CentralPublic Health Laboratory, London.
18. Primrose, S. B., and S. D. Ehrlich. 1981. Isolation of plasmiddeletion mutants and study of their instability. Plasmid 6:193-201.
19. Projan, S., S. Carlton, and R. P. Novick. 1983. Determination ofplasmid copy number by fluorescence densitometry. Plasmid9:182-190.
20. Silberstein, Z., and A. Cohen. 1987. Synthesis of linear multim-ers of OriC and pBR322 derivatives in Escherichia coli K-12:role of recombination and replication functions. J. Bacteriol.169:3131-3137.
21. Silberstein, Z., S. Maor, I. Berger, and A. Cohen. 1990. LambdaRed-mediated synthesis of plasmid linear multimers in Esche-richia coli K-12. Mol. Gen. Genet. 223:496-507.
22. Smith, G. R. 1988. Homologous recombination in procaryotes.Microbiol. Rev. 52:1-28.
23. Southern, E. 1975. Detection of specific sequences among DNAfragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
24. te Riele, H., B. Michel, and S. D. Ehrlich. 1986. Single-strandedplasmid DNA in Bacillus subtilis and Staphylococcus aureus.Proc. Natl. Acad. Sci. USA 83:2541-2545.
25. te Riele, H., B. Michel, and S. D. Ehrlich. 1986. Are single-stranded circles intermediates in plasmid DNA replication?EMBO J. 5:631-637.
26. Walker, G. C. 1987. The SOS response of Escherichia coli, p.1346-1357. In F. C. Neidhardt, J. L. Ingraham, K. B. Low, B.Magasanik, M. Schaechter, and H. E. Umbarger (ed.), Esche-
VOL. 174, 1992
on Novem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

178 DABERT ET AL.
richia coli and Salmonella typhimurium: cellular and molecularbiology, vol. 2. American Society for Microbiology, Washing-ton, D.C.
27. Willetts, N. S., and A. J. Clark. 1969. Characteristics of somemultiply recombination-deficient strains of Escherichia coli. J.Bacteriol. 100:231-239.
28. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. Improved
M13 phage cloning vectors and host strains: nucleotide se-quences of the M13mp18 and pUC19 vectors. Gene 33:103-119.
29. Yasbin, R. E., M. Stranathan, and K. Bayles. 1991. TherecE(A)+ gene of B. subtilis and its gene product: furthercharacterization of this universal protein. Biochimie 73:245-250.
J. BACTERIOL.
on Novem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from