hills district branch · cover photo: 2012 australian national 1st place normal greygreen owned by...
TRANSCRIPT

Hills District Branch BSNSW Inc.
Established 1974
NEWSLETTER JUNE 2012

Cover Photo: 2012 Australian National
1st Place Normal Greygreen Owned by The Welshman Family
Photo by Nola Bradford
Hills District Branch BS NSW Inc. Committee 2011-2012
Position Name Address P/Code Phone Email
Chairman Peter Dodd 7 Rocca St Denistone East 2112 9888 5631 [email protected]
Vice Chair. Harry Charalambous PO Box 990 Wahroonga 2076 O417209416 [email protected]
Vice Chair. Craig Buckingham 44 Barellan Ave Carlingford 2118 O408602381 [email protected]
Secretary Kathy Manton 44 Clifton Rd Marsden Park 2765 9627 7748 [email protected]
Treasurer Ian Manton 44 Clifton Rd Marsden Park 2765 9627 7748 [email protected]
Show Manager Craig Buckingham As above
Show Manager Richard Abraham 3 Edwards Rd Middle Dural 2158 9651 3689 [email protected]
Show Secretary Cliff Spare 10 Olive Lee St Quakers Hill 2763 9626 0300 [email protected]
Ass't Show Sec. Ross Selig 617 East Kurrajong Rd East Kurrajong 2758 4576 7093 [email protected]
Ass't Show Sec. Jeff Mansfield 59 Broughton St Strathfield 2135 9861 0393 [email protected]
Ass't Show Sec. Mark Valvason 130 Chapel Lane Baulkham Hills 2153 9862 6644 [email protected]
Ass't Show Sec. Harry Charalambous As above
Ass't Show Sec. Paul Brett 3 Spring Rd Kellyville 2155 9629 3210 [email protected]
Ass't Show Sec. Peter Kerr 7 A Laurel Close Hornsby 2077 9940 3385 [email protected]
Publicity Officer Ray Galbraith 5 Gardiner Rd Galston 2159 9653 1538 [email protected]
Editor Santo Calabrese 65 Tallowwood Ave Cherrybrook 2126 9875 2647 [email protected]
Ring Registrar Graeme Gordon 35 Watkins Rd Baulkham Hills 2153 9639 9603 [email protected]
Events CoOrd. Craig Buckingham As above
Librarian Paul Brett As above
Catering Officer Jo Zammit 7 Grevillea Rd St Clair 2759 9834 6541 [email protected]
M'gmnt. Delegate Craig Buckingham As above
M'gmnt. Delegate Stuart Williams 7 Kerry Rd Schofields 2762 9627 1415 [email protected]
Alt. Delegate Ray Galbraith As above
Trophy Steward Brian Findlay 14 Brabyn St Windsor 2756 4577 9535 [email protected]
Auditor Harry Charalambous As above
Auditor Helen Charalambous PO Box 990 Wahroonga 2076
New Member CoOrd. Ross Selig As above
New Member CoOrd. Lloyd Mullens 23 Flinders Rd North Ryde 2113 9802 0780
New Member CoOrd. Kevin Nesbitt 103 Blackwell Ave St Clair 2759 9834 1587 [email protected]
M'Ship Registrar Peter Dodd As above
Webmaster Daniel Child 11/105 Hammers Rd Northmead 2152 O410628691 [email protected]
Floor Member Andre Ozoux 79 Morrison Rd Gladesville 2111 9816 5914 [email protected]
Floor Member Ken Mitchell 22 Salter Court Harrington Park 2567 4647 4444 [email protected]
Floor Member Dave Zammit 7 Grevillea Rd St Clair 2759 9834 6541 [email protected]
Floor Member Tommy Hancock 71 Pitt St Richmond 2753 4578 1723 [email protected]
Returning Officer Paul Brett As above
Trustee Craig Buckingham As above
Trustee Daniel Child As above
Services Manager Stuart Williams As above

From The Editor
The last month has been quite a memorable one for the Hills Branch, with Hills Branch coming a very close second at the State Team Challenge after winning the previous 3 years in a row. Our cover this month is the National Greygreen winner from Hills member the Welshman family. Congratulations to all Hills members who did well at both the State Team Challenge and the Nationals. National results link is here – http://www.budgerigar.com.au/nationals/2012-National-Results.pdf The Hills Auction will be held on the 7th July 2012 and members will know how important it is in maintaining the Branches finances. Please be aware that we will require a strong turnout on the day to make it a success. Your Committee has planned as well as it can but we will need runners, clerical staff and a lot of bodies on the day, especially as the auction comes to the end. Please come to Friday night’s meeting and volunteer for the day. Hills member and veterinarian Phil Ashby will be presenting a talk on Genetics in Poultry and its relevance to budgerigar breeding. Phil is currently involved in the Poultry industry but it’s amazing how relevant to the budgerigar fancy poultry animal husbandry is.
Annual membership is due 1
st July! Don’t become
unfinancial. Membership renewals can be done at our meeting on Friday!

What's On in 2012? There are the events of the coming 12 months or so. Hills Branch Monthly Meetings are the 4th Friday of the month. ________________________________________
June
24th - Canary & Cage Bird / BRASEA Variety Show 24th - BIS Annual Show - 24th - Gulgong & District Avicultural Society Inc. Annual Show
July
1st - Mid North Coast Branch Annual Show - 7th - Hills Branch Auction 8th - Hunter Branch Annual Show - 9th - BSNSW Management Meeting, 8.00pm, Blacktown RSL Club, Second Ave, Blacktown. 14th - "Invitational Lawn Show' - Northside, Penrith Valley, West Sydney & Pied Society 21st - Mid State Annual Show - 29th - New England Branch Annual Show
August
1st - Closing date for Budgerigar Articles. 5th - West Sydney Branch Auction (TBC) 11th - Newcastle BS Auction 12th - Newcastle BS YBS 19th - St George Auction 26th - Canary & Cage Bird Federation - Annual Dinner
September
9th - Tamworth Budgerigar Society YBS 1oth - BSNSW Management Meeting, 8.00pm, Blacktown RSL Club, Second Ave, Blacktown. 16th - Northside Branch Annual Show
October
1st - Closing date for Budgerigar Articles. 14th - BRASEA Auction 21st - Shoalhaven Branch - Southern Cup YBS (TBC) 21st - New England Branch YBS 28th - St.George/Illawarra Branch YBS (TBC)
November
4th - Central Coast Branch YBS 4th - Canberra Branch YBS (TBC) 11th - Penrith Valley Branch Annual Show 12th - BSNSW Management Meeting, 8.00pm, Blacktown RSL Club, Second Ave, Blacktown. 25th - Sydney Annual Lawn Show

AVIAN NUTRITION
By Dr Ron Woodhead
Many Avian Vets are of the opinion Chronic Generalised Malnutrition is a primary underlying
factor of disease in the majority of birds presented to their Clinical Practice.
The physical appearance of a bird suspected of suffering chronic deficiencies of several
nutrients are as follows:
* Lackluster
* Loose Feathering
* Breakdown of interlocking barbules
* Stress lines on feathers
* Debris on microscopic examination
* Depigmentation of feathers
* Chronic heavy pinfeathering
* Easily broken wing and tail feathers
* Epithelial surfaces dry & flaky
* Beak & Nails long & rough
* Layering of beak
* Skin on legs & feet thickened & dull
* Obesity
* Poor Breeding Potential
* Liver Disease
* Pancreatitis
* Kidney Disease
* Diabetes
* Visceral Gout
The Nutritional requirements for aviary birds are poorly understood. Each species of bird having
its own unique nutritional requirement. They satisfy these requirements in nature by selecting a
wide variety of foods from their natural environment.
For good health and reproductive performance birds must be provided with the correct balance
of amino acids, fatty acids, cho’s, vitamins, minerals and water.
FACTORS INFLUENCING AVIAN MALNUTRITION
1. Food selection- the consumption of individual items in a mixed diet is extremely variable
often resulting in a disproportionate consumption of certain foods.
2. Type of food provided- inappropriate or not eaten.
3. Species of bird.
4. Changing requirements of birds.
Caged birds/ Aviary birds/ Breeding, Growth, Racing, Molting

FOODS COMMONLY FED TO BIRDS INCLUDE:
Seed
Dry Seed- Seeds are low in proteins, low in copper, zn, manganese, gross imbalance in Ca &
Pho’s. Usually high in P which inhibits the absorption of Ca.
Deficient in minerals – Iodine. The mineral composition is directly related to the soil quality
and type where the crop is grown. Sunflower, rape, linseed, niger are rich in fat –50%, cause
obesity and form insoluble soaps with Ca & Fe inhibiting their availability to the bird. Canary,
panicum, millet and hulled oats are rich in C.H.O’s.
Seeds are deficient in vitamins- Vit A, B5, B12, Biotin, Low in Riboflavin, niacin, pantothenic
acid & Vit K. Vit E Uncertain.
Seed is an excellent source of energy but only plays a limited role in a balanced diet.
Soaked seed:
No additional nutrient value, more easily digested in young birds.
Sprouted seed:
More protein
Less fat
Breeding
Seeding Grass:
Seed heads are rich in vitamins & minerals such as Fe & Ca.
Green food
High water content
High in vitamins/ minerals/ fibre.
Fibre maintains intestinal muscle tone and the natural balance of intestinal flora. Include
fruits/ vegetables/ branches of native plants.
Mineral Supplements:
Cuttle fish, shell grit = Ca
Charcoal = Iodine
Rock Salt = Sodium
Soluble grit
Insoluble grit (granite) = Gizzard
(Coarse sand)
Lorikeet & Honeyeaters lack a muscular gizzard, can’t survive on seed.
Water
Fresh, cool, clean filtration, black poly pipe.

Malnutrition in Aviculture manifests as
1. Specific deficiency disease.
2. I increased susceptibility to disease
3. Decreased response to therapy of other diseases
4. Obesity
5. Starvation
6. Decreased breeding performance
7. Poor growth
Specific deficiency diseases:-
1. Vit D3 = Ca/P – Ultra Violet light. Pets in cages & shaded aviaries.
2. Vit A- Mucous membranes, epithelial health, bumble foot.
3. Vitamin E- Cockatiel paralysis
4. Riboflavin- curled toe paralysis
5. Goiter- iodine = cabbage= gottragen
6. Gout- High protein
Low Vitamin A
7. Ca/P ratio- Rickets, young birds
8. Lysine- loss of green pigment in feathers= yellow
9. Toxicity – Avocado- Love birds
Obesity = fatty liver
Milk sap plants
How to minimise nutritional disorders:
1. Feed correct seed type for species
2. Minimal oil seeds
3. Wide selection on foods- observe what is eaten fresh
4. Vitamin/ mineral/ amino acids- in feed- on soft food- in water
5. Protein supplement for breeding & rearing young.
Dr Ron Woodhead

The Blackface: a New Mutation in the Budgerigar By: H.W.J. v.d. Linden (Judge)
Translated by: Inte Onsman, Research coordinator MUTAVI
A new mutation can be added to the long list of existing mutations in the Budgerigar: the blackface. According to the phenotype of this mutant, this name seems to be the most appropriate. Therefore the name blackface is proposed. Where and when this mutation originally was derived cannot be ascertained, but it is almost for sure that it was in the Netherlands. In any way, the blackface discussed here was discovered in The Netherlands.
The facts: Mr. Van Dijk, a Budgerigar breeder from the Netherlands, discovered two blue cocks having black undulated masks and black striped markings at their abdomen, amongst a wide range of varieties at a bird-market. He decided to buy the birds to breed with them and find out if the trait was a modification or inheritable. In the first year the cocks were mated to normal Australian grey hens. Out of these matings normal greys and sky blues were derived, cocks as well as hens. During the second year Mr. Van Dijk mated the young hens back to the original cocks. Only one mating - original blackface cock x F1 (grey) hen - produced offspring from which a few were blackface. After the second breeding season it was almost certain that the blackface mutation resides on an autosome and is recessive to the wild type. The testmatings Mr. Van Dijk carried out so far, confirm this point of view. At this moment there are no indications that the mutation is linked to any other known mutation in the Budgerigar. Mr. Van Dijk observed a much darker body colour as opposed to normal birds split blackface [2]. A blue blackface without dark factor is showing a considerable darker blue colour as is seen in normal cobalt blue Budgerigars. In grey blackface specimens without dark factor, a comparable darkening of the body colour is observed as opposed to normal grey birds. In green blackfaces the same phenomenon has been observed. By the way, all green blackfaces died. Only a few pictures are left. At this moment Mr. Van Dijk has one cinnamon-blackface, naturally with a brown mask and ditto striped markings on the abdomen. It was told by Mr. Van Dijk that the first two cocks bought in 1992 must have had a common recessive pied ancestor, because several recessive pieds were found in the offspring. In the past five years several youngsters were bred with a small head spot suggesting they are split for recessive pied. Breeding blackfaces is, according to Van Dijk, not as easy as one would like [2].
For this new mutation the symbol bf is proposed, derived from the English name given to this mutation. The wild type symbol is bf +.
Possible matings:
1. blackface x blackface = 100% blackface
2. blackface x normal/blackface = 50% normals/blackface and 50% blackface
3. blackface x normal = 100% normal/blackface
4. normal/blackface x normal/blackface = 25% normals, 50% norm./blackface and 25% blackface
5. normal/blackface x normal = 50% normal and 50% normal/blackface
Matings 4 and 5 are not recommended because one cannot see which youngsters are split blackface and which are not.

Expectations: Breeding blackfaces is, according to Van Dijk, not very easy. His stock contains about twelve blackfaces and some split blackfaces and "maybe" splits. It will take some time for this mutation to establish and will become available to a wider range of breeders. That is understandable, but on the other hand risky because if a fatal disease affects this stock or another disaster takes place, there is a chance the mutation will disappear again. On the long run, when sufficient birds are available, one could possibly achieve a smooth colour of the "mask" and body, and what to think of a fully coloured black belly-spot? If this is indeed a possibility we will have to wait to see what happens in the future.
Note from the MUTAVI Research and Advice Group: This mutation could be a mutation of the MC1R-locus which is not fully understood nor described in aviculture. Therefore it is recommended to carry out further test matings. It is obviously a pigment distribution factor resulting in some kind of melanism [1,3,4]. We have not seen these birds alive and our latest information is that they are extinct.
Literature:
[1] Bruckner J.H., (1939) The Inheritance of Melanism in Pheasants Journal of Heredity Vol.30; pp.45-52 [2] Dijk v., (1998) Personal communication [3] Moore J.W., Smyth J.R., (1971) Melanotic: Key to a Phenotypic Enigma in the Fowl Journ.of Heredity Vol.62; 215-219 [4] Sage B.L., (1962) Albinism and Melanism in Birds British Birds Vol.55 no.6; pp.201-225
©Inte Onsman
MUTAVI Research & Advice Group

The Greywing by Ghalib Al-Nasser
Beauty is in the eye of the beholder". This applies to any Budgerigar variety and without a doubt the Greywing variety has got that certain appeal to many Budgerigar breeders. As a breeder of this variety I found the challenge to breed a good specimen as rewarding as breeding a good Light Green or a Skyblue.
The first Greywings were established in around 1919 in Europe and were of the green series. They were initially called Apple Green or Jade. But when the Greywing of the blue series was established, initially in Austria in 1927 and then afterwards in the UK in 1928, the Colour Committee of the Budgerigar Society grouped them together and gave them the name "Greywings" in 1929.
It is easy to recognise the variety when viewing it in daylight by the grey marking on the bird in comparison to that of the black on the normal varieties. By comparing a Greywing Light Green to a normal Light Green the spots, undulations at the back of the head, markings on the wings and primary wing flights are all of a grey colour instead of black in the normal varieties. The cheek patches are of light violet and the body colour is diluted by 50% of the normal body colour. Also the colour of the tail is grey with a bluish tinge instead of dark blue in the normal varieties.
At times fanciers find it difficult to distinguish between the Dilutes (Yellows and Whites) and the Greywings, especially if both varieties are of the grey factor (Greys and Grey Greens). Here a closer look at the body colour and wing marking of the Grey Yellow and Grey White when compared to Greywing Grey Green and Greywing Grey is that the body colour of the former is that much further diluted than the latter. The tail feathers of the Dilutes are off white to pale grey while those of the Greywing Grey and Grey Green are grey. Without a doubt Greywings are more appealing on dark factor birds such as Dark Greens and Cobalts.
It was found that a recessive gene controls the Greywing when mated to normal varieties. So when a Greywing is mated to a normal all the young will be normal but carrying the Greywing gene in a hidden (split) form; Normal/Greywing. If one of these splits is mated back to a Greywing then&nbp;50% of the young will be split for Greywing and the other 50% will be visual Greywings. Mating two Greywings together will result in all the young being Greywings. Because of the recessive gene it does not matter much which sex is the visual Greywing as both cocks and hens can be mated and produced according to the above three types of mating. Also because of that recessive gene, a bird can be carrying the Greywing factor in a hidden form for many generations.
This is why on occasions a Greywing can pop up in a nest of normal parents in a stud that does not carry this mutation and the fancier is at a loss as to how the Greywing appeared. Following the Mendel Theory of Inheritance, pairing two normals that are split for Greywing will result in 25% of the young being Greywings. Another 50% will be normal/Greywing and the remainder 25% being pure Normal. Suddenly the Greywing factor has been established in this stud.
Early breeders established, using Laws of Genetics, that although the variety is recessive to the normal, it is dominant to the Dilutes (Yellows and Whites), the same as the Clearwings (Yellow-wings and White wings) are dominant to the Dilutes, even though all the three varieties are recessive. In fact all the three varieties are separate mutations of the same gene and form what is known as a multiple allelomorph.
If a Greywing is paired to a Dilute all the chicks will be Greywings but split for Dilutes. The same will apply if a Clearwing is paired to a Dilute. But something quite unusual happens when pairing a Greywing to a Clearwing. Here the resulting young will possess the combination of both varieties in one bird and are called "full bodied colour Greywings". This means that they have the full body colour of the Clearwing variety with the Greywing marking; quite an unusual and beautiful coloration. These full-bodied colour Greywings are split for Clearwing but cannot be split for Dilutes.
If a full-bodied colour Greywing is paired to a Dilute then the results will be 50% Greywings and 50% Clearwings all split for Dilutes. However, if a full bodied colour Greywing is paired to either a Greywing or a Clearwing then the results will be 50% full bodied colour Greywings and 50% Greywings (or Clearwings).

The Involvement of Recessive Pied in the Origin of the Dark Eyed Clears in the Budgerigar
By: Inte Onsman, Research coordinator
MUTAVI Research & Advice Group, The Netherlands
Recessive pied The first pied mutation in Budgerigars was established in 1932. The mutation appeared to be recessive and was called Danish pied. These birds were also referred to as Finnish pieds for a short time because the first species who reached England came from Finland [17]. The very first mutant was discovered in a mixed coloured flock of Budgerigars breeding as a colony. Therefore it was not possible to find out from which parents this bird was derived. One of the first aviculturists who understood that this actually was an important new mutation was C. af Enehjelm, head of the Helsinki zoo in Finland. Thanks to his interest and dedication this mutation was quickly distributed through the whole of Europe. During World war II Danish pieds almost died out but the Danish fancier Walter Langberg saved the mutation from total extinction.
Dutch pied The source of the first Dutch pieds cannot be established for sure. In "The Handbook for Budgerigar Breeders" written by W.Beckmann (1966) is written that Mr. Van Dijk, head of "Animali" zoo in Eindhoven (The Netherlands) was already breeding Dutch pieds before World war II. Also foreign sources mention the year 1939 to be the year in which the first Dutch pieds appeared and even in America the mutation is known as "Dutch pied". Many authors consider the clear flight to be a selective form of Dutch pied and both in England and America these clear flights are known as "continental clear flights". The first specimens were developed by R. Raemaker from Bruxelles who claimed it took him several years to breed the first clear flights from birds who initially had only a few unpigmented feathers.
During the late Forties and the early Fifties, people from Europe mentioned black-eyed "albinos" and "lutinos". During several years it was presumed that these birds could only be bred by cross breeding them, but later on evidence was found that this was not true. Experimental matings with these birds eventually cleared the matter. They were in fact a combination of recessive pied and continental clear flights (a selective form of Dutch pied). These birds also did not have black eyes but the plum eyes of the recessive pieds. Yet they were known as dark eyed clears. Unfortunately Dutch pieds never became very popular in the Netherlands unlike America and Germany where some fanciers breed them with great enthusiasm [22,23,24]. There are some remarkable similarities between Dutch pieds and recessive pieds to be found in literature. If a DF Dutch pied is mated to a normal, one expects 100% Dutch pieds. However, some authors report the appearance of a single normal youngster among the offspring of such mating (...). Male Dutch pieds show more pied feathers than female Dutch pieds. This is also the case in Australian pieds and recessive Danish pieds. In the USA and in England one distinguishes Dutch pieds from continental clear flights. Some people state that continental clear flights could be a selective form of Dutch pied and having seen my own results with Dutch pieds I am willing to believe that this is actually the case. The main question is why the combination Danish- / Dutch pied produces a dark eyed clear bird and the combination Danish / Australian does not. To find the answer to this question, we have to return to the early embryonical state and the source where melanoblasts, the precursors of melanocytes, are produced.

The research The origin of pigmentation in vertebrates has always been a favourite subject of scientists all over the world [7,9,10,11,15]. The first authors were very much impressed by the star shaped cells containing melanin they found in the skin. Already in 1860 Kölliker described migration of pigment cells in the skin which were derived from mesenchyme (brain) tissue. This point of view was already confirmed by Kerbert in 1877 for reptiles and birds, again for humans and birds by Riehl in 1885 and several mammals and again birds by Ehrmann. In 1890 Haecker studied pigmentation in bird feathers and came to the conclusion that feather pigmentation is caused by migrating pigment cells derived from mesenchyme sources. (The mesenchyme is a network made of embryonic tissue from which in a later state the ordinary tissues, blood and lymph vessels are derived.) Soon it became clear that the source of pigment cells could not be found under the microscope because melanoblasts (pigment cell precursors) are not to be distinguished from other cells. Experimental transplants involving amphibians showed notable different conclusions, for pigment cells do not derive from mesenchyme tissue, or as some people stated, in the skin itself, but derive from the neural crest. In 1935 this was definitely confirmed by Dushane [3] who did research on transplanted parts of the neural crest from Ambystoma (a Mexican salamander). In the fowl this problem was first investigated in 1936 and 1939 by Dorris [2] and also in 1941 by Ris [18]. By studying cultivated tissues and transplantation experiments, they concluded that the neural crest was able to produce pigment cells and that the mesoderm as well as the skin were not able to do so.
The neural crest The neural crest in fowl embryos was first described by His in 1868. In a very early stage the neural walls are formed. These neural walls rise and close in order to form the neural tube, the precursor of the central nerve system and the spinal cord. The neural crest is situated on the backside of the neural tube and immediately starts dispersing celprecursors. Along both sides of the neural tube somites are to be found. Somites are blockshaped embryonic tissue elements which turn into a dermatome from which skin segments are formed. All skin segments together eventually will form the entire skin.
Pied genes Genes causing nonuniform pigmentation patterns have been found and investigated in many vertebrates. In mice more than fifteen different pied mutations are known, some of them are even multiple allelic. Pied genes are able to manifest themselves in different ways as many research projects that have been done over the past years have shown. Deol investigated the role of the tissue environment in the expression of pied genes in mice [1] and found that some pied genes affect melanoblasts (pigment cell precursors). Others affect the tissue environment in which the melanoblasts reside. During this research particularly interest was taken in the eye pigmentation of several piebald mutations. Some of these mutations affect eye pigmentation and others do not.
In the fowl the migratory behaviour of neural crest cells was studied extensively by Noden [14]. He found that the migratory behaviour of cells derived from the neural crest, is influenced dramatically by certain interactions with their environment. All neural crest cells, with regard to which part they came from, do possess the ability to recognize specific genetical signals, react on them and so determine the direction of their migration. In 1982 Hirobe published a paper in which he described investigations on the regulation of melanoblast / melanocyte populations in the skin of newborn mice [5]. He came to the conclusion that the distribution of melanoblasts / melanocytes into the skin, is

under control of a number of (up to now) unknown semi dominant genes. In 1992 he publishes the results of a follow-up where he investigated the proliferation and differentiation of melanocytes [6] and announced research on the number of genes involved.
Eye pigmentation The research of Deol inspired me some years ago to compare eye pigmentation from Danish- and Australian pieds using a light microscope. The following items are important to know in this respect. Melanocytes or pigment cells derive from three different sources: 1) The neural crest from where they migrate to all parts of the body including the choroid and the outer layer of the iris of the eye. 2) The outer ridge of the optic cup which eventually will form the pigment epithelium of the retina. 3) The neural tube from where neuromelanins are formed who mainly are to be found in brain tissues.
Summarized the melanocytes of the eye have two sources. To be precise, the melanocytes of the choroid and the iris stroma (frontside of the iris) are derived from the neural crest, the melanocytes of the pigment epithelium (retina) and the inner (back) side of the iris are derived from the edge of the optic cup. The optic cup is the precursor state of the eye, the definite eyeball and lens are yet to be formed. The pigment epithelium exists of honeycomb shaped highly specialized melanocytes, residing between the retina and the choroid. The melanocytes of the choroid reside amongst the blood vessels of this layer which is situated between the pigment epithelium and the scleral cartilage (outer layer of the eye in birds). The difference between eye melanocytes and epidermal (skin) melanocytes is that the melanocytes of the eye do not disperse their pigment granules into neighbouring tissues unlike skin melanocytes. The results of my investigations showed that the eyes of Danish pieds do have unpigmented areas in the choroid, and the outer layer (frontside) of the iris almost totally lacks pigment cells. The retinal pigment epithelium and the inner layer (backside) of the iris are unaffected. The eyes of Australian pieds were completely unaffected and eyes from Dutch pieds have not yet been investigated but are presumed to be unaffected as well.
Conclusion: Having seen these results I came to the conclusion that in Danish pieds, exclusively melanocytes derived from the neural crest are affected by the s-locus. This suggest a possible defect of the neural crest itself causing a disturbed production of too few, abnormal and also normal pigment cell precursors. Obviously the s-locus (Danish pied) acts in the neural crest and not in the skin. In Australian pieds, the Pb-locus could act in the former mentioned dermatomes or in certain skin areas in which as a result melanoblasts are unable to differentiate into melanocytes leaving those areas unpigmented. The Dutch pied gene (Pi-locus) could affect e.g. the proliferative capacities of melanoblasts because often wingtops, being most far apart from the neural crest, show pied areas. It could also be possible that the Dutch pied locus is involved in determining the direction of the migrating melanoblasts. The real dark eyed clears (or should we say plum-eyed) can only be achieved by the interaction of Danish- and Dutch pied. Apparently these two loci are playing a major role in pigment distribution in Budgerigars. The combination Australian- / Dutch pied will never develop a dark eyed clear phenotype that is quite obvious. The mottle is deliberately disregarded in this article, because the cause of the mottled phenotype differs significant from the other pied mutations. Breeding Dutch pieds should be recommended strongly. They are very attractive in appearance, have bright colours just as Danish pieds and body size is easy to improve.

Rare Budgerigar Varieties, The Clearbody by Ghalib Al-Nasser
Since my early days in the Budgerigar fancy (early seventies), I had heard of the American mutation, the Clearbody. The late Cein Roberts of Lancing, Sussex had seen the variety and was in the process of importing some to the U.K. but that process never materialised. His nearest description to the variety was a futuristic artist's impression by R A Vowles, which appeared in Dr M D S. Armour's book Exhibition Budgerigars. Dr Armour predicted the appearance of a variety with a clear body, normal wing markings and black tail and referred to it as a Laced Yellow.
The Clearbody originated in America in the early 1950's. Initially there were three separate types of Clearbodies but one (Terraneo Clearbody) seems to have disappeared. The other two are the Texas Clearbody, because it originated in the State of Texas, and the Easley Clearbody, named after C F Easley from the State of California who established the variety. The two varieties differ in both appearance and breeding pattern.
While I was in Australia in 1994, on my lecturing and judging tour with the late Mick Wheeler, I came across the Australian version of the Clearbody in Sydney, which they say has been in existence since the early fifties. Here is an example of how a mutation has the ability to appear in different parts of the World at about the same time. The Australian version was referred to as a Greywing Dilute. I found this definition difficult to accept as the Greywing variety is dominant to the Dilute (Yellows and Whites) even though both varieties are recessive and therefore a Greywing can not be masking a Dilute but can be split for it.
The Texas Clearbody is more common and was first imported to the UK in 1989 by Jeff Attwood and then to Europe. The Budgerigar Society provided a provisional colour standard for this variety in 1997. The general appearance of this variety in green and blue is similar to normal and but with the following differences. The flight feathers are pale grey instead of black, the body colour is suffused and may vary in intensity from minimum through to almost 50% of normal body colour depth and increase in intensity downwards and towards the rump area.
The Texas Clearbody is sex-linked recessive in its breeding inheritance but has an unusual relationship when paired to Ino (Lutino and Albino) as it was found to act as dominant. Because of this relationship an Ino cannot be masking a Clearbody as it can with other varieties and a normal cannot be split for both Clearbody and Ino, yet a Clearbody can be split for Ino.
Because of this relationship there are a number of matings that can produce a Clearbody depending on the partner used. The table below outlines this permutation. The term "Clearbody" used in this table refers to the Texas sex-linked variety for simplicity:
No. Pairing Expectation
1 Ino cock × Clearbody hen 50% Clearbody/Ino cocks 50% Ino hens
2 Clearbody cock × Ino hen 50% Clearbody/Ino cocks 50% Clearbody hens
3 Clearbody/Ino cock × Ino hen
25% Clearbody/Ino cocks 25% Ino cocks 25% Clearbody hens 25% Ino hens

4 Clearbody/Ino cock × Clearbody hen
25% Clearbody cocks 25% Clearbody/Ino cocks 25% Clearbody hens 25% Ino hens
5 Normal/Clearbody cock × Ino hen
25% Normal/Clearbody cocks 25% Normal/Ino cocks 25% Clearbody hens 25% Normal hens
6 Clearbody cock × Normal hen 50% Normal/Clearbody cocks 50% Clearbody hens
7 Clearbody/Ino cock × Normal hen
25% Normal/Clearbody cocks 25% Normal/Ino cocks 25% Clearbody hens 25% Ino hens
8 Normal/Clearbody cock × Clearbody hen
25% Normal/Clearbody cocks 25% Clearbody cocks 25% Clearbody hens 25% Normal hens
9 Normal/Clearbody cock × Normal hen
25% Normal/Clearbody cocks 25% Normal cocks 25% Clearbody hens 25% Normal hens
10 Normal cock × Clearbody hen 50% Normal/Clearbody cocks 50% Normal hens
11 Clearbody cock × Clearbody hen 50% Clearbody cocks 50% Clearbody hens
It is worth knowing that all the Normals and split cocks for either Clearbody or Ino are indistinguishable and only by test mating can their identity be revealed. The use of the Ino in the above matings lightens the body colour, while the Normals add extra size and head features desirable for a show specimen.
I was delighted to have met with Tom Easley at the 1998 All American Show in San Diego and had quite a long chat with him about the variety, which was established by his father over forty years ago. Mr Easley did inform me that the present Easley Clearbody did not resemble the mutation his father established. The present Easley Clearbody (referred to sometimes as Laced Clearbody) generally has jet black wing markings, flight feathers, tail feathers and mask spots because of the excess black melanin. Their cheek patches are a shade of grey/silver regardless of their body colour. The body colour also tends to have less suffusion as found with the Texas mutation. However, Tom Easley did say that the cheek patches of the original stock were pale violet.
Ken Gray published an article about the Easley Clearbody in a recent publication of the Rare Variety & Colour BS magazine. The article being based on a letter sent by the late C F Easley to the late Cyril Rogers, which gave full details of this variety. Mr Easley described the first so-called Easley Clearbody, which appeared in his aviary in January 1954 as an Opaline Greywing Dark Green hen with a yellow body colour. The parents of that bird were an Opaline Dark Green cock and a Cobalt hen. When this hen was paired the following year to a normal Dark Green cock two more of this new mutation appeared, both cocks, a normal and an opaline. Both fitted the above description of the Easley Clearbody but with pale violet cheek patches.
Mr Easley also established that the variety is dominant in its breeding pattern and can be produced in both sexes from any mating of which one partner must be visual. Because of the dominant gene, the variety can be established in both a single and double factor. The double factor version tends to have a much diluted body colour. Although I have seen examples of this variety in both America and Europe, to date the variety has not been imported to the UK.

MINUTES OF THE MONTHLY MEETING HILLS BRANCH - B.S.N.S.W. (Inc.)
Meeting held at Don Moore Community Centre on Friday 25th May 2012
MEETING OPENED: 8.20pm Harry Charalambous presided. PRESENT: As per attendance sheet. NEW MEMBERS: Tione Plowman and Kevin French. Welcome tonight to new members Tione and Kevin, and to James Matthews who joined last month. VISITORS: Craig Sutherland, Abi Haider, and John Hogarth APOLOGIES: Peter Dodd, Brian Findlay, Craig Buckingham, Andre Ozoux, Richard Abraham and Stuart Williams Moved that the apologies be accepted. Moved: M. Hill Seconded: L. Mullins PREVIOUS MINUTES: Moved that minutes are a true and accurate record. Moved: T. Hancock Seconded: K. Nesbitt BUSINESS ARISING FROM PREVIOUS MINUTES: STCC. Thank you to everyone who made their birds available, and congratulations to those who had birds selected for Hills, and to those who had birds picked for the NSW Team. TREASURER’S REPORT: All accounts approved at previous General / Committee meetings have been paid.
Income from last General / Committee meeting: Nil
Accounts paid since last General / Committee meeting: - BSNSW (STCC entry fee) $ 100.00 Accounts received for payment tonight:
- S. Calabrese $ 41.70 Moved that accounts be paid and Treasurer’s Report be accepted. Moved: K. Nesbittt Seconded: R. Galbraith CORRESPONDENCE IN:
Hunter Branch Schedules
BIS Show schedules
BRASEA Auction catalogues
Kelwyn Kakoschke auction catalogue
Email correspondences via the website
CORRESPONDENCE OUT:
Reply emails to website queries Moved that the correspondence be accepted Moved: V. Tarpis Seconded: I. Manton GENERAL BUSINESS:
State Team Challenge. Kathy gave a report on the STCC and STS.
Awards Night- where it could be held, what night it best be
Distribution of the 4 x ANBC Standards that were allocated to Hills Moved that we take this matter to the Committee for a decision Moved: R. Galbraith Seconded: P. Brett
Website – need updated profile for the Mantons. Request for bird photos, etc.
Look at our strength and weaknesses with varieties and the possibility of purchasing birds to improve the Branches weak varieties
Bird swapping / mentoring / ring swapping
Lloyd has 60 gold rings available
Kakoschke auction on Sunday. Need help on Saturday afternoon to set up.
MEETING CLOSED: 9.45 pm HARRY CHARALAMBOUS KATHY MANTON VICE CHAIRMAN SECRETARY (ACTING CHAIRMAN)

MINUTES OF THE COMMITTEE MEETING HILLS BRANCH - B.S.N.S.W. (Inc.) Meeting held at Dural Country Club, Dural on Tuesday 12th Jun 2012. MEETING OPENED: 8.01 pm Peter Dodd presided. PRESENT: Peter Dodd (Chairman), Stuart Williams, Kathy Manton, Ra Galbraith, Paul Brett, Harry Charalambous, Santo Calabrese, Richard Abraham, Graeme Gordon, Craig Buckingham, Ian Manton and Ray Galbraith. APOLOGIES: Tom Hancock an, Andre Ozoux Moved that the apologies be accepted. Moved: P. Brett Seconded: C. Spare PREVIOUS MINUTES: Moved that the minutes of the previous meeting as printed in the Newsletter are a true and accurate record. Moved: H. Charalambous Seconded: S. Williams BUSINESS ARISING FROM PREVIOUS MINUTES: Purchase of birds – hold over to General Business TREASURER’S REPORT: All accounts approved at previous meeting have been paid. Income from last General / Committee meeting: Raffle of Show Cage $70.00 Reimbursement of gas bottle refill-BS Annual $34.95 Membership $50.00 Accounts paid since last General / Committee meeting: Westpac Merchant fee $ 24.00 North Rocks Community Centre $ 80.60 Balance of Accounts as at 11.06.12 • Cheque Account: $19,701.29 • Petty Cash: $ 239.85 Accounts received for payment: Bird Boxes Galore $190.00 (incl seed for STCC, seed for Auction, Table Show certificates & Annual Show) Moved that the Treasurer’s report be accepted and accounts be paid – Moved: R. Galbraith Seconded: C. Spare CORRESPONDENCE OUT: Nil CORRESPONDENCE IN: • 10 email requests for Auction catalogue Moved that the Correspondence be accepted- Moved: C. Buckingham Seconded: G. Gordon GENERAL BUSINESS Hills Auction
Daniel to update Catalogue Stickers – Kathy Santo – update “how to” form Staging – Jeff BBQ food – Ray BBQ- Teanna + Michelle Phone bidders- Craig, Harry, Santo, Stuart Absent bidders form Registration forms Airport pickups – Harry Projector – check at North Rocks? Security – Ross?

Runners – John Halas? Set up 7.00am sharp Biscuits for morning tea – Ray Milk – pick up on the day Show Cages – (need 81) Paul x 12, Craig x 10, Peter x 24, Manton x 30 – still need more EFTPOS has been ordered Robert Mead – AV Gear Monitor – Kathy Clerical – Cliff sales tickets, Graeme, Tom & Ross – registrations Sticking birds – Paul Interstate birds – Craig, Peter
AGM – Peter & Cliff not re-standing Sale Night – members birds only Newsletter – Memberships now due 4 x allocation of ANBC Standards – keep one copy in the Library Moved that the remaining 3 Standards be drawn from a hat Moved: C. Spare Seconded: I. Manton Purchasing of Variety Birds – for Hills Club. Further discussions included: Need Greywings, Clearbodies, Black Eyed Self, Fallows, and Normals / Cinnamonwings. Initially have two senior breeders per variety with possibly mentoring involved also. Suggest that one or two Open breeders be involved in the initial purchases, within a specified budget, by end of 2012. June Meeting: Lecture by Phil Ashby, Richard Abraham to judge. BSNSW AGM – Complete a postal vote if unable to attend the AGM Presentation / Awards Night – Saturday 25th August St George Maronite Catholic Church Hall at Thornleigh - $350 hall hire – penciled in at this stage. Catering / menu options approximately $25 - $30 per head. To be considered by the members at the next Branch meeting. Moved that Club pay the hall hire, members to pay for meals. BYO drinks Moved: C. Spare Seconded: S. Calabrese Aviary Tour – Dubbo area – possible aviaries include: John Scoble, John Boss, Mark Hawke, Wayne Cusak and Terry O’Brien. MEETING CLOSED: 10.36 PM. PETER DODD KATHY MANTON CHAIRMAN SECRETARY
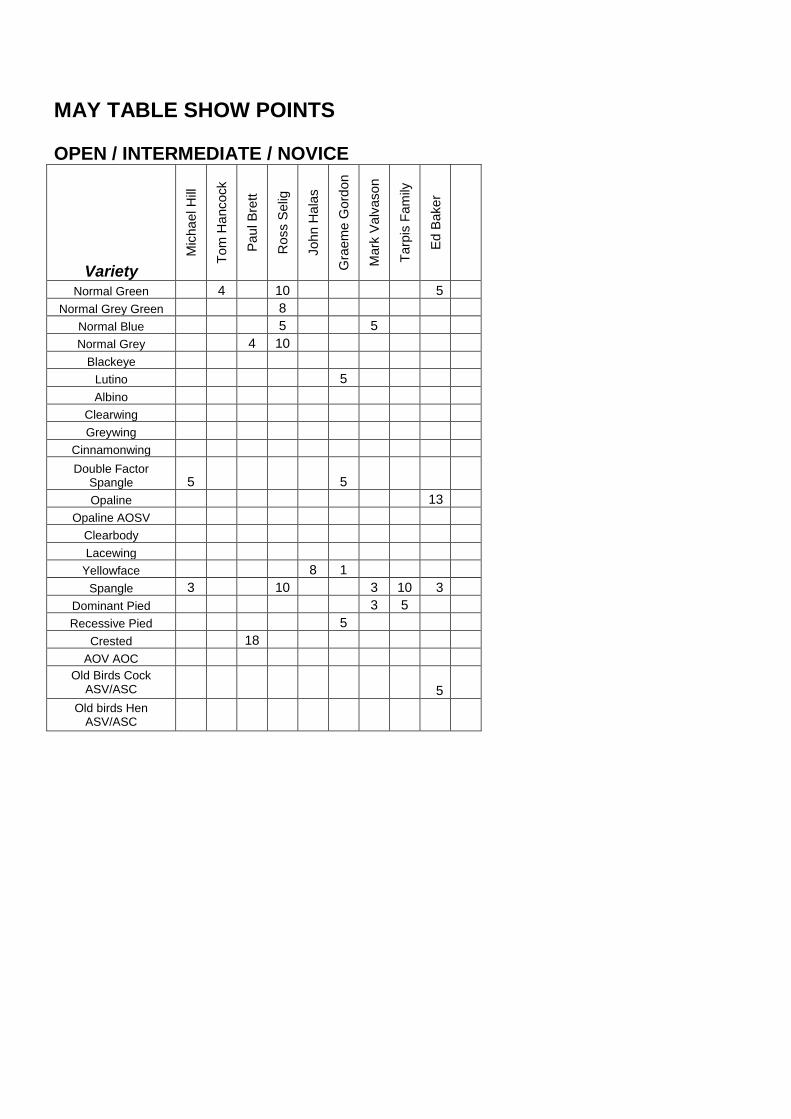
MAY TABLE SHOW POINTS
OPEN / INTERMEDIATE / NOVICE
Variety
Mic
ha
el H
ill
Tom
Ha
ncock
Pau
l B
rett
Ross S
elig
John H
ala
s
Gra
em
e G
ord
on
Mark
Va
lvason
Tarp
is F
am
ily
Ed B
aker
Normal Green 4 10 5
Normal Grey Green 8
Normal Blue 5 5
Normal Grey 4 10
Blackeye
Lutino 5
Albino
Clearwing
Greywing
Cinnamonwing
Double Factor Spangle 5 5
Opaline 13
Opaline AOSV
Clearbody
Lacewing
Yellowface 8 1
Spangle 3 10 3 10 3
Dominant Pied 3 5
Recessive Pied 5
Crested 18
AOV AOC
Old Birds Cock ASV/ASC 5
Old birds Hen ASV/ASC

Best in Variety Judge : Kathy Manton
Date: May 2012
Variety Open Novice
1 Normal Green R Selig E Baker
2 Normal Grey Green R Selig ----
3 Normal Blue R Selig M Valvason
4 Normal Grey R Selig ----
5 Blackeye ---- ----
6 Lutino ---- G Gordon
7 Albino --- ----
8 Clearwing ---- ----
9 Greywing ---- ----
10 Cinnamonwing ---- ----
11 Double Factor Spangle M Hill G Gordon
12 Opaline ---- E Baker
13 Opaline AOSV --- ----
14 Clearbody ---- ----
15 Lacewing ---- ----
16 Fallow ---- ----
17 Yellowface ---- J Halas
18 Spangle R Selig Tarpis Family
19 Dominant Pied ---- Tarpis Family
20 Recessive Pied ---- G Gordon
21 Crest P Brett ----
22 AOV AOC ---- ----
Best Cock R Selig Tarpis Family
Variety Spangle Spangle
Best Hen R Selig Ed Baker
Variety Spangle Opaline
Name Variety
Best Open R Selig Spangle cock
Best Novice Ed Baker Opaline hen
Bird of the Night R Selig Spangle cock
Opposite Sex Ed Baker Opaline hen
NUMBER OF BIRDS
Open / Inter 22
Novice 18
Old birds 1
TOTAL 41

2011 / 2012 CUMULATIVE TABLE SHOW POINTS as at May 2012
Variety
Bak
er_
E_J
Bre
tt_P
Bu
ckin
gham
- C
Cauchi_
L
Cia
nta
r_C
Hal
as_
J
Do
dd
- P
Fin
dla
y_B
Gal
bra
ith
- R
Go
rdo
n -
G
Han
cock
_T
Hill
- M
Ken
nedy_
N
Man
ton
- I
& K
AR
ISTO
N_A
Mit
chel
l - K
& D
NES
BIT
T Fa
mily
Selig
_ R
KER
R_P
Val
vaso
n -
M
Zam
mit
_ D
ave
& J
o
Zam
mit
_D &
D
AP
RI_
L?
Tarp
is fa
mily
M_A
ngus
Normal Green 28 3 14 15 5
Normal Grey Green 6 10 3 3 18 5
Normal Blue 6 5 8 5 30 8 18 5 5
Normal Grey 31 5 3 43 15
Blackeye 15
Lutino 5 23 9
Albino 10 8 5 5 5
Clearwing 21 16
Greywing
Cinnamonwing 10 6 35 1
Double Factor Spangle 5 15 5 11 5
Opaline 43 3 5 3 11 28 5 3 3 18 5 5
Opaline AOSV 5 5 10 5
Clearbody 5
Lacewing 3 23 38 3
Yellowface 25 11 10
Spangle 29 5 8 1 1 11 7 16 12 55 11 56
Dominant Pied 6 16 19 8 45 13 5 3 15 14 15
Recessive Pied 25 5
Crested 65 59 13
AOV AOC 5
Old Birds Cock ASV/ASC 25 5 10 5 8 6 3
Old birds Hen ASV/ASC 12 1 4 1 8 15
TOTALS 137 121 15 55 86 36 93 54 68 55 120 47 39 39 6 63 5 214 20 38 13 5 3 96 5
**NB Novice Spangle error has been corrected in this table

BIRD OF THE NIGHT TROPHY 2011 / 2012
August September October November January February March April May June TOTAL
Name Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Bird o
f th
e N
igh
t
Opposite s
ex
Peter Dodd 1 2 3
Ross Selig 2 1 2 2 2 1 2 12
Ed Baker 1 1
Paul Brett 1 1
C Buckingham 1 1
I & K Manton 0
Laurie Cauchi 2 1 3
Ray Galbraith 0
Ken Mitchell 1 1
Brian Findlay 0
Tarpis Family 2 1 3
Nick Kennedy 2 2