home range, movements, and denning sites of raccoons
TRANSCRIPT

HOME RANGE, MOVEMENTS, AND DENNING SITES OF RACCOONS
ON THE HIGH PLAINS OF TEXAS
by
JESSE J. JUEN B.S.
A THESIS
IN
WILDLIFE SCIENCE
Submitted to the Graduate Faculty of Texas Tech University in Partial Fulfillment of the Requirements of
the Degree of
MASTER OF SCIENCE
Approved
Accepted
December, 1981

jr
tJC'» !^'^'' TABLE OF CONTENTS
dZ<^P^^ Page
ACKNOWLEDGEMENTS iv
ABSTRACT v
LIST OF TABLES vi
LIST OF FIGURES vii
CHAPTER ONE
INTRODUCTION 1
CHAPTER TWO
STUDY AREA 3
CHAPTER THREE
METHODS AND MATERIALS 6
Live Trapping 6
Immobilization, Handling, and Measuring . . . 6
Sexing and Aging 6
Marking and Radio Collaring 7
Home Range and Movement 7
Den-Site Selection 8
Habitat Use 8
Transplant Raccoon 9
CHAPTER FOUR
HOME RANGE AND MOVEMENT 10
RESULTS 10
Adult Females 10
Adult Males 12
Juvenile Females 14
Juvenile Males 14
SUM IARY 19
CHAPTER FIVE
DEN SITES 21
RESULTS 21
Playa Basins 21
Agriculture 22
Pastures 22
Man-made Structures 23 • m
11

Page
SUMMARY 23
CHAPTER SIX
HABITAT RELATIONSHIPS 26
SUMMARY 28
CHAPTER SEVERN
TRANSPLANT RACCOON 30
RESULTS 30
SUMMARY 32
LITERATURE CITED 35
APPENDIX 38
111

ACKNOWLEDGEMENTS
I wish to thank my committee members Dr. Robert Baker and Dr. Fred
S. Guthery for the helpful suggestions and review of the manuscript.
I would especially like to thank my committee chairman. Dr. Eric G,
Bolen* for his guidance and friendship, and also Dr. C. David Simpson,
who essentially chaired the thesis except for the final stages of
completion.
I am truly indebted to all the landowners for their kindness and
generosity to me and to this research. I wish to thank F. A. and
Lucille Smith; Preston, Lavern, and Reent Upshaw; Jimmy Ray, Greta, and
Jami Davis, and Richard Ellis for their friendship, hospitality, and
kindness extended to me.
Much appreciation is extended to Elouise Phillips, Kay Arellano,
Becky Watkins, and Nila Brazell for their clerical help and also to
graduates and undergraduates who assisted in data collection.
Special thanks go to Karl Launchbaugh, Bok Sowell, and Ben Koerth
for their many diversions, distractions, music, and lifetime friendship.
Most of all, love and appreciation go to my wife Maggie for her
support, patience and encouragement throughout this research.
iv

ABSTRACT
Raccoons occur throughout the state of Texas. With the recent
dispersal of raccoons in Castro County and increased interest in
conservation of wetlands, there is a need for data on the importance
of playa basins to raccoons as well as other wildlife in the area.
This study was designed to evaluate the importance of playa basins to
raccoons and document the significance of these basins with respect
to; (1) home range and movements of raccoons and, (2) the den-site
selection of raccoons. The study area selected was Castro County,
Texas, and portions of Lamb and Hale Counties, Texas.
Raccoon home ranges varied from 795-3,845 ha. Adult males had
the largest home range areas and juvenile females had the smallest.
Adults and males moved significantly greater (P<0.10) distances than
juveniles and females. However, there was no significant interaction
(P>0.10) between sex and age classes, indicating these characteristics
acted independently of each other.
Sixty eight percent of the den sites were located on playa
basins, 19% in croplands, 3% in pastures, and 10% in man-made structures
All den sites located in vegetation were surrounded by cover 1.0 m or
taller.
Playa basins were used significantly more (P<0.10) in proportion
to their availability while croplands were used significantly less
(P<0.10) in proportion to their availability. Pastures were used in
proportion to their availability.
Playa basins were very important to raccoon use throughout the
year, providing food, water, and cover; however, croplands, especially
com fields were seasonally important to raccoon use.

LIST OF TABLES
Page
1. Occurrence of den/rest site observations in 4 habitats
for raccoons in Castro County, Texas 25
2. Habitat use by radio collared raccoons compared to
proportion availability to show habitat preferences in
Castro County, Texas 27
3. Habitat use by a radio collared raccoon transplanted to
the Castro County, Texas study area from Donley County,
Texas. Comparison with habitat availability indicates
preference 33
VI

LIST OF FIGURES
Page
1. Castro, Lamb, and Hale counties: s±udy area . . . . . 5
2. Home range of AF raccoon #12, 24 April - 4 September
1980, in Castro County, Texas, encompassing 1,896 ha. 11
3. Monthly home range shifts for AF raccoon #12 in Castro
County, Texas 13
4. Home range of AM raccoon #22, 15 August - 18 December
1980, in Castro County, Texas, encompassing 3,854 ha. 15
5. Home range of JF raccoon #17, 5 June - 12 October
1980, in Castro County, Texas, encompassing 1,285 ha. 16
6. Home range of JM raccoon #20, 21 July - 6 September
1980, in Castro County, Texas, encompassing 3,248 ha. 18
7. Home range of transplant raccoon, 11 May - 28 June
1980, in Castro County, Texas, encompassing 288 ha. . 31
Vll

CHAPTER I
INTRODUCTION
The raccoon (Procyon lotor) occurs throughout the United States
except for parts of New Mexico, Arizona, California, Nevada, Colorado,
Montana, and Wyoming (Burt and Grossenheider 1976). Distribution on the
Southern High Plains of Texas is based on 4 county records (Davis 1974).
Castro County, Texas, had no record of raccoon occurrence prior to this
study.
Although considerable research has been published on various aspects
of raccoon ecology (Stuewer 1943, Sanderson 1961, Fritzell 1978), little
is known about its life history in Texas. Previous research has addressed
food habits in east Texas (Baker et al. 1945, Wood 1954), reproduction
and rate of increase in the post oak region (Wood 1955), parasitic helminths
in east Texas (Chandler 1942), and serology of infectious diseases in
south Texas (Cook et al. 1965). The lack of information of raccoons in
other regions of Texas points to the need for further research.
Stuewer (1943) and Fleming (1975) have shown the importance of water
to raccoons. However, its importance to raccoons in the Texas High Plains
is not known. Sources of available water for raccoon use within the High
Plains are draws, tailwater recovery pits, and playa basins. Prior to the
1930's. in the Castro County study area, playa basins were largely ephemeral
and draws were permanent, spring-fed creeks (F. A. Smith pers. commun.).
Today, many basins have been modified to hold water for longer periods,
and, in some instances, contain permanent water. Deep-well irrigation
has lowered the underlying Ogallala Aquifer level to the point where draws
are no longer spring-fed. Water in draws is now ephemeral from seasonal
precipitation and irrigation runoff.
In Castro County, long-term residents independently corroborated
that few raccoons existed in draws or playa basins prior to the 1930's.
After establishment of deep-well irrigation in the 1950's, raccoon numbers
and distribution appeared to increase. Thus, despite adequate water
resources from draws prior to the 1930's, raccoons were apparently not
abundant in this area. However, since 1950 they may have become more
widely dispersed in association with modern agricultural development and
longer-lasting surface water in playa basins.
1

2
With the recent dispersal of raccoons in Castro County and increased
interest in conservation of wetlands, there is a need for data on the
importance of playa basins to raccoons as well as other wildlife in the
area. This study was designed to evaluate the importance of playa basins
to raccoons and document the significance of these basins with respect to
home range, movements, and den-site selection.

CHAPTER II
STUDY AREA
The study area was principally located in Castro County, Texas, in
the Southern High Plains. The topography consists of flatland, broken by
draws and playa basins (Frye and Byron 1957). The county is approximately 2
2,300 km with elevation ranging from 1,100-1,200 m. Average annual
precipitation is 44.0 cm with 78% of this falling during May through
October. Winters are mild and dry with an average of 193 frost-free days
per year (Bruns 1974).
The study area consisted of intensive agriculture and some pastures
with no trees other than a few remnant species in draws, playa basins,
and limited homestead shelterbelts. Major agricultural crops included
corn, wheat, cotton, grain sorghum, and various vegetable crops (Bruns
1974). To a lesser extent, beef and dairy production are also important
in the area, using locally-grown grain for feedlots (Moore 1980).
Farming practices whithin the study area consist of deep-well
irrigation and some dry-land farming. Water supplies for irrigation are
obtained from the underlying Ogallala Aquifer, which extends from the
Southern High Plains of Texas and New Mexico northward through portions
of Oklahoma, Colorado, and Kansas.
The extensive study area encompassed approximately 67,770 ha and was
located in southeastern Castro and portions of northern Lamb and Hale
counties. The area within which all collared raccoon locations were
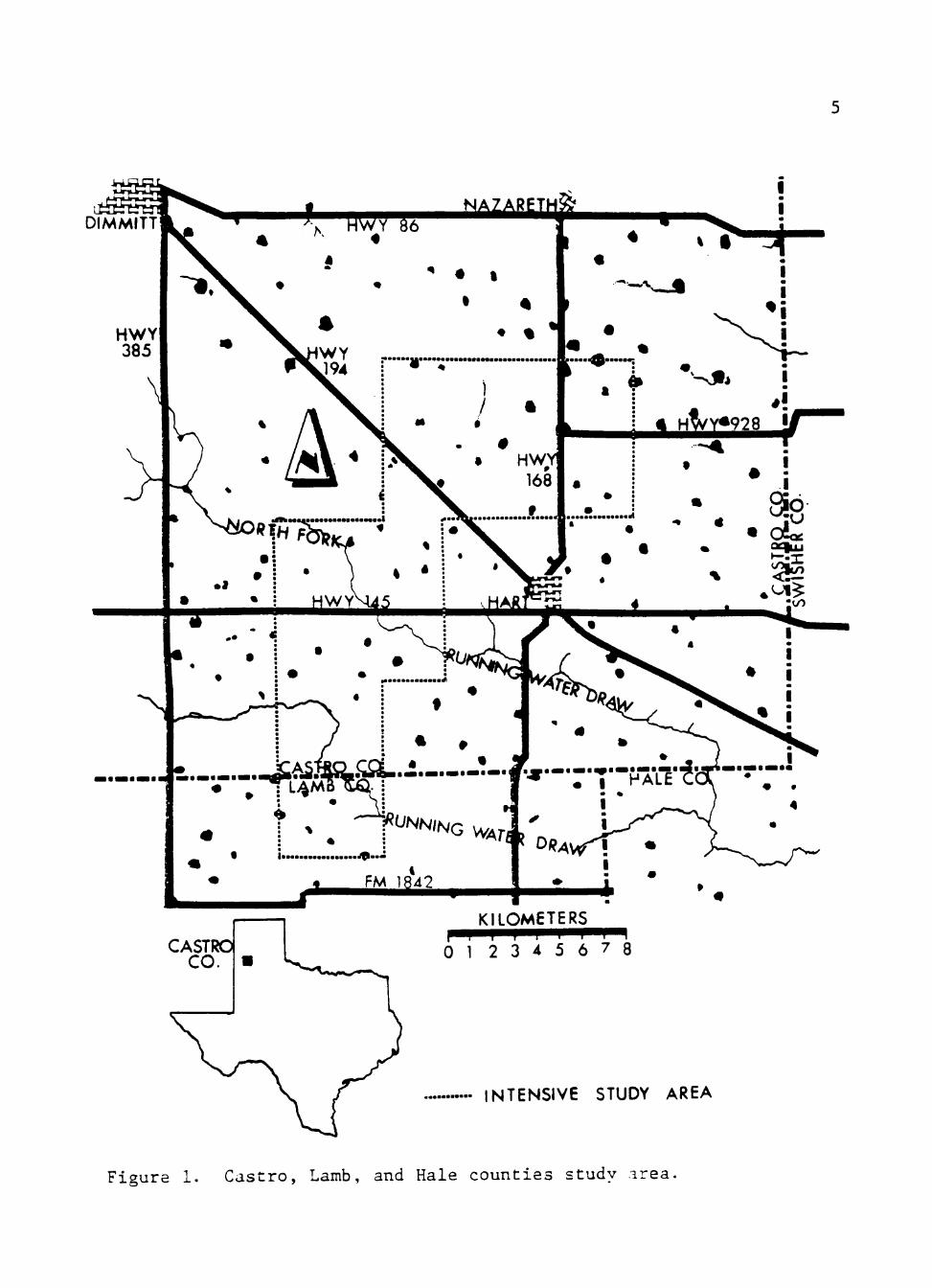
obtained was the intensive study area (Fig. 1).
Within the 23,872-ha intensive study area there were 2 major draws.
North Fork of Running Water Draw and Running Water Draw, and 64 playa
basins. These playa basins are pluvial basins consisting of Randall clays
(Bruns 1974) that act as permanent or ephemeral reservoirs for water.
Because of the slow soil permeability of Randall clays, these basins lose
much of their water by evaporation.
Major vegetation associated with basins that were not cultivated
largely consisted of emergents such as bulrush (Scirpus spp.), cattail
(Typha spp.), smartweed (Polygonum spp.), and spikerush (Eleocharis spp.).
Kochia (Kochia scoparia), and barnyard grass (Echinochloa crusgalli),
3

4
were also found in close association with basins. In some cases pondweed
(Potamogeton spp.) was found in basins with permanent water.
NTENSIVE STUDY AREA
Figure 1. C a s t r o , Lamb, and Hale coun t i e s study area .

CHAPTER III
METHODS AND MATERIALS
Live Trapping
Forty 18 gauge wire Tomahawk live traps (Tomahawk Co., Tomahawk,
Mich.) were used to capture raccoons. Sardines, catfish, tuna fish, duck
carcasses, jelly, apples, and shiny materials were used for bait. The
most successful bait was apples.
The trapping period extended from August 1979 to November 1980.
Traps were placed on playa basins, tailwater recovery pits, and possible
den/rest sites. Traps were checked every morning and evening, and trapping
effort was recorded as one trap/night for each setting.
During the trapping period human interference was encountered. This
included trap damage, theft of animals, or theft of traps. In some
instances traps were damaged beyond repair. At the end of the trapping
period only 22 out of 40 traps remained, thereby affecting trapping effort
and coverage.
Immobilization, Handling, and Measuring
Captured animals were confined in the trap and their movements
restricted with a wooden plunger (E. G. Bolen, pers. commun.). Ketamine
hydrochloride (Parke-Davis, Detroit, Mich.) was used to anesthetize
captured individuals because of its broad safety margins and short recovery
time (Bigler and Hoff 1974). Each animal was injected intramuscularly in
the posterior region of the body with a dosage of 22 mg/kg body weight.
The time period from injections to a tranquilized state was recorded for
each anesthetized raccoon (Appendix A).
Standard measurements and weight were taken for each animal. These
were right ear length, right hind foot length, tail length, total body
length, and total body weight. Weight and growth differences were also
measured on all recaptured animals.
Sexing and Aging
Sex determination was based on the presence or absence of an os penis
and by the locations of urinary openings (Sanderson 1950). Penis
6

7
extrusability and size, teat size and color (Sanderson 1961), and tooth
wear (Grau 1968) were used to age raccoons. Animals were placed into 5
age classes; 0-14 months, 15-38 months, 39-57 months, 58-86 months, and
86 months or older (Grau 1968). Animals less than 14 months were
referred to as juveniles and those older than 14 months as adults.
Marking and Radio Collaring
Captured raccoons were ear tagged following Butterfield (1944). A
numbered commercial metal poultry tag was placed in each ear to identify
retrapped animals and mortalities. During the first 4 months of trapping,
colored ear streamers were used in conjunction with metal ear tags. Ear
streamers were discontinued after 1979 when it was found that streamers
were missing from retrapped individuals and marked mortalities. Raccoons
also were toe clipped in a numerical series to aid identification by
tracks.
Thirteen raccoons were collared with L2B5 transmitters attached to
the animals with a special cable-collar configuration (Telonics, Mesa,
Az.). Collars were fastened with an alimiinum lock bolt and weighed
approximately 80 g. When the collar was attached, enough space was
allowed to permit the animal to function normally. Collars were checked
with the receiver to assure proper working order before raccoons were
released.
Home Range and Movement
A 148 Mhz frequency receiver and a hand held yagi antenna were used
to monitor locations of 9 radio-collared raccoons for home range and
movement data. Located animals were monitored at approximately 30-minute
intervals throughout each monitoring night. Also, one diurnal location
was obtained between 1200 and 1600 h whenever possible. Triangulation
was used to pinpoint animal locations.
Daily locations and movements were plotted on county maps, then
transposed onto USGS topographic maps each month. Home range was delimited
by the modified minimum home range method (Harvey and Barbour 1965).
Home range size was determined for each radio-collared raccoon using
only telemetric locations obtained from date of release to date of radio
failure.

8
Telemetric locations, recaptures, mortalities, and identified tracks
were combined to provide movement data. Distance moved was calculated
by summation of all locations for an individual from the time of departure
from a rest site until the next rest site was established (Siniff and
Tester 1965). Marked tracks were used to continue monitoring movements
outside home range areas subsequent to radio failure. A split-plot
analysis and Duncan's multiple range test (Steel and Torie 1978) were
used to analyze movement data and differences between means for sex and
age classes.
Den-Site Selection
Den-site analysis did not include qualitative separation between
den and rest sites because of the lack of defined differences. Therefore,
den and rest sites are used synonymously in this report.
Information taken at each den/rest site, was surface temperature,
air temperature, and distance to nearest surface water. Each den/rest
site was located with telemetry equipment between 1200 and 1600 h.
Analysis of den/rest site data was based on percent occurrence of all
sites within those habitats used for denning/resting by raccoons.
Habitat Use
Habitat use by raccoons was investigated to determine preferences
using a non-mapping utilization vs. availability technique (Marcum and
Loftsgaarden 1980). Assumptions associated with the technique are that
all habitats are available for selection by each animal and that animal
locations be determined randomly.
The null hypothesis formulated was that each raccoon used each
habitat in proportion to its availability within the individual's home
range. A Chi-square test was conducted on the recorded use of each
habitat, using playa basins, pastures, and agricultural land as primary
habitat categories, and corn, wheat, cotton, milo, sunflower, alfalfa,
oats, and fallow fields as subheadings under agricultural land.
To examine habitat preferences, simultaneous confidence intervals
were constructed using the proportion of each habitat and observed number
of raccoon locations in each habitat (Marcum and Loftsgaarden 1980).

9
If the confidence interval for any habitat included 0, that habitat was
used in proportion to its availability. If 0 was not within the confidence
interval and both end points were positive, the habitat was used
significantly less (P<0.10) in proportion to its availability. If 0 was
not within the confidence interval and both end points were negative,
the habitat was used significantly more (P<0.10) in proportion to its
availability.
Transplant Raccoon
During the course of this study the opportunity arose to instrument
and compare data from a transplant raccoon to data collected on resident
raccoons. A juvenile female was transplanted from Donley County, Texas,
to the Castro County, Texas study area. The same methodology was applied
to monitoring the transplant female as described earlier for resident
raccoons.

CHAPTER IV
HOME RANGE AND MOVEMENT
Home range for raccoons has been researched by numerous workers.
Lotze (1979) compiled a thorough review of these studies showing author,
state, method used, sex and age, number of data points, duration of study,
and home range size. Researchers also have shown great differences in
movement patterns for raccoons throughout the United States. In east-
central Illinois, Ellis (1964) found raccoons moved an average minimum
distance of 196 m per daylight tracking period. In contrast, Fritzell
(1978) found raccoons in the prairie pothole region of North Dakota
traveled mean distances of 1.2 km to 3.7 km during nocturnal hours.
Urban (1970) and Fleming (1975), working in Ohio and Louisiana, respectively,
reported similar findings to Fritzell (1978).
RESULTS
Adult Females
Radio contact was lost with 2 of the 3 adult females (AF) within 1
week of release. Only 1 individual was monitored long enough for adequate
home range and movement analysis.
AF #7 was collared 16 September 1980. On 28 September 1980 the collar
was found 1.3 km from the release site submerged in a runoff pit under
water and mud. The presence of her marked tracks around the pit and
condition of the radio collar, which was retrieved from the water, led to
the assumption that she was taken by a local trapper and the collar
discarded into the pit. AF #41 was collared 4 November 1980. On 9
November 1980 the collar was found inside a closed trap, indicating AF #
41 had been retrapped and removed from the area. Trap appearance, marked
tracks, and collar condition again led to the assumption of removal by a
local trapper.
AF #12 was instrumented 24 April 1980 and monitored regularly over
a 4-month period until radio contact was lost after 4 September 1980.
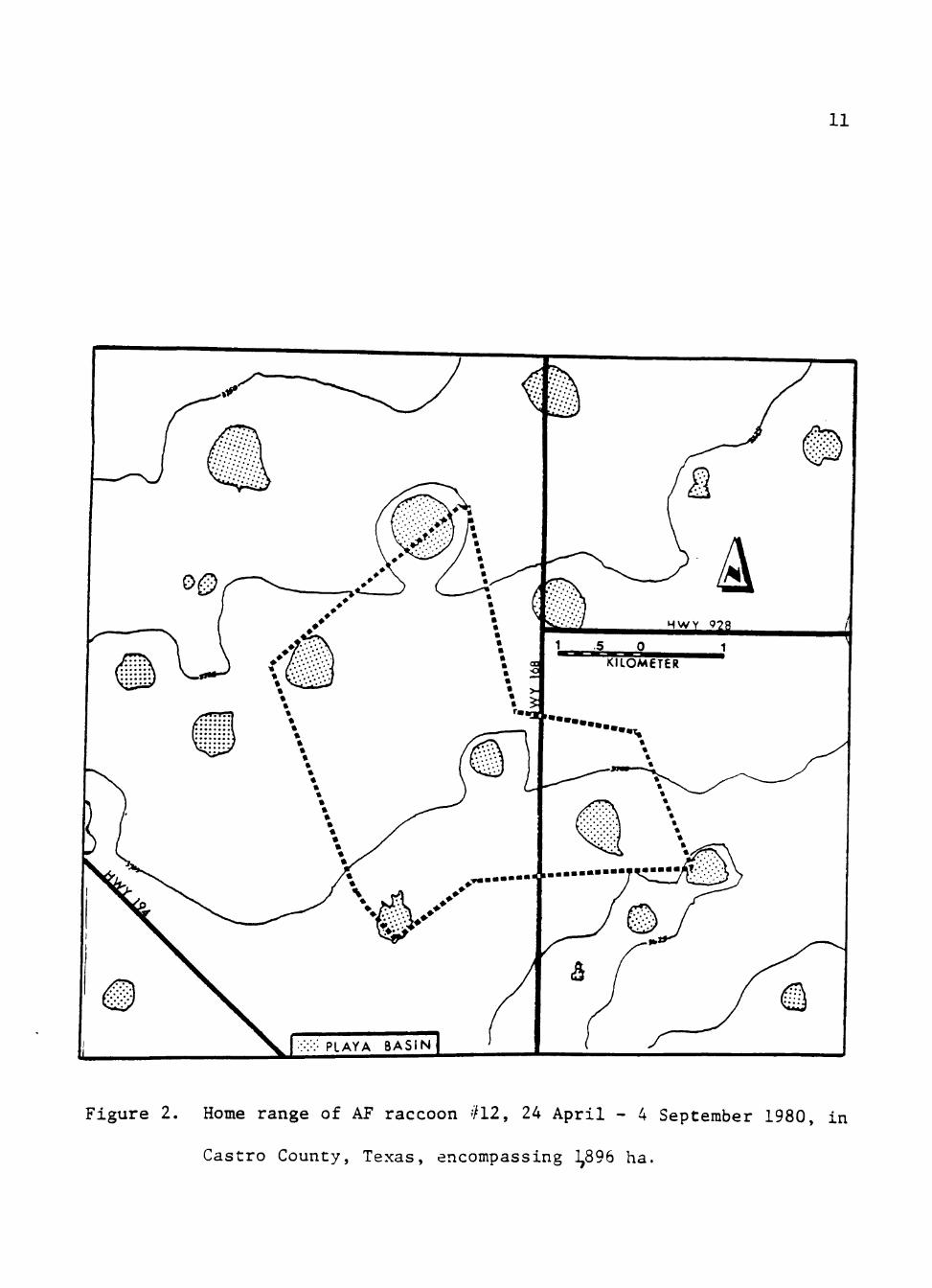
Her established home range during this period was 1,896 ha (Fig. 2).
This animal exhibited a mean nightly movement of 2.7 km ranging from 0.6-
11.6 km
10
11
Figure 2. Home range of AF raccoon #12, 24 April - 4 September 1980, in
Castro County, Texas, encompassing ],896 ha.

12
AF #12 gave birth to 6 kits on approximately 22 April 1980 in a
haystack located next to a playa basin. Her movements apparently were
not restricted by the presence of her kits. During the first 2 weeks
after parturition the female traveled up to 5.3 km from the nest at night
but always returned and denned there just before or shortly after dawn.
In the second 2 weeks the female traveled similar distances up to 5.5 km
from the nest. At this time she began returning to the nest on alternate
evenings, denning at the nest overnight on 2 occasions and visiting the
den on 3 other occasions. During the final 2 weeks and 4 days before the
nest was abandoned, the female continued movements up to 4.9 km from the
nest but only visited for short periods of time during night. During the
day she denned under an abandoned house once and an abandoned shed twice,
both of which were close to the nest.
Marked tracks in June showed AF #12 was traveling with all 6 kits.
This situation continued until 5 August 1980 when only 4 kits were recorded
traveling with her. AF #12's use of playa basins with cover, food, and
water remained constant throughout the monitoring period, but the area
used changed from month-to-month (Fig. 3). Similar shifts were indicated
by all raccoons monitored long enough to map these changes. Monthly locations
showed heavy use of playa basins that contained areas of dense vegetation.
Areas between basins were traveled through corn fields where standing
crops also provided cover and food for raccoons.
Adult Males
Only 2 of the 3 collared adult males (AM) were monitored long enough
to offer adequate data for analysis.
AM #19 was killed by an automobile 3 days after instrumentation and
release. The collar was retrieved and used again. AM #18 was collared
18 June 1980 and monitored until radio contact was lost 31 July 1980.
The animal's home range for the period was 1,049 ha. Mean nightly
movement for this individual was 2.7 km, ranging from 0.4-3.9 km.
Concentration of telemetric locations were on playa basins having abundant
cover, food, and water from June through August, and in standing com
fields adjacent to these basins.
AM #22 was collared 15 August 1980 and monitored until loss of radio
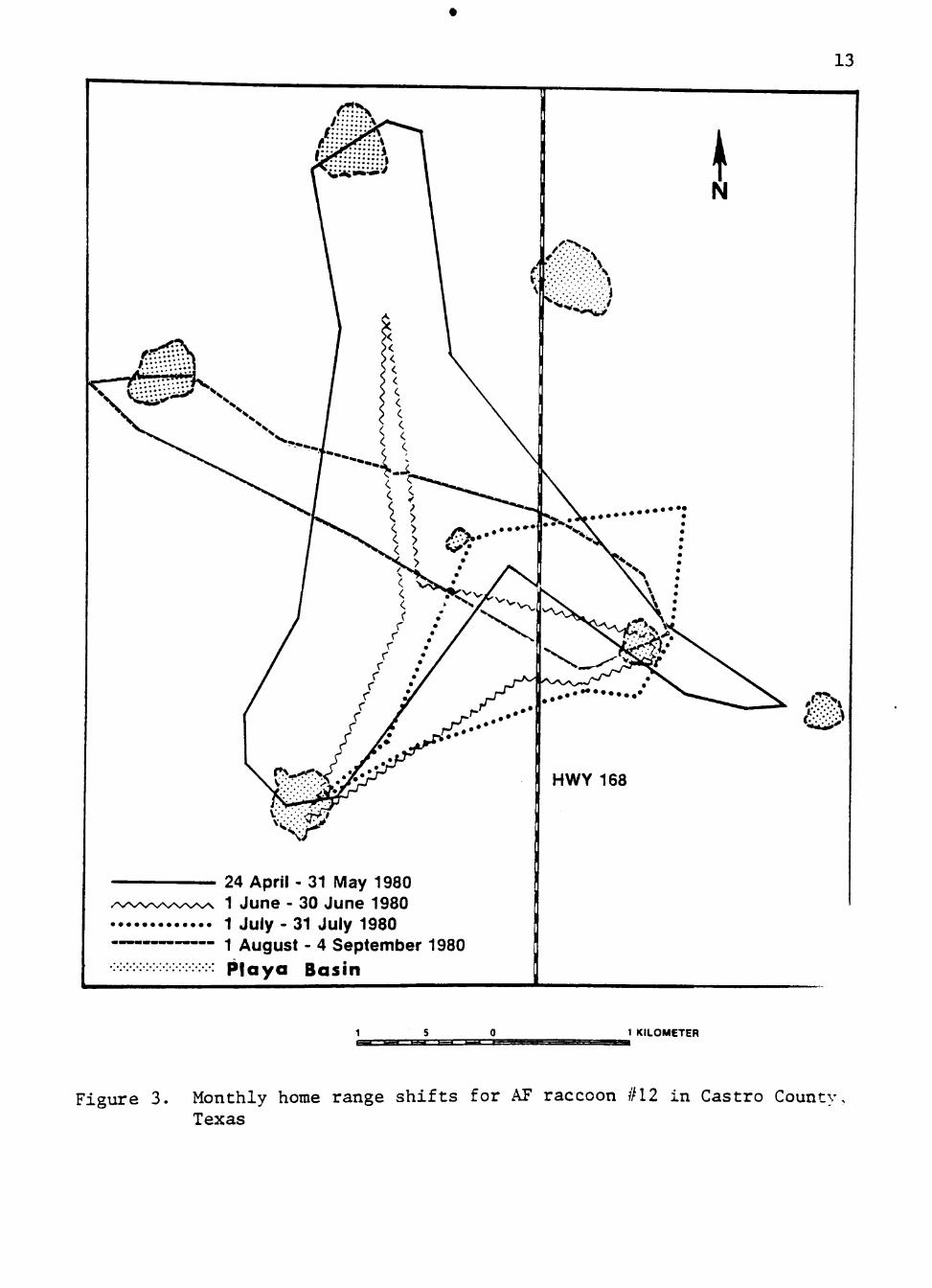
13
24 April - 31 May 1980 1 June - 30 June 1980 1 July - 31 July 1980 1 August - 4 September 1980
Playa Basin
1 KILOMETER
Figure 3 . Monthly home range s h i f t s for AF raccoon #12 in Cast ro County, Texas

14
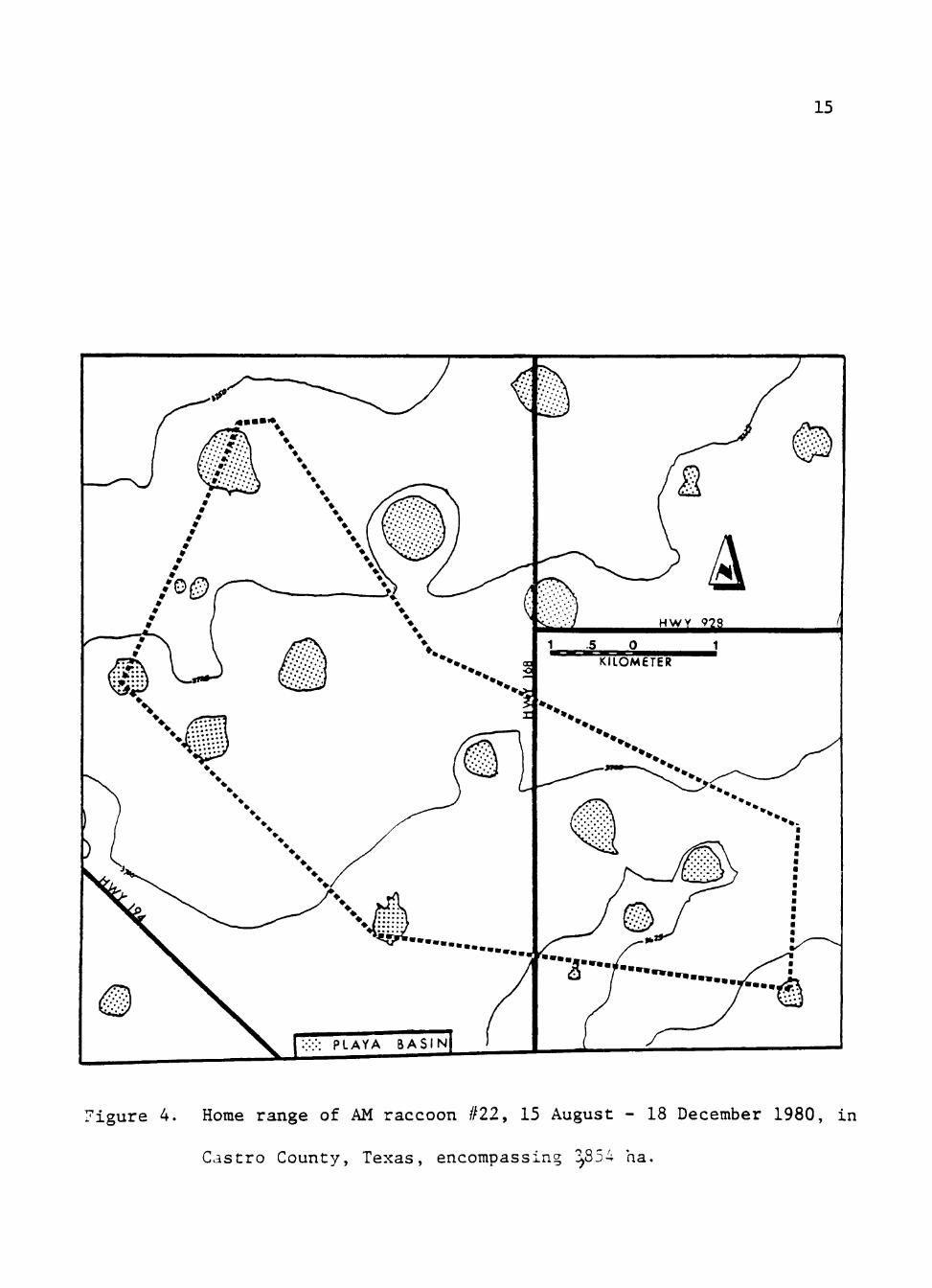
AM #22 was collared 15 August 1980 and monitored until
loss of radio contact 18 December 1980. This individual had
the largest home range of all instrumented raccoons,
encompassing 3854 ha (Fig. 4). Mean movement was 5.6 km,
ranging from 0.7-13.7 km. This was, again, the largest mean
nightly distance traveled for collared individuals. Telemetric
locations were concentrated on playa basins and adjacent corn
fields. The area used by this animal varied monthly but close
proximity to playa basins seemed important.
Both monitored AM's showed heavy use of playa basins and
associated standing crops. However, AM #22 later restricted
his movements and the size of his home range after October,
following corn and cotton harvests.
Juvenile Females
Juvenile female (JF) #25 was collared 2 October 1980 and
monitored until radio contact was lost 11 November 1980. Home
range for this animal was 890 ha and mean nightly movement
was 3.8 km, ranging from 2.3-6.5 km. On 1 January 1981 JF
#25 was located in Hale County approximately 39.5 km southeast
of her monitored home range after being killed by an automobile
JF # 17 was trapped, instrumented, and released on 5
June 1980. The monitoring period ended 12 October 1980 when
radio contact was lost. Home range for this individual was
1,283 ha (Fig. 5). Mean nightly distance traveled was 3.0 km,
ranging from 0.2-6.7 km. Both basins and standing corn fields
were used frequently by this animal.
Both juvenile females had similar home range areas and
mean movement distances during their overlapping monitored
periods. JF #17 increased her home range size in September
and October, which may have represented greater movement
because of more limited availability of cover.
Juvenile Males
Juvenile male (JM) #15 was collared 4 June 1980 and
monitored until 20 June 1980 when radio contact was lost.
15
figure 4. Home range of AM raccoon #22, 15 August - 18 December 1980, in
Castro County, Texas, encompassing ! 854 ha.

]6
KILOMETER
PLAYA BASIN
Figure 5. Home range of JF raccoon #17, 5 June - 12 October 1980, in
Castro County, Texas, encompassing 1,285 ha,

17
For this short period JM #15 had a home range of 1,057 ha,
which suggested dispersal movement more than nightly foraging
activities within an established home range. Mean nightly
movement for this animal was 3.6 km, ranging from 0.7-6.6 km.
Locations for JM #15 were not concentrated around playa basins
as was found for other instrumented animals.
Marked tracks of this animal were sighted on 5 and 7
February 1981. Both track sightings were 11.4 km and 7.3 km
northeast of the monitored home range, respectively. This
suggested that the animal had enlarged its home range by
moving long distances to and from the original home range or
had established a new home range beyond the study area.
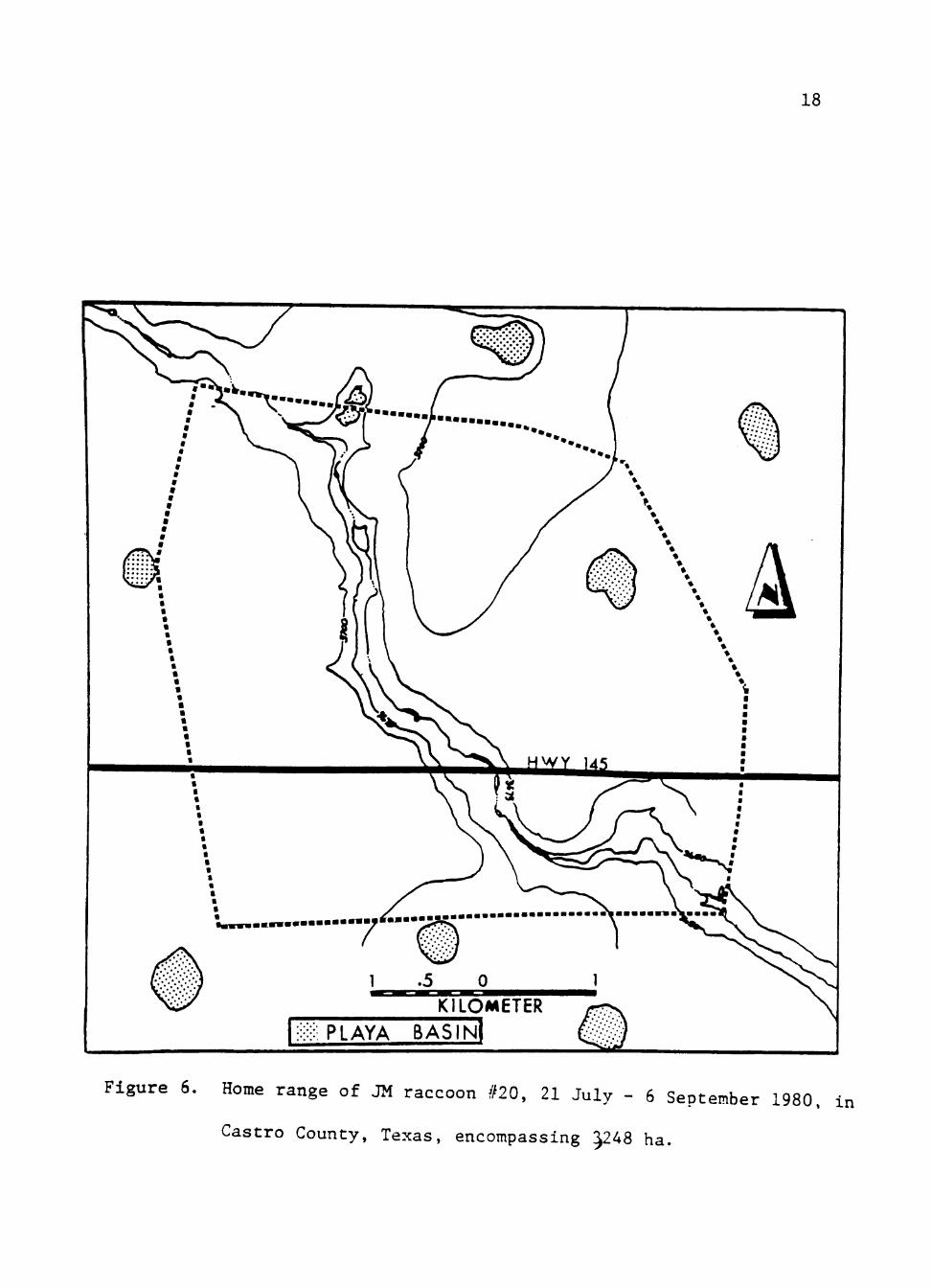
JM #20 was collared and released 21 July 1980 and
monitored until radio contact was lost 6 September 1980. Home
range for the period was 3,248 ha (Fig. 6 ) , larger than any
instrumented raccoon except AM #22. Mean nightly movement
was 5.2 km ranging from 1.4-10.9 km. JM #20 consistently
used pastures associated with draws and standing corn fields
even though playa basins existed within the home range area.
JM #20 was seldom found on playa basins in the area. In
Januarv he was found dead in a sheltered haystack 6.8 km west
of his monitored home range.
JM #23 was collared and released 7 September 1980. He
was monitored until 24 October 1980 and exhibited a home range
of 795 ha. Mean nightly movement was 1.8 km ranging from
0.5-5.1 km. In contrast to JM #20, this individual showed
a high association with playa basins.
After loss of radio contact, marked tracks were sighted
for JM #23 on 3 occassions: 17 October 1980, 2.3 km east of
the monitored home range; 9 and 10 December 1980, 3.5 km and
3.3 km also east of the monitored home range, respectively.
These tracks indicated that he was still close to his original
home range during October and December.
18
PLAYA BASIN]
Figure 6. Home range of JM raccoon #20, 21 July - 6 September 1980, in
Castro County, Texas, encompassing J248 ha.

19
SUMMARY
Area used by raccoons ranged from 795 ha to 3,845 ha, with AM's
having the largest home range areas and JF's the smallest. AF home
ranges were slightly larger than those recorded for JM's. Of all sex and
age classes, JM's showed the most contrast in home range size and area
used. In the case of JM #15, the short monitoring period and extent of
movement indicated dispersal. His marked tracks showed continued long
distance movements after loss of radio contact. JM #23 exhibited the
smallest home range for all monitored raccoons in contrast to JM #20,
which exhibited a large home range, but JM #20 was monitored longer than
#23 was. Lotze's (1979) review indicated that only 1 study (Fritzell
1978) recorded home ranges as large as those found in this study. When
compared to Fritzell (1978), home range sizes in this research were
larger for all sex and age classes except AM's.
Overlapping home ranges occurred for all collared individuals except
AM #18. It was not uncommon to monitor 2 or 3 different raccoons on the
same playa basin during 1 night. Raccoons in Michigan, observed by marked
tracks, also showed overlapping home ranges (Stuewer 1943).
Movement data for raccoons in this study showed there was a significant
difference (P<0.10) between males and females and between adults and
juveniles. Adults moved more than juveniles and males moved more than
females. There was no significant (P>0.10) interaction between sex and
age classes, indicating these characteristics acted independently of each
other.
No restriction of movement was evident among either sex or age
classes. Even AF #15 showed extensive movement away from the nest after
parturition and during postnatal care. Contrary to this study, Ellis
(1964), Schneider et al. (1971), and Fritzell (1978) reported shorter
movements of females after parturition and during postnatal care.
Rlange and movements for raccoons changed monthly during this study.
Research on raccoons in Louisiana and Minnesota also reflected this
characteristic (Mech and Turkowski 1966, Fleming 1975). Long-range
dispersal and extension of monthly home range areas were especially
noticeable from October through December. This period covers a season
in which there is little food, cover, or water available to raccoons.

20
Within the study area, crop harvesting was completed by October. This
left barren fields over a large portion of the county. Irrigation
decreased and surface water was restricted to 3 permanent playa basins,
tailwater pits, and cattle watering facilities which froze intermittently.
By December, cover was restricted to man-made structures, burrows, playa
basins, and some pastures. This depletion of the environment during the
winter months may have forced range extension and dispersal as raccoons
searched for food, cover or water. Monitored raccoons were feeding in
corn fields after harvest, but from December through February large
concentrations of marked and unmarked raccoon tracks were found on playa
basins that had permanent water. Scats of raccoons at this time showed
no corn contents.
Gels (1966) showed evidence of JM dispersal between August and
November but the maximum dispersal distance from release sites was only
3 km. Stuewer's (1943) data on raccoons in Michigan indicated some JM's
and JF's dispersed during summer months.
Datum in this study suggest home range and dispersal of resident
raccoons within the study area were disimilar to previous studies on
raccoon. What seems to be winter dispersal may actually be a constant
seasonal range extension that includes changing areas to find adequate
food, water, and cover. Since wititer is the most exigent season, range
extension becomes similar to dispersal movements in all sex and age groups.
When crops were planted and irrigation practices increased during spring
and summer the activity range for resident raccoons decreased in size.

CHAPTER V
DEN SITES
Den/rest sites used by raccoons have been studied by Giles (1942),
Stuewer (1943), Mech et al. (1966) and Berner and Gysel (1967). Sites
differ from region to region depending on location and availability.
Stuewer (1943) stated that raccoons in Michigan denned in various types
of tree dens including oaks (Quercus spp.), maples (Acer spp.), ash
(Fraxinus spp.), butternut (Juglans cinerea), and sycamores (Platanus
spp.). In contrast, Dorney (1954) found 90% of 81 raccoons on the Horicon
Marsh in Wisconsin used ground dens on islands or ditch-banks, 5% denned
in frozen muskrat houses, and 5% used tree dens.
RESULTS
Playa basins received varied use as den/rest sites by raccoons. All
sex and age classes occupied den/rest sites in playa basins and were
observed occupying new and re-using specific basins within their home range
areas. Movement between basins and re-use of the same den/rest sites were
frequent. In contrast, 2 collared raccoons showed no specific association
with 1 or 2 basins and seldom used the same rest site more than once during
the monitored period. Fleming (1975) and Schnell (1970), studying raccoons
in Louisiana and Minnesota, respectively, supported the fact raccoons
searched for certain den/rest sites and randomly chose sites as well.
Schnell (1970) suggested that selection was dependent upon the degree of
"foraging or investigatory behavior" of the raccoon.
Playa Basins
Den/rest sites on basins were located under or in tall, dense
vegetation such as cattails, bulrush, willow, smartweed, dock (Rumex spp.),
and Johnsongrass (Sorghum halepense). In these areas, den/rest sites were
observed on soil, matted vegetation above ground, and matted vegetation
above water.
Of the 128 sites located on playa basins, average vegetation height
was 2.1 m, ranging from 1.6-2.7 m. Mean den/rest site temperature for
July and August was 27.7°C while mean air temperature was 31.5 C. For
21

22 o.
September and October mean aite temperature was 22.5 C and air temperature
averaged 28.1 C. During November and December the mean den/rest site and
air temperatures were 10.3 C and 17.3 C respectively.
Agriculture
In cropland areas raccoons were observed denning/resting only in
standing crops. The number of sites in cropland areas represented 19.1%
of all site locations (N=189) with 31 locations in corn, 2 in wheat, 2 in
sorghum, and 1 in a sunflower field.
AM's were telemetrically located in cropland areas but den/rest sites
were not observed in agriculture for this sex/age category. Collared
individuals from all other categories used cropland areas and denned/rested
under tall stalks upon moist irrigated soil or matted vegetation above
wet soil. Mean crop height was 2.0 m, ranging from 1.7-2.3 m. Mean site
temperature for all cropland areas was 28.8 C while mean air temperature
was 30.0°C.
No sites were observed in cotton fields; therefore it was assumed
that cotton fields did not suit the den/rest site requirements of raccoons.
Also, no sites were recorded in cropland areas after standing crops were
harvested in October, although many stubble fields remained through
December. Presumably this habitat no longer afforded enough cover.
Pastures
All sex and age classes except AF's were observed denning/resting
in pastures. Two sites were located in Johnsongrass/cocklebur (Xanthium
strumarium) patches and 4 were located in ground burrows. These 6 den/
rest sites represented 3.2% of the total sites located for all raccoons.
Mean site and air temperature for both Johnsongrass/cocklebur sites was
25.8°C and 29.4°C, respectively. Mean site temperature for burrows was
24.1°C while mean air temperauture was 36.9 C.
Vegetation surrounding sites in the Johnsongrass/cocklebur areas
averaged 1.5 m in height. However, burrows were surrounded with sparse
cover or short grasses and bare ground.

23
Man-made Structures
Structures such as barns:, abandoned houses and sheds, irrigations
wells, haystacks, wood piles, and silage pits, represented 10% of all
monitored den/rest sites. However, accounts by local farmers and residents
indicate higher use of structures by raccoons. Stuewer (1943) and Mech
and Turkowski (1966) reported use or artificial structures as den sites
by raccoons during winter.
SUMMARY
Raccoons used playa basins most often as daytime den/rest sites in
areas with dense, tall vegetation. These basins seemed to provide secure
cover as well as supplies of food and water. In playa basins and croplands
vegetation height and site temperature may be important factors in den/
rest site selection. Other than ground burrows and man-made structures,
all sites were located in vegetation no less than 1.0 m high, which
indicates importance of tall cover.
In agricultural areas, corn and wheat were irrigated during May-
August. This irrigation provided water and cool resting/denning sites
for raccoons. Although raccoons were not recorded denning/resting in any
agricultural crop other than corn or wheat, other irrigated fields were
available.
All temperature readings were cooler at the actual den/rest sites
than air temperatures 2 m above the sites. Readings were not taken after
December, but temperature readings would probably be warmer at the actual
site during cold winter weather. This difference is important and could
govern thermoregulation and behavior of individual animals in site
selection at specific times of the year.
Lack of sites in pastures may be attributed to low sparse vegetation
because of grazing pressure and seasonal flooding of draws bordering
pastures. However, ground burrows used as den/rest sites may provide
protection and comfortable denning/resting temperatures. Use of ground
burrows also may increase with the decrease of vegetational cover and the
onset of severe weather in winter.
Use of man-made structures as den/rest sites was not common except
for breeding females. This specific use may be characteristic of pregnant

24
females that use these structures for better protection of their young.
No use of man-made structures was evident after the young were old enough
to leave the nest site CTahle 1).

25
on
y
CO
o
5 UJ CO
a UJ
CO
CO UJ
UJ Q
o 8 CJ
<Z5
hO LQ r O (ZD
?d CD CSI CNI
OO CVI CSI i-H
OD
CO
<-Q
1 - ^
CO < CQ
$
3 Q _
CO
PLA
N
o <5
CO UJ
? CO <
a_
CO. UJ Ql
p y f -
C/?'

CHAPTER VI
HARITAT RELATIONSHIPS
Raccoons can adapt to many habitats. Studies have shown raccoons
exclusively using mixed hardwood forests and bottomland habitats (Stuewer
1943, Mech et al. 1966). Recently, habitat use by raccoons has been
investigated in coastal marsh areas CFleming 1975) and prairie pothole
regions (Fritzell 1978). These studies indicate their adaptability to
varied habitats for denning, feeding, and resting by extending home range
areas and by employing certain behavioral characteristics. In this study,
determining the importance of playa basins to raccoons was a primary
objective.
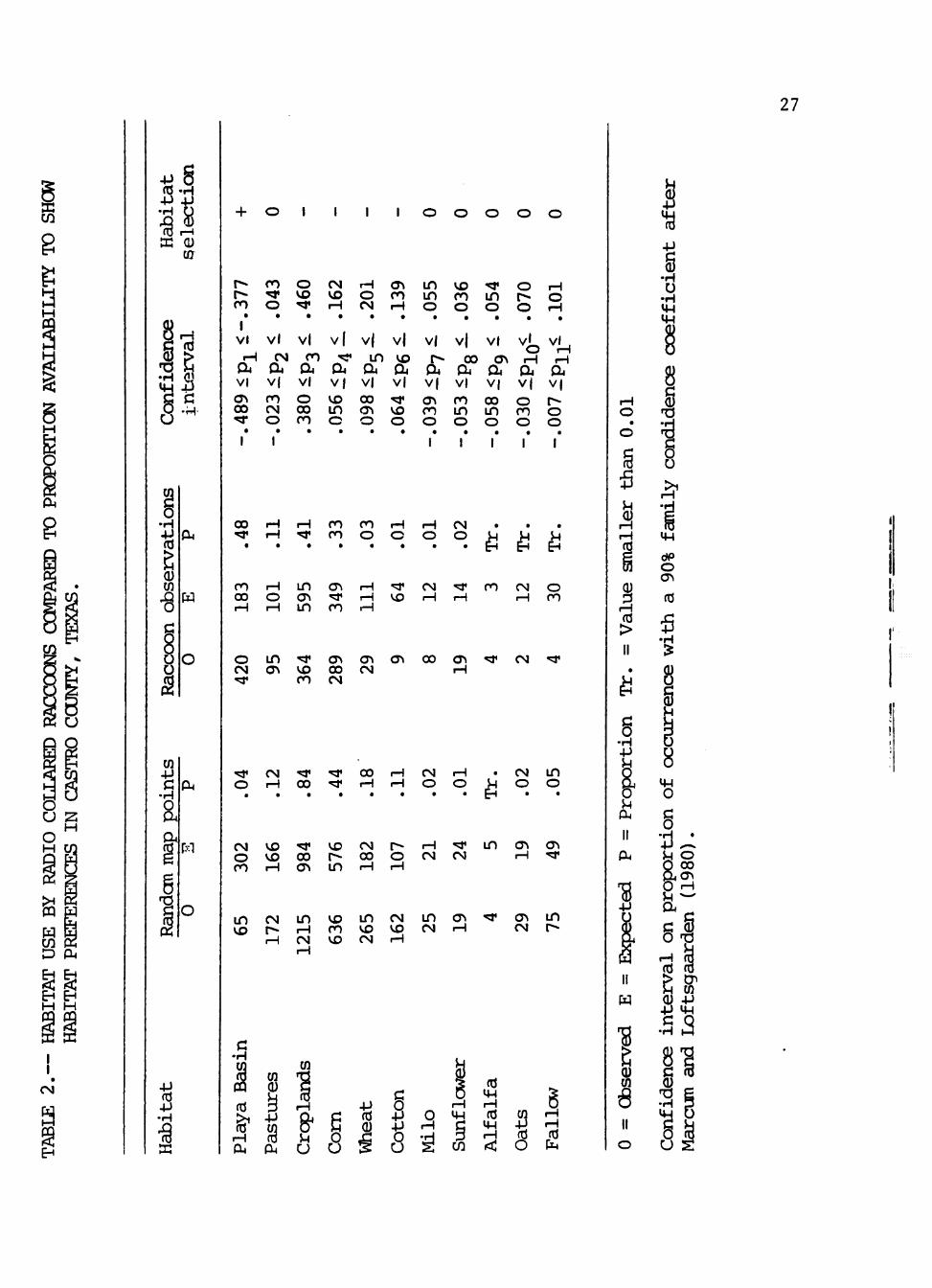
The expected number of raccoon observations differed significantly
(P<0.10) from the availability of all habitats within raccoon home ranges.
Playa basins were the only habitat used significantly more than in !
proportion to their availability (Table 2), including ephemeral and
permanently watered basins. Basins comprised 4% of home range areas
whereas 48% of all telemetric locations occurred in playa basins.
Corn, wheat, and cotton were the major crops and were used less than
in proportion to their availability. However, milo, sunflower, alfalfa,
oats, and fallow fields were minor crops and were used proportionately.
Croplands comprised 84% of raccoon home ranges and 41% of all
observations occurred in these habitats. Movements through most agricultural
areas were generally direct and rapid, indicating use as travel lanes.
However, taller crops such as corn were used for feeding and denning/resting
sites with 33% of all locations occurring in corn fields.
Pastures were used in proportion to their availability with 11% of
all observations occurring in these habitats. Telemetric observations of
movements showed direct and rapid travel across pastures, but these areas
were sometimes used as feeding and denning/resting sites.
Availability could not be measured for artificial structures so
preference of these habitats was not determined. Only 18 locations were
observed in man-made structures. Raccoon use of these structures was
never observed during night monitoring. However, tracks and scat data
suggested greater use of artificial structures than telemetry indicated.
26
27
I I
CM
i)S (0 -H
-9 fD r-l ac 0)
m
infi
den
ce
rX - O
+
r r CO
1 VI iH a VI
cr» 00 'r
o
en •^ o
vl CM
Cu v| CO CN o
1
o vo ^
v( en a vl o 00 en
1
CN vo iH
vl ^
QA
v| vo in c
1
rH O CM
v| in &. vl 00 cr» o
1
<T\ en rH
vl VO QA
vl •^ vo o
o
in in o
vl r> a v( (J CO o
o
vo en o
v| 00 a vl CO in o
o
•^ in o
VI a\
Ch V| 00 in o
o
o r* o
rH a v| o en o
o
rH o rH
Vl I "
r" a v| r* o o
0 •H
1 (U
§ I
cu
w
cu
r :!
o
1 4-> • H
00
183
rH iH
rH O rH
iH
in
in
en en
CO
en o
rH rH
rH
o
vo
rH O
CM iH
CN O
rH
^ ^ ^
en csi
o i n ' * c r > ( y > c T » o o o ^ « « * CM CJN vo 00 CM rH '^r CO CM
^
a 8 « Q i 4H
MH rH fO
MH
O en
CM "^r
o
302
in vo
CM rH
VO vo rH
Csl
00
00 C3^
in rH CM rH
VO
in
vo CO
vo
00
CM 00 rH
in vo CM
o
CM VO rH
CM O
rH CM
in CM
o
CM
Tr.
in
^
CM
o
cr>
CM
in
o
in
3 s d rH fd Id C/3 (< O t^
o o
fO
fd >
§ •H -P
a o u OA
PA a:

28
Personal communications with, landowners on us£ of open-well irrigation
pipes by raccoons supported this.
SUMMARY
Playa basins were the only habitat preferred by raccoons during the
study. These habitats had dense vegetation, available water, food, cover;
playas were used for den/rest sites and travel lanes. These characteristics
provided habitat that was used by raccoons as often as pasture and cropland
habitats combined.
The importance of basins may become most critical during the winter
months when water and food become scarce. Stuewer (1943), Dorney (1954),
and Fleming (1975) have all related the importance of water to raccoons
but nothing has been reported on its significance during the winter months
in the High Plains of Texas. Stuewer (1943) suggested that water was not
important to raccoons in Michigan when denning occurred during harsh winter
months. By contrast, water seemed important to raccoons in this study.
Irrigation ceased and precipitation rates decreased during fall, causing
ephemeral basins to dry. Except for occasional precipitation and winter
wheat pre-plant irrigation, dry conditions persisted through winter
resulting in decreased aquatic resources. Therefore, a limited number
of modified playas and tailwater pits within the study area, provided
water throughout winter. Evidence for continued use of permanent basins
only, was determined from the large numbers of marked and unmarked tracks
on these basins, especially during the December-February period. Tracks
were not observed on tailwater pits in January and February.
Several foods used by raccoons were closely linked to playa basins,
although many items were temporal in availability. Fleming (1975) stated
that the annual diet of coastal-marsh raccoons in Louisiana varied with
precipitation, tide levels, and water salinity, and suggested a close link
between food availability and water. In Castro County, food availability
was limited during winter compared to other seasons. Increased use of
permanent basins during winter also may have occurred because of their
available food sources as opposed to pastures or croplands.
Other than possible sources for food and water, vegetation on basins
was dense and rank in fall and winter months providing cover, and possible

29
den/rest sites.
In croplands, grain was consumed by raccoons and fields were used as
travel lanes. Several researchers have found grain in varying amounts in
raccoon diets (Whitney 1952, Johnson 1970, Fleming 1975). Giles (1930)
stated that raccoon diets in Iowa contained 68.5% corn from March through
December, whereas raccoon diets in New York contained 19.2% grain by
frequency and 14.7% by volume (Hamilton 1951). Examination of fresh scats
in the study area showed raccoons consumed large amounts of corn between
July and December but none in January and February.
Locations were monitored for raccoons in pastures only from May through
October. Raccoon use of pastures was limited to travel lanes, den/rest
sites, and possibly some feeding. More importantly, pastures provided
abundant food sources during the warm season. Foods such as insects, bird
eggs, and crustaceans were readily available to raccoons. These have
previously been reported in raccoon diets (Hamilton 1951, Fleming 1975).
In the spring and summer of 1980, a grasshopper infestation of pastures
occurred in the study area. Examination of fresh scats showed grasshoppers
comprised part of raccoon diets during the summer months, but quantitative
data were not available to evaluate their importance in the diet.
Recorded use of man-made structures by raccoons was minimal compared
to other habitats, but these structures may have provided an important
function as winter den/rest, breeding, and nest sites. In this study
several landowners corroborated that wells, barns, haystacks and silage
pits were used by raccoons, especially during the winter season. As
mentioned, vegetational cover is greatly reduced in winter and man-made
structures could easily serve as winter cover. Also, landowner information
on use of open wells by female raccoons with kits indicated that these
afforded greater protection against predators and inclement weather than
natural nesting sites.

CHAPTER VII
TRANSPLANT RACCOON
Trapping and transplanting of raccoons have been conducted by various
state game departments. Removal of raccoons from waterfowl nesting areas,
distribution of raccoons to refuges for sport hunting, or restocking of
raccoons in certain areas are a few reasons why trapping and transplanting
have taken place. However, the behavior of transplanted raccoons has
rarely been reported.
In this study, a JF raccoon was obtained from Clarendon in Donley
County, Texas, 125 km northeast of the Castro study area. Clarendon's
principal industry is ranching, but cultivated fields of wheat and rye
(Secale cereale) do occur (Koerth 1981). Playa basins are not present in
the Clarendon area but year round water is available in spring fed-creeks
and stock tanks.
In May the animal was transported to the Castro study area and kept
in a holding pen for 3 days preceeding release. After release methodology
followed procedures applied to radio-collared native raccoons.
RESULTS
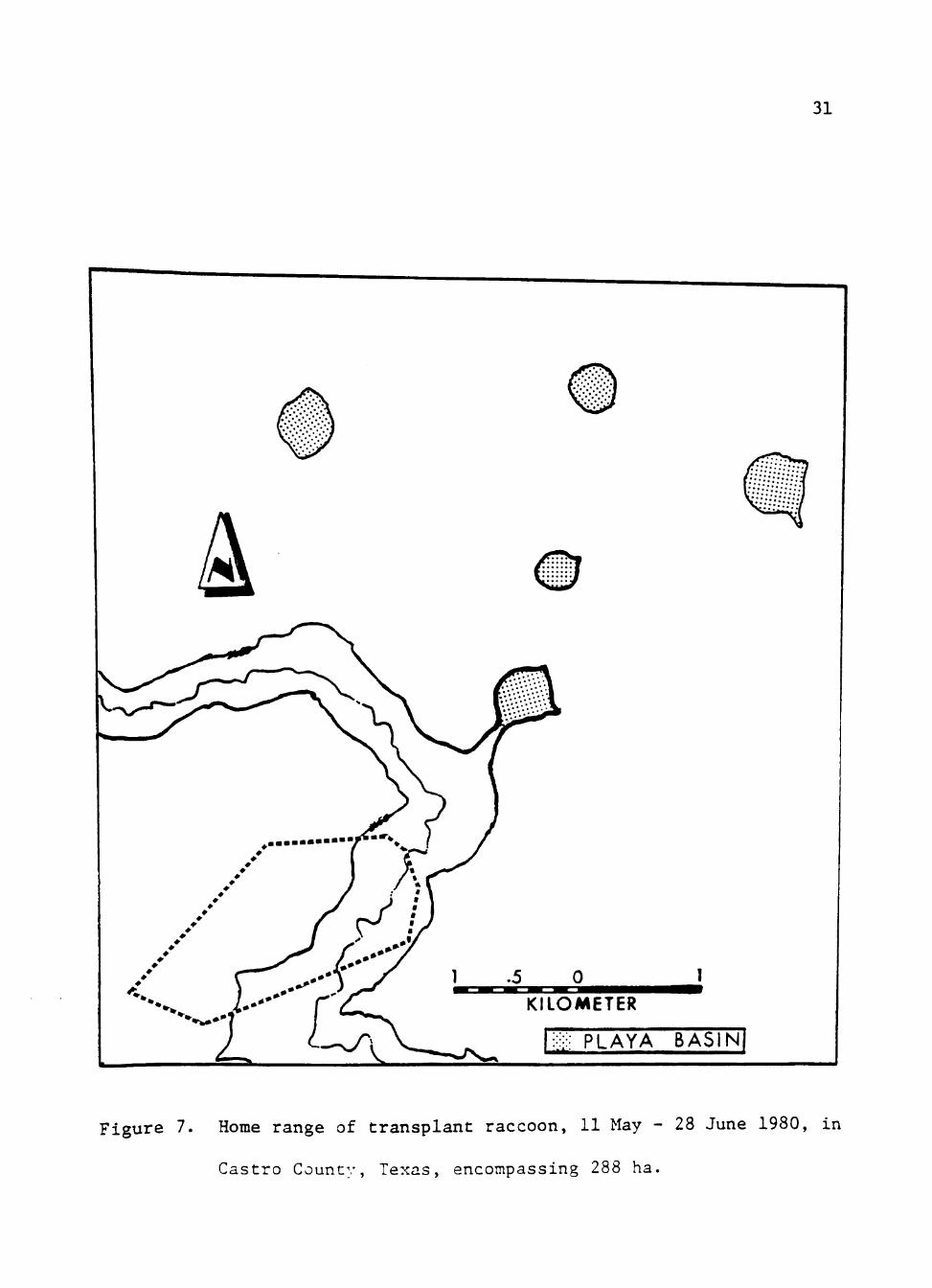
The transplant JF was monitored from 11 May 1980 through 28 June 1980.
The animal was released in an abandoned barn in Running Water Draw to
allow the individual immediate security and cover. Playa basins were
located within 3-5 km from this release site.
The transplanted JF had a monitored home range of 288 ha, which was
smaller than any of the resident raccoon home range areas (Fig. 7). Mean
movement was 2.2 km, ranging from 1.0-5.0 km. This was less than average
movements recorded for native collared raccoons.
Sixteen den/rest sites were located for the transplanted raccoon.
Of these, 75% were located in standing wheat fields, 12.5% in abandoned
burrows, and 12.5% in haystacks. During her monitored period, this animal
denned/rested regularly in the same wheat field until it was harvested.
After harvest 2 sites were recorded in a haystack and 2 more in burrows.
The haystack and burrows were located on pastures in Running Water Draw
close to the wheat field she had previously used. From her release in
30
31
LOMETER
PLAYA BASIN
Figure 7. Home range of transplant raccoon, 11 May - 28 June 1980, in
Castro County, Texas, encompassing 288 ha.

32
April until radio failure in June-, the transplanted raccoon used pastures,
wheat, and cotton fields. Using a Chl-square test of homogeneity, the
expected number of raccoon locations differed significantly (P<0.10) from
the occurrence of habitat within the animal's home range. Preferences for
habitat were calculated as for resident raccoons (Table 3). Pastures and
wheat fields were used in proportion to their availability. Cotton fields
were used less than their availability, the same as for resident raccoons.
SUMMARY
During the monitored period the JF did not visit a playa basin. Most
of her time was spent in standing wheat fields and pastures bordering
Running Water Draw. The small home range recorded for the monitored
period after release, along with relatively restricted movements may
indicate a period of adjustment to the release site. In June 1980, wheat
fields were harvested, removing"; a large portion of cover she had been
using within the monitored home range. Radio contact was lost 1 week
after the wheat fields were harvested.
Although both food and water were available to this animal, the food
base in the study area may have been very different than the JF's endemic
area. In Clarendon, fruits such as wild plum, grapes, and berries were
present during summer, whereas, the study area had no wild furits or
berries. Possible food items that were similar to Clarendon were amphibians,
insects, reptiles, eggs, crustaceans, and invertebrates (B. H. Koerth,
pers. commun.).
The JF spent much of her time in surroundings similar to those found
in Clarendon. Observed den/rest sites in the sides of old silage pits
and wheat fields may reflect denning habits along the river breaks country
and wheat fields in Clarendon.
The cotton fields in which telemetric locations occurred were located
between a wheat field and a tailwater pit next to a wheat field. The 2
recorded locations occurred when the raccoon was traveling to or from the
pit. Avoidance of cotton fields was probably because of the lack of cover
and unfamiliarity with this type (Clarendon area has no cotton).
No further information was obtained on this animal after June until
14 January 1981 when her marked tracks were located on a playa basin

33
w
w eu
•H O (U
m <u CO
§ ^ §§
' ^ « •H 3 MH 4J S c: Q-H O
in CM
•
V» rH
Cb v< o r H
rH CM
•
vl CM
QA
v/ CM i n
CM
•
vl CO
QA
Vl vo t-i
o
I
eu
H
eu
w
1
•
o en
00 CM
in •
in en
en
in o
•
o
•^
in rH ^ * * CN ro
• • •
^ ^
CO VO o
(0
I 1 I
I
cu
I • H MH MH
U
• H MH
§
MH

34
40.3 km northwest of her monitored home range area. Ellis C1964) reported
the movements of an adult male transplanted from Georgia to tlllnois.
Eight months after releas-e the raccoon was found 19.3 km away. Dispersal
of the JF raccoon in this study showed similar behavior. It is difficult
to draw conclusions from a single observation: hovever, it is possible
that when the wheat harvest removed the security cover in the JF's newly
established home range the animal began moving in search of a more
suitable and familiar environment.

LITERATURE CITED
Baker, R. H. , c. C. Newman, and F. Wilke. 1945. Food habits of the
raccoon in eastern Texas. J. Wlldl. Manage. 9Q):45-48.
Berner, A., and L. W. Gysel. 1967. Raccoon use of large tree cavities
and ground burrows. J. Wlldl. Manage. 31(4):706-714.
Bigler, W. J., and G. L. Hoff. 1974. Anesthesia of raccoons with
ketamine hydrochloride. J. Wlldl. Manage. 38(4): 364-366.
Bruns, H. E. 1974. Soil snrvey of Castro County, Texas. Soil Cons.
Serv. 41pp.
Burt, W. H., and R. P. Grossenheider. 1976. A field guide to the
mammals. Houghton Mifflin Co., Boston. 289pp.
Butterfield, R. T. 1944. Populations, hunting pressure, and movement
of Ohio raccoons. Trans. N. Am. Wlldl. Conf. 9:334-337.
Chandler, A. C. 1942. The helminths of raccoons in east Texas. J.
Parasit. 28:255-268.
Cook, R. S., D. 0. Trainer, W. C. Glazener, and B. D. Nassif. 1965.
A serological study of infectious diseases of wild populations in
south Texas. Trans. N. Am. Wlldl. Conf. 30:142-155.
Davis, W. B. 1974. The mammals of Texas. Texas Parks and Wildl. Dept.
Bull. 41. 294pp.
Dorney, R. S. 1954. Ecology of marsh raccoons J. Wildl. Manage. 18(2):
217-225.
Ellis, R. J. 1964. Tracking raccoons by radio. J. Wildl. Manage.
28(2):363-368.
Fleming, D. M. 1975. Movement patterns of the coastal marsh raccoon
in Louisiana and notes on its life history. M. A. Thesis,
Louisiana State Univ. 90pp.
Fritzell, E. K. 1978. Habitat use by prairie raccoon during the
waterfowl breeding season. J. Wildl. Manage. 42(1):118-127.
Frye, J. C , and L. A. Byron. 1957. Studies of cenozoic geology along
eastern margin of Texas High Plains, Armstrong to Howard counties
Univ. Texas Bur. Econ. Geology, Rep. 32. 60pp.
Gels, G. L. 1966. Mobility and behavior of raccoons in eastern South
Dakota. M. S. Thesis, South Dakota State Univ., Brookinas, S. D. 40pp
35

36
Giles, L. W. 1940. Food habits of the raccoon in eastern Iowa. J. Wildl.
Manage. 4(4):375-382.
. 1942. Utilization of rock exposures for dens and escape by
raccoons. Am. Midi. Nat. 27(1):171-176.
Grau, G. A. 1968. Age determination in the raccoon (Procyon lotor).
M. A. Thesis, Univ. Missouri. 78pp.
Hamilton, Jr., W. J. 1951. Warm weather foods of the raccoon in New
York state. J. Mammal. 32(3):341-344.
Harvey, M. J. and R. W. Barbour. 1965. Home range of Microtus ochragaster
as determined by a modified minimum area method. J. Mammal. 46(3):
398-402.
Johnson, A. S. 1970. Biology of the raccoon (Procyon lotor varius Nelson
and Goldman) in Alabama Agri. Exp. Stat. Bull. 402, Auburn Univ.
148pp.
Koerth, B. H. Jr. 1981. Habitat use, herd ecology, and seasonal
movements of mule deer in the Texas panhandle. M. A. Thesis. Texas
Tech Univ. 103pp.
Lotze, J. H. 1979. The raccoon (Procyon lotor) on St. Catherines Islands,
Georgia. 4 comparisons of home ranges determined by live-trapping
and radio-tracking. Am. Mus. Novit. No. 2664. 25pp.
Marcum, L. C. and D. 0. Lof tsgaarden. 1980. A nonmapping technique for
studying habitat preferences. J. Wildl. Manage. 44(4):963-968.
Mech, D. L., J. R. Tester, and D. W. Warner. 1966. Fall daytime resting
habits of raccoons as determined by telemetry. J. Mammal. 47(3):
450-466. and F. J. Turkowski. 1966. Twenty-three raccoons in one
winter den. J. Mammal. 47:529-530
Moore, R. L. 1980. Aspects of the ecology and hunting economics of
migratory waterfowl on the Texas High Plains. M. S. Thesis, Texas
Tech Univ. 80pp. Sanderson, G. C. 1950. Methods of measuring productivity in raccoons.
J. Wildl. Manage. 14(4):389-402.
1961. The lens as an indicator of age in the raccoon.
Amer. Midi. Nat. 65(2):481-485.

37
Schneider, D. G., L. D. Mech., and J. R, Tester. 1971, Movements of
female raccoons and their young as determined by radio tracking,
Anim. Behav. Monogr. 4(1):1-43.
Schnell, J. H. 1970. Rest site selection by radio-tagged raccoons. J.
Minn. Acad. Sci. 36:83-88.
Siniff, D. B., and J. R. Tester. 1965. Computer analysis of animal
movement data obtained by telemetry. Biosci. 15(2):104-108.
Steel, R. G. D., and J. H. Torie. 1978. Principles and proceedures of
statistics a biometrical approach. 2nd edit. McGraw-Hill. New York,
N. Y. 636pp.
Stuewer, F. W. 1943. Raccoons; their habits and management in Michigan.
Ecol. Monogr. 13(2):203-257.
Urban, D. 1970. Raccoon populations, movement patterns, and predation
on a managed waterfowl marsh. J. Wildl. Manage. 34(2):372-383.
Whitney, L. F. , and A. B. Underwood. 1952. The raccoon. Practical Sci.
Publ. Co., Orange Conn. 177pp.
Wood, J. E. 1954. Food habits of furbearers of the Upland Post Oak Region
in Texas. J. Mammal. 35(3):406-415.
1955. Notes on reproduction and rate of increase of raccoons
in the Post Oak Region of Texas. J. Wildl. Manage. 19(3) :409-410.

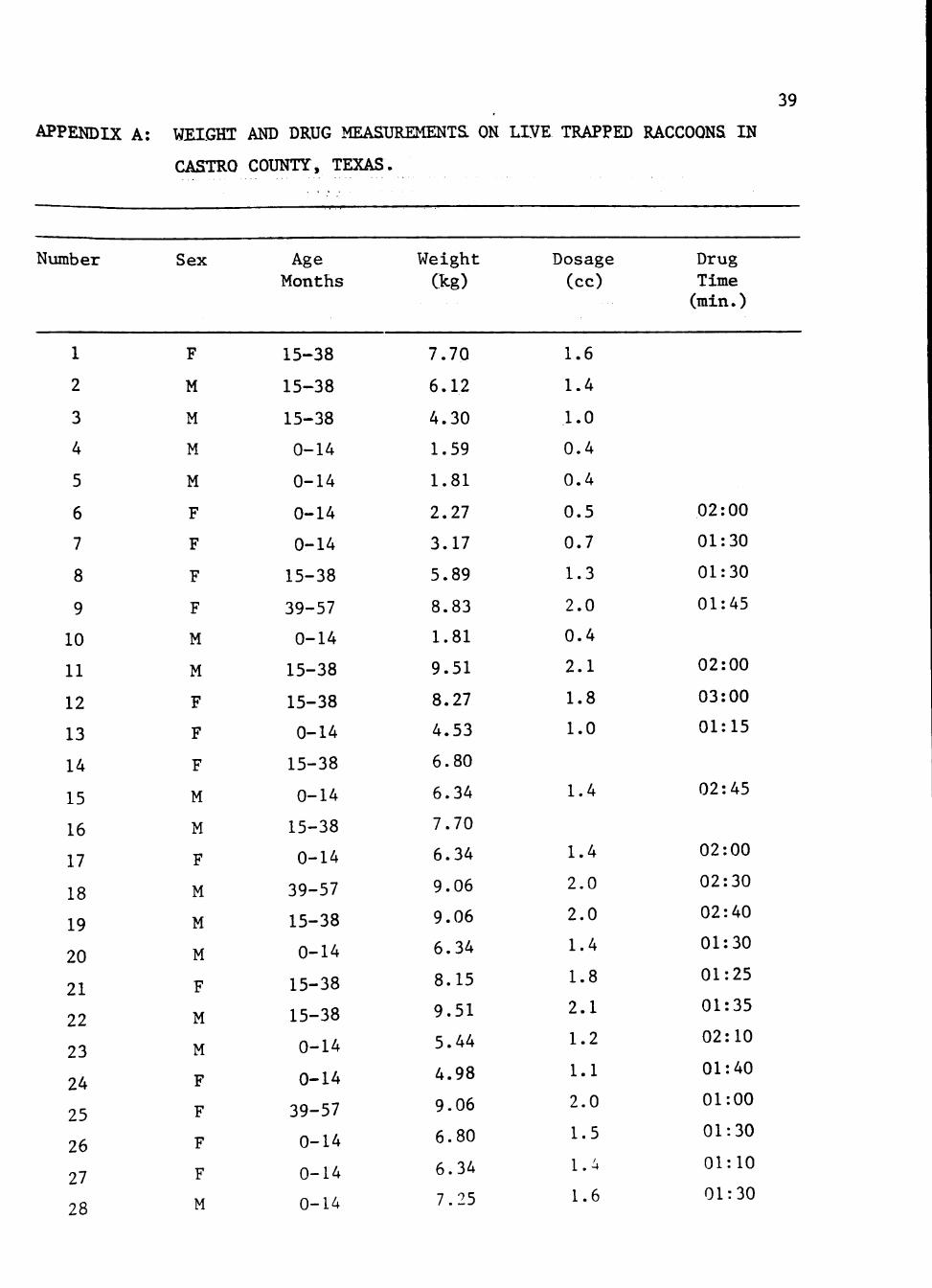
APPENDIX
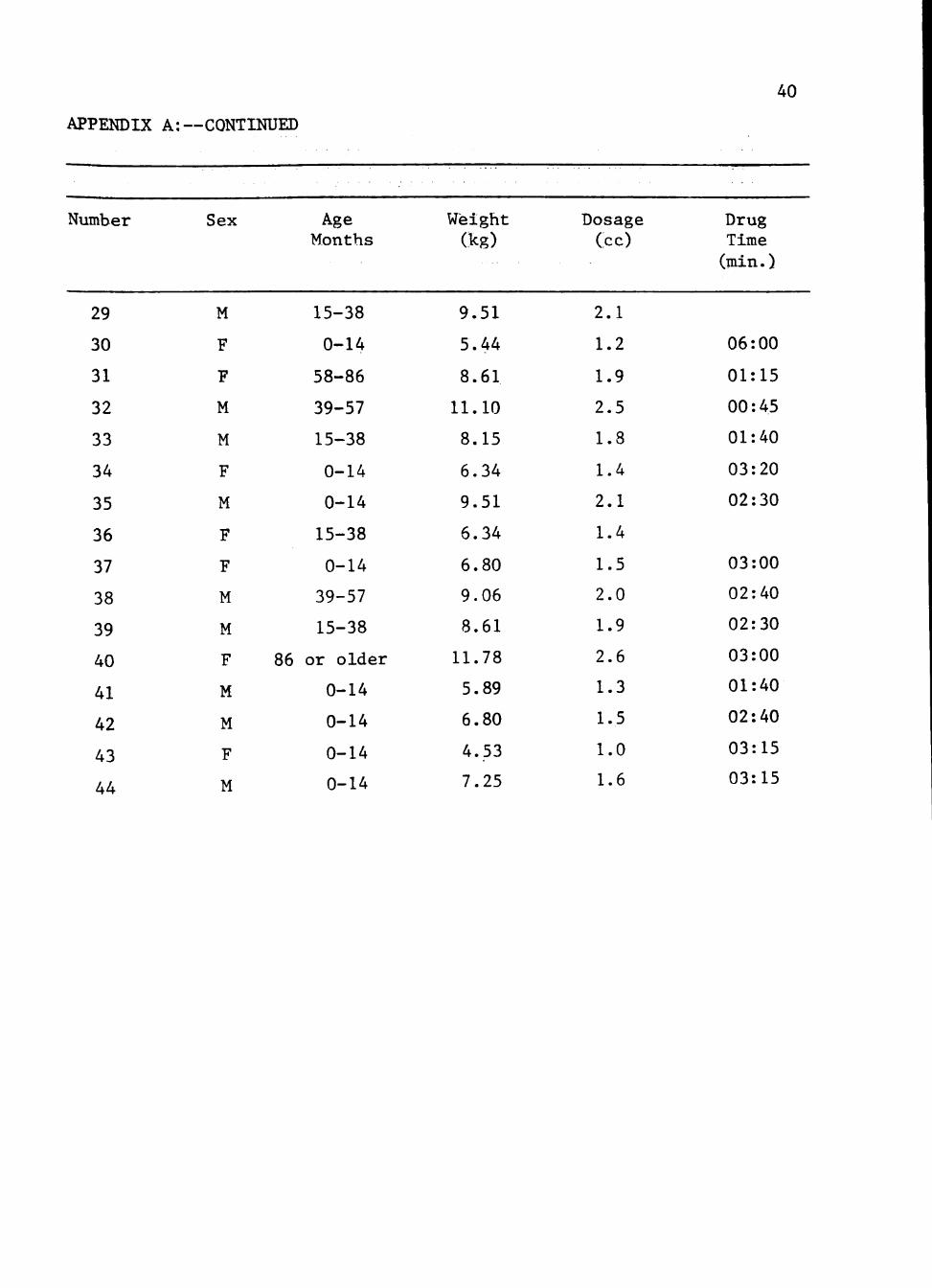
A. Weight and drug measurements on live trapped raccoons in Castro
County, Texas.
38
APPENDIX A: WEIGHT AND DRUG MEASITREMENTS. ON LIVE TRAPPED RACCOONS IN
CASTRO COUNTY, TEXAS.
39
Number
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
Sex
F
M
M
M
M
F
F
F
F
M
M
F
F
F
M
M
F
M
M
M
F
M
M
F
F
F
F
M
Age Months
15-38
15-38
15-38
0-14
0-14
0-14
0-14
15-38
39-57
0-14
15-38
15-38
0-14
15-38
0-14
15-38
0-14
39-57
15-38
0-14
15-38
15-38
0-14
0-14
39-57
0-14
0-14
0-14
Weight
(kg)
7.70
6.12
4.30
1.59
1.81
2.27
3.17
5.89
8.83
1.81
9.51
8.27
4.53
6.80
6.34
7.70
6.34
9.06
9.06
6.34
8.15
9.51
5.44
4.98
9.06
6.80
6.34
7.25
Dosage (cc)
1.6
1.4
1.0
0.4
0.4
0.5
0.7
1.3
2.0
0.4
2.1
1.8
1.0
1.4
1.4
2.0
2.0
1.4
1.8
2.1
1.2
1.1
2.0
1.5
1.4
1.6
Drug Time (min.)
02:00
01:30
01:30
01:45
02:00
03:00
01:15
02:45
02:00
02:30
02:40
01:30
01:25
01:35
02:10
01:40
01:00
01:30
01:10
01:30
APPENDIX A:—CONTINUED
40
Number
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
Sex
M
F
F
M
M
F
M
F
F
M
M
F
M
M
F
M
Age Months
15-38
0-14
58-86
39-57
15-38
0-14
0-14
15-38
0-14
39-57
15-38
86 or older
0-14
0-14
0-14
0-14
Weight (kg)
9.51
5.44
8.61
11.10
8.15
6.34
9.51
6.34
6.80
9.06
8.61
11.78
5.89
6.80
4.53
7.25
Dosage (cc)
2.1
1.2
1.9
2.5
1.8
1.4
2.1
1.4
1.5
2.0
1.9
2.6
1.3
1.5
1.0
1.6
Drug Time (min.)
06:00
01:15
00:45
01:40
03:20
02:30
03:00
02:40
02:30
03:00
01:40
02:40
03:15
03:15