homeobox genes gsx1 and gsx2 differentially - proceedings of the
TRANSCRIPT

Homeobox genes Gsx1 and Gsx2 differentiallyregulate telencephalic progenitor maturationZhenglei Peia, Bei Wanga,1, Gang Chenb, Motoshi Nagaoa, Masato Nakafukua, and Kenneth Campbella,2
aDivision of Developmental Biology and bDivision of Pulmonary Biology, Perinatal Institute, Cincinnati Children’s Hospital Medical Center, University ofCincinnati College of Medicine, Cincinnati OH 45229
Edited by Pasko Rakic, Yale University, New Haven, CT, and approved December 15, 2010 (received for review June 21, 2010)
Homeobox genes Gsx1 and Gsx2 (formerly Gsh1 and Gsh2) areamong the earliest transcription factors expressed in neuronal pro-genitors of the lateral ganglionic eminence (LGE) in the ventraltelencephalon. Gsx2 is required for the early specification of LGEprogenitor cells and recently has been shown to specify differentLGE neuronal subtypes at distinct time points. In Gsx2 mutants,Gsx1 compensates, at least in part, for the loss of Gsx2 in thespecification of LGE neuronal subtypes. Because no specific phe-notype has been described in Gsx1 mutants, it is unclear what rolethis factor plays in the development of the ventral telencephalon.Here, we used a gain-of-function approach to express either Gsx1or Gsx2 throughout the telencephalon and found that Gsx1 func-tions similarly to Gsx2 in the specification of LGE identity. How-ever, our results show that Gsx1 and Gsx2 differentially regulatethe maturation of LGE progenitors. Specifically, Gsx2 maintainsLGE progenitors in an undifferentiated state, whereas Gsx1 pro-motes progenitor maturation and the acquisition of neuronal phe-notypes, at least in part, through the down-regulation of Gsx2.These unique results indicate that the two closely related Gsxgenes similarly regulate LGE patterning but oppositely controlthe balance between proliferation and differentiation in the neu-ronal progenitor pool.
cell cycle kinetics | self-renewal
The proper functioning of the vertebrate CNS depends on thecorrect assembly of neural circuits containing the appropriate
types of neurons produced in the right numbers, at the righttime, and in the right place. In the telencephalon, this assembly isachieved by coordinated control of patterning, proliferation, anddifferentiation of neural progenitors (1). The molecular mech-anisms underlying the orchestration of these processes remainunclear. However, vertebrate homeobox genes, such as Pax6 (2–5) and Emx2 (6–8) have been shown to play important rolesin the control of both patterning and proliferation of dorsaltelencephalic progenitors.Gsx (previously known as Gsh) genes (Gsx1 and Gsx2) rep-
resent a family of homeobox genes, which are expressed in dis-crete progenitor cells of the embryonic brain, including theventral telencephalon where they govern the early specificationof lateral ganglionic eminence (LGE) progenitors (3, 4, 9–15).More specifically, Gsx genes are required for the development ofstriatal projection neurons and olfactory bulb interneurons,which are the two major derivatives of the LGE. Similar to Pax6and Emx2 in the dorsal telencephalon, Gsx genes are not onlyrequired for the patterning of LGE progenitors but also for thecontrol of their proliferative characteristics (3, 4, 14).Despite the fact that both Gsx genes are expressed in the LGE
and the medial ganglionic eminence (MGE), they display largelycomplementary patterns of expression. From embryonic day (E)12.5 and onward, Gsx2 is expressed at a high level in progenitorsof the dorsal LGE (dLGE) and relatively lower level in theventral LGE (vLGE) and MGE progenitors, whereas Gsx1 isexpressed mainly in the MGE and vLGE (4, 10, 14–18). Thegraded Gsx2 expression pattern in LGE progenitors has recentlybeen implicated in the distinct neuronal output of the dLGE
versus the vLGE (17). In Gsx2 mutants, the expression of Gsx1expands throughout the dorsal extent of the LGE (14, 18). De-spite this, however, Gsx1 only partially compensates for the lossof Gsx2 in the development of the Gsx2 mutant striatum andolfactory bulb. To date, no specific telencephalic defects havebeen reported in Gsx1 mutants (14, 18, 19) and thus the re-lationship between Gsx1 and Gsx2 function in the developingtelencephalon remains unclear. In this study, we have takena gain-of-function approach to uncover distinct roles for Gsx1and Gsx2 in regulating patterning and maturation of telence-phalic progenitors.
ResultsGsx1 Is Localized to a Subset of Telencephalic Progenitor Cells. Un-like Gsx2 (3, 17, 18, 20), Gsx1 protein has never been localizedspecifically in telencephalic progenitors due to lack of a well-characterized antibody. Thus, we obtained Gsx1-EGFP BACtransgenic mice from GENSAT (www.gensat.org) and charac-terized the EGFP-expressing cells using antibodies that recog-nize either Gsx2 (3) or Gsx1 and -2 (12) at various embryonicstages. At E12.5, EGFP staining in Gsx1-EGFP embryos wasmost prevalent in the subventricular zone (SVZ) and mantleregions of the MGE (Fig. 1 A and B). This expression likelyrepresents perdurance of the EGFP because Gsx2 or Gsx1/2staining is largely confined to the ventricular zone (VZ) (Fig. 1 Aand B). Indeed, scattered EGFP+ cells were observed in theventricular zone (VZ) of both the vLGE and MGE with a con-centration near the VZ/SVZ boundary (Fig. 1 A and B). Veryfew of these EGFP+ VZ cells were observed to coexpress Gsx2at any of the embryonic stages examined (Fig. 1 A, C, D, and H).However, Gsx1/2 immunostaining revealed double labeling withEGFP specifically in progenitor cells that were mostly concen-trated around the VZ/SVZ boundary of the vLGE (and MGE) atE12.5 (Fig. 1B) as well as at later stages (Fig. 1 E, F, I, and J).Thus, whereas Gsx2 is detected in most VZ cells throughout theapical–basal axis of the LGE and MGE, Gsx1 marks a sub-population of cells clustered basally at the VZ/SVZ boundary.
Gsx2 Expression in the Gsx1 Mutant Telencephalon. Previous studieshave shown that Gsx2 is expressed in a high dorsal to lowventral gradient in LGE VZ cells (4, 17) (see also Fig. 1 A, C,and G). It remains unclear, however, what mechanism(s)functions to establish and maintain the gradient of Gsx2 ex-pression along the dorsal–ventral axis. The inverse relationshipbetween cells expressing high levels of Gsx2 and those in whichEGFP is driven from the Gsx1 BAC (Fig. 1) might suggest that
Author contributions: Z.P., M. Nakafuku, and K.C. designed research; Z.P. performed re-search; B.W., G.C., M. Nagao, and M. Nakafuku contributed new reagents/analytic tools;Z.P., M. Nakafuku, and K.C. analyzed data; and Z.P. and K.C. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1Present address: Genentech, 1 DNA Way, South San Francisco, CA 94080.2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1008824108/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1008824108 PNAS | January 25, 2011 | vol. 108 | no. 4 | 1675–1680
NEU
ROSC
IENCE
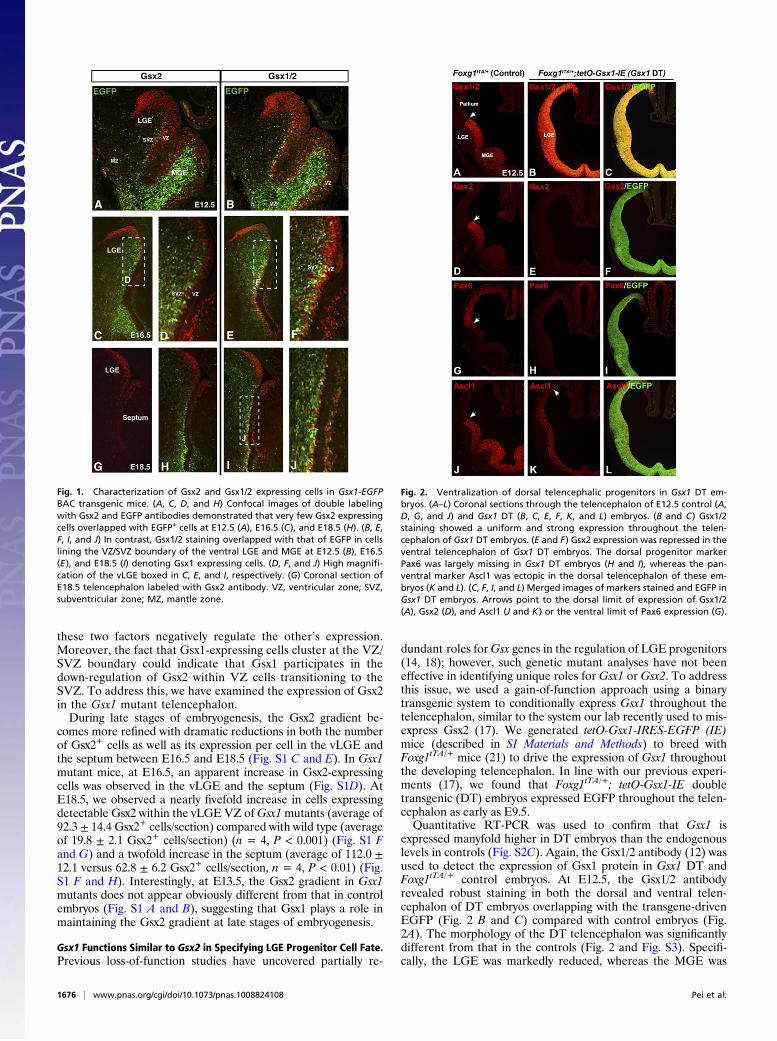
these two factors negatively regulate the other’s expression.Moreover, the fact that Gsx1-expressing cells cluster at the VZ/SVZ boundary could indicate that Gsx1 participates in thedown-regulation of Gsx2 within VZ cells transitioning to theSVZ. To address this, we have examined the expression of Gsx2in the Gsx1 mutant telencephalon.During late stages of embryogenesis, the Gsx2 gradient be-
comes more refined with dramatic reductions in both the numberof Gsx2+ cells as well as its expression per cell in the vLGE andthe septum between E16.5 and E18.5 (Fig. S1 C and E). In Gsx1mutant mice, at E16.5, an apparent increase in Gsx2-expressingcells was observed in the vLGE and the septum (Fig. S1D). AtE18.5, we observed a nearly fivefold increase in cells expressingdetectable Gsx2 within the vLGE VZ ofGsx1mutants (average of92.3 ± 14.4 Gsx2+ cells/section) compared with wild type (averageof 19.8 ± 2.1 Gsx2+ cells/section) (n = 4, P < 0.001) (Fig. S1 Fand G) and a twofold increase in the septum (average of 112.0 ±12.1 versus 62.8 ± 6.2 Gsx2+ cells/section, n = 4, P < 0.01) (Fig.S1 F and H). Interestingly, at E13.5, the Gsx2 gradient in Gsx1mutants does not appear obviously different from that in controlembryos (Fig. S1 A and B), suggesting that Gsx1 plays a role inmaintaining the Gsx2 gradient at late stages of embryogenesis.
Gsx1 Functions Similar to Gsx2 in Specifying LGE Progenitor Cell Fate.Previous loss-of-function studies have uncovered partially re-
dundant roles forGsx genes in the regulation of LGE progenitors(14, 18); however, such genetic mutant analyses have not beeneffective in identifying unique roles for Gsx1 or Gsx2. To addressthis issue, we used a gain-of-function approach using a binarytransgenic system to conditionally express Gsx1 throughout thetelencephalon, similar to the system our lab recently used to mis-express Gsx2 (17). We generated tetO-Gsx1-IRES-EGFP (IE)mice (described in SI Materials and Methods) to breed withFoxg1tTA/+ mice (21) to drive the expression of Gsx1 throughoutthe developing telencephalon. In line with our previous experi-ments (17), we found that Foxg1tTA/+; tetO-Gsx1-IE doubletransgenic (DT) embryos expressed EGFP throughout the telen-cephalon as early as E9.5.Quantitative RT-PCR was used to confirm that Gsx1 is
expressed manyfold higher in DT embryos than the endogenouslevels in controls (Fig. S2C). Again, the Gsx1/2 antibody (12) wasused to detect the expression of Gsx1 protein in Gsx1 DT andFoxg1tTA/+ control embryos. At E12.5, the Gsx1/2 antibodyrevealed robust staining in both the dorsal and ventral telen-cephalon of DT embryos overlapping with the transgene-drivenEGFP (Fig. 2 B and C) compared with control embryos (Fig.2A). The morphology of the DT telencephalon was significantlydifferent from that in the controls (Fig. 2 and Fig. S3). Specifi-cally, the LGE was markedly reduced, whereas the MGE was
Fig. 1. Characterization of Gsx2 and Gsx1/2 expressing cells in Gsx1-EGFPBAC transgenic mice. (A, C, D, and H) Confocal images of double labelingwith Gsx2 and EGFP antibodies demonstrated that very few Gsx2 expressingcells overlapped with EGFP+ cells at E12.5 (A), E16.5 (C), and E18.5 (H). (B, E,F, I, and J) In contrast, Gsx1/2 staining overlapped with that of EGFP in cellslining the VZ/SVZ boundary of the ventral LGE and MGE at E12.5 (B), E16.5(E), and E18.5 (I) denoting Gsx1 expressing cells. (D, F, and J) High magnifi-cation of the vLGE boxed in C, E, and I, respectively. (G) Coronal section ofE18.5 telencephalon labeled with Gsx2 antibody. VZ, ventricular zone; SVZ,subventricular zone; MZ, mantle zone.
Fig. 2. Ventralization of dorsal telencephalic progenitors in Gsx1 DT em-bryos. (A–L) Coronal sections through the telencephalon of E12.5 control (A,D, G, and J) and Gsx1 DT (B, C, E, F, K, and L) embryos. (B and C) Gsx1/2staining showed a uniform and strong expression throughout the telen-cephalon of Gsx1 DT embryos. (E and F) Gsx2 expression was repressed in theventral telencephalon of Gsx1 DT embryos. The dorsal progenitor markerPax6 was largely missing in Gsx1 DT embryos (H and I), whereas the pan-ventral marker Ascl1 was ectopic in the dorsal telencephalon of these em-bryos (K and L). (C, F, I, and L) Merged images of markers stained and EGFP inGsx1 DT embryos. Arrows point to the dorsal limit of expression of Gsx1/2(A), Gsx2 (D), and Ascl1 (J and K) or the ventral limit of Pax6 expression (G).
1676 | www.pnas.org/cgi/doi/10.1073/pnas.1008824108 Pei et al.

essentially missing in all Gsx1 DT embryos. Indeed, Nkx2.1 ex-pression in these embryos was never observed to be ectopic andwas drastically reduced or missing (Fig. S3D), similar to that inGsx2 DT embryos (17). Remarkably, in all Gsx1 DT embryos,Gsx2 expression was essentially lost (Fig. 2 E and F and Fig. S2D)compared with control embryos (Fig. 2D and Fig. S2D). Thisresult indicates that Gsx1 dramatically down-regulates Gsx2 ex-pression in ventral telencephalic progenitors. Interestingly, inGsx2 DT embryos, the expression of Gsx1 mRNA in the ventraltelencephalon was significantly reduced (Fig. S2 A and B), whichis in accordance with the observed opposing gradients of Gsx2and Gsx1-EGFP in the LGE (Fig. 1).Our recent work (17) found that misexpression of Gsx2
induces ventral telencephalic identity in the telencephalon by up-regulating pan-ventral markers, such as Ascl1 and Dlx (Dlx1, -2,-5, and -6) proteins, while repressing dorsal telencephalic regu-lators, such as Pax6 and Tbr1. In Gsx1 DT embryos, the ex-pression of Pax6 (Fig. 2 H and I) and Tbr1 (Fig. S3F) wasconsiderably repressed in the dorsal telencephalon, comparedwith controls (Fig. 2G and Fig. S3E). Furthermore, Ascl1 (Fig. 2K and L) and pan-Dlx (Fig. S3B) were found ectopicallyexpressed in the dorsal telencephalon of Gsx1 DT embryos,unlike the case in controls (Fig. 2J and Fig. S3A). Thus, Gsx1misexpression confers generic ventral telencephalic identitysimilar to Gsx2. However, unlike the case in Gsx2 DT embryos(17), the VZ of Gsx1 DT embryos, as marked by Ascl1 (Fig. 2K),pan-Dlx (Fig. S3B), and Sox2 (Fig. S3H) is thinner than itscounterpart in controls (Fig. 2J and Fig. S3 A and G).Previous studies have shown that the lateral ganglionic emi-
nence (LGE) contains two distinct progenitor domains, the dLGE,which gives rise to olfactory bulb interneurons and the vLGE,which generates striatal projection neurons (4, 22). The zincfinger transcription factor Sp8 (23) and the LIM homeoboxprotein Islet-1 (Isl1) (22) are markers of dLGE and vLGEderivatives, respectively. We recently demonstrated that Gsx2specifies vLGE (i.e., Isl1) identity at early stages of telencephalicneurogenesis and dLGE (i.e., Sp8) cell fates at later time points(17). To test whether Gsx1 might play a similar temporal role inregulating vLGE versus dLGE fates, we examined Isl1 and Sp8expression, respectively, inGsx1 DT embryos at E14.5. Similar tothe misexpression results with Gsx2 (17), we found that early(i.e., E9.5 onward) misexpression of Gsx1 resulted in dorsal ex-pansion of the vLGE marker Isl1 with only scattered cellsexpressing the dLGE factor, Sp8 (Fig. S4 B and D). As dem-onstrated in our previous Gsx2 misexpression study (17), we wereable to repress Gsx1 transgene activation until E13, by admin-istering doxycycline (Dox) to the pregnant dam in the drinkingwater from E7–9. Dox-treated Gsx1 DT embryos examined atE15.5 showed a severe reduction in Isl1 expression within thevLGE and a concomitant expansion of Sp8 expression bothwithin the dorsal telencephalon as well as within the vLGE (Fig.S4 F and H). These results are similar to what is observed inDox-treated Gsx2 DT embryos (17), albeit less robust. Thus, inagreement with earlier mutant analyses, our gain-of-functionresults indicate that Gsx1 specifies LGE progenitors similarto Gsx2.
Gsx1 and Gsx2 Differentially Regulate Progenitor Maturation andProliferation. Given the distinct expression patterns exhibited byGsx1 and Gsx2 in ventral telencephalic progenitors, we won-dered whether each of these factors plays a unique role in thematuration of LGE progenitors. During telencephalic neuro-genesis, two proliferative populations exist. The primary pro-liferative population (24, 25) emerges in the VZ and asdevelopment proceeds, a secondary proliferative population (26,27) that occupies the SVZ arises, which adjoins the VZ. In thetelencephalon, both VZ and SVZ cells are mitotically activeprogenitors (25, 26). However, SVZ cells are progeny of VZ cells
(28, 29) and are thought to be more restricted in their de-velopmental potential. In Gsx2 mutants, the SVZ does not formproperly at early stages (i.e., E12.5) (3, 4, 14); however, at laterdevelopmental stages, Gsx1 expands throughout the entireGsx2 mutant LGE and this is concomitant with the reappear-ance of a proliferative SVZ (14, 18). Thus, the restricted ex-pression of Gsx1 at the VZ/SVZ boundary may function topromote the maturation of VZ progenitors to an SVZ andultimately neuronal fate.To test this hypothesis, we first examined the expression of
β-III tubulin, an early neuronal differentiation marker, in Gsx1and Gsx2 DT embryos. We limited our analysis to the dorsaltelencephalon, because neither of the Gsx genes is endogenouslyexpressed there and both factors can effectively ventralize theseprogenitors (Fig. 2, Fig. S3, and ref. 17). At E12.5 in controls,β-III tubulin staining was confined to the mantle zone through-out the dorsal telencephalon (Fig. 3B) with very few labeled cellsin the germinal zones. Misexpression of Gsx2 in the dorsal tel-encephalon clearly reduced the intensity of β-III tubulin stainingat this stage (Fig. 3A). Conversely, misexpression of Gsx1 dra-matically increased β-III tubulin staining in the mantle region(Fig. 3C) and even resulted in the appearance of many β-IIItubulin+ cells within the VZ. These results, therefore, suggest
Fig. 3. In vivo cell cycle kinetics studies in Gsx1 or Gsx2 DT embryos. (A–C)β-III tubulin staining of coronal sections of E12.5 dorsal telencephalon.Compared with control (B), Gsx2 misexpression reduced β-III tubulin staining(A), whereas Gsx1 misexpression dramatically increased β-III tubulin staining(C). Note, the precocious β-III tubulin+ cells in the cortical VZ of Gsx1 mis-epxressing embryos (C). Insets in A–C are higher magnification of dashedboxes. (D–I) Pregnant females were exposed to a single pulse of BrdU andchased either 30 min (D–F) or 24 h (G–I). Sections were stained with anti-bodies to Ki67 and BrdU. (F) Quantification of the cell cycle length (i.e., 30-min BrdU labeling) experiment shown in D and E at three different ros-trocaudal levels of the dorsal telencephalon. (I) Quantification of the cellcycle retention (i.e., 24-h BrdU labeling) experiment shown in G and H atthree different levels of the dorsal telencephalon. Data shown in F and Irepresent the mean ± SEM (n = 3); *significant compared with controls atP < 0.01 (one-way ANOVA, Tukey HSD post hoc test). CP, cortical plate; VZ,ventricular zone.
Pei et al. PNAS | January 25, 2011 | vol. 108 | no. 4 | 1677
NEU
ROSC
IENCE

that Gsx2 limits neurogenesis, whereas Gsx1 enhances this pro-cess in telencephalic progenitors.During neural development, the proliferation and differenti-
ation of neuronal progenitors must be tightly balanced throughthe regulation of the cell cycle (1, 5, 30–32) and cell survival (33)to ensure the generation of appropriate neuronal fates in theright time at the right place. No changes in cell death, asdetected by activated caspase 3 staining, were detected in Gsx1or Gsx2 DT embryos at E12.5 (Fig. S5).Cell cycle lengthening has been closely correlated with the
maturation of neuronal progenitor cells and enhanced neuro-genesis (31). During neocortical neurogenesis, cell cycle durationis lengthened from about 8 h at the beginning of neurogenesis tonearly 20 h at the end (34). To determine whether misexpressionof Gsx1 or Gsx2 might cause changes in cell cycle length, weperformed a short-term (i.e., 30 min) BrdU pulse together withstaining for the cycling progenitor marker Ki67 (see ref. 32). Cellcycle length index was determined as the percentage of neuronalprogenitors that incorporated BrdU (BrdU+Ki67+/Ki67+) dur-ing 30 min of labeling. Reductions in the cell cycle length indexare indicative of cell cycle lengthening. Ectopic expression ofGsx1 in the dorsal telencephalon led to a significant reduction inthe cell cycle length index (% BrdU+Ki67+/Ki67+) from ≈45%in controls to about 30% at each rostrocaudal level examined(n= 3, P < 0.01) (Fig. 3F). Conversely, misexpression of Gsx2 hadno significant effect on cell cycle length index at any of the dorsaltelencephalic levels examined (Fig. 3F). Of note, BrdU staining incontrol embryos revealed the typical basal localization of labelednuclei in S phase (Fig. 3D) in the developing cortex (35). However,in Gsx1 double transgenic embryos, BrdU+ nuclei were positionedrandomly throughout the apical–basal extent of the germinal zone,supporting a deregulation of cell cycle duration (Fig. 3E). Thus,misexpression of Gsx1 in dorsal telencephalic progenitors signifi-cantly increased their cell cycle length.To examine the balance between cell cycle retention and exit,
we performed 24 h BrdU pulse-chase experiments together withKi67 staining (see ref. 32). The cell cycle retention index wasdetermined as the percentage of BrdU+ cells that still retainedthe expression of Ki67 (BrdU+Ki67+/BrdU+) 24 h after a BrdUpulse. A high retention index indicates that more cells remain ascycling progenitors. Gsx2 misexpression caused a significant in-crease of the cell cycle retention index (Fig. 3 G–I). In thecontrol dorsal telencephalon, the cell cycle retention indexranged from 32% in rostral regions to 27% in more caudalregions compared with 45% at rostral levels and 35% at caudallevels in the Gsx2-expressing embryos (n = 3, P < 0.01) (Fig. 3I).In this case, Gsx1 misexpression showed no difference in the cellcycle retention index at any of the levels examined (Fig. 3I).Indeed, in control embryos, many BrdU+ Ki67− cells were seenin the developing cortical plate (Fig. 3G), which are likely to bepostmitotic cells undergoing further neuronal differentiation. InGsx2 DT embryos, however, the number of BrdU+ Ki67− cells inthe region of the cortical plate was severely reduced, with mostBrdU+ cells remaining in the VZ (Fig. 3H). In summary, Gsx1misexpression leads to a lengthening of the cell cycle in telen-cephalic progenitors thus promoting neurogenesis (31), whereasmisexpression of Gsx2, at least partially, maintains cells as di-viding progenitors, thus expanding the progenitor pool.
In Vitro Differentiation Assay and Clonal Analysis of ProgenitorsMisexpressing Gsx1 and Gsx2. To further address the roles of Gsx1and Gsx2 in the maintenance of the progenitor pool, we gener-ated neurosphere cultures from control, Gsx1, and Gsx2 DTembryos at E12.5. As was the case in vivo, only neurospheresgrown from DT embryos expressed the transgene-driven EGFP(Fig. 4 A and B).To examine neuronal differentiation, primary neurospheres
were dissociated to single cells and grown in differentiation
medium for 5 d. Sox2+ staining was used to identify the neuralstem/progenitor cell population, whereas neuronal productionwas detected by β-III tubulin staining. Compared with controls,Gsx2-expressing cells showed a significant reduction in neuronaldifferentiation in both the cortical (−40%; Fig. 4 D, F, and G)and LGE cultures (−30%; Fig. 4H) (n = 3, P < 0.01). Moreover,the proportion of Sox2+ cells remaining in the differentiationcultures was higher in the Gsx2-expressing cultures than in thecontrols from either the cortex (+20%; Fig. 4 C, E, andG) or theLGE (+19%; Fig. 4H) (n = 3, P < 0.01). Unlike the Gsx2-expressing cultures, those expressing Gsx1 did not show anychange in the proportion of β-III tubulin+ neurons or Sox2+ cellscompared with controls (Fig. S6).Our in vivo data demonstrated that misexpression of Gsx2 (but
not Gsx1) increases the number of progenitors that remain in the
Fig. 4. In vitro neurosphere assays. Compared with control (A), spheresderived from the cortex of E12.5 Gsx2 DT embryos expressed EGFP (B). Insetsin A and B are phase contrast images of the primary sphere cultures. Sox2 (Cand E) and β-III tubulin (D and F) staining of in vitro differentiated cellsderived from control (C and D) and Gsx2 DT cortices (E and F). (G and H)Quantification of β-III tubulin+ and Sox2+ cells in differentiation culturesderived from the cortex (G) and the LGE (H) of control and Gsx2 DT embryos.Data shown are the mean ± SEM (n = 3); Student’s t test, Gsx2 DT cortex β-IIItubulin/DAPI: P < 0.001; Gsx2 DT cortex Sox2/DAPI: P < 0.001; Gsx2 DT LGEβ-III tubulin/DAPI: P < 0.01; Gsx2 DT LGE Sox2/DAPI: P < 0.01. (I) Quantifi-cation of secondary sphere formation in clonal assays. Values were expressedas percentage of control set to 100%. Data shown represent the mean ±SEM (n = 3 for Gsx2 DT; n = 5 for Gsx1 DT); Student’s t test, Gsx2 DT cortex:P < 0.05; Gsx1 DT cortex: P < 0.05; Gsx1 DT LGE: P < 0.01. (J) A model showingthe proposed roles of Gsx genes in the maturation of LGE progenitor cells.
1678 | www.pnas.org/cgi/doi/10.1073/pnas.1008824108 Pei et al.

cell cycle, suggesting that Gsx2 might function, in part, tomaintain neural stem/progenitor identity in LGE cells. The ca-pacity for self-renewal is a hallmark of neural stem/progenitor cells(36). To examine self-renewal of neurospheres expressing eitherGsx1 or Gsx2, single cell suspensions were prepared from primaryneurospheres at a clonal density and cultured until secondaryspheres were formed (37). Compared with control cultures, Gsx2misexpression enhanced the formation of clonally generated sec-ondary spheres that were derived from the double transgeniccortex by 243% from controls (n= 3, P < 0.05); however, the LGEcultures overexpressing Gsx2 did not show a significant change inthe formation of secondary spheres (Fig. 4I). In contrast to Gsx2,expression of Gsx1 significantly reduced secondary sphere for-mation in cultures that were derived from both the DT cortex(77% of control; n = 5, P < 0.05) and LGE (66% of control; n =5, P < 0.01) (Fig. 4I). These in vitro results, therefore, indicate thatGsx1 and Gsx2 oppositely regulate the self-renewal capacity oftelencephalic progenitors and provide further evidence thatthese two factors differentially regulate the balance betweenproliferation and differentiation in telencephalic progenitors.
DiscussionIn this study, we provide unique data concerning the cellularexpression of Gsx1 in progenitors of the ventral telencephalon.Our data indicate that Gsx1 and Gsx2 exhibit complementaryexpression patterns on two different axes, namely the apical–basal and dorsal–ventral axes. Here we show that Gsx1 is foundin a subset of progenitors concentrated basally at the VZ/SVZborder of the vLGE and MGE. Additionally, Gsx1 expressionappears to be complementary to the previously described highdorsal to low ventral expression of Gsx2 within the LGE (4, 17).Although previous loss-of-function studies have indicated par-tially redundant roles for Gsx genes in the specification of LGEprogenitors (14, 18), the distinct expression patterns revealedhere suggest that unique roles for Gsx1 and Gsx2 in the regu-lation of LGE progenitor development may exist. Using a gain-of-function approach, we show that Gsx1 can regulate LGEspecification in a manner similar to that previously observed byGsx2 (17). However, our data also revealed that Gsx1 and Gsx2regulate progenitor maturation oppositely, with Gsx2 maintain-ing telencephalic progenitors in an undifferentiated state andGsx1 promoting progenitor maturation toward neurogenesis.The expression of Gsx2 at both the mRNA and protein level
has been thoroughly examined (3, 17, 18, 20). However, thespecific cellular expression of Gsx1 protein has not been de-scribed due to the lack of a well-characterized antibody. Ourprevious study found that Gsx1, as detected by Gsx1/2 antibody(12), is expressed by a subset of progenitors in the Gsx2 mutantLGE, which are concentrated basally at the VZ/SVZ boundary(16). Using Gsx1-EGFP BAC transgenic mice (together with theGsx1/2 antibody), we show here that, even in wild-type embryos,Gsx1-expressing progenitors of the vLGE and MGE appear to beclustered at the VZ/SVZ boundary. This is quite distinct fromGsx2, which is expressed to some level in most cells throughoutthe apical–basal extent of the VZ. Interestingly, Gsx1 isexpressed in regions of the ventral telencephalon where Gsx2 isfound at moderate to low levels (i.e., vLGE and MGE). More-over, our findings indicate that the loss of Gsx1 gene functionresults in increased numbers of Gsx2 cells within the VZ of thevLGE, at late stages of embryogenesis. This is in line with ourresults showing that overexpression of Gsx1 throughout the tel-encephalon essentially abolishes Gsx2 expression and suggeststhat Gsx1 may function as a repressor of Gsx2 gene expression.Thus the Gsx2 gradient within the LGE, at least at late stages ofembryogenesis, appears to be maintained by Gsx1. Moreover,the cross-talk between Gsx genes may play essential roles in theultimate differentiation capacity of LGE progenitors.
Some redundancy between Gsx1 and Gsx2 gene function in thespecification of the ventral telencephalic identity has previouslybeen suggested by studies in Gsxmutants (14, 18). Our results here,using a gain-of-function approach, support these findings and in-dicate that ectopic Gsx1 imparts ventral identity on dorsal telence-phalic progenitors in a manner very similar to that of Gsx2 (17).Moreover, misexpression of Gsx1 appeared to promote vLGE anddLGE specification with a similar temporal profile to that recentlyshown inGsx2 gain-of-function studies (17). It is not surprising, how-ever, that Gsx1 would regulate patterning similar to Gsx2 becausetheir consensus DNA binding sites are quite similar (9, 10).The apparent accumulation of Gsx1-expressing progenitors at
the VZ/SVZ boundary suggests that this factor may participate inthe transition of VZ cells to the SVZ, ultimately leading to theirdifferentiation within the mantle. In accordance with this notion,misexpression of Gsx1 led to an increase in neurogenesis in vivo.We did not see an increase in neurogenesis within Gsx1-expressing cultures; however, this assay may be too rapid to ac-curately determine the effect of Gsx1, in vitro. In fact, the numberof neurochemically defined neurons in the control cultures wasalready maximal at 2 d in vitro. Conversely, overexpression ofGsx2 reduced neurogenesis both in vivo and in vitro. It should bementioned, however, that the levels of Gsx1 and Gsx2 in the re-spective DT embryos are considerably higher than their endoge-nous levels (Fig. S2) and thus could result in nonphysiologicalconsequences. However, we do not believe that this is the case,because Gsx proteins bind to similar DNA sequences (9, 10) andyet the overexpression of Gsx1 and Gsx2 leads to opposite effectson progenitor maturation. These opposing roles for Gsx genes inLGE progenitors are in line with the observed phenotype inGsx2mutants (14, 18). Although Gsx1 can restore the molecularidentity of progenitors within the Gsx2 mutant LGE, it remainsconsiderably smaller than a wild-type LGE and the resultingstriatum is less than half the normal size (18). This suggests thatGsx1 can specify LGE progenitors similar to Gsx2 but that it doesnot expand the LGE progenitors to the same extent.In fact, the first evidence suggesting that Gsx genes differen-
tially control LGE progenitor maturation was that Gsx1 DTembryos exhibited less prominent ventral elevations (i.e., MGEand LGE) and a thinner telencephalic VZ, unlike that observedin Gsx2 DT embryos (17) and controls. Our gain-of-functionapproach was necessary to uncover this unique role for Gsx1,because Gsx1 mutants do not display overt telencephalic phe-notypes (14, 18, 19). It must be, therefore, that other factors cansubstitute for the absence of Gsx1 in driving the differentiationof LGE progenitors. In this respect, Ascl1 (Mash1), which isknown to play an important role in LGE progenitor maturation(38, 39), is required for the partial compensatory role of Gsx1 inthe Gsx2 mutant telencephalon (16).To uncover the cellular mechanisms underlying the differen-
tial regulation of progenitor maturation by Gsx genes, we used invivo cell cycle kinetic studies to examine cell cycle length and exitas well as in vitro neurosphere assays for self-renewal capacity.Our data show that Gsx1 misexpression leads to lengthening ofthe cell cycle, whereas Gsx2 misexpression had no effect on thisparameter. Cell cycle lengthening correlates well with progenitormaturation during neocortical neurogenesis (31, 34). A longercell cycle is thought to increase the time during which cell fatedeterminants can act, particularly in G1, thereby increasing thechance that the cell will undergo neurogenesis versus remainingas a cycling progenitor (31). Thus the increased cell cycle lengthinduced by misexpression of Gsx1, correlates well with the en-hanced neurogenesis observed.Cell cycle retention is obviously an important characteristic of
neural stem/progenitor cells and an essential aspect of generat-ing the correct number of differentiated progeny (e.g., neurons)or the appropriate size of brain structures (32, 36). Unlike Gsx1,Gsx2 misexpression decreased the proportion of cells exiting the
Pei et al. PNAS | January 25, 2011 | vol. 108 | no. 4 | 1679
NEU
ROSC
IENCE

cell cycle. Our in vitro results using neurosphere culture dem-onstrate opposing effects of Gsx1 and Gsx2 on the self-renewalcapacity of telencephalic progenitors. Whereas Gsx2 increasedthe self-renewal capacity of neurospheres derived from thedorsal telencephalon (but not the LGE where Gsx2 is endoge-nously expressed), Gsx1 significantly reduced this measure inboth cortical and LGE cultures. Nagao et al. (37) have recentlyshown that overexpression of the protooncoprotein Myc in-creased self-renewal in telencephalic progenitors. In Drosophila,the Gsx homolog Ind, is a direct target of dMyc in intermediateneuroblasts (40). Thus Myc may regulate self-renewal in LGEprogenitors, at least in part, through the up-regulation of Gsx2.Our results demonstrate that Gsx1 and Gsx2 oppositely regulate
telencephalic progenitor maturation. This differential regulation isin concordance with the negative cross-regulation that Gsx1 andGsx2 exhibit with each other. In this manner, high levels of Gsx2would limit Gsx1 expression and maintain progenitors in a lessdifferentiated state, whereas the up-regulation of Gsx1 would leadto the down-regulation of Gsx2 and the subsequent maturation ofprogenitors. In summary, we propose amodel for progenitors withinthe LGE (Fig. 4J) in which Gsx2 promotes self-renewal of VZprogenitors. Up-regulation of Gsx1 in cells expressing Ascl1 leadsto the repression of Gsx2 and the transition from the VZ to theSVZ. The loss of Gsx1 and coexpression of Ascl1 and Dlx proteinstogether define neurogenic LGE progenitors.
Materials and MethodsTransgenic Mice. TetO-Gsx1-IE mice were generated as described in the SIMaterials and Methods. Doxycycline hyclate (Sigma) was used to delay the
activation of transgene expression in Foxg1tTA/+; tetO-Gsx1-IE (i.e., Gsx1 DT)embryos. For a detailed description, see ref. 17.
Animals. Genotyping details concerning tetO-Gsx1-IE, tetO-Gsx2-IE, Foxg1tTA/+,Gsx1-EGFP and and Gsx1 mice and embryos are provided in SI Materials andMethods. For staging of embryos, the morning of vaginal plug detection wasdesignated E0.5.
Histology and Immunohistochemistry. Embryos were fixed briefly in 4%paraformaldehyde for 6 h, rinsed extensively in PBS, and cryoprotected in30% sucrose in PBS before being sectioned at 12 μM on a cryostat. For in vivocell cycle length and cell cycle exit studies, cryoprotected embryos weresectioned at 8 μM on the cryostat. Immunohistochemistry was done on slide-mounted sections as described previously (17, 22). For a detailed descriptionof the antibodies used, see SI Materials and Methods.
Cell Cycle Length and Cell Cycle Exit Analysis and Quantification. Analyses wereperformed according to ref. 32, as described in SI Materials and Methods.
RNA Purification and Quantitative RT-PCR Assays. Analyses were performed asdescribed in SI Materials and Methods.
Neurosphere Culture. A modified protocol was used to culture neurospheresfrom E12.5 mouse embryos (37). For a detailed description, see SI Materialsand Methods.
ACKNOWLEDGMENTS. We thank Ron Waclaw for helpful comments andStewart Anderson and Eseng Lai for the Foxg1tTA mice. We also thankT. Edlund, M. Goulding, J. Johnson, and J. Kohtz for antibodies. This workwas supported by National Institutes of Health Grants NS044080 andNS069893.
1. Dehay C, Kennedy H (2007) Cell-cycle control and cortical development. Nat RevNeurosci 8:438–450.
2. Warren N, et al. (1999) The transcription factor, Pax6, is required for cell proliferationand differentiation in the developing cerebral cortex. Cereb Cortex 9:627–635.
3. Toresson H, Potter SS, Campbell K (2000) Genetic control of dorsal-ventral identity inthe telencephalon: Opposing roles for Pax6 and Gsh2. Development 127:4361–4371.
4. Yun K, Potter S, Rubenstein JL (2001) Gsh2 and Pax6 play complementary roles indorsoventral patterning of themammalian telencephalon.Development 128:193–205.
5. Quinn JC, et al. (2007) Pax6 controls cerebral cortical cell number by regulating exitfrom the cell cycle and specifies cortical cell identity by a cell autonomous mechanism.Dev Biol 302:50–65.
6. Heins N, et al. (2001) Emx2 promotes symmetric cell divisions and a multipotential fatein precursors from the cerebral cortex. Mol Cell Neurosci 18:485–502.
7. Bishop KM, Garel S, Nakagawa Y, Rubenstein JL, O’Leary DD (2003) Emx1 and Emx2cooperate to regulate cortical size, lamination, neuronal differentiation, developmentof cortical efferents, and thalamocortical pathfinding. J Comp Neurol 457:345–360.
8. O’Leary DD, Chou SJ, Sahara S (2007) Area patterning of the mammalian cortex.Neuron 56:252–269.
9. Hsieh-Li HM, et al. (1995) Gsh-2, a murine homeobox gene expressed in thedeveloping brain. Mech Dev 50:177–186.
10. Valerius MT, et al. (1995) Gsh-1: A novel murine homeobox gene expressed in thecentral nervous system. Dev Dyn 203:337–351.
11. Szucsik JC, et al. (1997) Altered forebrain and hindbrain development in mice mutantfor the Gsh-2 homeobox gene. Dev Biol 191:230–242.
12. Kriks S, Lanuza GM, Mizuguchi R, Nakafuku M, Goulding M (2005) Gsh2 is requiredfor the repression of Ngn1 and specification of dorsal interneuron fate in the spinalcord. Development 132:2991–3002.
13. Corbin JG, Gaiano N, Machold RP, Langston A, Fishell G (2000) The Gsh2homeodomain gene controls multiple aspects of telencephalic development.Development 127:5007–5020.
14. Toresson H, Campbell K (2001) A role for Gsh1 in the developing striatum andolfactory bulb of Gsh2 mutant mice. Development 128:4769–4780.
15. Carney RS, Cocas LA, Hirata T, Mansfield K, Corbin JG (2009) Differential regulation oftelencephalic pallial-subpallial boundary patterning by Pax6 and Gsh2. Cereb Cortex19:745–759.
16. Wang B, Waclaw RR, Allen ZJ 2nd, Guillemot F, Campbell K (2009) Ascl1 is a requireddownstream effector of Gsx gene function in the embryonic mouse telencephalon.Neural Dev 4:5.
17. Waclaw RR, Wang B, Pei Z, Ehrman LA, Campbell K (2009) Distinct temporalrequirements for the homeobox gene Gsx2 in specifying striatal and olfactory bulbneuronal fates. Neuron 63:451–465.
18. Yun K, Garel S, Fischman S, Rubenstein JL (2003) Patterning of the lateral ganglioniceminence by the Gsh1 and Gsh2 homeobox genes regulates striatal and olfactorybulb histogenesis and the growth of axons through the basal ganglia. J Comp Neurol461:151–165.
19. Li H, Zeitler PS, Valerius MT, Small K, Potter SS (1996) Gsh-1, an orphan Hox gene, isrequired for normal pituitary development. EMBO J 15:714–724.
20. Corbin JG, Rutlin M, Gaiano N, Fishell G (2003) Combinatorial function of thehomeodomain proteins Nkx2.1 and Gsh2 in ventral telencephalic patterning.Development 130:4895–4906.
21. Hanashima C, Shen L, Li SC, Lai E (2002) Brain factor-1 controls the proliferation anddifferentiation of neocortical progenitor cells through independent mechanisms. JNeurosci 22:6526–6536.
22. Stenman J, Toresson H, Campbell K (2003) Identification of two distinct progenitorpopulations in the lateral ganglionic eminence: Implications for striatal and olfactorybulb neurogenesis. J Neurosci 23:167–174.
23. Waclaw RR, et al. (2006) The zinc finger transcription factor Sp8 regulates thegeneration and diversity of olfactory bulb interneurons. Neuron 49:503–516.
24. Smart IH (1976) A pilot study of cell production by the ganglionic eminences of thedeveloping mouse brain. J Anat 121:71–84.
25. Bhide PG (1996) Cell cycle kinetics in the embryonic mouse corpus striatum. J CompNeurol 374:506–522.
26. Takahashi T, Nowakowski RS, Caviness VS, Jr. (1995) Early ontogeny of the secondary pro-liferative population of the embryonic murine cerebral wall. J Neurosci 15:6058–6068.
27. Sheth AN, Bhide PG (1997) Concurrent cellular output from two proliferative populationsin the early embryonic mouse corpus striatum. J Comp Neurol 383:220–230.
28. Halliday AL, Cepko CL (1992) Generation and migration of cells in the developingstriatum. Neuron 9:15–26.
29. Noctor SC, Martínez-Cerdeño V, Ivic L, Kriegstein AR (2004) Cortical neurons arise insymmetric and asymmetric division zones and migrate through specific phases. NatNeurosci 7:136–144.
30. Cremisi F, Philpott A, Ohnuma S (2003) Cell cycle and cell fate interactions in neuraldevelopment. Curr Opin Neurobiol 13:26–33.
31. Salomoni P, Calegari F (2010) Cell cycle control of mammalian neural stem cells:Putting a speed limit on G1. Trends Cell Biol 20:233–243.
32. Chenn A, Walsh CA (2002) Regulation of cerebral cortical size by control of cell cycleexit in neural precursors. Science 297:365–369.
33. Kuan CY, Roth KA, Flavell RA, Rakic P (2000) Mechanisms of programmed cell death inthe developing brain. Trends Neurosci 23:291–297.
34. Takahashi T, Nowakowski RS, Caviness VS, Jr. (1995) The cell cycle of thepseudostratified ventricular epithelium of the embryonic murine cerebral wall. JNeurosci 15:6046–6057.
35. McConnell SK (1995) Constructing the cerebral cortex: Neurogenesis and fatedetermination. Neuron 15:761–768.
36. Temple S (2001) The development of neural stem cells. Nature 414:112–117.37. Nagao M, et al. (2008) Coordinated control of self-renewal and differentiation of
neural stem cells by Myc and the p19ARF-p53 pathway. J Cell Biol 183:1243–1257.38. Casarosa S, Fode C, Guillemot F (1999) Mash1 regulates neurogenesis in the ventral
telencephalon. Development 126:525–534.39. Yun K, et al. (2002) Modulation of the notch signaling by Mash1 and Dlx1/2 regulates
sequential specification and differentiation of progenitor cell types in the subcorticaltelencephalon. Development 129:5029–5040.
40. Orian A, et al. (2007) A Myc-Groucho complex integrates EGF and Notch signaling toregulate neural development. Proc Natl Acad Sci USA 104:15771–15776.
1680 | www.pnas.org/cgi/doi/10.1073/pnas.1008824108 Pei et al.