hong kong baptist university master's thesis biological
TRANSCRIPT

Hong Kong Baptist University
MASTER'S THESIS
Biological control of golden apple snails (Pomacea canaliculata) infreshwater wetland using black carp (Mylopharyngodon piceus)Ip, Ka Lok Kelvin
Date of Award:2013
Link to publication
General rightsCopyright and intellectual property rights for the publications made accessible in HKBU Scholars are retained by the authors and/or othercopyright owners. In addition to the restrictions prescribed by the Copyright Ordinance of Hong Kong, all users and readers must alsoobserve the following terms of use:
• Users may download and print one copy of any publication from HKBU Scholars for the purpose of private study or research • Users cannot further distribute the material or use it for any profit-making activity or commercial gain • To share publications in HKBU Scholars with others, users are welcome to freely distribute the permanent URL assigned to thepublication
Download date: 04 Dec, 2021

Biological Control of Golden Apple Snails (Pomacea canaliculata)
in Freshwater Wetland
using Black Carp (Mylopharyngodon piceus)
IP Ka Lok Kelvin
M. Phil. Thesis
Department of Biology
Hong Kong Baptist University
October 2013


Biological Control of Golden Apple Snails (Pomacea canaliculata)
in Freshwater Wetland
Using Black Carp (Mylopharyngodon piceus)
IP Ka Lok Kelvin
A thesis submitted in partial fulfilment of the requirements
for the degree of
Master of Philosophy
Principal Supervisor: Dr. QIU Jianwen
Hong Kong Baptist University
October 2013

i
Declaration
I hereby declare that this thesis represents my own work which has been done
after registration for the degree of MPhil at Hong Kong Baptist University, and
has not been previously included in a thesis, dissertation submitted to this or
other institution for a degree, diploma or other qualification.
Signature
October, 2013

ii
Abstract
The apple snail Pomacea canaliculata Lamarck is a native of South America but
has invaded Hong Kong since early 1980s. Its feeding has resulted in a
tremendous loss in semi-aquatic agriculture, especially rice (Oryza sativa L.) and
other aquatic crops such as taro (Colocasia esculenta L.) and water spinach
(Ipomoea aquatica Forssk). While spreading to freshwater wetlands, its feeding
threatens macrophyte diversity. Owing to its voracious appetite, this invasive
snail has also become a competitor of lowland indigenous mollusks. On
ecosystem level, over-grazing by high density of apple snails could also induce
excessive release of nutrients from macrophytes to water bodies, thus promoting
phytoplankton growth and primary production.
Measures to control invasive apple snails fall into three categories:
mechanical / cultural, chemical, and biological. Among them, biological control
methods are appealing because they are usually considered relatively less labor-
intensive and more cost-effective. However, both the control efficacy and
potential non-target effects should be carefully evaluated before adopting a
species in biological control. Although various fish species have been proposed
as biological control agents for apple snails, their effectiveness and non-target
effects on wetland flora and fauna are largely unknown.
This study investigated the feasibility of black carp (Mylopharyngodon
piceus Richardson) as bio-control agent for apple snails in both laboratory and
field experiments.

iii
The laboratory experiment compared the feeding of black carp, common
carp (Cyprinus carpio L.) and white-spotted catfish (Clarias fuscus Lacepède) on
apple snails. These three species are indigenous and widely aquacultured in
southern China. The three species of fish of comparable body length were each
offered apple snails of various sizes ad libitum in aquaria. Black carp (fork length:
165 mm; maximum gap width: 16 mm) was the most effective predator, with a
predatory rate of 70.5 apple snails in 48 hours. Common carp and white-spotted
catfish of similar fork lengths consumed only 58.6 and 15.7 apple snails on
average within the same experimental period. Apple snails preyed upon by black
carp and common carp were juveniles, with their respective shell length ranged
from 3 - 16mm and 3 - 17mm, while that for white-spotted catfish ranged from 3
- 21mm.
An 8-week mesocosm experiment was conducted in a constructed wetland
during the dry season of 2011 to determine whether black carp (fork length: 170 -
185 mm) is as effective as common carp (fork length: 170 - 195 mm) as a bio-
control agent for apple snails, but causes less herbivory to macrophytes and
predation to non-Pomacea snails. Both species of carp preyed effectively on P.
canaliculata, removing almost all apple snail individuals (~ 200 per enclosure)
that were small enough to fit into their mouths. The effects of the two fish
species on macrophytes were different. Black carp reduced herbivory on
macrophytes through reducing apple snail density. However, common carp
reduced apple snail density but did not result in a lower level of herbivory
because it also grazed on macrophytes. Non-target mollusk density was reduced
by both fish species.

iv
A one-year whole-pond experiment was also conducted in June 2012 to
June 2013 to investigate the applicability of black carp as a biological control
agent of apple snails in constructed freshwater wetlands. Three separate
constructed freshwater wetlands were used as replicates of the experiment. Each
wetland was divided into a control side without black carp and a treatment side
with black carp. Four individuals of black carp (fork length 260 - 310 mm) were
released to the side of wetland assigned as treatment. Prior to starting the
experiment and every three months, density of apple snails and other macro-
invertebrates, apple snail egg clutch size and abundance, water quality
parameters (total nitrogen, ammonia nitrogen, total phosphorus and reactive
phosphorus) were recorded. Black carp was highly tolerant to the low dissolved
oxygen in the shallow stagnant waters. It was an effective predator of juvenile
apple snails (<5 – 25mm), but it did not result in significant reduction of adult
apple snails (shell length >25mm) nor affected their reproduction. In addition,
black carp preyed on non-apple snail macro-invertebrates, especially mollusks.
In conclusion, our study has shown that juvenile black carp (minimum total
length: 300mm) is a suitable bio-control agent of apple snails in shallow water
wetlands as it is tolerant of stagnant poor water quality and is an effective
predator of apple snails. A major decline of 89.2% in average overall density of
apple snail has been recorded in the treatment plots of the three experimental
sites after one year. Juvenile snails would be eradicated before they get to mature
minimum size (male SL: 25.2 ± 3.3mm; female: 29.8 ± 3.6mm) for reproduction.
Given the longevity of black carp, a low stocking density (80-89 individuals ha-1
)
is sufficient to control apple snail populations. However, black carp reduces the

v
abundance and diversity of non-target macro-invertebrates. Therefore the
benefits of the biological control must be weighed against the potential
undesirable effects on wetland diversity before adopting in the pest management.
To maximize the control efficacy, mechanical methods to eradicate adult snails,
for instance hand-picking in the shallow water, should be implemented with
biological control effort in an integrated apple snail management program.

vi
Acknowledgements
I would like to take this opportunity to thank my principal supervisor Dr.
Jianwen Qiu for his insightful guidance, generous sharing of knowledge,
continuous support and patience throughout my course of study. I would like to
thank whole-heartedly for his encouragement and support for me in pursuing this
degree with such an inspiring topic that could provide practical management
recommendations for wetlands in future. Special thanks to my co-supervisor Dr.
Yan Liang who has provided vital comments for my study and manuscripts as
well.
I must thank the Mass Transit Railway Corporation Ltd. for providing
financial support for this research. I would like to deliver my thanks to Dr. Kam
Foon Chan, Mr. Peter Choi, Ms. Catherine Leung, Ms. Ida Yu and Mr. John
Allcock who has provided technical comments and all necessary on-site support
for field experiments in Kam Tin compensation wetlands.
I would also like to thank Ms. Yingxuan Li for her kind assistance in
handling the technical and financial problems for us throughout the study. I am
thankful to my dear labmates, Dr. Jin Sun, Ms. Yanan Sun, Mr. Paul Fong, Mr.
James Xie, Ms. Huawei Wu who have provided support in field work,
encouragements, comfort and laughter whenever we chat. I am also grateful to
receive useful comments from two previous students of my principal supervisor:
Mr. King Lun Allan Kwok, Mr. Pak Ki Wong who conducted their in-depth
studies and contributed to understanding in ecology of apple snails in Hong Kong.
Special thanks to Mr. Mak who provided us with precious black carp specimens;
Mr. Aaron Au and Dr. Dickey Lau for assistance in contacting local fish farmers;

vii
Mr. Tim Chung, Ms. Phoebe Lam, Mr. Lam Ngo Lun Alan and Ms. Priscilla Li
for their assistance to my tedious field work in summer.
I would like to extend my thanks to my parents. I am forever indebted to
their love, care, encouragement and understanding in my current research and
previous 25 years. Their support for my freedom in exploring personal academic
interest and career pathway has always been the most precious gift of my lifetime.
I would also like to thank Ms. Flora Leung for her love and encouragement for
me in the last four years, whilst I hope that would last till ever.
Last but not least, thanks to all indigenous fauna and flora of Hong Kong.
You have strived through the time where even few trees could ever survive after
wartime. With your contribution, we are proud of being one of the wealthiest
cities in the world with such huge biodiversity. Though we have deprived your
rights to thrive within several decades by over-exploitation, habitat degradation,
and introduction of invasive species, I hope our efforts and enthusiasm would
finally bridge the knowledge gaps in biodiversity conservation and ecological
consequences brought forth by rapid development.

viii
Table of Contents
Declaration .......................................................................................................... i
Abstract.............................................................................................................. ii
Acknowledgements ........................................................................................... vi
Table of Contents ............................................................................................ viii
List of Tables ..................................................................................................... x
List of Figures ................................................................................................... xi
Chapter 1 Introduction........................................................................................ 1
1.1 Natural distribution range and pathway of introduction of apple snails .. 1
1.2 Distribution and identity of apple snails in Hong Kong .......................... 2
1.3 Impact of apple snails as invasive species to Chinese wetlands .............. 4
1.4 Current control methods of apple snails in Hong Kong .......................... 8
1.5 Biological control and indigenous predators of apple snails in Hong
Kong ......................................................................................................... 10
1.6 Research objectives and general organization of thesis ........................ 13
Chapter 2 Predatory potential of indigenous fish as bio-control agent of apple
snails ................................................................................................................ 19
2.1 Background ......................................................................................... 19
2.2 Material and method ............................................................................ 22
2.3 Results ................................................................................................ 24
2.4 Discussion ........................................................................................... 25
Chapter 3 Biological control of invasive apple snails in a freshwater wetland by
two carp species: subtle differences matter ....................................................... 32
3.1 Background ......................................................................................... 32
3.2 Materials and methods ......................................................................... 35
3.3 Results ................................................................................................ 39
3.4 Discussion ........................................................................................... 42
Chapter 4 Wetland scale application of biological control of apple snails using
black carp: efficacies and subtle effect to ecosystem......................................... 51
4.1 Background ......................................................................................... 51
4.2 Study site ............................................................................................ 54
4.3 Method ................................................................................................ 55

ix
4.4 Results ................................................................................................ 61
4.5 Discussion ........................................................................................... 67
5.1 Summary of information on local invasion history and possible
indigenous predators of apple snails .................................................... 94
5.2 Predatory potential of different local fish species on apple snails ......... 96
5.3 Mesocosm studies on functional practicability of fish: testing efficacies
and subtle effects in a nutshell ............................................................. 97
5.4 Wetland scale application of biological control of apple snails using
black carp: efficacies and subtle effect to ecosystem............................ 98
5.5 Implications on integrated pest management ....................................... 99
Appendices .................................................................................................... 100
References ..................................................................................................... 104
Curriculum Vitae............................................................................................ 128

x
List of Tables
Page
Table 1.1 List of possible indigenous predators of aquatic mollusks
in Hong Kong wetlands
15
Table 2.1Results of one-way ANOVA and post-hoc Tukey test on
total abundance of consumed apple snails after the treatment 28
Table 4.1 Fish survival in three experimental sites in June 2012-
September 2013
74
Table 4.2 Results of two-way ANOVA testing the difference in
estimated hatchlings between treatments and among sampling time
75
Table 4.3 Effect of black carp on density of different size range of
apple snail and their reproductivity
76
Table 4.4 Hatchability of collected egg clutches in laboratory 77
Table 4.5 Effect of black carp to non-target gastropods
78
Table 4.6 Effect of black carp on abundance of non-target benthic
invertebrates except gastropods. Data are total abundance of each
family of invertebrates in 1.5m2 quadrat area
79
Table 4.7 Effect of black carp on abundance of non-target benthic
invertebrates except gastropods. Data are total abundance of each
family of invertebrates in 1.5m2 quadrat area
80
Table 4.8 Results of two-way ANOVA testing the difference in
nutrient parameters in individual sites between treatment and among
sampling period. Nutrient parameters abbreviations: NH3 =
ammoniacal nitrogen; TN = total nitrogen; P = phosphates; TP =
total phosphorus
81
Table 4.9 The black carp experiment. Effect of black carp and apple
snails on water quality
Times 1-5 represented June 2012, September 2012, December 2012,
March 2013 and June 2013 respectively. Data are means with SE in
parentheses
82
Table 4.10 Effect of fish and apple snails on plant abundance. Data
are mean of eight quadrats with S.D. in parentheses. Time T1 and
T2 represented June 2012 and June 2013 respectively
84

xi
List of Figures
Page
Fig. 2.1 Size-dependent predation of three fish species on apple
snails in aquaria. Each graph for each species showed total
abundance of snails survived in six aquaria before (t=0hrs) and after
(t = 48hrs) the experiment
29
Fig. 2.2 Mean percentage of survival for golden apple snails in six
aquaria, after individual treatments with three species of fish
30
Fig. 2.3 Comparison of mean amount of snails consumed per
experimental tank among three treatments. Letters above the bars
are results of one-way ANOVA and the Tukey test comparing the
difference in mean amount of golden apple snails consumed per
tank among treatments. Different letters and asterisk indicate
significant difference at p < 0.05
31
Fig. 3.1 Size frequency distribution of Pomacea canaliculata at the
end of the 8-week experiment. Abbreviations: AS = treatment
containing apple snails; AS+CC = treatment containing apple snails
and common carp; AS+BC = treatment containing apple snails and
black carp
47
Fig. 3.2 Abundance of non-Pomacea mollusks at the end of the 8-
week experiment. Each bar shows the mean number of mollusks per
replicate for each treatment
48
Fig. 3.3. Size-frequency distribution of the two most abundant non-
target mollusks – Sinotaia quadrata and Melanoides tuberculata.
Each graph shows the pooled data from five replicate enclosures for
each treatment
49
Fig. 3.4. Biomass of three species of macrophytes at the end of the
8-week experiment. Each bar represents the mean ± S.D. of all
available (3-5) replicate enclosures. Letters above the bars are
results of the Tukey tests comparing the difference in biomass
among treatments
50
Fig. 4.1 Mean densities of apple snails in each plot from June 2012
to June 2013
85
Fig. 4.2 Number of egg clutches encountered during walkover
survey from June 2012 – June 2013 86
Fig. 4.3 Abundance of apple snail hatchlings (individuals m-2
) from
June 2012 to March 2013
87

xii
Fig. 4.4 Size-frequency distribution of Pomacea canaliculata in
experimental and control plots from June 2012 to March 2013
88
Fig. 4.5 Densities of three major species of non-target mollusks
(individuals m-2
) in June 2012 – March 2013
89
Fig. 4.6 Comparison of physical water quality parameters within
June 2012 to June 2013. Mean values of each data were shown with
standard deviations as error bars
90
Fig. 4.7 Comparison of chemical water quality parameters within
June 2012 to June 2013. Mean values of each data were shown with
standard deviations as error bars
91
Fig. 4.8 2D nMDS plots showing impact of black carp and apple
snail on vegetation structure from initial (June 2012 – June 2013),
using three sites as replicates
92

1
Chapter 1 Introduction
1.1 Natural distribution range and pathway of introduction of apple snails
Pomacea (Ampullariidae) is a large genus of apple snails with more than 100
described species (Cowie and Thiengo, 2003). They are indigenous to freshwater
marshes in South America. At least three species (i.e. Pomacea canaliculata
Lamarck, P. maculata Perry and P. scalaris D’ Orbigny) have been introduced
into Asia since the 1980s (Anderson, 1993; Jhang, 1985; Joshi, 2007; Yusa and
Wada, 1999; Joshi et al., 2001; Chen et al., 2004; Cowie et al., 2006; Hayes et al.,
2008; Hayes et al., 2012; Lv et al., 2013). P. canaliculata was the first species
introduced to Asia, through importation of egg clutches to establish snail
aquaculture in Taiwan in 1980. This species was soon introduced to Japan, China
and several countries in Southeast Asia including the Philippines, Vietnam,
Cambodia and Thailand (Hamada and Matsumoto, 1985; Cha, 1989; Mochida,
1991; Joshi and Sebastian, 2006). In addition to P. canaliculata, P. maculata and
P. scalaris were also introduced into Asia (Cowie et al. 2006; Hayes et al. 2008;
2012). In China, P. canaliculata and P. maculata exhibited a wide and mosaic
distribution in southern China (Lv et al., 2009; Lv et al., 2013). In Taiwan, P.
canaliculata is widely distributed, but the distribution of P. scalaris is restricted
to southern part (Chen, 2011; Lee and Wu, 1996; Wu et al., 2011). Due to their
rapid growth and high protein content, apple snails were initially widely
promoted as human food and protein supplement in animal feed. However, apple
snail aquaculture farms soon stopped operation due to a number of reasons: lack
of tenderness of apple snail meat, high processing cost, and infestation by
eosinophilic meningitis causing rat lungworm Angiostrongylus cantonensis

2
(Mochida, 1991; Yu et al., 2001; Lv et al., 2008). Abandoned snails soon
established large populations in various freshwater habitats through active
crawling and passive transportation by water flow in drainage systems (Mochida,
1991; Yusa and Wada, 1999; Cowie, 2002). In this chapter we provide a review
of the ecology (distribution, secondary production, and impact to wetland flora
and fauna and wetland function) and management of apple snails in the
agricultural and non-agricultural areas of Hong Kong.
1.2 Distribution and identity of apple snails in Hong Kong
Apple snails were first reported in northern New Territories as a result of a
territory-wide survey conducted in 1988 by Yipp et al. (1991), who estimated
that these snails were introduced to agricultural area with many vegetable
gardens and abandoned rice paddies in 1980 to 1983. This estimation was
consistent with the results of an earlier territory-wide survey of mollusks
conducted in 1980-1981 by Yipp (1983), who did not find apple snails. There is
no consensus about the source and invasion pathway for apple snails in Hong
Kong. Because the first report of apple snails in the agricultural area of the New
Territories of Hong Kong was very close in timing of the first report of apple
snails in Guangdong (Cai, 1990), and the agricultural areas of Hong Kong and
Shenzhen (southern Guangdong) are connected by a network of streams and
drainage channels, it was difficult to determine apple snails were first introduced
to which of these two areas.
Within the subsequent two decades of their invasion, apple snails spread to
most lowland wetlands in the New Territories including streams, ponds,
freshwater marshes, abandoned rice paddies, vegetable gardens and drainage

3
channels in the New Territories. The periodic flooding during the summer rainy
season must have helped apple snail dispersion in the lowland wetlands, which
are connected through streams and drainage channels. A territory-wide survey of
apple snails conducted in 2006 (Kwong et al., 2008) revealed the wide-
distribution of apple snails in the New Territories (Fig. 1). The sites inhabited by
apple snails typically had high levels of phosphate and alkalinity, but apple snails
were also occasionally found in streams where nutrient and alkalinity levels were
low. It was also found that geographical isolation and a lack of agricultural
activities may have halted the spread of apple snails to other areas with apparent
suitable habitats in Hong Kong. Specifically, although the water quality
parameters of sites, i.e. abandoned rice paddies, on Lantau Island were
apparently suitable for apple snails, they had not been colonized due to a lack of
physical connection with the New Territories. However, the building of bridges
connecting the New Territories and Lantau Island in 1997 has greatly enhanced
the traffic, and there is concern of introducing apple snails to Lantau Island
through replanting abandoned rice paddies with agricultural crops, which may
carry apple snail eggs, as a result of the development of ecotourism. There was
some confusion regarding the identity of apple snails introduced into Hong Kong
(Kwong et al., 2008). Cha (1989) and Yipp et al. (1991) identified two species of
apple snails - Ampullaria levior and A. gigas. Although both species were found
in northern New Territories, they showed some differences in their distribution
(Cha, 1989). Yipp et al. (1991) concluded that A. gigas preferred habitats with
bottom sediment of coarser grain sizes while A. levior was a dominant grazer in
finer ones. Lam (1994) identified the apple snails collected from a stream in the

4
New Territories as P. levior, whereas Dudgeon and Corlett (2004) considered P.
lineata to be the species of apple snails that are widely spread in Hong Kong. To
determine the species identity of apple snails in Hong Kong, selective snails
collected from different locations (Kwong et al., 2008) showing the greatest
morphological variations with respect to size, shell color and shell shape were
sent to Robert Cowie and Kenneth Hayes of University of Hawaii, who
successfully amplified a segment of the cytochrome oxidase I gene for 12
individuals. An analysis of these mitochondrial DNA sequences showed that all
of these individuals were P. canaliculata (Kenneth Hayes, pers. comm.),
therefore P. canaliculata is likely the only species of Pomacea currently present
in Hong Kong. Based on shell morphology, this species shall be A. levior in Cha
(1989) and Yipp et al. (1991), P. levior in Lam (1994) and P. lineata in Dudgeon
and Corlett (2004). A. gigas in Cha (1989) and Yipp et al. (1991) was likely P.
maculata, which can reach a larger maximum body size than P. canaliculata
(Hayes et al. 2012). P. maculata has successfully established in some areas of
southern China (Lv et al., 2013), but may have become locally extinct in Hong
Kong.
1.3 Impact of apple snails as invasive species to Chinese wetlands
Natural existence of apple snails in agricultural wetlands of South America has
been reported to be harmless (Cazzaniga, 2006). Yet, it has been widely regarded
as pests to freshwater agricultural plants in many tropical and sub-tropical
countries, especially to rice (Oryza sativa) and other aquatic crops such as taro
(Colocasia esculenta) and water spinach (Ipomoea aquatica ) (Anderson, 1993;
Joshi, 2007; Naylor, 1996; Yusa and Wada, 1999a; Yusa and Wada, 1999b). In

5
Taiwan, an annual loss of US $3.89 million has been estimated, with 100,000 ha
of agricultural lands devastated by apple snails yearly (Huang, 2005).
Approximately 660,000 ha and 167,000 ha of rice paddies has been infested with
apple snails in Guangdong and Guangxi province in 2006 respectively, and apple
snail population is expanding further north at rate of 8-10 km per year (Yin et al.,
2006).
In fact, the impact of apple snails to agricultural lands is not restricted to
taros and rice. Grazing by apple snails also significantly reduce the yield of
various aquatic crops in shallow wet agricultural lands, and crops that are planted
immediately adjacent to paddies for instance watercress (Nasturtium officinale)
(Hue et al., 1997) and Manchurian wild rice (Zizania latifolia) (Dong et al.,
2012). Since these wet agricultural lands remain key habitats of farmland birds in
Asia (Amano et al., 2008), infestation of apple snails as an invasive species could
further reduce incentives for conserving these habitats (Pascual and Perrings,
2007) due to their potential impacts on wetland flora and fauna. Apple snails can
disrupt the natural growth cycle of phytoplankton and macrophyte in shallow
wetlands, such as ponds, lakes and floodplains. In natural and artificial wetlands,
macrophytes are a key component in habitat complexity. In general, they provide
surface for the growth of cyanobacteria and periphyton, refuge for animals
against predation, nutrient sink (phosphorus and nitrogen) and reduce re-
suspension of sediments. In shallow wetlands, there are few invertebrate grazers
of macrophytes in southern China including Hong Kong (Dudgeon 1999c). There
is however plenty of shredders that slowly consume allochthonous leaf litter. Yet,
periphyton attachs and grows on the surface of macrophytes constitutes a large

6
portion in the diet of scrapers, while detritus from decomposition of dead
macrophytes comprise a large portion in diet of collector-gatherers (which
include most of the snails) (Dudgeon and Yipp 1983). Previous studies have
reported that exotic apple snails could consume macrophytes and promote the
growth of phytoplankton, thus shifting wetland ecosystems towards a state of low
macrophyte diversity and high nutrient concentration (Carlsson et al., 2004a;
Carlsson and Lacoursière, 2005; Fang et al., 2010).
Apple snails may have out-competed local freshwater invertebrates including
mollusks by predation. Studies have shown that apple snails could approach its
prey in a much quicker rate than the other snails (Kwong et al., 2009) and graze
heavily on fresh aquatic plants with low phenolic content and dry matter content
(Fang et al., 2010; Qiu and Kwong, 2009; Wong et al., 2010). In addition, apple
snails are generalists in feeding and could utilize a much broader spectrum of diet
than other indigenous snails (Kwong et al. 2010). As a highly productive
consumer, its secondary production estimate is approximately 10-20 folds more
than that of Melanoides tuberculata, the most productive exotic snail in the
lowland lentic habitats of Hong Kong (Kwong et al., 2010). Cha (1989) reported
that apple snails could influence the directional movement of indigenous
mollusks, especially Sinotaia quadrata. But it is still unclear whether the feeding
efficiency of S. quadrata would be affected by apple snails in the field. Apple
snails could feed on juveniles and eggs of local freshwater snails as a part of its
diet in supplement to nutrients obtained from macrophytes (Kwong et al., 2009).
Apple snails could complete three generations under favorable conditions
before cold spells come in winter (Kwong et al., 2010; Yin et al., 2006). With the

7
ability to bury in substratum, the adults could hibernate and survive through
winter and become active in the next wet season. Adult apple snail that survive
through winter could reproduce when conditions become favorable, therefore
once apple snails invade an area, they often become established and their impact
can persist for many years.
Although apple snails are notorious invasive species in Asia, there were
advocates on the application of apple snails as bio-control agents, but it was later
concluded that the side-effects of apple snails on non-target flora and fauna are
difficult to remediate. The release of apple snails and their close relatives has
been reported to be beneficiary in some countries, where they were used to
control exotic weeds of weeds (Perera and Walls, 1996; Porte et al. 2006). But
they were also intended to be introduced to countries with severely infestation of
water-borne schistosomiasis disease (Pointier and David, 2004; Hofkin et al.
1991; Frandsen 1987). Parasitic trematodes in the genus Schistosoma
(Platyhelminthes; Trematoda; Digenea; Schistosomatidae) are responsible for
this disease after infesting human through contact or ingestion of contaminated
water (Collins III et al. 2013). Mollusks like Oncomelania spp. and
Biomphalaria spp. are intermediate host of human-related trematodes to
complete its lifecycle (Huang et al., 2010; Tang, 1985). There are advocates on
the use of apple snails and their close relatives as biological control agents for
trematode-hosting mollusks. Marisa cornuarietis, a close relative of apple snails,
was the most widely used snail in controlling Biomphalaria spp. (Haridi and
Jobin, 1985; Nguma et al., 1982; Pointier and Jourdane, 2000; Pointier et al.
1994; Pointier et al. 1991). P. canaliculata has also been used in field

8
experiments in Thailand to control trematode-hosting mollusks (Kruatrachue and
Upatham, 1993). Although these studies may have achieved certain extent of
success in controlling the target snails, the ecological impacts on non-target
macrophytes and invertebrates are difficult to estimate (Cowie, 2001). In addition,
P. canaliculata is the intermediate host of the rat lungworm (Angiostrongylus
cantonensis), which was responsible for a mass infection of over 100 citizens of
Guangxi province (Hao et al., 2007) and 8 citizens of Fujian province (Chen et
al., 2003).
1.4 Current control methods of apple snails in Hong Kong
Since their first invasion in the 1980s, apple snails have become a major pest of
rice in Asia and taro in Hawaii (Cowie, 2002). Although Hong Kong is one of
the most densely populated cities with more than seven million people living on
small area of 1,108 km2, there is still approximately 51 km
2 agricultural land.
While existing rice paddies (approximately 10% of agricultural land) are mainly
for demonstration of tradition rice cultivation in Yuen Long, small-scale farms
still provide 8% of vegetables for local consumption. Several species of semi-
aquatic vegetable, especially Ipomoea aquatica (water spinach), Nasturtium
officinale (watercress), Nelumbo nucifera (Indian lotus) and Sagittaria
sagittifolia (Chinese arrowhead) are commonly cultivated species. Apple snails
are widely distributed in these semi-aquatic vegetable gardens and drainage
channels connecting them. In response to the infestation of apple snails in
vegetable gardens, local farmers have adopted cultural and chemical control
methods. The cultural method involved simple hand picking of apple snail eggs
and large individuals throughout the year, especially the summer when apple

9
snails reach their peak of reproduction and growth (HKBWS, 2006). The
chemical methods involved application of lime and / or tea seed cake (residue of
seeds in the Camellia family, after oil extraction, which contains saponin)
between two crops to control apple snails (Lau, S.P., pers. comm.).
Due to the increasing development pressure, many wetlands in the New
Territories have been lost to construction for residential buildings and associated
roads and highways. As required by the Environmental Impact Assessment
Ordinance (Cap 499), compensation wetlands of a similar or larger size of those
destroyed were created with various macrophytes used to build up the floral
community (Lau et al., 2004). An example of such wetland loss is the large-scale
(220 ha) residential development in Tin Shui Wai for a population of 340,000
(Cha, 2004). As part of the compensation, Hong Kong Wetland Park (60 ha) was
created with channels of different depths connecting many ponds separated by
weirs for flow and water depth control, and macrophytes were planted to filter
intake water and to create habitats of different floral diversity. The invasion of
apple snails to these constructed wetlands may have greatly reduced floral
biodiversity, and their value as habitats of animals, such as birds. Current
wetland management activities in Mai Po Nature Reserve (managed by World
Wide Fund, Hong Kong) and Hong Kong Wetland Park (managed by Agriculture,
Fisheries and Conservation Department and Development Bureau, HKSARG)
and West Rail wetlands in Kam Tin (managed by Mass Transit Railway
Corporation Ltd., Hong Kong) to control apple snail populations in constructed
wetlands mainly involve hand-picking, which can be an effective method, but is

10
labor-intensive and requires repeated efforts (Cowie, 2002; Hung T.T.H., pers.
comm.; Yau V.P.T., pers. comm.).
1.5 Biological control and indigenous predators of apple snails in Hong
Kong
A summary of all potential predators of golden apple snails in Hong Kong
wetlands based upon local and international literatures have been compiled as
table 1.1. Among all resident species of Hong Kong lowland wetlands, there are
only twenty indigenous species have recorded diet on gastropods. It should be
noted that most of these predators are omnivores and apple snails might only
comprised a portion of their diet. Yet, lack of any local studies provided practical
evidences in demonstrating importance of particular species of fauna or
integrated ecosystem in controlling apple snail population.
Biological control of snail species susceptible to contamination has been
practiced in Southeast Asia, including China, Taiwan, Japan, Vietnam, Thailand,
Malaysia, Lao P.D.R., Sri Lanka, Israel and Bangladesh (Mukherjee et al. 1991).
Ducks are well known predators of pest snails in paddy fields (Cagauan et al.,
1999; Gallebu et al., 1992). Before 1970s, fish farmers in the whole North-
western New Territories, including the Deep Bay Area, once widely practiced
duck farming (Tuen Mun and Yuen Long District Planning Office, 2002; Carey
et al. 2001). Previous literature has cited about 58% and 8% of all fish farms
raised ducks and gooses respectively (Kausar, 1983; Sin and Cheng, 1976). Yet,
it has not been practiced anymore locally. In addition, wild Anatidae are only
winter passage migrants of Hong Kong through East Asia-Australian Flyway
(WWF, 2012) instead of resident species. Other than ducks, fish has been

11
suggested and implemented as agent for integrated pest control in paddyfields
(Fernando, 1993; FAO, 1998; Halwart, 1998; Lardans and Dissous, 1998; Teo,
2006; Ichinose et al., 2002; Yusa et al., 2006). Yet, few studies have considered
their potential subtle effects to wetland flora and fauna. Recent mesocosm
experiment by Wong et al. (2009) studied the effectiveness and non-target effects
of common carp (Cyprinus carpio L.) as a biological control agent against P.
canaliculata in a 2-month enclosure experiment. They examined the impact of
common carp on nine species of gastropods including the apple snail, and three
species of macrophytes in a constructed wetland in Hong Kong Wetland Park.
Their results showed that common carp completely eliminated apple snail
juveniles that are small enough to fit into their months. Although they could not
prey on large apple snails due to the use of small fish as constrained by the size
of the enclosures, its ability to grow in local ponds showed common carp is an
effective bio-control agent of apple snails. However, common carp also caused a
significantly reduction of plant biomass, as well as of the densities of most non-
Pomacea gastropods, indicating common carp’s strong non-target effects on
local wetland flora and fauna. To continue finding a cost-effective mean to
control invasive apple snails, this research focused on investigating the predatory
potential and practical control efficacy of black carp Mylopharyngodon piceus
Richardson. on apple snails. Black carp has been suggested in recent years as
remediation in particular to reduce pest mollusks (Ben-Ami and Heller, 2001;
Collins, 1996; Hung et al. 2013; Leventer, 1979; Mochida, 1991; Shelton et
al.,1995; Thomforde, 1999; Venable et al.,2001) and are traditionally reared in
farm ponds (Wu et al., 1964) , fish ponds (Hickling, 1971), rice paddies (Wang,

12
2010) and reservoirs (Leventer and Teltsch, 1990), while being food fish to
generate profit in complement with that from crops yield (Lin, 1955; Liu, 1955).

13
1.6 Research objectives and general organization of thesis
The present thesis consists of five chapters. Chapter 1 is a general introduction
and Chapter 5 is a summary of research findings. Chapter 2 to 4 describes the
laboratory (Chapter 2) and field experiments (Chapter 3-4) conducted to
evaluating the effectiveness and non-target effects of fish as predators of apple
snails with fish as predators. Details of three studies are as follows:
1. Predatory potential of indigenous fish as bio-control agent of apple snails
(Chapter 2) – This chapter mentioned an aquarium study (Chapter 2). It laid
the foundation for the field studies described in Chapters 3 and 4. Three
indigenous fish species were evaluated regarding their predatory potential on
apple snails. The results were useful for selecting a candidate for the field
experiments.
2. Biological control of invasive apple snails in a freshwater wetland by two
species of carp: subtle differences matter (Chapter 3) – this study aimed to
explore and compare the performance of two fish species that fed voraciously
on apple snails in laboratory experiments. In addition, I would like to
understand their possible side-effects on water quality, aquatic plants and
various epi- and benthic fauna in freshwater constructed wetland habitat. Two
species of fish, common carp C. carpio and black carp M. piceus were
selected due to their promising performance in the laboratory experiments
(Chapter 2). The field experiment lasted two months. The results were useful
for selecting a fish for the following whole wetland experiment.
3. Wetland scale application of biological control of apple snails using black
carp: efficacies and subtle effect to ecosystem (Chapter 4) – Black carp,
which was found to be a more desirable predator of apple snails from
previous field trial experiment (Chapter 3), was released in semi-natural
constructed wetlands and was tested upon its practical efficacy in reducing
apple snail abundance. Previous studies only used black carp in small scaled
trials and only its direct impact on the target mollusks were tested (Ben-Ami
and Heller, 2001; Collins, 1996; Mochida, 1991; Shelton et al., 1995;

14
Thomforde, 1999; Venable et al., 2001). They were conducted in semi-
natural habitats with low biodiversity especially shallow rice paddies, without
evaluating the effects on non-target benthic macroinvertebrates and water
quality, while only large scale study in Israel (Leventer 1979) failed to
provide many details of his experimental design and results. In our study,
side effects on water quality and co-existing macrobenthos were also
evaluated (Chapter 4). Macrophyte abundance and diversity were also
analyzed to test on treatment effect.
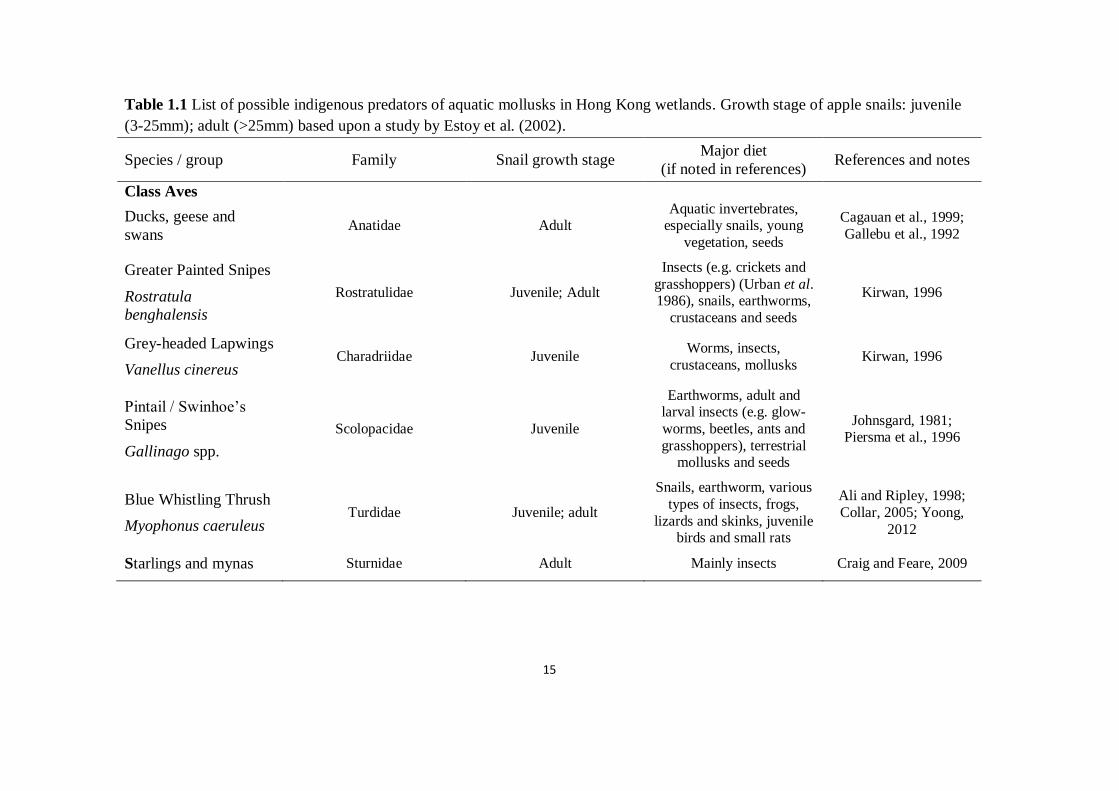
15
Table 1.1 List of possible indigenous predators of aquatic mollusks in Hong Kong wetlands. Growth stage of apple snails: juvenile
(3-25mm); adult (>25mm) based upon a study by Estoy et al. (2002).
Species / group Family Snail growth stage Major diet
(if noted in references) References and notes
Class Aves
Ducks, geese and
swans Anatidae Adult
Aquatic invertebrates, especially snails, young
vegetation, seeds
Cagauan et al., 1999;
Gallebu et al., 1992
Greater Painted Snipes
Rostratula
benghalensis
Rostratulidae Juvenile; Adult
Insects (e.g. crickets and
grasshoppers) (Urban et al. 1986), snails, earthworms,
crustaceans and seeds
Kirwan, 1996
Grey-headed Lapwings
Vanellus cinereus Charadriidae Juvenile
Worms, insects,
crustaceans, mollusks Kirwan, 1996
Pintail / Swinhoe’s
Snipes
Gallinago spp.
Scolopacidae Juvenile
Earthworms, adult and larval insects (e.g. glow-
worms, beetles, ants and
grasshoppers), terrestrial mollusks and seeds
Johnsgard, 1981; Piersma et al., 1996
Blue Whistling Thrush
Myophonus caeruleus Turdidae Juvenile; adult
Snails, earthworm, various
types of insects, frogs,
lizards and skinks, juvenile birds and small rats
Ali and Ripley, 1998;
Collar, 2005; Yoong,
2012
Starlings and mynas Sturnidae Adult Mainly insects Craig and Feare, 2009
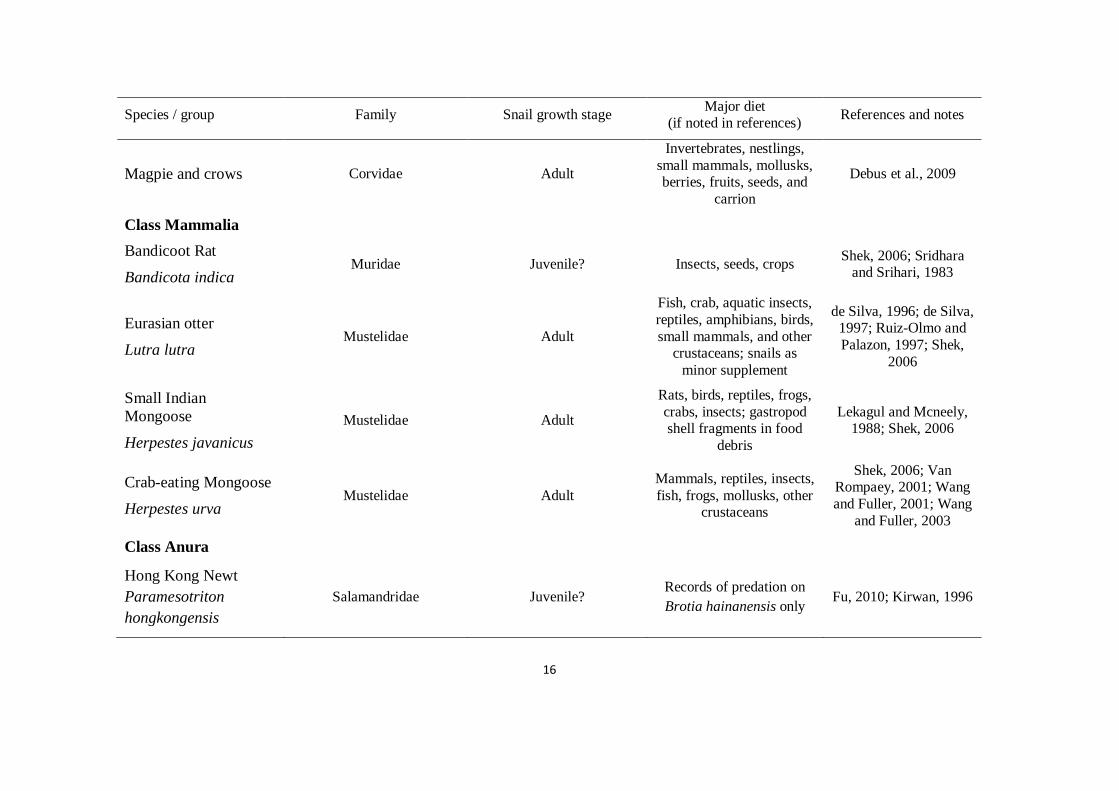
16
Species / group Family Snail growth stage Major diet
(if noted in references) References and notes
Magpie and crows Corvidae Adult
Invertebrates, nestlings,
small mammals, mollusks, berries, fruits, seeds, and
carrion
Debus et al., 2009
Class Mammalia
Bandicoot Rat
Bandicota indica Muridae Juvenile? Insects, seeds, crops
Shek, 2006; Sridhara and Srihari, 1983
Eurasian otter
Lutra lutra Mustelidae Adult
Fish, crab, aquatic insects,
reptiles, amphibians, birds,
small mammals, and other crustaceans; snails as
minor supplement
de Silva, 1996; de Silva, 1997; Ruiz-Olmo and
Palazon, 1997; Shek,
2006
Small Indian
Mongoose
Herpestes javanicus
Mustelidae Adult
Rats, birds, reptiles, frogs,
crabs, insects; gastropod shell fragments in food
debris
Lekagul and Mcneely, 1988; Shek, 2006
Crab-eating Mongoose
Herpestes urva Mustelidae Adult
Mammals, reptiles, insects,
fish, frogs, mollusks, other crustaceans
Shek, 2006; Van Rompaey, 2001; Wang
and Fuller, 2001; Wang
and Fuller, 2003
Class Anura
Hong Kong Newt
Paramesotriton
hongkongensis
Salamandridae Juvenile? Records of predation on
Brotia hainanensis only Fu, 2010; Kirwan, 1996
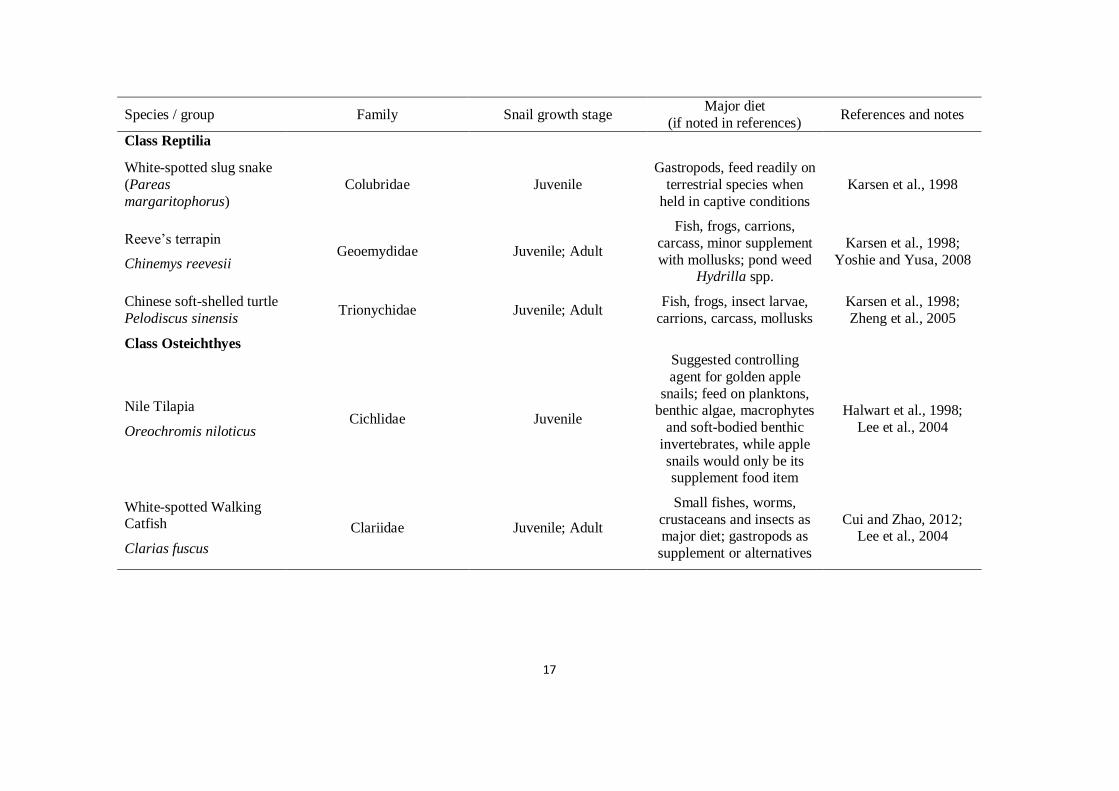
17
Species / group Family Snail growth stage Major diet
(if noted in references) References and notes
Class Reptilia
White-spotted slug snake
(Pareas
margaritophorus)
Colubridae Juvenile
Gastropods, feed readily on
terrestrial species when
held in captive conditions
Karsen et al., 1998
Reeve’s terrapin
Chinemys reevesii Geoemydidae Juvenile; Adult
Fish, frogs, carrions,
carcass, minor supplement
with mollusks; pond weed Hydrilla spp.
Karsen et al., 1998;
Yoshie and Yusa, 2008
Chinese soft-shelled turtle
Pelodiscus sinensis Trionychidae Juvenile; Adult
Fish, frogs, insect larvae,
carrions, carcass, mollusks
Karsen et al., 1998;
Zheng et al., 2005
Class Osteichthyes
Nile Tilapia
Oreochromis niloticus Cichlidae Juvenile
Suggested controlling
agent for golden apple
snails; feed on planktons, benthic algae, macrophytes
and soft-bodied benthic
invertebrates, while apple
snails would only be its supplement food item
Halwart et al., 1998;
Lee et al., 2004
White-spotted Walking Catfish
Clarias fuscus
Clariidae Juvenile; Adult
Small fishes, worms,
crustaceans and insects as major diet; gastropods as
supplement or alternatives
Cui and Zhao, 2012; Lee et al., 2004
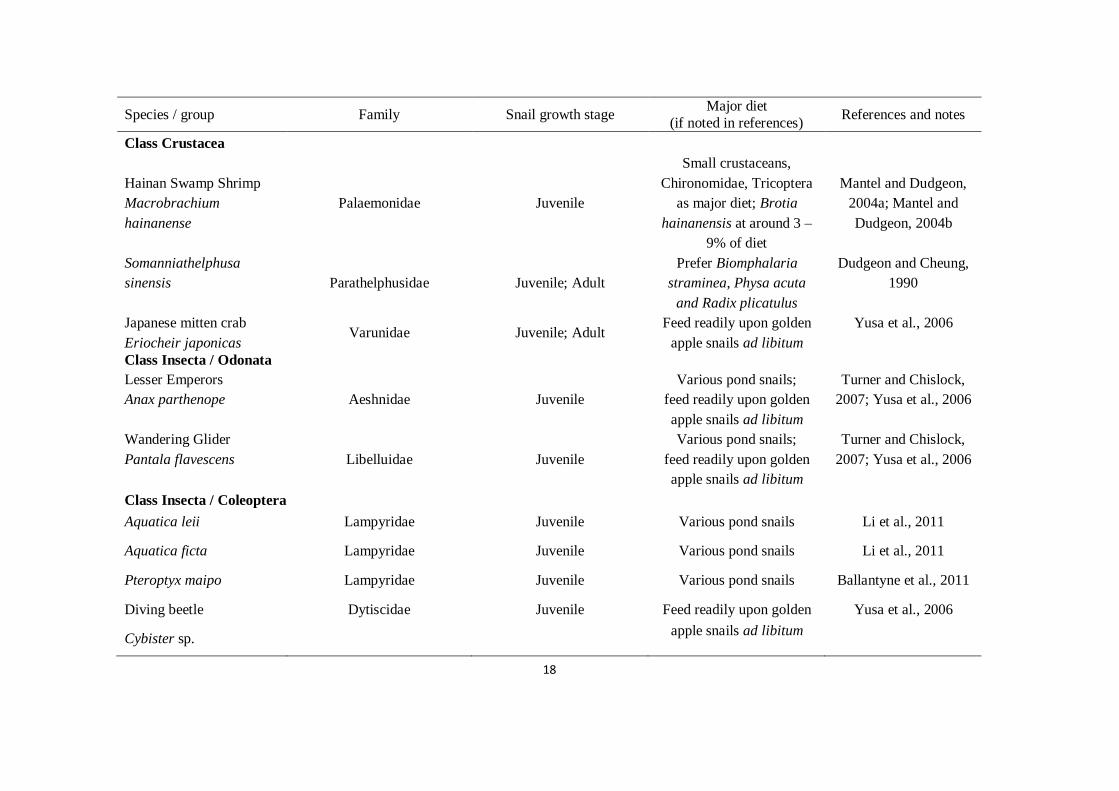
18
Species / group Family Snail growth stage Major diet
(if noted in references) References and notes
Class Crustacea
Hainan Swamp Shrimp
Macrobrachium
hainanense
Palaemonidae Juvenile
Small crustaceans,
Chironomidae, Tricoptera
as major diet; Brotia
hainanensis at around 3 –
9% of diet
Mantel and Dudgeon,
2004a; Mantel and
Dudgeon, 2004b
Somanniathelphusa
sinensis Parathelphusidae Juvenile; Adult
Prefer Biomphalaria
straminea, Physa acuta
and Radix plicatulus
Dudgeon and Cheung,
1990
Japanese mitten crab
Eriocheir japonicas Varunidae Juvenile; Adult
Feed readily upon golden
apple snails ad libitum
Yusa et al., 2006
Class Insecta / Odonata
Lesser Emperors
Anax parthenope Aeshnidae Juvenile
Various pond snails;
feed readily upon golden
apple snails ad libitum
Turner and Chislock,
2007; Yusa et al., 2006
Wandering Glider
Pantala flavescens Libelluidae Juvenile
Various pond snails;
feed readily upon golden
apple snails ad libitum
Turner and Chislock,
2007; Yusa et al., 2006
Class Insecta / Coleoptera
Aquatica leii Lampyridae Juvenile Various pond snails Li et al., 2011
Aquatica ficta Lampyridae Juvenile Various pond snails Li et al., 2011
Pteroptyx maipo Lampyridae Juvenile Various pond snails Ballantyne et al., 2011
Diving beetle
Cybister sp.
Dytiscidae Juvenile Feed readily upon golden
apple snails ad libitum
Yusa et al., 2006

19
Chapter 2 Predatory potential of indigenous fish as bio-control
agent of apple snails
2.1 Background
There has been an increasing incidence of exotic species becoming invasive
when it is removed from its natural enemies (predators or parasites) in its original
niche and upon introduction to a new favorable habitat (Forrest et al. 2012;
Higgins & Vander Zanden 2010; Hermoso et al., 2011; Mack et al., 2000). High
impact invaders established in new habitats and caused more chronic negative
impact to native biota than native species (in terms of diversity loss, change in
disturbance regime and retarding ecosystem succession) (Light and Marchetti,
2007; Simberloff et al., 2012; Paolucci et al., 2013). These species also degraded
ecosystem services of our native ecosystem (Pimentel et al., 2001; Pejchar and
Mooney, 2009) and eventually homogenize aquatic biodiversity (Lodge 1993;
Rahel 2002). Eradication of invasives was therefore crucial before they continue
to spread to suitable habitats and alter the ecosystem (Myers et al. 2000)
Apple snails (Pomacea canaliculata Lamarck.) have first infested Hong Kong
in 1970s and are well-known major pests of rice in Asia and taro in Hawaii (Cha
1989; Cowie, 2002). They have established sustainable population and
distributed widely in these semi-aquatic vegetable gardens, inter-connecting
drainage channels and some compensation wetlands (Kwong et al. 2008). In
response to the infestation of apple snails in vegetable gardens, local farmers
have adopted cultural and chemical control methods. The cultural method
involved detachment of apple snail egg clutches and hand-picking of large
individuals throughout the year, especially in summer when apple snails reach

20
their peak of reproduction and growth (HKBWS, 2006; Hung T.T.H., pers. comm.;
Yau V.P.T., pers. comm). The chemical methods involved application of lime or
tea seed cake (residue of seeds in the Camellia family, after oil extraction, which
contains saponin) between two crops to apple snails (Lau, S.P., pers. comm.).
However, these methods are usually labor-intensive. In addition, chemical
control could not be applied to wetlands with deeper water levels (>1.0m),
especially during wet season as rain-induced spate usually flood those areas and
provide favorable breeding grounds for apple snails.
Classical biological control mechanism that involves introducing predators
existing in the natural area of origin might not be a good solution for controlling
apple snails. These South America originated predators are namely snail kites
(Rostrhamus sociabilis), glossy ibis (Plegadis falcinellus), redear sunfish
(Lepomis microlophus), exotic fire ants (Solenopsis geminata) and introduced
duck varieties (Gallebu et al., 1992; Teo, 2001). Yet, none of these predators are
native in Hong Kong. In addition, there is no consent on an efficient agent that
could minimize their invasion by predation or competition temporally in global
scale.
Natural biological control would be a better option of management, in
which diversity and abundance of native species are manipulated to reduce the
invasion impact to minimal. It is yet difficult to locate a predator or competitor
specific for apple snails. Previous experiment has revealed secondary production
by apple snails outweighed all lowland wetlands fauna in Hong Kong (Kwong et
al., 2010a). In addition, though apple snails have invaded for more than a decade
in Hong Kong (Yipp et al., 1991), there are few observations on predators that

21
feed upon them, while these records focus on only a handful of omnivorous
wetland-dependent residents (Chapter 1, table 1).
Using single fish species as direct controlling agent or in complement with
ducks, is most widely studied bio-control mechanism against apple snail invasion
in Southeast Asia. It originated from studies on a number of cichlid species
(Slootweg et al., 1993; Slootweg et al., 1994), Nile tilapia (Oreochromis niloticus
Linnaeus.) and common carp (Cyprinus carpio L.) (Cagauan et al., 2000;
Halwart, 1994a). Biological control using common carp has then been widely
explored in the laboratory (Caguan 2002; Halwart et al., 1998; Teo, 2006; Yusa
et al. 2006), and rice fields in Taiwan (Mochida, 1991), Vietnam (FAO, 1998),
Japan (Ichinose et al., 2002) and Malaysia (Teo, 2006). In China, black carp
Mylopharyngodon piceus Richardson. was long considered as a potent
molluscivore in fish ponds (Hickling, 1971; Lin, 1955; Liu, 1955). Similar
researches in fish ponds have subsequently continued in Taiwan (Mochida, 1991),
Israel (Ben-Ami and Heller, 2001; Leventer, 1979) and Russia (Denisov, 1982).
Other than using black carp, various species of catfish with wide gape has also
been tested in different countries regarding their effectiveness in controlling
apple snails. These include broadhead catfish Clarias macrocephalus Günther.
(Cagauan and Joshi, 2002; Carlsson and BrÖnmark, 2006), African catfish C.
gariepinus Burchell. (Slootweg et al., 1993) and Philippine catfish C. batrachus
L. (Liao et al., 2004). Though African catfish might prey upon apple snails
(Coates 1984), it is classified as an invasive species in Asia (ISSG 2013). The
only resident species in Genus Clarias in Hong Kong is white-spotted catfish
(Clarias fuscus Lacépède.).

22
The current objective of the present study was to identify two species as
predators of apple snails in wetlands. As fish would be a preferable candidate at
the present stage, we compared the predatory potential of three indigenous fish
species on apple snails by an aquaria study. This study included estimation of the
maximum amount of ingestion and maximum size of snail consumed by the
tested fish species under optimum conditions. By comparing the figures, we
located two most suitable ones with high predatory potential on apple snails for
conducting field trial experiment to further explore their possible side-effects
upon release.
2.2 Material and method
Fish and apple snails used in aquaria experiment
We have used three species of fish for this experiment. They are black carp,
common carp and white-spotted walking catfish. Juvenile black carp (fork length
170-185 mm), common carp (fork length 170-195 mm) and white-spotted catfish
(fork length 150-180mm) were purchased from a commercial fish farm and an
aquarium shop, respectively. The gape width of each fish specimen was
measured by a digital caliper. They are acclimated for at least 2 weeks under
experimental conditions (25oC and in separate tanks for each specimen with 8.0L
dechlorinated water and water circulation system) before the experiment
commenced. All specimens selected for experiment are ensured to feed
simultaneously upon provision of juvenile apple snails reared in laboratory (8-
15mm), with daily supplement by commercial fish feed.
Juvenile (8-25mm) and adult (>25mm, minimum reproductive size due to
previous literature (Estebenet and Cazzaniga, 1992; Martin, 1986) apple snails

23
with variety of sizes were caught from drainage channels of a vegetable garden in
Long Valley (Yin Kong Tsuen, Sheung Shui), They were then transported to
laboratory and were allowed to acclimatize in three separate aquaria with 8.0L of
dechlorinated water and water circulation system for at least 7 days before use.
Aeration was provided in each aquarium and the snails were fed daily with
romaine lettuce (Lactuca sativa L.).
Aquaria set-up
There were six aquaria (size: 67cm*51cm*33.5cm) with 30.0L dechlorinated
water and water circulation system for each species of fish. A day prior to the
experiment, all fish specimens and snails were supplied with no food to allow
natural starvation. Similar size spectrum of approximately 100-140 apple snails
was first introduced into each experiment aquarium. Shell length of each snail
introduced into the tank has been measured by digital caliper to the nearest mm.
For each fish specimens, total length, fork length and gape width were measured.
Upon start of the experiment, only a single fish specimen was allowed to enter
each experimental tank. The experimental period then lasted for 48 hours.
After the experiment, fish specimens were first removed from the tanks.
Surviving snails were collected and measured individually for their shell length.
Dead snails were observed for signs of predation or natural death if exist with
entire shell.
An extra aquarium has been prepared for control treatment, where
approximately same quantity and size spectrum of apple snails was allowed to

24
stay in an aquarium without later introduction of fish. This would serve as a
comparison of snail’s survival with experimental treatments.
Data analysis
Total abundances of snails consumed in six replicates are compared among fish
species and control using one-way ANOVA. In order to investigate the effect of
size-dependent predation, amount of consumption within two size ranges of
apple snails (Juvenile: 3-25mm; adult: >25mm) is compared among three species
of fish and control with one-way ANOVA.
Relationship between fish total length and amount of apple snails
consumed was tested using linear regression analysis.
2.3 Results
Aquaria studies on predatory potential of three fish species on apple snails
All of the apple snails in control treatment survived throughout the experimental
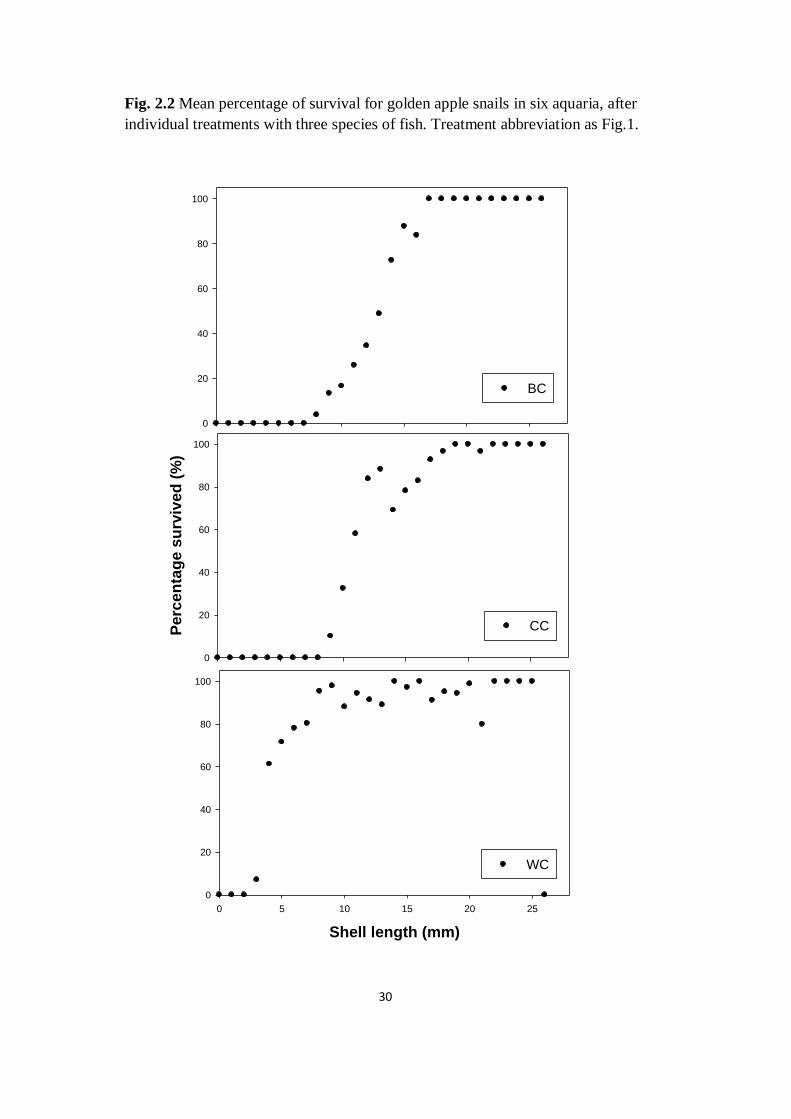
period. Among six replicates of each treatment, mean survival rate of apple snails
with presence of catfish was 86.1%, while that with presence of common carp
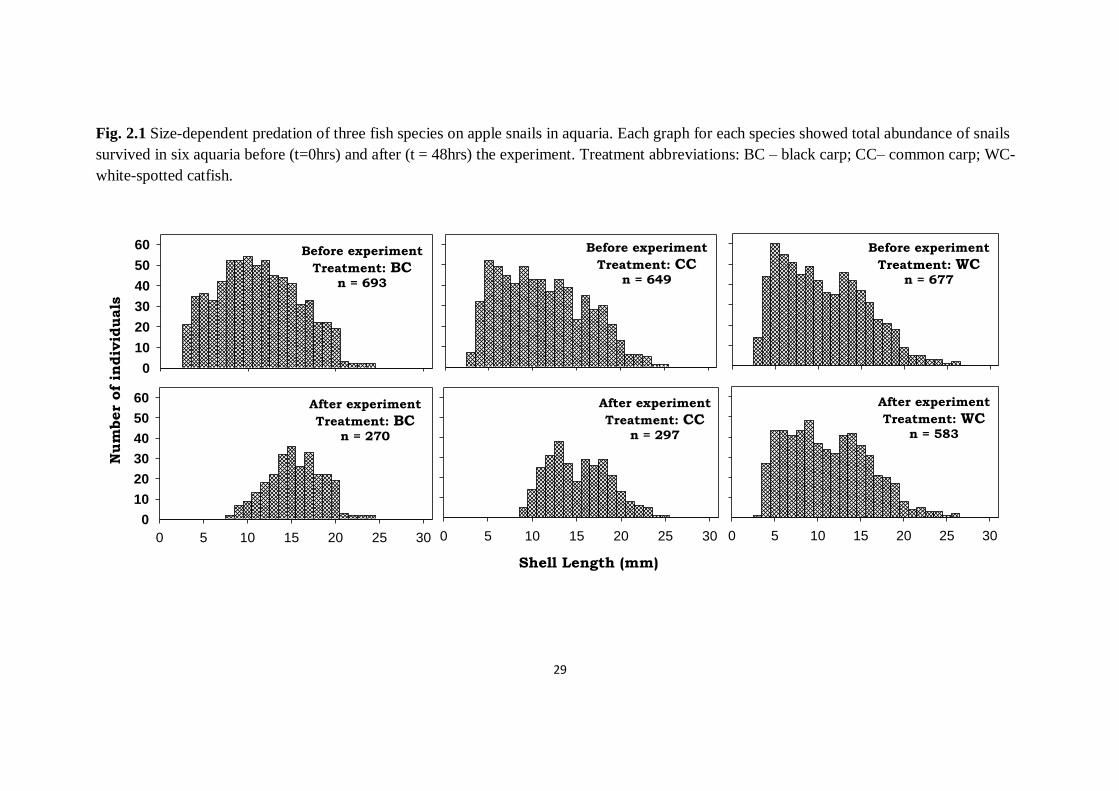
and black carp was 45.8% and 39.0% respectively (Fig. 2.1). Abundance of apple
snails after 48hours reduced significantly in all fish treatments, with compared to
that before treatments. Significant difference was found between total amount of
consumed apple snail among treatments (F3,20 = 118.50, p < 0.01). Results of
Tukey test comparing among treatments revealed significant difference in control
treatment and all treatments with fish. Among all with fish treatments, abundance
of apple snails consumed was not significantly different within treatments with
two species of carp, but that of treatment with catfish is significantly lower. On
average, estimated daily predatory rate of black carp was 35.25 individuals (SE =

25
6.28) while that of common carp was 29.33 individuals (SE = 3.78). Maximum
predatory rate for black carp and common carp was 45.0 and 31.5 individuals per
day respectively.
Predation of apple snails by two species of carp was size-dependent, with
juvenile snails being more susceptible. Snails within shell length (SL) of 3-8mm
were totally eradicated in both carp treatments (Fig. 2.2). Significant difference
was found in amount of juvenile snails (SL = 3 - 25mm) consumed in all
treatments (F3,20= 118.835, p <0.01*). Tukey test results indicated that both carp
species specimens consumed significantly larger amount of apple snails than that
by catfishes, though no significant differences were observed between results of
two species of carp (Fig. 2.3). In treatment tanks with catfish, only snails with
shell length below 6mm was consumed most significantly. This accounted for
50% of the mean total snail consumption while the rest scattered within juvenile
sized snails. For all fish species, no significant differences were drawn among
abundance of adult snails provided (SL >25mm) (F3,20= 1.0, p = 0.413).
Maximum size of ingestion by common carp was 18mm, which was higher
than 16mm in tanks with black carp. Yet, a point to note was that the maximum
size of ingestion by catfish was 26mm. This far exceeded that of the other two
carp species, though catfish consumed much less in quantity of snails.
2.4 Discussion
Fish in family Cyprinids do not have jaw teeth. Benthic dwellers, like black carp
and common carp share the same characteristics of being pharyngeal crushers,
having pharyngeal teeth, horny and callous pad specialized for preying upon

26
mollusks (Liu et al., 1990; Sibbing, 1988). In this study, black carp and common
carp demonstrated high predatory potential against apple snails. But common
carp possess protruding mouthparts allowing them to prey upon mollusks of
higher shell length, by sucking the soft tissues of mollusks with intact shells
(Sibbing et al. 1986). Given that omnivorous common carp has been
demonstrated as good example of biological control against apple snails (FAO
1998), molluscivorous black carp with similar predatory potential might also be
another option in controlling mollusk population with lower trophic interactions
with aquatic vegetation and other invertebrates. Further field manipulative
experiment would thus be needed for more in-depth study.
White-spotted walking catfish C. fuscus, an omnivorous native predator that
equipped with large broad flat head and huge mouthpart, has demonstrated weak
predatory effect to apple snails in this study. Indeed gastropods might only
compose a small portion in their natural diet. Local literatures noted small fish,
crustaceans, insects, planktons and rotten meat and plants as diet items of this
species (Man and Hodgkiss 1981; Lee et al. 2004). Though there were some
literatures supporting catfish including African catfish C. gariepinus (Coates
1984; Phonekhampheng et al. 2009) and Philippine catfish C. batrachus (Liao
2005) as suitable bio-control agents for snails, catfish are indeed opportunistic
predators having ontogenic diet shift upon different habitats. Moving targets,
including small fishes, worms, crustaceans and insects might be their optimal
prey items instead of gastropods (Cagauan and Joshi, 2002; Cui and Zhao, 2012).
To conclude, the present study has clearly demonstrated high potential of
common carp and black carp as predators of the golden apple snails, and that

27
white-spotted catfish has comparatively limited ability in preying upon the snails.
Both carp species are gape-limited predators and snails were first engulfed and
crushed before being swallowed. Indigenous White-spotted catfish have a wider
gape but have low diet preference towards exotic apple snails.
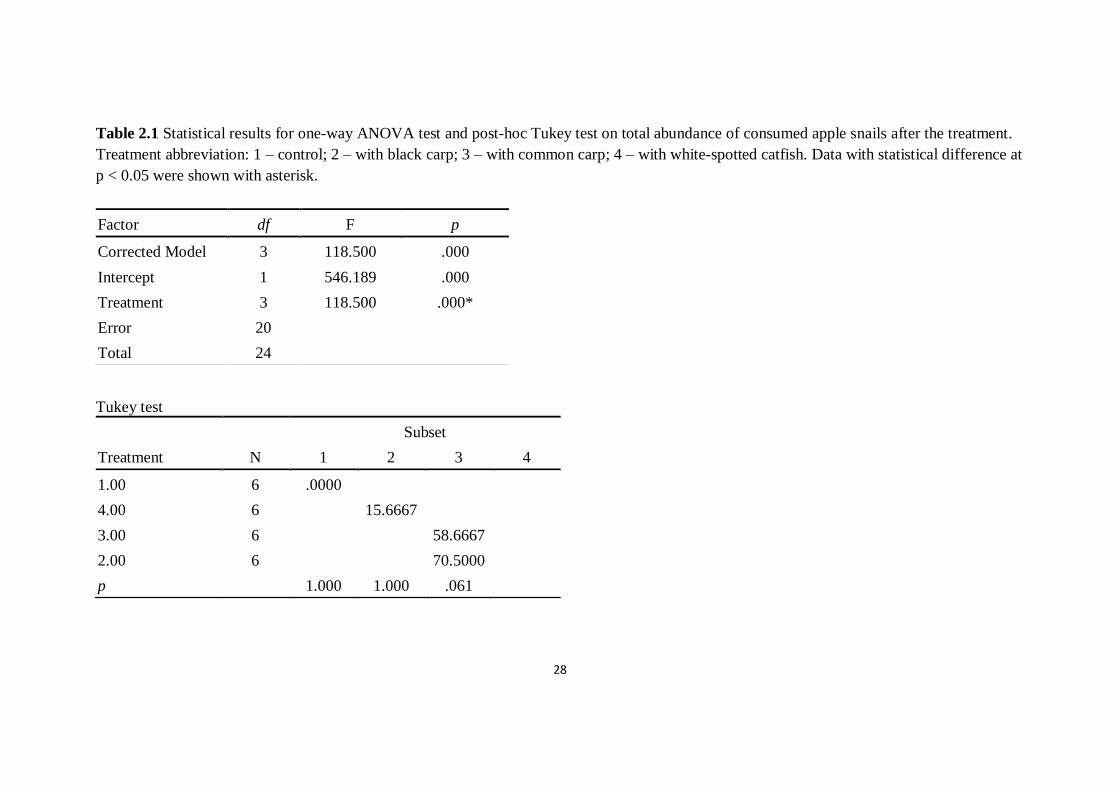
28
Table 2.1 Statistical results for one-way ANOVA test and post-hoc Tukey test on total abundance of consumed apple snails after the treatment.
Treatment abbreviation: 1 – control; 2 – with black carp; 3 – with common carp; 4 – with white-spotted catfish. Data with statistical difference at
p < 0.05 were shown with asterisk.
Factor df F p
Corrected Model 3 118.500 .000
Intercept 1 546.189 .000
Treatment 3 118.500 .000*
Error 20
Total 24
Tukey test
Treatment N
Subset
1 2 3 4
1.00 6 .0000
4.00 6 15.6667
3.00 6 58.6667
2.00 6 70.5000
p 1.000 1.000 .061
29
Fig. 2.1 Size-dependent predation of three fish species on apple snails in aquaria. Each graph for each species showed total abundance of snails
survived in six aquaria before (t=0hrs) and after (t = 48hrs) the experiment. Treatment abbreviations: BC – black carp; CC– common carp; WC-
white-spotted catfish.
Before experiment
Treatment: WCn = 677
0 5 10 15 20 25 30
After experiment
Treatment: WCn = 583
Shell Length (mm)
0 5 10 15 20 25 30
After experiment
Treatment: CCn = 297
Before experiment
Treatment: CCn = 649
Num
ber
of
indiv
iduals
0
10
20
30
40
50
60
0 5 10 15 20 25 30
0
10
20
30
40
50
60 After experiment
Treatment: BCn = 270
Before experiment
Treatment: BCn = 693
30
Fig. 2.2 Mean percentage of survival for golden apple snails in six aquaria, after
individual treatments with three species of fish. Treatment abbreviation as Fig.1.
0
20
40
60
80
100
BC
2D Graph 2
Pe
rce
nta
ge
su
rviv
ed
(%
)
0
20
40
60
80
100
CC
Shell length (mm)
0 5 10 15 20 25
0
20
40
60
80
100
WC

31
Fig. 2.3 Comparison of mean amount of snails consumed per experimental tank among three treatments. Treatment abbreviations: BC – Black
carp; CC – Common carp; WC: White-spotted walking catfish. Letters above the bars are results of Tukey test comparing the difference in mean
amount of golden apple snails consumed per tank among treatments. Individual analysis has been conducted for number of juvenile and adult
size class of apple snails consumed after the treatment. Different letter indicate significant difference at p < 0.05.
Treatment
BC CC WC
Mean
con
sum
ed
ap
ple
sna
il (indiv
idu
als
pe
r ta
nk)
0
20
40
60
80 Juvenile (3-25mm)
Adult (>25mm)
a
a
b
a a a

32
Chapter 3 Biological control of invasive apple snails in a
freshwater wetland by two carp species: subtle differences matter
3.1 Background
With the rapid development in travel and trade in the last century, there has been
a dramatic increase in the introduction of non-indigenous species worldwide
(Sala et al., 2000). While some introduced species can have economic benefits,
they can also become invasive, causing significant changes to the structure and
function of the invaded ecosystems through predation, competition, habitat
alteration and transmission of pathogens (Woodward and Hildrew, 2001;
Dudgeon et al., 2006; Gozlan et al., 2010; Paterson et al., 2011).
Biological control has been widely used as a tool to manage invasive
species, but there has been a long-standing debate regarding its benefits, risks
and costs (Cowie, 2001; Messing and Wright, 2006; Hulme, 2009; Strayer, 2010;
De Clercq et al., 2011). Critics of biological control argue that the risks are
difficult to predict due to the complexity of species interactions, and negative
impacts on non-target organisms and ecosystem functioning are often not
detected until it is too late (Howarth, 1991; Simberloff and Stiling, 1996).
Proponents of biological control emphasize that controlling or eradicating
invasive species benefits the economy, public health and ecosystems (Van
Driesche et al., 2010), and that precautionary procedures are sufficient to protect
non-target organisms (DeLoach, 1991). Very often, biological control is
perceived as more environmentally benign than chemical control, which is
usually expensive, non-selective and requires repeated application. This is
especially the case in the management of the apple snail, Pomacea canaliculata

33
Lamarck, an indigenous species of the freshwater wetlands of South America
that has invaded several Asian countries and Pacific islands including Hawaii
(Hayes et al., 2008). This species is a large freshwater gastropods and has a
voracious appetite for vegetation (Boland et al., 2008; Tamburi and Martín, 2009;
Qiu et al., 2011), causing tremendous losses to rice (Halwart, 1994b; Naylor,
1996) and taro agriculture (Cowie, 1995), as well as structural and functional
changes to non-agricultural wetlands (Carlsson et al., 2004a; Carlsson and
Lacoursière, 2005). Due to its huge economic and ecosystem impacts, this
species is listed among the 100 most notorious invasive species worldwide
(Lowe et al., 2000), and has become a prime target for control (Cowie, 2002;
Joshi and Sebastian, 2006).
Cultural, mechanical, chemical and biological methods have been used to
control populations of invasive apple snails (Litsinger and Estano, 1993; Cowie,
2002; Joshi and Sebastian, 2006). Cultural methods, such as changing from direct
seeding to transplanting older seedlings, and mechanical methods such as placing
grills at water inlets and hand-picking the snails, are effective but labour
intensive. Chemical methods, which involve the application of pesticides, are
expensive when used in large areas, and pose risks to human and ecosystem
health. Biological control using predators, especially common carp, has been
widely explored in the laboratory (Halwart et al., 1998; Teo, 2006), and rice
fields in Taiwan (Mochida, 1991), Vietnam (FAO, 1998), Japan (Ichinose et al.,
2002) and Malaysia (Teo, 2006). These experiments and field applications have
focused on effectiveness of control, without considering other potential
ecosystem effects. This focus is understandable as the studies were conducted in

34
rice paddies where biodiversity was low and the risk to non-target species might
be minor.
However, since P. canaliculata has also invaded non-agricultural wetlands
(Carlsson et al., 2004a; Kwong et al., 2008; Fang et al., 2010), the risks that
biological control agents may pose to non-target flora and fauna should be
carefully evaluated before releasing them to the field. Fish as predators of apple
snails in wetlands have been studied using enclosures, but the results were
unsatisfactory: the fish did not prey on apple snails but escaped from the
enclosures (Carlsson and Brönmark, 2006); the predation effect was not strong
enough to reduce herbivory by apple snails (Carlsson et al., 2004b); or the fish
were effective in controlling apple snails, but reduced macrophyte biomass as
much as did the apple snails and fed on macroinvertebrates in the sediment
(Wong et al., 2009). The present study was therefore conducted to compare the
performance of black carp (Mylopharyngodon piceus Richardson) and common
carp (Cyprinus carpio L.) as biological control agents of P. canaliculata in non-
agricultural wetlands. Black carp feeds on snails (Venable et al., 2000; Ben-Ami
and Heller, 2001). Five million juveniles of this species were released to control
P. canaliculata in rice planting areas in Taiwan, but its effectiveness was not
reported (Mochida, 1991). In this study, we compared not only the effectiveness
of the two biological control agents, but also their potential side effects on
wetland flora and fauna.

35
3.2 Materials and methods
Field site
The experimental site (22°26'40.69" N, 114°3'18.99" E) is in Kam Tin,
northwestern Hong Kong. The climate of Hong Kong is subtropical with a wet
season (April–September) and a dry season (October–March) (Dudgeon and
Corlett, 2004). The study lasted eight weeks from early October to early
December 2011, during which mean air temperatures declined from 23.7 °C in
the first week to 19.1°C in the last week. Total precipitation during the study
period was 301.3 mm, with 210.7 mm in the first 4 weeks and 86.1 mm in the
last 4 weeks. The experiment was conducted in a freshwater earthen wetland
created in 2002 as a compensation for the loss of the original wetland due to the
construction of a railway. The wetland was roughly oval, with a total area of 2.36
hectare. The bottom sloped gently from the edge to the centre, the deepest part
with over 2.0 m of water. The water is slightly alkaline (pH = 7.1), mildly
oxygenated (5.4 mg l-1
) and quite clear (turbidity = 2.4 NTU). The edge of the
wetland was vegetated with a variety of macrophytes, e.g. water dragon
(Ludwigia x taiwanensis Peng), water spinach (Ipomoea aquatica Forssk.),
bahiagrass (Paspalum sp.), knotgrass (Polygonum barbatum L.), common reed
(Phragmites australis Trin.). and bulrush (Typha angustifolia L.). From around
1.0 m to the centre, the wetland was unvegetated. The bottom was sandy clay,
with a community of macrobenthos dominated by the detritivorus snails Sinotaia
quadrata Benson (Viviparidae) and Melanoides tuberculata Müller (Thiaridae).
Pomacea canaliculata was present in the wetland at a low density (0.4 individual
m-2
, surveyed prior to the field experiment).

36
Experimental setup
The experiment was conducted in 18 1 x 1 x 1 m enclosures. There were five
replicates of three experimental treatments: with apple snails and black carp –
AS+BC; with apple snails and common carp – AS+CC; with apple snails but
without fish – AS; and three replicates of the control treatment without fish or
apple snails – C (originally five replicates but two enclosures were damaged by a
typhoon). The enclosures were placed in five haphazardly chosen blocks in the
shallow water (~ 0.7 m) near the shore (Supplementary material 1, Photographs
of the site showing the experimental enclosures), with each block containing one
enclosure of each of the four treatments (with the exception of two blocks in
which the original control enclosures had been damaged). Enclosures were made
of four woven plastic side-walls and a sewn nylon net bottom with a pore size of
200 m. An external frame, made of 4.3 cm PVC pipes and joints, was used to
support each enclosure. The lower part of the frame had four 15 cm legs, one at
each corner. To enhance the stability, the legs were pushed into the bottom
sediment, and a brick was tied to each lower corner of the enclosure. Surface
sediment (~ the top 3 cm of the substrate) was collected from the wetland for
introduction into the enclosures. The sediment was sieved through a 1.6 mm
mesh and washed into each enclosure to a depth of 3 cm. The residues retained
were then checked to remove apple snails by hand-picking. Other mollusks were
introduced into each enclosure at natural density (i.e. mollusks in 1 x 1 m surface
sediment). On the following three days, the enclosure walls and surface sediment
were swept to collect any apple snails that had been missed previously.
Afterwards, each enclosure was stocked with natural abundance of three species

37
of macrophytes common in local wetlands to test for the grazing effects of apple
snails and fish. Two of them were emergent (water dragon Ludwigia x
taiwanensis Peng, water spinach Ipomoea aquatica Forssk), whereas one was
floating (water lettuce Pistia striatiotes L.). Each plant was trimmed to
standardize the weight. The initial wet weight of the emergent species was 100
1 g, whereas that of the floating species was 50 1 g.
Black carp (fork length 170-185 mm) and common carp (fork length 170-
195 mm) were purchased from a commercial fish farm and an aquarium shop,
respectively. Both species are native to southern China, and are traditional
aquaculture species. The fish were allowed to acclimatize in enclosures in the
field for at least one week. One fish was introduced into each of the designated
enclosures. Enclosures were checked twice per week to ensure fish survival.
Replacement of fish was done immediately after dead specimen was observed.
Apple snails of different sizes were introduced into the enclosures. The
juveniles (shell length (SL): 8-15 mm) and adults (SL: 30-40 mm) were collected
from drainage channels of a vegetable garden in Yin Kong Tsuen, Sheung Shui
(22°30'22.08" N, 114°6'38.88" E), 15 km from the experimental wetland. They
were allowed to acclimatize in the field in an empty enclosure for at least 3 days
before use. Four adults (2 males and 2 females) and 20 juveniles were introduced
into each of the experimental enclosure in treatments AS, AS+BC and AS+CC
along with ten neonates (SL: 3 mm), obtained by allowing field collected egg
clutches to hatch in the laboratory. An additional 200 neonates were introduced

38
into each of these enclosures in the fifth week of the experiment to simulate the
hatching of one clutch of apple snail eggs.
Enclosure maintenance, data collection and analysis
During the experiment, the enclosures were covered with a nylon net (1 mm
mesh) to exclude birds and prevent the apple snails escaping. The enclosures
were inspected daily at the start of the experiment and roughly weekly after one
month to examine their integrity, the presence of grazing marks on the
macrophyte leaves and the health of the fish. Dead fish would float on the water
surface. Dead fish were removed and replaced with fish with the same species
and size ( 1 cm fork length of the dead one). Apple snail egg clutches laid on
the enclosure walls were recorded, removed from site and brought back to
laboratory. The number of eggs in each egg clutch was then counted after
separating the eggs in 1 M sodium hydroxide solution (Joshi et al., 2002)
At the end of the 8-week experiment, macrophytes were collected from
each enclosure, blotted dry with a paper towel, and each species weighed to the
nearest 0.1 gram. Macrophytes from each enclosure were weighted separately in
the field and then placed in separate Ziploc bags. They were brought back to the
laboratory, stored temporarily in a cold room at 4C, and later examined to
collect juvenile apple snails hidden among the roots. After removing the
macrophytes, the enclosures were detached from the supporting frames and
hauled ashore. Fish were collected and their length and gape width measured.
The enclosures were then transported to the laboratory where they were stored at
4C, and within 3 days the contents were washed into a sieve with a pore size of

39
500 m, in order to retrieve mollusks. The contents retained on the sieve were
stained with 0.01% Rose Bengal to facilitate sorting and preserved in 70%
ethanol. The preserved samples were then sorted to recover mollusks, which
were then identified to the lowest possible taxonomic level counted and the shell
length and width measured to the nearest 0.01 cm using digital calipers.
Differences among the four treatments were compared using one-way
randomized block ANOVA followed by the Tukey test. Parameters compared
included the number of survivors for each of the two size classes of apple snails
(juveniles and adults), the number of egg clutches deposited and the total number
of eggs, the biomass of each of the three species of macrophytes, total
macrophyte biomass, and the abundance of non-Pomacea mollusks. The t-test
was used to compare difference in predation of the two most abundant non-
Pomacea mollusks by the two species of carp.
3.3 Results
Fish survival and effect on apple snail abundance
All black carp in the enclosures survived through the 8-week experiment.
Common carp died in three of the five replicates, and in total six individuals died.
The mortality occurred during the first month and thereafter the replacement
individuals survived well.
Of the 234 apple snails introduced into each of the enclosures, the mean
survival rate was as high as 93% in the control, but only 5% in the common carp
treatment and 2% in the black carp treatment. Apple snail mortality was size-
dependent, with smaller individuals suffering the most severe mortality (Fig. 3.1).

40
In the AS+CC treatment, the survivors were restricted to individuals ≥ 0.8 cm
shell width (SW). In the AS+BC treatment, except for 6 individuals of 0.3 cm
SW, all survivors were ≥ 2.0 cm SW. Statistical analysis showed that the
abundances of juvenile (F2,12 = 165.619, p < 0.001) snails in the treatments with
carp were significantly reduced when compared with the control. However, the
abundance of adult apple snails that survived through the experiment did not
differ significantly among the treatments (F2,12 = 0.105, p = 0.901). The Tukey
tests showed that the abundance of the two size classes of apple snails did not
differ significantly between the two fish treatments, even for juvenile snails for
which the mean value in the AS+BC treatment (1.2 individuals) was much lower
than the mean value in the AS+CC treatment (8.6 individuals) due to the great
variation among replicates (range: 0–17 individuals; S.D.: 7.02). The number of
apple snail egg clutches and the total number of eggs laid during the experiment
varied substantially among replicates in each treatment (AS: 1–3 clutches,146–
393 eggs/enclosure; AS+CC: 0–3 clutches, 0–396 eggs/enclosure; AS+BC: 0–2
clutches, 0–316 eggs/enclosure) but there was no significant difference in either
reproductive parameter among the three treatments (number of clutches: F2,12 =
0.20, p = 0.821; number of eggs: F2,12 = 0.426, p = 0.663).
Predation on non-Pomacea mollusks by carp
Seven species of non-Pomacea mollusks were recovered from the enclosures,
two bivalve species (Corbicula fluminea Müller., Anodonta woodiana Lea.), four
operculate snail species (Sinotaia quadrata, Melanoides tuberculata, Bithynia sp.,
Segmentina succinea Gredl.) and one pulmonate snail species (Physella acuta
Draparnaud.). M. tuberculata ranked first and S. quadrata ranked second in

41
abundance, representing 44% and 33% of the overall abundance, respectively
(Fig. 3.2). The overall abundance of these non-Pomacea mollusks differed
significantly among the four treatments (one-way ANOVA, F3,14 = 8.628, p =
0.002). The Tukey tests showed that the four treatments were clustered into two
subsets (C, AS) and (AS+CC, AS+BC).
Comparison of the number of the two most abundant non-Pomacea
mollusks across the treatments showed that black carp exerted a stronger
predatory effect than common carp, and in both treatments M. tuberculata was
more susceptible than S. quadrata to fish predation (Fig. 3.2). Comparing with
the treatment with apple snails, the abundance of S. quadrata was reduced by
74.3% in enclosures with black carp, while only by 31.8% in enclosures with
common carp. In contrast, the abundance of M. tuberculata reduced by 95.7%
and 67.5% in enclosures with black carp and common carp, respectively. The t-
test detected significantly fewer S. quadrata (t = 3.373, p = 0.012) and M.
tuberculata (t = 2.657, p = 0.033) in the black carp treatment than in the common
carp treatment. To determine whether there was size-dependent predatory effect
on the two most abundant snails, abundance data from all replicates from each
treatment were pooled to show the size frequency distribution (Fig. 3.3). For S.
quadrata, the small individuals (< 1 cm SW) were clearly eaten by both fish
species. For M. tuberculata, there appeared to be no size refugium from
predation due to its elongate shell with a small maximum width, but the mortality
of this snail in the black carp treatment was much higher than in the common
carp treatment.

42
Treatment effects on macrophytes
The three species of macrophytes had different patterns of growth and loss to
herbivory (Appendix: Plate 1). In the control, the biomass of water spinach (I.
aquatica) and water dragon (L. x taiwanensis) increased by 0.5- and 3-fold
respectively during the experiment (from the initial 100 g per replicate), but that
of water lettuce (P. stratiotes) declined to only 41% of its initial value (50g) (Fig.
3.4). Of these three species, the greatest reduction due to herbivory by apple
snails or apple snails plus carp occurred in L. x taiwanensis, the species that had
the highest increase in biomass in the control. There was a great increase in the
total macrophyte biomass in the control, from 250 g in the beginning to 542.6 g
at the end (Fig. 3.4). Macrophyte biomass in treatments with apple snails or apple
snails plus carp was remarkably lower than the control: only 30% in the AS
treatment, 32.5% in the AS+CC treatment and 56% in the AS+BC treatment. The
total plant biomass was significantly different among treatments (F3,16 = 38.746,
p < 0.001). The Tukey tests showed that the difference between the AS and
AS+CC treatments was not significant (p = 0.988), but plant biomass in the
AS+BC treatment was significantly higher than that of the AS and AS+CC
treatments (p < 0.05).
3.4 Discussion
Our study shows that the invasive apple snail P. canaliculata can exert a strong
and selective grazing pressure on wetland macrophytes. This is consistent with
the results from previous field enclosure studies (Carlsson et al., 2004a; Wong et
al., 2009). Laboratory experiments show that P. canaliculata can consume many
species of macrophtytes with clear feeding selectivity (Estebenet, 1995; Lach et

43
al., 2000; Boland et al., 2008; Tamburi and Martín, 2011), with its feeding rate
being determined by the nutrient (nitrogen and phosphorus) and defense
chemical (phenolics) contents of the macrophyte leaves (Qiu and Kwong, 2009;
Wong et al., 2010; Qiu et al., 2011). Such strong and selective grazing may alter
floral composition, and thus highlights the need to control P. canaliculata
population in non-agricultural freshwater wetlands.
Although most attempts to control apple snail populations by biological
methods have been undertaken in agricultural areas due to the obvious need to
protect crops (Liao et al., 2005; Teo, 2006), with the realization that apple snails
can reduce macrophtyte biomass and alter floral structure in non-agricultural
wetlands (Carlsson et al., 2004a), biological control of apple snails in these more
natural ecosystems has been explored in two mesocosm studies conducted in the
field (Carlsson et al., 2004b; Carlsson and Brönmark, 2006). In one of the studies,
Carlsson et al. (2004b) used climbing perch Anabas testudineus (Bloch) as a
predator of P. canaliculata, and examined the amelioration of the snail’s
herbivory due to the biological control. Their results were encouraging as the
perch showed some level of control by feeding on the neonate apple snails with
2-3 mm SL. Predation on these small apple snails, however, did not reduce the
overall herbivory on macrophytes when compared with the no-fish treatment.
Furthermore, potential predation on non-target benthic invertebrates was not
examined in these studies.
The present study not only determined the effectiveness of common carp
and black carp as potential predators of apple snails, but also examined the side-
effects on wetland benthic invertebrates and macrophytes. In a previous study

44
using similar experimental design that included common carp as the only
predator (Wong et al., 2009), we found that common carp removed almost all the
small and medium-sized P. canaliculata from the enclosures, being much more
effective than the climbing perch and catfish (Carlsson et al., 2004b; Carlsson
and Brönmark, 2006). However, common carp also greatly reduced the biomass
of macrophytes by herbivory, as well as the abundance of non-Pomacea snails by
predation. The present study, conducted in a different wetland with somewhat
different floral and faunal composition in the enclosures, confirmed common
carp’s desirable predatory effect on apple snails and undesirable side effects on
macrophytes and non-target invertebrates.
Using black carp as another potential biological control agent for apple
snails in this study has allowed us to conduct a side-by-side comparison with
common carp. Both species are effective in removing the apple snails, but
perhaps black carp is a stronger predator of the snails as it fed on more apple
snails than common carp. Although common carp had a wider gape size than
black carp (mean ± S.D.: 17.1± 0.6 mm vs. 15.9 ± 0.5mm), black carp still
consumed a wider size spectrum and higher quantity of apple snails than
common carp. Both fish were unable to feed on large-sized apple snails, but their
long life span of up to 13 years (Nico et al., 2005), their ability to consume larger
apple snails as the carp grow, and the high natural mortality of large-sized apple
snails (Kwong et al., 2010) should make both species of carp effective biological
control agents in Asian freshwater wetlands. Compared with common carp,
however, black carp appeared to be more beneficial as it preys on invertebrates
only, whereas common carp feed on both invertebrates and macrophytes.

45
Black carp was used to control snails (Melanoides tuberculata and Physella
acuta) in an outdoor concrete pond in Isreal (Ben-Ami and Heller, 2001), as well
as Planorbella trivolvis in a commercial catfish culture pond in the USA
(Venable et al., 2000). However, in a field application involving the use of both
carp species to control P. canaliculata in the paddy fields of Vietnam, black carp
had a lower survivorship than common carp (FAO, 1998). This is probably
because black carp has adapted to live in lakes and middle or lower stretches of
rivers (Pan et al., 1991). It is more sensitive to the overheating commonly
associated with the shallow water of paddies during summer in Southeast Asia.
Thus the water level was raised to 40 cm when black carp were used to control P.
canaliculata in a water bamboo field in Taiwan (Liao et al., 2005). In the present
study, overheating was probably not a problem as the part of the wetland where
we conducted the experiment was around 0.7 m deep, and there was no mortality
of black carp. However, common carp introduced into three of the five
enclosures died. Since the mortality occurred during the first month of study,
longer acclimation in the field might have improved the survival of common carp.
Although we replaced the fish as soon as possible, in one of the enclosures where
the replacement fish also died, substantial number of non-Pomacea snails
survived to the end of the experiment (498 individuals). Nevertheless, in the
other four replicates, common carp clearly exerted strong control of the non-
Pomacea mollusks, with only 160 ± 45 individuals (mean ± S.D.) surviving to
the end of the experiment.
In conclusion, our study has clearly shown that both black carp and
common carp are effective predators of the invasive apple snail P. canaliculata

46
in non-agricultural wetlands. However, the two fish species exhibited subtle
differences in their effects on non-target organisms. Black carp prey on non-
target mollusks, whereas common carp feed on both non-target mollusks and
macrophytes. These subtle differences should be considered before adopting the
fish as management tools for invasive apple snails. For instance, if wetland
macrophyte diversity is a major management concern, black carp would be a
better choice than common carp. If wetland invertebrate biodiversity is a major
concern, mechanical and cultural control measures should be included in long
term management plan, and perhaps neither species of carp should be used to
control apple snails. Our empirical study, driven by the need to control an
invasive species, has provided solid evidence to support the call for considering
the complex predator-prey interactions when evaluating the potential of a
predator as a bio-control agent (Cowie, 2002).
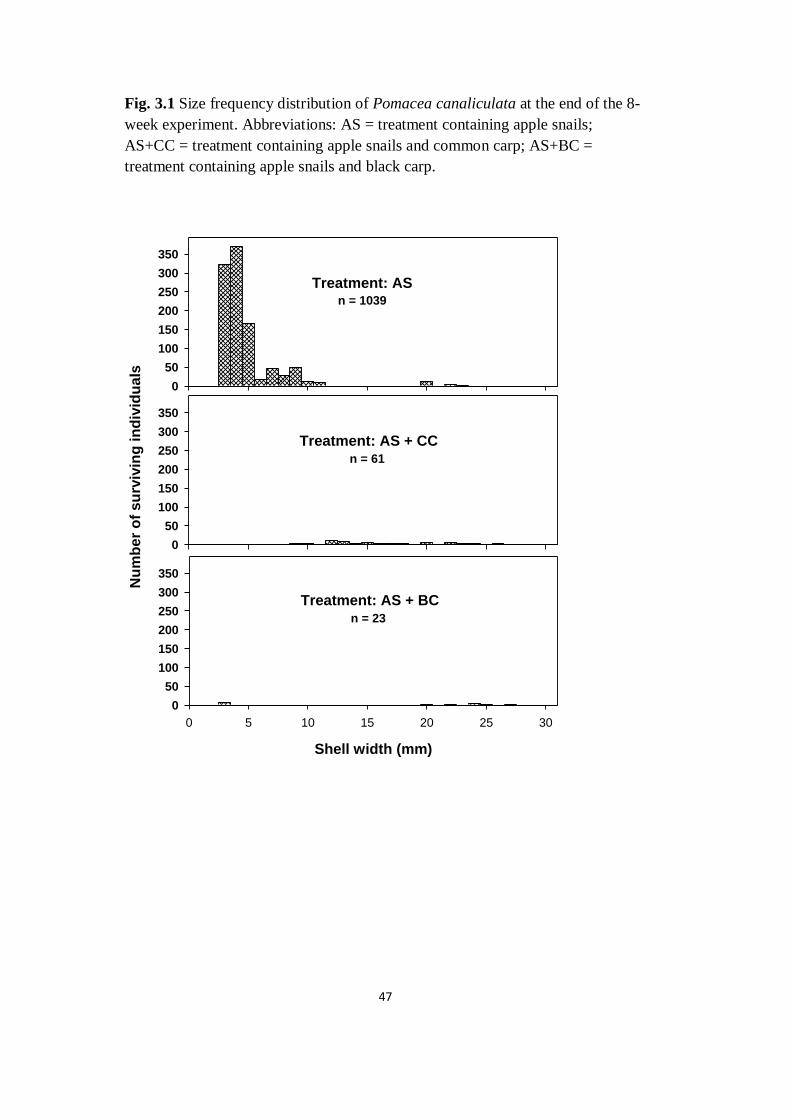
47
Fig. 3.1 Size frequency distribution of Pomacea canaliculata at the end of the 8-
week experiment. Abbreviations: AS = treatment containing apple snails;
AS+CC = treatment containing apple snails and common carp; AS+BC =
treatment containing apple snails and black carp.
0
50
100
150
200
250
300
350
Treatment: ASn = 1039
Nu
mb
er
of
su
rviv
ing
in
div
idu
als
0
50
100
150
200
250
300
350
Treatment: AS + CCn = 61
Shell width (mm)
0 5 10 15 20 25 30
0
50
100
150
200
250
300
350
Treatment: AS + BCn = 23
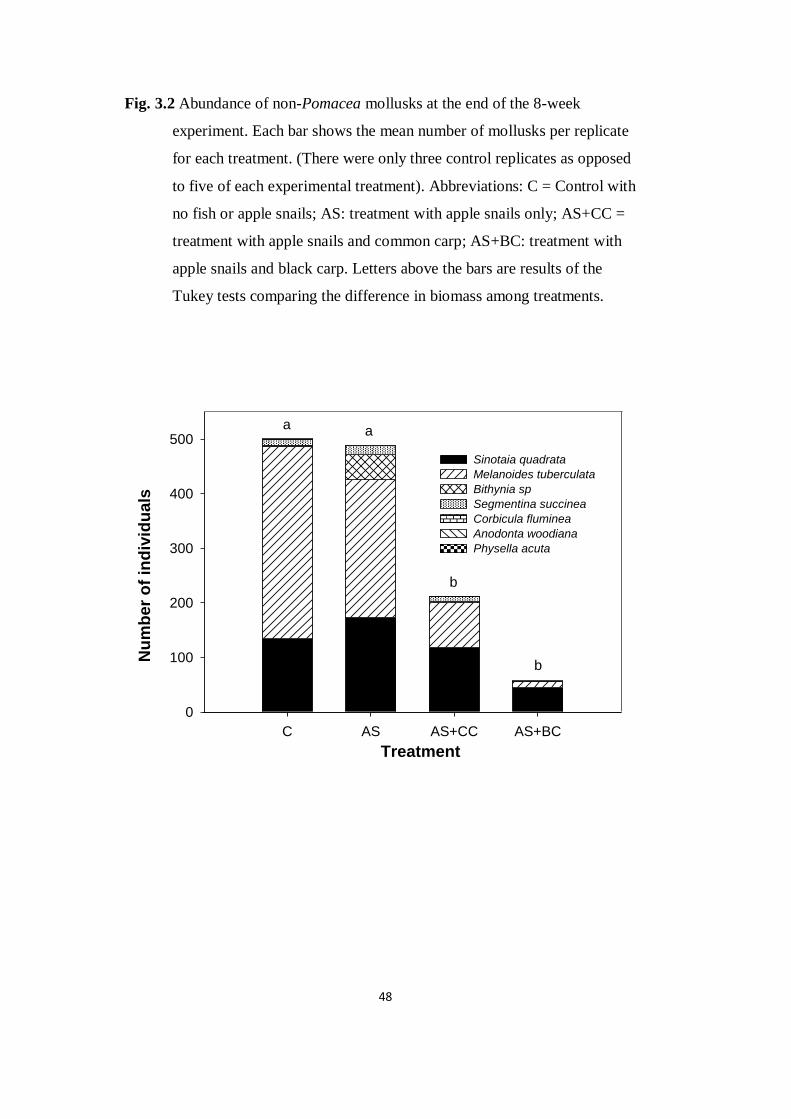
48
Fig. 3.2 Abundance of non-Pomacea mollusks at the end of the 8-week
experiment. Each bar shows the mean number of mollusks per replicate
for each treatment. (There were only three control replicates as opposed
to five of each experimental treatment). Abbreviations: C = Control with
no fish or apple snails; AS: treatment with apple snails only; AS+CC =
treatment with apple snails and common carp; AS+BC: treatment with
apple snails and black carp. Letters above the bars are results of the
Tukey tests comparing the difference in biomass among treatments.
C AS AS+CC AS+BC
0
100
200
300
400
500
Sinotaia quadrata
Melanoides tuberculata
Bithynia sp
Segmentina succinea
Corbicula fluminea
Anodonta woodiana
Physella acuta
Treatment
Nu
mb
er
of
ind
ivid
uals
a a
b
b
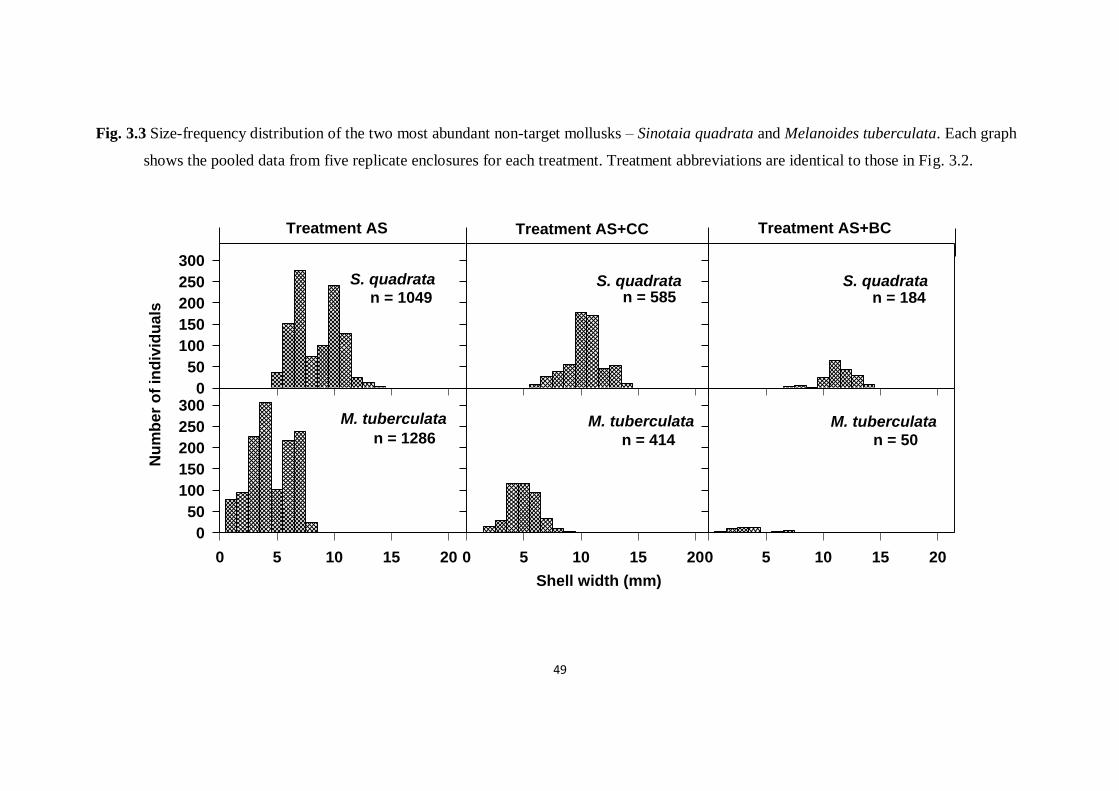
49
Fig. 3.3 Size-frequency distribution of the two most abundant non-target mollusks – Sinotaia quadrata and Melanoides tuberculata. Each graph
shows the pooled data from five replicate enclosures for each treatment. Treatment abbreviations are identical to those in Fig. 3.2.
M. tuberculata
0 5 10 15 20
0
50
100
150
200
250
300M. tuberculata
0 5 10 15 200 5 10 15 20
S. quadrata
0
50
100
150
200
250
300
n = 1049
n = 414
Shell width (mm)
Treatment AS Treatment AS+CC Treatment AS+BC
n = 1286
Nu
mb
er
of
ind
ivid
uals
n = 50
S. quadratan = 585 n = 184
S. quadrata
M. tuberculata
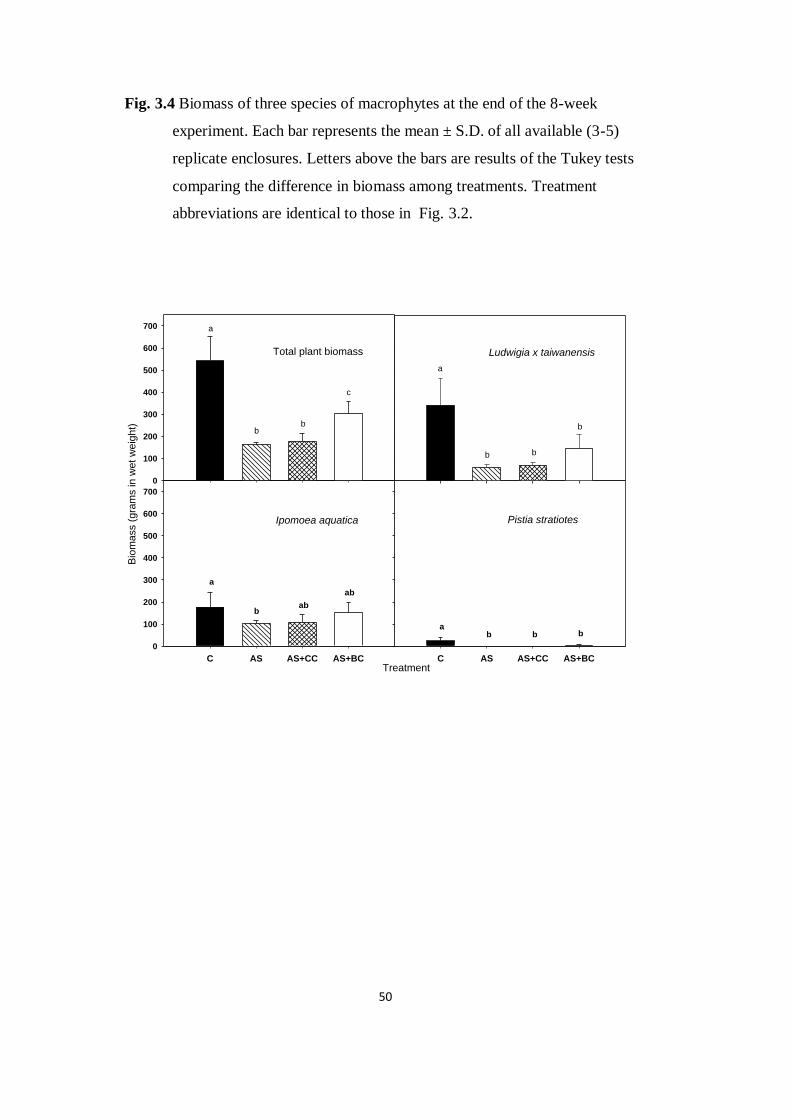
50
Fig. 3.4 Biomass of three species of macrophytes at the end of the 8-week
experiment. Each bar represents the mean ± S.D. of all available (3-5)
replicate enclosures. Letters above the bars are results of the Tukey tests
comparing the difference in biomass among treatments. Treatment
abbreviations are identical to those in Fig. 3.2.
TreatmentC AS AS+CC AS+BC
0
100
200
300
400
500
600
700
C AS AS+CC AS+BC
Total plant biomassLudwigia x taiwanensis
Ipomoea aquatica Pistia stratiotes
a
bb
c
a
bab
ab
a
b
a
a
ab b b
Bio
mass (
gra
ms in w
et w
eig
ht)
0
100
200
300
400
500
600
700 a
bb
c
a
b b
b
Total plant biomass Ludwigia x taiwanensis

51
Chapter 4 Wetland scale application of biological control of apple
snails using black carp: efficacy and subtle effect to ecosystem
4.1 Background
Invasive mollusks have altered many aquatic ecosystems, either by replacing
indigenous species through predation or competition, coexisting with indigenous
species though facilitation, or colonizing modified habitats where indigenous
species have already been eradicated by abrupt environmental changes (Cowie,
1998; Ricciardi et al., 1998). However, whether an introduced species could
eventually establish to become invasive is dependent on both the characteristics
of the recipient ecosystem such as diversity, resource abundance, frequency and
scale of disturbances (Davis and Pelsor, 2001) and also propagule pressure
(Simberloff, 2009) and life-history traits of the introduced species such as dietary
flexibility and tolerance to adverse environmental conditions (Kolar and Lodge,
2001). In Asia, three species of apple snails (Pomacea canaliculata Lamarck, P.
maculata Perry and P. scalaris D’Orbigny) have been introduced from South
America (Anderson, 1993; Jhang, 1985; Joshi, 2007; Yusa and Wada, 1999;
Joshi et al., 2001; Chen et al. 2004; Cowie et al., 2006; Hayes et al. 2008; Hayes
et al., 2012; Lv et al., 2013). Among them, two (P. canaliculata and P. maculata)
have become highly successful agricultural pests (Joshi, 2005; Joshi, 2007), but
their impact has currently proven to extend beyond agricultural context once
established and spread. As unique grazers with enlarged jaws with radula and a
digestive system with amylase for macrophyte digestion, these apple snails have
invaded natural wetlands by reversing succession pressure, promoting growth of
phytoplankton by releasing nutrients stored in living aquatic plants (Carlsson et

52
al., 2004) and drive shallow lakes towards turbid conditions (Jones and Sayer
2003; Carlsson and Lacoursière, 2005). In addition, wetland plant and
phytoplankton diversity can be altered by the feeding selectivity of P.
canaliculata that exhibit clear selection over macrophytes with high nitrogen
content, low dry matter content and chemical defense (Qiu and Kwong, 2009;
Wong et al., 2010). As most indigenous snails are mainly detritivores (Dudgeon
and Yipp, 1985), apple snails have successfully established its own niche in
consumption of underutilized wetland resource - riparian macrophytes in Hong
Kong and South-east China.
Fish, especially carp, is natural predators that regulate macroinvertebrate
density and diversity. Use of fish as bio-control agents for trematode and
schistosomiasis-hosting snails was common practice in human-modified habitats,
like fishponds and paddyfields (FAO, 1998; Lardans and Dissous, 1998). In this
aspect, common carp (Cyprinus carpio L.), Nile tilapia (Oreochromis niloticus L.)
and catfishes (Clarias spp.) have been most widely studied and implemented in
Asia (Ali, 1992; Fernando, 1993; Halwart, 1998; Liao et al. 2005; Teo, 2006).
Yet, they are omnivores that feed upon macroinvertebrates and aquatic plants
(Batzer et al., 2000), which in turn might alter original ecosystem functioning.
Chinese carp species are indigenous to riverine and floodplain habitats and have
high degree of niche segregation including specialized diet regime (Dudgeon,
1999b). Black carp (Mylopharyngodon piceus Richardson) are one of the most
abundant residents of the Great rivers (Nanjing Institute of Geography, Academia
Sinica, 1981; Liu et al., 1986; Qiu et al., 2002). They are traditionally reared in
farm ponds (Wu et al., 1964), fish ponds (Hickling, 1971) and rice paddies

53
(Wang, 2010). Other than being food fish to generate profit in complement with
that from crops yield (Lin, 1955; Liu, 1955), Black carp is also widely used as
biocontrol agents of pest mollusks.
Previous field experiment using black carp as controlling agent, however,
only addressed the applicability of this fish to slow down the invasion of snails
bearing disease-causing agents in semi-natural wetlands and within a short term
(less than a year). Experiments were conducted mainly in deeper waters, such as
nursery ponds (Hung et al., 2013), fish ponds (Venable et al. 2000) and reservoirs
(Ben-Ami and Heller, 2001). Only one experiment was conducted with apple
snails in Taiwan in shallow water paddy fields (Liao et al .2004). A large-scaled
project to control pest snails was implemented in Israeli reservoirs using black
carp, but the effects of black carp on non-target macroinvertebrates and water
quality were not evaluated (Leventer, 1979). Above all, no study has been
conducted to evaluate the application of black carp as a bio-control agent in
shallow water non-agricultural wetlands where non-target effects on macrophytes
and indigenous invertebrates might be of management concern. Nor did they
consider the effects of season on apple snail reproduction and development, and
consequently on the effectiveness of the biological control.
This field study was conducted in a constructed freshwater wetland being
infested by apple snails. Specifically, the objectives for experiment were: first, to
determine the effectiveness of black carp on apple snails in natural settings;
second, to determine the direct predatory effect of black carp on non-target
macroinvertebrates and indirect effect of predation of apple snails on macrophyte
diversity and biomass, as well as macroinvertebrates and water quality.

54
4.2 Study site
The study was conducted in three constructed freshwater wetlands that are nearly
permanent lentic ponds. The study lasted one year encompassing the wet season
(June - September 2012 & April - June2013) and dry season (October 2012 -
March 2013). Mean monthly air temperature declined from a maximum of
30.5°C in June 2012 to a minimum of 15.9 °C in December 2012. Total
precipitation during the study period was 2595.5 mm (till June 2013), with
1855.2 mm in the wet season and 307.1 mm in the dry season. These constructed
wetlands are all freshwater earthen ponds created in 2002 as a compensation for
the loss of the original wetlands due to the construction of the West Rail. Mean
water depth ranged from 1.1 – 2.2 m in the wet season; and 0.6 – 1.3 m in the dry
season. Site 1 and site 2 were roughly rectangular, with a total area of 2525
m2and 1000 m
2 respectively.
A partition was set up with woven nylon clothes supported by a steel frame
in the middle of each of these two sites, creating one experimental plot and one
control plot. The partitions prevented snails from migrating and minimized water
exchange between the control and treatment plots. Site 3 was comprised of two
separated but immediately adjacent ponds; one with an area of 450 m2 served as
control and the other with an area of 625 m2 as treatment. In general, the water in
sites 1 and 2 was mildly acidic (6.4 ± 0.44), mildly oxygenated (4.8 ± 2.3 mg/L)
and quite turbid (20.3 ± 11.9 NTU). Site 3 was quite hypo-oxygenated (2.39 ±
2.44 mg/L) at the initial stage. In subsequent months during dry season 2012,
mechanical water pumping was conducted to maintain stable water levels within
all plots above 0.5 m. Dissolved oxygen levels in Site 3 had thus been raised and
maintained at approximately 3 mg/L until the experiment ended in June 2013.

55
Several emergent macrophytes dominated the sites. These included
Panicum dichotomiflorum Michaux, Paspalum distichum Linnaeus, Polygonum
barbatum Linnaeus, P. lapthifolium L. and Ipomoea aquatica Forsskål.
Apple snail P. canaliculata inhabited the three ponds at initial mean
densities of 24.67 – 39.3 individuals per m2, and were similar between each of
the control and treatment pairs. Indigenous snails co-existing with apple snails
within our study site were all common residents at lowland lentic wetlands of
Hong Kong. These included Sinotaia quadrata Benson, Melanoides tuberculata
Müller and Bithynia sp. in sites 1 and 2. For site 3, only exotic snail Segmentina
succinea co-existed with apple snails. In April and May 2012 before experiment
commenced, various attempts of stocking S. quadrata and M. tuberculata were
made in Site 3 as these two species are very common in local lentic water. It was
thus important to include these two species to test the non-target effects of carp
predation. However, since subsequent field surveys did not show that these two
species successfully colonized Site 3, no further reintroduction was conducted.
4.3 Method
Black carp specimen preparation and collection
Thirty individuals of black carp were obtained from a local fish farm in Sheung
Shui, New Territories. They were kept in an indoor aquarium and fed with
commercial fish feed for at least one month before the experiment. One day prior
to the experiment, no food was given to all specimens. 12 specimens were
randomly selected and transported to site. Four black carp were released into
each experimental plot (treatment), whereas no fish were introduced into the
control. The average total length and wet weight for black carp was 294 mm

56
(S.D.: 17.40mm) and 231.3g (S.D.: 42.13g), respectively. Experiment started
immediately once black carp were released to each experimental plot. Fish
specimens were not fed during the experimental period.
At the end of the experiment, fish were extracted by drying the whole site and
subsequent extensive netting. They were euthanized and freezed as soon as they
were returned to laboratory. Respective total length, weight and gape width were
measured for each retrieved fish specimen. Within the subsequent three days
after retrieval, fish extracted from each of the three sites were dissected to
remove their stomach. Each stomach was then preserved in 10% formalin. Prey
composition was determined by dissecting the foregut of each stomach, where
there was least active amount of digestion and allowed easier identification. Gut
fullness was estimated visually by a dissecting microscope. Indirect volumetric
analysis was deployed in estimating the percentage of different food items in
comparing with the total volume of all stomach content (Hyslop 1980).
Seasonal field surveys
Field surveys were conducted from June 2012 to June 2013 to test the
effectiveness of black carp as a bio-control agent for apple snails, as well as their
non-target effects on macrophytes and benthos. In these surveys, macrobenthic
community analysis and water quality measurement were conducted every three
months, while macrophyte composition was determined only in the beginning
and at the end of the study, due to low water levels in the dry season which
prevented apple snails from feeding on the macrophytes.
Macrobenthic survey

57
The benthic surveys were conducted at fixed points marked with bamboo sticks.
In each plot, six 0.5 m2 were sampled by enclosing the area with a throw trap and
intensively collecting the macrobenthos on the sediment surface, in the sediment
as well as on the macrophytes using a hand-net with a mesh size of 500μm. A
total of 36 benthic samples were collected from the six sites (i.e. three control
and three treatment sites) during each sampling. The samples stored in numbered
Ziploc bags and were transported to Hong Kong Baptist University, and kept in a
cold room at 4ºC before further processing.
Sieving was conducted with a metallic sieve (pore size 500μm) within three
days after sampling to remove fine sediment. Macrobenthos were extracted from
the residuals retained on the sieve. They were identified into the lowest
taxonomic level and counted. One sample for each identified taxon was
preserved in 70% ethanol for detailed identification. For any species of identified
mollusk, its amount within each sample was enumerated, with shell length of
each individual measured. Same procedures applied to any species of identified
invertebrates, except that their sizes were not measured.
Determination of water quality parameters
Water quality parameters were either measured in-situ or in the laboratory. In-
situ measurements were conducted for temperature, pH, salinity, dissolved
oxygen, conductivity and turbidity. Temperature, pH, salinity, dissolved oxygen,
conductivity and turbidity were measured using a TPS 90-FLMV meter, whereas
turbidity was measured using a Thermo Scientific Orion AQ4500 meter. A total
of three measurements were taken for each parameter in each site. Two 1-L water
samples were also collected from each treatment and control site for laboratory

58
measurement of nutrient parameters (ammonia nitrogen, total nitrogen, reactive
phosphate and total phosphorus). A total of 12 water samples were collected
from three sites during each sampling. They were kept in a cool box with ice and
immediately transported to the laboratory for analysis using standard methods
(Eaton and Franson, 2005): total nitrogen (TN) using a flow injection analyzer
(Method 4500 N B); ammoniacal nitrogen (NH3) using a flow injection analyzer
(Method 4500-NH3 H); total phosphorus (TP) and total phosphate (P) using the
ascorbic acid method (Method 4500-P B & E; Method 4500-P E respectively).
Abundance of apple snail hatchlings
The number of neonates that might enter the pond was estimated by determining
the number of eggs available in the ponds and estimating their hatchability. The
number of eggs available was determined by counting the actual number of egg
clutches observed (a walkover survey throughout the whole pond bund during
each sampling time). Once an egg clutch was detected, we measured the
maximum length and width to the nearest 0.1 mm using digital calipers. This was
repeated until all egg clutches in each site were measured accordingly. The
number of eggs in each egg clutch was estimated by a regression relationship
between maximum area (maximum length* maximum width) and the actual
number of eggs in each egg clutch from previous literature as follows (Ichinose
et al., 2002).
Y = 0.1 * X1.24
(r2 = 0.75, n=272),
where X is the product of the maximum length and width of the egg masses and
Y is the number of eggs.

59
Hatchability of a given egg mass was estimated by hatching field-
collected egg clutches in the laboratory. In each month of the experiment, 25 - 30
egg masses were collected from wetlands within 200 m from our experimental
ponds. Five egg clutches were placed on a steel rack on top of a plastic container
(total number of containers = 5 - 6) with de-chlorinated water, and allowed to
hatch for 21 days at a rooftop. At the end of the experiment, the hatched neonates
and non-hatched eggs were counted. Monthly mean hatchability data were then
obtained. The estimated hatchling number from all individual egg clutches was
then estimated by the following equation:
N = Σ Hhatch * Y
where Hhatch is the monthly mean hatchability and Y is the number of eggs
estimated from egg clutch.
Macrophytes
Macrophyte surveys were conducted to understand the indirect effects of black
carp on macrophytes, or whether controlling apple snails would enhance
macrophyte biomass and diversity. Since macrophytes were present only on the
edges of the ponds during the wet season when the water levels are sufficiently
high, surveys were conducted in the beginning and at the end of the experiment
only. Eight 1 m x 1 m permanent quadrats were set up in each plot of each site
with the corners marked with bamboo sticks. A total of 48 1m2 quadrats were
established along in the six plots. In each quadrat, vegetative cover (%) was
estimated by visual observation, and the species composition was recorded.
Data analysis

60
Hatchling numbers were compared among sampling seasons and treatments
using a two-way Analysis of Variance (ANOVA). Change in apple snail
abundance over time from the same set of replicates was compared using
repeated measures ANOVA.
Abundance of juvenile apple snails was compared between the control and
treatment among the three sites using repeated measures ANOVA to determine
the effects of black carp. Abundances of three major non-target mollusks –
Sinotaia quadrata, Melanoides tuberculata and Bithynia sp. were compared
among sampling period and treatments by repeated measures ANOVA.
Physical and chemical water quality parameters were compared in each
individual sites using two-way ANOVA to determine the significance of
treatment and period effect.
Vegetation coverage data were square-root transformed. Non-metric
multidimensional scaling ordination (nMDS) was used to ordinate the Bray-
Curtis similarities computed from transformed vegetation data according to
treatment and period (data taken at the start versus that at the end of experiment).
One-way Analysis of Similarities (ANOSIM) was used to determine the
significance of treatment effect in each site. To determine whether particular
species was susceptible to treatment effects, individual one-way ANOVA test
were done on abundance in the first four dominant aquatic plant species of
aquatic plant observed before and after the treatment.

61
4.4 Results
Fish growth and survival
All fish were captured using landing nets, after full water extraction of all three
sites by September 2013 (Table 4.1). Mean monthly growth rate for length and
weight was 3.27 cm (S.D.: 0.20 cm) and 279.24g (S.D. = 20.26 g). While
average total length increment of more than one fold was recorded, fish specimen
weights increased in average for 3350.85g (S.D.: 243.15g) within one year.
Average survival rate was 66.67% (8 out of 12 individuals survived throughout
treatment). All four specimens were retrieved in site 2, while only two specimens
were retrieved from site 1 and 3 respectively. Death of two black carp was
detected in site 1 during September 2012. Even after replenishment of two new
juveniles in September 2012, we failed to retrieve them by the end of the
experimental period. For site 3, one specimen died in October 2012 and another
two specimens in April 2013 was found to be dead. Release of another two
specimens was done by mid-April 2013. Yet, none of newly released specimens
survived by September 2013.
Eggs clutches in the field
Eggs were laid most abundantly in June 2012 and then declined continuously to
minimum in March 2013, thereafter egg abundance rose again till June 2013
(Table 4.3). In treatment site 1 and 2, amount of egg clutches was reduced in
higher proportions than control sites during each sampling period until June 2013.
By the end of the experiment, a reduction of 55.8% of egg clutch abundance was
recorded in treatment site 1 by comparing the data of June 2012 and 2013. In
control site 1, amount of egg clutches in control site increased by three folds

62
within the same period. Same degree of reduction was not observed in treatment
site 2 by June 2013. Egg clutches abundance sharply rose for 2 folds during
March to June 2013 and reached similar amount to that in control site. Yet in
March 2013, percentage of egg clutches abundance in treatment site still
decreased by 15.38% since the trough reached during December 2012, compared
to an increase of more than one fold in control within the same period. No
comparable differences due to treatment were observed in data of site 3 (Fig. 4.2).
Hatchability of egg clutches in laboratory
The number of successful hatched eggs per clutch ranged from 90 to 355 (n =
25 – 30 egg clutches per sampling period). Mean hatchability ranged from 40%
in December 2012 to 83% in March 2013 (Table 4.4).
Apple snail and hatchling densities
Mean snail densities ranged from 1.33 – 92.0 individuals per m2 throughout the
experimental period (Fig. 4.1). There was a major decline of 96.6% (site 1),
94.1% (site 2) and 77.0% (site 3) in overall apple snail density in sites with black
carp introduced within the first three months (Jun 2012 – Sept 2012), while it
remained fairly stable till the breeding season in June 2013. Snail densities in all
three sites were significantly different both between treatments (F1,16 = 5.590, p <
0.05*) and among sampling time (F3,16 = 3.456, p < 0.05*).
Total estimated abundance of hatchlings ranged from 612.6 to 23973.0
individuals per month, and exhibited a clear seasonal pattern, with high values in
wet season (June – September 2012; April – June 2013) and low values in the dry
season (October – March 2013) (Fig. 4.3). A gradual decline in hatchlings was
recorded in both the control and treatment sites from June to December 2012, but

63
the number of hatchlings rose since March 2013. A comparison using three sites
as replicates showed significant differences between treatments (F1,16 = 4.450, p
< 0.05*) but not among sampling times (F3,16 = 2.011, p = 0.132) nor interaction
between the two factors (F3,16 = 0.0475, p = 0.766) (Table 4.2).
From the data of snail densities and hatchling abundance, we could
summarize that there was a clear seasonal differences in density and reproduction.
During June 2012, apple snail density reached a peak due to the high recruitment
from egg clutches. In September to December 2012, snail densities declined
which reduced the amount of egg clutches. Snail densities rose again in March
2013, thus causing the egg clutches abundance to grow again until it peaked in
June 2013.
For the juvenile apple snails (SL: <5 – 25mm), there were significant
differences in snail density recorded between treatments (F1,7 = 4.633, p < 0.05*)
but not among sampling times (F3,7 = 1.765, p = 0.194). For the adults,
significant differences in densities were detected only among sampling times
(F1,7 = 4.759, p = 0.01*) but not among treatments (F1,7 = 1.681, p = 0.213).
These results suggested size-dependent predation of juvenile snails by black carp
in experimental plot, while adults population was not affected (Fig. 4.4).
Fish gut content analysis
Among all eight specimens retrieved, mean gut fullness was 36.25% (S.D. =
30.67%). No distinguishable features in bolus could be identified to species level
even under dissection microscope.
Effects to non-target mollusks and other invertebrates

64
Mean densities of S. quadrata, M. tuberculata and Bithynia sp. ranged from 0.7 -
250, 0.0 - 126 and 0 - 110 individuals per m2, respectively (Fig. 4.5 & Table 4.6).
While no statistical differences in all parameters were detected in Bithynia sp.
densities, significant differences were detected in S. quadrata densities between
treatments (F1,7 = 6.229, p < 0.05*) and among sampling times (F3,7 = 4.477, p <
0.05). Densities of M. tuberculata differed between treatments (F1,7 = 9.236, p =
0.01*) and among sampling times (F3,7 = 5.301, p = 0.01*). In experimental site
1, average densities of S. quadrata and M. tuberculata reduced by 98.8% and
100% by June 2013 when compared to data in one year before, while that in
control site 1 decreased by 42.13% and increased by 33.33% respectively.
Similar magnitude of reduction was detected in experimental site 2. Average
densities for S. quadrata and M. tuberculata declined by 94.34% and 87.88%
respectively, while only a minor reduction of 0.01% in densities of S. quadrata
and 41.27% for that of M. tuberculata were recorded in control site 2.
No significant reductions due to treatment were found in abundance of
other non-target invertebrates. This was probably because of a high seasonal
variation in amount of benthic invertebrates, especially insects in ponds (Table
4.6).
Fish influence on water quality
There were significant seasonal differences among all physical water quality
parameters in all three sites (Fig. 4.6 & Table 4.9). Three parameters showed
additional significant difference between treatments (Table 4.7). These include
dissolved oxygen (site 2 and 3), pH (site 1, 2 and 3) and turbidity (site 2 and 3). It
was noted that turbidity levels shown significant differences between treatments

65
in both site 2 and 3. In site 2, treatment pond was 4.74% more turbid than control
pond in June 2012. By March and June 2013, treatment pond was 19.78% and
31.58% clearer than control pond respectively. Yet, in site 3, it was found that
treatment pond is 364.28% and 84.31% more turbid than control during
September and December 2012. Turbidity levels in treatment pond turned
53.60% and 17.85% clearer than control pond in two subsequent sampling
periods (March and June 2013).
In all three sites, nutrient level fluctuated throughout the experimental
period (Fig. 4.7). In general, total phosphorus (TP) and total nitrogen (TN) levels
reached maximum in June 2012, then dropped to minimum in December, while
they rose again thereafter. Among three sites, site 3 showed the highest level in
total phosphorus (≥ 0.3 mgL-1
), no matter in experiment or control treatments.
Ranges for four nutrient parameters were 0.0 – 1.45 mgL-1
(NH3); 0.8 – 2.25
mgL-1
(TN); 0.0 – 0.24 mgL-1
(PO4-) and; 0.01 – 0.45 mgL
-1 (TP) (Table 4.9).
While seasonal effect was highly significant in all parameters among sites, only
data of P and TP in site 2 and 3 showed significant differences among treatments
(Table 4.8). In site 2, significantly lower phosphate concentrations (F1,4 = 5.000,
p < 0.05*) and total phosphorus concentration (F1,4 = 8.333, p = 0.01*) were
recorded in treatment pond, while interactions with period effect was both
significant. In site 3, significant differences were also found in two phosphorus
parameters. Significantly lower concentrations of phosphates (F1,4 = 96.729, p <
0.01) and total phosphorus (F1,4 = 34.416, p < 0.01) was recorded by June 2013,
while positive interactions with period effect were recorded as well (Table 4.8).
Vegetation abundance

66
Vegetation diversities were quite consistent across three sites. From mean
abundance of each plant species among three sites, dominant vegetation before
the experiment was mostly emergent plants comprised of Paspalum conjugatum
(30.71%), Paspalum distichum (12.50%), Ipomoea aquatica (12.29%), Panicum
dichotomiflorum (11.73%). P. dichotomiflorum (25.19%), Paspalum distichum
(17.18%), Ipomoea aquatica (13.14%, especially at site 3), and Paspalum
conjugatum (12.43%) became the most dominant plant species after the treatment
(Table 4.10). Ordination of plant abundance communities before and after the
treatment yet gave poor representation in three nMDS plots (stress = 0.1) (Fig.
4.8). No clear change in overall community structure was found after fish
implementation in all three experimental plots. Results from one-way ANOSIM
indicated that there was unclear segregation of results due to treatment (site 1:
Global R = 0.088, p = 3.8%; site 2: Global R = 0.119, p = 2.1%; site 3: Global R
= 0.126, p = 1.5%).
Though no significant changes in vegetation community structure were
recorded, particular plant species grew vigorously in treatment with fish. Yet,
these changes were restricted in one site, while not observed in all three
replicates. By comparing plant abundances from same eight quadrats in June
2012 and June 2013, average abundances of P. dichotomiflorum increased
dramatically by 100% in experimental site 1, while that in control reduced by
63%; in experimental site 3, mean abundance of I. aquatica increased by 400%
in treatment with black carp and decreased by 99.1% in control treatment.
Significant statistical differences in abundance of P, dichotomiflorum (F1,14 =
23.33, p < 0.01*) in site 1 and I. aquatica (F1,14 = 4.876, p < 0.05*) in site 3 were

67
shown individually after treatment with black carp for one year, while no
significant differences were observed by comparing them on June 2012 (P.
dichotomiflorum: F1,14 = 0.066, p = 0.801; I. aquatica: F1,14 = 7.177, p = 0.059).
Comparison on maximum height of these two emergent aquatic plant species
showed same pattern of results. Mean maximum height of P. dichotomiflorum in
quadrats of experimental site 1 reached 1.3 m (S.D. = 0.063, n = 8), while that in
control has only reached 0.91 m (S.D. = 0.13, n = 2). In site 3, maximum height
of I. aquatica has reached 0.82 m on average (S.D. = 0.21, n = 5) in experimental
site and 0.50 m (S.D. = 0.11, n = 4) in control site.
4.5 Discussion
In natural shallow water bodies, presence of predatory fish, especially
molluscivorous specialists, impose top-down control on mollusk populations in
aquatic ecosystems (Brönmark, 1988; Brönmark and Hansson, 2005; Martin et
al., 1992). In Hong Kong, due to the absence of large water bodies and artificial
channelization of natural lowland streams, there are insufficient predators to
control apple snail populations (refer to Chapter 1, table 1.1). In addition, escape
of edible exotic fish fingerlings from fish ponds and purposeful release facilitate
invasion of exotic predatory and omnivorous fish, like omnivorous Nile tilapia
(Oreochromis niloticus Linnaeus) and piscivorous snakehead murrels (Channa
striata Bloch), in disturbed rivers and channels (Dudgeon, 1996; Hodgkiss &
Man, 1977; Lee et al., 2004). However, among all existing predatory fish in local
freshwater habitats, none are specialized molluscivores (Lee et al., 2004). Re-
introduction of an indigenous mollusk specialist might thus be a viable solution
in controlling apple snails, especially in lowland wetlands with impoverished
biodiversity.

68
Black carp (Mylopharyngodon piceus) is a highly tolerant species that
migrates annually from riverine habitats to lakes and floodplains to complete
reproductive cycle. This species has longevity of 13 – 15 years, making it a good
candidate for biological control (Nico et al., 2005). Due to synchronization with
annual flooding events, it has broad environmental tolerances. For example, it
can survive in a wide range of water temperature from 4 to 30oC (Chu and Chen,
1989; Gangstad, 1980; Li and Fang, 1990). In our study, even under the poor
conditions of site 3, 2 of the 4 individuals released survived throughout the whole
study period. The water depth at this site was as low as 0.4 m during the dry
season, while temperature reached 31.1oC and dissolved oxygen level was
0.41mgL-1
in the wet season. At the start of the experiment, dissolved oxygen
was even as low as 0.16 mgL-1
but no fish specimens were observed to die within
the first week after release even being reared in stagnant water.
Our study is the first pond-scale experiment that has examined the
effectiveness of black carp as a biological control agent of apple snails. The one-
year experimental period was essential to determine how seasonal changes in
apple snail life-history characteristics could modulate the predatory effect of
black carp. Within the first three months of experiment (June – September 2012),
apple snail density had already diminished by an average of 89.25%, while that
ranged from 77.0% (site 3) to 96.6% (site 1). The maximum reduction in snail
density of 98.3% was achieved in December 2012 in site 1, where no more
neonates were recorded until the final survey conducted in June 2013. It was also
in site 1 where the abundance of juvenile snails (< 25 mm) declined most quickly
and remained low until the end of experiment. In March 2013, this size range of

69
snails was also depleted by black carp in site 3 after low recruitment rate in dry
season, though the level rose again by June 2013. This re-elevation in snail
densities might be related to the availability of favorable habitat structure for
apple snails. During wet season in June 2013, the site was flooded by rainwater
and generated additional shallow habitats (<0.2m). These habitats were known to
increase apple snails’ fitness (Karunaratne et al., 2006), but low water depth
increased difficulties for black carp in reaching their prey. This could be proved
from body depths of all retrieved black carp, in which it ranged from 0.13 -
0.14m. In site 3, large number of adult snails, which were too big to fit into the
mouth of the fish released into the pond, sustained a huge population of over
20000 individuals of hatchlings in June 2012 and 2013. Though carp could
significantly reduce the abundance of juvenile snails, eradication of the whole
population could only be achieved after the adult snails died out after completing
their lifespan. Nevertheless, since the adults suffered great over-winter mortality,
even measures that can control only the abundance of juveniles should be
effective for the whole population control by limiting new recruits from reaching
adulthood. Gut content analysis of black carp specimens were insufficient in
reflecting the diet of black carp. Even in the foregut where food items were least
digested, no distinguishable features among all prey present within the habitat
could be found. This is probably because pharyngeal mastication (Sibbling, 1982)
and repeated re-suspension of food materials with mucus in pharynx (Sibbling,
1991) has highly increased the feeding efficiency in all cyprinid fishes.
By controlling the population of herbivorous apple snails, black carp might
have indirect effects to macrophytes and nutrients. Grazing by apple snails could

70
release nitrogen and phosphorus originally stored in macrophytes (Carlsson et al.,
2004a), although the sediment might re-absorb some of the nutrients. Previous
studies conducted in wetlands with greater macrophyte diversity showed that the
existence of juvenile apple snails with high abundance might become more
competitive than adult ones by having higher feeding efficiency (Carlsson and
Brönmark, 2006; Boland et al. 2008). It is therefore possible that significant
reduction of juvenile apple snails could eventually minimize nutrient flux after
the adults died, thus preventing shift of primary production from macrophytes to
phytoplanktons. Results from our present study showed significantly different
phosphates and total phosphorus concentrations in two out of three treatment
sites than control sites since treatment started. However, comparing between
results of June 2012 and June 2013, we could only observe a significant
reduction in phosphorus concentrations in site 3, but not among the rest two
treatment sites. Insignificant results in only one but not all treatment sites might
be due to continued addition of contaminated water source from an adjacent
channel. As there was only one reliable water source, constant water supply was
provided to only site 1 and 2 to maintain sufficient water levels during November
2012 – February 2013 from this channel. As a high population of tilapia and
apple snails was sustained inside this channel, water inside this channel might
had a steadily high concentrations of nitrogen and phosphorus. It would therefore
highly possible that fluctuation in dissolved nutrient concentrations in our
treatment sites was caused by this process. Our results from present study were
therefore inconclusive in explaining reduction in population of a voracious
consumer in aquatic plants could reduce excessive nutrient influx to water bodies.

71
Although black carp is known to be mollusk-eating, most studies conducted
using this fish species as subject however provided insufficient data in evaluating
the fish performance as a bio-control agent. In one of publication at Fuxian Lake
at Yunnan, China, bivalves namely Margarya spp. and Corbicula fluminea have
been cited as food items of black carp (Yang and Chen, 1995), though no
experiment was cited as reference. A number of experimental studies have shown
that black carp is an effective predator of molluscs that are hosts of human
parasites, such as Planorbella spp. (Ledford and Kelly, 2006), Physa acuta and
Melanoides tuberculata (Ben-Ami and Heller, 2001) Ram-horn snails Heliosoma
sp.and Physa sp. (Collins, 1996; Thomforde, 2000). Though some of these
experiments were conducted in field, there were no data evaluating densities of
co-existing benthic organisms. The only field based experiment that considered
subtle effects in controlling exotic snails on existing indigenous mollusks was
conducted in Israeli reservoirs by Leventer (1979). From the results, it has been
reported that visibly all target and non-target specimens of snails including
Lymnea auricularia, Bulinus truncates, M. tuberculata, Melanopsis praemorsa
and M. costata plus one bivalve Corbicula fluminea were eliminated. Yet, only
their abundance data estimated after the experiment were used to justify the
success of deploying fish as effective bio-control agent, while seasonal predatory
effect was not considered. Results from our experiment indicated that black carp
would prey upon golden apple snails and at least three species of non-target
snails including Sinotaia quadrata, M. tuberculata and Bithynia sp., though
significant reduction was not recorded in Bithynia sp. due to variation in densities
among plots. Other benthic invertebrates including various species of dragonflies
and damselflies also existed within the sites. But no significant changes could be

72
concluded from densities of any co-exiting soft-bodied invertebrates other than
mollusks between two treatments.
Only some species of aquatic plants but not all showed increased growth
performance by reduction in apple snail densities. In this study, though black
carp succeeded in controlling abundance of juvenile apple snails, only two
emergent or floating macrophytes P. dichotomiflorum and I. aquatica were
shown to have benefited. Reduced abundance of apple snails as voracious grazers
had promoted their heights and densities in sites with fish treatment. Wong et al.
(2010) has suggested I. aquatica to be preferably consumed by apple snails
according to their relatively high nutrient content and low dry matter content.
Recent mesocosm experiment (Ip et al. submitted) had also shown increased
abundance and biomass of I. aquatica when black carp were in coexistence with
apple snails. For P. dichotomiflorum, its habitat is restricted to shallow water and
marshy places. It is a hydrophious herbaceous plant with emergent and floating
culms (Lin, 2009; Xia et al., 2011). No current data was available considering its
nutrient data and physical constituents. Yet their differences in growth
performance might be related to their natural growth strategies. Since our present
study did not devise sufficient replicates to compare growth performances among
plant species and between treatments, we could not reach conclusive summary on
reduction of apple snail abundance could lead to more favorable growth
performance of co-existing aquatic plants.
In conclusion, black carp is a highly tolerant species that can survive in
shallow stagnant waters. This species could be used as a biological control agent
of apple snails due to its high effectiveness as a predator in shallow freshwater

73
wetlands, which are common habitat in southern China. Given the longevity of
black carp, a low stocking density (80 - 89 individuals ha-1
; stocking density for
site 2 and 3 in this experiment) will be sufficient to control apple snail
populations. Additionally, black carp culture in wetlands can provide commercial
incentives as it is an edible fish. Yet implementation of black carp to control
apple snails should only be carried out in habitats with low biodiversity of
macro-invertebrates, especially mollusks. Juvenile (total length: at least
approximately 300mm) specimens are most preferable option for release as they
could reach a broader range of habitats including shallow riparian area where
snails aggregate, while still beholding their high adaptability to adverse
environmental conditions in shallow wetlands.
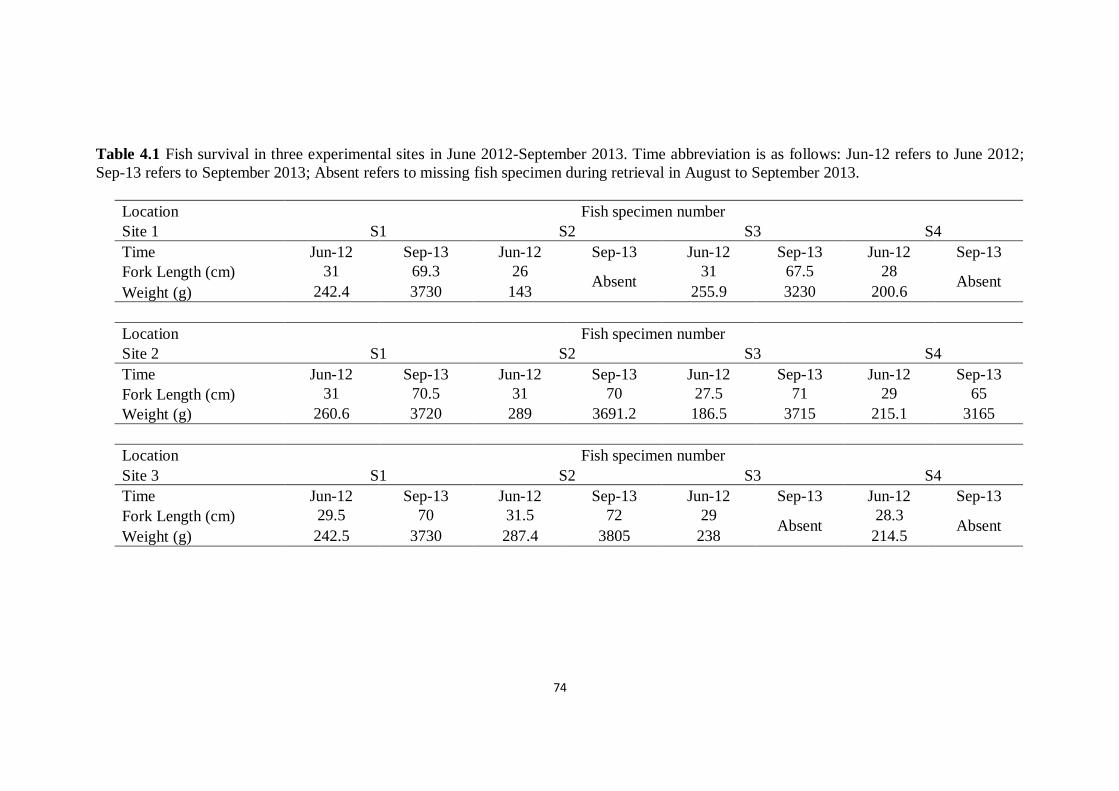
74
Table 4.1 Fish survival in three experimental sites in June 2012-September 2013. Time abbreviation is as follows: Jun-12 refers to June 2012;
Sep-13 refers to September 2013; Absent refers to missing fish specimen during retrieval in August to September 2013.
Location Fish specimen number
Site 1 S1 S2 S3 S4
Time Jun-12 Sep-13 Jun-12 Sep-13 Jun-12 Sep-13 Jun-12 Sep-13
Fork Length (cm) 31 69.3 26 Absent
31 67.5 28 Absent
Weight (g) 242.4 3730 143 255.9 3230 200.6
Location Fish specimen number
Site 2 S1 S2 S3 S4
Time Jun-12 Sep-13 Jun-12 Sep-13 Jun-12 Sep-13 Jun-12 Sep-13
Fork Length (cm) 31 70.5 31 70 27.5 71 29 65
Weight (g) 260.6 3720 289 3691.2 186.5 3715 215.1 3165
Location Fish specimen number
Site 3 S1 S2 S3 S4
Time Jun-12 Sep-13 Jun-12 Sep-13 Jun-12 Sep-13 Jun-12 Sep-13
Fork Length (cm) 29.5 70 31.5 72 29 Absent
28.3 Absent
Weight (g) 242.5 3730 287.4 3805 238 214.5

75
Table 4.2 Results of 2-way ANOVA testing the difference in estimated
hatchlings between treatments and among sampling time.
Factor df F P
Treatment 1 4.450 0.048*
Sampling time 4 2.011 0.132
Treatment * Sampling time 4 0.457 0.766
Error 20
Data with statistical differences (P < 0.05) are shown with asterisk.
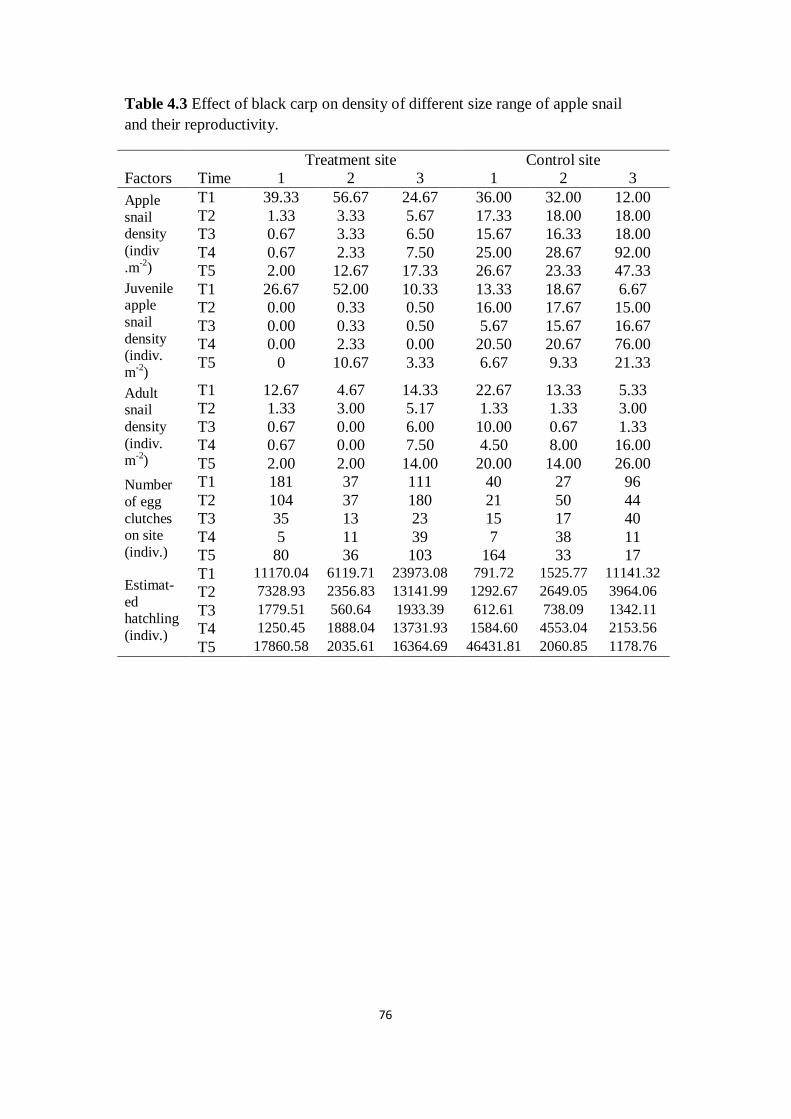
76
Table 4.3 Effect of black carp on density of different size range of apple snail
and their reproductivity.
Treatment site Control site
Factors Time 1 2 3 1 2 3
Apple
snail density
(indiv
.m-2
)
T1 39.33 56.67 24.67 36.00 32.00 12.00
T2 1.33 3.33 5.67 17.33 18.00 18.00
T3 0.67 3.33 6.50 15.67 16.33 18.00
T4 0.67 2.33 7.50 25.00 28.67 92.00
T5 2.00 12.67 17.33 26.67 23.33 47.33 Juvenile apple
snail
density (indiv.
m-2
)
T1 26.67 52.00 10.33 13.33 18.67 6.67
T2 0.00 0.33 0.50 16.00 17.67 15.00
T3 0.00 0.33 0.50 5.67 15.67 16.67
T4 0.00 2.33 0.00 20.50 20.67 76.00
T5 0 10.67 3.33 6.67 9.33 21.33
Adult snail
density
(indiv.
m-2
)
T1 12.67 4.67 14.33 22.67 13.33 5.33
T2 1.33 3.00 5.17 1.33 1.33 3.00
T3 0.67 0.00 6.00 10.00 0.67 1.33
T4 0.67 0.00 7.50 4.50 8.00 16.00
T5 2.00 2.00 14.00 20.00 14.00 26.00
Number
of egg
clutches on site
(indiv.)
T1 181 37 111 40 27 96
T2 104 37 180 21 50 44
T3 35 13 23 15 17 40
T4 5 11 39 7 38 11
T5 80 36 103 164 33 17
Estimat-
ed hatchling
(indiv.)
T1 11170.04 6119.71 23973.08 791.72 1525.77 11141.32
T2 7328.93 2356.83 13141.99 1292.67 2649.05 3964.06
T3 1779.51 560.64 1933.39 612.61 738.09 1342.11
T4 1250.45 1888.04 13731.93 1584.60 4553.04 2153.56
T5 17860.58 2035.61 16364.69 46431.81 2060.85 1178.76
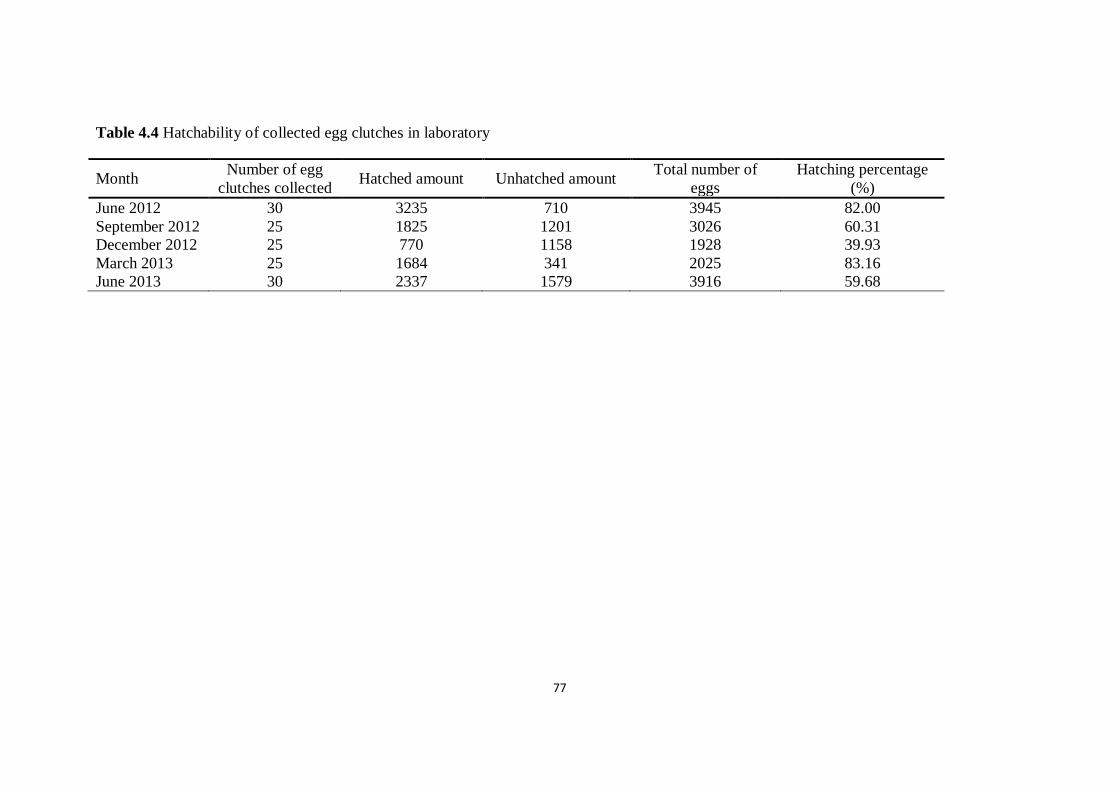
77
Table 4.4 Hatchability of collected egg clutches in laboratory
Month Number of egg
clutches collected Hatched amount Unhatched amount
Total number of
eggs
Hatching percentage
(%)
June 2012 30 3235 710 3945 82.00
September 2012 25 1825 1201 3026 60.31
December 2012 25 770 1158 1928 39.93
March 2013 25 1684 341 2025 83.16
June 2013 30 2337 1579 3916 59.68
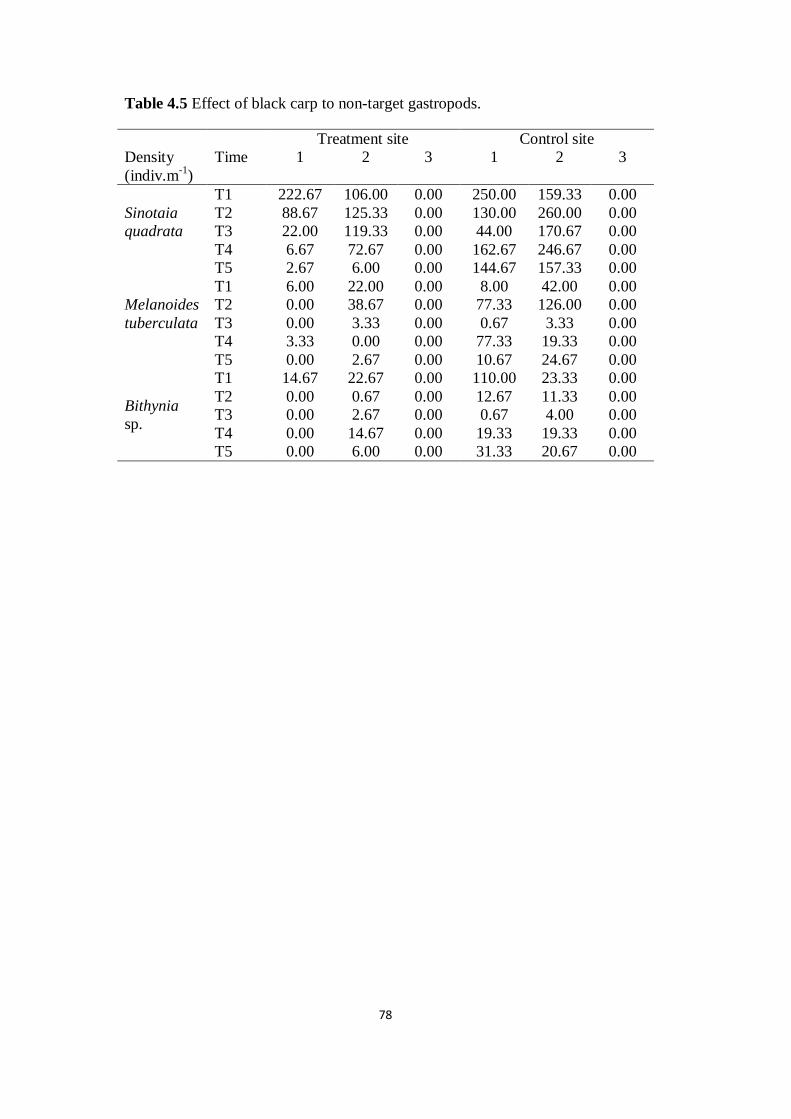
78
Table 4.5 Effect of black carp to non-target gastropods.
Treatment site Control site
Density
(indiv.m-1
)
Time 1 2 3 1 2 3
Sinotaia
quadrata
T1 222.67 106.00 0.00 250.00 159.33 0.00
T2 88.67 125.33 0.00 130.00 260.00 0.00
T3 22.00 119.33 0.00 44.00 170.67 0.00
T4 6.67 72.67 0.00 162.67 246.67 0.00
T5 2.67 6.00 0.00 144.67 157.33 0.00
Melanoides
tuberculata
T1 6.00 22.00 0.00 8.00 42.00 0.00
T2 0.00 38.67 0.00 77.33 126.00 0.00
T3 0.00 3.33 0.00 0.67 3.33 0.00
T4 3.33 0.00 0.00 77.33 19.33 0.00
T5 0.00 2.67 0.00 10.67 24.67 0.00
Bithynia
sp.
T1 14.67 22.67 0.00 110.00 23.33 0.00
T2 0.00 0.67 0.00 12.67 11.33 0.00
T3 0.00 2.67 0.00 0.67 4.00 0.00
T4 0.00 14.67 0.00 19.33 19.33 0.00
T5 0.00 6.00 0.00 31.33 20.67 0.00
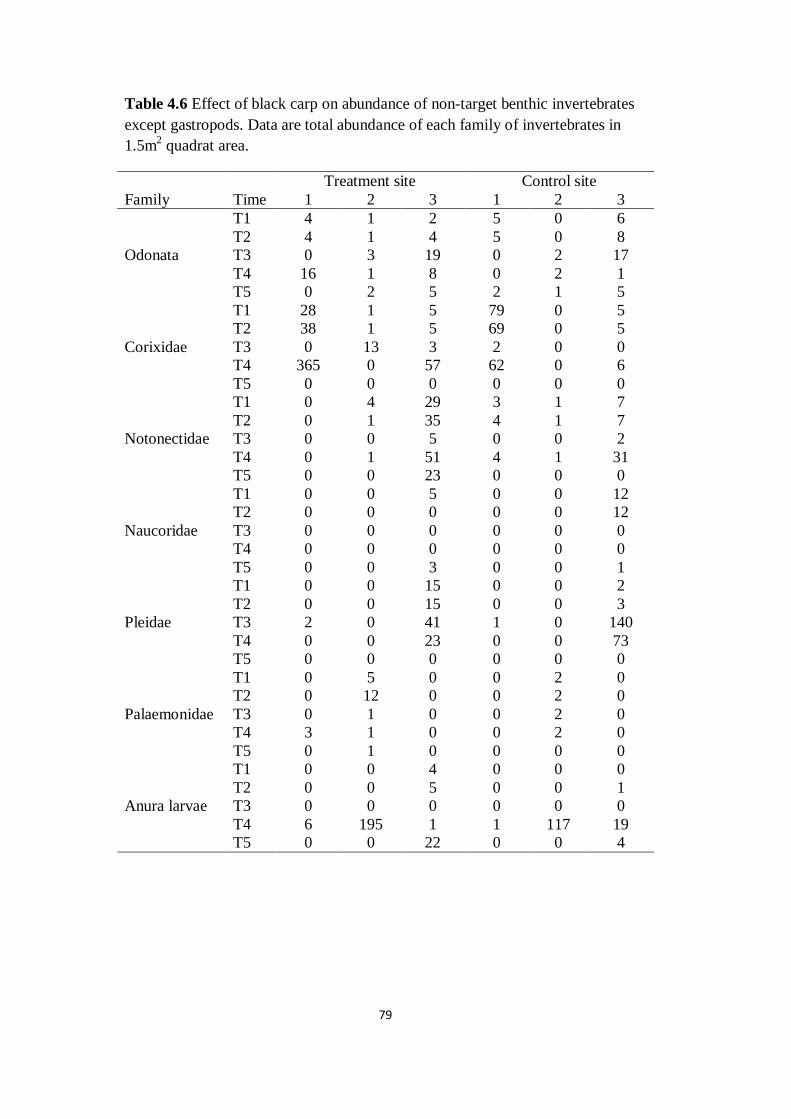
79
Table 4.6 Effect of black carp on abundance of non-target benthic invertebrates
except gastropods. Data are total abundance of each family of invertebrates in
1.5m2 quadrat area.
Treatment site Control site
Family Time 1 2 3 1 2 3
Odonata
T1 4 1 2 5 0 6
T2 4 1 4 5 0 8
T3 0 3 19 0 2 17
T4 16 1 8 0 2 1
T5 0 2 5 2 1 5
Corixidae
T1 28 1 5 79 0 5
T2 38 1 5 69 0 5
T3 0 13 3 2 0 0
T4 365 0 57 62 0 6
T5 0 0 0 0 0 0
Notonectidae
T1 0 4 29 3 1 7
T2 0 1 35 4 1 7
T3 0 0 5 0 0 2
T4 0 1 51 4 1 31
T5 0 0 23 0 0 0
Naucoridae
T1 0 0 5 0 0 12
T2 0 0 0 0 0 12
T3 0 0 0 0 0 0
T4 0 0 0 0 0 0
T5 0 0 3 0 0 1
Pleidae
T1 0 0 15 0 0 2
T2 0 0 15 0 0 3
T3 2 0 41 1 0 140
T4 0 0 23 0 0 73
T5 0 0 0 0 0 0
Palaemonidae
T1 0 5 0 0 2 0
T2 0 12 0 0 2 0
T3 0 1 0 0 2 0
T4 3 1 0 0 2 0
T5 0 1 0 0 0 0
Anura larvae
T1 0 0 4 0 0 0
T2 0 0 5 0 0 1
T3 0 0 0 0 0 0
T4 6 195 1 1 117 19
T5 0 0 22 0 0 4
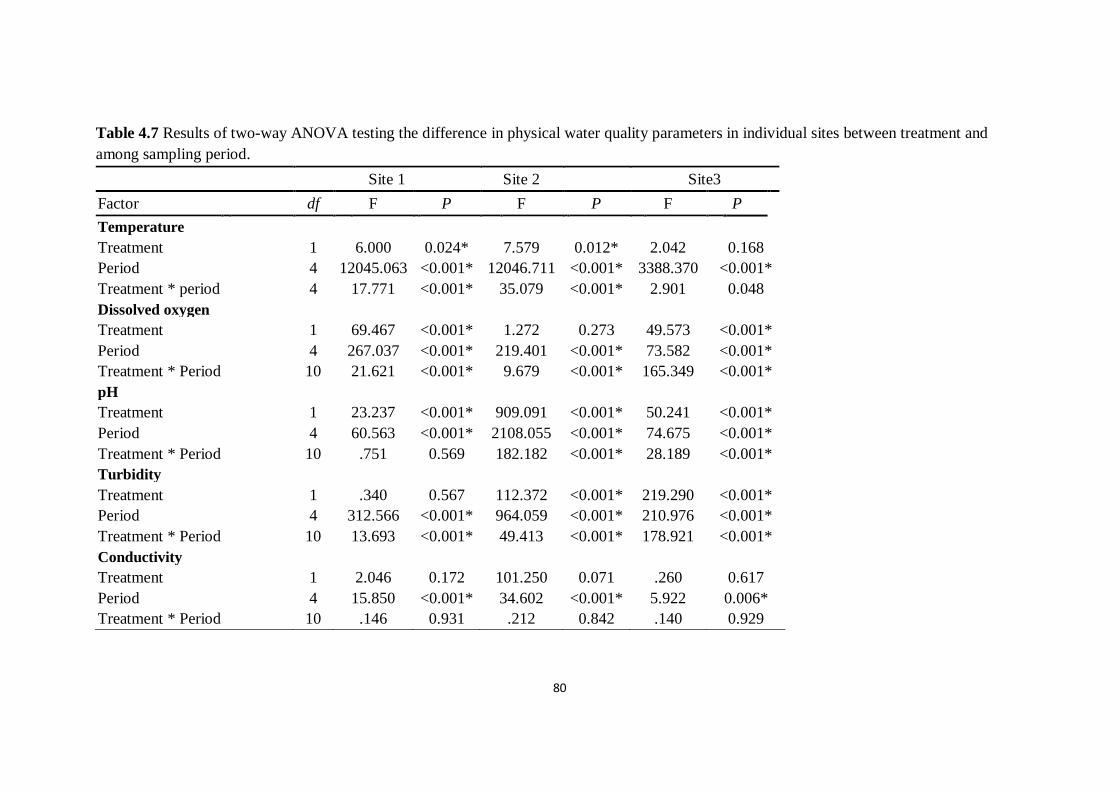
80
Table 4.7 Results of two-way ANOVA testing the difference in physical water quality parameters in individual sites between treatment and
among sampling period.
Site 1 Site 2 Site3
Factor df F P F P F P
Temperature
Treatment 1 6.000 0.024* 7.579 0.012* 2.042 0.168
Period 4 12045.063 <0.001* 12046.711 <0.001* 3388.370 <0.001*
Treatment * period 4 17.771 <0.001* 35.079 <0.001* 2.901 0.048
Dissolved oxygen
Treatment 1 69.467 <0.001* 1.272 0.273 49.573 <0.001*
Period 4 267.037 <0.001* 219.401 <0.001* 73.582 <0.001*
Treatment * Period 10 21.621 <0.001* 9.679 <0.001* 165.349 <0.001*
pH
Treatment 1 23.237 <0.001* 909.091 <0.001* 50.241 <0.001*
Period 4 60.563 <0.001* 2108.055 <0.001* 74.675 <0.001*
Treatment * Period 10 .751 0.569 182.182 <0.001* 28.189 <0.001*
Turbidity
Treatment 1 .340 0.567 112.372 <0.001* 219.290 <0.001*
Period 4 312.566 <0.001* 964.059 <0.001* 210.976 <0.001*
Treatment * Period 10 13.693 <0.001* 49.413 <0.001* 178.921 <0.001*
Conductivity
Treatment 1 2.046 0.172 101.250 0.071 .260 0.617
Period 4 15.850 <0.001* 34.602 <0.001* 5.922 0.006*
Treatment * Period 10 .146 0.931 .212 0.842 .140 0.929
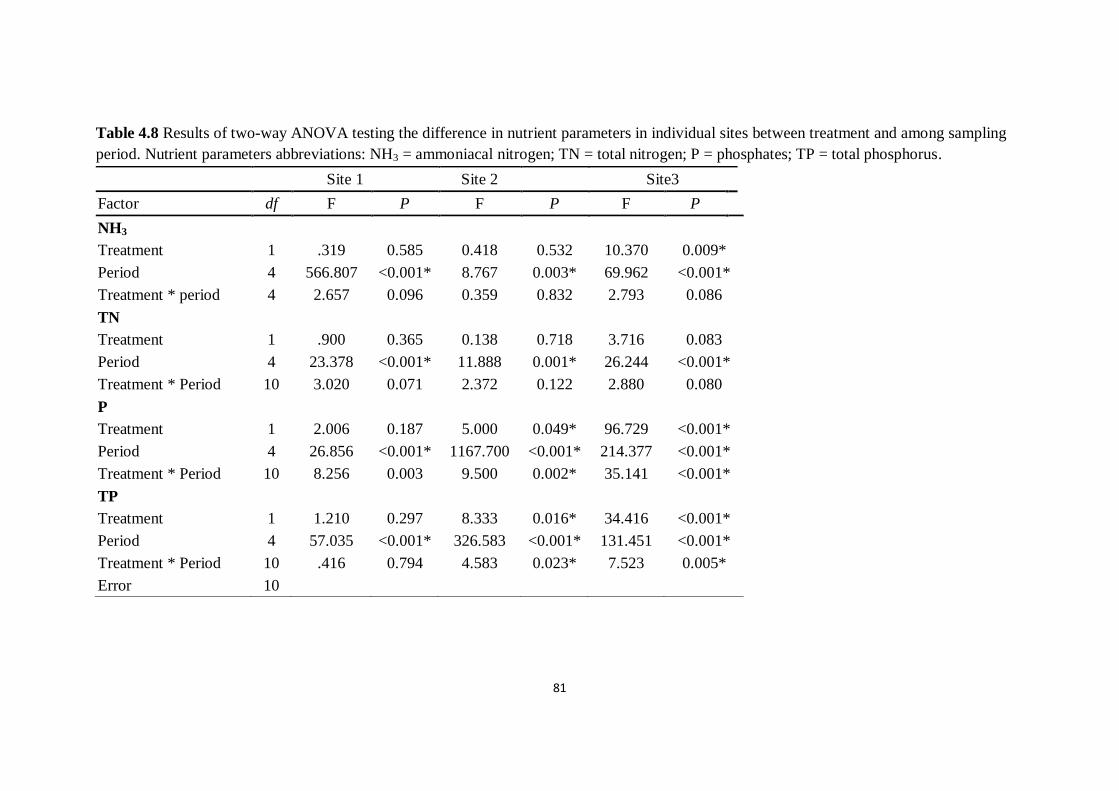
81
Table 4.8 Results of two-way ANOVA testing the difference in nutrient parameters in individual sites between treatment and among sampling
period. Nutrient parameters abbreviations: NH3 = ammoniacal nitrogen; TN = total nitrogen; P = phosphates; TP = total phosphorus.
Site 1 Site 2 Site3
Factor df F P F P F P
NH3
Treatment 1 .319 0.585 0.418 0.532 10.370 0.009*
Period 4 566.807 <0.001* 8.767 0.003* 69.962 <0.001*
Treatment * period 4 2.657 0.096 0.359 0.832 2.793 0.086
TN
Treatment 1 .900 0.365 0.138 0.718 3.716 0.083
Period 4 23.378 <0.001* 11.888 0.001* 26.244 <0.001*
Treatment * Period 10 3.020 0.071 2.372 0.122 2.880 0.080
P
Treatment 1 2.006 0.187 5.000 0.049* 96.729 <0.001*
Period 4 26.856 <0.001* 1167.700 <0.001* 214.377 <0.001*
Treatment * Period 10 8.256 0.003 9.500 0.002* 35.141 <0.001*
TP
Treatment 1 1.210 0.297 8.333 0.016* 34.416 <0.001*
Period 4 57.035 <0.001* 326.583 <0.001* 131.451 <0.001*
Treatment * Period 10 .416 0.794 4.583 0.023* 7.523 0.005*
Error 10
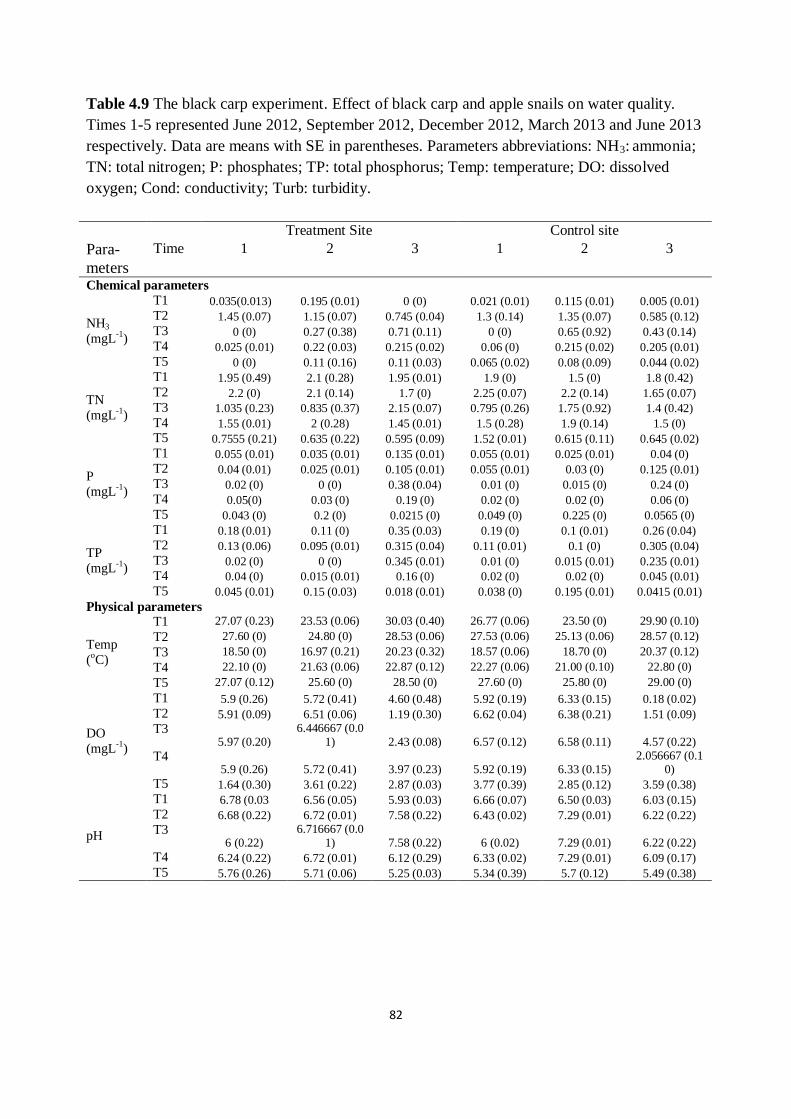
82
Table 4.9 The black carp experiment. Effect of black carp and apple snails on water quality.
Times 1-5 represented June 2012, September 2012, December 2012, March 2013 and June 2013
respectively. Data are means with SE in parentheses. Parameters abbreviations: NH3: ammonia;
TN: total nitrogen; P: phosphates; TP: total phosphorus; Temp: temperature; DO: dissolved
oxygen; Cond: conductivity; Turb: turbidity.
Treatment Site Control site
Para-
meters
Time 1 2 3 1 2 3
Chemical parameters
NH3
(mgL-1)
T1 0.035(0.013) 0.195 (0.01) 0 (0) 0.021 (0.01) 0.115 (0.01) 0.005 (0.01)
T2 1.45 (0.07) 1.15 (0.07) 0.745 (0.04) 1.3 (0.14) 1.35 (0.07) 0.585 (0.12)
T3 0 (0) 0.27 (0.38) 0.71 (0.11) 0 (0) 0.65 (0.92) 0.43 (0.14)
T4 0.025 (0.01) 0.22 (0.03) 0.215 (0.02) 0.06 (0) 0.215 (0.02) 0.205 (0.01)
T5 0 (0) 0.11 (0.16) 0.11 (0.03) 0.065 (0.02) 0.08 (0.09) 0.044 (0.02)
TN
(mgL-1)
T1 1.95 (0.49) 2.1 (0.28) 1.95 (0.01) 1.9 (0) 1.5 (0) 1.8 (0.42)
T2 2.2 (0) 2.1 (0.14) 1.7 (0) 2.25 (0.07) 2.2 (0.14) 1.65 (0.07)
T3 1.035 (0.23) 0.835 (0.37) 2.15 (0.07) 0.795 (0.26) 1.75 (0.92) 1.4 (0.42)
T4 1.55 (0.01) 2 (0.28) 1.45 (0.01) 1.5 (0.28) 1.9 (0.14) 1.5 (0)
T5 0.7555 (0.21) 0.635 (0.22) 0.595 (0.09) 1.52 (0.01) 0.615 (0.11) 0.645 (0.02)
P
(mgL-1)
T1 0.055 (0.01) 0.035 (0.01) 0.135 (0.01) 0.055 (0.01) 0.025 (0.01) 0.04 (0)
T2 0.04 (0.01) 0.025 (0.01) 0.105 (0.01) 0.055 (0.01) 0.03 (0) 0.125 (0.01)
T3 0.02 (0) 0 (0) 0.38 (0.04) 0.01 (0) 0.015 (0) 0.24 (0)
T4 0.05(0) 0.03 (0) 0.19 (0) 0.02 (0) 0.02 (0) 0.06 (0)
T5 0.043 (0) 0.2 (0) 0.0215 (0) 0.049 (0) 0.225 (0) 0.0565 (0)
TP
(mgL-1)
T1 0.18 (0.01) 0.11 (0) 0.35 (0.03) 0.19 (0) 0.1 (0.01) 0.26 (0.04)
T2 0.13 (0.06) 0.095 (0.01) 0.315 (0.04) 0.11 (0.01) 0.1 (0) 0.305 (0.04)
T3 0.02 (0) 0 (0) 0.345 (0.01) 0.01 (0) 0.015 (0.01) 0.235 (0.01)
T4 0.04 (0) 0.015 (0.01) 0.16 (0) 0.02 (0) 0.02 (0) 0.045 (0.01)
T5 0.045 (0.01) 0.15 (0.03) 0.018 (0.01) 0.038 (0) 0.195 (0.01) 0.0415 (0.01)
Physical parameters
Temp
(oC)
T1 27.07 (0.23) 23.53 (0.06) 30.03 (0.40) 26.77 (0.06) 23.50 (0) 29.90 (0.10)
T2 27.60 (0) 24.80 (0) 28.53 (0.06) 27.53 (0.06) 25.13 (0.06) 28.57 (0.12)
T3 18.50 (0) 16.97 (0.21) 20.23 (0.32) 18.57 (0.06) 18.70 (0) 20.37 (0.12)
T4 22.10 (0) 21.63 (0.06) 22.87 (0.12) 22.27 (0.06) 21.00 (0.10) 22.80 (0)
T5 27.07 (0.12) 25.60 (0) 28.50 (0) 27.60 (0) 25.80 (0) 29.00 (0)
DO
(mgL-1)
T1 5.9 (0.26) 5.72 (0.41) 4.60 (0.48) 5.92 (0.19) 6.33 (0.15) 0.18 (0.02)
T2 5.91 (0.09) 6.51 (0.06) 1.19 (0.30) 6.62 (0.04) 6.38 (0.21) 1.51 (0.09)
T3 5.97 (0.20)
6.446667 (0.01) 2.43 (0.08) 6.57 (0.12) 6.58 (0.11) 4.57 (0.22)
T4 5.9 (0.26) 5.72 (0.41) 3.97 (0.23) 5.92 (0.19) 6.33 (0.15)
2.056667 (0.10)
T5 1.64 (0.30) 3.61 (0.22) 2.87 (0.03) 3.77 (0.39) 2.85 (0.12) 3.59 (0.38)
pH
T1 6.78 (0.03 6.56 (0.05) 5.93 (0.03) 6.66 (0.07) 6.50 (0.03) 6.03 (0.15)
T2 6.68 (0.22) 6.72 (0.01) 7.58 (0.22) 6.43 (0.02) 7.29 (0.01) 6.22 (0.22)
T3 6 (0.22)
6.716667 (0.01) 7.58 (0.22) 6 (0.02) 7.29 (0.01) 6.22 (0.22)
T4 6.24 (0.22) 6.72 (0.01) 6.12 (0.29) 6.33 (0.02) 7.29 (0.01) 6.09 (0.17)
T5 5.76 (0.26) 5.71 (0.06) 5.25 (0.03) 5.34 (0.39) 5.7 (0.12) 5.49 (0.38)
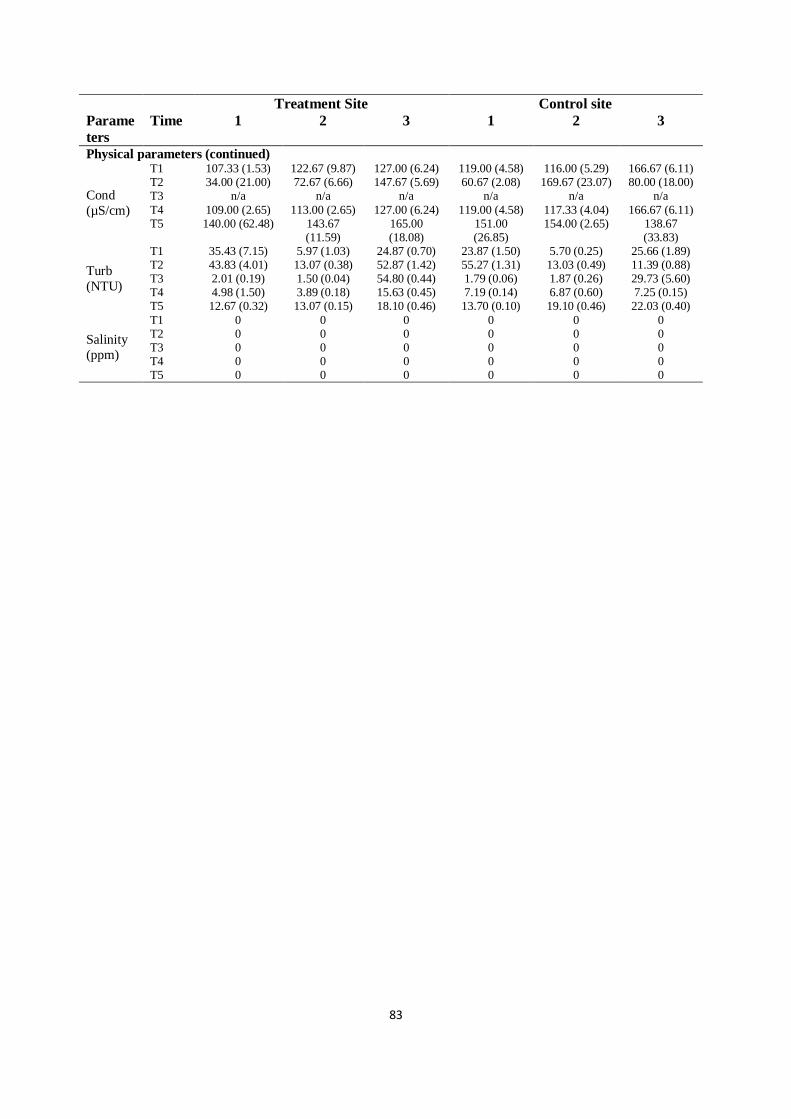
83
Treatment Site Control site
Parame
ters
Time 1 2 3 1 2 3
Physical parameters (continued)
Cond
(µS/cm)
T1 107.33 (1.53) 122.67 (9.87) 127.00 (6.24) 119.00 (4.58) 116.00 (5.29) 166.67 (6.11) T2 34.00 (21.00) 72.67 (6.66) 147.67 (5.69) 60.67 (2.08) 169.67 (23.07) 80.00 (18.00) T3 n/a n/a n/a n/a n/a n/a
T4 109.00 (2.65) 113.00 (2.65) 127.00 (6.24) 119.00 (4.58) 117.33 (4.04) 166.67 (6.11) T5 140.00 (62.48) 143.67
(11.59) 165.00 (18.08)
151.00 (26.85)
154.00 (2.65) 138.67 (33.83)
Turb
(NTU)
T1 35.43 (7.15) 5.97 (1.03) 24.87 (0.70) 23.87 (1.50) 5.70 (0.25) 25.66 (1.89) T2 43.83 (4.01) 13.07 (0.38) 52.87 (1.42) 55.27 (1.31) 13.03 (0.49) 11.39 (0.88) T3 2.01 (0.19) 1.50 (0.04) 54.80 (0.44) 1.79 (0.06) 1.87 (0.26) 29.73 (5.60) T4 4.98 (1.50) 3.89 (0.18) 15.63 (0.45) 7.19 (0.14) 6.87 (0.60) 7.25 (0.15) T5 12.67 (0.32) 13.07 (0.15) 18.10 (0.46) 13.70 (0.10) 19.10 (0.46) 22.03 (0.40)
Salinity
(ppm)
T1 0 0 0 0 0 0 T2 0 0 0 0 0 0 T3 0 0 0 0 0 0 T4 0 0 0 0 0 0 T5 0 0 0 0 0 0
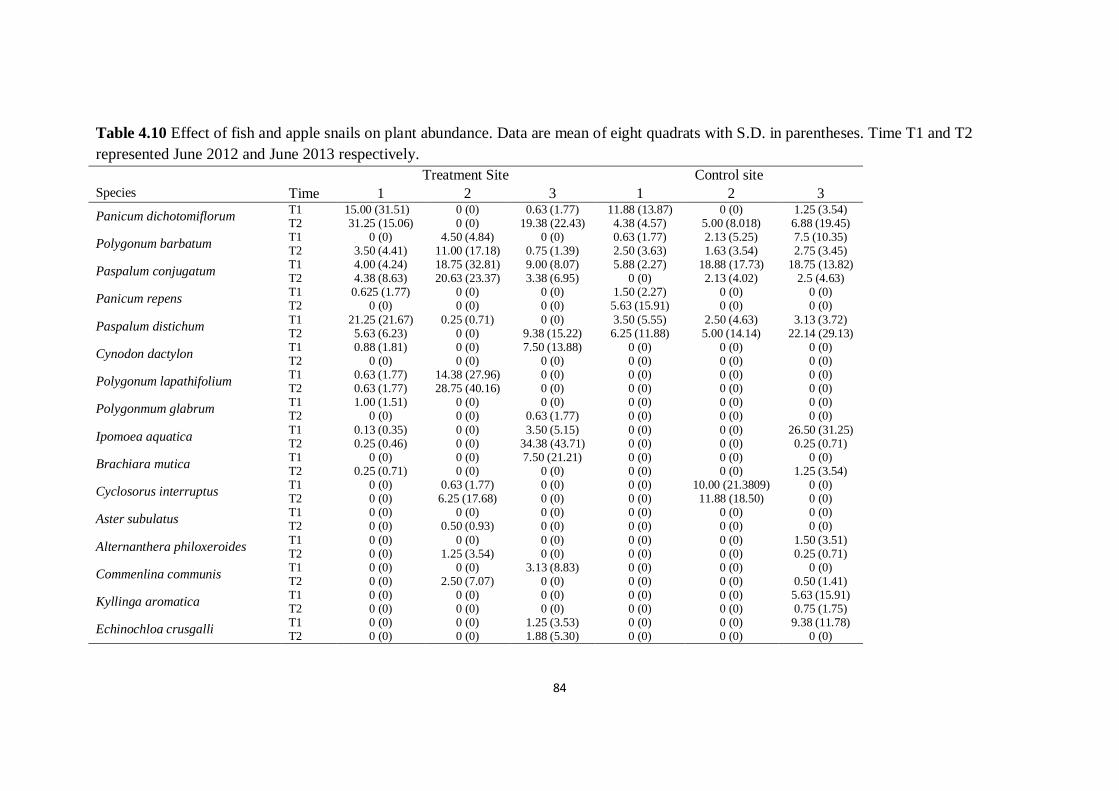
84
Table 4.10 Effect of fish and apple snails on plant abundance. Data are mean of eight quadrats with S.D. in parentheses. Time T1 and T2
represented June 2012 and June 2013 respectively.
Treatment Site Control site
Species Time 1 2 3 1 2 3
Panicum dichotomiflorum T1 15.00 (31.51) 0 (0) 0.63 (1.77) 11.88 (13.87) 0 (0) 1.25 (3.54) T2 31.25 (15.06) 0 (0) 19.38 (22.43) 4.38 (4.57) 5.00 (8.018) 6.88 (19.45)
Polygonum barbatum T1 0 (0) 4.50 (4.84) 0 (0) 0.63 (1.77) 2.13 (5.25) 7.5 (10.35) T2 3.50 (4.41) 11.00 (17.18) 0.75 (1.39) 2.50 (3.63) 1.63 (3.54) 2.75 (3.45)
Paspalum conjugatum T1 4.00 (4.24) 18.75 (32.81) 9.00 (8.07) 5.88 (2.27) 18.88 (17.73) 18.75 (13.82) T2 4.38 (8.63) 20.63 (23.37) 3.38 (6.95) 0 (0) 2.13 (4.02) 2.5 (4.63)
Panicum repens T1 0.625 (1.77) 0 (0) 0 (0) 1.50 (2.27) 0 (0) 0 (0) T2 0 (0) 0 (0) 0 (0) 5.63 (15.91) 0 (0) 0 (0)
Paspalum distichum T1 21.25 (21.67) 0.25 (0.71) 0 (0) 3.50 (5.55) 2.50 (4.63) 3.13 (3.72) T2 5.63 (6.23) 0 (0) 9.38 (15.22) 6.25 (11.88) 5.00 (14.14) 22.14 (29.13)
Cynodon dactylon T1 0.88 (1.81) 0 (0) 7.50 (13.88) 0 (0) 0 (0) 0 (0) T2 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
Polygonum lapathifolium T1 0.63 (1.77) 14.38 (27.96) 0 (0) 0 (0) 0 (0) 0 (0) T2 0.63 (1.77) 28.75 (40.16) 0 (0) 0 (0) 0 (0) 0 (0)
Polygonmum glabrum T1 1.00 (1.51) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) T2 0 (0) 0 (0) 0.63 (1.77) 0 (0) 0 (0) 0 (0)
Ipomoea aquatica T1 0.13 (0.35) 0 (0) 3.50 (5.15) 0 (0) 0 (0) 26.50 (31.25) T2 0.25 (0.46) 0 (0) 34.38 (43.71) 0 (0) 0 (0) 0.25 (0.71)
Brachiara mutica T1 0 (0) 0 (0) 7.50 (21.21) 0 (0) 0 (0) 0 (0) T2 0.25 (0.71) 0 (0) 0 (0) 0 (0) 0 (0) 1.25 (3.54)
Cyclosorus interruptus T1 0 (0) 0.63 (1.77) 0 (0) 0 (0) 10.00 (21.3809) 0 (0) T2 0 (0) 6.25 (17.68) 0 (0) 0 (0) 11.88 (18.50) 0 (0)
Aster subulatus T1 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) T2 0 (0) 0.50 (0.93) 0 (0) 0 (0) 0 (0) 0 (0)
Alternanthera philoxeroides T1 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1.50 (3.51) T2 0 (0) 1.25 (3.54) 0 (0) 0 (0) 0 (0) 0.25 (0.71)
Commenlina communis T1 0 (0) 0 (0) 3.13 (8.83) 0 (0) 0 (0) 0 (0) T2 0 (0) 2.50 (7.07) 0 (0) 0 (0) 0 (0) 0.50 (1.41)
Kyllinga aromatica T1 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 5.63 (15.91) T2 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0.75 (1.75)
Echinochloa crusgalli T1 0 (0) 0 (0) 1.25 (3.53) 0 (0) 0 (0) 9.38 (11.78) T2 0 (0) 0 (0) 1.88 (5.30) 0 (0) 0 (0) 0 (0)
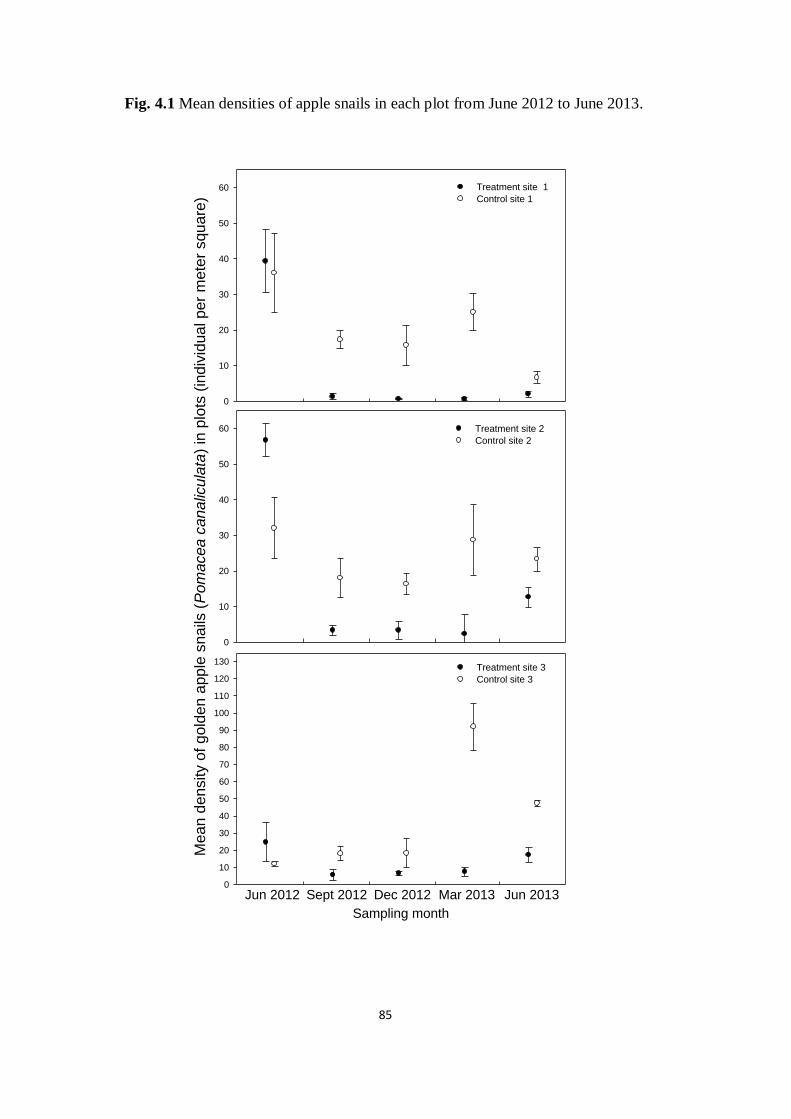
85
Fig. 4.1 Mean densities of apple snails in each plot from June 2012 to June 2013.
0
10
20
30
40
50
60 Treatment site 1
Control site 1M
ean d
ensity o
f g
old
en a
pple
snails
(P
om
acea c
analic
ula
ta)
in p
lots
(in
div
idual per
mete
r square
)
0
10
20
30
40
50
60 Treatment site 2
Control site 2
Sampling month
0
10
20
30
40
50
60
70
80
90
100
110
120
130Treatment site 3
Control site 3
Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013
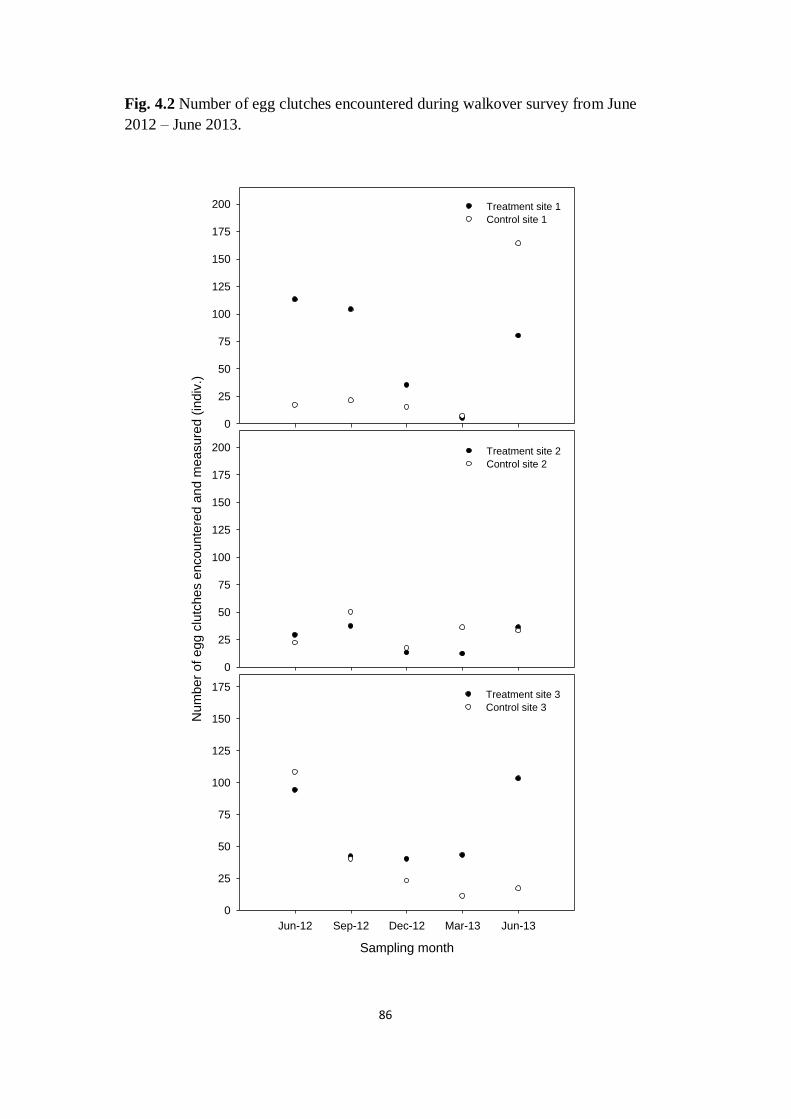
86
Fig. 4.2 Number of egg clutches encountered during walkover survey from June
2012 – June 2013.
0
25
50
75
100
125
150
175
200 Treatment site 1
Control site 1
Nu
mb
er
of
eg
g c
lutc
he
s e
nco
un
tere
d a
nd
me
asu
red
(in
div
.)
0
25
50
75
100
125
150
175
200 Treatment site 2
Control site 2
Sampling month
Jun-12 Sep-12 Dec-12 Mar-13 Jun-13
0
25
50
75
100
125
150
175Treatment site 3
Control site 3
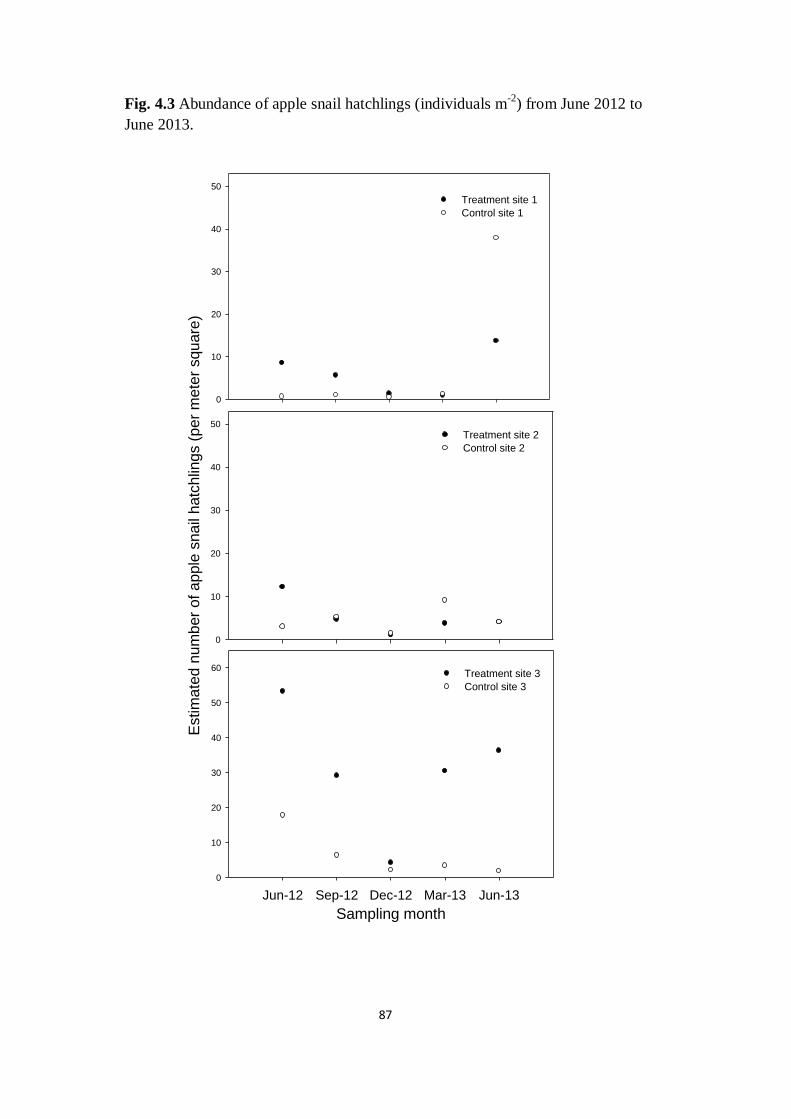
87
Fig. 4.3 Abundance of apple snail hatchlings (individuals m-2
) from June 2012 to
June 2013.
0
10
20
30
40
50
Treatment site 1
Control site 1
Estim
ate
d n
um
ber
of
apple
sna
il h
atc
hlin
gs (
per
mete
r squ
are
)
0
10
20
30
40
50
Treatment site 2
Control site 2
Sampling month
Jun-12 Sep-12 Dec-12 Mar-13 Jun-13
0
10
20
30
40
50
60Treatment site 3
Control site 3

88
Fig. 4.4 Size-frequency distribution of Pomacea canaliculata in experimental and control plots from June 2012 to June 2013. Each bar
of the graph represented sum of total number of apple snail in 1.5m2.
<5
5-1
0
11
-15
16
-20
21
-25
26
-30
31
-35
36
-40
41
-45
46
-50
51
-55
56
-60
Fre
qu
en
cy o
f o
ccu
ren
ce
(in
div
idu
als
)
0
5
10
15
20
25
30
35
0
5
10
15
20
25
30
35
0
5
10
15
20
25
30
35
0
5
10
15
20
25
30
35
<5
5-1
0
11-1
5
16-2
0
21-2
5
26-3
0
31-3
5
36-4
0
41-4
5
46-5
0
51-5
5
56-6
0
0
5
10
15
20
25
30
35
Snail shell length (mm)
<5
5-1
0
11
-15
16
-20
21
-25
26
-30
31
-35
36
-40
41
-45
46
-50
51
-55
56
-60
<5
5-1
0
11-1
5
16-2
0
21-2
5
26-3
0
31-3
5
36-4
0
41-4
5
46-5
0
51-5
5
56-6
0
<5
5-1
0
11-1
5
16-2
0
21-2
5
26-3
0
31-3
5
36-4
0
41-4
5
46-5
0
51-5
5
56-6
0
0
5
10
15
20
25
30
35
40
45
50
0
5
10
15
20
25
30
35
40
45
50
0
5
10
15
20
25
30
35
40
45
50
0
5
10
15
20
25
30
35
40
45
50
<5
5-1
0
11-1
5
16-2
0
21-2
5
26-3
0
31-3
5
36-4
0
41-4
5
46-5
0
51-5
5
56-6
0
0
5
10
15
20
25
30
35
40
45
50
Treatment site 1 Control site 1 Treatment site 2 Control site 2 Treatment site 3 Control site 3
Jun-12 (59)
Sept-12 (2)
Dec-12 (1)
Mar-13 (1)
Jun-13 (3)
Jun-12 (54)
Sep-12 (24)
Dec-12 (23)
Mar-13 (38)
Jun-13 (10)
Jun-12 (87)
Sep-12 (5)
Dec-12 (6)
Mar-13 (11)Mar-13 (8)
Jun-13 (19)
Jun-12 (41)
Sep-12 (15)
Dec-12 (28)
Mar-13 (43)
Jun-13 (35)
Jun-12 (37)
Sep-12 (8)
Dec-12(40)
Mar-13(11)
Jun-13 (26)
Jun-12 (18)
Sep-12 (21)
Dec-12 (28)
Mar-13 (138)
Jun-13(71)
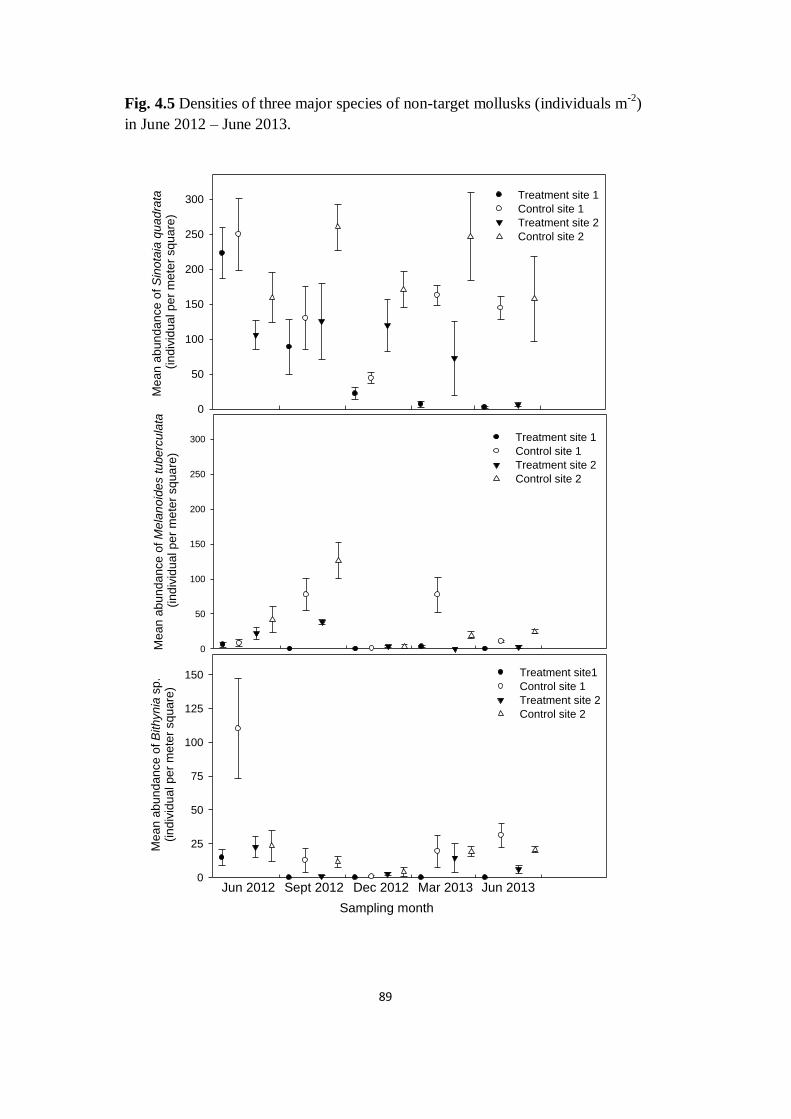
89
Fig. 4.5 Densities of three major species of non-target mollusks (individuals m-2
)
in June 2012 – June 2013.
M
ea
n a
bu
nd
an
ce
of M
ela
no
ide
s tub
erc
ula
ta
(in
div
idu
al pe
r m
ete
r sq
ua
re)
Me
an
ab
un
da
nce
of B
ithyn
ia s
p.
(in
div
idu
al pe
r m
ete
r sq
ua
re)
0
25
50
75
100
125
150 Treatment site1
Control site 1
Treatment site 2
Control site 2
Sampling month
Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013
Me
an
ab
un
da
nce
of S
ino
taia
qu
ad
rata
(in
div
idu
al p
er
me
ter
sq
ua
re)
0
50
100
150
200
250
300 Treatment site 1
Control site 1
Treatment site 2
Control site 2
0
50
100
150
200
250
300 Treatment site 1
Control site 1
Treatment site 2
Control site 2
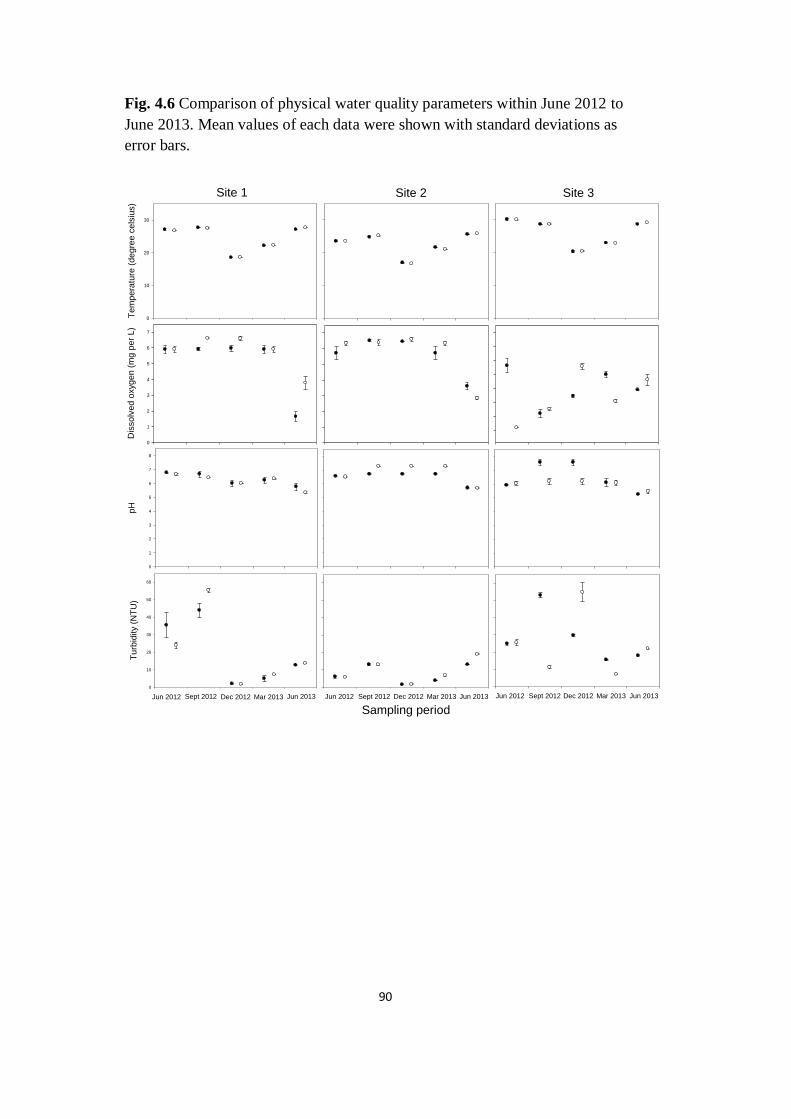
90
Fig. 4.6 Comparison of physical water quality parameters within June 2012 to
June 2013. Mean values of each data were shown with standard deviations as
error bars.
Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013 Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013 Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013
Site 1 Site 2 Site 3
Tem
pera
ture
(degre
e c
els
ius)
0
10
20
30
Turb
idity (
NT
U)
0
10
20
30
40
50
60
Dis
solv
ed o
xygen (
mg p
er
L)
0
1
2
3
4
5
6
7
pH
0
1
2
3
4
5
6
7
8
Sampling period
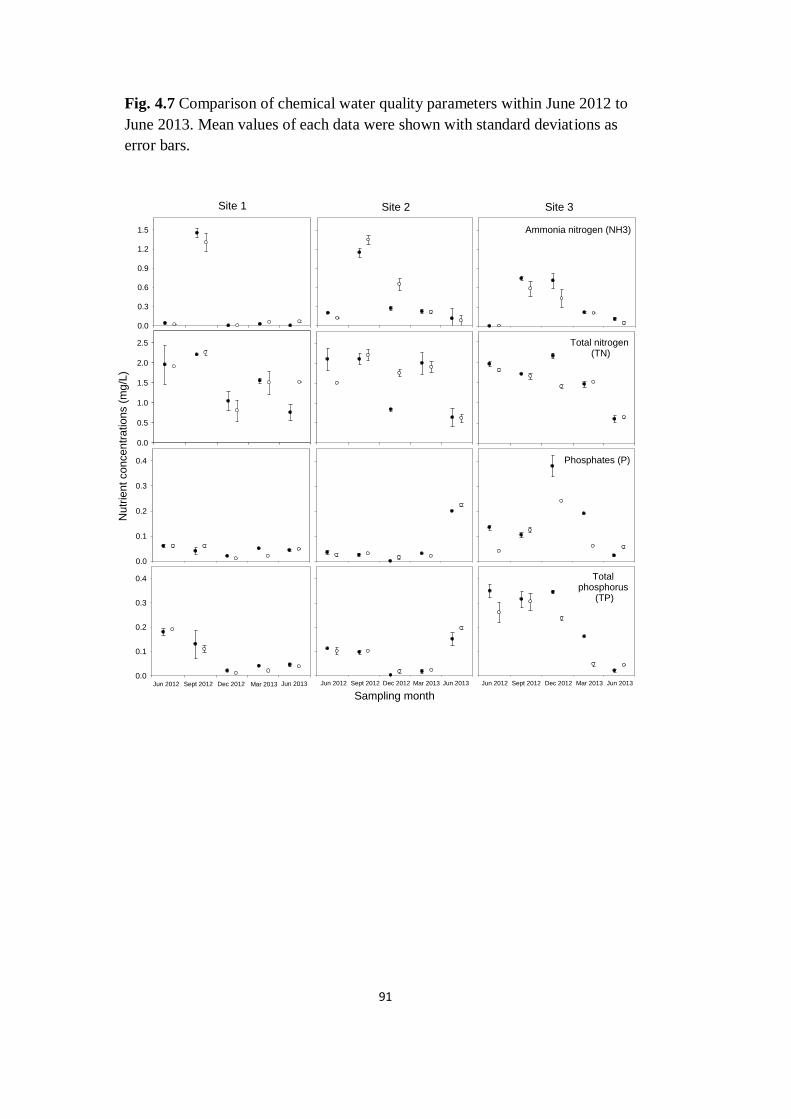
91
Fig. 4.7 Comparison of chemical water quality parameters within June 2012 to
June 2013. Mean values of each data were shown with standard deviations as
error bars.
Site 1 Site 2 Site 3
0.0
0.3
0.6
0.9
1.2
1.5
0.0
0.1
0.2
0.3
0.4
0.0
0.5
1.0
1.5
2.0
2.5
Nutr
ient concentr
ations (
mg/L
)
0.0
0.1
0.2
0.3
0.4
Sampling month
Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013 Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013 Jun 2012 Sept 2012 Dec 2012 Mar 2013 Jun 2013
Ammonia nitrogen (NH3)
Total nitrogen (TN)
Phosphates (P)
Total phosphorus
(TP)
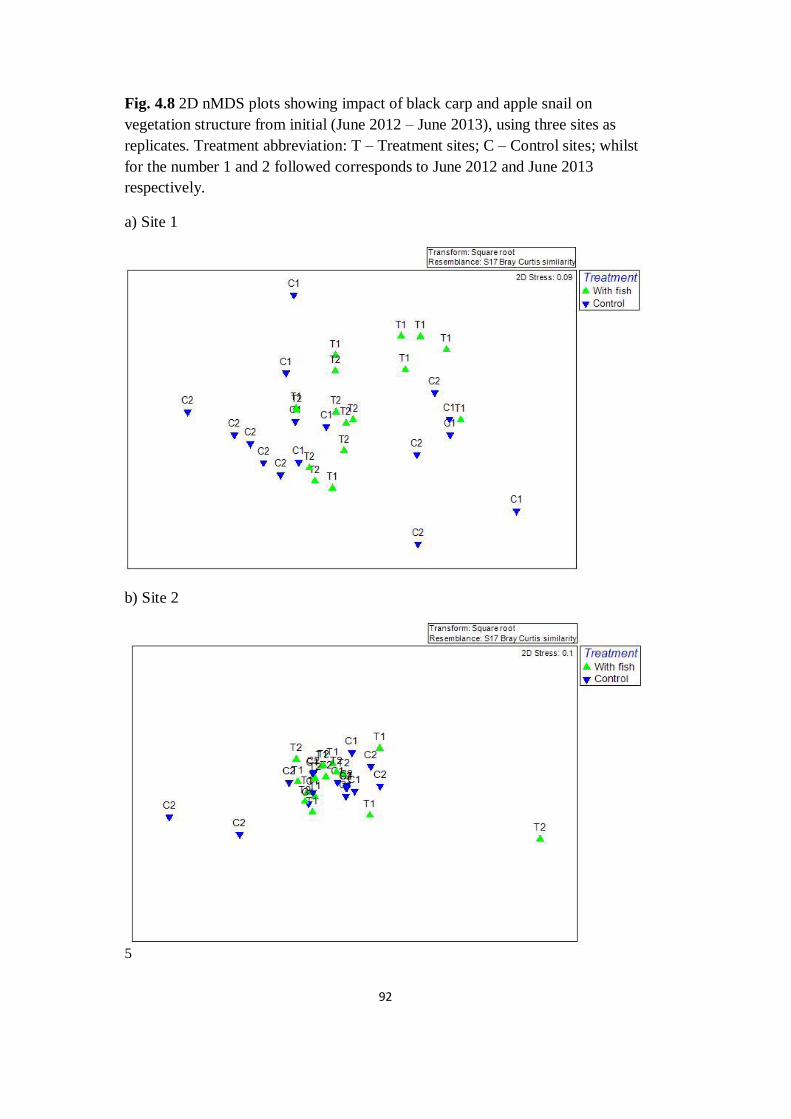
92
Fig. 4.8 2D nMDS plots showing impact of black carp and apple snail on
vegetation structure from initial (June 2012 – June 2013), using three sites as
replicates. Treatment abbreviation: T – Treatment sites; C – Control sites; whilst
for the number 1 and 2 followed corresponds to June 2012 and June 2013
respectively.
a) Site 1
b) Site 2
5
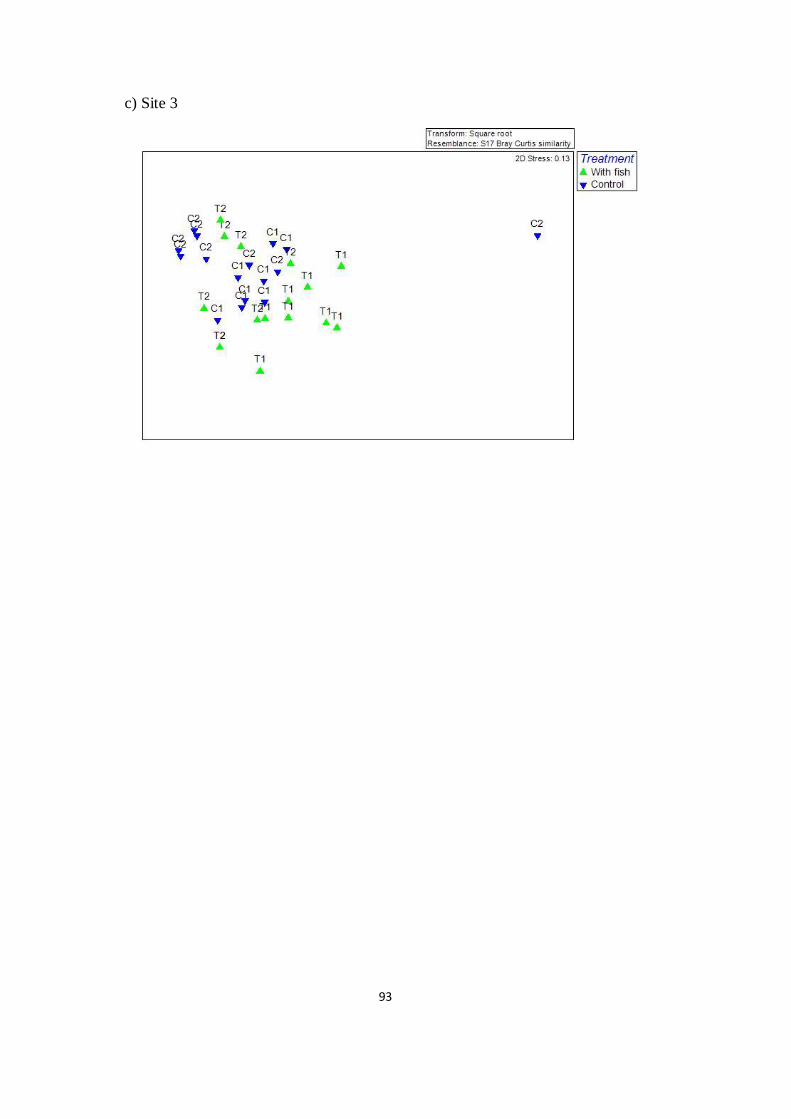
93
c) Site 3

94
Chapter 5 General summary and discussion
5.1 Summary of information on local invasion history and possible
indigenous predators of apple snails
Studies since the 1980s have showed that two species of apple snails (P.
canaliculata Lamarck and P. maculata) may have invaded Hong Kong wetlands
(Hayes et al. 2008; Hayes et al. 2012; Kwong et al. 2008; Lv et al.2013; Mochida
1991). They have first been reported to present in water channels, in which are
often organically enriched in Hong Kong. The high tolerance to physical stress
might have allowed these apple snails to adapt and breed in channels, as
numerous egg clutches are seen on various solid surfaces and on vegetation
during wet season (Ip pers. obs.). This inevitably adds propagule pressure in
facilitating its subsequent spread in the northern New Territories through the
interconnected water channels (Simberloff 2009). During a systematic survey
conducted in 2008 (Kwong et al. 2008), apple snails were found to be widespread
in New Territories and cause economic loss by devouring semi-aquatic
vegetables in farmlands. In non-agricultural wetlands, the presence of apple
snails resulted in dramatic changes to the lowland agricultural and non-
agricultural freshwater ecosystems. Apple snails graze native and exotic aquatic
macrophytes (Qiu and Kwong 2009; Wong et al. 2009; Wong et al. 2010).
Grazing could trigger excessive influx of nutrients from macrophytes into water,
and shifting succession pressure to favor growth of phytoplanktons (Carlsson et
al. 2004a; Fang et al. 2010). In addition, apple snail grazes selectively on more
nutritious aquatic plants with lower dry matter content (Wong et al. 2010).
Among them, some native macrophytes are preferred over exotic ones from
South America, the home region for apple snails. This finding supports the

95
notion that apple snails are not a safe bio-control agent without invasion risk
(Cowie, 2001). Apple snails are unique invasive species that have devoured
living aquatic plants, directly compete with other grazing macroinvertebrates
through more efficient feeding, resulting in a much higher secondary production
estimate than any other invertebrates recorded within the territory (Dudgeon
1999; Kwong et al. 2009). In short, apple snails possess multiple traits of a
superior competitor and a high impact invader (van der Valk 2012).
Current management of apple snails in Hong Kong is effective but labor
intensive and time-consuming. Control of apple snails in the agricultural area
relies on application of lime and natural molluscides with saponin. In non-
agricultural wetlands, managers mainly rely on hand-picking of adult snails and
detachment of their egg masses, but bio-control of gastropods using fish has
never been explored in local wetlands. A literature review and communication
with local wetland managers suggested there are no effective natural predators in
local waters. Ducks have been demonstrated to be effective predators of apple
snails in Malaysia (Teo, 2001), Philippines (Cagauan et al., 2000), Japan (Yusa et
al., 2006) and China (Liang et al., 2013). But duck farming industry (Carey et al.
2001; Kausar, 1983; Sin and Cheng, 1976; Tuen Mun and Yuen Long District
Planning Office, 2002) had stopped in two decades ago, and since 2006 it has
become illegal for individual residents to keep rearing ducks due to the issue of
public health as to Avian Flu infections (Department of Justice, HKSAR, 2013).
Various species of fish, especially carp, are known to be effective predators
of apple snails (Carlsson et al., 2004b; Halwart et al., 1998; Ichinose et al., 2002;
Liao et al., 2004; Teo, 2006; Wong et al., 2009; Yusa et al., 2006). However, no

96
study has been conducted to evaluate the application of black carp as a bio-
control agent in shallow water non-agricultural wetlands where non-target effects
on macrophytes and indigenous invertebrates might be of management concern.
Nor have previous studies considered the effects of season on apple snail
reproduction and development, and consequently on the effectiveness of the
biological control. We have conducted laboratory test and field implementation
trials to select indigenous fish as biological control agents against apple snails
and to detect possible side-effects of these fish on non-target wetland flora and
fauna.
5.2 Predatory potential of different local fish species on apple snails
An indoor experiment has been conducted in October 2011 in Hong Kong
Baptist University to screen out effective predators of apple snails. Three
indigenous freshwater fish species were selected due to their promising results
based on literature review. The experiment was conducted in separate aquaria
using one individual of each species of fish. Six aquaria of each fish species were
inspected after 48 hours. It aimed to compare the predatory potential of black
carp (Mylopharyngodon piceus), common carp (Cyprinus carpio) and white-
spotted catfish (Clarias fuscus). Both black and common carp are highly
effective predators of apple snails. Results from our aquarium experiment
suggested the two species of carp are more effective predators on apple snails
than catfish. With 100-140 individuals of apple snails offered ad libitum, an
average of 54.2% and 61.0% were predated by common carp and black carp
respectively. Both species are gape limited predators and snails were engulfed
and crushed by their pharyngeal teeth before being swallowed. Catfish crushed a

97
significantly lower amount of apple snails than both carp species, in which
86.1% on average survived.
5.3 Mesocosm studies on functional practicability of fish: testing efficacies
and subtle effects in a nutshell
Molluscivorous fish, especially carp, have been adopted as biological control
agents of golden apple snail, but previous studies have mainly focused on their
effectiveness, with little attention paid to their undesirable side effects on the
non-target plants and animals. To further explore fish as a bio-control agent of
apple snails, a mesocosom study was conducted using black carp
(Mylopharyngodon piceus). The aim of the study was to determine whether black
carp is as effective as common carp as a bio-control agent for apple snails, and
whether it will graze on macrophytes and prey on non-Pomacea snails. The
experimental setup was very similar with that in Wong et al. (2009), except using
two fish species, common carp and black carp in this study. The experimental
duration was also 2 months. Both species of carp preyed effectively on P.
canaliculata, removing almost all individuals that were small enough to fit into
their mouths. Of the 234 apple snails introduced into each of the enclosures, the
mean survival rate was as high as 93% in the control, but only 5% in the common
carp treatment and 2% in the black carp treatment. Apple snail mortality was
size-dependent, with smaller individuals (≤2.0 cm SW) suffering the most severe
mortality.
The effects of the two fish species on macrophytes were different. There
was a great increase in the total macrophyte biomass in the control, from 250 g in
the beginning to 542.6 g at the end. Macrophyte biomass in treatments with apple

98
snails or apple snails plus carp was remarkably lower than the control: only 30%
in the AS treatment, 32.5% in the AS+CC treatment and 56% in the AS+BC
treatment. This experiment proved the ability of black carp to reduce herbivory
on macrophytes through minimizing apple snail density. However, common carp
reduced apple snail density but did not result in a lower level of herbivory
because it also grazed on macrophytes. Non-target mollusk density was reduced
by both fish species.
5.4 Wetland scale application of biological control of apple snails using black
carp: efficacies and subtle effect to ecosystem
In order to demonstrate the practical efficacies of black carp against apple snails
and its subtle effects to organisms and ecosystem, a large-scale field experiment
was conducted during June 2012- June 2013 in three pieces of separated
freshwater wetland as replicates.
We found that black carp is a highly tolerant species that can survive in
shallow stagnant waters. This species can be used as a biological control agent of
apple snails as a predator of juvenile apple snails (SL: <5 – 25mm) in shallow
freshwater wetlands that are common in southern China. A major decline of
89.2% in apple snail overall densities have been recorded on average in three
experimental sites with treatment after one year. Juvenile snails would be
eradicated before they get to mature minimum size (male SL: 25.2 ± 3.3mm;
female: 29.8 ± 3.6mm (Estoy et al. 2002)) for reproduction. Given the longevity
of black carp, a low stocking density (80 - 89 individuals ha-1
) will be sufficient
to control apple snail populations, as adults die out. Additionally, black carp
culture in wetlands can provide commercial incentives as it is an edible fish. Yet

99
implementation of black carp to control apple snails should only be carried out in
habitats with low biodiversity of macro-invertebrates, especially mollusks.
Juvenile specimens (total length: at least approximately 300mm) are most
preferable option for release as they could reach a wider range of habitats
including shallow riparian area where snails aggregate, while still beholding their
high adaptability to adverse environmental conditions in shallow wetlands. Our
results also indicated that application of black carp to control apple snail
populations reduced phosphorus nutrients influx thus preventing aquatic
macrophyte community from shifting towards plankton-dominating state.
5.5 Implications on integrated pest management
A call for integrated pest management has prevailed in recent years (Peshin and
Dhawan 2009). Our study has implications on methods to eradicate pest snails in
shallow water wetlands, with much lower input of labor and management-
associated cost than hand-picking. Using fish as a single controlling agent is a
viable method in permanent and seasonal wetlands infested with apple snails,
where pre-treatment options like natural molluscicides application could not be
applied. To maximize treatment efficacies, it should be supplemented with
mechanical methods, including regular removal of adult snails and detachment of
egg clutches. Supplement methods above should be practiced in the riparian
region of wetland with very shallow water levels and dense vegetation that could
not allow fish access. In seasonality aspect, hand-picking should be conducted
most frequently during wet season (April – September annually) where plenty of
shallow pools generate as the ponds are in spate with rainwater.

100
Appendices
Plate 1Site photos and general growth condition of three species of macrophytes
at the end of 8-week mesocosm experiment. A: Blocks of enclosures at
experimental site; B: Enclosures with meshed nylon cover; C: Control enclosure;
D: Enclosure with AS treatment; E: Enclosure with AS+BC treatment; F:
Enclosure with AS+CC treatment.

101
Plate 2 Photos showing the three sites used as replicates in large-scaled field
implementation experiment in Kamtin, New Territories.
Control plot 1 Experimental plot 1
Control plot 2 Experimental plot 2
Control plot 3 Experimental plot 3

102
Plate 3 Photos showing a list of common benthic macro-invertebrates found in
samples.
Sinotaia quadrata
Melanoides tuberculata
Class Aeshnidae
Darner dragonfly
Orthetrum sabina
Green Skimmer
Ischnura senegalensis
Common Bluetail
9mm
17mm 16mm
Bithynia sp.

103
Pantala flavescens
Wandering Glider
Family Pleidae
Pygmy backswimmer
Family Naucoridae
Creeping waterbug
Family Notonectidae
Backswimmer
Family Palemonidae
Shrimp
Family Corixidae
Water Boatmen

104
References
Ali, A.B., 1992. Rice-fish farming development in Malaysia: past, present and
future. In: Dela Cruz, C.R., Lightfoot, C., Costa-Pierce, B.A., Carangal,
V.R., Bimbao, M.P., (Eds.), Rice-fish Research and Development in Asia.
Proceedings of 24th ICLARM Conference. Manila. pp. 69-74.
Ali, S., Ripley, S.D., 1998. Handbooks of the birds of Indian and Pakistan
together with those of Bangladesh, Nepal, Sikkim, Bhutan and Sri Lanka,
no. 9. Second edition. Oxford University Press, Kolkata.
Allcock, J., Carey, G., Chow, G., Welch, G., 2013. Hong Kong Bird Report. The
Hong Kong Bird Watching Society. 288p.
Amano, T., Kusumoto, Y., Tokuoka, Y., Yamada, S., Kim, E.Y., Yamamoto, S.,
2008. Spatial and temporal variations in the use of rice-paddy dominated
landscapes by birds in Japan. Biological Conservation 141, 1704-1716.
Anderson, B., 1993. The Philippine snail disaster. The Ecologist 23, 70.
Ballantyne, L., Fu, X.H., Shih, C.H., Cheng, C.Y., Yiu, V., 2011. Pteroptyx
maipo Ballantyne, a new species of bent-winged firefly (Coleoptera:
Lampyridae) from Hong Kong, and its relevance to firefly biology and
conservation. Zootaxa 2931, 8-34.
Batzer, D.P., Pusateri, C.R., Vetter, R., 2000. Impacts of fish predation on marsh
invertebrates: direct and indirect effects. Wetlands 20, 307-312.
Ben-Ami, F., Heller, J., 2001. Biological control of aquatic pest snails by the
black carp Mylopharyngodon piceus. Biological Control 22, 131-138.
Billard, R., 1999. Biology of the cyprinids. In: Billard, R., (Ed.), Carp - biology
and culture. Praxis Publishing Ltd., Chichester, U.K., INRA, Paris,
France, pp. 1-8.
Birdlife International, 2001. Malaysian Whistling-Thrush Myophonus robinsoni.
Threatened birds of Asia: the BirdLife International Red Data Book.
BirdLife International, Cambridge, pp. 1993-1994.
Boland, B.B., Meerhoff, M., Fosalba, C., Mazzeo, N., Barnes, M.A., Burks, R.L.,
2008. Juvenile snails, adult appetites: contrasting resource consumption
between two species of apple snails (Pomacea). Journal of Molluscan
Studies 74, 47-54.

105
Brönmark, C., 1988. Effects of vertebrate predation on freshwater gastropods an
exclosure experiment. Hydrobiologia 169, 363-370.
Brönmark, C., Hansson, L.A., 2005. Food web interactions in freshwater
ecosystems. Chapter 5. The biology of lakes and ponds. Second edition.
Oxford University Press Inc. New York, United States, pp. 163-206.
Cagauan, A.G., Branckaert, R.D.S., Van Hove, C., 1999. Rice-duck farming in
Asia: Increasing its production potential by integration with fish and the
nitrogen fixing aquatic fern Azolla. The scope and effect of family
poultry research and development: INFPDE-conferences, Animal
production and health division, FAO agriculture department Additional
paper no. 4.
Cagauan, A.G., Branckaert, R.D.S., Van Hove, C., 2000. Integrated fish and
Azolla into rice-duck farming in Asia. Naga, the ICLARM Quarterly 23,
4-10.
Cagauan, A.G., Joshi, R.C., 2002. Predation of freshwater fish on golden apple
snail, Pomacea canalicuala Lam., under screenhouse conditions.
International Rice Research Notes 27, 24-26.
Carey, G., Chalmers, M.L., Diskin, D.A., Kennerley, P.R., Leader, P.J., Leven,
M.R., Lewthwaite, R.W., Melville, D.S., Turnbill, M., Young, L., 2001.
The avifauna of Hong Kong. Hong Kong Bird Watching Society, Hong
Kong. 563p.
Carpenter, S.R., Lodge, D.M., 1986. Effects of submerged macrophytes on
ecosystem process. Aquatic Botany 26, 341-370.
Carlsson, N.O.L., Brönmark, C., 2006. Size-dependent effects of an invasive
herbivorous snail (Pomacea canaliculata) on macrophytes and periphyton.
Freshwater Biology 51, 695-704.
Carlsson, N.O.L., Brönmark, C., Hansson, L.A., 2004a. Invading herbivory: the
golden apple snail alters ecosystem functioning in Asian wetlands.
Ecology 85, 1575-1580.
Carlsson, N.O.L., Kestrup, Å., Mårtensson, M., Nyström, P., 2004b. Lethal and
non-lethal effects of multiple indigenous predators on the invasive golden
apple snail (Pomacea canaliculata). Freshwater Biology 49, 1269-1279.

106
Carlsson, N.O.L., Lacoursière, J.O., 2005. Herbivory on aquatic vascular plants
by the introduced golden apple snail (Pomacea canaliculata) in Lao PDR.
Biological Invasions 7, 233-241.
Cazzaniga, N.J., 2006. Pomacea canaliculata: Harmless and useless in its natural
realm (Argentina). In: Joshi, R.C., Sebastian, L.S., (Eds.), Global
advances in ecology and management of golden apple snails. 37-61pp.
Nueva Ecija: Philippine Rice Research Institute.
Cha, M.W., 1989. Interspecific interactions among freshwater gastropoda in
Hong Kong. MPhil. thesis. Department of Applied Biology and Chemical
Technology. Hong Kong Poytechnic University, Hong Kong.
Cha, M.W., 2004. The application of constructed wetlands for water quality
improvement in the deep bay catchment of Hong Kong. In: Wong, M.H.,
(Ed.), Wetlands Ecosystems in Asia: Function and Management. Elsevier,
Amsterdam, pp. 401-412.
Chen, T.H., 2006. Distribution and status of the introduced red-eared slider
(Trachemys scripta elegans) in Taiwan. In: Koike, F., Clout, M.N.,
Kawamichi, M., De Poorter, M., Iwatsuki, K., Eds.), Assessment and
control of biological invasion risks. Shoukadoh Books Sellers, Kyoto,
Japan and the World Conservation Union (IUCN), Gland, Switzerland,
pp. 187-195.
Chen, W.D., 2011. Pomacea scalaris (D’ Orbigny, 1835). In: Freshwater
molluscs of Taiwan. National Museum of Marine Biology and Aquarium.
216-218pp.
Chen, B.J., Li, Y.S., Lin, J.X., 2003. Investigation on outbreaking human cases
and natural infectious focus of Angiostrongylus cantonensis infection in
Changle city, Fujian province. Parasitoses and Infectious Diseases 1, 6-9.
Chen, Y.H., Yang, P.S., Chen, W.T., Chen, Y.H., 2004. The impact of golden
apple snail (GAS) on Taiwan's agricultural and ecological environment:
An economic approach. Proceedings of APEC Syposium on the
Management of the Golden Apple Snail. Institute of Science and
Technology, National Pingtung University.

107
Cheung, S.M., Dudgeon, D., 2006. Quantifying the Asian turtle crisis: market
surveys in southern China, 2000-2003. Aquatic Conservation: Marine
and Freshwater Ecosystem 16, 751-770.
Chu, X.L., Chen, Y.R., 1989. The fishes of Yunnan, China. Part I. Cyprinidae.
Science Press, Beijing.
Clarke, K.R., Gorley, R.N., 2000. Primer 6: user manual / tutorial. Primer-E Ltd.,
Plymouth, United Kingdom, pp. 189.
Collar, N., 2005. Family Turdidae (Thrushes). In: del Hoyo, J., Elliott, A.,
Christie, D.A., Eds.), Volume 10 Cuckoo-shrikes to Thrushes. Handbook
of the birds of the world. Lynx Edicions, Barcelona, pp. 514-807.
Collins, C.B., 1996. The Chinese Black carp: a potential biological control for
snails in warmwater fish production ponds. Aquaculture Magazine May /
June 1996, 83-86.
Collins III, J.J., Wang, B., Lambrus, B.G., Tharp, M.E., Iyer, H., Newmark, P.A.,
2013. Adult somatic stem cells in the human parasite Schistosoma
mansoni. Nature 494, 476-479.
Cowie, R.H., 1995. Identity, distribution and impacts of introduced
Ampullariidae and Viviparidae in the Hawaiian Islands. Journal of
Medical and Applied Malacology 5, 61-67.
Cowie, R.H., 1998. Patterns of introduction of non-indigenous non-marine snals
and slugs in the Hawaiian Islands. Biodiversity and Conservation 7, 349-
368.
Cowie, R.H., 2001. Can snails ever be effective and safe biocontrol agents?
International Journal of Pest Management 47, 23-40.
Cowie, R.H., 2002. Apple snails as agricultural pests: their biology, impacts and
management. In: Barker, G.M., (Ed.), Molluscs as crop pests. CAB
International, Wallingford, UK., pp. 144-192.
Craig, A.J.F.K., Feare, C.J., 2009. Family Sturnidae (Starlings). In: Del Hoyo, J.,
Elliott, A., Christie, D.A., Eds.), Volume 14 Bush-shrikes to Old World
sparrows. Handbook of the birds of the world. Lynx Edicions, Barcelona,
pp. 650-709.

108
Cui, K., Zhao, H., 2012. Clarias fuscus. In: IUCN, (Ed.), IUCN Red List of
Threatened Species. Version 2012.2.
Davis, M.A., Pelsor, M., 2001. Experimental support for a resource-based
mechanistic model of invasibility. Ecological Letters 4, 421-428.
De Clercq, P., Mason, P.G., Babendreier, D., 2011. Benefits and risks of exotic
biological control agents. Biocontrol 56, 681-698.
De Graaf, G., Janssen, H., 1996. Chapter 3 Biology. Artificial reproduction and
pond rearing of the African catfish Clarias gariepinus in sub-Saharan
Africa - a handbook. FAO Fisheries technical paper 362. Food and
Agriculture Organization of the United Nations, Rome.
de Silva, P.K., 1996. Food and feeding habits of the Eurasian Otter Lutra lutra L.
(Carnivora, Mustelidae) in Sri Lanka. Journal of South Asian Natural
History 2, 81-90.
de Silva, P.K., 1997. Seasonal variation of the food and feeding habits of the
Eurasian otter Lutra lutra L.(Carnivora, Mustelidae) in Sri Lanka.
Journal of South Asian Natural History 2, 205-216.
Debus, S.J.S., Madge, S.C., Marzluff, J.M., 2009. Family Corvidae (Crows). In:
Del Hoyo, J., Elliott, A., Christie, D.A., Eds.), Volume 14 Bush-shrikes
to Old World sparrows. Handbook of the birds of the world. Lynx
Edicions, Barcelona, pp. 494-640.
DeLoach, C.J., 1991. Past successes and current prospects in biological control
of weeds in the United States and Canada. Natural Areas Journal 11,
129-142.
Denisov, A.I., 1982. The epizootiology and prophylaxis of diplostomiasis.
Verterinariya (Moscow, USSR) 8, 40-42. (In Russian).
Department of Justice, HKSAR, 2013. Public health (animals and birds)
Ordinance. Chapter 139L. Ordinances and subsidiary legislation of Hong
Kong.
Dong, S.Z., Zheng, G.W., Yu, X.P., Fu, C.H., 2012. Biological control of golden
apple snail, Pomacea canaliculata by Chinese soft-shelled turtle,
Pelodiscus sinensis in the wild rice, Zizania latifolia field. Scientia
Agricola 69, 142-146.

109
Dudgeon, D., 1996. Anthropogenic influences on Hong Kong streams.
GeoJournal 40, 53-61.
Dudgeon, D., 1999a. Decapoda: Brachyura. Tropical Asian streams: zoobenthos,
ecology and conservation. Hong Kong University Press, Hong Kong, pp.
177-186.
Dudgeon, D., 1999b. Ecological overview - fish. Tropical Asian streams -
zoobenthos, ecology and conservation. Hong Kong University Press,
Hong Kong, pp. 75-82.
Dudgeon, D., 1999c. Ecological overview – functional organization and the
River Continuum Concept. Tropical Asian streams - zoobenthos, ecology
and conservation. Hong Kong University Press, Hong Kong, pp. 63-70.
Dudgeon, D., 2003. 2- Hillstreams. The Department of Ecology and Biodiversity,
The University of Hong Kong.
Dudgeon, D., Arthington, A.H., Gessner, M.O., Kawabata, Z., Knowler, D.,
Lévêque, C., Naiman, R.J., Prieur-Richard, A.H., Soto, D., Stiassny,
M.L.J., Sullivan, C.A., 2006. Freshwater biodiversity: importance, threats,
status and conservation challenges. Biological Reviews 81, 163-182.
Dudgeon, D., Cheung, C.P.S., 1990. Selection of gastropod prey by a tropical
freshwater crab. Journal of Zoology, London 220, 147-155.
Dudgeon, D., Corlett, R., 2004. The ecology and biodiversity of Hong Kong.
Friends of the Country Parks & Joint Publishing, Hong Kong.
Dudgeon, D., Lam, P.K.S., 1985. Freshwater gastropod foraging strategies:
interspecific comparisons. In: Morton, B., Dudgeon, D., (Eds.), The
Malacofauna of Hong Kong and Southern China II. Hong Kong
University Press, Hong Kong, Hong Kong, pp. 591-600.
Dudgeon, D., Yipp, M.W., 1985. The diets of Hong Kong freshwater gastropods.
In: Morton, B., Dudgeon, D., (Eds.), The Malacofauna of Hong Kong and
Southern China II. Hong Kong University Press, Hong Kong, Hong Kong,
pp. 491-512.
Eaton, A.D., Franson, M.A.H., 2005. Standard methods for the examination of
water and wastewater. 21st Edition. American Public Health Association.

110
Estebenet, A.L., 1995. Food and feeding in Pomacea canaliculata (Gastropoda:
Ampullariidae). The Veliger 38, 277-283.
Estebenet, A.L., Cazzaniga, N.J., 1992. Growth and demography of Pomacea
canaliculata (Gastropoda: Ampullariidae) under laboratory conditions.
Malacological Review 25, 1-12.
Estebenet, A.L., Martin, P.R., 2002. Pomacea canaliculata (Gastropoda:
Ampullaridae): life-history traits and their plasticity. Biocell 26, 83-89.
Fang, L., Wong, P.K., Lin, L., Lan, C.Y., Qiu, J.W., 2010. Impact of invasive
apple snails in Hong Kong on wetland macrophytes , nutrients,
phytoplankton and filamentous algae. Freshwater Biology 55, 1191-1204.
FAO, 1998. Fish-farming in Vietnamese rice fields flights golden apple snail pest.
FAO, News & Highlights.
Fernando, C.H., 1993. Rice-field ecology and fish culture – an overview.
Hydrobiologia 259, 91-113.
Forrest, A.L., Wittmann, M.E., Schmidt, V., Raineault, N.A., Hamilton, A., Pike,
W., Schladow, S.G., Reuter, J.E., Laval, B.E., Trembanis, A.C., 2012.
Quantitative assessment of invasive species in lacustrine environments
through benthic imagery analysis. Limnology and Oceanography:
methods 10, 65-74.
Frandsen, F., 1987. Control of schistosomiasis by use of biological control of
snail hosts with special reference to competition. Memórias do Instituto
Oswaldo Cruz 82, 129-133.
Frei, M., Khan, M.A.M., Razzak, M.A., Hossain, M.M., Dewan, S., Becker, K.,
2007. Effects of a mixed culture of common carp, Cyprinus carpio L.,
and Nile tilapia, Oreochromis niloticus (L.), on terrestrial arthropod
population, benthic fauna, and weed biomass in rice fields in Bangladesh.
Biological Control 41, 207-213.
Froese, R., Pauly, D., 2011. FishBase - Mylopharyngodon piceus (Richardson,
1846) Black carp. World Wide Web electronic publication.
Fu, W.K., 2010. Population dynamics, diet and morphological variation of the
Hong Kong Newt (Paramesotriton hong kongensis). M.Phil. thesis. The
University of Hong Kong.

111
Fu, X.H., Ohba, N., Fredric V, V., Lei, C.L., 2005. Structure, behavior and the
lifecycle of an aquatic firefly, Luciola substraiata, in China. The
Canadian Entomologist 137, 83-90.
Fu, X.H., Ohba, N., Zhang, Y., Lei, C.L., 2006. A rearing apparatus and diet for
the aquatic firefly Luciola leii (Coleoptera: Lampyridae). The Canadian
Entomologist 138, 399-406.
Gallebu, A.U., Jover, P.C., Bongolan, F.F., 1992. Integration of ducks to lowland
culture as biological control of the golden apple snail. Philippine Journal
of crop science 17, 27.
Gangstad, E.O., 1980. Weed control methods for public health applications. CRC
Press, Boca Raton, Florida.
Gozlan, R.E., Britton, J.R., Cowx, I., Copp, G.H., 2010. Current knowledge on
non-native freshwater fish introductions. Journal of Fish Biology 76, 751-
786.
Halwart, M. 1994a. Fish as bio-control agents in rice. Tropical Agroecology 8,
169. Margraf Verlag. Germany.
Halwart, M., 1994b. The golden apple snail Pomacea canaliculata in Asian rice
farming systems: present impact and future threat. International Journal
of Pest Management 40, 199-206.
Halwart, M., Litsinger, J.A., in press. Efficacy of fish as biocontrol agents of rice
prests in the Philippines. International Journal of Pest Management.
Halwart, M., Poethke, H.J., Kaule, G., 2006. Golden snail population ecology in
rice-fish culture and rice monoculture: a modeling approach. In: Joshi,
R.C., Sebastian, L.S., Eds.), Global advances in ecology and management
of golden apple snails. Nueva Ecija: Philippine Rice Research Institute,
Philippines.
Halwart, M., Viray, M.C., Kaule, G., 1998. Cyprinus carpio and Oreochromis
niloticus as biological control agents of the golden apple snail Pomacea
canaliculata - effects of predator size, prey size and prey density. Asian
Fisheries Science 11, 31-42.
Hamada, Y., Matsumoto, T., 1985. Apple snails in Kumamoto Prefecture.
Kyushu-no-kai 24, 5-12.

112
Hao, L., Wu, K., Cheng, L., Wang, Q., Chen, X.G., 2007. Investigation of
Angiostrongylus cantonensis infection in Pomacea canaliculata in
Guangzhou and other districts. Journal of Tropical Medicine 1.
Haridi, A.A.M., Jobin, W.R., 1985. Estimated risks and benefits from
introducing Marisa cornuarietis into the Sudan. Journal of Tropical
Medicine and Hygiene 88, 145-151.
Hayes, K.A., Cowie, R.H., Thiengo, S.C., Strong, E.E. 2012. Comparing apples
with apples: clarifying the identities of two highly invasive Neotropical
Ampullariidae (Caenogastropoda). Zoological Journal of the Linnean
Society 166, 723-753.
Hayes, K.A., Joshi, R.C., Thiengo, S.C., Cowie, R.H., 2008. Out of South
America: Multiple origins of non-native apple snails in Asia. Diversity
and Distributions 14, 701-712.
Hermoso, V., Clavero, M., Blanco-Garrido, F., Prenda, J., 2011. Invasive species
and habitat degradation in Iberian streams: an analysis of their role in
freshwater fish diversity loss. Ecological Applications 21, 175-188.
Hickling, C.E., 1971. Fish culture. Faber and Faber Ltd., London.
Higgins, S.N., Vander Zanden, M.J., 2010. What a differencea species makes: a
meta-analysis of dreissenid mussle impacts on freshwater ecosystem.
Ecological Monographs 80, 179-196.
HKBWS, 2006. Handbook to eco-tour guides on Long Valley ecology. Birds and
humans in harmony – a sustainable management scheme in Long Valley.
Ho, J.Z., Chiang, P.H., Wu, C.H., Yang, P.S., 2010. Life cycle of the aquatic
firefly Luciola ficta (Coleoptera: Lampyridae). Journal of Asia-Pacific
Entomology 13, 189-196.
Hodgkiss, I.J., Man, H.S.H., 1977. Stock density and mortality assessment of
Sarotherodon mossambicus (Cichlidae) in Plover Cove Reservoir, Hong
Kong. Environmental Biology of Fishes 1, 171-180.
Hofkin, B.V., Stryker, G.A., Koech, D.K., Loker, E.S., 1991. Consumption of
Biomphalaria glabrata egg masses and juvenilesby the ampullariid snails
Pila ovata, Lanistes carinatus and Marisa cornuarietis. Acta Tropica 49,
37-44.

113
Howarth, F.G., 1991. Environmental impacts of classical biological control.
Annual Review of Entomology 36, 485-509.
Huang, D.C., 2005. Current situation and management of exotic species.
National Policy Foundation Research Report, Taiwan.
Huang, C.F., Zou, J.X., Li, S.Q., Zhou, X.M., 2010. 我国钉螺控制研究在血吸
虫病防治中的应用. Chinese Journal of Zoonoses 10, 963-965.
Hue, N.V., Guo, F.M., Zhang, G.Q., Yost, R.S., Miyasaka, S.C., 1997. Reactions
of copper sulfate with wetland-taro soils in Hawaii. Communications in
Soil Science and Plant Analysis 28, 849-862.
Hulme, P.E., 2009. Trade, transport and trouble: managing invasive species
pathways in an era of globalization. Journal of Applied Ecology 46, 10-18.
Hung, N.M., Duc, N.V., Stauffer Jr., J.R., Madsen, H., 2013. Use of black carp
(Mylopharyngodon piceus) in biological control of intermediate host
snails of fish-borne zoonotic trematodes in nursery ponds in Red River
Delta, Vietnam. Parasites and Vectors 6, 142.
Ichinose, K., Tochihara, M., Wada, T., Suguiura, N., Yusa, Y., 2002. Influence
of common carp in apple snail in a rice field evaluated by a predator-prey
logistic model. International Journal of Pest Management 48, 133-138.
ISSG, 2013. Ecology of Clarias gariepinus. Global Invasive Species Database.
SSC, IUCN.
Ip, K.K.L., Liang, Y., Wu, H.X., Xue, J.Z., Qiu, J.W., submitted. Biological
control of invasive apple snails by two species of carp: effects on non-
target species matter.
Jhang, W.C., 1985. Study of golden apple snail from ecological aspect. Bulletin
of Malacology 11, 43-51.
Johnsgard, P.A., 1981. The plovers, sandpipers and snipes of the world.
University of Nebraska Press, U.S.A. and London, U.K.
Joshi, R.C., 2005. Managing invasive alien mollusc species in rice. International
Rice Research Notes 30, 5-13.
Joshi, R.C., 2007. Problems with the Management of the Golden Apple Snail
Pomacea canaliculata: an Important Exotic Pest of Rice in Asia. In:

114
Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.), Area-Wide
Control of Insect Pests. IAEA, pp. 257-264.
Joshi, R. C., De La Cruz, M.S., Duca, A.V., 2002. Ovicidal effect of a
molluscicide on golden apple snail in the Philippines. International Rice
Research Notes 27: 26-27
Joshi, R. C., M. S. De La Cruz, E. C. Martin, J. C. Cabigat, R. G. Bahatan, E. H.
Abayao, Joe Chow-Awon, N. P. Chilagan and A. B. Cayong. 2001.
Current status of the golden apple snail in the Ifugao Terraces,
Philippines. Journal of Sustainable Agriculture 18(2/3), 71-90.
Joshi, R. C., Sebastian, L. S. (editors), 2006. Global advances in ecology and
management of golden apple snails. Philippine Rice Research Institute,
Nueva Ecija.
Karsen, S.J., Lau, M.W.N., Bogadek, A., 1998. Hong Kong amphibians and
reptiles. Second edition. Provisional Urban Council, Hong Kong.
Karunaratne, L.B., Darby, P.C., Bennetts, R.E., 2006. The effects of wetland
habitat structure on Florida apple snail density. Wetlands 26, 1143-1150.
Kausar, R., 1983. Review on recycling of animal eastes as a source of nutrients
for freshwater fish culture within an integrated livestock system. Paper
for UNDP/ FAO project PA/80/019 - Coordinated national programme
for livestock feed resources and nutrition. FAO Fisheries and Aquaculture
Department.
Kirwan, G.M., 1996. Family Rostratulidae (Painted Snipes). In: del Hoyo, J.,
Elliott, A., Sargatal, J., Eds.), Handbook of the birds of the world -
volume 3. Lynx Edicions, Spain, pp. 821.
Kolar, C.S., Lodge, D.M., 2001. Progress in invasion biology: predicting
invaders. Trends in ecology and evolution 16, 199-204.
Kondo, A., Tanaka, F., 1989. An experimental study of predation by the larvae of
the firefly, Luciola lateralis Motschulsky (Coleoptera: Lampyridae) on
the Apple snail, Pomacea canaliculata Lamarck
(Mesogastropoda:Pilidae). Japanese Journal of Applied Entomology and
Zoology 33, 211-216.

115
Kruatrachue, M., Upatham, E.S., 1993. Biological control of schistosomiasis-
mediating snails in Thailand: simulated field experiments. Journal of
Medical and Applied Malacology 5, 117-128.
Kutty, M.N., 1987. Fish culture in rice fields. United Nations development
programme, Food and Agriculture Organizaion of the United Nations and
Nigerian Institute for oceanography and marine research. Project
RAF/82/009., Available:
http://www.fao.org/docrep/field/003/AC180E/AC180E00.htm#TOC.
Kwong, K.L., Chan, R.K.Y., Qiu, J.W., 2009. The potential of the invasive snail
Pomacea canaliculata as a predator of various life-stages of five species
of freshwater snails. Malacologia 51, 343-356.
Kwong, K.L., Dudgeon, D., Wong, P.K., Qiu, J.W., 2010. Secondary production
and diet of an invasive snail in freshwater wetlands: implications for
resource utilization and competition. Biological Invasions 12, 1153-1164.
Kwong, K.L., Wong, P.K., Lau, S.S.S., Qiu, J.W., 2008. Determinants of the
distribution of apple snails in Hong Kong: Two decades after their initial
invasion. Malacologia 50, 293-302.
Lach, L., Britton, D.K., Rundell, R.J., Cowie, R.H., 2000. Food preference and
reproductive plasticity in an invasive freshwater snail. Biological
Invasions 2, 279-288.
Lam, P.K.S., 1994. Intraspecific life-history variation in Radix plicatulus
(Gastropoda: Pulmonata: Lymnaeidae). Journal of Zoology 232, 435-446.
Lardans, V., Dissous, C., 1998. Snail control strategies for reduction of
Schistosomiasis transmission. Parasitology Today 14, 413-417.
Lau, S.S.S., Lane, S.N., 2002. Biological and chemical factors influencing
shallow eutrophication: a long term study. The Science of the Total
Environment 288, 167-181.
Leader, P., 2009. Inland wetlands. In: Wong, C.L.C., Lam, V.W.Y., Ades,
G.W.J., (Eds.), Ecology of Hong Kong birds, pp. .69-86. Kadoorie Farm
and Botanic Garden.

116
Ledford, J.J., Kelly, A.M., 2006. A comparison of black carp, Redear sunfish and
Blue catfish as biological controls of snail populations. North American
Journal of Aquaculture 68, 339-347.
Lee, V.L.F., Lam, S.K.S., Ng, F.K.Y., Chan, T.K.T., Young, M.L.C., 2004. Field
guide to the freshwater fish of Hong Kong. Friends of the country parks
& Cosmos books Ltd., Agriculture, Fisheries and Conservation
Department, HKSAR.
Lee, Y.C., Wu, W.L., 1996. Study on Pomacea canaliculata (Lamarck, 1819) in
Taiwan. Bulletin of Malacology, Taiwan, 20, 43-50.
Lekagul, B., Mcneely, J.A., 1988. Mammals of Thailand. White Lotus Press,
Bangkok, Thailand.
Lester, P.J., Harmsen, R., 2002. Functional and numerical responses do not
always indicate the most effective predator for biological control: an
analysis of two predators in a two-prey system. Journal of Applied
Ecology 39, 455-468.
Leventer, H., 1979. Biological control of reservoirs by fish. Mekoroth Water
Company, Jordan District, Nazareth Elit, Israel.
Leventer, H., Teltsch B., 1990. The contribution of silver carp
(Hypophthalmichthys molitrix) to biological control of Netofa reservoirs.
Hydrobiologia 191,47-55.
Li, A.P.L., Shih, R.S.H., Chan, E.K.W., Cheng, J.C.Y., 2011. Aquatica leii: a
new record of an aquatic firefly species in Hong Kong. In: Chan, S.K.F.,
Chan, A.S.W., Eds.), Hong Kong Biodiversity: Agriculture, Fisheries
and Conservation Department newsletter. Agriculture, Fisheries and
Conservation Department, HKSARG, Hong Kong, pp. 9-11.
Li, S.Z., Fang, F., 1990. On the geographical distribtuion of the four kinds of
pond-cultured carps in China. Acta Zoologica Sinica 36, 244-250.
Liang, K.M., Zhang, J.E., Fang, L., Zhao, B.L., Luo, M.Z., Parajuli,P., Ouyang,
Y., 2013. The biological control of Pomacea canaliculata population by
rice-duck mutualism in paddy fields. Biocontrol Science and Technology
23, 674-690.

117
Liao, C.T., Lin, C.S., Chang, C.C., 2005. Biological control of golden apple snail
(Pomacea canaliculata) by black carp (Mylopharyngodon piceus) and
walking catfish (Clarias batrachus). In: Lai, P.Y., Chang, Y.F., Cowie,
R.H., (Eds.), Proceedings of the APEC Symposium on the management
of the golden apple snail, 6-14 September 2004, Pingtung, National
Pingtung University of Science and Technology, Taiwan.
Light, T., Marchetti, M.P., 2007. Distingusihing between invasions and habitat
changes as drivers of diversity loss among California’s freshwater fishes.
Conservation Biology 21, 434-446.
Lin, C.J., 2009. Panicum paludosum. A field guide to aquatic and wetland plants
of Taiwan 3: Monocotyledon. Commonwealth Publishing Group, Taiwan,
pp. 252-253.
Lin, S.Y., 1955. Chinese systems of pond stocking. Indo-Pacific Fisheries
Council Proceedings, pp. 113-125.
Litsinger, J.A., Estano, D.B., 1993. Management of the golden apple snail
Pomacea canaliculata (Lamarck) in rice. Crop Protection 12, 363-370.
Liu, C.K., 1955. On productivity of two experimental fish ponds managed with
traditional method of Chinese pisciculture. Acta Hydrobiologica Sinica 1,
25-43.
Liu, L.H., Wu, G.X., Chao, W.X., Wang, Z.L., 1986. Studies on the ecological
effect on spawning of the black carp, the grass carp, the silver carp and
the bighead carp in the Changjiang River after the constructions of the
Gezhouba hydroelectric project. Acta Hydrobiologica Sinica 10, 353-364.
Liu, H.L., Li, H., Zhao, B.X., Liu, W., 1990. Post-larval development of the
masticating apparatus of Black Carp Mylopharyngodon piceus
(Richardson). Acta Hydrobiologica Sinica 14, 310-320.
Lodge, D.M., 1993. Biological invasions: lessons for ecology. Trends in Ecology
and Evolution 8, 133-136.
Lowe, S., Browne, M., Boudjelas, S., De Poorter, M., 2000. 100 of the world’s
worst invasive alien species. The Invasive Species Specialist Group.
IUCN, Auckland.

118
Lv, S., Zhang, Y., Liu, H.X., Hu, L., Yang, K., Steinmann, P., Chen, Z., Wang,
L.Y., Utzinger, J., Zhou, X.N., 2009. Invasive snals and an emerging
infectious disease: results from the first national survey on
Angiostrongylus cantonensis in China. Plos Neglected Tropical Diseases
3, e368.
Lv, S., Zhang, Y., Liu, H.X., Hu, L., Liu, Q., Wei, F.R., Guo, Y.H., Steinmann,
P., Hu, W., Zhou, X.N., Utzinger, J. 2013. Phylogenetic evidence for
multiple and secondary introductions of invasive snails: Pomacea species
in the People’s Republic of China. Diversity and distribution 19, 147-156.
Lv, S., Zhang, Y., Steinmann, P., Zhou, X.N., 2008. Emerging angiostrongyliasis
in mainland China. Emerging Infectious Diseases 14, 161-164.
Mack, R.N., Simberloff, D., Lonsdale, W.M., Evans, H., Clout, M., Bazzaz, F.A.,
2000. Biotic invasions: causes, epidemiology, global consequences, and
control. Ecological Applications 10, 689-710.
Man, S.H., Hodgkiss, I.J., 1981. Hong Kong Freshwater Fishes. The Urban
Council, Hong Kong. 75pp.
Mantel, S.K., Dudgeon, D., 2004a. Dietary variation in predatory shrimp,
Macrobrachium hainanense (Palaemonidae), in Hong Kong forest
streams. Archiv für Hydrobiologie 160, 305-328.
Mantel, S.K., Dudgeon, D., 2004b. Effects of Macrobrachium hainanense
predation on benthic community functioning in tropical Asian streams.
Freshwater Biology 49, 1306-1319.
Martin, P.R., Estebenet, A.L., Cazzaniga, N.J., 2001. Factors affecting the
distribution of Pomacea canaliculata (Gastropoda: Ampullariidae) along
its southernmost natural limit. Malacologia 43, 13-23.
Martin, S.M., 1986. Ciclo reproductivo de Ampullaria canaliculata (Gastropoda;
Ampullariidae) en el área rioplatense. Neotropica 32, 171-181.
Martin, T.H., Crowder, L.B., Dumas, C.F., Burkholder, J.M., 1992. Indirect
effects of fish on macrophytes in bays mountain lake evidence for a
littoral trophic cascade. Oecologia 89, 476-481.

119
Messing, R.H., Wright, M.G., 2006. Biological control of invasive species:
solution or pollution. Frontiers in Ecology and the Environment 4, 132-
140.
Mochida, O., 1991. Spread of freshwater Pomacea snails (Pilidae, Mollusca)
from Argentina to Asia. Micronesia Suppl. 3, 51-62.
Mukherjee, T.K., Moi, P.S., Panandam, J.M., Yang, Y.S., 1992. Integrated
livestock-fish production systems. Proceedings of the FAO/IPT
Workshop on Integrated Livestock-Fish Production Systems. Organising
Secretariat, FAO/IPT Workshop on Integrated Livestock-Fish Production
Systems and Institute of Advanced Studies, University of Malaya, Kuala
Lumpur.
Myers, J.H., Simberloff, D., Kuris, A.M., Carey, J.R., 2000. Eradication revisited:
dealing with exotic species. Trends in Ecology and Evolution 15, 316-320.
Nanjing Institute of Geography, Academia Sinica, 1981. A survey on the
spawning grounds of black carp, grass carp, silver cap, bighead carp and
bream in Huan He. Journal of Fisheries of China 5, 361-367.
Naylor, R., 1996. Invasion in agriculture: assessing the cost of the golden apple
snail in Asia. Ambio 25, 443-448.
Nguma, J.F.M., McCullough, F.S., Masha, E., 1982. Elimination of
Biomphalaria pfeifferi, Bulinus tropicus and Lymnaea natalensis by
ampullarid snail, Marisa cornuarietis, in a man-made dam in northern
Tanzania. Acta Tropica 39, 85-90.
Nico, L.G., Williams, J.D., Jelks, H.L., 2005. Black carp: Biological synopsis
and risk assessment of an introduced fish. American Fisheries Society
special publication 32. American Fisheries Society, Bethesda, Maryland,
USA.
Pan, J.H., Zhong, L., Zheng, C.Y., Wu, H.L., Liu, J.Z., 1991. Mylopharyngodon
Peters, 1880. Leuciscinae. Cypriniformes., The freshwater fishes of
Guangdong province. Guangdong Science and Technology Press,
Guangzhou, pp. 80-82.

120
Paolucci, E.M., MacIsaac, H.J., Ricciardi, A., 2013. Origin matters: alien
consumers inflict greater damage on prey populations than do native
consumers. Diversity and Distribution 19, 988-995.
Pascual, U., Perrings, C., 2007. Developing incentives and economic
mechanisms for in situ biodiversity consevation in agricultural landscapes.
Agriculture, Ecosystems and Environment 121, 256-268.
Paterson, R.A., Townsend, C.R., Poulin, R., Tompkins, D.M., 2011. Introduced
brown trout alter native acanthocephalan infections in native fish. Journal
of Animal Ecology 80, 990-998
Pejchar L., Mooney, H.A., 2009. Invasive species, ecosystem services and
human well-being. Trends in Ecology and Evolution 24, 497-504.
Perera, G., Yong, M., Ferrer, J., 1996. Apple Snails in Aquarium. TFH
Publications, Inc.. Neptune City, New Jersey. 121pp.
Peshin, R., Dhawan, A.K., 2009. Integrated pest management: innovation-
development process. Volume 1 and 2. Springer Science and Business
Media B.V..
Piersma, T., van Gils, J., Wiersma, P., 1996. Family Scolopacidae (Sandpipers,
Snipes and Phalaropes). In: del Hoyo, J., Elliott, A., Sargatal, J., Eds.),
Volume 3 Hoatzin to Auks. Handbook of the birds of the world. Lynx
Edicions, Barcelona, pp. 444-533.
Pimentel, D., McNair,S., Janecka, J., Wrighman, J., Simmonds, C., O’Connell,
C., Wong, E., Russle, L., Zern, J., Aquino, T., Tsomondo, T., 2001.
Economic and environemtnal threats of alien plants, animal, and microbe
invasions. Agriculture, Ecosystems and Environment 84, 1-20.
Pointier, J.P., David, P., 2004. Biological control of Biomphlaria glabrata, the
intermediate host of schistosomes, by Marisa cornuarietis in ponds of
Guadeloupe: long term impact on the local snail fauna and aquatic flora.
Biological Control 29, 81-89.
Pointier, J.P., Jourdane, J., 2000. Biological control of the snail host of
schistosomiasis in areas of low transmission: the example of the
Caribbean area. Acta Tropica 77, 53-60.

121
Pointier, J.P., Théron, A., Imbert-Establet, D., Borel, G., 1991. Eradication of a
sylvatic focus of Schistoma mansoni using biological control by
competitor snails. Biological Control 1, 244-247.
Porte, D., Verzola, R., Uphoff, N., 2006. Using golden apple snails as weed
control. In: Joshi, R.C., Sebastian, L.S., Eds.), Global advances in
ecology and management of golden apple snails. Philippine Rice
Research Institute, pp.505-513.
Qiu, S.L., Liu, S.P., Huang, M.G., Chen, D.Q., Duan, X.B., 2002. Monitoring of
spawning sites of four major Chinese carps in the middle section of
Yangtze River. Acta Hydrobiological Sinica 26, 716-718.
Qiu, J.W., Chan, M.T., Kwong, K.L., Sun, J., 2011. Consumption, survival and
growth in the invasive freshwater snail Pomacea canaliculata: does food
freshness matter?. Journal of Molluscan Studies 77, 189-195.
Qiu, J.W., Kwong, K.L., 2009. Effects of macrophytes on feeding and life history
traits of the invasive apple snail Pomacea canaliculata. Freshwater
Biology 54, 1720-1730.
Rahel, F.J., 2002. Homogenization of freshwater faunas. Annual Review of
Ecology and Systematics 33, 291-315.
Ricciardi, A., Neves, R.J., Rasmussen, J.B., 1998. Impendin extinctions of North
American freshwater mussels (Unionoida) following the Zebra mussel
(Dreissena polymorpha) invasion. Journal of Animal Ecology 67, 613-
619.
Ruiz-Olmo, J., Palazon, S., 1997. The diet of the European otter (Lutra lutra L.,
1758) in Mediterranean freshwater habitats. Journal of Wildlife Research
2, 171-181.
Sala, O.E., Chapon, F.S.I., Armesto, J.J., Berlow, R., Bloomfield, J., Dirzo, R.,
Huber-Sanwald, E., Huenneke, L.F., Jackson, R.B., Kinzig, A., Leemans,
R., Lodge, D., Mooney, H.A., Oesterheld, M., Poff, N.L., Sykes, M.T.,
Walker, B.H., Walker, M., Wall, D.H., 2000. Global biodiversity
scenarios for the year 2100. Science 287, 1770-1774.
Shek, C.T., 2006. A field guide to the terrestrial mammals of Hong Kong.
Friends of the Country Parks & Cosmos Books Ltd., Hong Kong.

122
Shelton, W.L., Soliman, A., Rothbard, S., 1995. Experimental observations on
feeding biology of black carp (Mylopharyngodon piceus). The Israeli
journal of aquaculture - Bamidgeh 47, 59-67.
Shen, P.R., 1989. Chapter 1. The biology of major freshwater-cultivated fishes in
China. In: FAO, (Ed.), Integrated fish farming in China, pp. 294.
Sibbing, F.A., 1982. Pharyngeal mastication and food transport in the carp
(Cyprinus carpio L.): A cineradiographic and electromyographic study.
Journal of Morphology 172, 223-258.
Sibbing, F.A., 1988. Specialization and limitations in the utilization of food
resources by the carp, Cyprinus carpio: a study of oral food processing.
Environmental Biology of Fishes 22, 161-178.
Sibbing, F.A., 1991. Food capture and oral processing. In: Winfield, I.J., Nelson,
J.S., (Eds.), Cyprinid fishes,Fish and Fisheries Series. Springer
Netherlands, pp. 377-412.
Sibbing, F.A., Osse, J.W.M., Terlouw, A., 1986. Food handling in the carp
(Cyprinus carpio): its movement patterns, mechanisms and limitations.
Journal of Zoology 210, 161-203.
Simberloff, D. 2009. The role of propagule pressure in biological invasions.
Annual Review of Ecology, Evolution and Systematics 40, 81-102.
Simberloff, D., Stiling, P., 1996. How risky is biological control? Ecology 77,
1965-1974.
Simberloff, D., Souza, L., Nunẽz, M.A., Barrios-Garcia, N., Bunn, W., 2012. The
natives are restless, but not often and mostly when disturbed. Ecology 93,
598-607.
Sin, A.W., Cheng, K.W.J., 1976. Management systems of inland fish culture in
Hong Kong. In: Dunn, I.G., (Ed.), Proceedings of the Seventeenth Indo-
Pacific Fisheries Council Fisheries Symposium. FAO Regional Office for
Asia and the Far East Bangkok, Thailand, Colombo, Sri Lanka, pp. 390-
395.
Slootweg, R., Vroeg, P.A., Wiersma, S.J., 1993. Effects of molluscivorous fish,
water quality and pond management on the development of schistomiasis

123
vector snails in aquaculture ponds. Aquaculture and Fisheries
Management 24, 123-128.
Slootweg, R., 1994. The biological control of snail intermediate hosts of
schistosomiasis by fish. Reviews in Fish Biology and Fisheries 4, 67-90.
Southeast ecological science center, U.S.G.S., 2005. Foreign nonindigenous
carps and minnows (Cyprinidae) in the United States - a guide to their
Identification, distribution, and biology. Scientific investigation report
2005-5041. Available: http://fl.biology.usgs.gov/Carp_ID/index.html.
Sridhara, S., Srihari, K., 1983. Food preference studies of the larger bandicoot rat,
Bandicota indica (Bechstein). Proceedings: Animal Sciences 92, 43-48.
Strayer, D.L., 2010. Alien species in fresh water: ecological effects, interactions
with other stressors, and prospects for the future. Freshwater Biology 55,
152-174.
Sung, Y.H., 2012. Impacts of illegal trapping and plantation forestry on
herpetofaunal populations. Department of Ecology and Biodiversity,
Faculty of Science, University of Hong Kong, Hong Kong, pp. 211.
Sung, Y.H., Karakker, N.E., Hau, B.C.H., 2012. Terrestrial herpetofaunal
assemblages in secondary forests and exotic Lophostemon confertus
plantations in South China. Forest Ecology and Management 270, 71-77.
Tam, T.W., Leung, K.K., Kwan, B.S.P., Wu, K.K.Y., Tang, S.S.H., So, I.W.Y.,
Cheng, J.C.Y., Yuen, E.F.M., Tsang, Y.M., Hui, W.L., 2011. The
dragonflies of Hong Kong. Friends of the Country Parks & Cosmos
Books Ltd.
Tamburi, N.E., Martín, P.R., 2009. Feeding rates and food conversion
efficiencies in the apple snail Pomacea canaliculata (Caenogastropoda:
Ampullariidae). Malacologia 51, 221-232.
Tamburi, N.E., Martín, P.R., 2011. Effects of food availability on reproductive
output, offspring quality and reproductive efficiency in the apple snail
Pomacea canaliculata. Biological Invasions 13, 2351-2360.
Tang, C.T., 1985. A survey of Biomphalaria straminea (Dunker 1848)
(Planorbidae) for trematode infection, with a report on larval flukes from
other gastropoda in Hong Kong. In: Morton, B., Dudgeon, D., Eds.), The

124
Malacofauna of Hong Kong II. Hong Kong University Press, Hong Kong,
pp. 393-408.
Teo, S.S., 2001. Evaluation of different duck varieties for the control of the
golden apple snail (Pomacea canaliculata) in transplanted and direct
seeded rice. Crop Protection 20, 599-604.
Teo, S.S., 2006. Evaluation of different species of fish for biological control of
apple snail Pomacea canaliculata (Lamarck) in rice. Crop Protection 25,
1004-1012.
Thomforde, H., 2000. Black carp control grub in Fathead Minnows. Aquaculture
Magazine March/April, 23-24.
Tuen Mun and Yuen Long District Planning Office, P.D., 2002. Planning with
vision - Yuen Long. Planning Department, HKSARG.
Turner, A.M., Chislock, M.F., 2007. Dragonfly predators influence biomass and
density of pond snails. Oecologia 153, 407-415.
van der Valk, A.G., 2012. Chapter 7 Invasive Species. The biology of freshwater
wetlands. Second edition. Oxford University Press Inc., New York,
United States, pp. 173-193.
van Driesche, R.G., Carruthers, R.I., Center, T., Hoddle, M.S., Hough-Goldsteine,
J., Morin, L., Smith, L., Wagner, D.L., Blossey, B., Brancatini, V.,
Casagrande, R., Causton, C.E., Coetzee, J.A., Cuda, J., Ding, J., Fowler,
S.V., Frank, J.H., Fuester, R., Goolsby, J., Grodowitz, M., Heard, T.A.,
Hill, M.P., Hoffmann, J.H., Huber, J., Julien, M., Kairo, M.T.K., Kenis,
M., Mason, P., Medal, J., Messing, R., Miller, R., Moore, A.,
Neuenschwander, P., Newman, R., Norambuen, H., Palmera, W.A.,
Pemberton, R., Perez Panduro, A., Prattc, P.D., Rayamajhi, M., Salom, S.,
Sands, D., Schooler, S., Schwarzländer, M., Sheppard, A., Shaw, R.,
Tipping, P.W., van Klinken, R.D., 2010. Classical biological control for
the protection of natural ecosystems. Biological Control 54, 2-33.
van Rompaey, H., 2001. The crab-eating mongoose, Herpestes urva. Small
Carnivore Conservation 38, 12-17.
Venable, D.L., Gaudé, A.P.I., Klerks, P.L., 2000. Control of the trematode
Bolbophorus confusus in Channel Catfish Ictalurus punctatus ponds

125
using salinity manipulation and polyculture with Black Carp
Mylopharyngodon piceus. Journal of World Aquaculture Society 31, 158-
166.
Wang, C.L., 2010. 動物性資材-魚禽類. In: Wang, C.L., Yu, J.Z., Lu, C.T., Lin,
F.C., Shih, H.T., Eds.), Nonsynthetic resources for pest management.
Taiwan Agricultural Research Institute, Taiwan.
Wang, H., Fuller, T.K., 2001. Notes on the ecology of sympatric small carnivores
in southeastern China. Mammalian Biology 66, 251-255.
Wang, H., Fuller, T.K., 2003. Food habits of four sympatric carnivores in
southeastern China. Mammalia 67, 513-519.
Wong, L.C., 1999. Resource use by egrets and herons in Starling Inlet, Hong
Kong. M.Phil. thesis. Department of Ecology and Biodiversity.
University of Hong Kong, pp. 156.
Wong, P.K., Kwong, K.L., Qiu, J.W., 2009. Complex interactions among fish,
snails and macrophytes: implications for biological control of an invasive
snail. Biological Invasions 11, 2223-2232.
Wong, P.K., Liang, Y., Liu, N.Y., Qiu, J.W., 2010. Palatability of macrophytes
to the invasion of apple snails Pomacea canaliculata: differential effects
of multiple plant traits. Freshwater Biology 55, 2023-2031.
Wood, T.S., Anurakpongsatorn, P., Chaichana,R., Mahujchariyawong, J.,
Satapanajaru, T., 2006. Heavy predation on freshwater Bryozoans by the
Golden Apple Snail, Pomacea canaliculata Lamarck, 1822
(Ampullariidae). The Natural History Journal of Chulalongkorn
University 6, 31-36.
Woodward, G., Hildrew, A.G., 2001. Invasion of a stream food web by a new top
predator. Journal of Animal Ecology 70, 273-288.
Wu, X.W., Yang, G.R., Yue, P.Q., Huang, H.J., 1964. The economics animals of
China: freshwater fishes. Science Press, Beijing, China.
WWF, H.K., 2012. Mai Po wildlife fact sheet - Duck satellite tracking project
2008-2011. Hong Kong. Available:
http://awsassets.wwfhk.panda.org/downloads/satellite_duck_tracking_en
g.pdf (online).

126
Xia, N.H., Yu, H., Li, J., Zhao, N.X., Peng, H., 2011. 27. Panicum L.. Poaceae
(Gramineae). In: Hong Kong Herbarium, A., Sciences, S.C.B.G.C.A.o.,
Eds.), Flora of Hong Kong. Volume 4. Agriculture, Fisheries and
Conservation Department, HKSARG, Hong Kong, pp. 162-163.
Yang, J.X., Chen, Y.R., 1995. The biology and resource utilization of the fishes
of Fuxian Lake, Yunnan. Yunnan Science and Technology Press,
Kunming, China.
Yang, P.S., Chen, Y.H., Lee, W.C., Chen, Y.H., 2006. Golden apple snail
management and prevention in Taiwan. In: Joshi, R.C., Sebastian, L.S.,
Eds.), Global advances in ecology and management of golden apple
snails. Philippine Rice Research Institute, pp. 169-179.
Yin, Y., Jiang, H., Guo, D., Wang, Z.K., 2006. Golden apple snails in China. In:
Joshi, R.C., Sebastian, L.S., Eds.), Global advances in ecology and
management of golden apple snails. Nueva Ecija: Philippine Rice
Research Institute, pp. 299-314.
Yipp, M.W., 1983. The ecology of Biomphalaria straminea (Dunker, 1848)
(Gastropoda: pulmonata) introduced into Hong Kong. Ph.D thesis.
Department of Zoology, University of Hong Kong.
Yipp, M.W., Cha, M.W., Liang, X.Y., 1991. A preliminary impact assessment of
the introduction of two species of Ampullaria (Gatropoda: Ampullariidae)
into Hong Kong. Proceedings of the Tenth International Malacology
Congress. Tübingen 1989.
Yoong, K.S., 2012. Meat-eating chats: more observations on Blue Whistling
Thrushes Myophonus caeruleus in Peninsular Malaysia. BirdingASIA 17,
60-64.
Yoshie, H., Yusa, Y., 2008. Effects of predation on the exotic freshwater snail
Pomacea canaliculata (Caenogastropoda: Ampullariidae) by the
indigenous turtle Chinemys reevesii (Testudines: Geoemydidae). Applied
Entomology and Zoology 43, 475-482.
Yu, X.P., Wada, T., Li, Z.F., Lu, Z.X., Sun, L.P., Zhu, Y.H., Chen, J.M., Zhen,
X.S., Xu, H.X., 2001. Occurence of golden apple snail, Pomacea

127
canaliculata (Lamarck), in paddy fields and its management. Acta
Agriculturae Zhejiangensis 13, 247-252.
Yusa, Y., Sugiura, N., Wada, T., 2006. Predatory potential of freshwater animals
on an invasive agricultural pest, the apple snail Pomacea canaliculata
(Gastropoda: Ampullariidae) in southern Japan. Biological Invasions 8,
137-147.
Yusa, Y., Wada, T., 1999. Impact of the introduction of apple snails and their
control in Japan, Naga. Naga, The ICLARM Quarterly 22, 9-13.
Zheng, X.S., Lv, Z.X., Chen, J.M., Xu, H.X., Zhang, J.F., Chen, L.Z., Yu, X.P.,
2005. Trials of Pomacea canaliculata (Lamarck) control with stocking of
Soft-shelled Turtle (Trionyx sinensis). Journal of Zhejiang Agricultural
Sciences 1, 61-62.

128
Curriculum Vitae
Academic qualifications of the thesis author, Mr.IP Ka Lok Kelvin:
Received the degree of Bachelor of Science in Environmental
Life Science (Honors, second class division 2) from the
University of Hong Kong, August 2009.
Research experiences:
2013/April: Attended the 18th
International Conference on
Aquatic Invasive Species (ICAIS), organized by Invasive
Species Centre in Canada – Oral presentation.
2008-2009: BSc final year project at University of Hong Kong
on “Fruit and Seed Resources for birds in Long Valley, Hong
Kong” with Dr. Hau Chi Hang Billy.
2008/August: Summer Research Assistant, Division of
Ecology and Biodiversity, University of Hong Kong.
2008/June-July: Voluntary work in assisting ground survey
with Forest Wardens in Yinggeling Nature Reserve Hainan,
supervised by Dr. Chan Pui Lok, Bosco.
October2013