hum. reprod.-2011-fang-1252-8
DESCRIPTION
Hum. Reprod.-2011-Fang-1252-8TRANSCRIPT

ORIGINAL ARTICLE Reproductive genetics
The p53-HDM2 gene–genepolymorphism interaction is associatedwith the development of missedabortionYan Fang1, Beihua Kong1,*, Qifeng Yang2, Daoxin Ma3, and Xun Qu4
1Department of Obstetrics and Gynecology, Qilu Hospital, Shandong University, 107 Wenhua Xilu, Ji’nan, 250012 Shandong, China2Department of General Surgery, Qilu Hospital, Shandong University, 107 Wenhua Xilu, Ji’nan, 250012 Shandong, China 3Department ofHematology, Qilu Hospital, Shandong University, 107 Wenhua Xilu, Ji’nan, 250012 Shandong, China 4Department of Basic Medicine,Qilu Hospital, Shandong University, 107 Wenhua Xilu, Ji’nan, 250012 Shandong, China
*Correspondence address. Tel: +86-531-82169008; Fax: +86-531-86927544; E-mail: [email protected]
Submitted on September 3, 2010; resubmitted on December 22, 2010; accepted on January 12, 2011
background: The interaction between p53 and human double minute 2 (HDM2) plays an important role in apoptosis; therefore, func-tional polymorphisms in these genes might have adverse effects in early pregnancy. In this study, we investigated whether p53 codon 72 andHDM2 promoter (SNP309) polymorphisms were associated with the development of missed abortion.
methods: Women with missed abortions (n ¼ 60) and healthy controls (n ¼ 64) were included in the study. Genotyping of the p53codon 72 and HDM2 SNP309 (T . G) polymorphisms was performed by PCR with sequence-specific primers and PCR-restriction fragmentlength polymorphism analysis, respectively, using villous samples. The mRNA and protein levels for p53 and HDM2 were measured by real-time PCR and semi-quantitative immunohistochemistry, respectively.
results: For the p53 codon 72 polymorphism, no difference in genotype or allele frequencies was observed in women with missed abor-tion versus controls. However, for the HDM2 SNP309 (T . G) polymorphism, G/G genotype was associated with a higher risk of missedabortion compared with the T/T + T/G genotypes (P ¼ 0.043). Women carrying the HDM2 G/G genotype or p53 Pro/Pro genotype hadhigher HDM2 mRNA (P ¼ 0.04 and P ¼ 0.013, respectively) and protein (P ¼ 0.001 and P ¼ 0.037, respectively) levels than womenwith other HDM2 SNP309 and p53 codon 72 genotypes.
conclusions: The genotypes HDM2 SNP309 G/G and p53 codon 72 Pro/Pro can induce high levels of HDM2, which may be associ-ated with missed abortion.
Key words: missed abortion / p53 / human double minute 2 / polymorphism / pregnancy
IntroductionMissed abortion refers to a pregnancy in which there is a fetaldemise without outside intervention but the uterine activity isabsent to expel the products of conception before 20 weeks of ges-tation (Griebel et al., 2005). Appropriate apoptosis has been shownto be critically important for the successful development of normalpregnancy (Chatzaki et al., 2001; Jerzak et al., 2002; Savion et al.,2002; Choi et al., 2003). Spontaneous pregnancy disorders areassociated with the excessive apoptosis of trophoblasts (Halperinet al., 2000).
p53, as an important tumor suppressor gene, is called the ‘gatekeeperof cellular genome’. p53 expression has been demonstrated in first tri-mester placenta (Quenby et al., 1998). When a cell is exposed to DNA
damage, p53 can induce cell cycle arrest in G1 phase (DNA presyntheticphase) and apoptosis. Increased human double minute 2 (HDM2;human ortholog of murine double minute 2) levels were shown tolead to attenuation of the p53 DNA damage response, and HDM2 isa key negative regulator of p53 (Zauberman et al., 1995; Ries et al.,2000). Enhanced p53 expression and activity subsequently leads toinduction of HDM2, which may act as an oncogene (Fakharzadehet at., 1991; Dubs-Poterszman et al., 1995; Jones et al., 1998) or toinhibit growth (Brown et al., 1998). Specifically, enhanced HDM2levels have been shown to cause p53 degradation in surviving cellsand, consequently, attenuation of the p53-mediated DNA damageresponse (Zauberman et al., 1995; Ries et al., 2000), thereby forminga negative feedback loop (Michael et al., 2003).
& The Author 2011. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. All rights reserved.For Permissions, please email: [email protected]
Human Reproduction, Vol.26, No.5 pp. 1252–1258, 2011
Advanced Access publication on February 20, 2011 doi:10.1093/humrep/der017
by guest on Novem
ber 14, 2015http://hum
rep.oxfordjournals.org/D
ownloaded from

Single nucleotide polymorphisms (SNPs) in the p53 gene might havefunctional relevance. At least two forms of wild-type p53 protein existamong human populations; these forms are ascribed to amino acidreplacement at codon 72 of Arg (CGC) by Pro (CCC) in the transac-tivation domain of the p53 protein. The arginine and proline isoformshave different biological and biochemical effects: the Arg-p53 couldinduce apoptosis more efficiently than the Pro-p53 (Thomas et al.,1999; Dumont et al., 2003), while the Pro-p53 could induce ahigher level of G1 arrest than the Arg-p53 (Pim et al., 2004). Thep53 codon 72 polymorphism has been proposed as a candidate forincreasing the chance of miscarriage in otherwise healthy women (Pie-trowski et al., 2005). HDM2 is vital in the regulation of p53 yetsequence variations in the HDM2 promoter may result in alteredexpression of the HDM2 protein. Recently, polymorphism SNP309(a T to G change at nucleotide 309 in the first intron) was found inthe promoter of HDM2. In our previous report, we found that theHDM2 SNP309 G/G genotype was associated with a high risk ofmissed abortion (Fang et al., 2009). Cells carrying the G/G genotype,owing to an enhanced affinity for binding the stimulatory protein (Sp)1, show a heightened HDM2 expression and significant attenuation ofthe p53 pathway compared with those carrying the T/T genotype.Importantly, this HDM2 SNP309 G/G polymorphism enhanced pro-moter recognition by the transcription factor Sp1, which in turncaused elevated HDM2 expression and attenuation of thep53-mediated apoptotic response to cellular stresses, includingDNA damage (Bond et al., 2004; Hong et al., 2005). Polymorphismsin the p53 and HDM2 genes have therefore been shown to be of func-tional significance.
Based on the fact that the p53-HDM2 interaction plays an impor-tant role in apoptosis, we hypothesized that the functional polymorph-isms in p53 codon 72 and HDM2 SNP309 might be associated with arisk of missed abortion, and tested this hypothesis in this study
Materials and Methods
DiagnosisBy ultrasound examination, the first-trimester missed abortion was definedas an intact gestational sac lacking any fetal cardiac activity [6 weeks afterlast menstrual period (LMP)], intrauterine gestational sac with the largestdiameter exceeding 10 mm but devoid of yolk sac or an empty gestationalsac with a confirmed gestational age of no ,6 weeks (Griebel et al., 2005).
PatientsThis prospective observational study involved 60 pregnant women diag-nosed with first-trimester missed abortion. All the pregnancies terminatedat the first trimester ,10 weeks from the LMP. Prior to inclusion in thestudy, all subjects underwent a standard diagnostic work-up to rule outany verifiable cause of missed abortion. The women were examinedusing ultrasonography for uterine abnormalities, and blood was drawnfor testing for chromosomal abnormalities, immunologic factors (such aspositive anticardiolipin antibody and positive antinuclear antibody) andinfections, with these analyses resulting in an unexplained etiology.Among the 60 women who suffered from missed abortion, 26 had atleast two prior miscarriages, 24 had one prior miscarriage and 10 experi-enced miscarriage for the first time. None of these women had had a suc-cessful pregnancy. The control group consisted of 64 women in earlypregnancy with a healthy, viable intrauterine fetus and no prior
miscarriage. Fetal cardiac activity and gestational age were confirmed byultrasound. Written informed consent was obtained from all participatingsubjects. The study design was approved by the Ethical Committee ofShandong University.
SpecimensVillous samples (n ¼ 60, one per woman) from the missed-abortion groupwere collected by curettage or manual vacuum aspiration. Villous samples(n ¼ 64, one per woman) from the control group were obtained byvacuum aspiration from women undergoing elective abortion at 7–10weeks of gestation for social reasons. Each villous sample was dividedinto two parts: one part was stored at 280oC before genomic DNAand RNA isolation, the other part (only 30 from missed-abortion groupand 31 from control group, randomly selected) was stored in 4% formal-dehyde at room temperature overnight for immunohistochemistryanalysis.
DNA preparationDNA was extracted using a DNA isolation kit (Tiangen, Beijing, People’sRepublic of China). The DNA content and purity of each sample wereanalyzed by ultraviolet spectrophotometry (the E260/280 ratio rangingbetween 1.6 and 1.8), and a 10 ng DNA aliquot of each sample wasused for PCR amplification. DNA samples were routinely stored at2208C.
Genotyping of p53 codon 72 and HDM2SNP309PCR analysis of p53 codon 72 polymorphismAnalysis of p53 genotype at codon 72 was performed as described(Pietrowski et al., 2005). Sequence-specific primers (SSPs) for the Proallele are F: 5′-GCC AGA GGC TGC TCC CCC-3′; R: 5′-CGT GCAAGT CAC AGA CTT-3′. Primers of Arg allele are F: 5′-TCC CCC TTGCCG TCC CAA-3′; R: 5′-CTG GTG CAG GGG CCA CGC-3′. In each25-ml reaction, 10 ng genomic DNA was mixed with 1.25 U Taq PlatinumPolymerase (Tiangen), 250 mmol/l each dNTP, 25 mmol/l Tris–HCl (pH8.7), 10 mmol/l KCl, 2 mmol/l MgCl2 (Tiangen) and 10 mmol/l of eachprimer (previously described). PCR conditions for the Pro allele were asfollows: 948C for 5 min, 5 cycles of 948C for 1 min, 618C for 50 s,728C for 50 s and 35 cycles of 948C for 1 min, 528C for 50 s, 728C for50 s and a final extension of 728C for 7 min. PCR conditions for the Argallele were as follows: 948C for 5 min, 35 cycles of 948C for 1 min,648C for 45 s, 728C for 50 s and a final extension of 728C for 10 min.PCR products (10 ml) were subjected to electrophoresis on 3% agarosegels. The gels were stained with ethidium bromide and photographedusing an ultraviolet light transilluminator. The Pro product was 178 bpand the Arg product was 136 bp.
The possible outcomes were: (i) if a PCR product was obtained onlywith the Pro-specific primers, the patient was considered as Pro homozy-gote (Pro/Pro); (ii) if a PCR product was obtained only with the Arg-specificprimers, the patient was considered as Arg homozygote (Arg/Arg); (iii) ifthe sample showed amplification with both two primers, the patientwas considered as heterozygote (Arg/Pro). Several PCR products weresequenced to further validate the PCR results.
PCR-restriction fragment lengthpolymorphism analysis of HDM2 SNP309polymorphismPrimers used in this study have been described: 5′-CGCGGGAGTTCAGGGTAAAG-3′ and 5′-AGCTGGAGACAAGTCAGGACTTAAC-3′
p53/HDM2 gene polymorphisms and risk of missed abortion 1253
by guest on Novem
ber 14, 2015http://hum
rep.oxfordjournals.org/D
ownloaded from

(Ohmiya et al., 2006). The reagents were the same as for PCR-SSP above.The PCR conditions for HDM2 SNP309 were as follows: 948C for 7 min,35 cycles of 948C for 40 s, 608C for 40 s, 728C for 40 s and a final exten-sion of 728C for 7 min. The assay for the HDM2 polymorphism utilized theenzyme MSPA1I (5′ . . . CNG�CKG . . . 3′) (NEB, Beijing, People’sRepublic of China). After DNA restriction, 10 ml of digested sampleswere subjected to electrophoresis on 3% agarose gels. The gels werestained with ethidium bromide and photographed using an ultravioletlight transilluminator.
The possible outcomes were: (i) if only one DNA fragment of 237 bpwas observed, the patient was considered as T homozygous (T/T); (ii) ifonly two DNA fragments of 189 and 48 bp were observed, the patientwas considered as G homozygous (G/G); (iii) if three DNA fragments of237, 189 and 48 bp were observed, the patient was considered as hetero-zygous (T/G). Several PCR products were sequenced to further validatethe PCR results.
RNA extraction and RT reactionTotal RNA was extracted from villous samples using the Trizol reagent(Takara, Dalian, People’s Republic of China). The quantity of RNA wasassessed spectrophotometrically. The OD260/280 of the RNA samplesranged between 1.80 and 2.00. The total RNA was reverse transcribedinto cDNA using the Rever Tra Ace Kit (Takara) in a volume of 20 ml, includ-ing 4 ml of 5 × reverse transcriptase buffer, 2 ml dNTP (10 mmol/l), 1 mlRNAse inhibitor, 1 ml Rever Tra transcriptase, 1 ml Oligo (dT) 18(0.5 mg/ml) and 5 ml ample RNA in a thermal cycler (room temperature10 min, 428C for 1 h, and 708C for 10 min). The mRNA samples werestored at 2808C before analysis.
Real-time PCR analysis of p53 and HDM2mRNAThe Quantitative SYBR-Green PCR mix kit (Toyobo, Shanghai, People’sRepublic of China) was used for the quantification of p53 and HDM2mRNA expression. Relative gene expression quantification for p53 andHDM2, with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) wasan internal reference gene, was carried out using the Roche sequencedetection system (Roche, Shanghai, People’s Republic of China). Theprimers used for p53 were: forward: 5′-ACTAAGCGAGCACTGCCCAAC-3′, and reverse: 5′-CCTCATTCAGCTCTCGGAACATC-3′. Theprimers used for HDM2 were: forward: 5′-GGACAAGAACTCTCAGATGAAGATG-3′, and reverse: 5′-ATCTGTTGCAATGTGATGAAG-3′. ThePCR mixture consisted of 10 mM of each primer, 1 ml, 10 ml Mix SYBRGreen I (Toyobo) and 500 ng cDNA to a final volume of 20 ml. For nega-tive controls, we used a complete DNA amplification mix in which thetarget cDNA template was replaced by water. Cycling parameters werethe following: denaturation for one cycle at 958C for 10 s, 45 cycles (temp-erature transition of 208C/s) of 958C for 0 s, 588C for 10 s and 728C for10 s and fluorescence reading taken at 728C, and melting curve analysiswith continuous fluorescence reading. Individual p53 and HDM2 measure-ments was calculated relative to expression of GAPDH using a modifi-cation of the method described by Lehmann et al. (2001).
ImmunohistochemistryVillous samples (61: 30 from women with missed abortion and 31 fromcontrols) were fixed in 4% formaldehyde overnight at room temperature,embedded in paraffin wax and four-micron sections were cut (Zhongshan,Guangzhou, People’s Republic of China). Tissue sections were deparaffi-nized and rehydrated through graded alcohol. Endogenous peroxidaseactivity was quenched by incubation in 3% hydrogen peroxide for10 min. After three rinses with phosphate-buffered saline, sections were
blocked with normal goat serum to suppress non-specific backgroundstaining. Mouse monoclonal antibody for wild-type p53 (Zhongshan,diluted 1:50) and HDM2 antibody (Zhongshan, diluted1:50) wereapplied to the sections separately overnight at 48C. The sections wereincubated with biotinylated goat anti-rabbit immunoglobulin (Ig) G for20 min at 378C and then processed according to the SP kit (Zhongshan)protocol. In the negative control for staining, sections were incubatedwith isotype-matched non-specific antibodies (MsIgG). Nuclear stainingof cells was regarded as a positive result. The percentages of p53- andHDM2-positive cells were estimated by counting the number of immuno-reactive cells in five microscopic fields at high-power magnification (×400)in areas with the highest visually determined positive immunoreactivity.The absence of any positive cells was scored as negative, ,5% immuno-positive cells as +, 5–20% as ++, .20% as +++.
Statistical analysisChi-squared test was used to analyze the genotype distribution of p53codon 72 and HDM2 SNP309 polymorphisms between women withmissed abortion and control groups. Chi-squared test was also used toanalyze the association of p53 and HDM2 polymorphisms with the immu-nohistochemistry results for p53 and HDM2. Student’s t-test was used tocompare the p53 and HDM2 mRNA levels in villous samples. All thevalues were presented as mean+ SD. The odds ratio (OR) was usedto measure the association between the allele frequencies and risk ofmissed abortion. All P-values are two-tailed and 95% confidence intervals(CIs) were calculated. Significant difference was defined as P , 0.05.
Results
The genotypes and allele frequencies of p53codon 72 polymorphismArg/Pro (60%) was a dominant genotype, while the frequency of Pro/Pro genotype was only 20%. No significant difference in frequencywas observed between the Pro/Pro and the Arg/Arg + Arg/Pro geno-types in women with missed abortion versus controls (P ¼ 0.330;OR, 1.750; 95% CI, 0.661–4.636). For comparisons of the frequencyof Arg and Pro alleles, there was also no difference between womenwith missed abortion and controls (P ¼ 0.373; OR, 0.778; 95% CI,0.472–1.282).
The genotypes and allele frequencies ofHDM2 SNP309 polymorphismThe G/G genotype was more in women with missed abortion(28.33%) than controls (12.50%). In the comparison of frequency ofT/T + T/G and G/G genotypes, a significant difference was seenbetween women with missed abortion and controls (P ¼ 0.043;OR, 2.767; 95% CI, 1.092–7.011). A higher frequency of the Gallele was detected in women with missed abortion (48.33%) thancontrols (35.94%) (P ¼ 0.054; OR, 1.668; 95% CI, 1.003–2.773).
The mRNA levels of p53 and HDM2 invillous samplesThe relationship of the p53 codon 72 and HDM2 SNP309 polymorph-isms to the levels of p53 and HDM2 mRNA were examined in villoussamples (n ¼ 124). In the comparison of the p53 codon 72and HDM2 SNP309 polymorphisms, samples from women with thePro/Pro genotype and G/G genotype had a higher HDM2 mRNA
1254 Fang et al.
by guest on Novem
ber 14, 2015http://hum
rep.oxfordjournals.org/D
ownloaded from
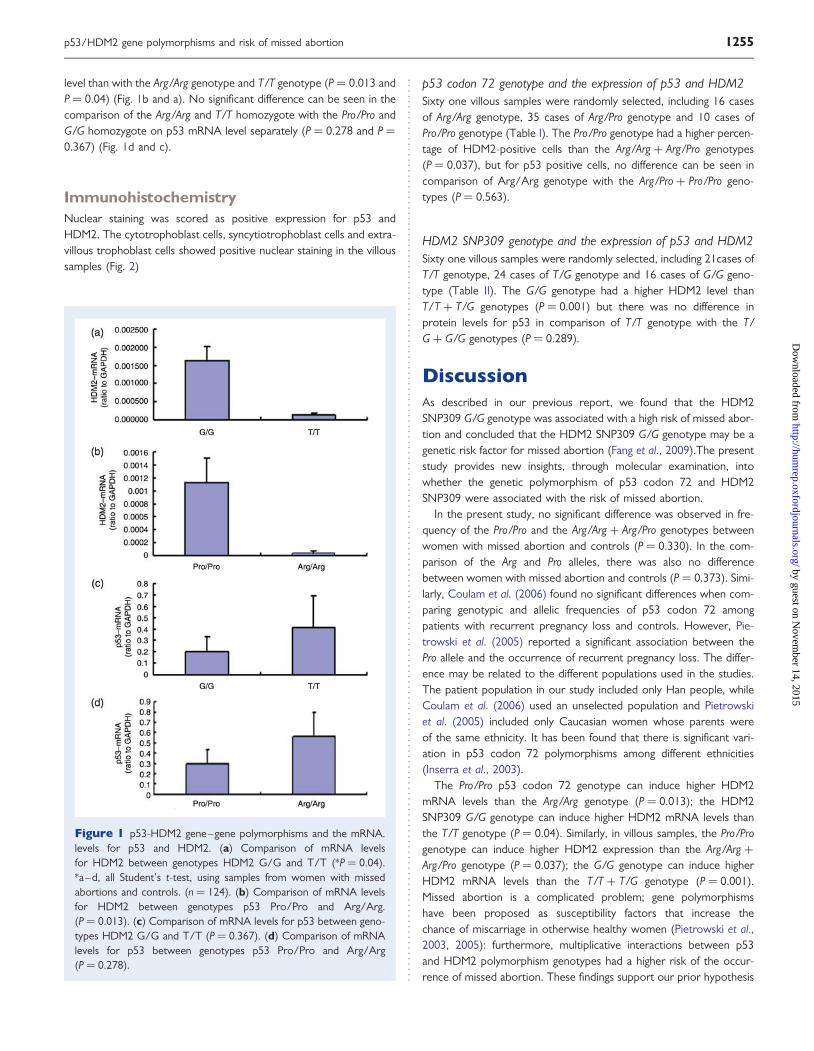
level than with the Arg/Arg genotype and T/T genotype (P ¼ 0.013 andP ¼ 0.04) (Fig. 1b and a). No significant difference can be seen in thecomparison of the Arg/Arg and T/T homozygote with the Pro/Pro andG/G homozygote on p53 mRNA level separately (P ¼ 0.278 and P ¼0.367) (Fig. 1d and c).
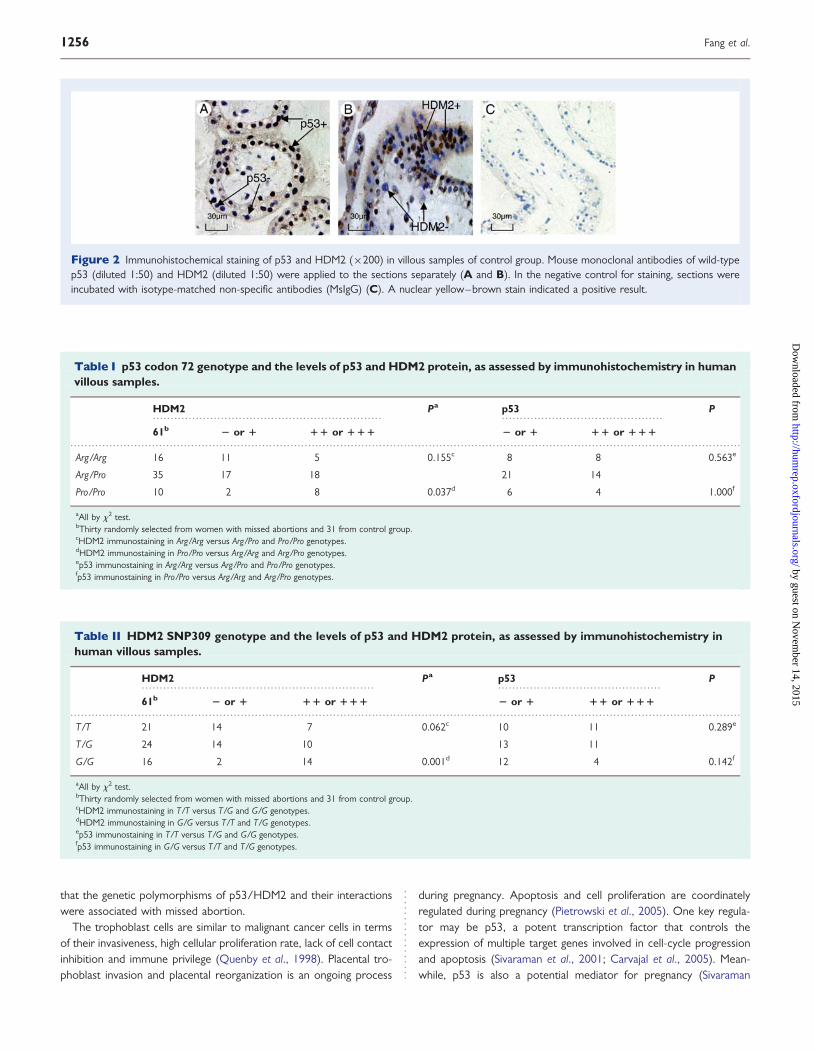
ImmunohistochemistryNuclear staining was scored as positive expression for p53 andHDM2. The cytotrophoblast cells, syncytiotrophoblast cells and extra-villous trophoblast cells showed positive nuclear staining in the villoussamples (Fig. 2)
p53 codon 72 genotype and the expression of p53 and HDM2Sixty one villous samples were randomly selected, including 16 casesof Arg/Arg genotype, 35 cases of Arg/Pro genotype and 10 cases ofPro/Pro genotype (Table I). The Pro/Pro genotype had a higher percen-tage of HDM2-positive cells than the Arg/Arg + Arg/Pro genotypes(P ¼ 0.037), but for p53 positive cells, no difference can be seen incomparison of Arg/Arg genotype with the Arg/Pro + Pro/Pro geno-types (P ¼ 0.563).
HDM2 SNP309 genotype and the expression of p53 and HDM2Sixty one villous samples were randomly selected, including 21cases ofT/T genotype, 24 cases of T/G genotype and 16 cases of G/G geno-type (Table II). The G/G genotype had a higher HDM2 level thanT/T + T/G genotypes (P ¼ 0.001) but there was no difference inprotein levels for p53 in comparison of T/T genotype with the T/G + G/G genotypes (P ¼ 0.289).
DiscussionAs described in our previous report, we found that the HDM2SNP309 G/G genotype was associated with a high risk of missed abor-tion and concluded that the HDM2 SNP309 G/G genotype may be agenetic risk factor for missed abortion (Fang et al., 2009).The presentstudy provides new insights, through molecular examination, intowhether the genetic polymorphism of p53 codon 72 and HDM2SNP309 were associated with the risk of missed abortion.
In the present study, no significant difference was observed in fre-quency of the Pro/Pro and the Arg/Arg + Arg/Pro genotypes betweenwomen with missed abortion and controls (P ¼ 0.330). In the com-parison of the Arg and Pro alleles, there was also no differencebetween women with missed abortion and controls (P ¼ 0.373). Simi-larly, Coulam et al. (2006) found no significant differences when com-paring genotypic and allelic frequencies of p53 codon 72 amongpatients with recurrent pregnancy loss and controls. However, Pie-trowski et al. (2005) reported a significant association between thePro allele and the occurrence of recurrent pregnancy loss. The differ-ence may be related to the different populations used in the studies.The patient population in our study included only Han people, whileCoulam et al. (2006) used an unselected population and Pietrowskiet al. (2005) included only Caucasian women whose parents wereof the same ethnicity. It has been found that there is significant vari-ation in p53 codon 72 polymorphisms among different ethnicities(Inserra et al., 2003).
The Pro/Pro p53 codon 72 genotype can induce higher HDM2mRNA levels than the Arg/Arg genotype (P ¼ 0.013); the HDM2SNP309 G/G genotype can induce higher HDM2 mRNA levels thanthe T/T genotype (P ¼ 0.04). Similarly, in villous samples, the Pro/Progenotype can induce higher HDM2 expression than the Arg/Arg +Arg/Pro genotype (P ¼ 0.037); the G/G genotype can induce higherHDM2 mRNA levels than the T/T + T/G genotype (P ¼ 0.001).Missed abortion is a complicated problem; gene polymorphismshave been proposed as susceptibility factors that increase thechance of miscarriage in otherwise healthy women (Pietrowski et al.,2003, 2005): furthermore, multiplicative interactions between p53and HDM2 polymorphism genotypes had a higher risk of the occur-rence of missed abortion. These findings support our prior hypothesis
Figure 1 p53-HDM2 gene–gene polymorphisms and the mRNA.levels for p53 and HDM2. (a) Comparison of mRNA levelsfor HDM2 between genotypes HDM2 G/G and T/T (*P ¼ 0.04).*a–d, all Student’s t-test, using samples from women with missedabortions and controls. (n ¼ 124). (b) Comparison of mRNA levelsfor HDM2 between genotypes p53 Pro/Pro and Arg/Arg.(P ¼ 0.013). (c) Comparison of mRNA levels for p53 between geno-types HDM2 G/G and T/T (P ¼ 0.367). (d) Comparison of mRNAlevels for p53 between genotypes p53 Pro/Pro and Arg/Arg(P ¼ 0.278).
p53/HDM2 gene polymorphisms and risk of missed abortion 1255
by guest on Novem
ber 14, 2015http://hum
rep.oxfordjournals.org/D
ownloaded from
that the genetic polymorphisms of p53/HDM2 and their interactionswere associated with missed abortion.
The trophoblast cells are similar to malignant cancer cells in termsof their invasiveness, high cellular proliferation rate, lack of cell contactinhibition and immune privilege (Quenby et al., 1998). Placental tro-phoblast invasion and placental reorganization is an ongoing process
during pregnancy. Apoptosis and cell proliferation are coordinatelyregulated during pregnancy (Pietrowski et al., 2005). One key regula-tor may be p53, a potent transcription factor that controls theexpression of multiple target genes involved in cell-cycle progressionand apoptosis (Sivaraman et al., 2001; Carvajal et al., 2005). Mean-while, p53 is also a potential mediator for pregnancy (Sivaraman
................................................................ .............................................
.............................................................................................................................................................................................
Table I p53 codon 72 genotype and the levels of p53 and HDM2 protein, as assessed by immunohistochemistry in humanvillous samples.
HDM2 Pa p53 P
61b 2 or 1 11 or 111 2 or 1 11 or 111
Arg/Arg 16 11 5 0.155c 8 8 0.563e
Arg/Pro 35 17 18 21 14
Pro/Pro 10 2 8 0.037d 6 4 1.000f
aAll by x2 test.bThirty randomly selected from women with missed abortions and 31 from control group.cHDM2 immunostaining in Arg/Arg versus Arg/Pro and Pro/Pro genotypes.dHDM2 immunostaining in Pro/Pro versus Arg/Arg and Arg/Pro genotypes.ep53 immunostaining in Arg/Arg versus Arg/Pro and Pro/Pro genotypes.fp53 immunostaining in Pro/Pro versus Arg/Arg and Arg/Pro genotypes.
................................................................. ..............................................
.............................................................................................................................................................................................
Table II HDM2 SNP309 genotype and the levels of p53 and HDM2 protein, as assessed by immunohistochemistry inhuman villous samples.
HDM2 Pa p53 P
61b 2 or 1 11 or 111 2 or 1 11 or 111
T/T 21 14 7 0.062c 10 11 0.289e
T/G 24 14 10 13 11
G/G 16 2 14 0.001d 12 4 0.142f
aAll by x2 test.bThirty randomly selected from women with missed abortions and 31 from control group.cHDM2 immunostaining in T/T versus T/G and G/G genotypes.dHDM2 immunostaining in G/G versus T/T and T/G genotypes.ep53 immunostaining in T/T versus T/G and G/G genotypes.fp53 immunostaining in G/G versus T/T and T/G genotypes.
Figure 2 Immunohistochemical staining of p53 and HDM2 (×200) in villous samples of control group. Mouse monoclonal antibodies of wild-typep53 (diluted 1:50) and HDM2 (diluted 1:50) were applied to the sections separately (A and B). In the negative control for staining, sections wereincubated with isotype-matched non-specific antibodies (MsIgG) (C). A nuclear yellow–brown stain indicated a positive result.
1256 Fang et al.
by guest on Novem
ber 14, 2015http://hum
rep.oxfordjournals.org/D
ownloaded from

et al., 2001). Investigations have shown that a large amount of p53protein is produced by the human placenta in abnormal pregnancies;thus, p53 is suspected as an important factor for the pathogenesis ofplacental disorders through the induction of the trophoblastic apopto-sis (Fulop et al., 1998; Qiao et al., 1998; Levy et al., 2002; Hu et al.,2006). The major regulator of p53 activity is the HDM2 protein,which also feeds back to promote p53 degradation (Alarcon-Vargaset al., 2002; Michael et al., 2002). Elevated HDM2 protein binds top53 to inactivate its transcriptional activity and facilitate the destabili-zation of p53. This interplay of p53 and HDM2 has been characterizedas the p53-HDM2 autoregulatory feedback loop (Wu et al., 1993).Post-translational modifications of HDM2 play a dynamic role in regu-lating the ability of HDM2 to destabilize p53 (Khosravi et al., 1999;Mayo et al., 2001; Goldberg et al., 2002).
At least two forms of wild-type p53 protein exist among majorhuman population groups, such as Caucasian and Han Chinese, anda number of studies have revealed the functional differences of thetwo polymorphisms (Hong et al., 2005; Pietrowski et al., 2005). ThePro/Pro genotype is believed to induce higher levels of G1 arrestthan the Arg/Arg genotype. The Arg/Arg genotype seems to induceapoptosis with faster kinetics and to suppress transformation moreefficiently than the Pro/Pro genotype (Thomas et al., 1999; Pimet al., 2004). In our study, the homozygous Pro/Pro genotype wasfound more often among women with missed abortion comparedwith controls. Allele frequencies may differ slightly when analyzingspecific ethnicities, and there were significant variants of p53 codon72 polymorphisms among different ethnicities (Inserra et al., 2003).
For the HDM2 polymorphism, Bond et al. (2004) showed that aftertreatment with etoposide to induce DNA damage, which activates thep53 pathway leading to DNA repair, cell cycle arrest and apoptosis,significant death was observed in cells with the HDM2 T/T genotypebut not in cells with the HDM2 G/G genotype. In our study, wefound that the G/G genotype can induce higher mRNA and proteinlevels of HDM2 (P ¼ 0.04 and P ¼ 0.001, respectively).
The maintenance of placenta is essential for the exchange betweenthe fetus and its mother and apoptosis and cell proliferation are majorprocesses contributing to a successful pregnancy. One explanation forthe observed increase of missed abortion for p53 Pro and HDM2 G/Gcarriers might be their higher potential to resist apoptosis. p53 is acti-vated by stressful stimuli, including DNA damage and hypoxia. Wesuggest that p53 Pro carriers might be at a disadvantage, with alater onset of response to stress or a lower level of the required phys-iological apoptosis. The lower level of apoptosis might lead to mis-guided growth of cells and high expression of HDM2. The HDM2G/G carriers might more easily respond to stress, leading to thehigh expression of HDM2. Specifically, enhanced HDM2 levelshave been shown to cause p53 degradation in surviving cells andconsequently attenuate the p53-mediated DNA damage response(Ries et al., 2000; Zauberman et al., 1995), which forms a negativefeedback loop.
Researchers found that both p53 codon 72 polymorphism andHDM2 SNP309 were associated with the age at onset of oralcarcinoma (Yoon et al., 2008), early development of hepatocellularcarcinomar in patients with chronic hepatitis B virus infection (Hanet al., 2008), predicting the clinical outcome of advanced non-smallcell lung cancer and influencing the clinical outcome of transitionalcell carcinoma of the bladder (Horikawa et al., 2008).This
epidemiologic study investigated whether the p53 and HDM2 poly-morphisms were associated with a risk of missed abortion. The inves-tigated polymorphisms of p53 and HDM2 have been shown to be offunctional significance. In our study, we observed that the woman whocarry the G/G or Pro/Pro genotype had significantly higher expressionlevels of HDM2. Missed abortion may be related to trophoblast cellsundergoing severe DNA damage. The high expression of HDM2 canattenuate the apoptosis reaction following DNA damage, possiblythen resulting in the death of the embryo. This study supports ourprior hypothesis that polymorphism variations in p53 and HDM2might have a relationship with missed abortion.
In conclusion, our findings support the hypothesis that the poly-morphisms of p53 codon 72 and HDM2 SNP309 and their multiplica-tive gene–gene interaction are associated with an increased risk ofmissed abortion. Further study will be necessary to determine themechanisms of missed abortion.
AcknowledgementsThe authors thank Qingshen Li and Qiuling Zhu from the Maternal andChild Health hospital of Ji’nan City for help with collecting the bloodand villous samples from the pregnancy women. The authors thankProf. Shukang Wang from the School of Public Health, Shandong Uni-versity for help with statistical analysis.
FundingThis work was supported partly by National Natural Science Foun-dation of China (No. 30872738).
ReferencesAlarcon-Vargas D, Ronai Z. p53-MDM2—the affair that never ends.
Carcinogenesis 2002;23:541–547.Bond GL, Hu W, Bond EE, Robins H, Lutzker SG, Arva NC, Bargonetti J,
Bartel G, Taubert H, Wuerl P et al. A single nucleotide polymorphism inthe MDM2 promoter attenuates the p53 tumor suppressor pathwayand accelerates tumor formation in humans. Cell 2004;119:591–602.
Brown DR, Thomas CA, Deb SP. The human oncoprotein MDM2 arreststhe cell cycle: elimination of its cell-cycle-inhibitory function inducestumorigenesis. EMBO J 1998;17:2513–2525.
Carvajal D, Tovar C, Yang H, Vu BT, Heimbrook DC, Vassilev LT.Activation of p53 by MDM2 antagonists can protect proliferating cellsfrom mitotic inhibitors. Cancer Res 2005;65:1918–1924.
Chatzaki E, Makrigiannakis A, Margioris AN, Kouimtzoglou E, Gravanis A.The Fas/FasL apoptosis pathway is involved in kappa-opioid-inducedapoptosis of human endometrial stromal cells. Mol Hum Reprod 2001;7:867–874.
Choi HK, Choi BC, Lee SH, Kin JW, Cha KY, Baek KH. Expression ofangiogenesis- and apoptosis-related genes in chorionic villi derivedfrom recurrent pregnancy loss patients. Mol Reprod Dev 2003;1:24–31.
Coulam CB, Kay C, Jeyendran RS. Role of p53 codon 72 polymorphism inrecurrent pregnancy loss. Reprod Biomed Online 2006;12:378–382.
Dubs-Poterszman MC, Tocque B, Wasylyk B. MDM2 transformation inthe absence of p53 and abrogation of the p107 G1 cell-cycle arrest.Oncogene 1995;11:2445–2449.
Dumont P, Leu JI, Della Pietra ACr, George DL, Murphy M. The codon 72polymorphic variants of p53 have markedly different apoptotic potential.Nat Genet 2003;33:357–365.
p53/HDM2 gene polymorphisms and risk of missed abortion 1257
by guest on Novem
ber 14, 2015http://hum
rep.oxfordjournals.org/D
ownloaded from

Fakharzadeh SS, Trusko SP, George DL. Tumorigenic potential associatedwith enhanced expression of a gene that is amplified in a mouse tumorcell line. EMBO J 1991;10:1565–1569.
Fang Y, Kong B, Yang Q, Ma D, Qu X. MDM2 309 polymorphism isassociated with missed abortion. Hum Reprod 2009;6:1346–1349.
Fulop V, Mok SC, Genest DR, Szigetvari I, Cseh I, Berkowitz RS. c-myc,c-erbB-2, c-fms and bcl-2 oncoproteins: expression in normalplacenta, partial and complete mole, and choriocarcinoma. J ReprodMed 1998;43:101–110.
Goldberg Z, Vogt Sionov R, Berger M, Zwang Y, Perets R, Van Etten RA.Tyrosine phosphorylation of MDM2 by c-Abl : implications for p53regulation. EMBOJ 2002;21:3715–3727.
Griebel CP, Halvorsen J, Golemon T, Day AA. Management ofspontaneous abortion. Am Fam Physician 2005;72:1243–1250.
Halperin R, Peller S, Rotschild M, Bukvsky I, Schneider D. Placentalapoptosis in normal and abnormal pregnancies. Gynecol Obstet Invest2000;50:84–87.
Han JY, Lee GK, Jang DH, Lee SY, Lee JS. Association of p53 codon 72polymorphism and MDM2 SNP309 with clinical outcome of advancednonsmall cell lung cancer. Cancer 2008;113:799–807.
Hong Y, Miao X, Zhang X, Ding F, Luo A, Guo Y, Tan W, Liu Z, Lin D.The role of p53 and MDM2 polymorphisms in the risk of esophagelsquamous cell carcinoma. Cancer Res 2005;65:9582–9587.
Horikawa Y, Nadaoka J, Saito M, Kumazawa T, Inoue T, Yuasa T.Clinical implications of the MDM2 SNP309 and p53 Arg72Pro poly-morphisms in transitional cell carcinoma of the bladder. Oncol Rep2008;20:49–55.Hu C, Smith SD, Pang L, Sadovsky Y, Nelson DM. Enhanced basal
apoptosis in cultured term human cytotrophoblasts is associated witha higher expression and physical interaction of p53 and Bak. Placenta2006;27:978–983.
Inserra P, Abrahamsen M, Papenfuss M, Giuliano AR. Ethnic variation ofthe p53 codon 72 polymorphism, HPV persistence, and cervicalcancer risk. Int J STD AIDS 2003;14:800–804.
Jerzak M, Bischof P. Apoptosis in the first trimester human placenta: therole in maintaining immune privilege at the maternal– foetal interfaceand in the trophoblast remodeling. Eur J Obstet Gynecol Reprod Biol2002;100:138–142.
Jones SN, Hancock AR, Vogel H, Donehower LA, Bradley A.Overexpression of MDM2 in mice reveals a p53-independent role forMDM2 in tumorigenesis. Proc Natl Acad Sci USA 1998;95:15608–15612.
Khosravi R, Maya R, Gottlieb T, Oren M, Shiloh Y, Shkedy D. RapidATM-dependent phosphorylation of MDM2 precedes p53accumulation in response to DNA damage. Proc Natl Acad Sci USA1999;96:14973–14977.
Lehmann U, Kreipe H. Real-time PCR analysis of DNA and RNAextracted from formalin-fixed and paraffin-embedded biopsies.Methods 2001;25:409–418.
Levy R, Smith SD, Yusuf K, Huettner PC, Kraus FT, Sadovsky Y.Trophoblast apoptosis from pregnancies complicated by fetal growth
restriction is associated with enhanced p53 expression. Am J ObstetGynecol 2002;186:1056–1061.
Mayo LD, Donner DB. A phosphatidylinositol 3-kinase/Akt pathwaypromotes translocation of MDM2 from the cytoplasm to the nucleus.Proc Natl Acad Sci USA 2001;98:11598–11603.
Michael D, Oren M. The p53 and MDM2 families in cancer. Curr OpinGenet Dev 2002;12:53–59.
Michael D, Oren M. The p53-MDM2 module and the ubiquitin system.Semin Cancer Biol 2003;13:49–58.
Ohmiya N, Taguchi A, Mabuchi N, Itoh A, Hirooka Y, Niwa Y, Goto H.MDM2 promoter polymorphism is associated with both an increasedsusceptibility to gastric carcinoma and poor prognosis. J Clin Oncol2006;24:4434–4440.
Pietrowski D, Tempfer C, Bettendorf H, Burkle B, Nagele F, Unfried G,Keck C. Angiopoietin-2 polymorphism in women with idiopathicrecurrent miscarriage. Fertil Steril 2003;4:1026–1029.
Pietrowski D, Bettendorf H, Riener EK, Keck C, Hefler LA, Huber JC,Tempfer C. Recurrent pregnancy failure is associated with apolymorphism in the p53 tumor suppressor gene. Hum Reprod 2005;4:848–851.
Pim D, Banks L. p53 polymorphic variants at codon 72 exert differenteffects on cell cycle progression. Int J Cancer 2004;108:196–199.
Qiao S, Nagasaka T, Harada T, Nakashima N. p53, Bax and Bcl-2expression, and apoptosis in gestational trophoblast of completehydatidiform mole. Placenta 1998;19:361–369.
Quenby S, Brazeau C, Drakeley A, Lewis-Jones DI, Vince G. Oncogeneand tumour suppressor gene products during trophoblastdifferentiation in the first trimester. Mol Hum Reprod 1998;4:477–481.
Ries S, Biederer C, Woods D, Shifman O, Shirasawa S, Sasazuki T,McMahon M, Oren M, McCormick F. Opposing effects of Ras onp53: transcriptional activation of MDM2 and induction of p19ARF. Cell2000;103:321–330.
Savion S, Lepsky E, Orenstein H, Carp H, Shepshelovich J, Torchinsky A,Fein A, Toder V. Apoptosis in the uterus of mice with pregnancy loss.Am J Reprod Immunol 2002;47:118–127.
Sivaraman L, Conneely OM, Medina D, BW OM. p53 is a potentialmediator of pregnancy and hormone-induced resistance to mammarycarcinogenesis. Proc Natl Acad Sci USA 2001;98:12379–12384.
Thomas M, Kalita A, Labrecque S, Pim D, Banks L, Matlashewski G. Twopolymorphic variants of wild-type p53 differ biochemically andbiologically. Mol Cell Biol 1999;19:1092–1100.
Wu X, Bayle JH, Olson D, Levine AJ. The p53-MDM2-2 autoregulatoryfeedback loop. Genes Dev 1993;7:1126–1132.
Yoon YJ, Chang HY, Ahn SH, Kin JK, Kang DR, Park JY. MDM2 and p53polymorphisms are associated with the development of hepatocellularcarcinoma in patients with chronic hepatitis B virus infection.Carcinogenesis 2008;29:1192–1196.
Zauberman A, Flusberg D, Haupt Y, Barak Y, Oren M. A functionalp53-responsive intronic promoter is contained within the humanMDM2 gene. Nucleic Acids Res 1995;23:2584–2592.
1258 Fang et al.
by guest on Novem
ber 14, 2015http://hum
rep.oxfordjournals.org/D
ownloaded from