human brca1 gene rescues the embryonic lethality of brca1 mutant mice
TRANSCRIPT

LETTER
Human BRCA1 Gene Rescues the Embryonic Lethality ofBrca1 Mutant MiceJennifer Chandler,1 Peter Hohenstein,2 Deborah A. Swing,1 Lino Tessarollo,1 andShyam K. Sharan1*1Mouse Cancer Genetics Program, National Cancer Institute, Frederick Cancer Research and Development Center,Frederick, Maryland2Medical Genetics Center, Department of Human and Clinical Genetics and Pathophysiology of Growth andDifferentiation, Department of Pathology, Leiden University Medical Center, Leiden, The Netherlands
Received 3 November 2000; Accepted 17 November 2000Published online 00 Month 2001
Summary: Half of all familial breast cancers are due tomutation in the BRCA1 gene. However, despite its im-portance, attempts to model BRCA1-induced disease inthe mouse have been disappointing. HeterozygousBrca1 knockout mice do not develop mammary tumorsand homozygous knockout mice die during embryogen-esis from ill-defined causes. Sequence analysis hasshown that the coding region, genomic organization,and regulatory sequences of the human and mousegenes are not well conserved. This has raised the ques-tion of whether the mouse can serve as an effectivemodel for functional analysis of the human BRCA1 gene.To address this question we have introduced a bacterialartificial chromosome containing the human BRCA1gene into the germline of Brca1 knockout mice. Surpris-ingly, we have found that the embryonic lethality ofBrca1 knockout mice is rescued by the human trans-gene. We also show that expression of human BRCA1transgene mirrors the endogenous murine gene. Our“humanized” transgenic mice can serve as a model sys-tem for functional analyses of the human BRCA1 gene.genesis 29:72–77, 2001. Published 2001 Wiley-Liss, Inc.†
Key words: BRCA1; humanized mice; bacterial artificialchromosome; transgenic mice; functional complementation
INTRODUCTION
Human breast cancer susceptibility gene BRCA1 ac-counts for almost 45% of all familial breast cancer cases(Easton et al., 1995; Miki et al., 1994). It has beenestimated that a BRCA1 carrier has over 70% risk ofdeveloping breast cancer and 63% risk of developingovarian cancer by the age of 70 (Ford et al., 1998). Tounderstand the biological functions and to create amouse model for this cancer susceptibility gene, severalstrains of Brca1 mutant mice have been generated (re-viewed by Hakem et al., 1998). With the exception ofmammary tumors observed in few Brca1 tissue-specificknockout mice, none of the heterozygous mutant micehave shown susceptibility to mammary tumor (Xu et al.,
1999). The amino acid sequences of mouse and humanBRCA1 proteins are 59% identical, which is low com-pared to most other tumor suppressor genes (Sharan etal., 1995). Sequence analysis also demonstrates differ-ences in the genomic organization and immediate cis-control elements of the human and mouse Brca1 genes(Xu et al., 1997). With the exception of a SP1 bindingsite and a CCAAT box, none of the human cis-controlelements, including the estrogen response element andthe AP1 site, are present in the murine gene (Xu et al.,1997). Could lack of regulatory sequence conservationbetween the murine and human genes result in differen-tial expression patterns? During the course of evolution,could the human and murine genes have divergedenough such that their functions have also changed? Wehave addressed these questions using transgenic micethat express the human gene under the control of itsendogenous promoter. We show that the human gene isfunctional in mice and complements an embryonic lethalmutation in the Brca1 gene. Our findings demonstratethat despite poor sequence conservation, the two pro-teins have similar functions. Thus transgenic mice carry-ing the human BRCA1 gene can serve as a mouse modelsfor functional studies of the human gene.
Rescue of Embryonic Lethality of Brca1 MutantMice by Murine Brca1 BAC
The regulatory elements required for spatial and tempo-ral regulation of the human and murine Brca1 geneshave not been characterized and the presence of remotecontrol elements cannot be ruled out. Therefore, priorto testing the function of the human BRCA1 gene inmice, we first examined the ability of a murine Brca1containing BAC to rescue the embryonic lethality of
* Correspondence to: Dr. Shyam Sharan, NCI, P.O. Box B, Frederick, MD21702. E-mail: [email protected]
Grant sponsor: National Cancer Institute, DHHS.†This article is a US government work and, as such, is in the public
domain in the United States of America.
Published 2001 Wiley-Liss, Inc. genesis 29:72–77 (2001)

Brca1 mutant mice. We used one of the BAC clonesdesignated as Mb1-493 with a 190-kb insert to generatetransgenic mice. This clone was obtained from a mousegenomic library and had large upstream as well as down-stream region based on relative end mapping of variousmurine Brca1 containing BACs (Fig. 1a). We crossedthree of the founder lines to mice that were heterozy-gous for Brca1 mutation. This mutation, Brca11700T, wasgenerated by insertion of the neomycin resistance geneinto exon 20 of Brca1 gene (Hohenstein, et al., In Press).Mice heterozygous for the mutation are viable and fertilebut the homozygotes die during the midgestation periodof embryogenesis. No viable homozygous mutant animal
has been obtained (Hohenstein et al., In Press). Whenwe crossed Brca11700T/1 transgenic mice carrying themurine Brca1 BAC 493 with Brca11700T/1 mice, weobtained viable and healthy homozygous mutant mice asshown by Southern analysis (Fig. 2a). This suggested thatall the regulatory sequences are located within the190-kb insert of Brca1 BAC.
Functional Test of the Human BRCA1 Genein Mice
Having demonstrated that the murine BAC rescues theembryonic lethality of Brca1 mutant mice, we next gen-erated transgenic lines with the human BRCA1 gene. Weused one of the human BRCA1 containing BAC clones,designated HB1-812 to generate transgenic lines (Fig.1b). The 200-kb insert has relatively large upstream anddownstream regions based on mapping of T7 and SP6ends relative to other BAC clones (Fig. 1b). We charac-terized the BAC clone HB1-812 by comparing the se-quence of the BAC ends to known genomic sequencesfrom the region (GenBank ID L78833.1). We estimatethat the BAC insert contains approximately 30-kb ofsequences located downstream of the BRCA1 gene andapproximately 80–90-kb of upstream sequence. (Fig.1c). Three of the transgenic founder lines carrying thisBAC were crossed to Brca11700T/1 mice. We examined2–3 F1 mice from each line for expression of the humangene in adult testis or ovary by Northern analysis. Wefound expression of the human gene in two of thetransgenic lines (data not shown) suggesting that thehuman gene is expressed in mice. Next we crossedBrca11700T/1 BAC transgenic mice expressing the hu-man gene with Brca11700T/1 mice and obtainedBrca11700T/Brca11700T BAC transgenic mice from eachof the two transgenic lines lines (Fig. 2b). These homozy-gous mutant mice are viable and both males and femalesare fertile and indistinguishable from their wild-type lit-termates (Fig. 2c). These are hemizygous for the transgeneand are obtained at the expected frequency (Table 1),suggesting that the human transgene is able to completelyrescue the embryonic lethality of the homozygous mutantmice. Some of these mice are over 18 months old and havenot shown any abnormality or tumor predisposition. Theembryonic lethality is therefore completely rescued by theBAC containing the human gene demonstrating that thehuman gene is functional in mice. Therefore, we concludethat the function of the human and murine Brca1 proteinsis conserved. This is in agreement with the findings of Laneet al. (2000).
Expression of the Human BRCA1 Gene inTransgenic Mice
The next question we addressed is whether the expres-sion of the human gene is spatially and temporally reg-ulated like the endogenous mouse gene. It has beensuggested that due to lack of conservation of cis-regula-tory elements, the human and murine Brca1 expressionpatterns may differ (Xu et al., 1997). We examined theexpression of the human BRCA1 gene in multiple tissues
FIG. 1. Mapping and characterization of BACs. a: relative order ofSp6 and T7 ends of BACs containing the entire Brca1 gene. “?”indicates that the SP6 and T7 ends of BAC 261 were not oriented.b: relative order of SP6 and T7 ends of BACs containing the entirehuman BRCA1 gene. c: characterization of human BAC HB1-812showing the location of the BRCA1 gene as well as the flankingregion containing NBR1, NBR2, and pseudo-BRCA1 gene. Arrowsindicate the direction of transcription and the solid box shows theregion between BRCA1 and NBR2 gene where the sequence ofhuman and murine cis-regulatory elements was compared.
73RESCUE OF Brca1 LETHALITY IN MICE
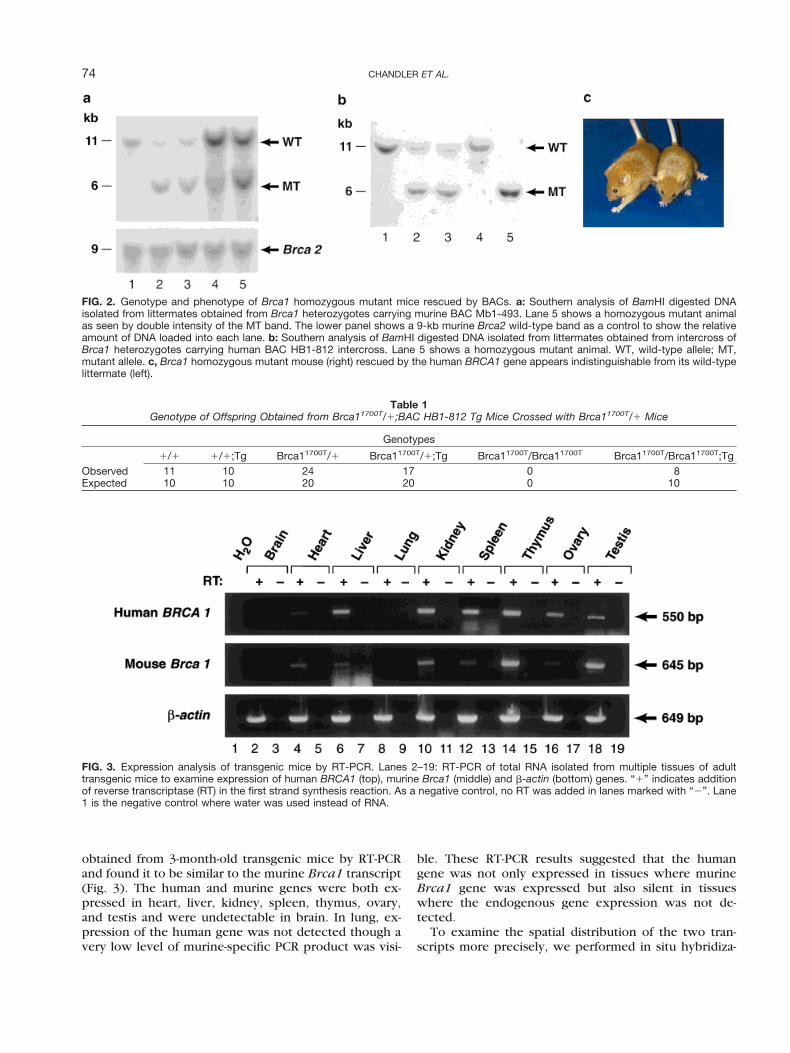
obtained from 3-month-old transgenic mice by RT-PCRand found it to be similar to the murine Brca1 transcript(Fig. 3). The human and murine genes were both ex-pressed in heart, liver, kidney, spleen, thymus, ovary,and testis and were undetectable in brain. In lung, ex-pression of the human gene was not detected though avery low level of murine-specific PCR product was visi-
ble. These RT-PCR results suggested that the humangene was not only expressed in tissues where murineBrca1 gene was expressed but also silent in tissueswhere the endogenous gene expression was not de-tected.
To examine the spatial distribution of the two tran-scripts more precisely, we performed in situ hybridiza-
FIG. 2. Genotype and phenotype of Brca1 homozygous mutant mice rescued by BACs. a: Southern analysis of BamHI digested DNAisolated from littermates obtained from Brca1 heterozygotes carrying murine BAC Mb1-493. Lane 5 shows a homozygous mutant animalas seen by double intensity of the MT band. The lower panel shows a 9-kb murine Brca2 wild-type band as a control to show the relativeamount of DNA loaded into each lane. b: Southern analysis of BamHI digested DNA isolated from littermates obtained from intercross ofBrca1 heterozygotes carrying human BAC HB1-812 intercross. Lane 5 shows a homozygous mutant animal. WT, wild-type allele; MT,mutant allele. c, Brca1 homozygous mutant mouse (right) rescued by the human BRCA1 gene appears indistinguishable from its wild-typelittermate (left).
Table 1Genotype of Offspring Obtained from Brca11700T/1;BAC HB1-812 Tg Mice Crossed with Brca11700T/1 Mice
Genotypes
1/1 1/1;Tg Brca11700T/1 Brca11700T/1;Tg Brca11700T/Brca11700T Brca11700T/Brca11700T;TgObserved 11 10 24 17 0 8Expected 10 10 20 20 0 10
FIG. 3. Expression analysis of transgenic mice by RT-PCR. Lanes 2–19: RT-PCR of total RNA isolated from multiple tissues of adulttransgenic mice to examine expression of human BRCA1 (top), murine Brca1 (middle) and b-actin (bottom) genes. “1” indicates additionof reverse transcriptase (RT) in the first strand synthesis reaction. As a negative control, no RT was added in lanes marked with “2”. Lane1 is the negative control where water was used instead of RNA.
74 CHANDLER ET AL.
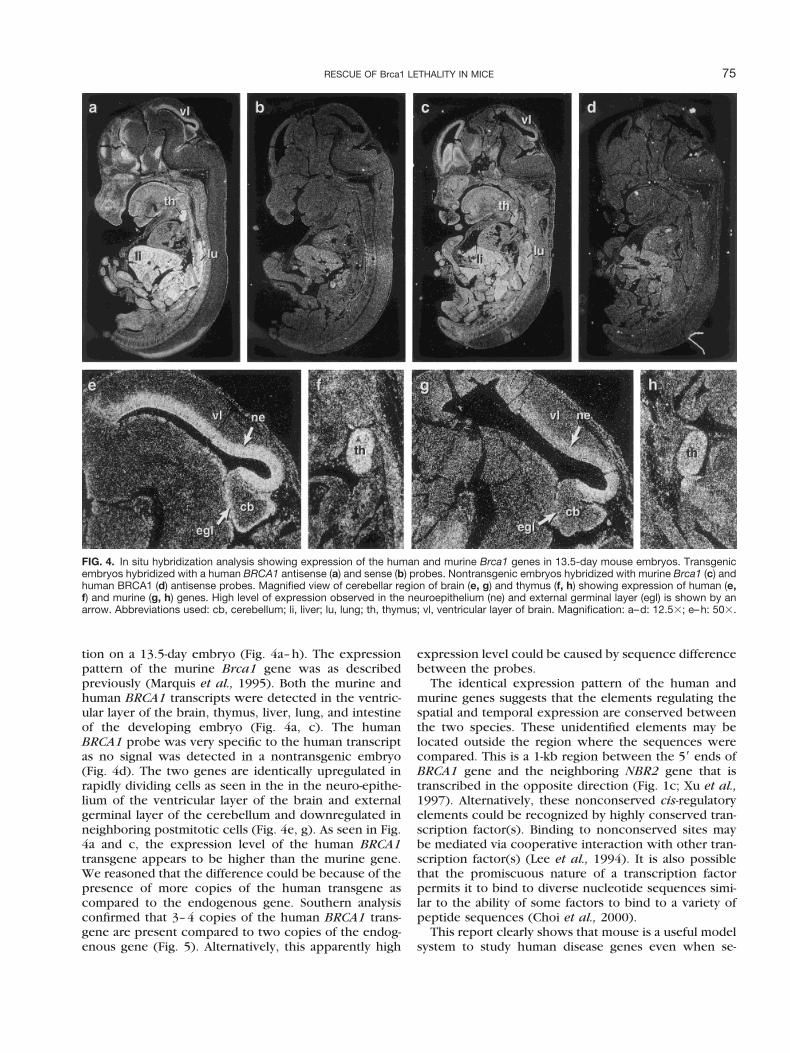
tion on a 13.5-day embryo (Fig. 4a–h). The expressionpattern of the murine Brca1 gene was as describedpreviously (Marquis et al., 1995). Both the murine andhuman BRCA1 transcripts were detected in the ventric-ular layer of the brain, thymus, liver, lung, and intestineof the developing embryo (Fig. 4a, c). The humanBRCA1 probe was very specific to the human transcriptas no signal was detected in a nontransgenic embryo(Fig. 4d). The two genes are identically upregulated inrapidly dividing cells as seen in the in the neuro-epithe-lium of the ventricular layer of the brain and externalgerminal layer of the cerebellum and downregulated inneighboring postmitotic cells (Fig. 4e, g). As seen in Fig.4a and c, the expression level of the human BRCA1transgene appears to be higher than the murine gene.We reasoned that the difference could be because of thepresence of more copies of the human transgene ascompared to the endogenous gene. Southern analysisconfirmed that 3–4 copies of the human BRCA1 trans-gene are present compared to two copies of the endog-enous gene (Fig. 5). Alternatively, this apparently high
expression level could be caused by sequence differencebetween the probes.
The identical expression pattern of the human andmurine genes suggests that the elements regulating thespatial and temporal expression are conserved betweenthe two species. These unidentified elements may belocated outside the region where the sequences werecompared. This is a 1-kb region between the 59 ends ofBRCA1 gene and the neighboring NBR2 gene that istranscribed in the opposite direction (Fig. 1c; Xu et al.,1997). Alternatively, these nonconserved cis-regulatoryelements could be recognized by highly conserved tran-scription factor(s). Binding to nonconserved sites maybe mediated via cooperative interaction with other tran-scription factor(s) (Lee et al., 1994). It is also possiblethat the promiscuous nature of a transcription factorpermits it to bind to diverse nucleotide sequences simi-lar to the ability of some factors to bind to a variety ofpeptide sequences (Choi et al., 2000).
This report clearly shows that mouse is a useful modelsystem to study human disease genes even when se-
FIG. 4. In situ hybridization analysis showing expression of the human and murine Brca1 genes in 13.5-day mouse embryos. Transgenicembryos hybridized with a human BRCA1 antisense (a) and sense (b) probes. Nontransgenic embryos hybridized with murine Brca1 (c) andhuman BRCA1 (d) antisense probes. Magnified view of cerebellar region of brain (e, g) and thymus (f, h) showing expression of human (e,f) and murine (g, h) genes. High level of expression observed in the neuroepithelium (ne) and external germinal layer (egl) is shown by anarrow. Abbreviations used: cb, cerebellum; li, liver; lu, lung; th, thymus; vl, ventricular layer of brain. Magnification: a–d: 12.53; e–h: 503.
75RESCUE OF Brca1 LETHALITY IN MICE

quence appears to be quite diverged. Hence, the biolog-ical functions of murine Brca1 gene can provide usefulinformation about human BRCA1 gene function. Failureto obtain mammary tumors in heterozygous mutant micemay be the result of the inability to lose the remainingwild-type allele in the cells to initiate the tumorigenesisprocess. This may be caused by smaller size of targettissue, reduced time because of shorter life span, orcertain unknown environmental factors but not inherentdifferences in functions of human and murine Brca1protein.
Humanized Mice as a Model System to StudyHuman Mutations
A wide range of mutations have been identified in thehuman BRCA1 gene in families with multiple cases ofbreast and ovarian cancer (Shen and Vadgama, 1999).Frameshift and nonsense result in a nonfunctional pro-tein due to premature truncation. About 25% of all themutations identified in this gene are missense mutationsthat are scattered throughout the length of the protein(Shen and Vadgama, 1999). However, the functionalsignificance of a majority of these mutations is unknown.Recently, a few functional assays have been designedthat have been used to test the significance of some ofthese missense mutations in vitro (Scully et al., 2000;Hayes et al., 2000). Our humanized transgenic miceprovide an experimentally tractable system to generatemutations identified in human breast cancer patients andanalyze how they result in tumorigenesis. Such mousemodels of human disease are necessary to evaluate ther-apeutic tools. BRCA1 has been implicated in severaldifferent biological processes including cell proliferationand recombination repair of DNA. Missense mutationsthat cause cancer can serve as valuable tools to dissectother functions and unveil the role of BRCA1 in otherbiological processes. We are currently generating trans-genic mice carrying missense mutations in the human
BRCA1 gene and we will analyze the phenotype associ-ated with these mutations in a Brca1 mutant back-ground.
METHODS
Isolation and Characterization of BACsMouse and human genomic BAC library filters obtainedfrom Roswell Park Cancer Institute were screened withmouse and human BRCA1 cDNA probes. PCR amplifiedDNA fragments from 59 (nucleotides 265–909) and 39 (nu-cleotides 4871–5452) ends of murine Brca1 cDNA(U35641) and 59 (nucleotides 281–769) and 39 (nucleotides5201–5600) ends of the human BRCA1 cDNA (U4680)were used as probes. Eleven clones were obtained usingmurine Brca1 probes and five hybridized to both 59 and 39probes. Three clones were obtained from the human li-brary and all hybridized to both end probes. Relative orderof the BAC ends was determined to identify the clone withlargest upstream and downstream regions. To identify sucha clone, T7 and SP6 ends of each BAC clone were se-quenced. The sequence information was used to designPCR primers and tested against each BAC DNA. (Sequenceof BAC ends and the primer pairs used will be providedupon request). The PCR primers from the end that wasmost distal amplified the corresponding region only fromthe BAC clone that was used to design them and noneother. Murine BAC designated Mb1-493 and human BRCA1BAC, HB1-812 were used to generate transgenic mice (Fig.1a, b). Insert size was estimated by running NotI digestedBAC DNA on pulsed field gel.
Generation of BAC Transgenic Mice
BAC DNA was extracted using standard alkaline lysismethod and then subjected to Cesium Chloride gradient onan ultracentrifuge. Supercoiled DNA was dialyzed over-night in 13 TE (10 mM Tris pH 7.6, 1 mM EDTA) buffer anddiluted to 0.5–1.0 ng/ml for pronuclear microinjection.Transgenic mice were identified by Southern analysis ofBamHI digested tail DNA using the chloramphenicol resis-tance gene present in the BAC vector as probe.
Genotyping Brca1 Mutant Miceby Southern Analysis
Brca11700H mutant mice were genotyped by hybridizingBamHI digested genomic DNA on a Southern blot with a400-bp ApaI-BamHI fragment from intron 21 of murineBrca1 gene (Hohenstein et al., In Press). The probe detectsan 11-kb wild-type and a 6-kb mutant band.
Expression Analysis by RT-PCR
Total RNA was extracted from multiple tissues of3-month-old BRCA1 BAC transgenic animals using RNA-zol B (Tel-Test, Inc.) reagent. RT-PCR was performedwith 3–5 mg of total RNA using the SuperScript Pream-plification system (Life Technologies). Primers used forgene specific PCR—Murine Brca1: forward primer 59-AAAGGGCCTTCACAATGTCC-39, reverse primer 59 TG-GCTCCACACACACATTTG, size of amplified fragment:
FIG. 5. Estimation of transgene copy number by Southern analysis.The 8-kb band corresponds to human BRCA1 transgene and the9-kb band corresponds to a two-copy endogenous Brca2 gene.Lane 1 contains DNA from a nontrangenic mouse, lanes 2 and 3contain DNA from mice that represent the two BAC HB1-812 trans-genic lines.
76 CHANDLER ET AL.

645-bp; Human BRCA1: forward primer 59-TCCAG-GAAATGCAGAAGAGG-39, reverse primer 59-ACTG-GAGCCCACTTCATTAG-39 size of amplified product:550-bp; b-actin: forward primer 59-TACCACAGGCATT-GTGATGG-39, reverse primer: 59-ATCGTACTCCTGCTT-GCTGA-39, size of amplified fragment: 649-bp product.
Expression Analysis by in situ Hybridization
The 13.5-day embryos were fixed in 4% paraformaldehydeand embedded in paraffin for sectioning. In situ hybridiza-tion was performed as described by Tessarollo and Parada(1995). Probes: PCR fragments containing regions of mu-rine Brca1 (nucleotides 265–909, U35641) and humanBRCA1 (nucleotides 2741–3290, U14680) cDNA werecloned in pGEM-T vector (Promega) and sequenced todetermine their orientation. 35S-labeled antisense humanBRCA1 probe was generated by in vitro transcription usingSP6 RNA polymerase and SacII linearized template. Thecorresponding sense probe was generated using T7 RNApolymerase and NotI linearized template. 35S-labeled anti-sense murine Brca1 probe was generated by using SP6RNA polymerase and SacII digested template DNA.
Estimation of Transgene Copy Number
Copy number of the human BRCA1 transgene was esti-mated by hybridizing a Southern blot containing 5 mg ofBamHI digested genomic DNA with a 407-bp humanBRCA1 gene-specific probe corresponding to nucleo-tides 37310–37716 (L78833) that detects an 8-kb frag-ment. As a control, a 440-bp (nucleotides 5461–5900,U65594) probe from murine Brca2 was used that de-tects a 9-kb fragment. The two probes were labeledusing T7QuickPrime Kit (Amersham Pharmacia Biotech,Inc.) and mixed in the same hybridization reaction. Theblot was exposed to a PhosphoImager (Molecular Dy-namics) screen for 16 h and subsequently analyzed toquantitate the relative intensity of the two bands.
ACKNOWLEDGMENTS
We thank Neal Copeland, Jairaj Acharya, Usha Acharya,and Mark Lewandoski for helpful discussions and criticalreview of the manuscript; Betty Martin and Elena Miroshfor technical assistance; and Richard Frederickson of thePublication Department, NCI-Frederick, for help withthe figures.
LITERATURE CITED
Choi Y, Asada S, Uesugi M. 2000. Divergent hTAFII31-binding motifshidden in activation domains. J Biol Chem 275:15912–15916.
Easton DF, Bishop DT, Ford D, Crockford GP. 1993. Genetic linkageanalysis in familial breast and ovarian cancer: results from 214
families. The Breast Cancer Linkage Consortium. Am J Hum Genet52:678–701.
Ford D, Easton DF, Stratton M, Narod S, Goldgar D, Devilee P, BishopDT, Weber B, Lenoir G, Chang-Claude J, Sobol H, Teare MD,Struewing J, Arason A, Scherneck S, Peto J, Rebbeck TR, Tonin P,Neuhausen S, Barkardottir R, Eyfjord J, Lynch H, Ponder BA,Gayther SA, Birch M, Lindblom A, Stoppa-Lyonnet D, Bignon Y,Borg A, Hamann U, Haites N, Scott RJ, Maugard CM, Vasen H, SeitzS, Cannon-Albright LA, Schofield A, Zelada-Hedman M, The BreastCancer Linkage Consortium. 1998. Genetic heterogeneity andpenetrance analysis of the BRCA1 and BRCA2 genes in breastcancer families. The Breast Cancer Linkage Consortium. Am J HumGenet 62:676–689.
Hakem R, de la Pompa JL, Mak TW. 1998. Developmental studies ofBrca1 and Brca2 knock-out mice. J Mammary Gland Biol Neoplasia3:431–445.
Hayes F, Cayanan C, Barilla D, Monteiro AN. 2000. Functional assayfor BRCA1: mutagenesis of the COOH-terminal region revealscritical residues for transcription activation. Cancer Res 60:2411–2418.
Hohenstein P, Kielman M, Breukel C, Bennett M, Wiseman R, Krimpen-fort P, Cornelisse C, Ommen G-J van, Devilee P, Fodde R. In Press.Embryonic lethality but no proliferation block in mice with a 39Brca1 mutation. Oncogene.
Lane TF, Lin C, Brown MA, Solomon E, Leder P. 2000. Gene replace-ment with the human BRCA1 locus: tissue specific expressionand rescue of embryonic lethality in mice. Oncogene 19:4085–4090.
Lee Y-H, Yano M, Liu SY, Matsunaga E, Johnson PF, Gonzalez FJA. 1994.Novel cis-acting element controlling the rat CYP2D5 gene andrequiring cooperativity between C/EBP beta and an Sp1 factor.Mol Cell Biol 14:1383–1394.
Marquis ST, Rajan JV, Wynshaw-Boris A, Xu J, Yin GY, Abel KJ, WeberBL, Chodosh LA. 1995. The developmental pattern of Brca1 ex-pression implies a role in differentiation of the breast and othertissues. Nat Genet 11:17–26.
Miki Y, Swensen J, Shattuck-Eidens D, Futreal PA, Harshman K, Tavti-gian S, Liu Q, Cochran C, Bennett LM, Ding W, Bell R, RosenthalJ, Hussey C, Tran T, McClure M, Frye C, Hattier T, Phelps R,Haugen-Strano A, Katcher H, Yakumo K, Gholami Z, Shaffer D,Stone S, Bayer S, Wray C, Bogden R, Dayanath P, Ward J, Tonin P,Narod S, Bristow PK, Norris FH, Helvering L, Morrison P, RosteckP, Lai M, Barrett JC, Lewis C, Neuhausen S, Cannon-Albrighr L,Goldgar D, Weisman R, Kamb A, Skolnick MH. 1994. A strongcandidate for the breast and ovarian cancer susceptibility geneBRCA1. Science 266:66–71.
Scully R, Ganesan S, Vlasakova K, Chen J, Socolovsky M, LivingstonDM. 2000. Genetic analysis of BRCA1 function in a defined tumorcell line. Mol Cell 4:1093–1099.
Sharan SK, Wims M, Bradley A. 1995. Murine Brca1: sequence andsignificance for human missense mutations. Hum Mol Genet4:2275–2278.
Shen D, Vadgama JV. 1999. BRCA1 and BRCA2 gene mutation analysis:visit to the Breast Cancer Information Core (BIC). Oncol Res11:63–69.
Tessarollo L, Parada LF. 1995. In situ hybridization. Methods Enzymol254:419–430.
Xu CF, Chambers JA, Solomon E. 1997. Complex regulation of theBRCA1 gene. J Biol Chem 272:20994–20997.
Xu X, Wagner KU, Larson D, Weaver Z, Li C, Ried T, Hennighausen L,Wynshaw-Boris A, Deng CX. 1999. Conditional mutation of Brca1in mammary epithelial cells results in blunted ductal morphogen-esis and tumour formation. Nat Genet 22:37–43.
77RESCUE OF Brca1 LETHALITY IN MICE