hyaluronidase 1 deficiency preserves endothelial function...
TRANSCRIPT

Sophie Dogné,1 Géraldine Rath,2 François Jouret,3 Nathalie Caron,1 Chantal Dessy,2
and Bruno Flamion1
Hyaluronidase 1 Deficiency PreservesEndothelial Function and GlycocalyxIntegrity in Early Streptozotocin-Induced DiabetesDiabetes 2016;65:2742–2753 | DOI: 10.2337/db15-1662
Hyaluronic acid (HA) is amajor component of the glycocalyxinvolved in the vascular wall and endothelial glomerularpermeability barrier. Endocytosed hyaluronidase HYAL1 isknown to degrade HA into small fragments in differentcell types, including endothelial cells. In diabetes, thesize and permeability of the glycocalyx are altered. Inaddition, patients with type 1 diabetes present increasedplasma levels of both HA and HYAL1. To investigate thepotential implication of HYAL1 in the development ofdiabetes-induced endothelium dysfunction, wemeasuredendothelial markers, endothelium-dependent vasodilation,arteriolar glycocalyx size, and glomerular barrier proper-ties in wild-type and HYAL1 knockout (KO) mice with orwithout streptozotocin (STZ)-induced diabetes. We ob-served that 4 weeks after STZ injections, the lack ofHYAL1 1) prevents diabetes-induced increases in solubleP-selectin concentrations and limits the impact of the dis-ease on endothelium-dependent hyperpolarization (EDH)–mediated vasorelaxation; 2) increases glycocalyx thick-ness and maintains glycocalyx structure and HA contentduring diabetes; and 3) prevents diabetes-induced glo-merular barrier dysfunction assessed using the urinaryalbumin-to-creatinine ratio and urinary ratio of 70- to 40-kDa dextran. Our findings suggest that HYAL1 contributesto endothelial and glycocalyx dysfunction induced by diabe-tes. HYAL1 inhibitors could be explored as a new therapeu-tic approach to prevent vascular complications in diabetes.
The glycosaminoglycan hyaluronic acid (HA), or hyaluronan,is a major component of the extracellular matrix. HA mediates
cell-cell and cell-matrix interactions and plays key roles incell migration, tumor growth and progression, inflamma-tion, and wound healing (1). HA is synthesized at theplasma membrane by different HA synthases and degradedby a family of endoglucosaminidases named hyaluronidases,mainly HYAL1 and HYAL2 in somatic tissues (2). HYAL1is the only hyaluronidase present in human and mouseplasma (3). In all cell types, its enzymatic activity occursat pH levels ,4.0, which requires the enzyme to undergoendocytosis (4).
HYAL1 deficiency in humans is a rare disease; it isassociated with bone erosions, synovitis, and polyarthritistogether with high plasma HA levels (5). A mouse modelof HYAL1 deficiency showed HA accumulation in serumwithout gross abnormalities except for a loss of proteo-glycans in knee joints (6).
In the vascular network, HA is a major component ofthe endothelial glycocalyx alongside heparan sulfate– andchondroitin sulfate–containing proteoglycans (7). In theglycocalyx, HA binds to its receptor CD44 but has no co-valent linkage and may freely exchange with the blood-stream. The glycocalyx is recognized as a major factor invascular physiology and pathology; it contributes to shearforce sensing and transduces these forces into intracellu-lar responses, such as nitric oxide (NO) release (7). Theglycocalyx also acts as a regulator of vascular permeability,a reservoir for various antithrombotic factors, and anantiadhesive barrier for leukocytes (8).
Through in vivo perfusion of hyaluronidase, whichremoves all HA in the endothelial surface layer, HA has
1Molecular Physiology Research Unit, Namur Research Institute for Life Sciences,University of Namur, Namur, Belgium2Pole of Pharmacology and Therapeutics, Institut de Recherche Expérimentale etClinique, Université Catholique de Louvain, Brussels, Belgium3Groupe Interdisciplinaire de Génoprotéomique Appliquée, Cardiovascular Sci-ences, University of Liège, Liège, Belgium
Corresponding author: Sophie Dogné, [email protected].
Received 13 December 2015 and accepted 23 May 2016.
C.D. and B.F. contributed equally to this study.
© 2016 by the American Diabetes Association. Readers may use this article aslong as the work is properly cited, the use is educational and not for profit, andthe work is not altered. More information is available at http://diabetesjournals.org/site/license.
2742 Diabetes Volume 65, September 2016
PATHOPHYSIO
LOGY

been found to be essential to maintain glycocalyx integrityand the functional barrier (9). Hyaluronidase infusion alsoabolishes the NO-dependent response to increased shearstress in segments of pig iliac artery or dog coronary ar-teries but not the acetylcholine (ACh)-induced NO produc-tion (10,11).
In patients with type 1 and type 2 diabetes, endothelialdysfunction appears to be a consistent finding underlyingthe pathophysiology of macro- and microvascular compli-cations, and therefore contributes to the increased mortalityrates observed in the population with diabetes. Glycocalyxdefects may play a central role in diabetes pathogenesis bycontributing to the proinflammatory state implicated inimpaired skin wound healing and atherosclerosis (12). In-deed, the glycocalyx itself is disturbed during both acute(13) and chronic hyperglycemia in man (14,15). In addition,patients with type 1 and type 2 diabetes have increasedplasma HA levels (15,16) and hyaluronidase activity (14,15).
To date, the implication of elevated plasma HYAL1 and/orHA levels in the pathogenesis of diabetes remains unex-plored. As plasma HYAL1 is endocytosed into endothelialcells and could therefore modulate their function possiblythrough glycocalyx regulation, we decided to investigate thepotential role of HYAL1 in the development of diabetes-induced endothelial dysfunction. To this aim, diabetes wasinduced in wild-type (WT) and HYAL1 knockout (KO) miceusing streptozotocin (STZ) injections, and endothelial-dependent vasorelaxation, circulating endothelial markers, andthe size and HA content of the glycocalyx were measured.
RESEARCH DESIGN AND METHODS
AnimalsAll experiments were performed on 7–9-week-old maleC57Bl/6 (WT) mice and B6.129 3 1-Hyal1tm1Stn/Mmcd
(Hyal12/2 or KO) mice obtained from Mutant Mouse Re-gional Resource Centers (MMRRC) backcrossed onto aC57Bl/6 genetic background for nine generations. Theanimals were fed regular chow and tap water ad libitum.All experiments were approved by the local animal ethicscommittees of the University of Namur and the Univer-sité Catholique de Louvain (2012/UCL/MD/004).
Type 1 diabetes was induced by five daily intraperito-neal injections of 55 mg/kg STZ in 10 mmol/L citratebuffer, pH 4.5. Control mice received buffer alone. Fourweeks after treatment, glycemia was measured using OneTouch Vita test strips (LifeScan Europe, Zug, Switzerland;limited to an upper value of 600 mg/dL). Animals withglycemia $300 mg/dL were assigned to the groups withdiabetes for the experiments. Mean arterial blood pres-sure was measured using a noninvasive computerized tail-cuff method (CODA; Kent Scientific, Torrington, CT) innonanesthetized mice after acclimation (17).
Chemical AssaysBlood was collected through cardiac puncture into 0.2%EDTA tubes. Soluble intercellular cell adhesion molecule-1(sICAM1), vascular cell adhesion molecule-1 (sVCAM1),
and P-selectin (sP-selectin) were quantified using ELISAkits, and HA using an ELISA-like assay that allows detectionof HA molecules $15 kDa (18), all obtained from R&DSystems (Minneapolis, MN). Syndecan-1 was measured us-ing an ELISA kit from Diaclone (Besançon, France). Albu-min and creatinine concentrations were measured in urinesamples using Albuwell (Exocell, Philadelphia, PA) andcreatinine (Enzo Life Sciences, Lausen, Switzerland) kits,respectively.
Preparation of Aortic SamplesAortas were isolated, cleaned of fat on ice, frozen in liquidnitrogen, and stored at 280°C. They were then lyophi-lized during 16 h and treated with Pronase (at 3 mg/mLin 100 mmol/L ammonia/formic acid buffer, pH 7–8) for24 h at 55°C. After relyophilization, the samples wereresuspended in water to allow HA measurement. In someexperiments, aortas were first flushed on ice with a stronginjection of cold PBS (2 mL) by holding their extremity onthe needle using pliers.
Hyaluronidase ActivityPlasma HYAL1 activity was measured using two differentapproaches: 1) zymography in renatured and native con-ditions, as described previously (4), and 2) gel electropho-resis (19) followed by quantification of oligosaccharidebands using ImageJ (public domain, NIH).
Glycocalyx Staining in Myocardial ArteriolesThe method followed a procedure previously described(20). In brief, the aorta of anesthetized mice was retro-gradely cannulated and the vena cava transected. The fol-lowing solutions were infused at a flow rate of 0.4 mL/minand a pressure of 336 5 mmHg: a cardioplegic solution for3 min, a phosphate-buffered 4% paraformaldehyde/1% glu-taraldehyde (pH 7.4) fixative solution for 2 min, and fi-nally, the same solution containing 0.05% Alcian Blue 8GX(Sigma-Aldrich) for 30 min. The left ventricular wall wascut in 2-mm segments, fixed for 1 h in the fixative solution,postfixed in 1% osmium tetroxide and 1% lanthanumnitrate for 1 h, and then processed for transmission elec-tronic microscopy (TEM) using a standard procedure. Sec-tions were visualized with a FEI Tecnai microscope andphotomicrographs analyzed using ImageJ. The glycocalyxthickness of cardiac arterioles was calculated by dividingthe surface area by the underlying endothelium length.
Dextran ExcretionAnesthetized mice were intravenously injected with amixture of 10 mg/mL Texas Red/40-kDa neutral dextranand 2.5 mg/mL FITC/70-kDa anionic dextran (MolecularProbes, Eugene, OR). Urine was collected for 30 min andfluorescence was measured to determine glomerular perm-selectivity based on the ratio of 70- to 40-kDa dextran. Theurinary albumin-to-creatinine ratio (ACR) was also measured.
Endothelium-Dependent VasodilationSecond-order mesenteric arteries were isolated fromanimals under terminal anesthesia and placed in ice-cold
diabetes.diabetesjournals.org Dogné and Associates 2743
Tyrode solution. Arteries were cleared of fat and connectivetissue and then cut into ,2-mm rings and mounted in a wiremyograph (model 610M-DMT; Danish Myo Technology A/S,Aarhus, Denmark), as previously described (21). After a 45-minstabilization in Tyrode solution containing 1025 mol/L indo-methacin, tension normalization, and 60-min equilibration,vessels were contracted using 100 mmol/L KCl. Then, cumu-lative concentrations of ACh (1028 to 3 3 1025 mol/L) wereadded to induce endothelium-dependent relaxation. Afterwashout and stabilization, vessels were again contracted using3 3 1026 mol/L phenylephrine in the absence or presence of1024 mol/L N-nitro-L-arginine methyl ester (L-NAME). Cumu-lative amounts of ACh were again added and the percentage ofresidual contraction was calculated. To test small conductancepotassium channel-3 (SK3) activity, an SK3 opener, cyclohexyl-[2-(3,5-dimethyl-pyrazol-1-yl)-6-methyl-pyrimidin-4-yl]-amine (CYPPA; Sigma-Aldrich), was used in the presenceof indomethacin and L-NAME. Vessels were precon-tracted with 1027 mol/L U46,619 (Sigma-Aldrich),a thromboxane A2 agonist, instead of phenylephrine toobtain a stable contractile state. Cumulative amounts ofCYPPA (3 3 1027 to 6 3 1025 mol/L) were then added.
mRNA ExpressionTotal RNA was isolated from microdissected mesentericarteries using the RNeasy Micro Kit (Qiagen, Hilden,Germany) and treated with DNAse. Reverse transcriptionwas performed using random hexamers and Superscript IIMMLV reverse transcriptase (Invitrogen, Carlsbad, CA).The levels of expression of several genes were determinedusing real-time PCR (7300 Real Time PCR System; AppliedBiosystems, Cheshire, U.K.) with SYBR Green detection.mRNA levels were calculated using the 2-ddCT method.
ImmunohistologyMesenteric arteries were fixed in alcoholic Bouin solu-tion for 72 h and embedded in paraffin. Detection ofSK3 and von Willebrand factor (vWF) in 6-mm sectionswas performed with an anti-SK3 rabbit polyclonal anti-body (Sc28621; Santa Cruz Biotechnology, Santa Cruz,CA) and an anti-vWF rabbit polyclonal antibody (A0082;Dako A/S, Glostrup, Denmark) followed by biotinylatedsecondary antibodies and streptavidin-peroxidase. Quan-tification of the SK3 immunostaining was performed usingImageJ.
Statistical MethodsTwo-way ANOVA followed by Bonferroni post hoc testswas performed to compare the four groups of mice in eachexperiment.
RESULTS
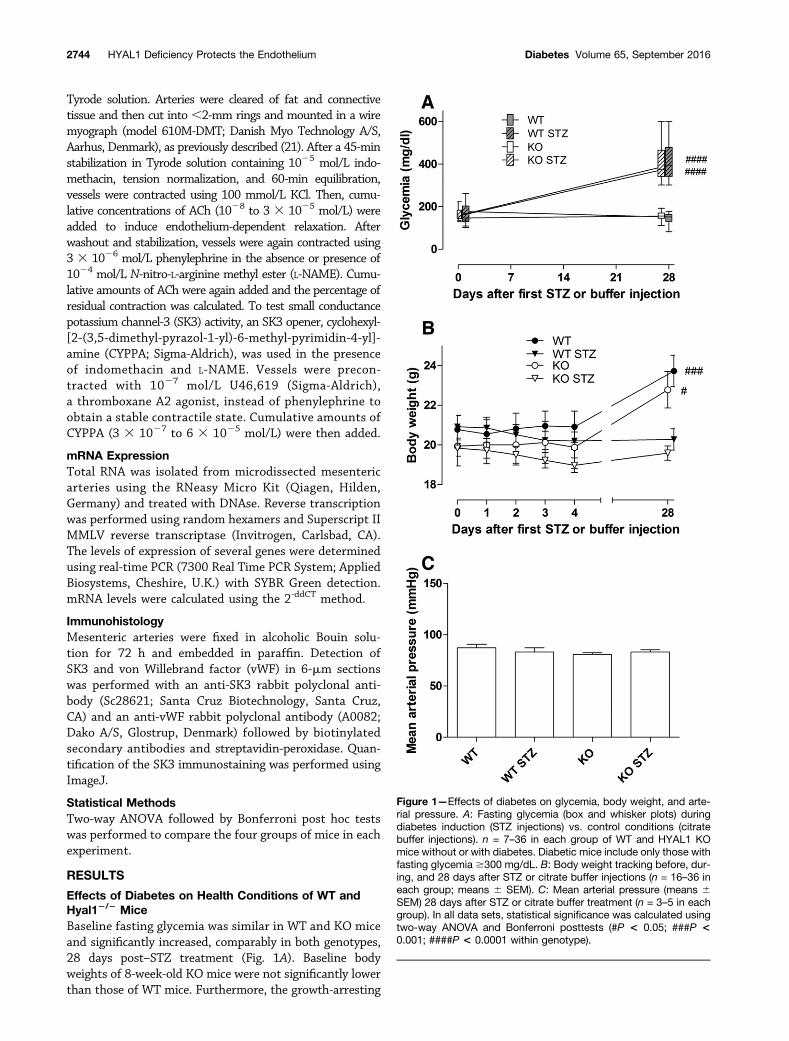
Effects of Diabetes on Health Conditions of WT andHyal12/2 MiceBaseline fasting glycemia was similar in WT and KO miceand significantly increased, comparably in both genotypes,28 days post–STZ treatment (Fig. 1A). Baseline bodyweights of 8-week-old KO mice were not significantly lowerthan those of WT mice. Furthermore, the growth-arresting
Figure 1—Effects of diabetes on glycemia, body weight, and arte-rial pressure. A: Fasting glycemia (box and whisker plots) duringdiabetes induction (STZ injections) vs. control conditions (citratebuffer injections). n = 7–36 in each group of WT and HYAL1 KOmice without or with diabetes. Diabetic mice include only those withfasting glycemia $300 mg/dL. B: Body weight tracking before, dur-ing, and 28 days after STZ or citrate buffer injections (n = 16–36 ineach group; means 6 SEM). C: Mean arterial pressure (means 6SEM) 28 days after STZ or citrate buffer treatment (n = 3–5 in eachgroup). In all data sets, statistical significance was calculated usingtwo-way ANOVA and Bonferroni posttests (#P < 0.05; ###P <0.001; ####P < 0.0001 within genotype).
2744 HYAL1 Deficiency Protects the Endothelium Diabetes Volume 65, September 2016

impact of diabetes was similar in both genotypes (Fig. 1B).There was no effect of either genotype or diabetes on meanblood pressure (Fig. 1C).
Effect of Diabetes on HA and Hyaluronidase Activityin WT and Hyal12/2 MiceA twofold increase in circulating HA levels was observedin diabetic versus nondiabetic WT mice (Fig. 2A). In KO
mice, baseline HA concentrations were four times higherthan in WT mice and did not further increase after in-duction of diabetes.
Zymography of serum samples revealed a single bandof hyaluronidase activity at ;80 kDa in both denaturingand native conditions, corresponding to the activity of theHYAL1 precursor, which did not increase after diabetesinduction (Fig. 2B and C). In aortic wall homogenates,
Figure 2—Plasma HA concentration and hyaluronidase activity. A: Plasma HA concentrations of WT and KO mice without or with STZ-induced diabetes (n = 4 or 5 in each group). Serum (B and C) and aortic wall (D) hyaluronidase activity measured using zymography undernative (B and D) or denaturing (C) conditions in each experimental group. Contrary to denaturing conditions, the native procedure does notallow the determination of the molecular weight of the HA-degrading enzyme. E–H: Measurement of HA oligosaccharide production byserum hyaluronidase activity using polyacrylamide gel electrophoresis. E: Rooster comb HA (5 mg) was incubated for 5 h with 1 mL of serumobtained from two animals of each group (lanes 1–8). The serum of KO mice (lanes 5–8) and undigested HA (lane 10) served as negativecontrols. HA incubated with 5 units of bovine testes hyaluronidase during 2 h (lane 9) served as positive control. F: A gradient of bovinetestes hyaluronidase at concentrations ranging from 0 (lane 1) to 5 units (lane 5), incubated for 2 h with the same amount of HA, was alsoperformed for comparison. G and H: Quantification of serum hyaluronidase activity. Polyacrylamide gel electrophoresis of HA samplesincubated with serum of WT mice without (lanes 1–4) and with (lanes 5–8) diabetes and used for quantification of hyaluronidase activity. Thenegative and positive controls used in G are similar to those used in E. H: Quantification of hyaluronidase activity; signal intensitymeasurements for successive and sufficiently distinct oligosaccharide bands (i.e., below the dashed line in G). Statistical analysis usestwo-way ANOVA (#P < 0.05; §P < 0.05; **P < 0.01; §§§§P < 0.0001; ****P < 0.0001).
diabetes.diabetesjournals.org Dogné and Associates 2745

zymography in native conditions showed two bands, corre-sponding to the precursor and cleaved forms of HYAL1 (4),again with no increase in hyaluronidase activity during dia-betes (Fig. 2D). Gel electrophoresis of HA solutions incubatedwith sera of nondiabetic and diabetic WTmice allowed a moreaccurate measurement of serum hyaluronidase activity andrevealed a slight but significant increase in the amount ofsmaller oligosaccharides produced using hyperglycemic sera(Fig. 2E–H), suggesting an increased HYAL1 activity (3) indiabetic WT mice. The same analysis performed on HYAL1KO mouse serum (Fig. 2E) confirmed the assay specificity.
HA on the Luminal Side of VesselsTo determine whether increased plasma HA concentrationsare accompanied by changes in glycocalyx HA content, wemeasured the amount of HA in flushed segments of theaortic wall, which almost completely removes the endothe-lium while leaving the underlying aortic wall intact, and com-pared it with that of unflushed segments. There was nodifference in the amount of HA in the unflushed aorticsegments between any of the groups (Fig. 3A). However,whereas the flushable/glycocalyx HA accounted for approx-imately one-quarter of the total aortic HA in healthy WTand KO mice, it was nearly absent in diabetic WT mice butwas preserved in diabetic KO mice.
Glycocalyx Thickness and IntegritySince HA is not the only component of glycocalyx, weexamined its structure in small vessels (ventriculararterioles) using a well-established electron microscopicmethod as described in RESEARCH DESIGN AND METHODS (Fig.3B and C). As summarized in Fig. 3D, glycocalyx thicknesswas greater than threefold higher in KO than WT mice,both in diabetic and nondiabetic mice. Diabetes did notinduce any significant change in glycocalyx thickness ineither genotype. Plasma syndecan-1 concentration (Fig.3E), measured to investigate glycocalyx shedding, wasnot impacted by either genotype or diabetes.
Glycocalyx Barrier FunctionThe relative permeability of the glomerular endothelialglycocalyx was then evaluated by measuring the concentrationratio of 70- to 40-kDa fluorescent dextran recovered in theurine after intravenous injection. As shown in Fig. 4, the ratioof 70- to 40-kDa excretion increased during diabetes in WTbut not in KO mice, suggesting that glomerular permselectiv-ity to high-molecular-weight dextran is altered in diabetic WTmice but not in diabetic KO mice. Similarly, the urinary ACRincreased in diabetic WT but not KO mice. This suggests thatthe absence of HYAL1 protects the glomerular endothelialglycocalyx against diabetes-induced functional damage.
Markers of Endothelium DysfunctionIn order to detect early signs of endothelial dysfunction,we measured the levels of circulating adhesion molecules.The level of sICAM1 was significantly upregulated duringdiabetes in both genotypes (Fig. 5B). The level of sP-selectinwas also upregulated during diabetes but only in WT mice
(Fig. 5A). The concentration of sVCAM1 was not affectedby diabetes in any genotype (Fig. 5C). The baseline plasmaconcentrations of sICAM1 and sVCAM1 were lower in KOthan WT mice, suggesting a healthier endothelial status inthe absence of HYAL1.
Endothelial-Dependent VasorelaxationEndothelial function was investigated ex vivo in small-diameter, second-branch mesenteric arteries, allowingassessment of both NO- and endothelium-dependent hy-perpolarization (EDH)–mediated relaxation. NO-dependentACh-induced vasodilation was similar in all groupsof mice (data not shown). EDH-dependent ACh-inducedvasodilation, on the other hand, was severely altered indiabetic WT mice, with only a remaining 34% EDH-dependent vasodilation, while partially preserved indiabetic KO mice, with a remaining 56% EDH-dependentvasodilation (Fig. 6). The difference between diabeticWT and KO mice was highly significant (P , 0.0001).There was no difference between healthy WT and KOmice.
Exploration of the EDH Pathway ComponentsThe mRNA expression level of several components of theEDH pathway (22) in mesenteric arteries was screenedusing real-time RT-PCR. No difference among experimen-tal groups was detected for connexins 37, 40, 43, and 45;SK channels SK1 and SK2; intermediate conductance po-tassium channel IK1; transient receptor potential (TRP)channels TRPV4 and TRPC1; and caveolin-1 (data notshown). Conversely, SK3 mRNA expression was signifi-cantly upregulated in KO mice independently of diabetes(Fig. 7A). Immunohistochemistry allowed the detection ofSK3 along the endothelium in a pattern similar to that ofvWF (Fig. 7B). Quantification of the staining confirmed in-creased expression of SK3 in KO versus WT vessels in-dependently of diabetes (Fig. 7C).
To determine whether the activity of SK3 was increasedin KO versus WT mice, mesenteric artery segments weremounted on wire myographs, contracted with U46,619,and exposed to increasing concentrations of the CYPPASK3 opener (23). SK3-dependent relaxation was more effi-cient in KO than in WT mice, whether the animals werehealthy or diabetic (Fig. 7D). SK3 overexpression may thusexplain the preservation of the EDH-dependent vasodila-tion observed in diabetic KO mice.
DISCUSSION
Increased serum hyaluronidase activity in diabetes hasbeen reported in man (15) and rats (24), but the implica-tion of these observations has been poorly studied. Thecurrent study confirms a slight increase in serum hyal-uronidase activity, which is likely due to HYAL1, in earlydiabetic WT mice (Fig. 2H). Furthermore, experimentsusing HYAL1-deficient mice at a relatively early stage oftype 1 diabetes (4 weeks after STZ injections) show, forthe first time, that HYAL1 may have a pathogenic role indiabetes-induced endothelial dysfunction.
2746 HYAL1 Deficiency Protects the Endothelium Diabetes Volume 65, September 2016

Lack of HYAL1 Prevents Endothelial DysfunctionEndothelial dysfunction in murine diabetes can be eval-uated by measuring plasma levels of sP-selectin, sICAM1,and sVCAM1, as well as ACh-dependent vasorelaxation.We showed that 4 weeks of STZ treatment in WT mice issufficient to increase sP-selectin and sICAM1 but notsVCAM1, suggesting that the latter may associate with a laterstage in diabetes development, as suggested previously (25).
Alteration in ACh-dependent vasorelaxation is anotherreliable marker of endothelial dysfunction, at least in ratmodels of type 1 and type 2 diabetes (26). In mice, loss ofendothelium-dependent relaxation appears only after 10weeks of hyperglycemia in STZ-induced diabetes (27).Concordant with these data and previous observationsin diabetic rats (28), we found no defect in the overall ACh-dependent vasorelaxation after 4 weeks of STZ injections.
Figure 3—Flushable HA in aortas of diabetic and nondiabetic WT and Hyal1 KO mice and structural evaluation of the endothelialglycocalyx. A: HA content of aortic homogenates (n = 5–7 in each group) standardized to dry weight before (white columns) and after(black columns) saline flushing. Statistical analysis was performed using two-way ANOVA and Bonferroni posttests to compare HA contentbefore vs. after flushing in each group (####P < 0.0001). Representative TEM images of myocardial arterioles of WT (B) and KO (C) micewithout or with STZ-induced diabetes after perfusion of the heart with Alcian Blue 8GX, in which the thickness of the glycocalyx can beappreciated. Bars correspond to 1 mm. D: Evaluation of glycocalyx thickness obtained by dividing the glycocalyx surface area by theendothelium length measured using ImageJ. n = 3–4 mice in each group, corresponding to ;50 TEM images of microvessels ($8 mm indiameter) in each group. Data are means 6 SEM of each experimental group. Statistical analysis was performed using two-way ANOVA(**P < 0.01). E: Plasma syndecan-1 levels in each experimental group (means 6 SEM, n = 7–10 in each group). Statistical analysis wasperformed using two-way ANOVA (ns).
diabetes.diabetesjournals.org Dogné and Associates 2747

Still, we demonstrated a clear reduction in EDH-dependentrelaxation.
Furthermore, HYAL1 deficiency may prevent the diabetes-induced increase in sP-selectin, but not in sICAM1, andlimit the impact of the disease on EDH-dependent vaso-relaxation. The data on sP-selectin and EDH-dependentvasorelaxation suggest functionally important cardiovascular-protective effects of HYAL1 deficiency during diabetes.
Interestingly, the absence of HYAL1 also modified thebaseline levels of circulating markers of endothelial dam-age, two of which (sICAM1 and sVCAM1) were lower inKO than in WT mice. This suggests that HYAL1-deficientendothelia may be less attractant to leukocytes. This hypoth-esis could be tested by measuring endothelial chemoattractionand diapedesis. Of note, HYAL1 deficiency did not prevent
Figure 4—Functional status of endothelial glycocalyx. A: Ratio of70- to 40-kDa dextran in urine 30 min after intrajugular injection of a1:4 mixture of 40- and 70-kDa dextran in WT (n = 17) and KO (n = 8)mice without or with diabetes. Statistical analysis was performedusing two-way ANOVA (*P < 0.05) and Bonferroni posttests withingenotypes (##P < 0.01). B: ACR measured in the same urine sam-ples. Statistical analysis was performed using two-way ANOVA(*P < 0.05) and Bonferroni posttests within genotypes (#P < 0.05)or across genotypes (§P < 0.05).
Figure 5—Endothelial injury markers. Evaluation of three markers ofendothelial damage in WT and KO mice before and after STZ-induced diabetes (n = 5–8 in each group). A: Serum sP-selectin levels.B: Plasma sICAM1 levels. C: Plasma sVCAM1 levels. Data aremeans 6 SEM. Statistical analysis was performed using two-wayANOVA (*P < 0.05; **P < 0.01; ****P < 0.0001) and Bonferroniposttests inside genotype (#P < 0.05; ##P < 0.01; ###P < 0.001)or across genotype (§P < 0.05).
2748 HYAL1 Deficiency Protects the Endothelium Diabetes Volume 65, September 2016

renal neutrophil and macrophage infiltration in a murinemodel of severe ischemia reperfusion injury (29), but this is acomplex acute lesion model in which multiple factors couldmodulate the phenotype.
Through a deeper analysis of several key endothelialproteins, we observed increased expression of SK3 inHYAL1-deficient endothelium. This endothelial potassiumchannel is reportedly a fundamental determinant of vasculartone and blood pressure (30) and a mainspring of the EDHpathway (31). In the absence of diabetes, however, the EDH-mediated vasodilation measured in mesenteric arteries wasnot enhanced by lack of HYAL1. This suggests baselineendothelial SK3 levels are not rate limiting for EDH-induced vasodilation in normal physiological conditionsbut could become so when the EDH pathway needs to beactivated, e.g., during early diabetes. SK3 downregula-tion was previously observed in STZ-induced diabeticApoE-deficient mice (22) or in the cavernous tissues ofdiabetic rats (32) but not in C57Bl/6 diabetic mice (27).In line with the latter results, SK3 was not downregu-lated by diabetes in our study. The long-term benefits ofthe HYAL1 deficiency–associated increase in SK3 expres-sion in diabetes remain to be demonstrated. The absenceof impact of diabetes and genotype on other EDH com-ponents at the RNA level does not preclude an effect atthe protein level. The small size of vascular samples hasprevented us from thoroughly examining this hypothesisin the current study.
Lack of HYAL1 Maintains Glycocalyx Structure andPrevents HA SheddingThe HA content of the glycocalyx is reportedly crucial forendothelial barrier function (9). A detrimental effect ofdiabetes or acute hyperglycemia on endothelial glycocalyxhas been demonstrated in several studies in man (13,15)
and mice (33). We postulated that the mechanism of en-dothelial protection in HYAL1-deficient mice is linked to astronger glycocalyx. In our study, 4-week STZ-induced di-abetes did not reduce the size of the endothelial glycocalyxin WT mice, as measured with a sensitive electron micro-scopic technique. Nevertheless, diabetes-exposed glycocalyxbecame HA depleted and thus potentially more vulnerable.HA seems to be incorporated within the glycocalyx in ashear stress–dependent way (34). It was therefore highlyrelevant to observe that HYAL1-deficient endothelial sur-faces, contrary to WT endothelia, maintained their HAcontent during diabetes (demonstrated in Fig. 3).
Furthermore, the absence of HYAL1 dramatically in-creased the thickness of the glycocalyx (in both healthyand diabetic glycocalyx). However, aortic flushes failed todemonstrate a higher HA content as the reason for theincreased size of glycocalyx. This may be due to insufficientsensitivity of the methods and/or additional factors. Onepossibility is a better anchoring of HA into the glycocalyxand a longer stretching of the HA chains in the absence ofHYAL1. This could lead to an apparent increased thicknessof the glycocalyx without HA accumulation.
Glycocalyx shedding under severe inflammatory condi-tions, such as postischemic reperfusion, can be preventedusing various treatments, e.g., hydrocortisone, antithrom-bin, or heparin (35,36). Sulodexide, a mix of heparan sul-fate and dermatan sulfate, increases glycocalyx thickness intype 2 diabetes (14). However, although sulodexide hadglobal beneficial effects on renal manifestations of experi-mental diabetes in C57Bl/6 mice (37), it failed to demon-strate renoprotection in overt type 2 diabetic nephropathyin man (38).
To our knowledge, such an increase in glycocalyx sizeand resistance as observed in our study using HYAL1 KOmice has never been described as a result of therapeuticintervention or genetic manipulation (39). A thickerglycocalyx may thus correspond to reduced access of cir-culating inflammatory cells to the endothelium (8) aswell as more efficient shear stress–induced signals. Theglycocalyx has indeed been suggested to mediate shearstress– or flow-induced NO production and vascularremodeling (40,41), although, perhaps surprisingly, itsrole in EDH-mediated vasorelaxation has not been ex-plored to date. A thicker glycocalyx could also explain thehigher baseline expression of SK3 observed in our study.
Finally, although a thicker glycocalyx correlated withbeneficial effects on diabetes-induced endothelial dys-function in the current study, this was only observed inthe absence of HYAL1. Thus, caution is warranted beforethe cardiovascular consequences of a thicker baselineglycocalyx can be described as indisputably favorable.
Lack of HYAL1 Prevents Glomerular Barrier DysfunctionDiabetes is a well-known aggressor of the glomerularglycocalyx and endothelial barrier, resulting in lossof permselectivity and ultrafiltration of albumin (42).Glycocalyx damage coincides with microalbuminuria in type 1
Figure 6—EDH-mediated vasodilation. Mesenteric arteriolar rings ofWT and KO mice without or with STZ-induced diabetes (n = 24–26rings in each group) were preconstricted with 3 3 1026 mol/L phen-ylephrine in the presence of 1024 mol/L L-NAME and 1025 mol/Lindomethacin. Vasodilation was then induced using increasing con-centrations (1028 to 33 1025 mol/L) of ACh. Data are means6 SEM.Statistical analysis, two-way ANOVA (****P < 0.0001).
diabetes.diabetesjournals.org Dogné and Associates 2749

diabetes (15). Obese diabetic db/db mice have an al-tered glycocalyx with higher access of a 70-kDa dextrantracer to the vessel wall and higher clearance of thistracer (43). In the current study, 4-week STZ-induceddiabetes significantly increased the urinary ACR andratio of 70- to 40-kDa dextran, confirming altered glomer-ular permselectivity. These effects were completely pre-vented in diabetic HYAL1 KO mice, bringing furtherarguments for a functionally preserved glycocalyx andendothelial barrier when diabetes develops in the ab-sence of HYAL1. Baseline ratios of albumin to creatinineand 70- to 40-kDa dextran were similar in WT andHYAL1-deficient mice.
Mechanisms of Endothelial Protection in the Absenceof HYAL1Table 1 summarizes the beneficial effects observed inHYAL1 KO mice exposed to STZ-induced diabetes as com-pared with WT mice. The main protective effects includelower release of P-selectin into the circulation, lower shed-ding of HA from the endothelial surface, preserved integrityof the glomerular endothelial glycocalyx, and conservationof EDH-mediated vasorelaxation.
As for the mechanisms involved in HYAL1-mediatedendothelial protection, our main hypothesis is that the absenceof HYAL1 prevents glycocalyx HA shedding during diabetesand, from there, affords protection against diabetes-induced
Figure 7—Expression and activity of SK3 channels in mesenteric arteries. A: SK3 relative mRNA expression in mesenteric arteries of WTand KO mice, without or with STZ-induced diabetes (normalized to b-actin, n = 8–10 in each group). Data are means 6 SEM. Statisticalanalysis was performed on ΔCT values using two-way ANOVA (***P< 0.001) and Bonferroni posttests across genotypes (§P< 0.05; §§P<0.01). B: SK3 and vWF immunostaining in mesenteric arteries of WT and KO mice without and with STZ-induced diabetes. C: Quantificationof SK3 immunostaining using ImageJ (n = 9–12 vessel sections in each group). Data are means 6 SEM. Statistical analysis was performedusing two-way ANOVA (***P < 0.01) and Bonferroni posttests across genotypes (§P < 0.05; §§P < 0.01). D: Residual contraction afterCYPPA activation of SK3 channels. Mesenteric rings of WT and KO mice without or with STZ-induced diabetes (n = 10–12 in each group)were preconstricted with 1027 mol/L U46,619 (PGH2 analog, vasocontractant) in the presence of 1024 mol/L L-NAME (NO synthaseinhibitor) and 1025 mol/L indomethacin (prostaglandin synthesis inhibitor). Vasodilation was then induced using increasing concentrations(33 1027 to 63 1025 mol/L) of CYPPA, an SK3 opener. Data are means6 SEM. Statistical analysis was performed using two-way ANOVAacross genotype (§§§P < 0.001; §§§§P < 0.0001).
2750 HYAL1 Deficiency Protects the Endothelium Diabetes Volume 65, September 2016

vascular damage and maintains a higher level of SK3expression. In turn, this would preserve EDH relaxationduring diabetes, when SK3 expression becomes rate limiting.
Another hypothesis is that elevated plasma HA levels inHYAL1 KO mice facilitate its reincorporation into theglycocalyx during diabetic injury. Previous studies haveshown that the glycocalyx is quickly (30 min) restored afterhyaluronidase treatment if HA and chondroitin sulfate areinfused in sufficiently high amounts, i.e., .100 times thebaseline plasma HA concentration (9). However, the differ-ence in plasma HA concentrations between diabetic WTmice and diabetic HYAL1 KO mice in our study was onlymoderate (+58% in the latter) and unlikely to explain thelarge difference in glycocalyx HA content during exposureof WT versus KO mice to hyperglycemia. Additionally, wehave shown similar molecular size profiles of circulatingHA in HYAL1 KO and WT mice (44). Finally, the hypoth-esis that higher baseline expression of SK3 would explaina thicker glycocalyx seems unlikely based on the knownfunction of SK3.
Conclusion and PerspectivesHYAL1 deficiency had several protective effects againstearly diabetes-induced endothelial and glycocalyx damage.Most, if not all, of these effects may result from a thickerand sturdier glycocalyx (with, e.g., a higher level of endo-thelial SK3 channels). This stronger glycocalyx may, in thelong run, protect against macro- and microvascular com-plications of diabetes, both of which have been linked toglycocalyx impairment. In ApoE-deficient atheromatosis-prone mice, for instance, HA synthesis inhibition using4-methylumbelliferone led to a loss of glycocalyx and en-hanced atherosclerosis (45). HYAL1 inhibition may have an
opposite, protective effect. In addition, the main diabetes-associated microvascular complications, i.e., retinopathy,neuropathy, and glomerular capillary injury, have beenlinked to a loss of glycocalyx. For instance, early diabetesimpacts glomerular permeability and glycocalyx alterationsare reported in diabetic mice even before the developmentof albuminuria (46). In early diabetic retinopathy in ratsand mice, both glycocalyx- and EDH-mediated vasodilationof retinal vessels are altered (47,48). Diabetic neuropathy inmice is accompanied by glycocalyx alteration in brain micro-vessels (49). Finally, diabetes significantly reduces EDH-mediated vasorelaxation in human penile resistance arteriesand downregulates SK3 and IK1 channels in rat corpuscavernosum tissues (32). HYAL1 inhibition could thus bea first step to prevent loss of glycocalyx during the develop-ment of diabetic nephropathy, retinal microangiopathy,blood-brain barrier damage, and erectile dysfunction.
Interestingly, even though HA has been shown toaccumulate in the aortic tunica media of patients withtype 2 diabetes (50), where it may have potential delete-rious effects, our study fails to demonstrate any HA ac-cumulation in the aorta of congenital HYAL1-deficientmice, either before or after induction of diabetes.
Overall, this report suggests that HYAL1 inhibitorscould potentially be explored as a new therapeutic approachto prevent endothelial dysfunction in diabetes. As a caveat,the beneficial effects of HYAL1 deficiency were observed inearly diabetes and should be confirmed over the course ofthe disease.
Acknowledgments. The authors offer sincere thanks to Laurence Jadin(University of Namur, Belgium) for reviewing the manuscript. This research used
Table 1—Beneficial effects observed in HYAL1 KO mice exposed to STZ-induced diabetes as compared with WT mice
Diabetic WTDiabetic KO
(HYAL1 deficiency)Likely mechanism of protection
in absence of HYAL1
Effect of diabetes on:EndotheliumPlasma P-selectin (↑) Diabetes-induced injuries Significant protection Endothelium is less activatedPlasma ICAM1 (↑) Diabetes-induced injuries No benefit No protection*Plasma VCAM1 (2) Absence of change Absence of change No effect of diabetes*EDH vasorelaxation (↓) Diabetes-induced injuries Significant protection Higher baseline SK3 or
thicker glycocalyxSK3 (2) Absence of change Absence of change No effect of diabetes*
GlycocalyxGlycocalyx HA (↓) Diabetes-induced injuries Significant protection Glycocalyx HA is more
adherentGlycocalyx size (2) Absence of change Absence of change No effect of diabetes*Syndecan1 shedding (2) Absence of change Absence of change No effect of diabetes*
Functional glomerular barrierUrinary ACR (↑) Diabetes-induced injuries Significant protection Preserved integrity of
glomerular glycocalyxUrinary ratio of 70- to
40-kDa dextran (↑)Diabetes-induced injuries Significant protection Preserved integrity of
glomerular glycocalyx
(↑), a significant increase in this parameter was measured 4 weeks after STZ-induced diabetes; (↓), a significant decrease; (2), absenceof change. *sICAM1 and sVCAM1 were lower, whereas SK3 expression was higher and glycocalyx was thicker, in HYAL1 KO micethan in WT mice.
diabetes.diabetesjournals.org Dogné and Associates 2751

resources of the Electron Microscopy Service located at the University of Namur.This Service is a member of the Plateforme Technologique Morphologie-Imagerie.Funding. C.D. is a senior research associate of the Fonds National de laRecherche Scientifique (FNRS). F.J. is an MD postdoctoral fellow of the FNRS.Duality of Interest. No potential conflicts of interest relevant to this articlewere reported.Author Contributions. S.D. designed and performed the experiments,contributed to the discussion, and wrote the manuscript. G.R. and F.J. designedand performed the experiments. N.C. contributed to the discussion. C.D. and B.F.designed and supervised the experiments, contributed to the discussion, andreviewed and edited the manuscript. S.D. is the guarantor of this work and, assuch, had full access to all the data in the study and takes responsibility for theintegrity of the data and the accuracy of the data analysis.Prior Presentation. Parts of this study were presented in poster form atthe 2014 Experimental Biology scientific meeting, San Diego, CA, 26–30 April2014, and at the 10th International Conference on Hyaluronan, Florence, Italy,7–11 June 2015.
References1. Hascall V, Esko JD. Hyaluronan. In Essentials of Glycobiology. 2nd ed. VarkiA, Cummings RD, Esko JD, et al., Eds. Cold Spring Harbor, NY, Cold SpringHarbor Laboratory Press, 20092. Csoka AB, Frost GI, Stern R. The six hyaluronidase-like genes in the humanand mouse genomes. Matrix Biol 2001;20:499–5083. Fiszer-Szafarz B, Litynska A, Zou L. Human hyaluronidases: electrophoreticmultiple forms in somatic tissues and body fluids. Evidence for conserved hy-aluronidase potential N-glycosylation sites in different mammalian species.J Biochem Biophys Methods 2000;45:103–1164. Puissant E, Gilis F, Dogné S, Flamion B, Jadot M, Boonen M. Subcellulartrafficking and activity of Hyal-1 and its processed forms in murine macrophages.Traffic 2014;15:500–5155. Natowicz MR, Short MP, Wang Y, et al. Clinical and biochemical manifes-tations of hyaluronidase deficiency. N Engl J Med 1996;335:1029–10336. Martin DC, Atmuri V, Hemming RJ, et al. A mouse model of human mu-copolysaccharidosis IX exhibits osteoarthritis [published correction appears inHum Mol Genet 2008;17:2919]. Hum Mol Genet 2008;17:1904–19157. Tarbell JM, Pahakis MY. Mechanotransduction and the glycocalyx. J InternMed 2006;259:339–3508. Mulivor AW, Lipowsky HH. Role of glycocalyx in leukocyte-endothelial celladhesion. Am J Physiol Heart Circ Physiol 2002;283:H1282–H12919. Henry CB, Duling BR. Permeation of the luminal capillary glycocalyx isdetermined by hyaluronan. Am J Physiol 1999;277:H508–H51410. Kelly R, Ruane-O’Hora T, Noble MI, Drake-Holland AJ, Snow HM. Differentialinhibition by hyperglycaemia of shear stress- but not acetylcholine-mediateddilatation in the iliac artery of the anaesthetized pig. J Physiol 2006;573:133–14511. Mochizuki S, Vink H, Hiramatsu O, et al. Role of hyaluronic acid glycos-aminoglycans in shear-induced endothelium-derived nitric oxide release. AmJ Physiol Heart Circ Physiol 2003;285:H722–H72612. Shakya S, Wang Y, Mack JA, Maytin EV. Hyperglycemia-induced changes inhyaluronan contribute to impaired skin wound healing in diabetes: review andperspective. Int J Cell Biol 2015;2015:70173813. Nieuwdorp M, van Haeften TW, Gouverneur MC, et al. Loss of endothelialglycocalyx during acute hyperglycemia coincides with endothelial dysfunctionand coagulation activation in vivo. Diabetes 2006;55:480–48614. Broekhuizen LN, Lemkes BA, Mooij HL, et al. Effect of sulodexide on en-dothelial glycocalyx and vascular permeability in patients with type 2 diabetesmellitus. Diabetologia 2010;53:2646–265515. Nieuwdorp M, Mooij HL, Kroon J, et al. Endothelial glycocalyx damage co-incides with microalbuminuria in type 1 diabetes. Diabetes 2006;55:1127–113216. Ceriello A, Giugliano D, Dello Russo P, Passariello N, Saccomanno F,Sgambato S. Glycosaminoglycans in human diabetes. Diabete Metab 1983;9:32–34
17. Krege JH, Hodgin JB, Hagaman JR, Smithies O. A noninvasive computerizedtail-cuff system for measuring blood pressure in mice. Hypertension 1995;25:1111–111518. Haserodt S, Aytekin M, Dweik RA. A comparison of the sensitivity, speci-ficity, and molecular weight accuracy of three different commercially availablehyaluronan ELISA-like assays. Glycobiology 2011;21:175–18319. Ikegami-Kawai M, Takahashi T. Microanalysis of hyaluronan oligosaccha-rides by polyacrylamide gel electrophoresis and its application to assay of hy-aluronidase activity. Anal Biochem 2002;311:157–16520. van den Berg BM, Vink H, Spaan JA. The endothelial glycocalyx protectsagainst myocardial edema. Circ Res 2003;92:592–59421. Ghisdal P, Godfraind T, Morel N. Effect of nitro-L-arginine on electrical andmechanical responses to acetylcholine in the superior mesenteric artery fromstroke-prone hypertensive rat. Br J Pharmacol 1999;128:1513–152322. Ding H, Hashem M, Wiehler WB, et al. Endothelial dysfunction in thestreptozotocin-induced diabetic apoE-deficient mouse. Br J Pharmacol 2005;146:1110–111823. Hougaard C, Eriksen BL, Jørgensen S, et al. Selective positive modulation ofthe SK3 and SK2 subtypes of small conductance Ca2+-activated K+ channels. BrJ Pharmacol 2007;151:655–66524. Ikegami-Kawai M, Suzuki A, Karita I, Takahashi T. Increased hyaluronidaseactivity in the kidney of streptozotocin-induced diabetic rats. J Biochem 2003;134:875–88025. Gustavsson C, Agardh CD, Zetterqvist AV, Nilsson J, Agardh E, Gomez MF.Vascular cellular adhesion molecule-1 (VCAM-1) expression in mice retinalvessels is affected by both hyperglycemia and hyperlipidemia. PLoS One 2010;5:e1269926. De Vriese AS, Verbeuren TJ, Van de Voorde J, Lameire NH, Vanhoutte PM.Endothelial dysfunction in diabetes. Br J Pharmacol 2000;130:963–97427. Matsumoto T, Miyamori K, Kobayashi T, Kamata K. Specific impairment ofendothelium-derived hyperpolarizing factor-type relaxation in mesenteric ar-teries from streptozotocin-induced diabetic mice. Vascul Pharmacol 2006;44:450–46028. De Vriese AS, Van de Voorde J, Blom HJ, Vanhoutte PM, Verbeke M,Lameire NH. The impaired renal vasodilator response attributed to endothelium-derived hyperpolarizing factor in streptozotocin–induced diabetic rats is restoredby 5-methyltetrahydrofolate. Diabetologia 2000;43:1116–112529. Colombaro V, Jadot I, Declèves AE, et al. Lack of hyaluronidases exacer-bates renal post-ischemic injury, inflammation, and fibrosis. Kidney Int 2015;88:61–7130. Taylor MS, Bonev AD, Gross TP, et al. Altered expression of small-conductance Ca2+-activated K+ (SK3) channels modulates arterial tone andblood pressure. Circ Res 2003;93:124–13131. Brähler S, Kaistha A, Schmidt VJ, et al. Genetic deficit of SK3 and IK1channels disrupts the endothelium-derived hyperpolarizing factor vasodilatorpathway and causes hypertension. Circulation 2009;119:2323–233232. Zhu JH, Jia RP, Xu LW, et al. Reduced expression of SK3 and IK1 channelproteins in the cavernous tissue of diabetic rats. Asian J Androl 2010;12:599–60433. Zuurbier CJ, Demirci C, Koeman A, Vink H, Ince C. Short-term hypergly-cemia increases endothelial glycocalyx permeability and acutely decreases linealdensity of capillaries with flowing red blood cells. J Appl Physiol (1985) 2005;99:1471–147634. Gouverneur M, Spaan JA, Pannekoek H, Fontijn RD, Vink H. Fluid shearstress stimulates incorporation of hyaluronan into endothelial cell glycocalyx. AmJ Physiol Heart Circ Physiol 2006;290:H458–H45235. Rubio-Gayosso I, Platts SH, Duling BR. Reactive oxygen species mediatemodification of glycocalyx during ischemia-reperfusion injury. Am J Physiol HeartCirc Physiol 2006;290:H2247–H225636. Chappell D, Hofmann-Kiefer K, Jacob M, et al. TNF-alpha induced sheddingof the endothelial glycocalyx is prevented by hydrocortisone and antithrombin.Basic Res Cardiol 2009;104:78–89
2752 HYAL1 Deficiency Protects the Endothelium Diabetes Volume 65, September 2016

37. Yung S, Chau MK, Zhang Q, Zhang CZ, Chan TM. Sulodexide decreasesalbuminuria and regulates matrix protein accumulation in C57BL/6 mice withstreptozotocin-induced type I diabetic nephropathy. PLoS One 2013;8:e5450138. Packham DK, Wolfe R, Reutens AT, et al.; Collaborative Study Group.Sulodexide fails to demonstrate renoprotection in overt type 2 diabeticnephropathy. J Am Soc Nephrol 2012;23:123–13039. Becker BF, Jacob M, Leipert S, Salmon AH, Chappell D. Degradation of theendothelial glycocalyx in clinical settings: searching for the sheddases. Br J ClinPharmacol 2015;80:389–40240. Pohl U, Herlan K, Huang A, Bassenge E. EDRF-mediated shear-induceddilation opposes myogenic vasoconstriction in small rabbit arteries. Am J Physiol1991;261:H2016–H202341. Florian JA, Kosky JR, Ainslie K, Pang Z, Dull RO, Tarbell JM. Heparan sulfateproteoglycan is a mechanosensor on endothelial cells. Circ Res 2003;93:e136–e14242. Salmon AH, Satchell SC. Endothelial glycocalyx dysfunction in disease: al-buminuria and increased microvascular permeability. J Pathol 2012;226:562–57443. Eskens BJ, Zuurbier CJ, van Haare J, Vink H, van Teeffelen JW. Effects oftwo weeks of metformin treatment on whole-body glycocalyx barrier properties indb/db mice. Cardiovasc Diabetol 2013;12:175
44. Bourguignon V, Flamion B. Respective roles of hyaluronidases 1 and 2 inendogenous hyaluronan turnover. FASEB J 2016;30:2108–211445. Nagy N, Freudenberger T, Melchior-Becker A, et al. Inhibition of hyaluronansynthesis accelerates murine atherosclerosis: novel insights into the role ofhyaluronan synthesis. Circulation 2010;122:2313–232246. Nagasu H, Satoh M, Kiyokage E, et al. Activation of endothelial NAD(P)Hoxidase accelerates early glomerular injury in diabetic mice. Lab Invest 2016;96:25–3647. Kumase F, Morizane Y, Mohri S, Takasu I, Ohtsuka A, Ohtsuki H. Glycocalyxdegradation in retinal and choroidal capillary endothelium in rats with diabetesand hypertension. Acta Med Okayama 2010;64:277–28348. Nakazawa T, Kaneko Y, Mori A, et al. Attenuation of nitric oxide- andprostaglandin-independent vasodilation of retinal arterioles induced by acetyl-choline in streptozotocin-treated rats. Vascul Pharmacol 2007;46:153–15949. Liao YJ, Ueno M, Nakagawa T, et al. Oxidative damage in cerebral vesselsof diabetic db/db mice. Diabetes Metab Res Rev 2005;21:554–55950. Heickendorff L, Ledet T, Rasmussen LM. Glycosaminoglycans in the humanaorta in diabetes mellitus: a study of tunica media from areas with and withoutatherosclerotic plaque. Diabetologia 1994;37:286–292
diabetes.diabetesjournals.org Dogné and Associates 2753