identification and evaluation of the probiotic potential of lactobacilli isolated from canine milk
TRANSCRIPT

The Veterinary Journal 185 (2010) 193–198
Contents lists available at ScienceDirect
The Veterinary Journal
journal homepage: www.elsevier .com/ locate/ tv j l
Identification and evaluation of the probiotic potential of lactobacilliisolated from canine milk
Rocío Martín a, Mónica Olivares b, Manuela Pérez a, Jordi Xaus b, Celina Torre c, Leonides Fernández a,Juan M. Rodríguez a,*
a Departamento de Nutrición, Bromatología y Tecnología de los Alimentos, Facultad de Veterinaria, Universidad Complutense de Madrid, 28040 Madrid, Spainb Department of Nutrition and Health, Puleva Biotech, 18004 Granada, Spainc Department of Research and Development, Affinity Petcare, 08174 Sant Cugat del Valles, Spain
a r t i c l e i n f o a b s t r a c t
Article history:Accepted 18 April 2009
Keywords:LactobacillusCanineBitchMilkProbioticsLactation
1090-0233/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.tvjl.2009.04.014
* Corresponding author. Tel.: +34 91 3943747; fax:E-mail address: [email protected] (J.M. Rodrígu
Canine milk protects puppies against infectious diseases through a variety of mechanisms. In this study,the presence of potentially probiotic lactic acid bacteria grown on MRS-Cys agar plates from milk of ninebitches was investigated. The Gram positive catalase negative bacilli identified in this study belonged tofour Lactobacillus species (Lactobacillus reuteri, Lactobacillus fermentum, Lactobacillus murinus, Lactobacillusanimalis), as well as one isolate that was identified as Weissella viridescens. Random amplified polymor-phic DNA profiling revealed 28 different genetic profiles among the lactobacilli isolates. Their probioticpotential was evaluated through different assays, including survival in conditions that resemble thoseexisting in the gastrointestinal tract, production of antimicrobial compounds, adherence to intestinalmucin, degradation of mucin and pattern of antibiotic sensitivity. Some strains showed potential forfuture applications as canine probiotics.
� 2009 Elsevier Ltd. All rights reserved.
Introduction
Lactation is a critical period in canine breeding and precociousweaning is usually associated with high mortality and morbidity.Canine milk not only fulfils all the nutritional requirements for rap-idly-growing puppies, but also protects them against infectiousdiseases. The protective effect of canine milk is due to the com-bined action of a variety of protective factors present in colostrumand milk, such as immunoglobulins, immunocompetent cells, anti-microbial fatty acids, polyamines, fucoylated oligosaccharides,lysozyme and lactoferrin.
Microbiological studies focussed on canine milk are scarce andhave been restricted to the identification of potential pathogenicbacteria in clinical perinatal infections, including lactational masti-tis in bitches and septicaemia in neonatal puppies (Jung et al.,2002; Schäfer-Somi et al., 2003; Ververidis et al., 2007). In contrast,human breast milk has been recently shown to be a source of pro-biotic lactic acid bacteria to the infant gut (Heikkilä and Saris,2003; Martín et al., 2003, 2007a, 2009; Olivares et al., 2006).
Probiotics are living micro-organisms which, upon ingestion incertain numbers, exert health affects beyond inherent basic nutri-tion (Guarner and Schaafsma, 1998). Probiotic bacteria selected for
ll rights reserved.
+34 91 3943743.ez).
practical applications must retain the characteristics for whichthey were originally selected (Salminen et al., 1996). These includegrowth and survival during production and commercialisation, aswell as during transit through the stomach and small intestine.Consequently, it is necessary to test the stability of these character-istics during manufacture, storage and/or ingestion to ensure thatthey are retained in different types of feeds and in the host (Leeand Salminen, 1995). Therefore, the initial screening and selectionof probiotic strains must include testing of the following importantcriteria: identification at the species level, carbohydrate utilisationpatterns, acid and bile tolerance, adhesion to mucin or intestinal-derived epithelial cells, production of antimicrobial substances,antibiotic resistance patterns and ability to influence metabolicactivities in the host (e.g. b-galactosidase activity or productionof vitamins) (Tuomola et al., 2001).
The objective of the present study was to determine whethercanine milk can also be a source of lactic acid bacteria to the puppygut and to evaluate the probiotic potential of such isolates.
Materials and methods
Isolation and enumeration of lactobacilli from canine milk
The protocol (B-16/07) was approved by the Ethical Committee on AnimalExperimentation of Universidad Complutense de Madrid, Spain.
The nine bitches from which the milk samples were obtained belonged to thefollowing breeds: Boston Terrier, Cocker Spaniel, Dalmatian, Golden Retriever(two bitches), Spanish Mastiff, Belgian Shepherd and Yorkshire Terrier (two

194 R. Martín et al. / The Veterinary Journal 185 (2010) 193–198
bitches). All bitches fulfilled the following criteria: (1) healthy bitches without pres-ent or past underlying conditions; (2) normal pregnancy; (3) no use of antibioticsand/or probiotics during the previous 2 months; and (4) absence of any kind of peri-natal problems in both puppies and bitches.
Nipples and mammary areolae were cleaned with soap and sterile water,soaked in chlorhexidine (Cristalmina, Salvat) for 5 min, then dried with a sterilecloth. Milk samples (days 7–12 after parturition) were then collected into a steriletube by manual expression using sterile gloves. The first drops (�500 lL) were dis-carded. The samples were kept on ice until delivery to the laboratory and processedwithin the first 2 h after collection.
Peptone water dilutions of the milk samples were plated in triplicate onto Man,Rogosa and Sharpe (MRS, Oxoid; a medium for the isolation of lactic acid bacteria)agar plates, which were aerobically incubated at 37 �C for 24 h. In parallel, the samesamples were also cultured on MRS supplemented with L-cysteine (0.5 g/L) (MRS-Cys) agar plates, which were incubated anaerobically (85% nitrogen, 10% hydrogen,5% carbon dioxide) in a anaerobic workstation (MINI-MACS, DW Scientific) at 37 �Cfor 48 h. In parallel, to evaluate potential faecal contamination, the samples werealso cultured on Violet Red Bile Agar (VRBA; Difco; a selective medium for the iso-lation of enterobacteria) agar plates, which were aerobically incubated at 37 �C for24 h. In both growth media, the lower limit of detection was 50 colony-formingunits (CFU)/mL (1.69 log10 CFU/mL).
Identification of bacterial isolates from canine milk
Initially, a total of 1660 colonies, including at least one representative of each mor-phological type, were selected from the agar plates. Subsequently, the colonies wereinoculated in to MRS broth tubes, which were incubated aerobically to exclude thosewith fastidious incubation requirements and, therefore, not suitable for practical appli-cations. The isolates (n = 490) that showed good growth (>108 CFU/mL in MRS brothafter 24 h) were examined by phase-contrast microscopy to determine cell morphol-ogy and Gram staining reaction and tested for oxidase and catalase activities.
The isolates were identified by 16S rRNA sequencing. For each strain, a singlebacterial colony was used as a template for PCR amplification of a segment of a16S rRNA gene variable region. The two primers used were those described by Kul-len et al. (2000), namely pbl16 (50-AGAGTTTGATCCTGGCTCAG-30) and mlb16 (50-GGCTGCTGGCACGTAGTTAG-30). The PCR conditions were as follows: 96 �C for30 s, 48 �C for 30 s and 72 �C for 45 s (40 cycles), then a final extension at 72 �Cfor 4 min. The amplified fragment was purified and sequenced using the aboveprimers. Sequencing reactions were prepared using the PRISM ABI BigDye ReadyReaction Terminator Cycle Sequencing kit with AmpliTaq DNA polymerase accord-ing to the manufacturer’s instructions and were run on an ABI 377A automated se-quencer (Applied Biosystems). The resulting sequences were used to searchsequences deposited in the EMBL database using the BLAST algorithm. The identityof the amplified strain was determined on the basis of the highest scores (>98%).
RAPD genotyping of lactobacilli isolated from canine milk
Once the isolates were identified, those belonging to the genus Lactobacilluswere genotyped by randomly amplified polymorphic DNA (RAPD)-PCR analysis.The RAPD technique involves the PCR amplification of random segments of genomicDNA with a single primer of arbitrary nucleotide sequence and allows the differen-tiation between genetically distinct individuals belonging to the same bacterial spe-cies. For this purpose, genomic DNA was isolated from 10 mL of overnight MRScultures using the DNeasy tissue kit (Qiagen), following the protocol recommendedby the supplier for isolation of genomic DNA from Gram positive bacteria. The DNAwas used in subsequent PCR amplifications that were performed using the Ready-to-Go RAPD Analysis kit (GE Healthcare). Then, 5 lL of the PCR mixtures were ana-lysed on 1.2% (w/v) agarose (Sigma) gels that were run for approximately 2 h at80 V and the DNA was visualised and analysed in a gel documentation system(Gel Doc 2000, Bio-Rad), using the Diversity Database software package (Bio-Rad).
Survival after exposition to different conditions
The concentrations of viable cells of the strains after their exposure to different con-ditions that can be of importance for probiotic applications were tested in MRS broth.The following growth conditions were assayed under aerobiosis: 20, 25, 32 and 42 �C(pH 6.2), 37 �C (pH 4.5), presence of canine bile salts (5%), freeze-drying and storageat 20 �C for 15 days and at 80 �C for 30 days in the presence of 15% (v/v) glycerol; inaddition, growth was also tested under anaerobiosis at 37 �C. All assays were per-formed in quadruplicate and the values were expressed as the mean ± standard devia-tion (SD). MRS cultures incubated at 37 �C under aerobiosis were used as controls.
Profiles of carbohydrate fermentation and enzymatic activities
Carbohydrate fermentation patterns were obtained with API Rapid CH fermen-tation strips (BioMérieux) in Lactobacillus Identification Medium (CHL broth, API 50CHL; BioMérieux) as specified by the manufacturer. Enzymatic activities weretested by using the APIzym system (BioMérieux), following the instructions pro-vided by the manufacturer.
Determination of antimicrobial activity
An overlay method (Magnusson and Schnürer, 2001) was used to determinethe ability of the strains to inhibit the growth of various species of bacteria,moulds and yeasts. It was performed using MRS agar plates on which the lactoba-cilli were inoculated as approximately 2 cm long lines and incubated at 37 �C for48 h in anaerobic jars (Oxoid). The plates were then overlaid with the indicatormicroorganisms in 10 mL of soft (0.7%) BHI (Oxoid; a general-purpose mediumsuitable for the cultivation of non-fastidious bacteria) at an approximate concen-tration of 104 CFU.
The following bacteria, from our own culture collection, were employed as indi-cator organisms: Lactococcus lactis MG1614, Lactobacillus sakei NCFB2714, Entero-coccus faecium P21, Enterococcus faecalis TAB28, Pediococcus acidilactici 347,Listeria monocytogenes ScottA, L. monocytogenes Ohio, Listeria seeligeri RdC,Staphylococcus aureus CECT 5191, Staphylococcus epidermidis CECT 231, Escherichiacoli CECT 4076 (O157:H7), E. coli RJM1, E. coli RJM2, E. coli K12 CECT 433, Salmonellacholeraesuis CECT4155, Klebsiella pneumoniae CECT 142, Proteus vulgaris CECT 484and Klebsiella oxytoca CECT 860T. The plates overlaid with bacterial indicators wereincubated at 32 or 37 �C (depending on the optimal temperature) for 48 h. Finally,the plates were examined for clear zones of inhibition (>2 mm) around the lactoba-cilli streaks. All experiments assaying inhibitory activity were performed intriplicate.
Production of bacteriocins
The lactobacilli were grown in MRS broth at 37 �C until the early stationaryphase (A620 �1.0). The cultures were centrifuged at 12,000 g for 10 min at 4 �Cand the supernatants were neutralised to 6.2 with 1 M NaOH, heated at 100 �Cfor 5 min and filter-sterilised through 0.22 lm pore size filters (Millipore). The bac-teriocinogenic activities of the cell-free supernatants were determined by an agarwell diffusion assay. Aliquots (100 lL) of the supernatants were placed in wells(7 mm diameter) cut in cooled BHI agar plates previously seeded (105 CFU/mL) withthe indicator strains. The plates were kept at 4 �C for 2 h, then incubated under opti-mal conditions for growth of the indicator.
The microorganisms employed as indicators of bacteriocinogenic activity werethe Gram positive bacteria used for determination of the antimicrobial spectrum:L. lactis MG1614, L. sakei NCFB2714, E. faecium P21, E. faecalis TAB28, P. acidilactici347, L. monocytogenes ScottA, L. monocytogenes Ohio, L. seeligeri RdC, S. aureus CECT5191 and S. epidermidis CECT 231.
Production of reuterin by the strains of Lactobacillus reuteri
The strains of L. reuteri were cultured in MRS broth at 37 �C for 16 h in anaerobicconditions. Cells were harvested by centrifugation at 4000 g for 5 min, resuspended(1:1 V/V) in a 250 mM glycerol solution and incubated for 3 h at 37 �C in anaerobicconditions. The cells were removed by centrifugation and the presence of reuterinin the cell-free supernatant was detected by the colorimetric method of Smiley andSobolov (1962). This method is based on the colorimetric determination of acroleinformed from the 3-hydroxypropionaldehyde (3-HPA) produced from glycerol byglycerol dehydratase. L. reuteri CECT 925 and Lactobacillus coryniformis CECT 5711(Martín et al., 2005b), two reuterin-producing strains, were used as a positivecontrols.
In parallel, since glycerol dehydratase is essential for reuterin production, PCRwas used to amplify a 279 base pair fragment of the gene encoding the 60-kDaglycerol dehydratase subunit. The primers (GD1 and GD2) and PCR conditions wereselected according to the criteria of Claisse and Lonvaud-Funel (2001). GD1 andGD2 are degenerate primers deduced from a domain of the protein subunit thatis conserved in Gram positive and Gram negative bacteria. L. reuteri CECT 925and L. coryniformis CECT 5711 were used as a positive control.
Production of riboflavin by the strains of L. reuteri
Surprisingly, several colonies of L. reuteri were yellow instead of white. There-fore, we investigated their ability to produce riboflavin (vitamin B2; a compoundthat is typically associated with such colour) when they were grown in MRS both.For this purpose, we followed the method described by Zafra-Gómez et al. (2006),which is based on a high performance liquid chromatography (HPLC) technique.The chromatographic separation was carried out in a 2695 Alliance chromatograph(Waters) equipped with a 2475 fluorescence detector and using a C18 SpherisorbODS-2 column (Waters) set at 40 �C. The fluorescence detection was set at 400/520 nm (excitation/emission) for riboflavin determination. Millennium 4.0 softwarewas used for data treatment.
Adherence to mucin
The adhesion of bacteria to mucin was determined according to the method de-scribed by Cohen and Laux (1995), with some modifications. One hundred microli-tres of a 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES)-bufferedHanks salt solution (HH) of canine mucin (1 mg/mL), obtained as described by

Table 1Enumeration and isolation of bacterial isolates grown on MRS agar plates inoculatedwith bitch milk.
Bitch (n = 9) MRS count(mean CFU/mL ± SD)
Initialisolatesa
(n = 1660)
Identifiedisolatesb
(n = 490)
Boston terrier 132 ± 6 64 26 (A: 26)Cocker spaniel 148 ± 5 72 56 (A: 25, B: 30, C: 1)Dalmatian 609 ± 18 298 45 (A: 27, B: 18)Golden retriever A 451 ± 12 222 48 (A: 28, B: 19, C: 1)Golden retriever B 471 ± 16 233 48 (A: 2, B: 18, C: 24)Spanish mastiff 507 ± 19 248 72 (A: 72)Belgian shepherd 466 ± 14 229 72 (B: 69, C: 3)Yorkshire terrier A 419 ± 10 207 70 (B: 70)Yorkshire terrier B 180 ± 8 87 53 (A: 43, B: 10)
a Fifty percent of the colonies observed in one of the MRS plates.b Initial isolates that showed a good growth in MRS broth incubated in aerobiosis;
A: Gram positive, catalase negative and oxidase negative rods; B: Gram positive,catalase negative and oxidase negative cocci and C: Gram positive, catalase positiveand oxidase negative bacteria.
Table 2Identification of the selected isolates.
Bitch Species Number of isolates
Boston terrier (n = 26) L. murinus 16L. animalis 8L. reuteri 1L. johnsonii 1
Cocker spaniel (n = 56) L. murinus 14L. animalis 8L. fermentum 3E. faecium 17E. faecalis 11Strept. salivarius 2Staph. epidermidis 1
Dalmatian (n = 45) L. murinus 5L. animalis 7L. reuteria 8
b
R. Martín et al. / The Veterinary Journal 185 (2010) 193–198 195
Rinkinen et al. (2003), were immobilised in polystyrene microtitre plates(Maxisorp; Nunc) by overnight incubation at 4 �C. The wells were washed twicewith 250 lL HH.
In parallel, bacteria were grown overnight at 37 �C in MRS broth and the bacte-rial pellets from 1 mL fractions were obtained by centrifugation and washed withHH. Then, 10 lL of 10 mM carboxifluorescein (Sigma) were added to the pelletsand the bacterial suspensions were incubated for 20 min at 37 �C. The bacterial cellswere washed three times with HH, then resuspended in 1 mL HH. A suspension of50 lL of the fluorescent-labelled bacteria (�5 � 107 CFU) was added to each well.After incubation for 1 h at 37 �C, the plates were washed twice with 250 lL of HHto remove unattached cells, then incubated for 1 h at 60 �C with 50 lL 1% sodiumdodecyl sulfate (SDS)–NaOH (0.1 mol/L) to release and lyse bound microorganisms.
Fluorescence was measured in a fluorescence microplate reader (Tecan). Adhe-sion was assessed as the percentage of the fluorescence retained in the wells afterthe washing steps when compared to that present in the labelled bacterial aliquotsoriginally added to the wells. The assays were performed in duplicate and the val-ues were expressed as the mean ± SD.
Degradation of mucin
The potential of the strains to degrade gastric mucin in vitro was evaluated fol-lowing the plate procedure developed by Zhou et al. (2001). Canine mucin and aga-rose type I-A (Sigma) were added to a minimal anaerobic culture medium withoutglucose (Ruseler-van Embeden et al., 1995) at concentrations of 0.5% and 1.5%(W/V), respectively. The plates were incubated at 37 �C anaerobically for 72 h andsubsequently stained with 0.1% amido black in 3.5 M acetic acid for 30 min. Then,they were washed with 1.2 M acetic acid until the mucin lysis zone around thecolony of positive control culture (faecal flora) appeared. These assays were per-formed in triplicate.
Sensitivity to antibiotics
Minimum inhibitory concentrations (MICs) of 12 antimicrobial agents weredetermined by microdilution using the newly developed and standardised ‘lacticacid bacteria susceptibility test medium’ (LSM) broth, consisting of a mixture ofIso-Sensitest (IST; Oxoid) broth (90%) and MRS broth (10%) adjusted to pH 6.7 aspreviously described (Klare et al., 2005). Nine of the antimicrobials tested (ampicil-lin gentamicin, streptomycin, quinupristin/dalfopristin, erythromycin, clindamycin,oxytetracycline, chloramphenicol, kanamycin) were those for which the panel onadditives and products or substances used in animal feed (FEEDAP) of the EuropeanFood Safety Authority (EFSA) has established microbiological breakpoints (cut-offvalues) that enable the distinction between strains of lactobacilli harbouring ac-quired antimicrobial resistances and susceptible strains (EFSA, 2008). MICs of threeadditional antimicrobials agents for which tentative cut-off values for lactobacillihave been suggested (Klare et al., 2007) were also determined.
The antimicrobials were tested in the concentration ranges (mg/L) given inparentheses: penicillin G (0.032-64), ampicillin (0.032-64), gentamicin (1-2048),streptomycin (2-4096), quinupristin/dalfopristin (tested as 30:70 ratio: 0.032-64),erythromycin (0.016-32), clindamycin (0.032-32), oxytetracycline (0.063-128), fu-sidic acid (0.063-128), linezolid (0.016-32), chloramphenicol (0.125-256) and kana-mycin (1-256).
L. reuteri 7E. faecalis 13Strept. bovis 5
Golden retriever A (n = 48) L. murinus 24L. animalis 3L. reuteri 1E. faecalis 10Strept. bovis 9Staph. simulans 1
Golden retriever B (n = 48) L. murinus 2E. faecium 18Staph. simulans 24Yeasts 4
Mastiff (n = 72) L. reuteri 72
Belgian shepherd (n = 72) Strept. bovis 69Staph. pseudointermedius 3
Yorkshire terrier A (n = 70) E. faecalis 70
Yorkshire terrier B (n = 53) L. fermentum 27L. reuteria 1L. reuterib 9W. viridescens 6E. faecium 10
L.: Lactobacillus; E.: Enterococcus; Strept.: Streptococcus; Staph.: Staphylococcus andW.: Weissella.
a White colonies.b Yellow colonies.
Results
Isolation and identification of lactobacilli from canine milk
Colonies were obtained from all the milk samples analysed,both in MRS and MRS-Cys plates, with MRS counts ranging from132 to 609 CFU/mL in different bitches (Table 1). No growth wasobserved on VRBA agar plates. Initially, 50% of the colonies growingon one of the MRS plates inoculated with each sample were sub-cultured, including at least one representative of each morphology(Table 1). All these colonies were inoculated in MRS broth andincubated at 37 �C under aerobiosis. Under such conditions, 490isolates showed good growth (>108 CFU/mL after 24 h) and wereselected for further screening. Most isolates were Gram positive,catalase negative and oxidase negative rods or cocci, while only afew of them were Gram positive, catalase positive and oxidase neg-ative cocci.
Identification of the isolates by 16S rDNA PCR sequencing re-vealed that the Gram-positive bacilli belonged to the GenusLactobacillus, along with one isolate that was classified as Weissellaspp. (Table 2). Coccoid isolates that showed catalase negative andoxidase negative reactions belonged to the genera Enterococcus or
Streptococcus. Catalase positive and oxidase negative cocci wereidentified as Staphylococcus spp. (Table 2). RAPD profiling revealedthe existence of 28 different profiles among the Lactobacillus spp.isolates (Table 3). Subsequently, one representative of eachLactobacillus profile (from now, ‘strain’) was selected for furthercharacterisation.
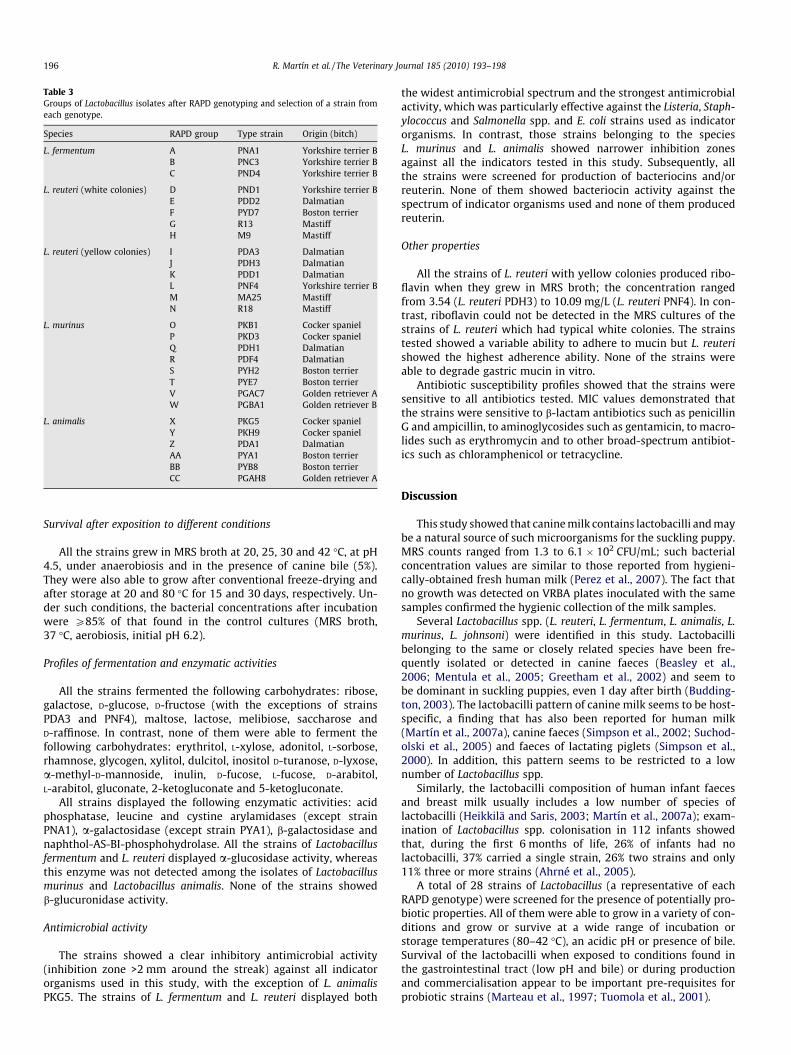
Table 3Groups of Lactobacillus isolates after RAPD genotyping and selection of a strain fromeach genotype.
Species RAPD group Type strain Origin (bitch)
L. fermentum A PNA1 Yorkshire terrier BB PNC3 Yorkshire terrier BC PND4 Yorkshire terrier B
L. reuteri (white colonies) D PND1 Yorkshire terrier BE PDD2 DalmatianF PYD7 Boston terrierG R13 MastiffH M9 Mastiff
L. reuteri (yellow colonies) I PDA3 DalmatianJ PDH3 DalmatianK PDD1 DalmatianL PNF4 Yorkshire terrier BM MA25 MastiffN R18 Mastiff
L. murinus O PKB1 Cocker spanielP PKD3 Cocker spanielQ PDH1 DalmatianR PDF4 DalmatianS PYH2 Boston terrierT PYE7 Boston terrierV PGAC7 Golden retriever AW PGBA1 Golden retriever B
L. animalis X PKG5 Cocker spanielY PKH9 Cocker spanielZ PDA1 DalmatianAA PYA1 Boston terrierBB PYB8 Boston terrierCC PGAH8 Golden retriever A
196 R. Martín et al. / The Veterinary Journal 185 (2010) 193–198
Survival after exposition to different conditions
All the strains grew in MRS broth at 20, 25, 30 and 42 �C, at pH4.5, under anaerobiosis and in the presence of canine bile (5%).They were also able to grow after conventional freeze-drying andafter storage at 20 and 80 �C for 15 and 30 days, respectively. Un-der such conditions, the bacterial concentrations after incubationwere P85% of that found in the control cultures (MRS broth,37 �C, aerobiosis, initial pH 6.2).
Profiles of fermentation and enzymatic activities
All the strains fermented the following carbohydrates: ribose,galactose, D-glucose, D-fructose (with the exceptions of strainsPDA3 and PNF4), maltose, lactose, melibiose, saccharose andD-raffinose. In contrast, none of them were able to ferment thefollowing carbohydrates: erythritol, L-xylose, adonitol, L-sorbose,rhamnose, glycogen, xylitol, dulcitol, inositol D-turanose, D-lyxose,a-methyl-D-mannoside, inulin, D-fucose, L-fucose, D-arabitol,L-arabitol, gluconate, 2-ketogluconate and 5-ketogluconate.
All strains displayed the following enzymatic activities: acidphosphatase, leucine and cystine arylamidases (except strainPNA1), a-galactosidase (except strain PYA1), b-galactosidase andnaphthol-AS-BI-phosphohydrolase. All the strains of Lactobacillusfermentum and L. reuteri displayed a-glucosidase activity, whereasthis enzyme was not detected among the isolates of Lactobacillusmurinus and Lactobacillus animalis. None of the strains showedb-glucuronidase activity.
Antimicrobial activity
The strains showed a clear inhibitory antimicrobial activity(inhibition zone >2 mm around the streak) against all indicatororganisms used in this study, with the exception of L. animalisPKG5. The strains of L. fermentum and L. reuteri displayed both
the widest antimicrobial spectrum and the strongest antimicrobialactivity, which was particularly effective against the Listeria, Staph-ylococcus and Salmonella spp. and E. coli strains used as indicatororganisms. In contrast, those strains belonging to the speciesL. murinus and L. animalis showed narrower inhibition zonesagainst all the indicators tested in this study. Subsequently, allthe strains were screened for production of bacteriocins and/orreuterin. None of them showed bacteriocin activity against thespectrum of indicator organisms used and none of them producedreuterin.
Other properties
All the strains of L. reuteri with yellow colonies produced ribo-flavin when they grew in MRS broth; the concentration rangedfrom 3.54 (L. reuteri PDH3) to 10.09 mg/L (L. reuteri PNF4). In con-trast, riboflavin could not be detected in the MRS cultures of thestrains of L. reuteri which had typical white colonies. The strainstested showed a variable ability to adhere to mucin but L. reuterishowed the highest adherence ability. None of the strains wereable to degrade gastric mucin in vitro.
Antibiotic susceptibility profiles showed that the strains weresensitive to all antibiotics tested. MIC values demonstrated thatthe strains were sensitive to b-lactam antibiotics such as penicillinG and ampicillin, to aminoglycosides such as gentamicin, to macro-lides such as erythromycin and to other broad-spectrum antibiot-ics such as chloramphenicol or tetracycline.
Discussion
This study showed that canine milk contains lactobacilli and maybe a natural source of such microorganisms for the suckling puppy.MRS counts ranged from 1.3 to 6.1 � 102 CFU/mL; such bacterialconcentration values are similar to those reported from hygieni-cally-obtained fresh human milk (Perez et al., 2007). The fact thatno growth was detected on VRBA plates inoculated with the samesamples confirmed the hygienic collection of the milk samples.
Several Lactobacillus spp. (L. reuteri, L. fermentum, L. animalis, L.murinus, L. johnsoni) were identified in this study. Lactobacillibelonging to the same or closely related species have been fre-quently isolated or detected in canine faeces (Beasley et al.,2006; Mentula et al., 2005; Greetham et al., 2002) and seem tobe dominant in suckling puppies, even 1 day after birth (Budding-ton, 2003). The lactobacilli pattern of canine milk seems to be host-specific, a finding that has also been reported for human milk(Martín et al., 2007a), canine faeces (Simpson et al., 2002; Suchod-olski et al., 2005) and faeces of lactating piglets (Simpson et al.,2000). In addition, this pattern seems to be restricted to a lownumber of Lactobacillus spp.
Similarly, the lactobacilli composition of human infant faecesand breast milk usually includes a low number of species oflactobacilli (Heikkilä and Saris, 2003; Martín et al., 2007a); exam-ination of Lactobacillus spp. colonisation in 112 infants showedthat, during the first 6 months of life, 26% of infants had nolactobacilli, 37% carried a single strain, 26% two strains and only11% three or more strains (Ahrné et al., 2005).
A total of 28 strains of Lactobacillus (a representative of eachRAPD genotype) were screened for the presence of potentially pro-biotic properties. All of them were able to grow in a variety of con-ditions and grow or survive at a wide range of incubation orstorage temperatures (80–42 �C), an acidic pH or presence of bile.Survival of the lactobacilli when exposed to conditions found inthe gastrointestinal tract (low pH and bile) or during productionand commercialisation appear to be important pre-requisites forprobiotic strains (Marteau et al., 1997; Tuomola et al., 2001).

R. Martín et al. / The Veterinary Journal 185 (2010) 193–198 197
API 50 CH profiles for the 28 strains of lactobacilli demonstratedphenotypic diversity, even among strains belonging to the samespecies. This result is not unusual, since application of carbohy-drate fermentation profiling to lactobacilli can lead to high intra-species diversity. In a previous study, none of the 18 isolates ofLactobacillus crispatus tested shared the same phenotypic pattern,20 isolates of Lactobacillus gasseri yielded 18 different patterns,20 isolates of Lactobacillus vaginalis isolates yielded 15 differentpatterns and 19 isolates of Lactobacillus jensenii produced 11different patterns (Boyd et al., 2005). In the same study, all theLactobacillus iners isolates and three of the L. vaginalis isolates werenon-reactive for all of the tests in the API 50 CH system.
It has been demonstrated that the use of the current API 50 CHdatabase for identification of commensal Lactobacillus spp. leads toa high frequency of misidentification (Nagy et al., 1991; Klein et al.,1998); therefore, its usefulness for identification of lactobacilli isvery limited and the use of genomic methods, such as 16S rRNAsequencing, is preferable (Wilks et al., 2004). However, carbohy-drate fermentation profiling may provide useful information todiscriminate among strains of lactobacilli belonging to the samespecies.
Lactobacilli have been long considered to constitute the primarymicrobiological barrier to infection by intestinal and urogenitalpathogens. The production of inhibitory substances, such as lacticacid, bacteriocins or reuterin, may affect undesirable or pathogenicbacteria. Bacteriocin or reuterin production could not be detectedin any of the lactobacilli tested in this study. Our results suggestthat differences among the antimicrobial activities displayed bythe strains tested in this work may merely be due to differencesin growth and production of organic acids. Although it has beensuggested that lactic acid can help to maintain a low pH in thegut (Aiba et al., 1998), its production and efficacy in vivo remainuncertain.
Another important property of potential probiotic strains istheir ability to adhere to intestinal or gastric mucin, since this isconsidered a pre-requisite for gut colonisation and may lead tothe competitive exclusion of pathogenic bacteria (Martín et al.,2005a). Some of the strains tested, such as L. fermentum PNA1, L.reuteri M9 and L. murinus PKB1, showed a strong ability to adhereto mucin, reaching values comparable to other highly adhesivestrains of lactobacilli (Olivares et al., 2006).
In addition, six strains of L. reuteri (isolated from three differentbitches) were able to synthesise riboflavin. This is a relevant obser-vation since, up to the present, it was thought that lactobacilli wereunable to produce this vitamin and, therefore, were dependent onexternal riboflavin for growth (Ledesma et al., 1976; Møretrø et al.,1998).
The verification that probiotic strains lack acquired antimicro-bial resistance properties is an important requisite when consider-ing them to be safe for animal consumption. Antimicrobialsusceptibility testing of lactobacilli can be performed by severalmethods, but dilution methods generally provide a more reliableindication of the intrinsic or acquired nature of a given resistancephenotype (Klare et al., 2007). All the strains of lactobacilli testedin this study did not demonstrate antimicrobial resistance whentested according to EFSA guidelines (EFSA, 2008).
The species of lactobacilli isolated from canine milk in thisstudy are among those considered as potential probiotic bacteria.L. reuteri, L. fermentum or L. animalis have already been tested as ca-nine probiotics (Manninen et al., 2006; Biagi et al., 2007). There-fore, the milk of healthy bitches may be a source of potentiallyprobiotic lactobacilli, with a role in protecting mothers and/or pup-pies against infectious diseases. Although cross-species efficacy hasbeen demonstrated for some probiotic strains, one criterion forselection of a probiotic is host species specificity. However, mostof the commercial probiotic strains for dogs are not of canine
origin. In a previous study, it was shown that Lactobacillus rhamnosusGG, a human isolate, survived gastrointestinal transit in dogs, butfaecal colonisation was less efficient than in humans (Weese andAnderson, 2002).
The origin of the lactobacilli that colonise the neonatal gut is asubject of debate. In the past, it was suggested that they were ac-quired by oral contamination with maternal lactobacilli during thetransit through the birth canal; however, molecular studies haveshown that human Lactobacillus colonisation is not significantlyrelated to the delivery method (vaginal delivery or Caesarean sec-tion) (Matsumiya et al., 2002; Martín et al., 2003; Ahrné et al.,2005). In contrast, breast milk constitutes a good source of lactoba-cilli (Heikkilä and Saris, 2003) and is responsible for the verticalmother-to-child transmission of lactobacilli and bifidobacteriainhabiting the gut (Matsumiya et al., 2002; Martín et al., 2003,2007b, 2009). Recently, it has been shown that live commensalbacteria can spread from the maternal gut to the epithelium ofthe lactating mammary gland through an endogenous routeinvolving dendritic cells and macrophages: the enteromammarypathway (Martín et al., 2004; Perez et al., 2007). Work is in pro-gress to compare the probiotic potential of strains of lactobacilliisolated from canine milk with that of strains recovered fromcanine faeces or gut.
Conclusions
Some strains of lactobacilli isolated from canine milk havepotential for future applications as canine probiotics. In particular,L. reuteri and L. fermentum exhibited high antimicrobial activities,high survival rates after exposure to different adverse physical con-ditions and desirable enzymatic activities (such as production ofa-glucosidase). In addition, these strains did not degrade mucinand the MICs of several antibiotics were within the values recom-mended by the EFSA.
Conflict of interest statement
C. Torre is employed by Affinity Petcare. None of the otherauthors have a financial or personal relationship with other peopleor organisations that could inappropriately influence or bias thecontent of the paper.
Acknowledgements
We are grateful to Elena Aguado (Clínica Veterinaria Sevilla laNueva, Spain) for the collection of the milk samples. This workwas funded by Affinity Petcare S.A. (Sant Cugat del Valles, Spain).
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.tvjl.2009.04.014.
References
Ahrné, S., Lönnermark, E., Wold, A.E., Aberg, N., Hesselmar, B., Saalman, R.,Strannegard, I.L., Molin, G., Adlerberth, I., 2005. Lactobacilli in the intestinalmicrobiota of Swedish infants. Microbes and Infection 7, 1256–1262.
Aiba, Y., Suzuki, N., Kabir, A.M.A., Takagi, A., Koga, Y., 1998. Lactic acid-mediatedsuppression of Helicobacter pylori by the oral administration of Lactobacillussalivarius as a probiotic in a gnotobiotic murine model. American Journal ofGastroenterology 11, 2097–2111.
Beasley, S.S., Manninen, T.J.K., Saris, P.-E.J., 2006. Lactic acid bacteria isolated fromcanine faeces. Journal of Applied Microbiology 101, 131–138.
Biagi, G., Cipollini, I., Pompei, A., Zaghini, G., Matteuzzi, D., 2007. Effect of aLactobacillus animalis strain on composition and metabolism of the intestinalmicroflora in adult dogs. Veterinary Microbiology 124, 160–165.

198 R. Martín et al. / The Veterinary Journal 185 (2010) 193–198
Boyd, M.A., Antonio, M.A.D., Hillier, S.L., 2005. Comparison of API 50 CH strips towhole-chromosomal DNA probes for identification of Lactobacillus species.Journal of Clinical Microbiology 43, 5309–5311.
Buddington, R.K., 2003. Postnatal changes in bacterial populations in thegastrointestinal tract of dogs. American Journal of Veterinary Research 64,646–651.
Claisse, O., Lonvaud-Funel, A., 2001. Primers and a specific DNA probe for detectinglactic acid bacteria producing 3-hydroxypropionaldehyde from glycerol inspoiled ciders. Journal of Food Protection 64, 833–837.
Cohen, P.S., Laux, D.C., 1995. Bacterial adhesion to and penetration of intestinalmucus in vitro. Methods in Enzymology 253, 309–314.
EFSA, 2008. Update of the criteria used in the assessment of bacterial resistance toantibiotics of human or veterinary importance. EFSA Journal 732, 1–15.
Greetham, H.L., Giffard, C., Hutson, R.A., Collins, M.D., Gibson, G.R., 2002.Bacteriology of the Labrador dog gut: a cultural and genotypic approach.Journal of Applied Microbiology 93, 640–646.
Guarner, F., Schaafsma, G.J., 1998. Probiotics. International Journal of FoodMicrobiology 39, 237–238.
Heikkilä, M.P., Saris, P.E.J., 2003. Inhibition of Staphylococcus aureus by thecommensal bacteria of human milk. Journal of Applied Microbiology 95, 471–478.
Jung, C., Wehrend, A., König, A., Bostedt, H., 2002. Untersuchung zu vorkommen,differenzierung und erregerspektrum caniner mastitiden. Praktischer Tierarzt83, 508–511.
Klare, I., Konstabel, C., Müller-Bertling, S., Reissbrodt, R., Huys, G., Vancanneyt, M.,Swings, J., Goossens, H., Witte, W., 2005. Evaluation of new broth media formicrodilution antibiotic susceptibility testing of lactobacilli, pediococci,lactococci, and bifidobacteria. Applied and Environmental Microbiology 71,8982–8986.
Klare, I., Konstabel, C., Werner, G., Huys, G., Vankerckhoven, V., Kahlmeter, G.,Hildebrandt, B., Müller-Bertling, S., Witte, W., Goossens, H., 2007. Antimicrobialsusceptibilities of Lactobacillus, Pediococcus and Lactococcus human isolates andcultures intended for probiotic or nutritional use. Journal of AntimicrobialChemotherapy 59, 900–912.
Klein, G., Zill, E., Schindler, R., Louwers, J., 1998. Peritonitis associated withvancomycin-resistant Lactobacillus rhamnosus in a continuous ambulatoryperitoneal dialysis patient: organism identification, antibiotic therapy, andcase report. Journal of Clinical Microbiology 36, 1781–1783.
Kullen, M.J., Sanozky-Dawes, R.B., Crowell, D.C., Klaenhammer, T.R., 2000. Use ofDNA sequence of variable regions of the 16SrRNA gene for rapid and accurateidentification of bacteria in the Lactobacillus acidophilus complex. Journal ofApplied Microbiology 89, 511–518.
Ledesma, O.V., Ruiz Holgado, A., Oliver, G., 1976. A synthetic medium forcomparative nutritional studies of lactobacilli. Journal of Applied Bacteriology42, 123–133.
Lee, Y.K., Salminen, S., 1995. The coming of age of probiotics. Trends in Food Science& Technology 6, 241–245.
Møretrø, T., Hagen, B.F., Axelsson, L., 1998. A new, completely defined medium formeat lactobacilli. Journal of Applied Microbiology 85, 715–722.
Magnusson, J., Schnürer, J., 2001. Lactobacillus coryniformis subsp. coryniformis strainSi3 produces a broad-spectrum proteinaceous antifungal compound. Appliedand Environmental Microbiology 67, 1–5.
Manninen, T.J.K., Rinkinen, M.L., Beasley, S.S., Saris, P.E.J., 2006. Alteration of thecanine small-intestinal lactic acid bacterium microbiota by feeding ofpotential probiotics. Applied and Environmental Microbiology 72,6539–6543.
Marteau, P., Minekus, M., Havenaar, R., Huis In’t Veld, J.H.J., 1997. Survival of lacticacid bacteria in a dynamic model of the stomach and small intestine: validationand the effects of bile. Journal of Dairy Science 80, 1031–1037.
Martín, R., Jiménez, E., Heilig, H., Fernández, L., Marín, M.L., Zoetendal, E.G.,Rodríguez, J.M., 2009. Isolation of bifidobacteria from breast milk andassessment of the bifidobacterial population by PCR-denaturing gradient gelelectrophoresis and quantitative real-time PCR. Applied and EnvironmentalMicrobiology 75, 965–969.
Martín, R., Langa, S., Reviriego, C., Jiménez, E., Marín, M.L., Xaus, J., Fernández, L.,Rodríguez, J.M., 2003. Human milk is a source of lactic acid bacteria for theinfant gut. Journal of Pediatrics 143, 754–758.
Martín, R., Langa, S., Reviriego, C., Jiménez, E., Marín, M.L., Olivares, M., Boza, J.,Jimenez, J., Fernandez, L., Xaus, J., Rodriguez, J.M., 2004. The commensalmicroflora of human milk: new perspectives for food bacteriotherapy andprobiotics. Trends in Food Science and Technology 15, 121–127.
Martín, R., Heilig, H.G., Zoetendal, E.G., Rodríguez, J.M., 2007a. Diversity of theLactobacillus group in breast milk and vagina of healthy women and potentialrole in the colonization of the infant gut. Journal of Applied Microbiology 103,2638–2644.
Martín, R., Heilig, H.G., Zoetendal, E.G., Jiménez, E., Fernández, L., Smidt, H.,Rodríguez, J.M., 2007b. Cultivation-independent assessment of the bacterial
diversity of breast milk among healthy women. Research in Microbiology 158,31–37.
Martín, R., Olivares, M., Marín, M.L., Fernández, L., Xaus, J., Rodríguez, J.M., 2005a.Probiotic potential of three lactobacilli strains isolated from breast milk. Journalof Human Lactation 21, 8–17.
Martín, R., Olivares, M., Marín, M.L., Xaus, J., Fernández, L., Rodríguez, J.M., 2005b.Characterization of a reuterin-producing Lactobacillus coryniformis strainisolated from a goat’s milk cheese. International Journal of Food Microbiology104, 267–277.
Matsumiya, Y., Kato, N., Watanabe, K., 2002. Molecular epidemiological study ofvertical transmission of vaginal Lactobacillus species from mothers to newborninfants in Japanese, by arbitrarily primed polymerase chain reaction. Journal ofInfection and Chemotherapy 8, 43–49.
Mentula, S., Harmoinen, J., Heikkila, M., Westermarck, E., Rautio, M., Huovinen, P.,Kononen, E., 2005. Comparison between cultured small-intestinal and fecalmicrobiotas in Beagle dogs. Applied and Environmental Microbiology 71,4169–4175.
Nagy, E., Petterson, M., Mardh, P.A., 1991. Antibiosis between bacteria isolated fromthe vagina of women with and without signs of bacterial vaginosis. ActaPathologica, Microbiologica et Immunologica Scandinavica 99, 739–744.
Olivares, M., Díaz-Ropero, M.P., Martín, R., Rodríguez, J.M., Xaus, J., 2006.Antimicrobial potential of four Lactobacillus strains isolated from breast milk.Journal of Applied Microbiology 101, 72–79.
Perez, P.F., Doré, J., Leclerc, M., Levenez, F., Benyacoub, J., Serrant, P., Segura-Roggero, I., Schiffrin, E.J., Donnet-Hughes, A., 2007. Bacterial imprinting ofthe neonatal immune system: lessons from maternal cells? Pediatrics 119,e724–e732.
Rinkinen, M., Westermarck, E., Salminen, S., Ouwehand, A.C., 2003. Absence of hostspecificity for in vitro adhesion of probiotic lactic acid bacteria to intestinalmucus. Veterinary Microbiology 97, 55–61.
Ruseler-van Embeden, J.G.H., Lieshcut-van, L.M.C., Gosselink, M.J., Marteau, P., 1995.Inability of Lactobacillus casei strain GG, L. acidophilus and Bifidobacteriumbifidum to degrade intestinal mucus glycoproteins. Scandinavian. Journal ofGastroenterology 30, 675–680.
Salminen, S., Isolauri, E., Salminen, E., 1996. Clinical uses of probiotics for stabilizingthe gut mucosal barrier: successful strains and future challenges. Antonie VanLeeuwenhoek 70, 347–358.
Schäfer-Somi, S., Spergser, J., Breitenfellner, J., Aurich, J.E., 2003. Bacteriologicalstatus of canine milk and septicaemia in neonatal puppies – a retrospectivestudy. Journal of Veterinary Medicine B 50, 343–346.
Simpson, J.M., Martineau, B., Jones, W.E., Ballam, J.M., Mackie, R.I., 2002.Characterization of fecal bacterial populations in canines: effects of age, breedand dietary fiber. Microbial Ecology 44, 186–197.
Simpson, J.M., McCracken, V.J., Gaskins, H.R., Mackie, R.I., 2000. Denaturing gradientgel electrophoresis analysis of 16S ribosomal DNA amplicons to monitorchanges in fecal bacterial populations of weaning pigs after introduction ofLactobacillus reuteri strain MM53. Applied and Environmental Microbiology 66,4705–4714.
Smiley, K.L., Sobolov, M., 1962. A cobamide-requiring glycerol dehydrase from anacrolein-forming Lactobacillus. Archives of Biochemistry and Biophysics 97,538–543.
Suchodolski, J.S., Ruaux, C.G., Steiner, J.M., Fetz, K., Williams, D.A., 2005. Assessmentof the qualitative variation in bacterial microflora among compartments of theintestinal tract of dogs by use of a molecular fingerprinting technique. AmericanJournal of Veterinary Research 66, 1556–1562.
Tuomola, E., Crittenden, R., Playne, M., Isolauri, E., Salminen, S., 2001. Qualityassurance criteria for probiotic bacteria. American Journal of Clinical Nutrition73, 393S–398S.
Ververidis, H.N., Mavrogianni, V.S., Fragkou, I.A., Orfanou, D.C., Gougoulis, D.A.,Tzivara, A., Gouletsou, P.G., Athanasiou, L., Boscos, C.M., Fthenakis, G.C., 2007.Experimental staphylococcal mastitis in bitches: clinical, bacteriological,cytological, haematological and pathological features. Veterinary Microbiology124, 95–106.
Weese, J.S., Anderson, M.E.C., 2002. Preliminary evaluation of Lactobacillusrhamnosus strain GG, a potential probiotic in dogs. Canadian VeterinaryJournal 43, 771–774.
Wilks, M., Wiggins, R., Whiley, A., Hennessy, E., Warwick, S., Porter, H., Corfield, A.,Millar, M., 2004. Identification and H2O2 production of vaginal lactobacilli frompregnant women at high risk of preterm birth and relation with outcome.Journal of Clinical Microbiology 42, 713–717.
Zafra-Gómez, A., Garballo, A., Morales, J.C., García-Ayuso, L.E., 2006. Simultaneousdetermination of eight water-soluble vitamins in supplemented foods by liquidchromatography. Journal of Agricultural and Food Chemistry 54, 4531–4536.
Zhou, J.S., Gopal, P.K., Hill, H.S., 2001. Potential probiotic lactic acid bacteriaLactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) andBifidobacterium lactis (HN019) do not degrade gastric mucin in vitro.International Journal of Food Microbiology 63, 81–90.