identification of a 14-kda laminin binding protein (hlbp14) in human melanoma cells that is...
TRANSCRIPT

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 297, No. 1, August 15, pp. 132-138, 1992
Identification of a 14-kDa Laminin Binding Protein (HLBP14) in Human Melanoma Cells That Is Identical to the 14-kDa Galactoside Binding Lectin
Vincent Castronovo,*‘l Frank Luyten,? Frederic van den Brtile,* and Mark E. Sobel* *Tumor Invasion and Metastasis Section, National Cancer Institute, Bethesda, Maryland 20892; and tBone Cell Biology Section, National Institute of Dental Research, Bethesda, Maryland 20892
Received February 17, 1992, and in revised form May 1, 1992
The carbohydrate moieties present on laminin play a crucial role in the multiple biological activities of this basement membrane glycoprotein. We report the iden- tification of a human laminin binding protein with an apparent molecular mass of 14 kDa on sodium dodecyl sulfate-polyacrylamide gels that was found, after puri- fication and amino acid microsequencing, to be identical to the previously described 14-kDa galactoside binding soluble L- 14 lectin. We have designated this human lam- inin binding protein as HLBP14. HLBP14 was purified from human melanoma cells in culture by laminin affinity chromatography and gel electroelution. We demonstrate that HLBP14 binds specifically to the poly-N-acetyllac- tosamine residues of murine laminin and does not bind to other glycoproteins that do not contain such structures, such as fibronectin. HLBP14 was eluted from a murine laminin column by lactose, N-acetyllactosamine, and ga- lactose but not by other control saccharides, including glucose, fucose, mannose, and melibiose. It did not bind to laminin treated with endo-&galactosidase. Lactose also eluted HLBP14 off a human laminin affinity column, implying that human laminin also contains poly-N-ace- tyllactosamine residues. On immunoblots, polyclonal an- tibodies raised against HLBP14 recognized HLBP14 as well as 31- and 67-kDa molecules that are also laminin binding proteins, indicating that these proteins share common epitopes. L-14, a dimeric lactose binding lectin, is expressed in a wide variety of tissues. Although the expression of this molecule has been linked to a variety of biological events, the elucidation of its specific func- tions has been elusive. The observation that HLBP14, a human cancer cell laminin binding protein, is identical to L-14 strongly suggests that the functions attributed to this lectin could be mediated, at least in part, through
1 To whom correspondence should be addressed at Laboratory of Pa- thology, National Cancer Institute, NIH, Building 10, Room 2A33, Be- thesda, MD 20892.
132
its ability to interact with the poly-N-acetyllactosamine residues of laminin. HLBP14 could potentially play a role during tumor invasion and metastasis by modulating the interactions between cancer cells and laminin. 0 1992
Academic Press, Inc.
The interaction between cells and laminin, a major basement membrane glycoprotein (l), is a determinant event during a large variety of normal and pathologic bio- logical processes (2-4). Laminin participates in the at- tachment, spreading, migration, proliferation, and differ- entiation of normal and malignant cells, as well as in neurite outgrowth and in tumor invasion and metastasis. Laminin is a macromolecule containing several functional domains (4). Previous studies using synthetic peptides and proteolytically generated fragments of laminin have identified several domains of the protein that mediate specific biological activities (4-6). Recent reports have shown that the carbohydrate moieties present on laminin also play an important role in mediating certain functions of this glycoprotein (7, 8). Indeed, laminin is a heavily glycosylated protein with up to 12-27% of its mass pro- vided by carbohydrate residues (9-14). Complex-type asn- linked oligosaccharides containing poly-N-acetyllacto- samine sequences ([3~Gal/31,4GlcNAc@l],) are the most commonly represented carbohydrates on laminin and ap- pear to be functionally important (12-14).
The functions of laminin are mediated through a variety of cell surface proteins (15-21). The first laminin binding protein to be identified was designated “laminin receptor” (15, 22, 23); it is a 67-kDa protein that is present on the surface of normal and neoplastic cells and has a high af- finity (& = 2 nM) for laminin (15). Other laminin binding proteins include members of the integrin family (16, 18, 21) and the galactosyltransferase (20). While we were preparing this manuscript, the murine Mac-2 (24) and
0003.9861/9‘2 $5.00 Copyright 0 1992 by Academic Press, Inc.
All rights of reproduction in any form reserved.

14.kDa GALACTOSIDE BINDING LECTIN BINDS TO LAMININ 133
the human Mac-2 (25) antigen as well as a 14-kDa calf heart lectin (26) were reported to be potential laminin binding proteins.
In the course of studying the non-integrin metastasis- associated 67-kDa laminin receptor in human cancer cells, we identified, purified, and characterized a laminin bind- ing protein with an apparent molecular mass of 14 kDa. It was revealed by microsequencing to be identical to a human 14-kDa galactoside binding soluble lectin (L-14) that has been characterized but whose physiological roles have not been clearly established (27). We have designated this human laminin binding protein as HLBP14 and, in this report, we present evidence that HLBP14 specifically interacts with the poly-N-acetyllactosamine residues present on laminin. Furthermore, we present immunologic evidence that HLBP14 shares common epitopes with 31- and 67-kDa laminin binding proteins. These three mol- ecules may represent a family of related cell surface mul- tifunction proteins that is part of the P-galactoside binding lectin family.
MATERIALS AND METHODS
Reagents. Laminin was purified from the mouse Engelbreth Holm Swarm tumor as described (1). Human laminin was obtained from Telios Pharmaceuticals, Inc. (San Diego, CA). A&Gel 10 resin was obtained from Bio-Rad (Richmond, CA). Human fibronectin was purchased from Collaborative Research, Inc. (Bedford, MA). Fetuin, asialofetuin, lactose, galactose, melibiose, mannose, glucose, jack bean P-galactosidase, green coffee bean a-galactosidase, and Vibrio cholerae neuraminidase were obtained from Sigma (St. Louis, MO). Endo-/3-galactosidase (Bacteroides fragilis) and Chaps’ were purchased from Boehringer-Mannheim (In- dianapolis, IN). [?S]Methionine (1062 Ci/mmol) was purchased from ICN (Irvine, CA). iZsI (13.7 mCi/yg) was obtained from Amersham Corp. (Arlington Heights, IL). Iodo-beads were purchased from Pierce Chem- ical Co. (Rockford, IL).
Cells and tissues. Human melanoma A2058 cells have been previously described (28). The cells were routinely grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum, penicillin (100 units/ml), and streptomycin (100 yg/ml) (GIBCO BRL, Grand Island, NY).
Laminin affinity chromatography. Laminin binding proteins were purified by affinity chromatography on laminin columns as described (29), except that (a) laminin was coupled to Affi-Gel 10 resin instead of CNBr-activated Sepharose 4B, (b) membrane protein extraction buffer contained 0.1% Chaps instead of 1% Nonidet-P40. Briefly, lo9 human melanoma cells were scraped with a rubber policeman, Dounce homog- enized, and incubated in the extraction buffer (25 mM Tris, pH 7.5, 150 mM NaCl, 0.1% Chaps, 3 mM MgClr, 1 mM CaCl,, 10 ag/ml aprotinin, 5 mM benzamidine, 50 yg/ml PMSF, 1 pM pepstatin) as described (29). The solubilized proteins were incubated overnight with the laminin col- umn. The laminin column was washed extensively at 4°C with 150 mM NaCl and finally with 400 mM NaCl. Specifically bound proteins were eluted using 10% acetic acid.
Protein purification and microsequencing. The column-eluted frac- tions containing laminin binding proteins were concentrated on centricon
’ Abbreviations used: Chaps, 3-[(3-cholamidopropyl)dimethylammo- niolpropanesulfonic acid, DMEM, Dulbecco’s modified Eagle’s medium; PMSF, phenylmethylsulfonyl fluoride; SDS, sodium dodecyl sulfate; BSA, bovine serum albumin; PBS, phosphate-buffered saline; PAGE, polyacrylamide gel electrophoresis; TBS, Tris-buffered saline.
10 filters (Amicon, Danvers, MA) and separated on a 12.5% SDS-poly- acrylamide gel according to Laemmli (30). A laminin binding protein with an apparent molecular mass of 14 kDa, designated HLBP14, was ultimately purified by electroelution according to a previously published protocol (31). Purity of the electroeluted protein was assessed on a 15% SDS-polyacrylamide gel. N-terminal amino acid sequencing of HLBP14 was unsuccessful. Therefore, HLBP14 (360 pmol) was digested with trypsin as described (32). Generated peptides were separated by reverse phase HPLC performed on a Vydac Cl8 column. Purified peptides were applied to a gas phase microsequencer (Applied Biosystem Model 4708 on line with HPLC Applied Biosystem 120). The amino acid sequences were examined for homology with previously described protein sequences using the Protein Identification Resource of the National Biomedical Research Foundation (Washington, DC).
El&ion of metabolically radiolabeled laminin bindingproteins with dif- ferent sugars. The carbohydrate structures involved in the interaction between laminin and HLBP14 were qualitatively investigated. A2058 cells were metabolically labeled using [35S]methionine as described (28). Total cell extracts were prepared as described (29) and were incubated with laminin coupled to Affi-Gel 10 resin as described above. After ex- tensive washes as described above, laminin binding proteins were eluted with 100 mM solutions of lactose, N-acetyllactosamine, glucose, galactose, melibiose, fucose, or mannose. The eluted fractions were concentrated on centricon 10 filters and separated on a 12.5% SDS-polyacrylamide gel. The radiolabeled laminin binding proteins were revealed by auto- radiography. The elution properties of the various sugars were quali- tatively evaluated. In sugar competition experiments, the cell extracts were incubated with the laminin affinity column in the presence of 300 mM lactose, galactose, melibiose, or mannose. After washes as described above, the proteins bound to the laminin column were eluted with 100 mM lactose. We also tested the possibility that human laminin may exhibit different binding properties than mouse EHS laminin. Radio- labeled cell extracts were applied to a human laminin-At&Gel chro- matography column following the same protocol used for murine laminin chromatography, and specifically bound molecules were eluted with 100 mM lactose. In some experiments, the cells extracts were applied to a column of fibronectin or fetuin prepared as described for the laminin affinity column.
Effect of the treatment of laminin by eroglycosidases or endo-(5galac- tosidase on its binding properties. Laminin coupled to Affi-Gel was treated with endo-a-galactosidase, a-galactosidase, or fi-galactosidase. For endo-fl-galactosidase and a-galactosidase digestion, 1 ml of laminin coupled to Afh-Gel was washed three times with 50 mM sodium acetate, pH 5.8, and incubated for 24 h at 37°C with 100 mU of the corresponding enzyme in the same buffer containing 100 pg bovine serum albumin (BSA)/ml. For P-galactosidase digestion, 1 ml of laminin coupled to At&Gel was washed three times with 50 mM sodium citrate buffer, pH 4, and incubated for 24 h at 37°C with 100 mU of enzyme in the same buffer containing 100 fig BSA/ml. After the enzyme digestion, the lam- inin-Affi-Gel beads were washed three times with PBS. The effect of the different enzyme treatments on the ability of laminin to bind laminin binding protein was assessed using the affinity chromatography protocols described above. The bound proteins were eluted with 100 mM lactose and examined by SDS-PAGE as described above.
Anti-HLBP14 antibodies and immunoblots. Polyclonal antibodies against HLBP14 were raised in rabbits by serial subcutaneous injection of polyacrylamide gel-purified proteins (BAbCO, Berkeley, CA). Affinity- purified antibody 4099 was raised against a synthetic peptide deduced from the cDNA sequence of the 67-kDa laminin receptor and has been previously described (28). Complement was heat inactivated in all sera. Anti-HLBP14 antibodies were tested in immunoblot experiments per- formed as described (28). Briefly, laminin binding proteins eluted with lactose from a laminin affinity column were separated on a 12.5% SDS- polyacrylamide gel and transferred to a PDVF membrane (Millipore, Bedford, MA) in a Trans.Blot apparatus (Bio-Rad, Richmond, CA). After blocking of the residual binding sites by incubation in a solution containing 5% nonfat dry milk, 1% ovalbumin, 5% fetal calf serum, and

134 CASTRONOVO ET AL.
7.5% glycine, the PDVF membranes were washed three times in washing solution (2% nonfat dry milk, 2% ovalbumin, 10% fetal calf serum in PBS). Anti-HLBP14 sera as well as affinity-purified anti-67-kDa laminin receptor synthetic peptide antibody 4099 were diluted 1:lOO in TBS (20 mM Tris, pH 7.5, 500 mM NaCl) an d incubated with the blotted PDVF membranes at 4°C overnight with gentle shaking. The blots were washed three times for 10 min each in the washing solution followed by incu- bation for 30 min at room temperature with affinity-purified goat anti- rabbit IgG conjugated to peroxidase (Boerhinger-Mannheim) diluted 1: 1000 in PBS. After three washes for 10 min each in the washing solution and one wash in TBS, the PDVF membranes were stained with HRP color development solution (Bio-Rad). In control experiments, the blots were incubated with corresponding preimmune sera.
RESULTS
Elution of Laminin Binding Proteins from Laminin Afinity Columns
Protein extracts from human melanoma cells were in- cubated with a laminin affinity column, and after ex- haustive washes, molecules specifically bound to the lam- inin column were eluted using 10% acetic acid. The major proteins in the acetic acid-eluted fractions had apparent molecular masses of 67,31, and 14 kDa as calculated from their mobility on SDS-polyacrylamide gels (Fig. 1A). The 67-kDa protein was found by immunoblot and immuno- precipitation to be the previously identified 67-kDa lam- inin receptor (15, 22, 23).
Purification and Microsequencing of the 14-kDa Laminin Binding Protein We decided to further purify and microsequence the
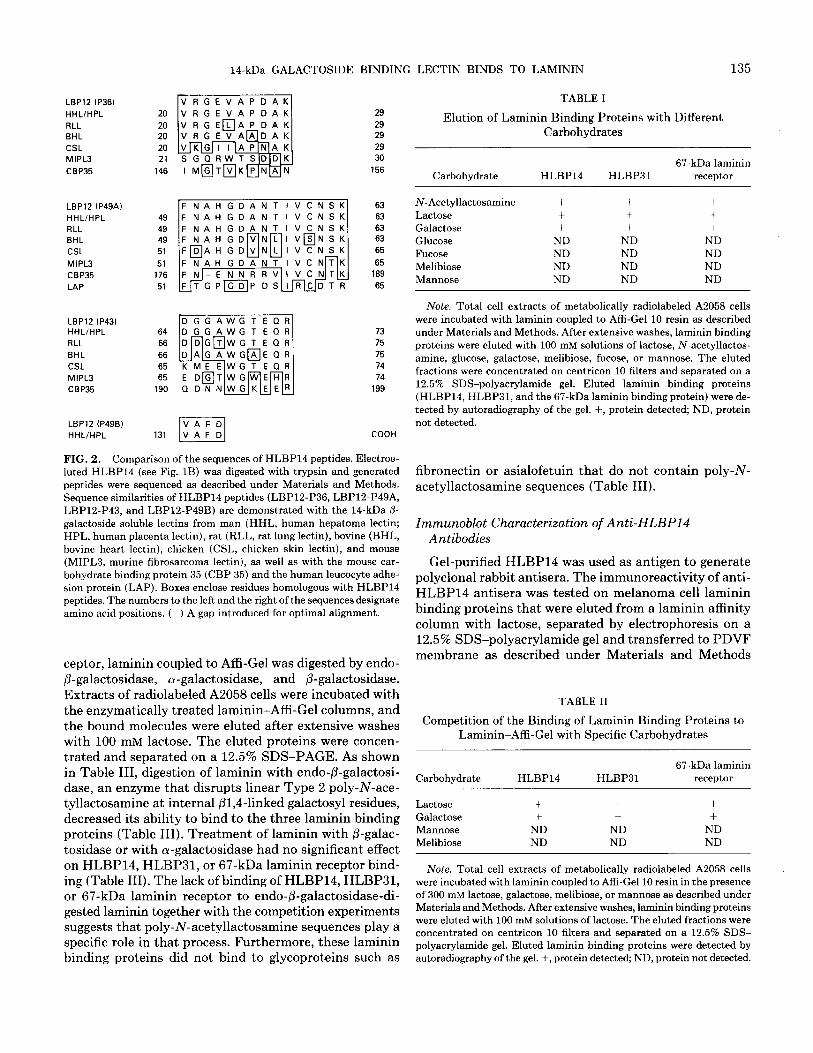
14-kDa protein since, at the time, such a molecule had not been described in human cells as a laminin binding protein. Apparent homogeneous preparations were ob- tained by electroelution (Fig. 1B). Peptides obtained from trypsin digestion of the electroeluted protein were purified by HPLC and microsequenced. Four sequences were gen- erated (VRGEVAPDAK, FNAHGDANTIVCNSK, DGGAWGTEQR, VAFD) and compared to a protein data base using the FASTA program. Computer analysis re- vealed that the peptides (Fig. 2) were homologous to the previously described 14.5-kDa human soluble P-galacto- side binding L-14 lectin (27,33, 34) and exhibited a high degree of sequence similarity to 14-kDa vertebrate /3-ga- lactoside binding lectins from rat (35), chicken (36), bo- vine (37), and mouse (38) (Fig. 2). We therefore designated this protein as HLBP14 based on its homology to L-14 and its function as a human laminin binding protein. It is worth noting that there were also some sequence sim- ilarities between some HLBP14 peptides and the mouse carbohydrate binding protein 35 (39) and the leukocyte adhesion protein (40) (Fig. 2).
HLBP14 Is Eluted from Laminin Affinity Columns by Specific Sugars and Binds to a Human Laminin Afinity Column The sequence similarities between HLBP14 and P-ga-
lactoside-binding lectins led us to test whether these mol-
68-
43-
29-
18-
FIG. 1. Laminin-Affi-Gel chromatography of human melanoma A2058 cell extracts and electroelution of HLBP14. (A) A2058 cell extracts were incubated with laminin-Affi-Gel resin overnight as described under Materials and Methods, and laminin binding proteins were eluted with 10% acetic acid. Elution fractions were concentrated and electrophoresed on a 12.5% SDS-polyacrylamide gel. (B) After electroelution, the purity of HLBP14 was assessed on a 15% SDS-polyacrylamide gel. Molecular weight markers in kilodaltons are noted in the margins.
ecules bind to laminin through its ,&galactoside residues. Extracts of 35S metabolically labelled A2058 cells were incubated with laminin affinity columns and the ability of various sugars to elute the laminin-bound proteins was tested. As summarized in Table I, N-acetyllactosamine, lactose, and galactose eluted HLBP14, as well as the 31- and the 67-kDa molecules, but mannose, fucose, glucose, or melibiose did not. Table II shows that in the presence of 300 mM lactose or galactose, neither HLBP14 nor the 31- and 67-kDa molecules bound to a laminin affinity col- umn. In contrast, 300 mM mannose or melibiose could not block binding activity. HLBP14 was specifically eluted with lactose from a human laminin affinity column (Fig. 3), suggesting that murine and human laminin have some similar carbohydrate residues.
HLBP14, HLBP31, and the 67-kDa Laminin Binding Protein Bind to Specific Carbohydrate Sequences on Laminin
To determine the role of poly-N-acetyllactosamine carbohydrate sequences in the interaction between lam- inin and HLBP14, HLBP31, or the 67-kDa laminin re-
14-kDa GALACTOSIDE BINDING LECTIN BINDS TO LAMININ 135
LBPlZ (P36)
HHLlHPL
FILL BHL
CSL MlPL3
CBP35
LBPlZ IP49A)
HHL/HPL RLL BHL
CSL
MIPL3 CBP35
LAP
LBPl2 IP43) HHL/HPL
FILL
BHL
CSL
MIPL3
CBP35
LBPl2 (P49Bl
HHL/HPL
49 FNAHGDANTIVCNSK
49 49 51
51 176
51
64
66
66
65 65
190
131
29
29 29
29 30
156
63 63
63 63
65
65 189
65
73
75
75
74
74 199
COOH
FIG. 2. Comparison of the sequences of HLBP14 peptides. Electroe- luted HLBP14 (see Fig. 1B) was digested with trypsin and generated peptides were sequenced as described under Materials and Methods. Sequence similarities of HLBPl4 peptides (LBP12-P36, LBP12-P49A, LBP12-P43, and LBP12-P49B) are demonstrated with the 14-kDa /3- galactoside soluble lectins from man (HHL, human hepatoma lectin; HPL, human placenta lectin), rat (RLL, rat lung lectin), bovine (BHL, bovine heart lectin), chicken (CSL, chicken skin lectin), and mouse (MIPL3, murine fibrosarcoma lectin), as well as with the mouse car- bohydrate binding protein 35 (CBP 35) and the human leucocyte adhe- sion protein (LAP). Boxes enclose residues homologous with HLBPl4 peptides. The numbers to the left and the right of the sequences designate amino acid positions. (-) A gap introduced for optimal alignment.
ceptor, laminin coupled to Affi-Gel was digested by endo- @-galactosidase, a-galactosidase, and P-galactosidase. Extracts of radiolabeled A2058 cells were incubated with the enzymatically treated laminin-Affi-Gel columns, and the bound molecules were eluted after extensive washes with 100 mM lactose. The eluted proteins were concen- trated and separated on a 12.5% SDS-PAGE. As shown in Table III, digestion of laminin with endo-fl-galactosi- dase, an enzyme that disrupts linear Type 2 poly-N-ace- tyllactosamine at internal fil,4-linked galactosyl residues, decreased its ability to bind to the three laminin binding proteins (Table III). Treatment of laminin with fl-galac- tosidase or with a-galactosidase had no significant effect on HLBP14, HLBP31, or 67-kDa laminin receptor bind- ing (Table III). The lack of binding of HLBP14, HLBP31, or 67-kDa laminin receptor to endo+galactosidase-di- gested laminin together with the competition experiments suggests that poly-N-acetyllactosamine sequences play a specific role in that process. Furthermore, these laminin binding proteins did not bind to glycoproteins such as
TABLE I
Elution of Laminin Binding Proteins with Different Carbohydrates
Carbohydrate 67-kDa laminin
HLBP14 HLBP31 receptor
N-Acetyllactosamine + + + Lactose t + + Galactose t + t Glucose ND ND ND Fucose ND ND ND Melibiose ND ND ND Mannose ND ND ND
Note. Total cell extracts of metabolically radiolabeled A2058 cells were incubated with laminin coupled to Affi-Gel 10 resin as described under Materials and Methods. After extensive washes, laminin binding proteins were eluted with 100 mM solutions of lactose, N-acetyllactos- amine, glucose, galactose, melibiose, fucose, or mannose. The eluted fractions were concentrated on centricon 10 filters and separated on a 12.5% SDS-polyacrylamide gel. Eluted laminin binding proteins (HLBP14, HLBP31, and the 67-kDa laminin binding protein) were de- tected by autoradiography of the gel. f, protein detected; ND, protein not detected.
fibronectin or asialofetuin that do not contain poly-N- acetyllactosamine sequences (Table III).
Immunoblot Characterization of Anti-HLBP14 Antibodies
Gel-purified HLBP14 was used as antigen to generate polyclonal rabbit antisera. The immunoreactivity of anti- HLBP14 antisera was tested on melanoma cell laminin binding proteins that were eluted from a laminin affinity column with lactose, separated by electrophoresis on a 12.5% SDS-polyacrylamide gel and transferred to PDVF membrane as described under Materials and Methods
TABLE II
Competition of the Binding of Laminin Binding Proteins to Laminin-Affi-Gel with Specific Carbohydrates
Carbohydrate HLBP14 HLBP31 67-kDa laminin
receptor
Lactose + + + Galactose + + + Mannose ND ND ND Melibiose ND ND ND
Note. Total cell extracts of metabolically radiolabeled A2058 cells were incubated with laminin coupled to Affi-Gel 10 resin in the presence of 300 mM lactose, galactose, melibiose, or mannose as described under Materials and Methods. After extensive washes, laminin binding proteins were eluted with 100 mM solutions of lactose. The eluted fractions were concentrated on centricon 10 filters and separated on a 12.5% SDS- polyacrylamide gel. Eluted laminin binding proteins were detected by autoradiography of the gel. +, protein detected; ND, protein not detected.

136 CASTRONOVO ET AL.
FIG. 3. HLBP14 binds to human laminin. Radiolabeled human mel- anoma A2058 cell extracts were applied to either a human (A) or an EHS murine (B) laminin affinity column and HLBPl4 was eluted by lactose and analyzed on a 15% SDS-polyacrylamide gel. Molecular weight markers are noted in the left margin.
(Fig. 4). Anti-HLBP14 antisera recognized HLBP14 and also immunoreacted with HLBP31 and the 67-kDa lam- inin receptor (B). Control experiments in which pre- immune sera were used as the first antibody were nega- tive (A).
DISCUSSION
Laminin is a multidomain macromolecule that exhibits several important biological functions (2-4). Therefore, the finding that several distinct molecules on the surface of normal and malignant cells are capable of interacting with different domains of laminin is not surprising. Sev- eral cell surface proteins are capable of interacting with laminin (X5-21), but the function(s) mediated by each specific laminin binding protein is not clearly established. Furthermore, the laminin domain(s) implicated in the in- teractions with specific laminin binding proteins is un- known. Several studies have suggested that specific lam- inin amino acid sequences can mimic some biological
97-
67-
43-
29-
18-
12-
FIG. 4. Immunoblots of lactose-eluted laminin binding proteins with anti-HLBP14 antibodies. Human melanoma A2058 laminin binding proteins were eluted from a laminin column by lactose, separated on a 12.5% SDS-polyacrylamide gel, transferred to a PDVF membrane, and immunoblotted as described under Materials and Methods. The eluted proteins were run alongside prestained protein markers (Bethesda Re- search Laboratories). (A) Preimmune serum; (B) anti-HLBP14.
activities of laminin, including cell attachment and neurite outgrowth (6,16). There is also a growing body of evidence suggesting that carbohydrate structures present on lam- inin play a crucial role in mediating interactions between cells and laminin (7, 8, 41).
Laminin is a heavily glycosylated molecule. The dis- sacharide [3/3GalPl, 4GlcNAc$31],, known as the poly-N- acetyllactosamine sequence, is one of the most repeated sequences present in branched oligosaccharides on lam- inin. The exact functions of these sequences is not clear, but it has been reported that they do not play a role in heparin binding to laminin or in the resistance of laminin to proteases (42). Studies have suggested that they may play a role in cellular adhesion to laminin (11, 20, 8).
TABLE III
Binding of Laminin Binding Proteins to Glycosidase-Digested Laminin and to Fibronectin or Asialofetuin
Affinity column HLBP14 HLBP31
Untreated laminin + + Laminin digested with @-galactosidase + t Laminin digested with endo+galactosidase ND ND Laminin digested with alpha-galactosidase t + Fibronectin ND ND Asialofetuin ND ND
67-kDa laminin receptor
+ +
ND t
ND ND
Note. Laminin coupled to Affi-Gel was treated with endo-&galactosidase, a-galactosidase, or fi-galactosidase. Total cell extracts of metabolically radiolabeled A2058 cells were incubated with the digested laminin-Affi-Gel column or with fibronectin- or asialofetuin-Affi-Gel columns as described under Materials and Methods. After extensive washes, laminin binding proteins were eluted with 100 mM lactose. The eluted fractions were concentrated on centricon 10 filters and separated on a 12.5% SDS-polyacrylamide gel. Eluted laminin binding proteins were detected by autoradiography of the gel. +, protein detected, ND, protein not detected.

14-kDa GALACTOSIDE BINDING LECTIN BINDS TO LAMININ 137
In this study, we purified and characterized HLBP14, a 14-kDa laminin binding molecule, from human mela- noma A2058 cells. The peptides of HLBP14 exhibit ho- mology with the human soluble 14-kDa /3-galactoside binding protein (L-14) (37) and share conserved sequences with the 14-kDa vertebrate lectin from mouse (38), rat (35), bovine (27), and chicken (36), suggesting that this molecule is derived from a common ancestral gene con- served through evolution. The 14-kDa lectin family of proteins has been implicated in important biological events including development, differentiation, cellular regulation, and tissue construction (42) but the specific mechanisms for these processes are not understood. Our data suggest that these functions may be all or partially explained by the capacity of these proteins to bind spe- cifically to laminin. Homology between HLBP14 peptides and the mouse carbohydrate protein 35 (39) as well as the human leucocyte adhesion protein (40) indicates the presence of conserved sequences in many different adhe- sion proteins. This pattern of conservation may very well have functional significance. Recently, a study reported that the 14-kDa S-type lectin from calf heart tissue was able to selectively bind to the poly-N-acetyllactosamine sequences of laminin (26). The apparent affinity of the 14-kDa calf heart lectin for laminin was determined by Scatchard analysis to be 9.92 X 1O-7 M (26). This finding adds weight to our conclusion that HLBP14 may play an important role during the interaction of cancer cells with laminin. While we were preparing this manuscript, Cooper et al. (44) reported that laminin is the major glycoprotein ligand for the murine L-14 and that this lectin might play a critical role during differentiation and fusion of murine myoblasts into tubular myofibers by regulating their de- tachment from laminin.
Immunoblot experiments performed using affinity pu- rified antibody raised against a synthetic peptide deduced from the 67-kDa laminin receptor precursor cDNA se- quence (28) strongly suggested that the 67-kDa molecule eluted from the laminin affinity column with lactose was the 67-kDa laminin receptor. Interestingly, anti-HLBP14 polyclonal antibodies reacted in immunoblot experiments not only with HLBP14, but also with the 31-kDa molecule and the 67-kDa laminin receptor. This suggests that these three proteins are immunologically related and share common epitopes. The ability of lactose to elute a 67-kDa elastin receptor that is either identical to, or that shares common epitopes with, the 67-kDa laminin receptor fur- ther supports our data (45). Recently, the elastin receptor and the laminin receptor were found to immunostain with antibodies raised against the rat lung 14.5-kDa /3-galac- toside binding lectin (46). Furthermore, HLBP31 was found after purification and microsequencing to be iden- tical to the 31-kDa IgE binding protein and to be very similar to the 31 human galactoside binding lectin (47, manuscript in preparation). One common functional do- main may be involved in the lectin binding properties of
all these molecules. However, protein and cDNA se- quencing of these three proteins clearly establishes that they are unique and are the products of specific, different genes (24, 28).
The identification of HLBP14 as a protein able to bind to laminin through its poly-N-acetyllactosamine residues suggests a function for these sequences and offers new evidence for the importance of laminin carbohydrate structures in cell surface-laminin interactions. Our data strongly suggest that HLBP14, as well as HLBP31 and the 67-kDa laminin receptor, selectively and specifically interacts with the poly-N-acetyllactosamine sequences present on laminin rather than with the terminal /3-ga- lactoside residues. The significant reduction in the binding of HLBP14 to laminin observed when the latter was treated with endo-fi-galactosidase, but not with a-galac- tosidase or P-galactosidase, confirms the role of poly-N- acetyllactosamine sequences in these interactions. The characteristics of the interactions between HLBP14 and laminin are similar to those between the S-type lectin from calf heart and laminin (26). The identification of HLBP14 as a laminin binding protein produced by human cancer cells suggests that this lectin might play an im- portant role in the interaction between cancer cells and laminin during tumor invasion and metastasis. Major dif- ferences in laminin glycosylation exist between species. It was therefore necessary to demonstrate that HLBP14 purified from human cancer cells also binds to human laminin. Our observation that HLBP14 is eluted off a human laminin affinity column by lactose strongly sug- gests that human laminin also contains poly-N-acetyl- lactosamine sequences. Therefore, HLBP14 probably plays a role in the interactions between human laminin and human cells, in uiuo. As suggested by the role of L- 14 in murine myoblast differentiation (44), HLBP14 could regulate coordinate detachment of malignant cells from the basement membrane glycoprotein laminin, a phe- nomenon which is necessary prior to the migration of invasive cells. We are currently investigating this hy- pothesis to determine if HLBP14 could be potentially used as an inhibitor of cancer cell attachment to laminin and, as such, as a possible antitumor invasion molecule.
REFERENCES 1.
2.
3. 4. 5.
6.
7.
Timpl, R., Rhode, H., Gehron-Robey, S. R., Foidart, J. M., and Martin, G. R. (1979) J. Biol. Chem. 254,9933-9937. Kleinman, H. K., Cannon, F. B., Laurie, G. W., Hassell, J. R., Au- mailley, M., Terranova, V. P., and Martin, G. R. (1982) J. Cell Biochem. 27, 317-325. Liotta, L. A. (1986) Cancer Res. 46, 1-7. Beck, K., Hunter, I., and Engel, J. (1990) FASEB J. 4, 148-160. Barsky, S. H., Rao, C. N., Williams, J. E., and Liotta, L. A. (1984) J. Clin. Znuest. 74, 843-848. McCarthy, J. B., Skubitz, A. P. N., Palm, S. L., and Furcht, L. T. (1988) J. Nutl. Cancer. Inst. 80, 108-115. Dean, J. W., III, Chandrasekaran, S., and Tanzer, M. V. (1990) J. Biol. Chem. 265, 12553312562.

138 CASTRONOVO ET AL.
8.
9.
10.
11.
12.
13.
14.
15.
16. 17.
18.
19.
20.
21.
22.
23. 24.
25.
26.
27.
Dean, J. W., III, Chandrasekaran, S., and Tanzer, M. V. (1990) Biochem. Biophys. Res. Commun. 156,411-416.
Shibata, S., Peters, B. P., Roberts, D. D., Goldstein, I. J., and Liotta, L. A. (1982) FEB.9 Z&t 142, 194-198.
Rao, C. N., Goldstein, I. J., and Liotta, L. A. (1983) Arch. Biochem. Biophys. 227, 118-124.
Dennis, J. W., Waller, C. A., and Schirrmacher, V. (1984) J. Cell Biol. 99,1416-1423.
Arumugham, R. G., Hsieh, T. C. Y., Tanzer, M. L., and Laine, R. (1986) Biochim. Biophys. Acta 883, 112-126.
Fujiwara, S., Shinkai, H., Deutzmann, R., Paulsson, M., and Timpl, R. (1988) Biochem. J. 252,453-461.
Knibbs, R. N., Perini, F., and Goldstein, I. J. (1989) Biochemistry 28,6379-6392.
Rao, C. N., Barsky, S. H., Terranova, V. P., Liotta, L. A. (1983) Biochem. Biophys. Res. Commun. 111,804-808. Albelda, S. M., and Buck, C. A. (1990) FASEB J. 4,2868-2880.
Horwitz, A., Duggan, K., Greggs, R., Decker, C., andBuck, C. (1985) J. Cell Biol. 101, 2134-2144.
Gehlsen, K. R., Dillner, L., Engvall, E., and Ruoslahti, E. (1988) Science 241,1228-1229. Kleinman, H. K., Ogle, R. C., Cannon, F. B., Little, C. D., Sweeney, T. M., and Luckenbill-Edds, L. (1988) Proc. N&l. Acad. Sci. USA 83,8182-8186. Ruynan, R. B., Versalovic, J., and Shur, B. D. (1988) J. Cell Biol. 107, X363-1871.
Kramer, R. H., McDonald, K. A., and Vu, M. P. (1989) J. Biol. Chem. 15642-15649.
Lesot, H., Kuhl, U., and von der Mark, K. (1983) EMBO J. 3,861- 865.
Malinoff, H., and Wicha, M. S. (1983) J. Cell Biol. 96, 1475-1480. Woo, H. J., Shaw, L. A., Messier, J. M., and Mercurio, A. M. (1990) J. Biol. Chem. 265,7097-7099.
Cherayil, B. Y., Chaitovitz, S. A., Wong, C., and Pillai, S. (1990) Proc. N&l. Acad. Sci. USA 87, 7324-7338.
Zhou, Q., and Cummings, R. D. (1990) Arch. Biochem. Biophys. 281,27-35. Abbott, W. M., Hounsell, E. F., and Feizi, T. (1988) Biochem. J. 252,283-287.
28.
29.
30. 31.
32.
33.
34. 35.
36.
37. Abbott, W. M., Mellor, A., Edwards, Y., and Feizi, T. (1989) Biochem. J. 359,283-290.
38. 39. 40.
41.
42. 43. 44.
Raz, A., and Lotan, R. (1987) Cancer Metastasis Rev. 6,433-452. Jia, S., and Wang, J. L. (1988) J. Btil. Chem. 263,6009-6011. Law, S. K. A., Gagnon, J., Hildreth, J. E., Wells, C. E., Willes, A. C., and Wong, A. J. (1987) EMBO J. 6,915-919. Castronovo, V., Colin, C., Parent, B., Foidart, J. M., Lambotte, R., and Mahieu, P. (1989) J. Natl. Cancer Inst. 81, 212-216. Howe, C. C. (1984) Mol. Cell Biol. 4, l-7. Barondes, S. H. (1984) Science 223, 1259-1264. Cooper, D. N. W., Massa, S. M., and Barondes, S. H. (1991) J. Cell Biol. 115, 1437-1448.
45.
46.
Mecham, R. P., Hinek, A., Griffin, G. L., Senior, R. M., and Liotta, L. A. (1989) J. Biol. Chem. 264,16652-16657. Hinek, A., Wrenn, D. S., Mecham, R. P., and Barondes, S. H. (1988) Science, 239, 1539-1541.
47. Raz, A., Pazerini, G., Carmi, P. (1990) Cancer Res. 49,3489-3493.
Rao, C. N., Castronovo, V., Schmitt, M. C., Wewer, U. M., Claysmith, A. P., Liotta, L. A., and Sobel, M. E. (1989) Biochemistry 28,7476- 7486. Wewer, U. M., Liotta, L. A., Jaye, M., Ricca, G. A., Drohan, W. N. Claysmith, A. P., Rao, C. N., Wirth, P., Coligan, J. E., Albrechtsen, R., Mudryj, M., and Sobel, M. E. (1986) Proc. Natl. Acad. Sci. USA 83, 7137-7141. Laemmli, U. K. (1970) Nature 227,680-685. Luyten, F. P., Cunningham, N. S., Ma, S., Muthukumaran, N., Hammonds, R. G., Nevins, W. B., Wood, W. I., and Reddi, A. H. (1989) J. Biol. Chem. 264, 13377-13380. Stone, K. L., LoPresti, M. B., Crawford, J. M., DeAngelis, R., and Williams, K. R. (1989) in A Practical Guide to Protein and Peptide Purification for Microsequencing (Matsudaire, P. T., Ed.), pp. 32- 47, Academic Press, San Diego. Gitt, M. A., and Barondes, S. H. (1986) Proc. Natl. Acad. Sci. USA 83, 7603-7607. Abbott, W. M., and Feizi, T. (1989) Biochem. J. 259, 291-294. Clerch, L. B., Whitney, P., Hass, M., Brew, K., Miller, T., Werner, R., and Massaro, D. (1988) Biochemistry 27, 692-699. Ohyama, Y., Hirabayashi, J., Oda, Y., Ohno, S. S., Kawasaki, H., Suzuki, K., and Kasai, K. (1986) Biochem. Biophys. Res. Commun. 134,51-56.