identification of enzymes and toxins in venoms of indian ... of some of the enzymes such as...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEM~TRY Vol. 236, No. 7, July 1961
Printed in U.S.A.
Identification of Enzymes and Toxins in Venoms of
Indian Cobra and Russell’s Viper after Starch Gel Electrophoresis
R. W. P. MASTER AND S. SRINIVASA RAO
From the Department of Immunology, Hafkine Institute, Bombay lW, India
(Received for publication, January 17, 1961)
Snake venoms are known to contain a number of enzymes (l-3). Attempts have been made to correlate the toxicity of snake venoms with their enzymic activity, e.g. between nucleases and toxicity (4), phospholipase A and neurotoxic action of cobra venom (5), and proteolytic and coagulant activity and toxicity (6-8). Most of these studies have been carried out on whole venoms or after elimination of some components by heat treat- ment (5). A number of methods, such as electrophoresis (9), paper electrophoresis (lo), and quite recently, ion exchange resins such as Amberlite IRC 50 (11) and cellulose ion-exchangers (12-14) have been used for fractionation. Ohsaka (15) has used electrophoresis on potato starch for separation of components of Habu venom, and Yang et al. (16, 17) have used a similar method for the fractionation of the venoms of Formosan cobra and Hyap- poda. Yang et al. (17) found that the neurotoxic activity of cobra venom did not coincide with any of the enzymes tested by them. In the case of Hyappoda venom, they found that pro- teases and phosphatases occurred in the same fractions as the toxic components.
We have studied the properties of venoms of Indian snakes, especially Indian cobra (Baja Izaja) and Russell’s viper (Vipera russelli). On a microscale we are able to obtain a fairly good separation of the components of both the venoms with the starch gel electrophoretic technique of Smithies (18). Use of paper and agar gel electrophoresis did not give sufficiently good separa- tion of the components. We have worked out rapid methods of detection of the enzymic components with only small quantities of venoms. The present communication is a report of the re- sults.
EXPERIMENTAL PROCEDURE
Materials and Methods-The pooled lyophilized venoms of cobra and Russell’s viper collected at the Haffkine Institute were used.
Potato starch for electrophoresis was hydrolyzed before use as described by Smithies (18).
Tris and L-histidine hydrochloride were purchased from Nu- tritional Biochemicals Corporation, Cleveland, Ohio. Calcium di-p-nitrophenyl phosphate was purchased from Sigma Chemical Company. Disodium p-nitrophenyl phosphate, and p-nitro- phenyl-UMP (barium salt) were generously supplied by Dr. H. G. Khorana, British Columbia University, Canada. AMP was a product of Pabst Laboratories. RNA (sodium salt) was purchased from Mann Research Laboratories, Inc., New York, and DNA (sodium salt) from L. Light and Company, (England).
Esters of 2,6-dichlorophenol indophenol were prepared according to the method described previously (19).
Rabbit cells were prepared by washing defibrinated rabbit blood several times with titrated 0.9% sodium chloride solution. Fresh human plasma was received each time for coagulation studies.
Starch Gel Electrophoresis-This was carried out according to the method of Smithies (18). Potato starch gel, 12% was pre- pared in Tris-citrate buffer, pH 8.6. The composition of the buffer solutions was the same as that recommended by Poulik (20) except that the pH of the borate buffer in the electrode compartments was adjusted to 8.6 with 20% sodium hydroxide solution. Four starch blocks (25 x 2.5 x 0.2 cm) were cast over cellophane paper on a glass plate. The ends were con- nected to the electrode compartments with filter paper strips. A 20% (weight per volume) solution of lyophilized cobra and Russell’s viper venoms in distilled water was used for electro- phoresis. The venom sample was put on the starch gel by first placing approximately 0.025 ml on a strip of Whatman No. 3 filter paper (0.2 X 2 cm). This was the procedure adopted when the gel was to be tested directly for the location of the enzyme by overlaying a suitable substrate on the starch gel. When the components were to be eluted from narrow segments after the electrophoresis, the above procedure was not suitable. In such cases, 0.025 ml of the venom was adsorbed on starch grains which were then inserted into a narrow furrow in the center of the starch gel according to the procedure of Smithies (18). Elec- trophoresis was carried out at approximately 200 volts and 19 ma for 5 to 6 hours. After electrophoresis, one of the strips was stained for proteins with Amidoblack 1OB (0.5% solution in a mixture of methanol-acetic acid-water in the ratio of 5 : 1:4) for 5 minutes and the excess of the dye was removed by repeated washing with 1% aqueous acetic acid. The remaining blocks were tested for enzymes as described below. When the separated components were to be eluted, the starch blocks were cut into IO-mm segments and marked as fl, $2, $3 etc., from the point of application towards the anode side and -1, -2, -3 etc., from the point of application towards the cathode side. These segments were kept frozen at -20” overnight; the next day, after thawing, they were put inside the barrel of a 20-ml syringe with the needle removed and pressed with the piston. Approximately 0.4 ml of fluid was obtained from each segment by this method, an amount quite sufficient for testing for en- zymes and toxicity. These experiments were repeated several times to confirm the identity of different components. For the
1986
by guest on Novem
ber 7, 2017http://w
ww
.jbc.org/D
ownloaded from

detection of some of the enzymes such as nuclease, phospho- diesterase, and cholinesterase, in which chromogenic substrates could be used, the starch gel after electrophoresis was covered with the substrate and the enzymes could be visualized and located directly by the color produced.
Enzyme Activities
Protease-Fluid from each segment, 0.1 ml, was placed on un- developed photographic film strips, kept in a covered Petri dish, and incubated at 37” in a humid incubator overnight. When the strips were flooded with water, the zones of the proteolytic activity were revealed by dissolution of the gelatin on the photo- graphic strip.
Coagulase-Fluid from each segment, 0.1 ml, was tested for coagulase activity by Quick’s method of prothrombin time meas- urement with recalcified human plasma (21). Anticoagulant activity was detected by the same procedure.
Phospholipase A-The method used was that of Gennaro and Ramsey (22) with slight modification. Numbered disks (1.0 cm in diameter) of Whatman No. 1 filter paper were soaked in the fluid expressed from the frozen and thawed segments and placed on sheep cell lecithin agar in Petri plates. The sheep cell lecithin agar was prepared by adding a mixture of 2.5 ml of 0.2 ‘+$ lecithin FIG. 2. Detection of L-amino acid oxidase in Russell’s viper solution in 0.9% sodium chloride solution and 2.5 ml of packed venom, by circular paper chromatography after electrophoresis.
sheep cells to 45 ml of 1% agar prepared in normal 0.9 “1” sodium chloride solution which had been melted and cooled to 40”. Each Phosphodiesterase, Monoesterase, and 5’-Nucleotidase-Phos-
Petri dish contained 10 ml of this mixture. The plates were phodiesterase activity was detected by the method of Sinsheimer
incubated overnight at 37”. Phospholipase A activity was and Koerncr (23) with calcium dinitrophenyl phosphate as the
located by the appearance of a zone of hemolysis around the substrate. It was further confirmed by covering the starch
filter paper disks. block with 0.1 y0 solution of p-nitrophenyl-UMP and incubating
Nucleases-A solution containing RNA (sodium salt) or DNA it at 25” in a water-saturated atmosphere. The diesterase ac-
(sodium salt) (3 g/100 ml), and agar (1 g/100 ml) was overlaid tivity was revealed by the appearance of an intense yellow zone
on the starch block after electrophoresis. The block was in- against a white background in about 10 minutes.
cubated overnight at 37” in a humid incubator after which it was Monoesterase activity was detected by incubating 0.4 ml of
flooded with 0.5 M hydrochloric acid. The nuclease activity was the fluid from each segment with 0.4 ml of 0.5% solution of
revealed by the appearance of a clear zone against a white pre- sodium p-nitrophenyl phosphate. The appearance of yellow
cipitate when placed on a black background (Fig. 1). Alter- p-nitrophenol after approximately 2 hours indicated the presence natively, Whatman No. 1 filter paper disks (1 cm in diameter) of the enzyme.
soaked in the fluid from each segment were kept on the surface 5’-Nucleotidasc activity was detected by incubating 0.4 ml of
of 1 To agar gel containing 3 y. RNA. The plates were incubated the fluid from each segment with AMP as the substrate and
at 37” overnight. The next day the filter paper strips were estimation of inorganic phosphate was carried out by the method
removed and the plates were flooded with 0.5 M HCl. The of Fiske and SubbaRow (24).
nuclease activity could be easily detected by clear spots where L-Amino Acid Oxidase-To 0.2 ml of the fluid from each seg-
the filter paper disk containing the nuclcascs had been kept. ment was added 0.5 ml of a solution of L-histidine hydrochloride (3 mg per ml). After incubation at 37” for 24 hours, 10 ml
+ aliquots were applied to Whatman No. 1 filter paper circles (24 cm in diameter). Circular paper chromatography was carried out with n-butanol-acetic acid-water in the ratio of 4 : 1: 5
as t.he solvent and diazotized sulfanilic acid as the spray reagent (25). The appearance of an additional band above that of L-
histidine revealed the presence of L-amino acid oxidase (Fig. 2). Cholinesterase-The method of Master (19) was followed.
The starch block after electrophoresis was covered by a mixture of 0.2 ml of 2,6-dichlorophenolindophenol acetate, butyrate, or caproatc in ethanol and 1.8 ml of Tris-citrate buffer, pH 8.6. The block was left at 25” in a water-saturated cabinet. The esterase was detected by the appearance of a blue band of 2,6- dichlorophenolindophenol on the starch block (Fig. 3) in about
July 1961 R. W. I’. Master and X. S. Rae
FIG. 1. IIetcct,ion of ribonuclense in the cobra venom. Upper 1 hour. Unsubstituted indophenol esters give equally good
Skip strained for proteins with Amidoblack 1OB and lower Strip results, and can be prepared by the method described by Kramer treated with ribonucleic acid as described in the text. et al. (26).
by guest on Novem
ber 7, 2017http://w
ww
.jbc.org/D
ownloaded from
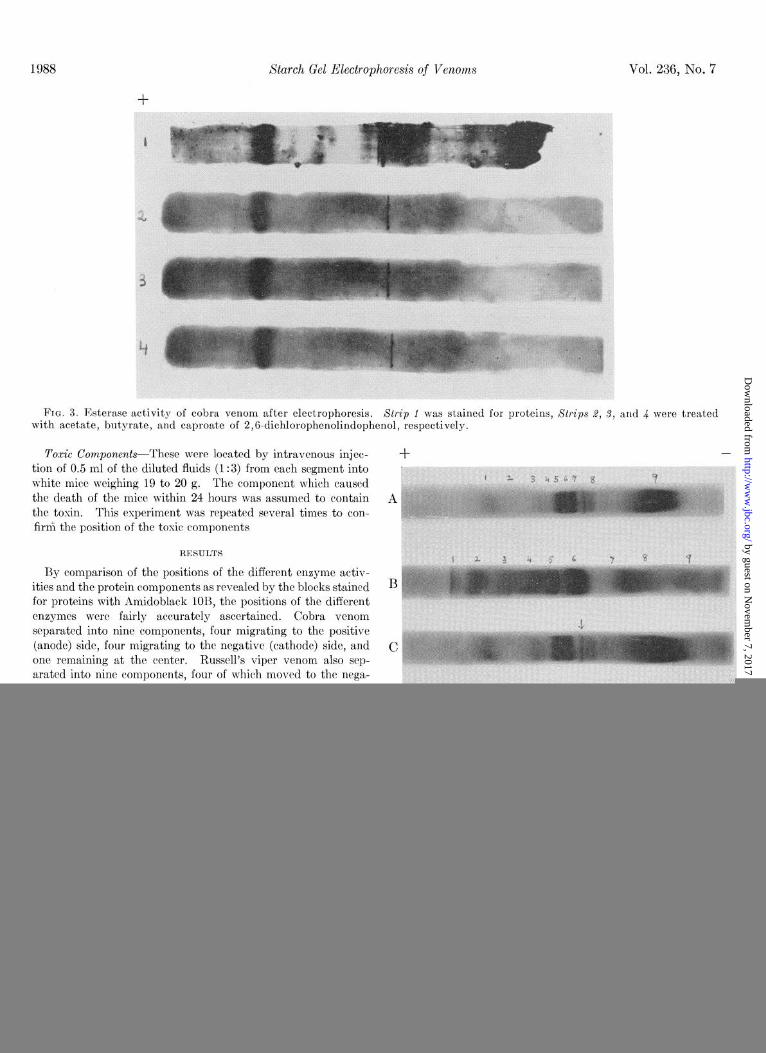
1988 Starch Gel Electrophoresis of Venoms Vol. 236, No. 7
FIG. 3. Esterase activity of cobra venom after electrophoresis. Xtrip 1 was stained for proteins, Strips 2, 3, and 4 were treated with acetate, butyrate, and caproate of 2,6-dichlorophenolindophenol, respectively.
Toxic Components-These were located by intravenous injec- tion of 0.5 ml of the diluted fluids (1:3) from each segment into white mice weighing 19 to 20 g. The component which caused
the death of the mice within 24 hours was assumed to contain the toxin. This experiment was repeated several times to con- firm the position of the toxic components
RESULTS
By comparison of the positions of the different enzyme activ- ities and the protein components as revealed by the blocks stained for proteins with Amidoblack IOB, the positions of the different enzymes were fairly accurately ascertained. Cobra venom separated into nine components, four migrating to the positive (anode) side, four migrating to the negative (cathode) side, and one remaining at the center. Russell’s viper venom also sep- arated into nine components, four of which moved to the nega- tive side and five to the positive side. The identification of the components of cobra and Russell’s viper venoms after starch gel electrophoresis is given in the legend to Fig. 4.
In both the venoms, it was found that phosphodiesterase and nuclease remained at the point of application and that 5’-nucle- otidase moved to the negative side. In Russell’s viper venom one component on the negative side also showed slight L-ammo acid oxidase activity.
When the toxicity of the fluids from all the segments after the electrophoresis of cobra venom was tested, it was found that the component in the third segment on the cathode side, correspond- ing to No. 8 in Fig. 4, killed the mice with the same typical symptoms of paralysis as the whole venom produced when in- j ected intravenously.
During electrophoresis of cobra venom, there occurred a pe- culiar sharp line at the position indicated by the arrow in Fig. 4 (Strip C) which was visible even when the starch block was not stained for proteins. The component adjacent to this line on the negative side was identified as the neurotosin. When the whole venom was heated to 100” for 15 minutes, centrifuged, and subjected to electrophoresis, this zone was present as the
D
FIG. 4. Starch gel electrophoresis of venoms of cobra and Rus- sell’s viper. (Strips A and 6, cobra venom; Strips B and D, Russell’s viper venom) stained with Amidoblack 1OB.
Cobra venom (Strip A) starting from the extreme component on the positive (anode) side: 1, choline&erase (also slight phos- phomonoesterase activity) ; 8, (not clear in the photograph) phos- pholipase A and anticoagulant; 3, L-amino acid oxidase; 4, prote- ase; 5, phosphodiesterase (ribonuclease and deoxyribonuclease); 6,5’-nucleotidase, 7, protease; 8, neurotoxin; and 9, phosphomono- esterase.
Russell’s viper venom (StGp B) : 1, unidentified; 2, phospho- lipase A; 3, protease; 4, L-amino acid oxidase and coagulase; 5, phosphodiesterase (ribonuclease and deoxyribonuclease) ; 6, 5’- nucleotidase; 7, r,-aminoacid oxidase, and& phosphomonoesterase.
only protein that resisted heat treatment. The peculiar sharp line made it possible to elute the component even without a reference block stained for proteins. The ncurotoxic component did not coincide with any of the enzymes detected in the present
by guest on Novem
ber 7, 2017http://w
ww
.jbc.org/D
ownloaded from

July 1961 R. W. P. Master and X. X. Rao
investigation. The protein responsible for the sharp line has not been identified.
In the case of Russell’s viper venom, interesting results were produced when the fluids obtained from each segment were tested for toxicity. About 0.4 ml of the fluids from each seg- ment was diluted to 1.0 ml, and 0.5 ml of this diluted solut.ion was injected intravenously into white mice weighing 19 to 20 g. This experiment was repeated several times. The expressed fluid from each of the segments was nontoxic although the amount of the Russell’s viper venom used for electrophoresis was approxi- mately 60 lethal doses. The fluids obtained from the segments on the positive side were pooled together into one lot and those from the negative side into another. The pooled fraction from the anode side was toxic and all the animals receiving injections died. The first five fractions on the positive side when com- bined, produced the toxic effect of the whole venom. These ex- periments were repeated four times and the same results were obtained. It was further confirmed that the aliquots of the very fractions, which were found to be nontoxic individually, became toxic when combined, in spite of the dilution. In the case of cobra venom, the fractions on the anode side which were also nontoxic individually did not become toxic when combined.
The toxicity experiments on Russell’s viper venom are sum- marized in Table I.
It was not possible to identify with certainty the components which should be mixed in order to restore the toxicity of the separated components because the amounts of fluids obtained mere very small. Further, a quantitative evaluation of the re- covery of the over-all toxicity after electrophoresis was not made. These venoms are now being fractionated on ion exchange cel- lulose columns.
DISCUSSION
Severai workers have tried to correlate the toxicity of snake venoms with their enzyme activities. Thus, the experiments of Taborda et al. (4) suggested that the nucleases might be responsi- ble for the toxic nature of the snake venoms. Braganca and Quastel (5) thought that phospholipase A of Indian cobra venom was responsible for the venom’s neurotoxic action inasmuch as both enzymes were stable to heat. However, these workers did not try to separate the two activities. In the present investiga- tion, the toxicity of cobra venom could not be attributed to any particular enzyme tested and certainly not to nucleases or phos- pholipase A. The toxic component which gave rise to typical paralytic symptoms of a neurotoxin was found to be electro- phoretically quite distinct from the other components. These results further support the results of previous workers (27, 28) who were able to separate phospholipase A from the neurotoxin in snake venoms. Phosphohpase A has been isolated by North and Doery (29) from snake venoms and used to neutralize staph- ylococcal toxin in vivo. Gitter et al. (10) have also separated the neurotoxin of Vipera xanthina palastine and found it to be distinct from the phospholipase A.
In the case of Russell’s viper venom, the components on the anode side eluted from segments 1 to 5 from the center seemed collectively to contribute to the toxicity of the venom. The enzymes present in them were coagulase, L-amino acid oxidase, protease, and phospholipase A; this is a good indication that some of the enzymes do enhance the action of other enzymes although by themselves they do not manifest signs of toxicity. Recently, Kochwa et al. (14) reported similar synergistic action
TABLE I Toxicity experiments on mice with pooled fractions after
electrophoresis of Russell’s viper venom
Exper;yent No. of fractions pooled No. of mice
surviving after 24 hrs.
Anode (+I to +lO) Cathode (-1 to -10) Anode (fl to +5) Anode (+6 to +10)
ml
1.0 o/4 1.0 4/4 0.5 o/3 0.5 3/3
* All injections were intravenous.
of two components separated by ion exchange cellulose columns from V. zanthina palastine venom which became more toxic when mixed. In the case of Russell’s viper venom, the toxicity has been attributed to the proteolytic enzymes (8). In the present study, on mixing the fractions on the anode side which contain coagulase, protease, n-amino acid oxidase etc., the tox- icity is restored. The effect of protease may be potentiated by the L-amino acid oxidase inasmuch as it removes the product of action of protease and perhaps of coagulase, as suggested by Zeller (1). Thus, the toxicity of Russell’s viper venom may be due to the combined action of protease, coagulase, and n-amino acid oxidase. The restoration of the toxicity on mixing the frac- tions could also be due to the recombination of an apoenzyme with its cofactor, which might have been separated during elec- trophoresis. The cofactor if present, is, however, nondialyzable, because dialyzed Russell’s viper venom has been found by us to have the same toxicity as the undialyzed venom.
The present investigation has revealed that in electrophoretic properties the 5’-nucleotidase is distinct from the diesterase, and occupies a position immediately on the negative side from the center in both the venoms.
The cholinesterase of cobra venom moves to the anode side which corresponds to the fastest component. This enzyme is absent in Russell’s viper venom. It hydrolyzes acetyl ester and also acts on higher esters such as butyrate and caproate (19). Hence, cholinesterase of cobra venom has a wider specificity. It has been found to be distinct from the neurotoxic component as reported by earlier workers (1).
Another interesting observation is that the electrophoretic mobilities of some enzymes such as phosphodiesterase, 5’-nucle- otidase, L-amino acid oxidase, protease, phospholipase A of cobra, and Russell’s viper venoms are more or less similar. The anode fractions of cobra and Russell’s viper venoms are similar except for a strong coagulase and L-amino acid oxidase activity in the Russell’s viper venom. However, when the anode fractions of cobra venom are mixed, they do not become toxic although this venom has a cholinesterase in addition. The toxicity of Rus- sell’s viper venom is therefore probably due to its higher content of protease and L-amino acid oxidase. The different modes of toxicity as evidenced by the present investigation throw some further light on their form of action. The cathode fractions of Russell’s viper venom are individually nontoxic and do not become toxic even when pooled together.
Although some of the common components of cobra and Rus- sell’s viper venoms have the same electrophoretic mobilities, they must be different immunologically because they do not cross react in the gel diffusion test, as observed by Kulkarni and Rao
(30).
by guest on Novem
ber 7, 2017http://w
ww
.jbc.org/D
ownloaded from

1990 Starch Gel Electrophoresis of Venoms Vol. 236, No. 7
SUMMARY
1. Venoms of cobra and Russell’s viper were subjected to starch gel electrophoresis and several different enzymic and toxic components were identified.
2. Methods are described for the detection of enzymes such as esterases, nucleases, n-amino acid oxidase, and phosphatases, after starch gel electrophoresis.
3. In the cobra venom, one component migrating to the cath- ode side was found to be responsible for the neurotoxic action of the venom. It did not coincide with any of the enzymes de- tected in the present investigation.
4. In the case of Russell’s viper venom, the individual com- ponents were found to be nontoxic after electrophoresis but, on mixing the fractions on the anode side, toxicity was restored.
Acknowledgment-The authors are grateful to Dr. A. K. Hazra,
Assistant Director, Haffkine Institute, and Dr. H. I. Jhala, Director, Haffkine Institute for their encouragement. One of us (R. W. P. Master) is grateful to the National Institute of Sciences of India and to the Government of India for the award of Fellowship.
REFERENCES
1. ZELLER, E. A., in F. F. NORD (Editor), Advances in enzymology, Vol. 8, Interscience Publishers, New York, 1948, p. 459.
2. BRAGANCA, B. M., Neurochemistry, Charles C Thomas, Spring- field, Ill., 1955, p. 623.
3. PORGES, N., Science, 117, 47 (1953). 4. TABORDA, A. R., TABORDA L. C., WILLIAMS, J. N., AND ELVE-
HJEM, C. A. J. Biol. Chem., 194, 227, (1952). 5. BRAGANCA, B. M., AND QUASTEL, J. H., Nature (London), 169,
695 (1952); Biochem. J., 63, 88 (1953). 6. KELLAWAY, C. H., in J. M. LUCK (Editor). Annual Review
of Biochemistry, Vol. 8, Annual Reviews, Inc., Stanford, Calif., 1939, p. 541.
7.
8.
9.
10.
11. 12.
13.
14.
15. OHSAKA, A., J. Biochem. (Tokyo), 45, %9, (1958): 16. YANG, C.. CHEN. C.. AND Su. C.. J. Biochem. ~Tokuo). 46.
17.
18. 19. 20. 21.
22.
23.
24. 25. 26.
27. 28.
29.
30.
DEUTSCH, H. F., AND DINIZ, C. R., J. Biol. Chem. 216, 17 (1955).
RAO, SHANTA, S., AND RAO, S. SRINIVASA, J. Sci. Ind. Research (India), 16C, 148 (1957).
GRASSET, E., BRECHBUHLER, T., SCHWARTZ, D. E., AND PONG- RATZ, E., Venoms, publication no. & of American Association Advancement of Science, Washington D. C. 1956. p. 153.
GITTER, S., KOCHWA, A., DE VRIES A., AND LEFFKOWITZ, M., Am. J. Trop. Med. Hyg., 6, 180 (1957).
DOERY H. M., Biochem. J., 70, 435 (1958). WALTER, B. AND BOMAN, H. G., Biochim. et Biophys. Acta., 34,
503 (1959). CARREY, J. E., AND WRIGHT, E. A., Nature (London), 186, 103
(1960). KOCHWA, S., PERLMUTTER, C., GITTER, S., RECHNIC, J., AND
DE VRIES, A., Am. J. Trap. Med. Hyg., 9, 374 (1960).
1201 (1959). ’ . “.I
YANG, C., Su, C., AND CHEN, C., J. Biochem. (Tokyo), 46, 1209 (1959).
SMITHIES, O., Biochem. J. 61, 629 (1955). MASTER, .R. %. P., Biochim. et Biophys. Acta, 39, 159 (1960). POULIK. M. D.. Nature (London). 180. 1477 (1957). PAGE, k. C., AND RUSSELL, H. I??:, J. Lab. Cl&. Med., 26, 1366
(1941). GENNARO, J. F., AND RAMSEY, H. W., Am. J. Trop. Med. Hyg.,
8, 546 (1959). SINSHEIMER, R. L., AND KOERNER, J. F. J. Biol. Chem. 198,
293 (1952). FISKE, C. H., AND SUBBAROW, Y., J. Biol. Chem., 66,375 (1925). MANN, T., AND LEONE, E., Biochem. J., 63, 140 (1953). KRAMER, D. N., GAMSON, R. M., AND MILLER, F. M. J. Org.
Chem., 24, 1742 (1959). BUSSARD A., AND COT& R., Compt. rend., 239, 915 (1954). NEUMANN, W. P., AND HABERMANN, E., Biochem. Z., 327, 170
(1955). NORTH, E. A., AND DOERY, H. M., Brit. J. Exptl. Path., XLI,
234 (1960). KULKARNI, M. E., AND RAO S. S., Venoms, publication no. 44
of the American Association Advancement of Science, Washing- ton, D. C., 1956, p. 175.
by guest on Novem
ber 7, 2017http://w
ww
.jbc.org/D
ownloaded from

R. W. P. Master and S. Srinivasa RaoViper after Starch Gel Electrophoresis
Identification of Enzymes and Toxins in Venoms of Indian Cobra and Russell's
1961, 236:1986-1990.J. Biol. Chem.
http://www.jbc.org/content/236/7/1986.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/236/7/1986.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on Novem
ber 7, 2017http://w
ww
.jbc.org/D
ownloaded from