igy antibodies anti-tityus caripitensis venom: purification and neutralization efficacy
TRANSCRIPT

ilable at ScienceDirect
Toxicon 74 (2013) 208–214
Contents lists ava
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
IgY antibodies anti-Tityus caripitensis venom: Purification andneutralization efficacy
Aurora Alvarez a, Yuyibeth Montero a, Eucarys Jimenez a, Noraida Zerpa a,Pedro Parrilla b, Caridad Malavé a,*
aCentro de Biociencias y Medicina Molecular, Instituto de Estudios Avanzados-IDEA, Apartado 17606, Caracas 1015-A, Venezuelab Laboratorio de Farmacología, Universidad de Oriente, Ciudad Bolívar, Venezuela
a r t i c l e i n f o
Article history:Received 5 November 2012Received in revised form 10 August 2013Accepted 15 August 2013Available online 29 August 2013
Keywords:Tityus caripitensisAnti-venomIgY antibodiesEgg yolk antibodies
* Corresponding author. Tel.: þ58 212 9035126; faE-mail addresses: [email protected], cmal
Malavé).
0041-0101/$ – see front matter � 2013 Elsevier Ltdhttp://dx.doi.org/10.1016/j.toxicon.2013.08.058
a b s t r a c t
Tityus caripitensis is responsible for most of scorpion stings related to human incidents inNortheastern Venezuela. The only treatment for scorpion envenomation is immuno-therapy based on administration of scorpion anti-venom produced in horses. Avian anti-bodies (IgY) isolated from chicken egg yolks represent a new alternative to be applied asanti-venom therapy. For this reason, we produced IgY antibodies against T. caripitensisscorpion venom and evaluated its neutralizing capacity. The anti-scorpion venom anti-bodies were purified by precipitation techniques with polyethylene glycol and evaluatedby Multiple Antigen Blot Assay (MABA), an indirect ELISA, and Western blot assays. Thelethality neutralization was evaluated by preincubating the venom together with the anti-venom prior to testing. The IgY immunoreactivity was demonstrated by a dose-dependentinhibition in Western blot assays where antibodies pre-absorbed with the venom did notrecognize the venom proteins from T. caripitensis. The anti-venom was effective inneutralizing 2LD50 doses of T. caripitensis venom (97.8 mg of IgY neutralized 1 mg of T.caripitensis venom). Our results support the future use of avian anti-scorpion venom as analternative to conventional equine anti-venom therapy in our country.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Scorpion envenomation is an important health problemin tropical and subtropical countries around the world.Venezuela is a countrywith a high incidence of scorpionism,mainlydue to stings by thegenus Tityus (Borges,1996).Tityuscaripitensis (T. caripitensis) is responsible for most accidentsby scorpionism in the Northeastern region of the country,being theMonagas state the regionwith the highest numberof cases in Venezuela (Borges and De Sousa, 2006). Scorpionenvenomation, especially in children, is often a severe clin-ical condition and is potentially lethal. This is characterizedby a systemic response that includes: hypertension and
x: þ58 212 [email protected] (C.
. All rights reserved.
hypotension, tachycardia and tachypnea, hypothermia,consciousness disturbances, seizures, dyspnea, miosis ormydriasis, heart failure, abdominal distension, pulmonaryedema, coma, convulsion and death (De Sousa et al., 2005).
The scorpion venoms are a complex mixture of neuro-toxic peptides, inhibitory enzymes, hyaluronidase, seroto-nin, histamine, nucleotides and lipids with variousphysiological and pharmacological lethally effects (Gweeet al., 2002; Müller, 1993). Venezuela produces scorpionanti-venom in horses which has proven to be an efficienttreatment to prevent the manifestation of envenomation(Poggioli De Scannone, 1996). However, the production ofthis anti-venom is very expensive and in some cases couldgenerate side effects in humans (Maya Devi et al., 2002;Karlson-Stiber and Persson, 1994).
The use of chicken antibodies (IgY) has increasedsignificantly in recent years, mainly because producing

A. Alvarez et al. / Toxicon 74 (2013) 208–214 209
antibodies in eggs is both cost effective and minimizes an-imal welfare concerns (Chacana et al., 2004). This method-ology has several advantages and follows “The principles ofHuman Experimental Technique”which are summarized inthree words: reduce, replace and refine (Russell and Burch,1959). Additionally, IgY isolated from chicken egg yolks of-fers the advantage of no complement cascade activation,removing one potential source of the reaction, and induceshigh titers of neutralizing antibodies (Schade et al., 2005). Atpresent, because of themanyadvantagesof IgY, it representsa newalternative to traditional anti-venomserum therapiesdeveloped in horses (Table 1). In this work, we producedIgY against T. caripitensis scorpion venom and evaluatedits neutralizing capacity.
2. Materials and methods
2.1. Venom
Adult T. caripitensis species scorpions were collected atnight around the villages of El Limon, Los Baños and theCruz Negra fromMonagas State, Venezuela, using UV lampsthat allowed distinguishing the specimens by their fluo-rescence (De Sousa et al., 2006). Scorpions were trans-ported live to the “Laboratorio de Alacranología”, Escuelade Ciencias de la Salud, Núcleo de Bolívar, Universidad deOriente, and maintained with water and fed with Tenebriomolitor larvae. Venom was extracted by electrical stimula-tion of the telson. Venom samples pooled from 30 to 40individuals were lyophilized and stored at�20 �C until use.
2.2. Animals
Sixteen-weeks-old Isabrown-laying hens (2 kg bodymass) were used for the production of anti-venom (IgY).C57BL/6 (18–20 g) mice were used to determine venomlethality and lethality neutralization. Animals were bred andmaintained at the animal facility from the Instituto de Estu-dios Avanzados and care according to the information con-tained in the guidelines issued by the European Communityfor the Use of Experimental Animals (L358-86/609/EEC).
2.3. Hens immunization
Twohenswere immunized intramuscularly in the breastregion at multiple sites with 50 mg of the T. caripitensis
Table 1Comparison of production of anti-venoms in horses and chickens.
Comparison items Horse anti-venoms IgG Chic
Time sustained antibodyproduction
From 8 to 10 weeks From
Costs of production Involves complexes and expensiveseveral steps
Invoinex
Cost of animal maintenance Expensive CheAntigen amount required
for immunizationInitial doses: 1–4 mg/horse InitBoosters: 5–10 mg/horse Boo
Probable adverse reactions Anaphylactic reactions andserum sickness
Low
Yield antibodies/years 75 L plasma/horse 3.9–
venom emulsified with complete Freund’s adjuvant (ACF)(1:1 v/v). Then, three boost (elevendays apart)were appliedwith 25 mg of venom emulsified with incomplete Freund’sadjuvant (IFA). Eggs were collected daily, individuallyidentified, and stored at 4 �C.
2.4. Isolation and purification of IgY
Isolation and purification of IgY from the yolk ofpreimmune and hyper-immunized eggs was done ac-cording to a modified Polson’s method (Polson et al.,1985). Briefly, 3.5% (w/v) polyethileneglycol (PEG 6000;Scharlau Chemie SA, European Union) was added to yolkdiluted in phosphate-saline buffer (PBS, 10 mM pH 7.2)(1:3 w/v) under stirring. The supernatant containing IgYwas collected by centrifugation at 3000 � g for 20 min at4 �C, filtered through sterile gauze, followed by theaddition of 20% chloroform, and centrifugation at2000 � g for 15 min. Finally, an IgY precipitation stepwith 8.5% PEG using the same conditions were made.Purified IgY were resuspended in PBS buffer and stored at�20 �C. The purity of these preparations was evaluatedby SDS-PAGE (Laemmli, 1970).
2.5. ELISA measurements
The optimal dilution of antibodies (Crowther, 2001) wasdetermined by enzyme linked immunosorbent assays(ELISA) according to Voller et al. (1974). Briefly, microtiterplates (Greiner Microlon 2HB) were coated with 0.312,0.625, 1.25, 2.5 and 5 mg/mL of T. caripitensis venom in PBSbuffer (pH 7.4). Plates were incubated overnight at 4 �C andthen, blocked with 3% BSA in PBS buffer (pH 7.4) for 2 h.Serial dilutions of IgY anti-venom (1:100 to 1:12,800) wereused in triplicates and incubated at 37 �C for 90 min. Plateswerewashed three times with 0.05% PBS-Tween 20 (PBS-T)buffer between each incubation step. Detection wasdeveloped with HRP-conjugated rabbit anti-chicken IgY (a-IgY-HRP THERMO, Illinois, U.S.A) diluted 1:20,000 in PBS-Tween 20, followed by 0.01% H2O2 with o-phenylenedi-amine (Pierce Biotechnology, Rockford, IL) in phosphate-citrate buffer (0.1 M pH 5.0). Absorbance was read at440 nm on a Synergy� HT reader (Bio-Tek, Vermont, USA).
The estimation of specific antibodies activity to nativescorpion venom was measured weekly by ELISA using theoptimaldilutionsof antibodies andvenomasdescribedabove.
ken anti-venoms IgY References
4 to 6 weeks Dias da Silva and Tambourgi, 2010;WHO, 2010
lves simple andpensive few steps
Dias da Silva and Tambourgi, 2010
ap Meenatchisundaram et al., 2008.ial doses: 30–50 mg/chicken Araújo et al., 2010ster: 25–50 mg/chicken WHO, 2010probability of occurring Maya Devi et al., 2002; Karlson-Stiber
and Persson, 1994; Paul et al., 20074.7 L egg-yolk/chicken Araújo et al., 2010

Fig. 1. Analysis of IgY antibodies purified from egg yolks by SDS-PAGE. Pu-rified IgY antibodies were run on a 10% SDS-PAGE gel and stained withCoomasie brilliant blue R250. Lane 1: Molecular weight marker. Lane 2: 6 mgof commercial purified IgY. Lane 3: 6 mg of IgY anti-T. caripitensis purifiedfrom chicken 1. Lane 4: 6 mg of IgY anti-T. caripitensis purified from chicken 2.Arrow indicated the heavy (68 kDa) and light (25 kDa) chains of IgYantibody.
A. Alvarez et al. / Toxicon 74 (2013) 208–214210
2.6. SDS-PAGE and immunoblotting
Venom and IgY anti-venom samples were analyzed bySDS-PAGE under reducing and dissociating conditions(Laemmli, 1970). Samples were heated for 3 min at100 �C in SDS-PAGE sample buffer (1% SDS, 1% b-mer-captoethanol, 0.001% bromophenol blue, 50 mM, Tris–HCl pH 6.8). The venom samples were separated on a15% SDS-PAGE gel and the IgY anti-venom on 10% gelsand were stained with Coomassie Blue. Recognition ofvenom proteins were carried out by Western blot ac-cording to Towbin’s method (Towbin et al., 1979). Pro-teins previously separated by electrophoresis wereelectroblotted onto nitrocellulose membranes in 25 mMTrizma base, 192 mM glycine and 20% methanol, pH 8.3,using a Mini Trans-Blot system (Bio-Rad, Hercules, CA)for 1 h at 100 V. After blotting, membranes were blockedovernight at 4 �C with blocking solution (5% non-fat milkin PBS buffer pH 7.4, containing 0.1% Tween 20). Blotswere then incubated at 4 �C for 1 h with diluted anti-T.caripitensis IgY (1:1000) in PBS-Tween with 1% non-fatdry milk. After three washes, blots were incubated for1 h with horseradish peroxidase-conjugated rabbit anti-chicken IgY (Pierce Biotechnology, Rockford, IL) diluted1:20,000 in PBS-Tween. Finally, immunoreactivity wasidentified by chemiluminescence (ECL Western BlottingAnalysis System, Amersham, U.K).
2.7. Competitive Western blot
To demonstrate the specificity of venom recognition, IgYanti-venom (diluted 1:1000 in blocking solution) waspreincubated for 30 min with various concentrations of T.caripitensis venom. Then, strips of nitrocellulose withelectroblotted venom proteins were incubated with thesedepleted anti-venom antibodies as described previously.Strips were washed and incubated with HRP-conjugatedrabbit anti-chicken IgY (Pierce Biotechnology, Rockford,IL) diluted 1:20,000 in PBS-Tween. Finally, immunoreac-tivity was identified by chemiluminescence (ECL WesternBlotting Analysis System, Amersham, U.K). The disappear-ance of a band in the Western blot was considered to be aspecific competitive assay.
2.8. Multiple antigen blot assay (MABA)
The cross-reactivity of IgY anti-T. caripitensis venomwasevaluated by dot blot assays (MABA) (Noya and Alarcón deNoya, 1998). Venoms (5 mg/mL) were immobilized onto anitrocellulose membrane, using an acrylic template (Mini-bloter� 28 S-L Immunetics Inc., Cambridge, MA). Twomillimeter width nitrocellulose strips, containing smallsquares from the total number of venoms investigated(different species from genus Tityus and Rhopalurus lat-icauda), were exposed to IgY antibody diluted 1:1000.Thereafter, a second antibody (a-IgY-HRP THERMO, Illinois,U.S.A) diluted 1:20,000 was added and developed by achemiluminescent substrate (Luminol� Amershan, LifeSciences, UK). Positive reactions to the antigen wererecorded as small black squares in a film (Hype film�-ECL,Amershan).
2.9. Neutralization of venom lethality by IgY anti-venomantibodies
The lethal toxicity (LD50) of T. caripitensis venom wasdetermined by intraperitoneal injection using differentdoses of crude venom in five groups of mice (18–20 g). Toassess the ability of the IgY anti-T. caripitensis venom toneutralize venom lethality, 2LD50 (0.2 mL) of venom sam-ple was incubated, for 30 min at 37 �C, with variousamounts of anti-venom IgY (mg). After that the mixturewas injected intraperitoneally into mice. Groups of 5 micewere used at each anti-venom dose. Control mice receivedthe same amount of venom without antibodies. The ED50was calculated from the number of deaths within 48 h ofinjection of the venom/anti-venom mixture and the datawere analyzed using the method of Spearman–Karber(Finney, 1978). The results provide insight into the amountof venom neutralized per mL of anti-venom.
3. Results
3.1. Purity of IgY anti-T. caripitensis venom
Evaluation of purified IgY antibodies by SDS-PAGE ana-lyses showed two typical protein bands with relative mo-lecular masses of 68 and 25 kDa coinciding with thecommercial control (Fig. 1). In addition, two bands (w43and 35 kDa) were observed.
3.2. IgY anti-T. caripitensis venom response
Antibodies levels from venom immunized chickenswere determined by ELISA assays. Based on checkerboardanalysis, the optimum titer for IgYanti-venomwas found tobe 1:100 dilution, with 5 mg/mL of immobilized T. car-ipitensis venom giving an absorbance of 1.0. Experimental
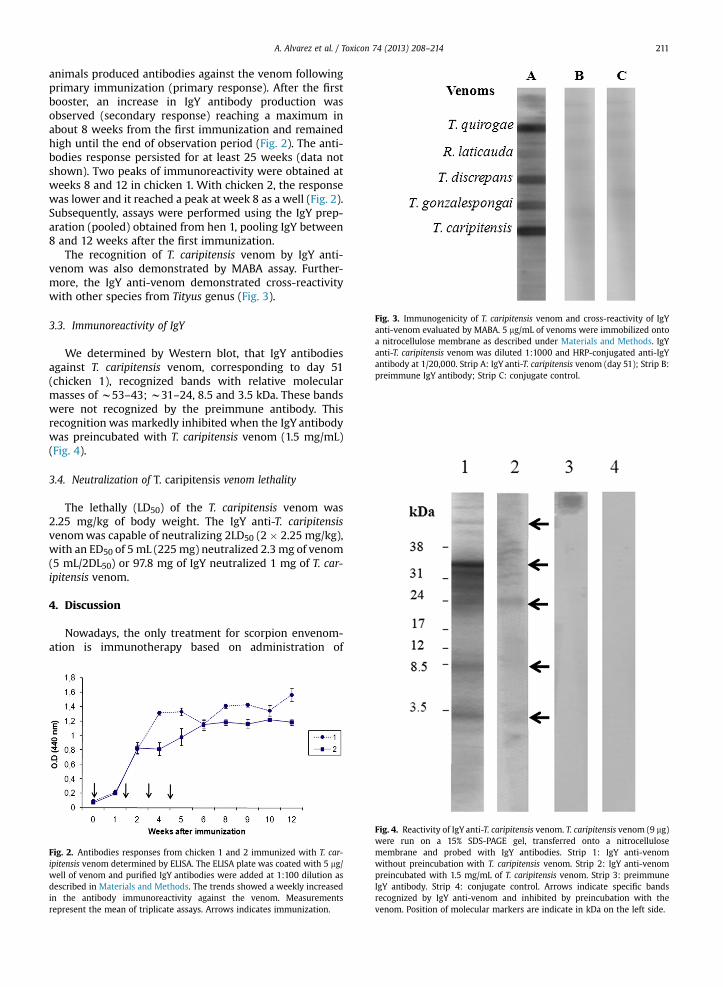
Fig. 3. Immunogenicity of T. caripitensis venom and cross-reactivity of IgYanti-venom evaluated by MABA. 5 mg/mL of venoms were immobilized ontoa nitrocellulose membrane as described under Materials and Methods. IgYanti-T. caripitensis venom was diluted 1:1000 and HRP-conjugated anti-IgYantibody at 1/20,000. Strip A: IgY anti-T. caripitensis venom (day 51); Strip B:preimmune IgY antibody; Strip C: conjugate control.
A. Alvarez et al. / Toxicon 74 (2013) 208–214 211
animals produced antibodies against the venom followingprimary immunization (primary response). After the firstbooster, an increase in IgY antibody production wasobserved (secondary response) reaching a maximum inabout 8 weeks from the first immunization and remainedhigh until the end of observation period (Fig. 2). The anti-bodies response persisted for at least 25 weeks (data notshown). Two peaks of immunoreactivity were obtained atweeks 8 and 12 in chicken 1. With chicken 2, the responsewas lower and it reached a peak at week 8 as a well (Fig. 2).Subsequently, assays were performed using the IgY prep-aration (pooled) obtained from hen 1, pooling IgY between8 and 12 weeks after the first immunization.
The recognition of T. caripitensis venom by IgY anti-venom was also demonstrated by MABA assay. Further-more, the IgY anti-venom demonstrated cross-reactivitywith other species from Tityus genus (Fig. 3).
3.3. Immunoreactivity of IgY
We determined by Western blot, that IgY antibodiesagainst T. caripitensis venom, corresponding to day 51(chicken 1), recognized bands with relative molecularmasses of w53–43; w31–24, 8.5 and 3.5 kDa. These bandswere not recognized by the preimmune antibody. Thisrecognition was markedly inhibited when the IgY antibodywas preincubated with T. caripitensis venom (1.5 mg/mL)(Fig. 4).
3.4. Neutralization of T. caripitensis venom lethality
The lethally (LD50) of the T. caripitensis venom was2.25 mg/kg of body weight. The IgY anti-T. caripitensisvenomwas capable of neutralizing 2LD50 (2 � 2.25 mg/kg),with an ED50 of 5 mL (225mg) neutralized 2.3 mg of venom(5 mL/2DL50) or 97.8 mg of IgY neutralized 1 mg of T. car-ipitensis venom.
4. Discussion
Nowadays, the only treatment for scorpion envenom-ation is immunotherapy based on administration of
Fig. 2. Antibodies responses from chicken 1 and 2 immunized with T. car-ipitensis venom determined by ELISA. The ELISA plate was coated with 5 mg/well of venom and purified IgY antibodies were added at 1:100 dilution asdescribed in Materials and Methods. The trends showed a weekly increasedin the antibody immunoreactivity against the venom. Measurementsrepresent the mean of triplicate assays. Arrows indicates immunization.
Fig. 4. Reactivity of IgY anti-T. caripitensis venom. T. caripitensis venom (9 mg)were run on a 15% SDS-PAGE gel, transferred onto a nitrocellulosemembrane and probed with IgY antibodies. Strip 1: IgY anti-venomwithout preincubation with T. caripitensis venom. Strip 2: IgY anti-venompreincubated with 1.5 mg/mL of T. caripitensis venom. Strip 3: preimmuneIgY antibody. Strip 4: conjugate control. Arrows indicate specific bandsrecognized by IgY anti-venom and inhibited by preincubation with thevenom. Position of molecular markers are indicate in kDa on the left side.

A. Alvarez et al. / Toxicon 74 (2013) 208–214212
antibodies produced in horses previously immunizedagainst the venom. However, side effects as hypersensi-tivity reactions and serum sickness have been reported(Maya Devi et al., 2002). Therefore, it is necessary togenerate new anti-venoms to prevent adverse reactions,keeping a low cost and without animal suffering. In thisstudy, we produced chicken egg yolk antibodies (IgY)against T. caripitensis scorpion venom and evaluated itsneutralizing capacity.
The isolation and purification of antibodies from eggyolks were efficient using the modified Polson’s method(Polson et al., 1985). SDS-PAGE analyses showed two bands(68 and 25 kDa) that correspond to heavy and light chainsfrom Y immunoglobulin. However, some other contami-nating proteins were also observed which could corre-spond to vitellogenins cleavage products, apolipovitellin II(35 kDa) identified by Klimentzou et al. (2006) and phos-vitin (40 kDa) (Mann andMann, 2008). In fact, the presenceof these impurities did not affect the effectiveness ofthe IgYs.
T. caripitensis venom induced an evident antibodyresponse in chicken models as demonstrated by the ELISAassays. The venom was capable of inducing an immuno-genic response as the antibodies recognized it in nativeform, being the best response obtained at weeks 8 and 12after the first immunization. Although there are few re-ports about IgY antibody production against scorpions(Thalley and Carroll, 1990), similar results have beenalready observedwith IgYanti-Echis carinatus snake venom(Paul et al., 2007). Furthermore, we have demonstrated byMABA assays that the IgY anti-T. caripitensis venom wascapable of recognizing venoms from different VenezuelanTityus species (Tityus quirogae, Tityus discrepans, Tityusgonzalespongai), a fact that can be explained by the pres-ence of toxins from common precursors shared amongthese scorpions (Borges et al., 2010). These results are veryimportant as broad spectrum antibodies are appealing tobe used as treatment against envenomation by differentspecies from the Tityus genus.
Western blot analysis showed that IgY antibodiesagainst T. caripitensis venom mainly recognized proteinswith relative molecular weights of 31–24, 8.5 and 3.5 kDa,being the low molecular weight polypeptides of medicalimportance (Petricevich, 2010; Borges et al., 2006). Thesmall toxins found in most scorpions mainly affect potas-sium channels (Borges and De Sousa, 2006; Rodriguez de laVega and Possani, 2007; Pimenta et al., 2003; Tytgat et al.,1999; Novello et al., 1996; Olamendi-Portugal et al., 1996;Martin-Euclaire and Couraud, 1995). However, thew8.5 kDa neurotoxin (Borges and De Sousa, 2006) modifysodium channels (Rodriguez de la Vega and Possani, 2007;Gazarian et al., 2005; Alami et al., 2001; Amaral andRezende, 2000; Martin-Euclaire and Couraud, 1995). TheIgY anti-venom also recognized a protein band of w53–43 kDa, which could correspond to enzymes such as hyal-uronidase, found in Tityus serrulatus, a protein capable offacilitating the spread of the toxin in the tissues (Pessiniet al., 2001). The specificity these antibodies was demon-strated by a competitive assay where the recognition ofthose polypeptides was abrogated by the preincubationwith the IgY anti-venom with T. caripitensis venom.
In this study, it was demonstrated that 225mg of the IgYanti-venom was effective in neutralizing 2.3 mg of T. car-ipitensis venom, corresponding to a 2LD50 challenge.Although several authors have described the neutralizationof toxins present in snake venoms using IgY antibodieswith satisfactory results (Araújo et al., 2010; Paul et al.,2007 Almeida et al., 1998; Akita et al., 1998; Hatta et al.,1993), there are few reports about the efficacy of IgY anti-bodies in neutralizing scorpion venoms lethality (Thalleyand Carroll, 1990). Therefore, this work represents acontribution to the generation of new anti-venoms in hensagainst scorpions of medical importance in our country.Certainly, chickens are a convenient and inexpensivesource of anti-venom. It has been reported that 18 immu-nized chickens were capable of producing the sameamount of anti-venom as one horse per year (Araújo et al.,2010). The high amount of anti-venom and the low main-tenance cost make the IgY production an attractive alter-native to produce large stocks of anti-venom fortherapeutic applications to scorpionism in Venezuela.
A drawback of the use of IgY in populations with animportant dietary component of poultry meat and eggs,cold be the risk of reducing the efficacy of IgY anti-venomand producing adverse reactions due the development ofIgGs against IgY, as pointed out by Sevcik et al. (2008).However, several studies indicate that antigens orallyingested can induce specific tolerance rather than an activeimmune response; oral tolerance can be defined as an in-hibition of specific immune response to subsequentparenteral injections of proteins to which an individual oranimal has been previously exposed via the oral route (Wuand Weiner, 2003; Mucida et al., 2005). Accumulated evi-dence shows that oral tolerance can be mediated by orallyactivated humoral and cellular factors (Gomes-Santos et al.,2012; Zemann et al., 2003). Some mechanisms have beenproposed for the development of oral tolerance: the dele-tion of antigen-specific T-cells (Chen et al., 1995), immunedeviation (Alpan et al., 2004), induction of anergy (VanHouten and Blake, 1996) and suppression by regulatory T-cells (Zhang et al., 2001). The fail of oral tolerance charac-terizes a number of pathological conditions includingchildhood onset Type I hypersensitivity, celiac disease andmany others autoimmune conditions. However, we shouldnot rule out the possibility of inducing an immune response(IgG/IgE), more than a tolerance, in individual with food-hypersensitivity reactions against eggs. It is important tohighlight that the reactions of hypersensitivity take placeagainst food-processed antigens (Sanchez and Fremont,2003). This processing may destroy existing epitopes on aprotein or may cause new ones to be formed (neoallergenformation) as a result of change in protein conformation(Vojdani, 2009). In the case of IgY anti-venom, it is lessprobably that hypersensitivity reactions might occurbecause it is a crude antigen. Nevertheless, is necessary toperform clinical trials for IgY tolerability and safety forfuture human applications. One strategy could be theevaluation of egg allergy with diagnostic tests measure-ment of egg-specific IgE using the skin prick test beforeapplying IgY anti-venom.
The present study demonstrates that the antibodiesraised were specific and capable of neutralizing the activity

A. Alvarez et al. / Toxicon 74 (2013) 208–214 213
of the T. caripitensis venom indicating the potential use-fulness of avian anti-scorpion venom as an alternative toconventional equine anti-venom in Venezuela.
Acknowledgments
This work was supported by Research Grant “ProyectoMisión Ciencia: Producción de Antivenenos” FONACIT No.2007000672. We are grateful to Msc. Hudefe, Msc. Parrilla,Mrs Tomas and Dra. Quiroga, for T. caripitensis collection(Pharmacology laboratory, Universidad de Oriente, Bolívar).We thank toMrGuevara,Mr.Mejias,Mr. Pérez,Mr. Toyo andMSc. Negrón for their assistance in animal care (Bioterio,IDEA- Venezuela.) We are grateful also for the DoctorsCecilia Castillo and JuanCarlosMartínez for English revision.
Conflict of interest
The authors declare that there are no conflicts ofinterest.
References
Akita, E.M., Li-Chan, E.C., Nakai, S., 1998. Neutralization of enterotoxogenicEscherichia coli heat-labile toxin by chicken egg yolk immunoglobulinY and its antigen-binding fragments. Food Agric. Immunol. 10, 161–172.
Alami, M., Ouafik, L., Céard, B., Legros, C., Bougis, P.E., Martin-Eauclaire, M., 2001. Characterization of the gene encoding the alpha-toxin Amm V from the scorpion Androctonus mauretonicus maur-etanicus. Toxicon 39, 1579–1585.
Almeida, C.M., Kanashiro, M.M., Rangel Filho, F.B., Mata, M.F., Kipnis, T.L.,Dias da Silva, W., 1998. Development of snake antivenom antibodiesin chickens and their purification from yolk. Vet. Rec. 143, 579–584.
Alpan, O., Bachelder, E., Isil, E., Arnheiter, H., Matzinger, P., 2004.‘Educated’ dendritic cells act as messengers from memory to naive Thelper cells. Nat. Immunol. 5, 615–622.
Amaral, C.F.S., Rezende, N.A., 2000. Treatment of scorpion envenomingshould include both a potent specific antivenom and support of vitalfunctions. Toxicon 38, 1005–1007.
Araújo, A.S., Lobato, Z.I.P., Chávez-Olórtegui, C., Velarde, D.T., 2010. Bra-zilian IgY- Bothrops antivenom: studies on the development of aprocess in chicken egg yolk. Toxicon 55, 739–744.
Borges, A., 1996. Scorpionism in Venezuela. Acta Biol. Venez. (Caracas) 16,65–76.
Borges, A., De Sousa, L., 2006. Escorpionismo en Venezuela: Una aprox-imación molecular, inmunológica y epidemiológica para su estudio.Rev. Fac. Farm. (Caracas) 1, 15–27.
Borges, A., García, C., Lugo, E., Alfonso, M., Jowers, M., Op den Camp, H.,2006. Diversity of long-chain toxins in Tityus zulianus and Tityusdiscrepans venoms (Scorpiones, Buthidae): molecular, immunologicaland mass spectral analyses. Comp. Biochem. Physiol. C 142, 240–252.
Borges, A., Bermingham, E., Herrera, N., Alfonzo, M.J., Sanjur, O.I., 2010.Molecular systematics of the neotropical scorpion genus Tityus(Buthidae): the historical biogeography and venom antigenic di-versity of toxic Venezuelan species. Toxicon 55, 436–454.
Chacana, P., Terzolo, E., Schade, R., 2004. Tecnología IgYo aplicaciones de losanticuerpos de yema de huevo de gallina: biología, propiedades y suaplicación enmedicina y veterinaria. Rev.Med. Vet. (Bs. As) 85,179–189.
Chen, Y., Inobe, J., Marks, R., Gonnella, P., Kuchroo, V.K., Weiner, H.L., 1995.Peripheral deletion of antigen-reactive T cells in oral tolerance. Na-ture 376, 177–180.
Crowther, J., 2001. Methods in Molecular Biology. In: The ELISA Guide-book. Humana Press Inc., Totowa. New Jersey, pp. 149–165.
De Sousa, L., Vásquez, D., Salazar, D., Valecillo, R., Vasquez, D., Rojas, M.,Parrilla Álvarez, P., Quiroga, M., 2005. Mortalidad en humanos porenvenenamientos causados por invertebrados y vertebrados en elestado Monagas, Venezuela. Investig. Clín. 46, 241–254.
De Sousa, L., Manzanilla, J., Parrilla-Alvarez, P., 2006. Nueva especie deTityus (Scorpiones: Buthidae) del Turimiquire, Venezuela. Rev. Biol.Trop. (Caracas) 54, 489–504.
Dias da Silva, W., Tambourgi, D.V., 2010. IgY: a promising antibody for usein immunodiagnostic and in immunotherapy. Vet. Immunol. Immu-nopathol. 135, 173–180.
Finney, D.J., 1978. Statistical Methods in Biological Assay, third ed. CharlesGriffin and Co. Ltd., London, p. 508.
Gazarian, K.G., Gazarian, T., Hernandez, R., Possani, L.D., 2005. Immu-nology of scorpion toxins and perspectives for generation of anti-venom vaccines. Vaccine 23, 3357–3368.
Gomes-Santos, A.C., Moreira, T.G., Castro-Junior, A.B., Horta, B.C.,Lemos, L., Cruz, D.N., Guimarães, M.A., Cara, D.C., McCafferty, D.M.,Faria, A.M., 2012. New insights into the immunological changes in IL-10-deficient mice during the course of spontaneous inflammation inthe gut mucosa. Clin. Dev. Immunol. 2012, 1–13.
Gwee, M.C.E., Nirthanan, S., Khoo, H.E., Gopalakrishnakone, P., Kini, R.M.,Cheah, L.S., 2002. Autonomic effects of some scorpion venoms andtoxins. Clin. Exp. Pharmacol. Physiol. 29, 795–801.
Hatta, H., Tsuda, K., Akachi, S., Kim, M., Yamamoto, T., 1993. Productivityand some properties of egg yolk antibody (IgY) against humanrotavirus compared with rabbit IgG. Biosci. Biotechnol. Biochem. 57,450–454.
Karlson-Stiber, C., Persson, H., 1994. Antivenom treatment in Vipera berusenvenoming– report of 30 cases. J. Intern. Med. 235, 57–61.
Klimentzou, P., Paravatou-Petsotas, M., Zikos, C., Beck, A., Skopeliti, M.,Czarnecki, J., Tsitsilonis, O., Voelter, W., Livaniou, E., Evangelatos, G.P.,2006. Development and immunochemical evaluation of antibodies Yfor the poorly, immunogenic polypeptide prothymosin alpha. Pep-tides 27, 183–193.
Laemmli, U., 1970. Cleavage of structural proteins during the assemble ofbacteriophage T4. Nature 227, 680–685.
Mann, K., Mann, M., 2008. The chicken egg yolk plasma and granuleproteomes. Proteomics 8, 178–191.
Martin-Euclaire, M.F., Couraud, F., 1995. Scorpion neurotoxins: effects andmechanisms. In: Handbook of Neurotoxicology. New York, MarcelDekker, pp. 683–716.
Maya Devi, C., Vasantha Bai, M., Krishnan, L.K., 2002. Development ofviper-venom antibodies in chicken egg yolk and assay of their antigenbinding capacity. Toxicon 40, 857–861.
Meenatchisundaram, S., Parameswari, G., Michael, A., Ramalingam, S.,2008. Neutralization of the pharmacological effects of Cobra and Kraitvenoms by chicken egg yolk antibodies. Toxicon 52, 221–227.
Mucida, D., Kutchukhidze, N., Erazo, A., Russo, M., Lafaille, J.J., Curotto deLafaille, M.A., 2005. Oral tolerance in the absence of naturallyoccurring Tregs. J. Clin. Invest. 115, 1923–1933.
Müller, G.J., 1993. Scorpionism in South Africa. A report of 42 seriousscorpion envenomations. S. Afr. Med. J. 83, 405–411.
Novello, J.C., Oliveira, B., Pereira, M.F., Giglio, J.R., Arantes, E.C.,Sampaio, S.V., Marangoni, S., 1996. The primary structure of TSTX IV.In: XXV Annual Meeting of the Brazilian Society of Biochemistry andMolecular Biology, Caxambu, MG M, pp. 138–141.
Noya, O., Alarcón de Noya, B., 1998. The multiple antigen blot assay(MABA): a simple immunoenzymatic technique for simultaneousscreening of multiple antigens. Immunology 63, 53–56.
Olamendi-Portugal, T., Gomez-Lagunas, F., Gurrola, G.B., Possani, L.D.,1996. A novel structural class of Kþ channel blocking toxin from thescorpion Pandinus imperator. Biochem. J. 315, 977–981.
Paul, K., Manjula, J., Deepa, E., Selvanayagam, G.K., Ubba, R.P., 2007. Anti-Echis carinatus venom antibodies from chicken egg yolk: isolation,purification and neutralization efficacy. Toxicon 50, 893–900.
Pessini, A.C., Takao, T.T., Cavalheiro, E.C., Vichnewski, W., Sampaio, S.V.,Giglio, R., Arantes, E.C., 2001. A hyaluronidase from Tityus serrulatusscorpion venom: isolation, characterization and inhibition by flavo-noids. Toxicon 39, 1495–1504.
Petricevich, V.L., 2010. Scorpion venom and the inflammatory response.Mediators Inflamm. 2010, 1–16, 903295.
Pimenta, A.M.C., Legros, C., Almeida, F.D.M., Mansuelle, P.M.E., Bougis, P.E.,Martin-Eauclarie, M.F., 2003. Novel structural class of four disulfide-bridged peptides from Tityus serrulatus. Biochem. Biophys. Res.Commun. 301, 1086–1092.
Poggioli De Scannone, J., 1996. Suero antiescorpiónico. Desarrollo y pro-ducción (Resumen). In: I Taller los Escorpiones y sus Toxinas, Biología,Clínica y Toxinología. Caracas, Venezuela. Ed IVIC, p. 11.
Polson, A., Coetzer, T., Kruger, J., Von Maltzahn, E., Van Merwe, K.J., 1985.Improvements in the isolation of IgY from the eggs laid by immu-nized chickens. J. Immunol. Invest. 14, 323–327.
Rodriguez de la Vega, R.C., Possani, L.D., 2007. Novel paradigms on scor-pion toxins that affects the activating mechanism of sodium channels.Toxicon 49, 171–180.
Russell, W.M.S., Burch, R.L., 1959. The Principles of Humane ExperimentalTechnique. Methuen, London, p. 238.

A. Alvarez et al. / Toxicon 74 (2013) 208–214214
Sanchez, C., Fremont, S., 2003. Consequences of heat treatment andprocessing of food on the structure and allergenicity of componentproteins. Rev. Fr Allergol. Immunol. Clin. 43, 13–20.
Schade, R., Calzado, E., Sarmiento, R., Chacana, P., Porankiewicz-Asplund, J., Terzolo, H., 2005. Chicken egg yolk antibodies (IgY-tech-nology): a review of progress in production and use in research andhuman and veterinary medicine. ATLA 33, 1–26.
Sevcik, C., Diaz, P., D0Suze, G., 2008. On the presence of antibodies againstbovine, equine and poultry immunoglobulins in human IgG prepa-rations, and its implications on antivenom production. Toxicon 51,10–16.
Thalley, B.T., Carroll, S., 1990. Rattlesnake and scorpion antivenoms fromthe egg yolk of immunized chickens. Biotechnology 8, 934–938.
Towbin, H., Stachelin, T., Gordon, J., 1979. Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: procedureand some applications. Proc. Natl. Acad. Sci. 9, 4350–4354.
Tytgat, J., Chandy, K.G., Garcia, M.L., 1999. A unified nomenclature forshort-chain peptides isolated from scorpion venoms: alpha-KTxmolecular subfamilies. Trends Pharmacol. Sci. 11, 444–447.
Van Houten, N., Blake, S.F., 1996. Direct measurement of anergy ofantigen-specific T cells following oral tolerance induction. J. Immunol.157, 1337–1341.
Vojdani, A., 2009. Detection of IgE, IgG, IgA and IgM antibodies againstraw and processed food antigens. Nutr. Metab. (Lond) 6, 22–39.
Voller, A., Bidwell, B., Hold, G., Engwall, E., 1974. A microplate method ofenzyme-linked-immunosorbent assay and its application to malaria.Bull. World Health Organ 51, 209–211.
World Health Organization, 2010. Guidelines for the Production, Controland Regulation of Snake Antivenom Immunoglobulins (Geneva).
Wu, H.Y., Weiner, H.L., 2003. Oral tolerance. Immunol. Res. 28, 265–284.Zemann, B., Schwaerzler, C., Griot-Wenk, M., Nefzger, M., Mayer, P.,
Schneider, H., de Weck, A., Carballido, J.M., Liehl, E., 2003. Oraladministration of specific antigens to allergy-prone infant dogs in-duces IL-10 and TGF-beta expression and prevents allergy in adultlife. J. Allergy Clin. Immunol. 111, 1069–1075.
Zhang, X., Izikson, L., Liu, L., Weiner, H.L., 2001. Activation of CD25(þ)CD4(þ) regulatory T cells by oral antigen administration. J. Immunol.167, 4245–4253.