immunocytochemical localization of polyamines during - biology
TRANSCRIPT

Immunocytochemical Localization ofPolyamines During Attachment and
Spreading of Retinal Pigment Epithelial andIntestinal Epithelial Cells
Dianna A. Johnson,1,2* Rajesh K. Sharma,1 Katherine Allan,1 Ramesh Ray,2
and Leonard R. Johnson1,2
1Department of Ophthalmology, University of Tennessee Health Science Center,Memphis
2Department of Physiology, University of Tennessee Health Science Center,Memphis
In order to form and maintain a protective barrier for photoreceptors, the retinalpigment epithelium relies on integrin signaling and related pathways to formadhesion complexes, undergo cell spreading, and establish a confluent cellularmonolayer. Polyamines are multifunctional polycations that are essential for cellattachment and spreading, although their exact mechanisms of action are as yetunclear. We report new immunocytochemical evidence suggesting that in the cellsof retinal pigment epithelium and also the intestinal epithelium, polyamines arepresent in a population of intracellular vesicles that appear transiently duringinitial stages of cell spreading. In newly attached cells with minimal spreading, thevesicles are seen near the nucleus, whereas in more highly spread cells, thevesicles are localized to the plasma membrane, near, but not precisely co-localizedwith an enzyme marker for adhesion complexes, focal adhesion kinase. We alsoobserve pronounced nuclear staining in newly attached cells that have not spread,whereas this staining is decreased in cells that have spread. Nuclear staining hasbeen previously reported in other cell types and has been attributed to DNAbinding of polyamines, which is known to stabilize chromatin structure. Wehypothesize that the appearance of polyamine vesicles near focal adhesions ofcells undergoing attachment and spreading may reflect the mechanism by whichpolyamine pools are targeted to appropriate interaction sites necessary for theassembly of adhesion complexes. Alternatively, the vesicles could represent themechanism by which polyamines are removed from the nucleus and possiblyreleased from the cell. Cell Motil. Cytoskeleton 58:269–280, 2004.© 2004 Wiley-Liss, Inc.
Key words: retinal pigment epithelium; intestinal epithelium cell; polyamines; focal adhesion kinase; cellspreading; cell migration
Contract grant sponsor: National Eye Institute; Contract grant num-bers: EYYO1655 and EY13080; Contract grant sponsor: NationalInstitute of Diabetes and Digestive and Kidney Diseases; Contractgrant number: DK52784; Contract grant sponsor: Research to PreventBlindness; Contract grant sponsor: Thomas A. Gerwin Endowment.
*Correspondence to: Dianna A. Johnson, Ph.D., Department of Oph-thalmology, University of Tennessee Health Science Center, 930 Mad-ison Ave., Memphis TN 38163. E-mail: [email protected]
Received 26 January 2004; Accepted 29 March 2004Published online in Wiley InterScience (www.interscience.wiley.com).DOI: 10.1002/cm.20014
Cell Motility and the Cytoskeleton 58:269–280 (2004)
© 2004 Wiley-Liss, Inc.

INTRODUCTION
Retinal pigment epithelium (RPE) serves multipleroles that are important for protecting and nurturing cellsof the neural retina. Arranged as a confluent monolayerof largely quiescent, post-mitotic cells, the RPE forms abarrier between the major vascular bed of the eye, pro-vides a uniform substrate for attachment of photoreceptorouter segments via a network of extracellular matrix, andprovides an osmotic barrier for movement of fluid in thesubretinal space. Thus, the ability of RPE cells to estab-lish and maintain attachments to adjacent cells in themonolayer and to their underlying substrate, calledBruch’s membrane, is critically important to their overallbarrier function and to their dominant role in maintaininga suitable environment for light detection by photorecep-tor cells. Loss of RPE function, with subsequent death ofphotoreceptors, is thought to be the causative factor inseveral retinal diseases, including age-related maculardegeneration and gyrate atrophy [Wang et al., 1996;Young, 1987].
RPE cells are attached (1) to an underlying sub-strate, formed by a basal lamina and Bruch’s membrane;(2) to each other through junctional complexes (zonulaoccludens, zonula adherens, gap junctions, and desmo-somes); and (3) to photoreceptor outer segments via anextracellular matrix structure, the interphotoreceptor ma-trix [Burnside and Bost-Usiner, 1998]. In addition toproviding an anchor for RPE cells, these points of attach-ment are thought to regulate intracellular signaling path-ways through activation of integrin receptors present at anumber of these attachment sites [Moszczynska andOpas, 1994]. Cytoskeletal elements are also associatedwith attachment sites, providing a cellular frameworkthat determines cell shape and, when needed, tractionpoints for cell spreading and migration. Because cy-toskeletal elements, such as actin filaments, can be boundto integrin receptors, the integrin signaling pathway func-tions as a major regulator of the cytoskeleton [Cooper,1991; Giancotti and Ruoslaht, 1999; Nobes and Hall,1995; Symons, 1996]. Thus, a primary means by whichextracellular conditions influence the intracellular envi-ronment can involve receptor ligands that are non-diffus-able (e.g., integrin binding to a substrate). The othermajor class of receptor ligands comprises diffusable sub-stances, including growth factors. Together, the solubleand substrate ligands, through their respective receptors,trigger intracellular pathways that regulate cell attach-ment and determine cell shape.
The polyamines spermidine and spermine and theirprecursor, putrescine, are found in virtually all cells ofhigher eukaryotes and are intimately involved in andrequired for cell growth, proliferation, and cell migration[Igarashi and Kashiwagi, 2000; Shaw, 1994]. Since there
is strong evidence that polyamines are necessary forformation/breaking of cell attachments during migration[McCormack et al., 1993, 1994a; Ray et al., 2000], wehypothesize that polyamines may represent an importantsoluble ligand necessary for formation of cellular attach-ments in RPE cells. Intracellular polyamine levels arehighly regulated and are dependent primarily on theactivity of ornithine decarboxylase (ODC), which cata-lyzes the first rate limiting step in polyamine biosynthe-sis, the decarboxylation of ornithine to form the diamine,putrescine [Pegg et al., 1988; Pegg et al., 1987; Seiler1994]. Spermidine and spermine are subsequentlyformed through the actions of spermidine and sperminesynthase, respectively, and S-adenoxylmethionine decar-boxylase (SAMdc). SAMdc forms decarboxylated S-ad-enosylmethione, thus providing the propylamine groupsneeded for polyamine synthesis.
DL-alpha-difluoromethylornithine (DFMO) is aspecific and irreversible inhibitor of ODC, having noeffects except those caused by inhibition of new poly-amine synthesis and the subsequent depletion of existingpools of polyamines through metabolism [McCormack etal., 1993, 1994a,b, 1999a]. We have previously shown inRPE that stimulation of ODC activity by exposure toserum is completely blocked by simultaneous addition ofDFMO. The DFMO effect appeared to be instantaneousand was readily observed at the initial assay time point at4 h [Johnson et al., 2002]. Similar results are seen inintestinal epithelial cells. For example, the inhibition ofODC with DFMO leads to the activation of STAT3within 4 h in the IEC-6 cell line [Pfeffer, et al 2000].
Polyamine depletion occurs at different rates de-pending on the specific polyamine. Putrescine and sper-midine were depleted in RPE within 8 h of exposure toDFMO, whereas spermine levels were unaffected [John-son et al., 2002]. Spermine is tightly bound to proteinsand other macromolecules, thus it is likely that free andavailable polyamines are depleted rapidly [Seiler, 1994].This, perhaps, explains rapid cellular responses toDFMO, such as STAT3 activation. All of the knowneffects of DFMO can be reversed by the addition ofexogenous polyamines [McCormack et al., 1994, 1999b;Russell, 1985] Thus, DFMO has provided the primarytool for the study of polyamine functions.
Recent studies in epithelial cell types (other thanRPE) have focused on the effect of polyamines on theRho family of GTPases, members of which (i.e., Cdc,Rac, and Rho) are known to control formation of filop-odia or microspikes [Symons 1996], lamellipodia [Ridleyet al., 1992], and assembly of focal adhesions and stressfibers [Ridley and Hall 1992], respectively. In intestinalepithelial cells, polyamines were shown either to activateRac1 directly or, more likely, to be required for a processnecessary for activation of Rac1 [Ray et al., 2003]. In
270 Johnson et al.

turn, Rac1 was shown to activate RhoA, but that activa-tion step did not require polyamines. Cdc42 is also acti-vated by Rac1, although the position of Cdc42 in theactivation hierarchy (upstream or downstream from Rho)was difficult to assess because of cross-talk. Thus, poly-amines may trigger a Rac1-activated cascade of eventsthat are required for cell attachment and cell spreading,as well as migration.
We have previously shown that, similar to intesti-nal epithelial cells, RPE cells in culture can be stimulatedto migrate in response to injury; they synthesize and storeendogenous polyamines, and depletion of the polyaminestores by DFMO inhibits cell migration [Johnson et al.,2002]. The addition of exogenous polyamines blocks theDFMO-induced inhibition. We also demonstrated thatthere was a general increase in polyamine immunoreac-tivity in RPE cells undergoing migration compared toquiescent cells in a confluent monolayer. In confluentcells, polyamine immunoreactivity was confined to mem-brane sites at points of contact between cells. The presentstudy examines polyamine localization in newly platedcells undergoing attachment and spreading. The threecell lines chosen for study (D407 and ARPE-19 humanRPE cells, and IEC-6 rat intestinal epithelial cells) havedifferent growth characteristics and different degrees ofcontact inhibition. In spite of these differences, all threecell lines show similar staining patterns. In newly placedcultures, we observe a transient population of polyamineimmunoreactive vesicles that initially appear within thethin rim of cytoplasm near the nucleus. With increasedcell spreading, the vesicles become distanced from thenucleus but remain closely associated with the plasmamembrane at sites adjacent to, but not precisely co-localized with, an enzyme marker for the focal adhesionkinase (FAK). Our results suggest that localization andlevels of polyamines change as the cell undergoes dif-ferent stages of attachment and spreading.
MATERIALS AND METHODS:
Cell Culture
Two different RPE cell lines (D407, a gift from Dr.Alberta Davis, Alcon Laboratories, Fort Worth, TX;[Davis et al., 1995] and ARPE-19, obtained from Amer-ican Type Culture Collection, ATTC, Manassas, VA)were maintained in T-25 culture flasks (Becton Dickin-son, Franklin Lake, NJ) at 37°C in an atmosphere of 95%air and 5% CO2. The culture medium contained Dulbec-co’s modified Eagles medium (DMEM, GIBCO, Invitro-gen Corp., Carlsbad, CA) with 5% neonatal calf serum,insulin (50 �g/mL), and an antibiotic-antimycotic mix-ture (10,000 U/mL penicillin G Sodium, 10 mg/mLstreptomycin sulfate, and 25 �g/mL amphotericin B).
Cells were passaged weekly and fed every other day.Culture passages 15–25 were used in the experiments.After reaching confluence, cells were removed from theflask by trypsinization (0.05% trypsin/EDTA for 5 min),washed with fresh medium, counted in a hemocytometer,and replated (250,000 cells/well) on laminin-coated glasscoverslips in 24 well plates. At the termination of theexperimental culture period, cultures were fixed in 4%paraformaldehyde, 4% sucrose (pH 7.2) for 1 h at roomtemperature.
For antibody staining, samples were preincubatedin blocking buffer (phosphate buffered saline with 1%Triton, 5% bovine serum albumin, and 5% normal serumto match the animal in which the primary antibody wasmade) for 1.5 h at room temperature or overnight at 4°C.Antibodies against polyamines were obtained from Dr.Jean Guy Delcros (Institute de Rechereche Contre leCancer, Rennes, France). The antibody has been wellcharacterized and was shown to have the highest bindingaffinity for spermine, with lesser affinity for spermidineand putrescine [Delcros et al., 1996; Bouille et al., 1999].Antibodies against FAK were obtained from Santa CruzBiotechnology, Santa Cruz, CA. Appropriately conju-gated secondary antibodies were purchased from JacksonImmunoresearch Laboratories, West Grove, PA. Sampleswere incubated in primary antibodies at appropriate di-lutions for 1 h at room temperature or overnight at 4°C,rinsed, incubated with appropriate fluorescence taggedsecondary antibody for 30 min at room temperature,rinsed again, and mounted in Vectashield� medium. ForFAK visualization, we used Cy3-conjugated anti-rabbitsecondary antibodies, which appear red; for polyaminevisualization, we used Cy2-conjugated anti-mouse sec-ondary antibodies, which appear green. For samples thatwere double labeled, the two primary antibodies werecombined for the initial incubation; the two secondaryantibodies were combined for the second incubation pe-riod. Samples were viewed either on Nikon fluorescencemicroscopes fitted with a Metaview� or Metamorph�Image analysis system or on a Bio-Rad confocal micro-scope.
For visualization of diaminobenzidine (DAB)-la-beled antibodies, cultures were rinsed several times inPBS and then incubated in the appropriate biotinylatedsecondary antibody for 30 min at room temperature. Allsecondary antibodies were obtained from Vector, Burlin-game, CA. Endogenous peroxidase activity was blockedby incubation in 0.3% hydrogen peroxide solution for 15min. Samples were then incubated in Vectastain, ABCsolution (Vector, Burlingame, CA) for 30 min followedby reaction with Vector DAB substrate for 4 min. Sam-ples were allowed to air dry and were mounted in Cy-toseal 60 mounting medium.
Polyamines in RPE Cell Spreading 271
Phalloidin Staining
After fixation, cells were washed with PBS andpermeabilized with 0.025% Triton X-100 in PBS for 5min. Cells were rinsed with PBS containing 0.3% BSAthree times and placed in a solution of Texas red labeledphalloidin (Molecular Probes T7471, Eugene, OR) di-luted 1:40 in PBS/0.3% BSA for 1 h. After rinsing inPBS, cells were mounted on glass slides with Fluoro-mount� mounting medium.
RESULTS
Changes in Morphology, F-Actin Cytoskeleton,and Polyamine Immunoreactivity in Newly PlatedD407 Cells
Cultured cells from the D407 human RPE cell lineundergo rapid changes in cell morphology within the firsthour after plating (Figs. 1 and 2). Concurrent withchanges in cytoskeletal organization and cell shape, there
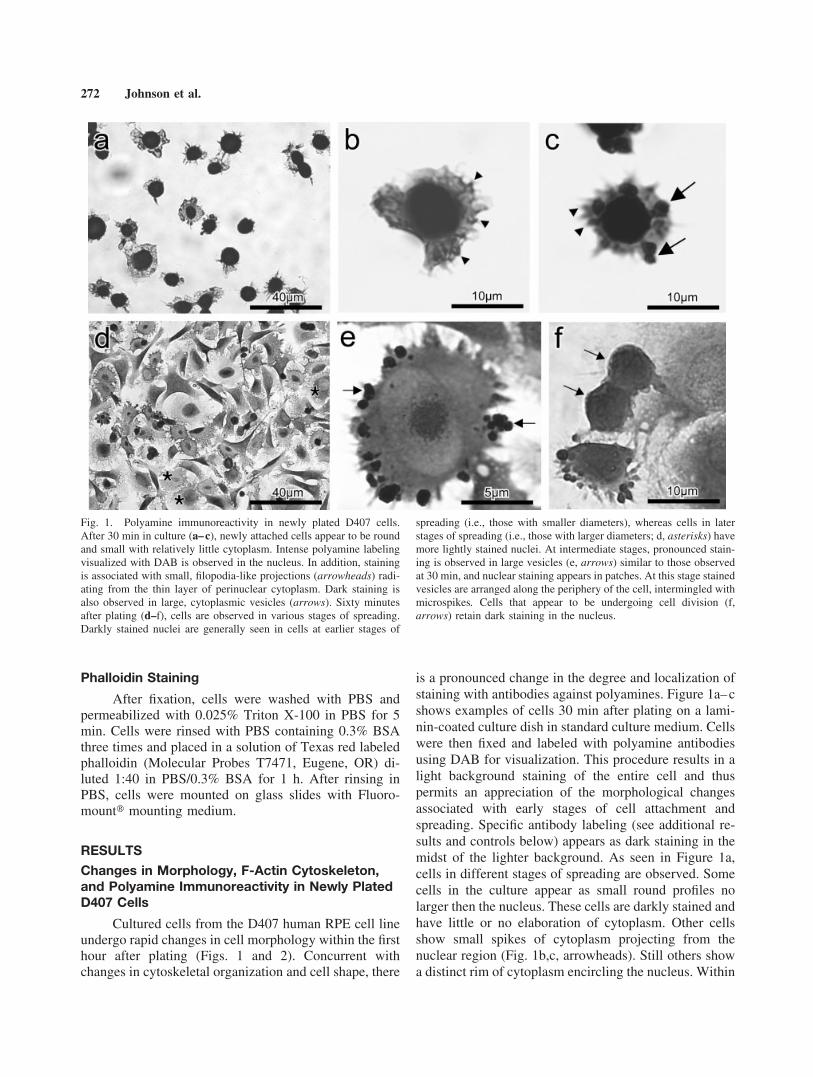
is a pronounced change in the degree and localization ofstaining with antibodies against polyamines. Figure 1a–cshows examples of cells 30 min after plating on a lami-nin-coated culture dish in standard culture medium. Cellswere then fixed and labeled with polyamine antibodiesusing DAB for visualization. This procedure results in alight background staining of the entire cell and thuspermits an appreciation of the morphological changesassociated with early stages of cell attachment andspreading. Specific antibody labeling (see additional re-sults and controls below) appears as dark staining in themidst of the lighter background. As seen in Figure 1a,cells in different stages of spreading are observed. Somecells in the culture appear as small round profiles nolarger then the nucleus. These cells are darkly stained andhave little or no elaboration of cytoplasm. Other cellsshow small spikes of cytoplasm projecting from thenuclear region (Fig. 1b,c, arrowheads). Still others showa distinct rim of cytoplasm encircling the nucleus. Within
Fig. 1. Polyamine immunoreactivity in newly plated D407 cells.After 30 min in culture (a–c), newly attached cells appear to be roundand small with relatively little cytoplasm. Intense polyamine labelingvisualized with DAB is observed in the nucleus. In addition, stainingis associated with small, filopodia-like projections (arrowheads) radi-ating from the thin layer of perinuclear cytoplasm. Dark staining isalso observed in large, cytoplasmic vesicles (arrows). Sixty minutesafter plating (d–f), cells are observed in various stages of spreading.Darkly stained nuclei are generally seen in cells at earlier stages of
spreading (i.e., those with smaller diameters), whereas cells in laterstages of spreading (i.e., those with larger diameters; d, asterisks) havemore lightly stained nuclei. At intermediate stages, pronounced stain-ing is observed in large vesicles (e, arrows) similar to those observedat 30 min, and nuclear staining appears in patches. At this stage stainedvesicles are arranged along the periphery of the cell, intermingled withmicrospikes. Cells that appear to be undergoing cell division (f,arrows) retain dark staining in the nucleus.
272 Johnson et al.
this cytoplasmic rim, darkly stained vesicles are oftenseen (Fig. 1c, arrows).
Cells maintained 60 min after plating show majorchanges in morphology (Fig. 1d–f). Variations in celldiameter and shape are observed, indicating that somecells (larger diameter) have undergone significantspreading, while others have not. Many cells have diam-eters that are increased several-fold over those of cellsharvested after 30 min (Fig. 1a–c). Polyamine staining inthe nuclei of these larger cells is noticeably decreased.(Fig.1d, asterisks) In many of the larger cells (for exam-ple, see Fig. 1e), residual staining in the nucleus isrestricted to a small patch, with the remaining nucleararea showing only background staining. A number of thelarger cells still retain populations of strongly immuno-reactive vesicles. These vesicles are generally locatednear the plasma membrane where spike-like projectionsappear (Fig. 1e, arrows). In contrast to large-diametercells, other cells in the 60-min culture appear to beundergoing mitosis (Fig. 1f). In these cases, the nucleusis darkly stained.
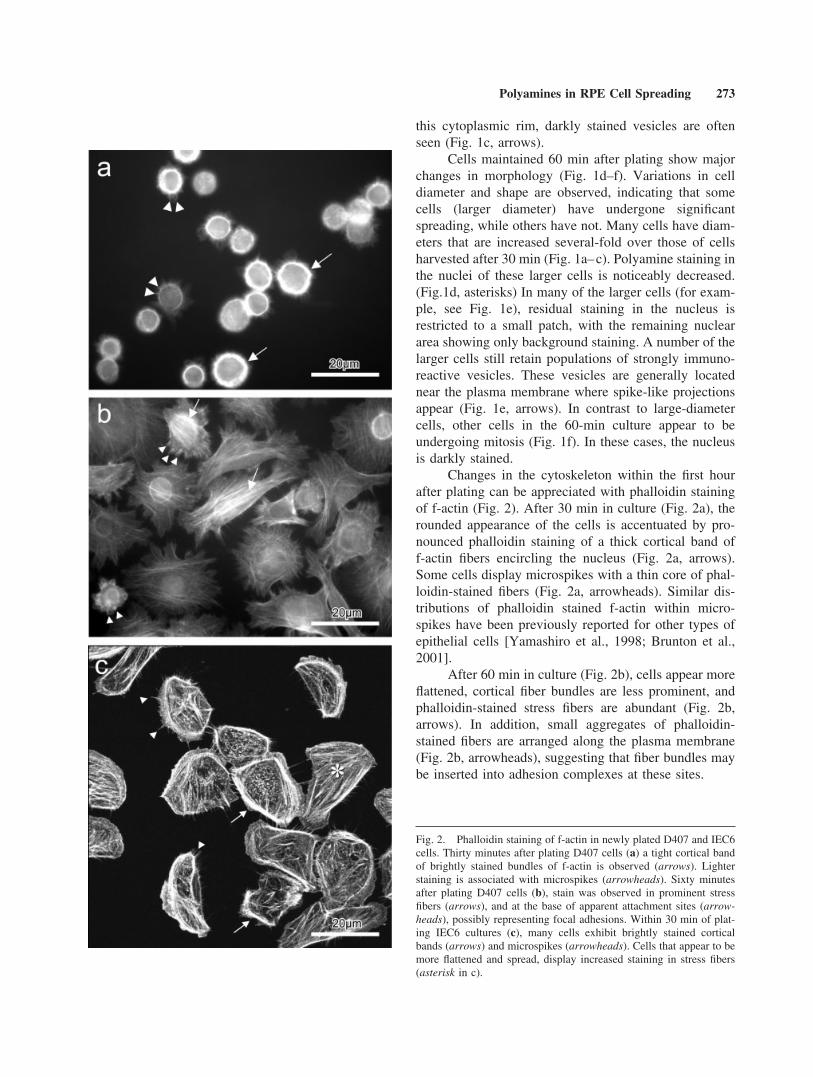
Changes in the cytoskeleton within the first hourafter plating can be appreciated with phalloidin stainingof f-actin (Fig. 2). After 30 min in culture (Fig. 2a), therounded appearance of the cells is accentuated by pro-nounced phalloidin staining of a thick cortical band off-actin fibers encircling the nucleus (Fig. 2a, arrows).Some cells display microspikes with a thin core of phal-loidin-stained fibers (Fig. 2a, arrowheads). Similar dis-tributions of phalloidin stained f-actin within micro-spikes have been previously reported for other types ofepithelial cells [Yamashiro et al., 1998; Brunton et al.,2001].
After 60 min in culture (Fig. 2b), cells appear moreflattened, cortical fiber bundles are less prominent, andphalloidin-stained stress fibers are abundant (Fig. 2b,arrows). In addition, small aggregates of phalloidin-stained fibers are arranged along the plasma membrane(Fig. 2b, arrowheads), suggesting that fiber bundles maybe inserted into adhesion complexes at these sites.
Fig. 2. Phalloidin staining of f-actin in newly plated D407 and IEC6cells. Thirty minutes after plating D407 cells (a) a tight cortical bandof brightly stained bundles of f-actin is observed (arrows). Lighterstaining is associated with microspikes (arrowheads). Sixty minutesafter plating D407 cells (b), stain was observed in prominent stressfibers (arrows), and at the base of apparent attachment sites (arrow-heads), possibly representing focal adhesions. Within 30 min of plat-ing IEC6 cultures (c), many cells exhibit brightly stained corticalbands (arrows) and microspikes (arrowheads). Cells that appear to bemore flattened and spread, display increased staining in stress fibers(asterisk in c).
Polyamines in RPE Cell Spreading 273
Similar Staining Patterns Observed in IEC6Intestinal Epithelial Cells
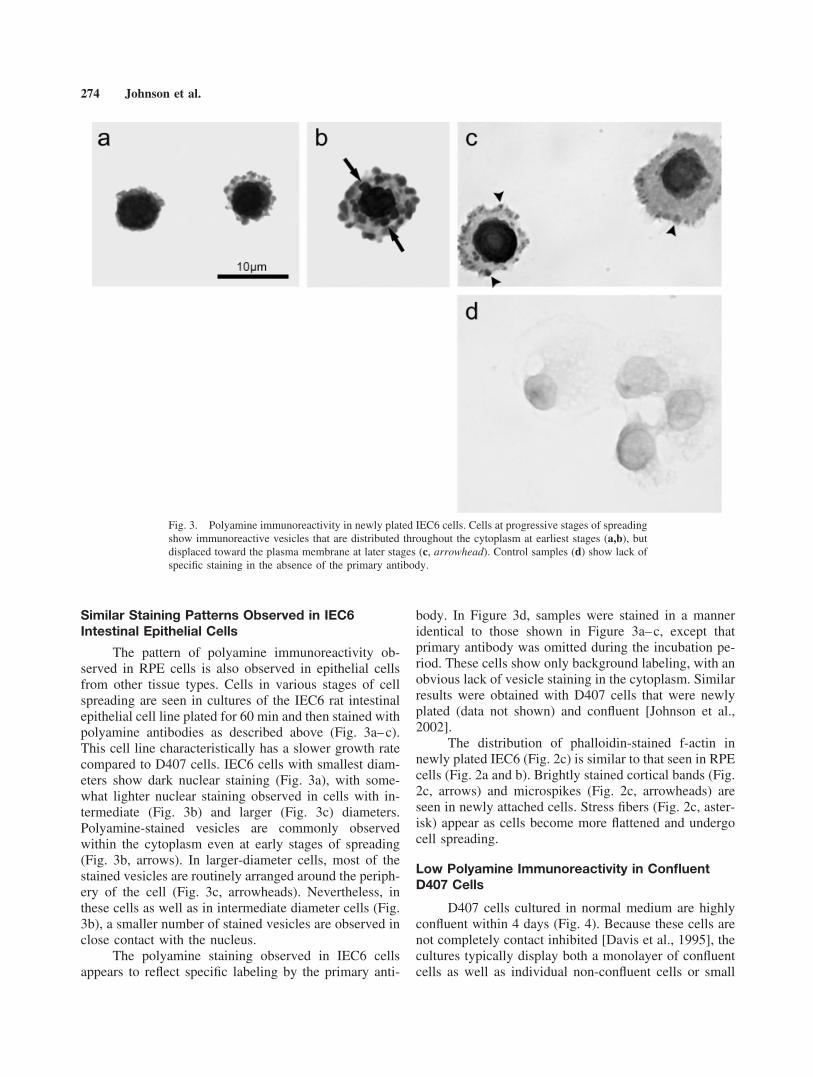
The pattern of polyamine immunoreactivity ob-served in RPE cells is also observed in epithelial cellsfrom other tissue types. Cells in various stages of cellspreading are seen in cultures of the IEC6 rat intestinalepithelial cell line plated for 60 min and then stained withpolyamine antibodies as described above (Fig. 3a–c).This cell line characteristically has a slower growth ratecompared to D407 cells. IEC6 cells with smallest diam-eters show dark nuclear staining (Fig. 3a), with some-what lighter nuclear staining observed in cells with in-termediate (Fig. 3b) and larger (Fig. 3c) diameters.Polyamine-stained vesicles are commonly observedwithin the cytoplasm even at early stages of spreading(Fig. 3b, arrows). In larger-diameter cells, most of thestained vesicles are routinely arranged around the periph-ery of the cell (Fig. 3c, arrowheads). Nevertheless, inthese cells as well as in intermediate diameter cells (Fig.3b), a smaller number of stained vesicles are observed inclose contact with the nucleus.
The polyamine staining observed in IEC6 cellsappears to reflect specific labeling by the primary anti-
body. In Figure 3d, samples were stained in a manneridentical to those shown in Figure 3a–c, except thatprimary antibody was omitted during the incubation pe-riod. These cells show only background labeling, with anobvious lack of vesicle staining in the cytoplasm. Similarresults were obtained with D407 cells that were newlyplated (data not shown) and confluent [Johnson et al.,2002].
The distribution of phalloidin-stained f-actin innewly plated IEC6 (Fig. 2c) is similar to that seen in RPEcells (Fig. 2a and b). Brightly stained cortical bands (Fig.2c, arrows) and microspikes (Fig. 2c, arrowheads) areseen in newly attached cells. Stress fibers (Fig. 2c, aster-isk) appear as cells become more flattened and undergocell spreading.
Low Polyamine Immunoreactivity in ConfluentD407 Cells
D407 cells cultured in normal medium are highlyconfluent within 4 days (Fig. 4). Because these cells arenot completely contact inhibited [Davis et al., 1995], thecultures typically display both a monolayer of confluentcells as well as individual non-confluent cells or small
Fig. 3. Polyamine immunoreactivity in newly plated IEC6 cells. Cells at progressive stages of spreadingshow immunoreactive vesicles that are distributed throughout the cytoplasm at earliest stages (a,b), butdisplaced toward the plasma membrane at later stages (c, arrowhead). Control samples (d) show lack ofspecific staining in the absence of the primary antibody.
274 Johnson et al.

groups of non-confluent cells that grow on top of themonolayer and continue to divide. In Figure 4, cells ofthe confluent monolayer show very light labeling in thenucleus and cytoplasm. As we have previously reported[Johnson et al., 2002], the only discrete polyamine stain-ing of confluent cells is seen in patches associated withthe plasma membrane at apparent points of contactamong cells. The non-confluent cells (i.e., those on top ofthe monolayer) display darker staining throughout thecytoplasm and lightly stained nuclei. Of particular inter-est are occasional cells of smaller diameter that also growatop the monolayer and exhibit vesicular staining (Fig. 4,arrows), reminiscent of that seen in newly plated cells.
Reduced Antibody Staining in DFMO TreatedCells
D407 cells that are plated and maintained in me-dium containing an inhibitor of polyamine synthesis(DFMO, 5 mM) for 60 min show decreased staining.Cultures in control medium (Fig. 5A) show the expectedlabeling patterns described above, with staining in thenucleus (arrows) and in cytoplasmic vesicles (arrow-heads). Cultures grown in the presence of DFMO (Fig.5B) show a reduction in nuclear staining and a verypronounced decrease in vesicular staining. When exog-enous polyamines (5 �M spermine) are added to theculture medium containing DFMO (Fig. 5C,D), levels ofimmunoreactivity are maintained at control levels, sug-
gesting that the added polyamines can be taken up by thecells to replace polyamine stores depleted by inhibitionof synthesis. Dark staining is observed in both nuclei(Fig. 5C, arrows) and vesicles (Fig. 5C and D, arrow-heads). The images in Figures 5A–C were obtained underidentical exposure conditions in order to allow directcomparison of staining intensity among the three images.These results add additional evidence for the specificityof the polyamine antibody used in our studies.
Double Labeling of ARPE-19 Cells WithAntibodies Against Polyamines and FAK
The ARPE-19 line of human RPE cells has growthcharacteristics that are somewhat distinct from those ofD407 cells; in particular, they exhibit more contact inhi-bition [Dunn et al., 1991; see Discussion section below).Nevertheless, polyamine staining patterns are similar be-tween the two cell lines. ARPE-19 cells that were platedfor 2 h in control medium and then harvested and stainedwith polyamine antibody and a Cy3-tagged secondaryantibody (Fig. 6A and C; red fluorescence) show varyingdegrees of spreading. There is light background stainingof the entire cell, with discrete polyamine labeling ofboth vesicles (arrows) and nuclei (arrowheads).
We also stained ARPE-19 cells with a marker foradhesion complexes, FAK. In these samples stained forFAK using a Cy2-tagged secondary antibody (Fig. 6B,D;green fluorescence), nuclei are brightly stained in manycells. Additionally, there are numerous patches of stain-ing associated with the plasma membrane (arrows).Thus, the pattern of FAK immunoreactive membranepatches we observe is similar to that described in previ-ous reports [Ray et al., 2000].
In an attempt to examine possible relationshipsbetween presumptive polyamine-labeled vesicles andFAK aggregates at adhesion complexes, we double-la-beled ARPE-19 cells with both FAK and polyamineantibodies (Fig. 6E). The merged confocal image showsdistinct labeling with the two antibodies but no evidencefor direct overlap between the two markers (which wouldresult in yellow). Patches of red and green staining areintermingled, perhaps indicating close association of theantibody binding sites that, based on size, could representboth nuclear as well as vesicular pools.
Figure 6F shows a merged confocal image ofARPE-19 cells plated and maintained in the presence of5 mM DFMO for 2 h, then double labeled with FAK(Cy2 label, green) and polyamines (Cy3 label, red) anti-bodies. As expected, inhibition of polyamine synthesisby DFMO leads to a significant reduction in polyamineimmunoreactivity (red) compared to that in cells grownin control medium (Fig. 6E). In addition, FAK immuno-reactivity (green) is diffusely distributed throughout thecell with little evidence of the stained patches seen in the
Fig. 4. Immunocytochemical localization of polyamines in confluentcultures of D407 cells. After 4 days in culture, a confluent monolayerof cells is formed, with actively dividing cells randomly distributed ontop. Staining of cells in the confluent layer is limited to small patchesof plasma membrane (arrowheads), possibly associated with sites ofattachment among neighboring cells. In contrast, actively dividingcells show enhanced staining throughout the cytoplasm, with occa-sional instances of cells with darkly stained vesicles (arrows) similarto that seen in non-confluent cultures (Fig. 1).
Polyamines in RPE Cell Spreading 275

control. These results support previous studies [Ray etal., 2000] suggesting that inhibition on polyamine syn-thesis blocks the normal activation/targeting of FAK tothe membrane.
DISCUSSION
Localization of Polyamine-ImmunoreactiveVesicles
A major new finding of this study is that a popu-lation of polyamine immunoreactive vesicles is presentin newly plated cells from two different RPE cell lines(D407 and ARPE-19) as well as from an intestinal epi-thelial cell line (IEC6). Distribution of the vesicles withinthe cell appears to change as each cell undergoes spread-ing after initial attachment. In cells that have spread to amodest degree and still retain a small cell diameter,vesicles are present in the thin rim of cytoplasm presentbetween the nucleus and the plasma membrane. In somecases, the vesicles appear to be adjacent to, or very near,the nucleus. In cells that have spread to attain a large
diameter, most polyamine vesicles are localized near theplasma membrane and are clearly distanced from thenucleus. In cells that have reached a maximum diameterand become confluent with neighboring cells, polyaminevesicles are not observed. In D407 cultures that containconfluent monolayers with individual, non-confluentcells growing on top of the confluent layer, we observedpolyamine vesicles only in the non-confluent cells, andnot in the confluent ones. Our interpretation is that poly-amine vesicles originate near the nucleus as the cellbecomes attached, and they subsequently move to theperiphery as the cell undergoes spreading. Alternatively,it is possible that the small rim of cytoplasm present inthe newly plated cells limits the initial distribution ofvesicles so that they, by necessity, appear close to boththe nucleus and the plasma membrane. In this case, thevesicles could originate from the plasma membrane andremain associated with it as the cell spreads. Observa-tions of cells at intermediate stages of spreading (Fig. 3band c) argue somewhat against this interpretation. Thelarger areas of cytoplasm present in these cells should not
Fig. 5. Polyamine immunoreactivity in ARPE-19 cells 1 h after plating in control medium (A), mediumcontaining 5 mM DFMO (B), and medium containing DFMO plus 5 �M spermine (C and at highermagnification, D). Immunoreactivity is reduced by DFMO inhibition of polyamine synthesis. This effect isblocked when spermine is added in the presence of DFMO. In this case, the staining pattern is similar to thatseen in controls, including the presence of stained vesicles located near the plasma membrane (D, arrows).
276 Johnson et al.
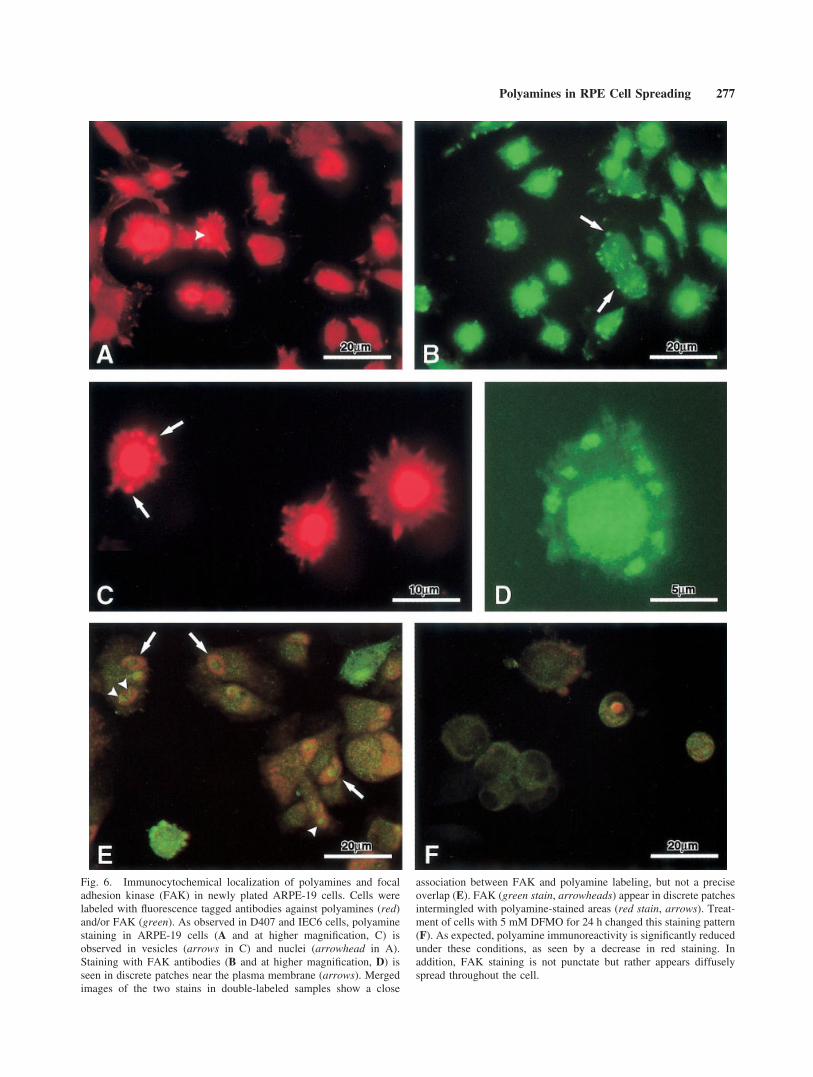
Fig. 6. Immunocytochemical localization of polyamines and focaladhesion kinase (FAK) in newly plated ARPE-19 cells. Cells werelabeled with fluorescence tagged antibodies against polyamines (red)and/or FAK (green). As observed in D407 and IEC6 cells, polyaminestaining in ARPE-19 cells (A and at higher magnification, C) isobserved in vesicles (arrows in C) and nuclei (arrowhead in A).Staining with FAK antibodies (B and at higher magnification, D) isseen in discrete patches near the plasma membrane (arrows). Mergedimages of the two stains in double-labeled samples show a close
association between FAK and polyamine labeling, but not a preciseoverlap (E). FAK (green stain, arrowheads) appear in discrete patchesintermingled with polyamine-stained areas (red stain, arrows). Treat-ment of cells with 5 mM DFMO for 24 h changed this staining pattern(F). As expected, polyamine immunoreactivity is significantly reducedunder these conditions, as seen by a decrease in red staining. Inaddition, FAK staining is not punctate but rather appears diffuselyspread throughout the cell.
Polyamines in RPE Cell Spreading 277

limit vesicle distribution, and yet they show most vesi-cles near the plasma membrane with a smaller numberstill attached to the nucleus. Whether the vesicles areactively transported through the cytoplasm duringspreading or they move passively with the expandingplasma membrane, the net result is that they are notretained near the nucleus. Furthermore, the vesicles ap-pear to be a transient feature of newly plated cells and arenot continuously generated after the cells stop spreadingand become confluent.
Staining of vesicles with polyamine antibodies hasbeen previously reported in a number of cell types in-cluding submandibular granular convoluted duct cells,pancreatic islet cells, anterior pituitary cells, and bloodplatelets [Hougaard and Larsson, 1986, 1988; Hougaardet al., 1986]. These results were obtained, not from cellcultures, but from sections of organ tissues or bloodsmears harvested from adult Wistar rats and BALB/cmice for analysis. The authors observed that not all celltypes contain polyamine vesicles; rather, vesicular stain-ing was most pronounced in cells types that secretecertain growth factors, such as epidermal growth factor(EGF), insulin, growth hormone, or platelet-derivedgrowth factor. In the two cases examined, polyamineswere localized with growth factors (EGF or insulin) insecretory granules. These authors suggest that poly-amines might facilitate the accumulation or storage ofvesicle contents and/or they may be released to act aloneor in concert with other released factors.
Polyamines and Cell Adhesions
It has been well established that polyamines areessential for cell migration and for formation of celladhesions. In intestinal epithelial (IEC6) cells, poly-amines are required for the activation RhoA and itssubsequent regulation of the activity and targeting ofFAK, the most prominent of the various protein tyrosinekinases that are linked to focal adhesion complexes [Rayet al., 2000; Guan, 1997; Parsons et al., 2000]. Uponstimulation of integrin, FAK undergoes phosphorylationand is subsequently targeted to the focal adhesion com-plex, where its additional phosphorylation provides bind-ing sites in the adhesion complex for other proteins, suchas paxillin. Paxillin, one of the actin-binding proteins,recruits actin to the complex, resulting in the formationof stress fibers [Burridge et al., 1992]. In the absence ofpolyamines, recruitment of FAK to the membrane, FAKphosphorylation, and the recruitment of paxillin to thefocal adhesion complex are all inhibited [Ray et al.,2000]. Thus, polyamine-depleted cells, even though theyare attached to the appropriate matrix, lack the integrinsignaling necessary to form viable focal adhesions andthe stress fibers essential for spreading, migration, andgrowth.
On the basis of these findings, we wished to deter-mine if polyamine vesicles are directly associated withfocal adhesions by using FAK immunoreactivity as amarker for the assembly of cell adhesions. Our analysisof RPE cells double-labeled with FAK and polyamineantibodies failed to show a strict overlap of the twostains. Thus, we have no evidence to suggest a directbinding of polyamines to activated FAK. Nevertheless,immunoreactivity of membrane-bound FAK and immu-noreactivity of polyamine vesicles were closely associ-ated, often intermingled, suggesting some degree ofphysical interaction between the two sites. If we deter-mine in future experiments that the vesicles are trulysecretory, as their general morphology, distribution, andtransient nature would suggest, we may be able to con-clude that polyamines are packaged in vesicles and thendelivered to the plasma membrane for release eitherextracellularly or intracellularly. It would also be ofinterest to determine if the proposed release is preferen-tially targeted to adhesion sites, or to emerging micro-spikes at or near adhesion sites. Our data do not demon-strate unequivocal targeting. In our confocal images,many but not all of the polyamine immunoreactive ves-icles are adjacent to (and in certain cases appear tosurround) some but not all FAK-labeled sites.
Polyamine Immunoreactivity in the Nucleus
In addition to labeled vesicles, the nuclei of mostnewly plated cells also showed specific labeling. Lessnuclear staining was observed in cells that had achievedsignificant spreading or that had reached confluence.Based on in vitro studies, polyamines are known to bindwith high affinity to nucleic acids [Davidson and Ander-son, 1960; Igarashi et al., 1977], to stabilize DNA [Flinkand Pettijohn, 1975], and to condense DNA into compactstructures [Gosule and Shellman, 1976]. Polyamines alsoappear to be required for packaging DNA in bacterio-phages and viruses [Cohen and McCormick, 1979]. Thetransient nuclear polyamine staining we observed duringearly stages of attachment suggests that nuclear bindingand stabilization by polyamines may reflect a cellularresponse to detachment (during harvesting) or attach-ment (during re-plating).
Nuclei of cells undergoing mitosis also showedstrong labeling, suggesting that polyamines in the nu-cleus may be associated with mitotic events. In previousstudies using HeLa cell cultures, there was strong stain-ing with polyamine antibodies in nuclei of cells in meta-phase and early anaphase [Hougaard et al., 1987], mitoticstages when chromatin is condensed. Staining decreasedduring anaphase and was undetectable in telophase, con-current with decondensation of chromatin. These find-ings, along with our observations, provide support for thesuggestion that in RPE and intestinal epithelial cells,
278 Johnson et al.

polyamines are concentrated in the nucleus and could beinvolved in chromatin stabilization or condensation dur-ing cell division.
Removal of polyamines from the nucleus duringdecondensation could occur through metabolism ortransport out of the nucleus. It is interesting to note thatpolyamine-labeled vesicles are found near the nucleus incells with small diameters and darkly stained nuclei (i.e.,in early stages of cell spreading), but that the vesicles arenot retained in this position as cell spreading proceeds.Rather, vesicles are aggregated near the plasma mem-brane in cells with large diameters and lightly stainednuclei (i.e., in later stages of cell spreading). Thus, re-moval of polyamines from the nucleus could conceivablybe achieved through vesicular transport from the nucleusto the periphery of the cell.
Effects of DFMO
Similar to our findings using explant cultures ofrabbit retina [Withrow et al., 2002], we also observe thatRPE cultures treated with DFMO (to inhibit polyaminesynthesis) showed a noticeable decrease in polyamineimmunoreactivity. Furthermore, addition of exogenouspolyamines along with the DFMO prevented the de-crease in both rabbit retina and RPE culture studies.These findings provide additional confirmation that theantibody staining we observe is specific for polyaminesand that exogenous polyamines accumulate in sites sim-ilar to those seen for endogenous polyamines.
The DFMO-induced decrease in endogenous poly-amines led to a decrease in the apparent targeting of FAKto the cell adhesion complex in RPE cells, suggestingthat polyamines are required for this process. This find-ing supports previous in vitro studies, which have usedDFMO to demonstrate that polyamines are required forFAK activation and targeting in intestinal epithelial cellcultures [Ray et al., 2000].
CONCLUSIONS
We hypothesize that polyamines transiently accu-mulate in the nuclei of cells that are newly harvested andre-plated and/or undergoing mitosis, at which time theymay facilitate the stabilization or condensation of chro-matin. We further speculate that polyamines are subse-quently removed from the nucleus by a process that is yetunknown, but that could involve transport, either pas-sively or actively, from the nucleus to the plasma mem-brane in vesicles. The disappearance of vesicles in cellsthat are highly spread or confluent may indicate thatpolyamines are transported and perhaps released near theplasma membrane where they could have potential intra-cellular or extracellular effects on cell attachment andformation of cell adhesions. Alternatively, polyamines
could act indirectly by influencing the accumulation/storage/release of factors that in turn have direct effectson the cell. This hypothesis is consistent with the well-documented requirement for polyamines during cell at-tachment, spreading, and migration.
ACKNOWLEDGMENTS
We are grateful to Tim O’Leary, Sarah Stinnett,and Marina Kedrov for technical assistance, to DannyMorse for assistance with preparation of the figures, andto Mary Douglas for assistance with preparation of themanuscript. This work was supported in part by theGerwin Scholar Award.
REFERENCES
Bouille N, Johnston J, Rouou A, Debroise I, Le Roch M, Renault J,Moulinoux JP, Delcros JG. 1999. Molecular analysis of thecombining site of a monoclonal antibody against spermine. MolImmunol 2:93–102.
Brunton VG, Fincham VJ, McLean GW, Winder SJ, Paraskeva C,Marshall JF, Frame MC. 2001. The protrusive phase and fulldevelopment of integrin-dependent adhesions in colon epithe-lial cells require FAK- and ERK-mediated actin spike forma-tion: deregulation in cancer cells. Neoplasia 3:215–226.
Burnside B, Bost-Usiner L. 1998. The retinal pigment epitheliumcytoskeleton. In: Marmor MF, Wolfensherger TJ, editors. Theretinal pigment epithelium. New York: Oxford UniversityPress. p 41–67.
Burridge K, Turner CE, Romer LH. 1992. Tyrosine phosphorylation ofpaxillin and pp 125FAK accompanies cell adhesion to extra-cellular matrix: a role in cytoskeletal assembly. J Cell Biol119:893–903.
Cohen SS, McCormick PP. 1979. Polyamines and virus replication.Adv Cirus Res 24:331–387.
Cooper JA. 1991. The role of actin polymerization in cell motility.Ann Rev Physiol 53:585–605.
Davidson DN, Anderson G. 1960. Chromosome coiling: abnormalitiesinduced by polyamines. Exp Cell Res 20:610–618.
Davis AA, Bernstein PS, Bok D, Turner J, Nachtigal S, Hunt RC.1995. A human retinal pigment epithelial cell line that retainsepithelial characteristics after prolonged culture. Invest Oph-thalmol Vis Sci 36:955–964.
Delcros JG, Clement S, Bouille N, Royou A, Debroise I, Thomas V,Moulinoux JP. 1996. Molecular requirements for polyaminesbinding to the antispermine monoclonal antibody Spm8-2. Hy-bridome 3:177–183.
Dunn KC, Aotaki-Keen AE, Putkey FR, Hjelmeland LM. 1991.ARPE-19, a human retinal pigment epithelial cell line withdifferentiated properties. Exp Eye Res 62:155–169.
Flink I, Pettijohn DE. 1975. Polyamines stabilize DNA folds. Nature253:62–63.
Giancotti FG, Ruoslaht E. 1999. Integrin signaling. Science 285:1028–1032.
Gosule LC, Shellman JA. 1976. Compact form of DNA induced byspermidine. Nature 259:333–335.
Guan JL. 1997. Role of focal adhesion kinase in integrin signaling. IntJ Biochem Cell Biol 29:1085–1096.
Hougaard DM, Larsson LI. 1986. Localization and possible functionof polyamines in protein and peptide secreting cells. Med Biol64:89–94.
Polyamines in RPE Cell Spreading 279

Hougaard DM, Larsson LI. 1988. Polyamines, molecules necessary forcell division, colocalize with peptide growth factors. Eur J CellBiol 48:18–28.
Hougaard DM, Nielsen JH, Larsson LI. 1986. Localization and bio-synthesis of polyamines in insulin-producing cells. Biochem J18:321–328.
Hougaard DM, Bolund L, Fujiwara K, Larsson LI. 1987. Endogenouspolyamines are intimately associated with highly condensedchromatin in vivo. A fluorescence cytochemical and immuno-cytochemical study of spermine and spermidine during the cellcycle and in reactivated nuclei. Eur J Cell Biol 44:151–155.
Igarashi K, Kashiwagi K. 2000. Polyamines: mysterious modulators ofcellular functions. Biochem Biophys Res Commun 271:559–564.
Igarashi K, Aoki Y, Hirose S. 1977. Base specificity of polyaminebinding to synthetic polynucleotides. J Biochem 81:1091–1096.
Johnson DA, Fields C, Fallon A, Fitzgerald MEC, Viar MJ, JohnsonLR. 2002. Polyamine-dependent migration of retinal pigmentepithelial cells. Invest Ophthalmol Vis Sci 43:1228–1233.
McCormack SA, Viar MJ, Johnson LR. 1993. Polyamines are neces-sary for cell migration by a small intestinal crypt cell line. Am JPhysiol Gastrointest Liver Physiol 264:G367–G374.
McCormack SA, Wang JY, Johnson LR. 1994a. Polyamine deficiencycauses reorganization of F-actin and tropomyosin in IEC-6cells. Am J Physiol 267:C715–C722.
McCormack SA, Wang JY, Viar MJ, Tague L, Davies PJA, JohnsonLR. 1994b. Polyamines influence transglutaminase activity andcell migration in two cell lines. Am J Physiol 267 (Cell Physiol.36):C706–C714.
McCormack SA, Blanner PM, Zimmerman BJ, Ray R, Poppleton HM,Patel TB, Johnson LR. 1999a. Polyamine deficiency alters EFTreceptor distribution and signaling effectiveness in IEC-6 cells.Am J Physiol Cell Physiol 274:C192–C205.
McCormack SA, Ray RM, Blanner PM, Johnson LE. 1999b. Poly-amine depletion alters the relationship of F-actin, G-actin, andthymosin b4 in migrating IEC-6 cells. Am J Physiol CellPhysiol 276:C459–C468.
Moszczynska A, Opas M. 1994. Regulation of adhesion-related pro-tein tyrosine kinases during in vitro differentiation of retinalpigment epithelial cells: Translation of pp60c-src to the nucleusis accompanied by downregulation of pp125FAK. BiochemCell Biol 72:43–48.
Nobes D, Hall A. 1995. Rho, rac, and Cdc42 GTPases regulate theassembly of multi molecular focal complexes associated withactin stress fibers, lamellipodia, and filopodia. Cell 81:53–62.
Parsons TJ, Martin KH, Slack JK, Taylor JM, and Weed SA. 2000.Focal adhesion kinase: a regulator of focal adhesion dynamicsand cell movement. Oncogene 19:5606–5613.
Pegg AE, McGovern KA, Weist L. 1987. Decarboxylation of alpha-difluoro-methyl ornithine by ornithnine decarboxylase. Bio-chem J 241:305–307.
Pegg AE, Kamejt T, Shirahata A, Stanley B, Madhubale R, andPajunen A. 1988. Regulation of mammalian S-adenosylmethi-onine decarboxylase. Adv Enzyme Regul 27:43–55.
Pfeffer LM, Yang CH, Pfeffer SR, Murti A, McCormack SA, JohnsonLR. 2000. Inhibition of ornithine decarboxylase inducesSTAT3 tyrosine phosphorylation and DNA binding in IEC-6cells. Am J Physiol Cell Physiol 278:C331–335.
Ray RM, Viar MJ, McCormack SA, Johnson LR. 2000. Focal adhe-sion kinase signaling is decreased in polyamine-depleted IEC-6cells. Am J Physiol Cell Physiol 281:C475–C485.
Ray RM, McCormack SA, Covington C, Viar MJ, Zheng Y, JohnsonLR. 2003. The requirement for polyamines for intestinal epi-thelial cell migration is mediated through rac1. J Biol Chem278:13039–13046.
Ridley AJ, Hall A. 1992. The small GRP-binding protein Rho regu-lates the assembly of focal adhesions and stress fibers in re-sponse to growth factors. Cell 70:389–399.
Ridley AJ, Paterson HF, Johnston CL, Diekmann D, Hall A. 1992. Thesmall of GRP binding protein Rac regulates growth-factorinduced membrane ruffling. Cell 70:401–410.
Russell DH 1985. Ornithine decarboxylase. A key regulatory enzymein growth. Proc Bong Res Ther 8:43–58.
Seiler N. 1994. Formation, catabolism and properties of the naturalpolyamines. In: Carter C, editor. Neuropharmacology of poly-amines. London: Harcourt Brace & Company. p 1–36.
Shaw GG. 1994. Polyamines as neurotransmitters or modulators. In:Carter C, editor. The neuro-pharmacology of polyamines. Lon-don: Harcourt Brace & Company. p 61–80.
Symons M. 1996. Rho family GTPases: the cytoskeleton and beyond.TIBS 21:178–181.
Wang T, Milam AH, Steel G, Valle D. 1996. A mouse model of gyrateatrophy of the choroid and retina. Early retinal pigment epithe-lium damage and progressive retinal degeneration. J Clin Invest97:2753–2762.
Withrow C, Ashraf S, O’Leary T, Johnson LR, Fitzgerald MEC,Johnson DA. 2002. Effect of polyamine depletion on conephotoreceptors of the developing rabbit retina. Invest Ophthal-mol Vis Sci 43:3081–3090.
Yamashiro S, Yamakita Y, Ono S, Matsumura F. 1998. Fascin, anactin-bundling protein, induces membrane protrusions and in-creases cell motility of epithelial cells. Mol Biol Cell 9:993–1006.
Young RW. 1987. Pathophysiology of age-related macular degenera-tion. Surv Ophthalmol 31:291–306.
280 Johnson et al.