immunostaining for substance p receptor labels gabaergic cells with distinct termination patterns in...
TRANSCRIPT

Immunostaining for Substance PReceptor Labels GABAergic Cells WithDistinct Termination Patterns in the
Hippocampus
L. ACSADY,1 I. KATONA,1 A.I. GULYAS, 1 R. SHIGEMOTO,2 AND T.F. FREUND1*1Institute of Experimental Medicine, Hungarian Academy of Sciences,
Budapest, H-1450, Hungary2Department of Morphological Brain Science, Faculty of Medicine,
Kyoto University, Kyoto 606-01, Japan
ABSTRACTA specific antiserum against substance P receptor (SPR) labels nonprincipal neurons in
the cerebral cortex of the rat (T. Kaneko et al. [1994], Neuroscience 60:199–211; Y. Nakaya etal. [1994], J. Comp. Neurol. 347:249–274). In the present study, we aimed to identify the typesof SPR-immunoreactive neurons in the hippocampus according to their content of neurochemi-cal markers, which label interneuron populations with distinct termination patterns.Markersfor perisomatic inhibitory cells, parvalbumin and cholecystokinin (CCK), colocalized with SPRin pyramidallike basket cells in the dentate gyrus and in large multipolar or bitufted cellswithin all hippocampal subfields respectively. A dense meshwork of SPR-immunoreactivespiny dendrites in the hilus and stratum lucidum of the CA3 region belonged largely toinhibitory cells terminating in the distal dendritic region of granule cells, as indicated by thesomatostatin and neuropeptide Y (NPY) content. In addition, SPR and NPY were colocalizedin numerous multipolar interneurons with dendrites branching close to the soma. Twenty-fivepercent of the SPR-immunoreactive cells overlapped with calretinin-positive neurons in allhippocampal subfields, showing that interneurons specialized to contact other gamma-aminobutyric acid-ergic cells may also contain SPR. On the basis of the known terminationpattern of the colocalized markers, we conclude that SPR-positive interneurons are function-ally heterogeneous and participate in different inhibitory processes: (1) perisomatic inhibitionof principal cells (CCK-containing cells, and parvalbumin-positive cells in the dentate gyrus),(2) feedback dendritic inhibition in the entorhinal termination zone (somatostatin andNPY-containing cells), and (3) innervation of other interneurons (calretinin-containing cells).J. Comp. Neurol. 378:320–336, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: tachykinins; neuropeptides; receptors; colocalization; interneurons
Combined morphological and electrophysiological stud-ies have revealed that interneurons with different termina-tion patterns are responsible for diverse inhibitory pro-cesses in the hippocampus (Lacaille et al., 1987; Lacailleand Schwartzkroin, 1988a,b; Buzsaki et al., 1992; Gulyaset al., 1993a,b; Soltesz and Deschenes, 1993; Buhl et al.,1994; McBain et al., 1994; Bragin et al., 1995; Buckmasterand Schwartzkroin, 1995; Cobb et al., 1995; Sık et al.,1995; Ylinen et al., 1995; Miles et al., 1996). The functionalclassification of the interneuron populations became in-creasingly difficult becausemorphologically identified typesand immunocytochemically characterized populationswereoften not or poorly correlated. The relationship to physi-ological characteristics was even more difficult to estab-lish, but there have been encouraging attempts (Sık et al.,
1995). Recent studies on the termination pattern of hippo-campal interneurons showed that on the basis of their rolein the inhibitory networks interneurons can be classifiedinto three basic categories, each consisting of severalsubtypes. The first two kinds of interneurons contactprincipal cells in two different membrane domains, i.e., inthe perisomatic region (basket and chandelier cells) and in
Contract grant sponsor: Human Frontier Science Program Organization;Contract grant sponsor: The Howard Hughes Medical Institute; Contractgrant sponsor: OTKA; Contract grant number: T 16942.*Correspondence to: Tamas F. Freund, Institute of Experimental Medi-
cine, Hungarian Academy of Sciences, Budapest, P.O. Box 67, H-1450,Hungary. E-mail: [email protected] 11 June 1996; Revised 18 September 1996; Accepted 27
September 1996
THE JOURNAL OF COMPARATIVE NEUROLOGY 378:320–336 (1997)
r 1997 WILEY-LISS, INC.

the dendritic region (Han et al., 1993; Gulyas et al.,1993a,b; Buhl et al., 1994; Miles et al., 1996), and the thirdkind is specialized to innervate other interneurons (Ac-sady et al., 1996b; Gulyas et al., 1996; Hajos et al., 1996).Electrophysiological and modeling studies ascribe differ-ent roles for somatic and dendritic inhibition and to thelocal inhibitory control of gamma-aminobutyric acid(GABA)-ergic cells. Basket and axo-axonic cells innervatethe perisomatic region of pyramidal cells (Somogyi et al.,1983, 1985; Gulyas et al., 1993a; Buhl et al., 1994) andhave profound effects on the timing and repetitive firing ofsodium-dependent action potentials of pyramidal cells(Cobb et al., 1995; Miles et al., 1996). The axons of den-dritic inhibitory cells innervate the dendritic surface of thepyramidal cells in conjunction with various excitatoryafferents (Gulyas et al., 1993a,b, 1996; Han et al., 1993;Buhl et al., 1994; Sık et al., 1995) andmay regulate calcium-dependent dendritic electrogenesis (Miles et al., 1996).Interneurons of the third category terminate almost exclu-sively on other interneurons (Acsady et al., 1996b; Gulyaset al., 1996; Hajos et al., 1996). Specific interactions amongdifferent sets of GABAergic cells may be involved in thehigh-frequency population oscillations and/or in disinhibi-tion (Muller and Misgeld, 1990; Michelson and Wong,1991; Buzsaki and Chrobak, 1995; Whittington et al.,1995). The neurochemical marker content (neuropeptidesand calcium-binding proteins) of interneurons shows re-markable correlation with these three categories. Parval-bumin and cholecystokinin (CCK) are present in twoseparate types of perisomatic inhibitory cells (Nunzi et al.,1985; Kosaka et al., 1987; Acsady et al., 1996a). Calbindin-immunoreactive interneurons innervate the proximal den-drites of pyramidal cells, and somatostatin- and neuropep-tide Y (NPY)-containing interneurons contact pyramidalcells in the distal dendritic region (Kohler et al., 1986;Deller and Leranth, 1990; Leranth et al., 1990; Buckmas-ter et al., 1994; Sık et al., 1995, 1996; Gulyas and Freund,1996; Katona et al., 1996). The majority of vasoactiveintestinal polypeptide- (VIP) and calretinin-containingcells selectively innervate other interneurons (Acsady etal., 1996b; Gulyas et al., 1996; Hajos et al., 1996). Thus,neurochemical identification provides a valuable tool tostudy large population of inhibitory cells with similarcharacteristics. Large numbers of nonprincipal cells havebeen visualized in the hippocampus by an antiserumagainst substance P receptor (SPR; Kaneko et al., 1994;Nakaya et al., 1994). Whether this antigen functions as areceptor for tachykinins in these locations remains to beestablished because there is considerable mismatch withthe distribution of the known endogenous ligands (Liu etal., 1994). However, the great detail of the morphologyprovided by the immunolocalization of SPR encourages itsuse as a marker in connectivity studies, even before thefunction of this protein is understood. The functionalclassification of SPR-immunoreactive neurons based on
the termination pattern was hindered by the lack of axonalstaining. In previous experiments, combined morphologi-cal and neurochemical studies of GABAergic cells wereuseful in separating functionally different sets of interneu-rons labeled by the same marker (Acsady et al., 1996a,b).Using similar methods, our aim in the present study wasto identify the morphologically heterogeneous SPR-immunoreactive neurons in the hippocampus according tothe content of neurochemical markers and to establish thetypes of inhibitory circuits they are involved in.
METHODS
Ten male Wistar rats (300–350 g, 2 months old; CharlesRiver, Budapest, Hungary) were deeply anesthetized withEquithesin (chlornembutal, 0.3 ml/100 g) and perfusedthrough the heart first with saline and then with phos-phate buffered (PB; 0.1 M) fixative containing 4% parafor-maldehyde, 0.2% picric acid, and 0.05% glutaraldehyde.Coronal sections (60 µm thick) were cut from the hippocam-pus on a Vibratome, kept in sequence, washed, cryopro-tected in 30% sucrose in 0.1 M PB overnight, and freezethawed over liquid nitrogen. Following extensive washes,the sections were incubated in one of the following anti-sera: for single immunostaining, rabbit anti-SPR (1:300,1.6 mg/ml; Shigemoto et al., 1993), rabbit anti-somatosta-tin (1:20,000; Lantos et al., 1995; or 1:300, Dakopatts),rabbit anti-NPY (1:20,000; Csiffary et al., 1990), rabbitanti-calretinin (1:5,000; Rogers, 1989), rabbit anti-VIP(1:10,000; Gulyas et al., 1990), rabbit anti-CCK (1:3,000;Gulyas et al., 1990), or rabbit anti-GABA (1:20,000; Beau-lieau et al., 1994) was used as primary antiserum (seeTable 1A). The secondary antibody was biotinylated anti-rabbit IgG made in goat (Vector; 1:200, 2 hours), followedby avidin biotinylated-horseradish peroxidase complex(ABC; Vector; 1.5 hours, 1:150). All the washes and dilu-tion of antisera were done in 0.05 M Tris buffered saline(TBS), pH 7.4. The immunoperoxidase reaction was devel-oped with 3,38-diaminobenzidine (DAB) as a chromogen.The sections were treated with 1% OsO4 in 0.1 M PB for 15minutes, dehydrated in ethanol and propylene oxide, andembedded in Durcupan (ACM, Fluka). The characteriza-tion of the SPR antiserum is described in Nakaya et al.(1994). Briefly, the antigen is a trpE-fusion protein thatcontains rat SPR sequence (amino acid residues 349–407).Cross reactivity with NK2 and NK3 receptors was ex-cluded by immonoblot and staining analyses of the recep-tor transfected cell lines (Shigemoto et al., 1993). Thedistribution of SPR immunoreactivity is generally veryconsistent with the autoradiography of SP-binding sites(Mantyh et al., 1989). The specificity of the other primaryantisera have been tested by the laboratories of origin (seereferences above). Replacing the primary antisera withnormal sera of the animals in which the primary antiserawere raised resulted in lack of immunostaining. SPR-immunoreactive cells were reconstructed with a drawingtube from serial 60-µm-thick sections. To study the coexist-ence of SPR with calretinin, CCK, somatostatin, VIP, NPY,and GABA, the mirror technique of Kosaka et al. (1985)was used. Because no obvious difference was found be-tween the dorsal and ventral parts of the hippocampus, theanalysis was carried out in the dorsal hippocampus (5–10sections/animal, 4–7 animals/colocalization studied). Adja-cent sections were treated for SPR or one of the otherantigens, and the cell bodies cut in half were identified on
Abbreviations
DG gyrus dentatusCCK cholecystokininCB calbindin D28kCR calretininNPY neuropeptide YPV parvalbuminSOM somatostatinVIP vasoactive intestinal polypeptide
SPR-CONTAINING INTERNEURONS IN THE HIPPOCAMPUS 321

the common surfaces of both sections by using capillariesas landmarks. First, bisected SPR-positive cell bodies wereidentified on the surface of the sections by using 1003oil-immersion objective. Second, the corresponding halvesof the cells were looked for on the matching surface of theadjacent section. Cells were only included in the analysis ifthe matching half could be unequivocally identified(whether negative or positive). In the case of antibodiesthat gave excellent dendritic staining, this method couldbe extended to examine cut main dendrites, as describedpreviously (Acsady et al., 1996a). For fluorescent doublestaining, the following mixtures of antisera were used (seeTable 1B): rabbit anti-SPR (1:200, 2.5 mg/ml) and mouseanticalbindin (1:2,000; Celio, 1990) or rabbit anti-SPR andmouse antiparvalbumin (1:500; Celio, 1990). In all cases,the second layer was lissamine-rhodamine sulfonyl chlo-ride (LRSC)-conjugated anti-rabbit IgG made in donkey(Jackson, 1:50) and fluorescein-conjugated anti-mouse IgGmade in goat (1:50; Dakopatts). The sections weremountedon glass slides in Vectashield (Vector) embedding medium.The results were obtained from 9–12 sections from threeanimals and evaluated with a Leitz Laborlux microscopewith Ploemopak fluorescence illuminators using filters forfluorescein isothiocyanate (type I3, exciting filter band-pass 450–490) and LRSC (type N2, exciting filter bandpass530–560).
RESULTS
Morphological characterization
Immunostaining for SPR labeled numerous, morphologi-cally heterogeneous nonprincipal cells in the hippocampusas reported previously (Nakaya et al., 1994). The immuno-precipitate was membrane-bound and visualized cell bod-ies and proximal and distal dendrites in a Golgi-like
manner. Various types of dendritic appendages were alsolabeled. Principal cells (granule, mossy, and pyramidalcells) were consistently devoid of SPR immunoreaction inthe hippocampus. Ventral subiculum showed dense neuro-pil staining. SPR-positive axons were not present in thematerial. The morphology of SPR-containing cells showedconsiderable variation within and among the hippocampalsubfields.
Dentate gyrus
The most frequently encountered SPR-positive cell typein the dentate gyrus belonged to the pyramidal-like cells(Fig. 1A,D). The medium-sized pyramidal-shaped cell bod-ies of this cell type were situated mainly in the deepstratum (str.) granulosum or at the border of str. granulo-sum and the hilus. Their single, pronounced apical den-drites crossed str. granulosum and branched profusely instr. moleculare. The two or three thinner basal dendritesentered the hilus. The dendrites of this cell type wereaspiny. Besides pyramidal-like cells, large multipolar orfusiform SPR-immunoreactive cells were also occasionallypresent in str. granulosum with aspiny dendrites enteringstr. moleculare and the hilus. Stratum moleculare con-tained a few stellate or fusiform SPR-positive cells, withthin primary dendrites largely confined to this layer.
Hilus of the dentate gyrus
The most prominent SPR-immunostaining was presentin the hilus (Fig. 1A). The profuse dendritic arbor ofnumerous SPR-immunoreactive cells formed a densemesh-work filling the hilus. Close examination of this regionshowed that the hilar plexus consists of morphologicallydifferent cell types. The most characteristic hilar SPR-containing cells were large, intensely stained, spiny neu-rons (Fig. 1B–C). Two three-thick primary dendrites origi-nated from the fusiform somata of these cells, runningparallel to the granule cell layer. The dendrites of the spinySPR-positive cells were restricted mainly to the hilus: theynever penetrated the granule cell layer but occasionallyentered str. radiatum of the CA3c region. The entiredendritic arbor and the somata of these cells were denselycovered with spines. The spines had thin, long, sometimesbranching, necks and small heads. Forty percent of all theSPR-containing cells belonged to this type in the hilus.Thorny excrescences, characteristic of mossy cells, wereencountered neither on this nor on any other hilar SPR-containing cell types. The hilus contained various smoothdendritic SPR-positive interneurons. Medium-sized multi-polar neurons with thick primary dendrites and stellate-like cells with thin primary dendrites were distinguish-able. In contrast to the spiny hilar SPR-positive cells, thedendrites of aspiny interneurons were radially orientedand regularly crossed str. granulosum and branched in str.moleculare.
The CA3 region
A large number of SPR-immunoreactive interneuronswere found in all strata of the CA3 region. The mostabundant cell type in all layers of the CA3 region wereaspiny multipolar SPR-immunoreactive cells, with radialthin primary dendrites (Fig. 2A). The 5–7 primary den-drites of these cells branched in close proximity to theround or irregularly shaped soma. Various larger multipo-lar or bitufted SPR-containing interneurons were alsoobserved with thick, more distally branching, primary
TABLE 1A. Antibodies Used for Preembedding Single Staining1
Primary antibodiesand dilutions
Source ofprimary antibody
Secondary antibody,source and dilution
Rabbit-anti Substance Preceptor (1:300, 1.6 µg/ml)
Shigemoto et al., 1993 Biotinylated anti-rabbit IgGmade in goat (1:200), Vector
Rabbit anti-somatostatin(1:20000)
Lantos et al., 1995
Rabbit anti-somatostatin(1:300)
(DAKOPATTS)
Rabbit anti-NPY (1:20000) Csiffary et al., 1990Rabbit anti-calretinin(1:5000)
Rogers, 1989
Rabbit anti-VIP (1:10000) Gulyas et al., 1990Rabbit anti-CCK (1:3000) Gulyas et al., 1990Rabbit anti-GABA (1:20000) Beaulieau et al., 1994
TABLE 1B. Antibodies Used for Double Immunofluorescence
Primary antibodyand its dilution
Source ofprimary antibody
Secondaryantibody andits dilution
Source ofsecondaryantibody
Rabbit-anti Sub-stance P receptor(1:200, 2.5 µg/ml)
Shigemoto et al.,1993
Lissamine-rho-damine sulfonylchloride-conju-gated anti-rabbitIgG made indonkey (1:50)
Jackson
Mouse anti-calbindin(1:2000)
Celio, 1990 Fluorescein-conju-gated anti-mouseIgG made in goat(1:50)
DAKOPATTS
Mouse anti-parval-bumin (1:500)
Celio, 1990 Fuorescein-conju-gated anti-mouseIgG made in goat(1:50)
DAKOPATTS
1Abbreviations: CCK: cholecystokinin, NPY: neuropeptide Y; VIP: vasoactive intestinalpolypeptide, SPR.
322 L. ACSADY ET AL.

Fig 1. A: Low magnification light micrograph of the dentate gyrusimmunostained for substance P receptor (SPR). Note the extremelydense meshwork of dendrites in the hilus, which is formed mainly byspiny dendrites of large fusiform cells (see B–C) and the pyramidal-like cells (arrowheads in D) at the hilar border of the granule cell layer.B,C: Spiny cells in the hilus restricted their dendrites to the hilus(arrowheads in B) and always proved to be positive for somatostatin.
D: In contrast, dendrites of SPR-positive pyramidallike cells in str.granulosum (s.g.) are smooth and extend into str. moleculare. Approxi-mately half of the SPR-immunoreactive cells in str. granulosumcontained parvalbumin (see Fig. 5), demonstrating that they areperisomatic inhibitory cells. Scale bars 5 100 µm in A,C, 30 µm in B,20 µm in D.

dendrites. A characteristic subgroup of these cells confinedthe majority of their dendrites to str. oriens and had a fewstubby spines. Stratum lucidum contained numerous SPR-positive spiny dendrites restricted to this layer and run-ning parallel to str. pyramidale. These dendrites origi-nated from large spiny cells, with morphology similar tohilar spiny SPR-positive neurons (Fig. 2B).
The CA1 region
The CA1 region contained multipolar cells of varioussizes (Fig. 2C). The aspiny multipolar cells looked similarto their counterparts in the CA3 region. Their aspinydendrites branched near the cell bodies and arborized inthe vicinity of the soma in a stellatelike manner (Fig. 3A).In contrast, the thick primary dendrites of large, darklystained, multipolar or bitufted cells (Fig. 2D) crossedseveral layers and occasionally bore large numbers ofspines. In the CA1 region, numerous SPR-positive den-drites ran horizontally at the border of str. radiatum andstr. lacunosum-moleculare, but relatively few entered str.lacunosum-moleculare. In summary, four major cell typeswere distinguished throughout the hippocampus on thebasis of location and dendritic morphology: (1) spiny,horizontal, fusiform cells were confined to the hilus andstr. lucidum of the CA3 region, (2) pyramidal-shaped cellsor fusiform cells, present only in str. granulosum of thedentate gyrus, (3) aspiny multipolar cells with thin radial,primary dendrites branching close to the soma, and (4)larger, more robust aspiny or sparsely spiny multipolar orbitufted cells with thick, more distally branching den-drites. Several SPR-positive cells showed mixed featuresof types 3 and 4. These multipolar cells were presentthroughout all subfields and all layers in the hippocampus.Because SPR immunocytochemistry provides no axonalstaining to investigate the role of various SPR-positiveinterneurons in the hippocampal network and to examinethe correlation of morphology and neurochemical charac-teristics, colocalization studies were carried out with neu-rochemical markers that label well-defined, functionallydistinct sets of GABAergic cells and with GABA itself.
Colocalization of SPR with GABA
The mirror technique of Kosaka et al. (1985; see Meth-ods) was used to establish the colocalization of SPR andGABA. From 103 SPR-immunoreactive interneurons, 92(89%) were unequivocally GABA positive. The remaining11 GABA-negative or weakly positive neurons all belongedto the spiny fusiform cell type situated in the hilus and str.lucidum of the CA3 region (Fig. 3B–C). This numberrepresented 67% of all the hilar spiny SPR-positive neu-rons (n 5 17). The hilar region contains a large number ofneurons with distant projections in which GABAcannot bedetected by immunocytochemical methods, at least in theirsoma. Thus, the GABA negativity of these cells might beexplained by their long projecting nature.
Colocalization with markers of interneuronsinnervating the perisomatic region ofpyramidal cells (CCK, parvalbumin)
Parvalbumin and CCK label discrete interneuron popu-lations with different calcium-binding protein and neuro-peptide content and with distinct intrahippocampal andextrahippocampal connections but with similar targetselectivity, i.e., the perisomatic region of pyramidal cells.In the hippocampus, parvalbumin is present in basket and
axo-axonic cells, whereas CCK labels another nonoverlap-ping population of basket cells (Gulyas et al., 1991, 1996;Acsady et al., 1996a). The CCK–SPR colocalization wasexamined by the mirror technique, and fluorescent doublestaining was used to establish the coexistence of parvalbu-min and SPR. The CCK and parvalbumin staining wassimilar to the results of others (Baimbridge and Miller,1982; Nunzi et al., 1985; Kosaka et al., 1987; Celio, 1990).Briefly, CCK labeled mainly basket cells scattered throughall layers and all regions of the hippocampus, and parval-bumin immunofluorescence was present in basket andaxo-axonic cells of the dentate gyrus, hilus, and in str.pyramidale and oriens of the Ammon’s horn. Nearly allCCK-immunoreactive cells (n 5 67) showed SPR immu-nopositivity in all regions, whcih represented 4–19% of allSPR-containing interneurons in the various subfields (seeTable 2). The majority of the CCK1/SPR1 cells in allregions were large, darkly stained, multipolar or bituftedcells with thick primary dendrites, which occasionallycrossed several layers without branching (Fig. 4). Thedendrites of CCK1/SPR1 interneurons rarely penetratedstr. lacunosum-moleculare and were occasionally deco-rated with substantial numbers of spines in the CA1region. Some of the SPR-immunoreactive pyramidal-shaped or fusiform cells in the dentate gyrus also con-tained CCK. Significant colocalization between SPR andparvalbumin was only found in the pyramidal-shaped orfusiform cells of the dentate gyrus (Fig. 5A–B). Parvalbu-min-containing cells in str. granulosum of the dentategyrus contained SPR in 90% of the cases (n 5 69), whichrepresented 59% of the SPR-immunoreactive cells in thislayer (n 5 78). In sharp contrast, only 5% of the SPR-positive interneurons colocalized parvalbumin in the hilusand the Ammon’s horn (n 5 190), which corresponds to 6%of parvalbumin-positive cells in these regions (n 5 129).
Colocalization with markers of interneuronsinnervating the dendritic region of pyramidal
cells (calbindin, somatostatin, NPY)
The colocalization of SPR-somatostatin and SPR–NPYwere studied by the mirror technique, and fluorescentdouble staining was used to establish the coexistence ofSPR and calbindin. The distribution of the interneuronslabeled by the markers were similar to the earlier results.Briefly, somatostatin-containing cells were present in thehilus and in str. oriens of CA3 and CA1 regions (Morrisonet al., 1982; Kohler and Chan-Palay, 1983). NPY-positivecells showed a similar distribution; however, a number ofcells were situated in str. pyramidale and radiatum of theCA1 region and at the base of granule cell layer (Kohler etal., 1986; Deller and Leranthh, 1990). Antisera to calbin-din stained the granule cells of the dentate gyrus, the CA1pyramidal cells, and scattered interneurons in the den-dritic region of the principal cells (Baimbridge and Miller,1982; Celio, 1990; Toth and Freund, 1992). SPR andsomatostatin colocalization showed large regional hetero-geneity (Table 2). In the hilus and str. lucidum of the CA3region, all large spiny fusiform SPR-containing cells (n 525) were immunoreactive for somatostatin (Fig. 6A–C). Inthe hilus, these cells represented 73% of all somatostatin-positive cells (n 5 25). In contrast, in the CA1 region only8% of the somatostatin-positive cells colocalized SPR. Inaddition to spiny cells in the hilus and str. lucidum of theCA3 region, somemultipolar SPR-positive cells also colocal-ized with somatostatin in str. oriens of CA3 region (Fig.
324 L. ACSADY ET AL.

Fig. 2. A: Low magnification light micrographs of the CA3 regionimmunostained for SPR. Stratum lucidum is outlined by a densenetwork of SPR-positive dendrites (arrowheads). The border betweenCA3 and CA1 is indicated by a dashed line. CA1 contains less intensivedendritic staining than CA3. B: The dense network of SPR-containingdendrites in str. lucidum (s.l.) mainly contains long, tangentiallyrunning spiny dendrites (arrowheads, inset) of large SPR-immunore-active cells. C: Low magnification light micrograph of the CA1 regionimmunostained for SPR showing the morphological heterogeneity of
positive cells. Arrows point to large multipolar cells (see Fig. 3), whichwere sparsely spiny and always cholecystokinin (CCK) positive.Arrowheads identify small smooth dendritic cells, which were eithercalretinin positive or contained neuropeptide Y (NPY). D: Lightmicrograph of a group of large multipolar SPR-positive cells in theCA1 region. This type proved to be CCK-positive basket cells. s.p., str.pyramidale. Scale bars 5 200 µm inA, 50 µm in B, 100 µm in C, 50 µmin D.

Fig. 3. A: Camera lucida drawings of small multipolar SPR-immunoreactive neurons in the CA1 region. These cells have manyprimary dendrites, pronounced dendritic tufts close to the cell body,and frequently contain NPY. B–C: Colocalization of SPR and gamma-aminobutyric acid (GABA) immunoreactivity in different neurons ofthe dentate gyrus. A large spiny SPR-positive cell (s1) is only faintly
positive for GABA, whereas an SPR-containing pyramidallike cell (s2)shows strong GABA immunoreactivity. Arrow points to a GABA-positive SPR-negative interneuron with a similar soma shape andlocation as s2. Granule cells are negative for both antigens. c1 and c2label capillaries that help identify the cells. str. rad., str. radicum; str.pyr., str. pyramidale; str. ori., str. oriens. Scale bars 5 20 µm.

6D–E). The thick, distally branching primary dendrites ofthese cells were confined to str. oriens and were occasion-ally decorated with stubby spines. Many of the spine-freedendrites belonging to SOM1SPR1 cells became spinyupon entering or crossing str. lucidum. The morphology ofinterneurons containing bothNPY and SPR differed amongthe different subfields of the hippocampus. In the dentategyrus, pyramidallike SPR-immunoreactive cells containedNPY in 28% of the cases (n 5 14). In the hilus and str.lucidum of CA3, nearly all large spiny SPR-positive cellsshowed NPY immunoreactivity (92%, n 5 25), also demon-strating the colocalization of NPY and somatostatin in thiscell type (Fig. 7). From the remaining smooth dendriticmultipolar SPR-positive cells, an average of 36% containedNPY (n 5 74) for the whole hippocampus. The highestcolocalization rate was found in the CA1 region (60%) andthe lowest in the CA3 region (12.5%). These SPR1/NPY1multipolar cells always belonged to the aspiny multipolarcell type, with thin primary dendrites branching close tothe soma, and showed consistently weaker immunostain-ing for NPY than did SPR-negative NPY-containing cells.Calbindin was present in only a very small fraction of theSPR-immunoreactive neurons (6%, n 5 318; Fig. 5C–F).These multipolar cells in the CA3 region emitted thick,rarely branching, dendrites largely confined to str. oriensand had a few stubby spines. The SPR immunoreactivity ofthe hilar calbindin-positive interneurons could not bereliably established due to the intense calbindin immuno-fluorescence of the mossy fibers.
Colocalization with markers of interneuronsspecialized to innervate other interneurons
(calretinin, VIP)
Themajority of calretinin- and VIP-containing interneu-rons selectively innervate other GABAergic cells in thehippocampus (Acsady et al., 1996b; Gulyas et al., 1996;Hajos et al., 1996). Their colocalization with SPR wasexamined by using the mirror technique. In the wholehippocampus, 25% of the SPR-positive cells (n 5 102)displayed calretinin immunoreactivity, with little regionalvariation, which represented 40% of the calretinin-containing cells. The SPR1/calretinin1 cells were aspinymultipolar cells with dendrites branching close to the
soma (Fig. 8). Multiple dendrodendritic contacts, a charac-teristic feature of some calretinin-positive cells (Gulyas etal., 1992), were rarely observed. Calretinin also labels spinycells (Gulyas et al., 1992), which display anatomical featuressimilar to spiny SPR-positive cells (i.e., dendrites are coveredwith long spines and are restricted to the hilus). However,from the 52 spiny SPR-immunoreactive cells, only four con-tained calretinin.VIP-containing cells formtwomajormorpho-logically and neurochemically different cell populations in thehippocampus (Acsady et al., 1996a,b). Small bipolar or oligopo-lar VIP-containing cells, which innervate other GABAergiccells, never contained SPR. However, large multipolar VIP-positive interneurons that innervate the perisomatic region ofpyramidal cells always showed SPR immunoreactivity. Thisobservation is consistent with earlier results, because VIP-containing basket cells were reported to colocalize CCK, andnearly all CCK-positive cells contain SPR.
DISCUSSION
In the present study, we have shown that in the hippo-campus SPR labels several morphologically, neurochemi-cally, and functionally heterogeneous interneuron popula-tions. GABAergic cells of all the three major interneuronclasses, i.e., perisomatic and dendritic inhibitory cells andGABAergic cells specialized to contact other interneurons,were found among the SPR-immunoreactive neurons.Nearly all CCK-positive interneurons contained SPR,whereas parvalbumin colocalized with SPR only in str.granulosum of the dentate gyrus but not in other subfields.In the hilus and str. lucidum of the CA3 region, somatosta-tin andNPYwere present in large spiny SPR-immunoreac-tive cells. In contrast, most of the somatostatin-positivecells were devoid of SPR in the CA1 region. Calretinin-immunoreactive cells also showed considerable overlapwith SPR-positive neurons in all regions. Previous studiesof interneuron populations labeled by various neurochemi-cal markers have shown that in most cases neuropeptidesand calcium-binding proteins label homogeneous popula-tion of GABAergic cells (Baimbridge and Miller, 1982;Somogyi et al., 1984; Roberts et al., 1984; Kosaka et al.,1987; Sloviter and Nilaver, 1987; Celio 1990; Gulyas et al.,1992). Moreover, the types of interneurons immunoreac-tive for a given marker were remarkably similar amonghippocampal subfields. For example, the calcium-bindingprotein parvalbumin is present in basket and axo-axoniccells in the whole hippocampus (Kawaguchi et al., 1987),whereas the dendritic and somatic distribution of somato-statin-containing cells is specialized for a role in feedbackinhibition of distal principal cell dendrites in all subfields(Baude et al., 1993; McBain et al., 1994; Blasco-Ibanez andFreud, 1995; Sık et al., 1995, 1996). Occasionally, the sameproteins and peptides (e.g., VIP and calretinin) may bepresent in a heterogeneous population of interneurons.However, in these cases, correlation of morphological andneurochemical features identify subpopulations with dis-tinct connectivity and function (Gulyas et al., 1992;Acsadyet al., 1996a,b; Hajos et al., 1996). The high degree ofheterogeneity of SPR-immunoreactive cells found in thisstudy is incomparable to that of any other markers.
SPR-immunoreactive cells may contain anyof the examined markers
In the present study, all seven neurochemical markersshowed various degrees of colocalization with SPR. The
TABLE 2. Coexistence of SPR With Calcium Binding Proteinsand Neuropeptides1
CCKn 5 164
PVn 5 268
SOMn 5 63
NPYn 5 134
CBn 5 138
CRn 5 102
VIPn 5 118
DG 4% 59% — 28% — — 0%Hilus 13% 0% 45% 60% — 25% 0%CA3 17% 9% 19% 12% 3% 23% 1%CA1 19% 0% 18% 60% 8% 30% 2%
A) Percentage of SPR-positive cells immunoreactive for different markers. N indicatesnumber of sampled SPR-immunoreactive cells.
CCKn 5 67
PVn 5 198
SOMn 5 103
NPYn 5 102
CBn 5 72
CRn 5 77
VIPn 5 60
DG 100% 90% — 80% — — 0%Hilus 100% 0% 73% 84% — 38% 0%CA3 81% 12% 70% 55% 14% 50% 7%CA1 90% 0% 8% 51% 10% 37% 20%
B) Percentage of calcium binding protein- and neuropeptide-containing cells immunore-active for SPR. N indicates number of sampled calcium binding protein- and neuropep-tide-containing cells.1Abbreviations: DG: gyrus dentatus; CCK: cholecystokinin; CB: calbindin; CR: calreti-nin; NPY: neuropeptide Y; PV: parvalbumin; SOM: somatostatin; VIP: vasoactiveintestinal polypeptide.
SPR-CONTAINING INTERNEURONS IN THE HIPPOCAMPUS 327

Fig. 4. A: Camera lucida drawings of multipolar or bituftedSPR-positive cells in the CA1 region also showing CCK immunoreactiv-ity. Note the large cell body and the thick primary dendrites. Nearly allCCK-positive cells also colocalized SPR.B,C:High magnification lightmicrographs of an SPR-positive cell that contains CCK as demon-strated by the mirror technique in the CA3 region. Arrow points to aSPR-positive CCK-negative cell body. Capillaries labeled by c1–c3
serve as landmarks. D,E: Large multipolar SPR-positive cell in str.pyramidale of the CA1 region cut in half on the surface of the section.The same cell shows CCK immunoreactivity in the adjacent section.The thick-cut primary dendrite (arrowhead) continues into the match-ing section. Arrow points to a SPR-positive CCK-negative cell body.Capillaries labeled by c1–c2 serve as landmarks. str. l.m., str. lacuno-sum-moleculare. Scale bars 5 20 µm.

Fig. 5. Light micrographs of sections with double-immunofluores-cent staining for SPR and the calcium-binding proteins parvalbuminand calbindin. A,B: SPR-positive pyramidallike basket cells containparvalbumin in str. granulosum of the dentate gyrus. Parvalbumin-positive interneurons were negative for SPR in all other subfields of
the hippocampus. C–D: Calbindin-immunoreactive cells showed SPRpositivity only in str. oriens of the CA3 region. The SPR-positive cell instr. radiatum of the CA1 region (E, arrow) is negative for calbindin(arrow in F). c, corresponding capillary. Scale bars 5 20 µm.
SPR-CONTAINING INTERNEURONS IN THE HIPPOCAMPUS 329

Figure 6

colocalization of these markers with each other havelargely been established. Parvalbumin does not colocalizewith any other markers, apart from a few of them thatcontain somatostatin or NPY (Gulyas et al., 1991; Miet-tinen et al., 1992; Gao and Fritschy 1994; Acsady et al.,1996a). Calbindin-containing cells in CA1 and CA3 str.oriens are mostly positive for somatostatin (Katona et al.,1996) or, particularly in the CA3 region, for CCK but notfor other markers (Gulyas et al., 1991; Miettinen et al.,1992). Calretinin colocalizes with somatostatin in spinycalretinin-positive cells (Katona et al., 1996), but 25% ofthe aspiny calretinin-immunoreactive cells contain VIP(Acsady et al., 1996a). VIP-immunoreactive interneuronscontaining calretinin innervate other GABAergic cells,whereas VIP and CCK coexist in parvalbumin-negativebasket cells (Acsady et al., 1996a,b). The majority ofCCK-positive cells are negative for all the other markersexcept VIP (Somogyi et al., 1984; Gulyas et al., 1991;Acsady et al., 1996a). The substantial overlap betweensomatostatin andNPY-immunoreactive cells was also dem-onstrated (Kohler et al., 1987). Based on these observa-tions and summing the proportion of SPR-immunoreactivecells colocalizing other markers (see Table 2), we canconclude that 80–90% of all the SPR-containing cells arelabeled by one of the markers or by a combination of them.Although simultaneous colocalization of SPR with twoother markers was not directly demonstrated, the coexist-ence of SPR with somatostatin and NPY in the hilar spinycells is highly probable. Similarly, VIP-immunoreactivebasket cells positive for colocalizing SPR are also likelycontain CCK. The large calbindin- and SPR-positive multi-polar cells in the CA3 str. oriens may also contain somato-statin or, occasionally, CCK.
SPR-positive perisomatic inhibitory cells
Morphologically distinct SPR-immunoreactive interneu-rons colocalized CCK and occasionally VIP but not theother markers. The majority of the cells in which SPR andCCK were colocalized had few thick primary dendrites.Previous studies have shown that CCK-positive interneu-rons innervate the perisomatic region of pyramidal cells;however, due to poor staining intensity, the extent of theirdendritic arbor could not be established. SPR immunostain-ing visualized the dendritic arbor of CCK-containing cellsin a Golgilikemanner. In the CA1 region, they were largelyconfined to strata oriens, radiatum, and pyramidale, withoccasional thin distal dendrites penetrating str. lacunosum-moleculare. The dendritic distribution of CCK-immunore-active cells suggests that the principal excitatory drive tothese cells is likely to originate from ipsi- and contralateral
Schaffer collaterals. They are unlikely to participate signifi-cantly in the perforant path-induced, feed-forward inhibi-tion, which is pronounced in the CA1 region (Buzsaki andEidelberg, 1982). Thus, this effect probably originates fromparvalbumin-containing basket and chandelier cells. Instr. granulosum of the dentate gyrus, all parvalbumin-positive (perisomatic inhibitory) cells colocalized SPR,whereas in the hilus and Ammon’s horn, parvalbumin-positive basket and chandelier cells did not contain SPR.Parvalbumin-positive cells show morphological diversityin the dentate gyrus (Baimbridge and Miller, 1982; Ribak,1992). In the present study, however, the location of theparvalbumin-containing cells rather than their morphol-ogy was directly related to SPR content, i.e., in str.granulosum all parvalbumin cells contained SPR regard-less of their pyramidal shape or fusiform morphology,whereas in the adjacent hilus none of the parvalbumin-positive cell were immunoreactive for SPR. This strikingdifference between parvalbumin-containing interneuronsin the hilus versus str. granulosum might be explained bythe pattern of substance P-containing innervation of thisregion. In contrast to parvalbumin-containing neurons,CCK-positive basket cells showed SPR immunoreactivityin all hippocampal subfields, further suggesting that func-tional differences exist between the two basket cell popula-tions (Acsady et al., 1996a).
SPR-positive dendritic inhibitory cells andSPR-positive interneurons innervating other
GABAergic cells
Large, fusiform, spiny cells in the hilus and str. lucidumof the CA3 region always contained somatostatin and NPYbut were negative for the other markers. Interneuronswith identical morphology have been described in previousGolgi studies and in vivo and in vitro intracellular labelingexperiments (Amaral, 1978; Han et al., 1993; Buckmasterand Schwartzkroin, 1995), but their NPY content has beendirectly demonstrated only for a singe cell (Sık et al.,1996). In immunocytochemical studies, somatostatin/NPY-containing cells projected to the outer two-thirds of themolecular layer and to innervate the contralateral dentategyrus (Kohler et al., 1986; Deller and Leranthh, 1990;Leranthh et al., 1990). In a recent study (Baude et al.,1993), the dendritic distribution of somatostatin-contain-ing cells were selectively visualized by immunostaining formetabotropic glutamate receptor 1a (mGluR1a). Baude etal. (1993) found only a few mGluR-positive dendritescrossing str. granulosum; the majority were restricted tothe hilus. In the present study, the dendritic distribution ofa large number of somatostatin/NPY-containing cells couldbe examined by SPR immunostaining. The dendrites ofspiny SPR-positive neurons were confined to the hilus andoccasionally penetrated str. radiatum of the CA3c regionbut never crossed str. granulosum. Based on this observa-tion, we can conclude that somatostatin/NPY-containing,spiny SPR-positive cells receive excitatory input almostexclusively from the mossy fiber collaterals and thus aredriven primarily in a feedback manner as their counter-part in the CA1 region (Blasco-Ibanez and Freund, 1995).In the hilus, spiny SPR-containing (i.e., somatostatin andNPY-positive) cells were many times faintly labeled ornegative for GABA, whereas in a recent study all somato-statin-containing cells contained the mRNA of GAD, the
Fig. 6. Somatostatin-containing neurons form a distinct group ofSPR-immunoreactive cells. A: Camera lucida drawing of a large spinyfusiform SPR-containing neuron in the hilus. The soma and theproximal and distal dendrites are densely covered with spines. All thecells with similar morphology examined were positive for somatosta-tin and for NPY. The arrow indicates the axon initial segment arisingfrom a proximal dendrite. B,C: High magnification light micrographsof two adjacent sections demonstrate that spiny SPR-positive cells inthe hilus also contain somatostatin. The dense meshwork of SPR-positive dendrites in the hilus mainly consists of spiny dendrites(arrowheads in B) of this cell type. c1 and c2 label capillaries used aslandmarks.D,E: Large aspiny or sparsely spiny SPR-immunoreactivecells also contained somatostatin (s1, s2) in str. oriens of the CA3region. c1–c3 labels corresponding capillaries. Scale bars 5 20 µm.
SPR-CONTAINING INTERNEURONS IN THE HIPPOCAMPUS 331
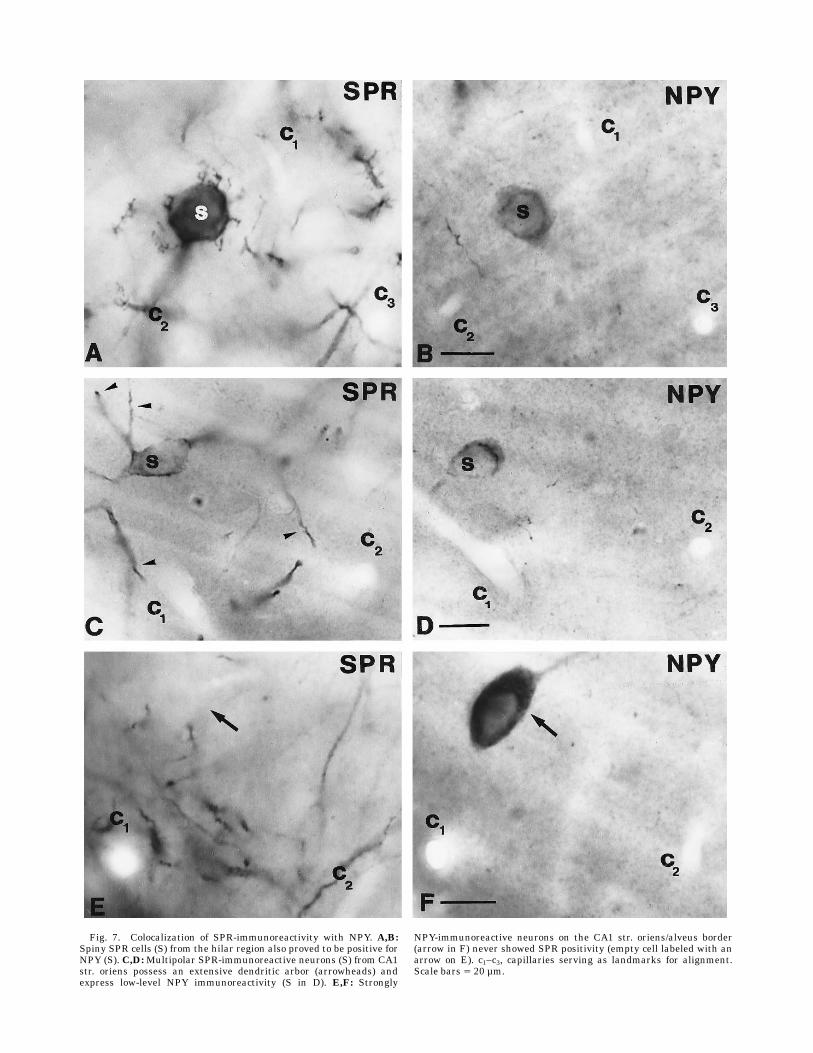
Fig. 7. Colocalization of SPR-immunoreactivity with NPY. A,B:Spiny SPR cells (S) from the hilar region also proved to be positive forNPY (S). C,D:Multipolar SPR-immunoreactive neurons (S) from CA1str. oriens possess an extensive dendritic arbor (arrowheads) andexpress low-level NPY immunoreactivity (S in D). E,F: Strongly
NPY-immunoreactive neurons on the CA1 str. oriens/alveus border(arrow in F) never showed SPR positivity (empty cell labeled with anarrow on E). c1–c3, capillaries serving as landmarks for alignment.Scale bars 5 20 µm.

synthesizing enzyme of GABA (Esclapez and Houser,1995). This apparent discrepancy between the in situhybridization technique and immunocytochemistry maybe explained by the lower sensitivity of immunohistochem-istry, which is unable to detect the low level of GABApresent in GABAergic cells with distant projection (Miet-tinen et al., 1992; Toth et al., 1993). In the hilus and theCA3 region, spiny SPR-positive cells contained somatosta-tin and NPY. In contrast, only a very small proportion ofthe somatostatin-containing cells colocalized SPR in theCA1 region. This neurochemical difference between somato-statin-positive neurons is again surprising, because theirrole (i.e., feedback inhibition in the distal dendritic regionof pricipal cells) is similar in all hippocampal subfields(Baude et al., 1993; Han et al., 1993; McBain et al., 1994;Blasco-Ibanez and Freund, 1995). In str. oriens of the CA3region, large SPR-immunoreactive cells with smooth thickprimary dendrites were positive for somatostatin; more-over, they contained calbindin, another marker for inter-neurons responsible for dendritic inhibition. In this sub-
field, calbindin is also present in some CCK-containing(SPR-positive) basket cells, whereas in the rest of thehippocampus calbindin-positive cells were negative forSPR. Multipolar SPR-immunoreactive cells with thin pri-mary dendrites branching close to the somata were hetero-geneous with regard to the colocalized marker. The major-ity of these cells contained either calretinin or NPY. Recentstudies on the postsynaptic target distribution of calretinin-positive interneurons showed that they selectively inner-vate other interneurons (Gulyas et al., 1996), whereasNPY-containing cells innervate the dendritic region ofpyramidal cells (Deller and Leranthh, 1990). In spite of theheterogeneity in termination pattern, the somadendriticmorphology of SPR-immunoreactive cells containing cal-retinin or NPY did not show consistent morphologicaldifferences. Comparison of the SPR-positive and negativecalretinin-immunoreactive interneurons showed that theSPR-positive cells rarely participate in dendrodendriticcontacts, which is a feature characteristic of numerouscalretinin-positive interneurons (Gulyas et al., 1992).
Fig. 8. A: Camera lucida drawing of SPR-immunoreactive interneurons also containing calretinin.Although the cells have prominent distal dendritic tufts, they have only few primary dendrites. B,C: AnSPR-positive interneuron in CA1 str. radiatum cut in half also shows calretinin immunostaining in theadjacent section. Scale bars 5 20 µm.
SPR-CONTAINING INTERNEURONS IN THE HIPPOCAMPUS 333

Correlation of the morphology andneurochemical marker content ofSPR-immunoreactive interneurons
For two subpopulations of SPR-immunoreactive neu-rons, morphological characteristics showed a direct corre-lation with marker content. The numerous, large, spinySPR-positive neurons in the hilus and str. lucidum of theCA3 region were always positive for somatostatin. Simi-larly large, multipolar, SPR-positve cells always containedCCK in the CA1 region. However, SPR-immunoreactiveinterneurons belonging to other morphologically estab-lished classes did not show characteristic neurochemicalmarker contents. Pyramidal-like and fusiform cells in thedentate gyrus contained parvalbumin, CCK, or NPY. Thesethree substances do not colocalize with each other and arelocalized in cell types with different termination patterns.Parvalbumin-positive cells innervate the perisomatic re-gion of granule cells (Kosaka et al., 1987; Soriano et al.,1990; Ribak et al., 1990), the majority of CCK-containingterminals form baskets in the hilar region (Leranth andFrotscher, 1986), and NPY axons mainly arborize in str.moleculare (Kohler et al., 1986; Deller and Leranth, 1990).This fact indicates that neurons with similar soma-dendritic morphology may have different axonal distribu-tion and target selectivity. Earlier Golgi studies haveshown that interneurons with pyramidal-like cell bodiesmay innervate the perisomatic and the dendritic regions ofthe granule cells (Amaral, 1978; Soriano and Frothcher,1993), which was confirmed by a recent intracellularlabeling study (Scharfman, 1995). Here, we have demon-strated that, in addition tomorphological and electrophysi-ological differences, pyramidal-shaped cells in the dentategyrus also show neurochemical heterogeneity. Similarly topyramidal-shaped cells, multipolar cells with thin primarydendrites in the CA1 and CA3 regions showed no consis-tent neurochemical characteristics because they containedeither NPY or calretinin.
Functional implications
The endogenous substrate of SPR is probably substanceP, although related tachykinins may also bind this recep-tor. Substance P-positive axons of extra- and intrahippo-campal origin have been described in a number of speciesincluding guinea pig, cat, monkey, and human (Gall andSelawski 1984; Yanagihara and Niimi, 1989; Seress andLeranth, 1996). However, the presence and number ofsubstance P-containing fibers and neurons in the hippocam-pus of the rat is still a controversial issue and seems todepend on the fixation protocol and the antibody used(Ljungdahl et al., 1978; Roberts et al., 1984; Davis andKohler, 1985; Iritani et al., 1989; Leranth and Borhegyi,personal communication). The extrahippocampal sub-stance P-containing fibers in the cat, guinea pig, andmonkey originate in the supramammillary nucleus (Galland Selawski, 1984; Ino et al., 1989; Yanagihara andNiimi., 1989; Leranth and Nitsch, 1994) and arborize inthe supragranular layer of the dentate gyrus and in theCA2 region. These subcortical fibers terminate almostexclusively on principal cells in the rat (Magloczky et al.,1994). Recent colocalization studies have shown that theintrahippocampal substance P-containing fibers derivefrom somatostatin-positive cells and arborize in the outertwo-thirds of the molecular layer in the monkey (Seressand Leranth, 1996), where they probably innervate gran-
ule cells. Thus, there seems to be a regional and cell-type-associated mismatch between the distribution of SPRimmunoreactivity and substance P-containing axons. SPR-positive dendrites are abundant in regions (i.e., hilus)where no intrahippocampal or extrahippocampal sub-stance P-positive fibers terminate. In addition, interneu-rons, which are not the principal targets of intrinsic orextrinsic substance P-positive fibers, express the receptor.The mismatch between substance P and SPR have beendirectly demonstrated (Liu et al., 1994), and a nonsynapticmechanism was suggested as the mode of action forsubstance P (Mantyh et al., 1995). Substance P diffused toconsiderable distance in the spinal cord (Duggan et al.,1990). The action of substance P is probably excitatory tononprincipal cells in the hippocampus (Dreifuss andRaggenbass, 1986). Our results suggest that diverse inhibi-tory mechanisms might be facilitated, by a diffuse releaseof substance P, through SPR, which is present on interneu-rons participating in perisomatic and dendritic inhibitionof principal cells and in the innervation of other GABAer-gic interneurons.
ACKNOWLEDGMENTS
This sudy was supported by grants from the HumanFrontier Science Program Organisation, the HowardHughes Medical Institute, and OTKA (T 16942) Hungary.We are grateful to Dr. K.G. Baimbridge and to Dr. M.R.Celio (calbindin and parvalbumin), Dr. T. Gorcs (CCK, VIP,NPY, and somatostatin), Dr. J.H. Rogers (calretinin), andDr. C.G. Beaulieau (GABA) for kind gifts of antisera. Theexcellent technical assistance of Mrs. E. Borok, Mrs. A.Z.Szabo, and Mr. G. Terstyanszky is also acknowledged.
LITERATURE CITED
Acsady, L., D. Arabadzisz, and T.F. Freund (1996a) Correlated morphologi-cal and neurochemical features identify different subsets of VIP-immunoreactive interneurons in rat hippocampus.Neuroscience 73:299–315.
Acsady, L., T.J. Gorcs, and T.F. Freund (1996b) Different populations ofVIP-immunoreactive interneurons are specialized to control pyramidalcells or interneurons in the hippocampus. Neuroscience 73:317–334.
Amaral, D.G., (1978) A Golgi study of cell types in the hilar region of thehippocampus in the rat. J. Comp. Neurol. 182:851–914.
Baimbridge, K.G., and J.J. Miller (1982) Immunohistochemical localizationof calcium-binding protein in the cerebellum, hippocampal formationand olfactory bulb of the rat. Brain Res. 245:223–229.
Baude, A., Z. Nusser, J.D. Roberts, E. Mulvihill, R.A. McIlhinney, and P.Somogyi (1993) The metabotropic glutamate receptor (mGluR1 alpha)is concentrated at perisynaptic membrane of neuronal subpopulationsas detected by immunogold reaction. Neuron 11:771–787.
Beaulieu, C., G. Campistron, and C. Crevier (1994) Quantitative aspects ofthe GABA circuitry in the visual cortex of adult rat. J. Comp. Neurol.339:559–572.
Blasco-Ibanez, J.M., and T.F. Freund (1995) Synaptic input of horizontalinterneurons in stratum oriens of the hippocampal CA1 subfield:Structural basis of feedback activation. Eur. J. Neurosci. 7:2170–2180.
Bragin, A., G. Jando, Z. Nadasdy, J. Hetke, K. Wise, and G. Buzsaki (1995)Gamma (40–100 Hz) oscillation in the hippocampus of the behaving rat.J. Neurosci. 15:47–60.
Buckmaster, P.S., and P.A. Schwartzkroin (1995) Interneurons and inhibi-tion in the dentate gyrus of the rat in vivo. J. Neurosci. 15:774–789.
Buckmaster, P.S., D.D. Kunkel, R.J. Robbins, and P.A. Schwartzkroin(1994) Somatostatin-immunoreactivity in the hippocampus of mouse,rat, guinea pig, and rabbit. Hippocampus 4:167–180.
Buhl, E.H., K. Halasy, and P. Somogyi (1994) Diverse sources of hippocam-pal unitary inhibitory postsynaptic potentials and the number ofsynaptic release sites. Nature 368:823–828.
334 L. ACSADY ET AL.

Buzsaki, G., and J.J. Chrobak (1995) Temporal structure in spatiallyorganized neuronal ensembles: A role for interneuronal networks. Curr.Opin. Neurobiol. 5:504–510.
Buzsaki, G., and E. Eidelberg (1982) Direct afferent excitation and long-termpotentiation of hippocampal interneurons. J. Neurophysiol. 48:597–607.
Buzsaki, G., Z. Horvath, R. Urioste, J. Hetke, and K. Wise (1992) High-frequency network oscillation in the hippocampus. Science 256:1025–1027.
Celio, M.R. (1990) Calbindin D-28k and parvalbumin in the rat nervoussystem. Neuroscience 35:375–475.
Cobb, S.R., E.H. Buhl, K. Halasy, O. Paulsen, and P. Somogyi (1995)Synchronization of neural activity in hippocampus by individual GABA-ergic interneurons. Nature 378:75–78.
Csiffary,A., T.J. Gorcs, andM. Palkovits (1990) NeuropeptideY innervationof ACTH immunoreactive neurons in the arcuate nucleus of rats: Acorrelated light and electronmicroscopic double immunolabelling study.Brain Res. 506:215–222.
Davis, S., and C. Kohler (1985) The substance P innervation of the rathippocampal formation. Anat. Embryol. 173:45–52.
Deller, T., and C. Leranth (1990) Synaptic connections of neuropeptide Y(NPY) immunoreactive neurons in the hilar area of the rat hippocam-pus. J. Comp. Neurol. 300:433–447.
Dreifuss, J.J., and M. Raggenbass (1986) Tachykinins and bombesin excitenon-pyramidal neurones in rat hippocampus. J. Physiol. 379:417–428.
Duggan, A.W., P.J. Hope, B. Jarrott, H.G. Schaible, and S.M. Fleetwood-Walker (1990) Relese spread and persistance of immunoreactive neuro-kinin A in the dorsal horn of the cat following noxious cutaneousstimulation. Studies with antibody microprobes. Neuroscience 35:195–202.
Esclapez, M., and C.R. Houser (1995) Somatostatin neurons are a subpopu-lation of GABA neurons in the rat dentate gyrus: Evidence fromcolocalization of pre-prosomatostatin and glutamate decarboxylasemessenger RNAs. Neuroscience 64:339–355.
Gall, C., and L. Selawski (1984) Supramammillary afferents to guinea pighippocampus contain substance P-like immunoreactivity. Neurosci.Lett. 51:171–176.
Gao, B., and J.M. Fritschy (1994) Selective allocation of GABAA receptorscontaining the alfa1 subunit to neurochemically distinct subpopula-tions of rat hippocampal interneurons. Eur. J. Neurosci. 6:837–853.
Gulyas, A.I., and T.F. Freund (1996) Calbindin-containing interneuronsinnervate pyramidal cell dendrites in the hippocampus. Hippocampus,(in press).
Gulyas, A.I., T.J. Gorcs, and T.F. Freund (1990) Innervation of differentpeptide-containing neurons in the hippocampus by GABAergic septalafferents. Neuroscience 37:31–44.
Gulyas, A.I., K. Toth, P. Danos, and T.F. Freund (1991) Subpopulations ofGABAergic neurons containing parvalbumin, calbindin D28k, andcholecystokinin in the rat hippocampus. J. Comp. Neurol. 312:371–378.
Gulyas, A.I., R. Miettinen, D.M. Jacobowitz, and T.F. Freund (1992)Calretinin is present in non-pyramidal cells of the rat hippocampus—I.A new type of neuron specifically associated with the mossy fibresystem. Neuroscience 48:1–27.
Gulyas, A.I., R. Miles, N. Hajos, and T.F. Freund (1993a) Precision andvariability in postsynaptic target selection of inhibitory cells in thehippocampal CA3 region. Eur. J. Neurosci. 5:1729–1751.
Gulyas, A.I., R. Miles, A. Sık, K. Toth, N. Tamamaki, and T.F. Freund(1993b) Hippocampal pyramidal cells excite inhibitory neurons througha single release site. Nature 366:683–687.
Gulyas, A.I., N. Hajos, and T.F. Freund (1996) Interneurons containingcalretinin are specialised to control other interneurons in the rathippocampus. J.Neurosci 16:3397–3411.
Hajos, N., L. Acsady, and T.F. Freund (1996) Target selectivity andneurochemical characteristics of VIP-immonopositive interneurons inthe rat dentate gyrus. Eur. J. Neurosci. 8:1415–1431.
Han, Z.S., E.H. Buhl, Z. Lorinczi, and P. Somogyi (1993) A high degree ofspatial selectivity in the axonal and dendritic domains of physiologi-cally identified local-circuit neurons in the dentate gyrus of the rathippocampus. Eur. J. Neurosci. 5:395–410.
Ino, T., K. Itoh, T. Sugimoto, T. Kaneko, H. Kamiya, and N. Mizuno (1989)The supramammillary region of the cat sends substance P-like immuno-reactive axons to the hippocampal formation and entorhinal cortex.Neurosci. Lett. 90:259–264.
Iritani, S., M. Fujii, and K. Satoh (1989) The distribution of substance P inthe cerebral cortex and hippocampal formation: An immunohistochemi-cal study in the monkey and rat. Brain. Res. Bull. 22:295–303.
Kaneko, T., R. Shigemoto, S. Nakanishi, and N. Mizuno (1994) Morphologi-cal and chemical characteristics of substance P receptor-immunoreac-tive neurons in the rat neocortex. Neuroscience 60:199–211.
Katona, I., L. Acsady, A. Gulacsi, and T.F. Freund (1996) Somatostatin-containing cells in the rat hippocampus: Connectivity and neurochemi-cal heterogeneity. Eur. J. Neurosci. 9(Suppl.): 176.
Kawaguchi, Y., H. Katsumaru, T. Kosaka, C.W. Heizmann, and K. Hama(1987) Fast spiking cells in rat hippocampus (CA1 region) contain thecalcium-binding protein parvalbumin. Brain Res. 416:369–374.
Kohler, C., and V. Chan-Palay (1983) Somatostatin-like immunoreactiveneurons in the hippocampus:An immunohistochemical study in the rat.Neurosci. Lett. 34:259–265.
Kohler, C., L. Eriksson, S. Davies, and V. Chan-Palay (1986) NeuropeptideY innervation of the hippocampal region in the rat andmonkey brain. J.Comp. Neurol. 244:384–400.
Kohler, C., L. Eriksson, S. Davies, and V. Chan-Palay (1987) Co-localizationof neuropeptide tyrosine and somatostatin immunoreactivity in neu-rons of individual subfields of the rat hippocampal region. Neurosci.Lett. 78:1–6.
Kosaka, T., K. Kosaka, K. Tateishi, Y. Hamaoka, N. Yanaihara, J.Y. Wu, andK. Hama (1985) GABAergic neurons containing CCK-8-like and/orVIP-like immunoreactivities in the rat hippocampus and dentate gyrus.J. Comp. Neurol. 239:420–430.
Kosaka, T., H. Katsumaru, K. Hama, J.Y. Wu, and C.W. Heizmann (1987)GABAergic neurons containing the Ca21-binding protein parvalbuminin the rat hippocampus and dentate gyrus. Brain Res. 419:119–130.
Lacaille, J.C., and P.A. Schwartzkroin (1988a) Stratum lacunosum-moleculare interneurons of hippocampal CA1 region. I. Intracellularresponse characteristics, synaptic responses, andmorphology. J. Neuro-sci. 8:1400–1410.
Lacaille, J.C., and P.A. Schwartzkroin (1988b) Stratum lacunosum-moleculare interneurons of hippocampal CA1 region. II. Intrasomaticand intradendritic recordings of local circuit synaptic interactions. J.Neurosci. 8:1411–1424.
Lacaille, J.C., A.L. Mueller, D.D. Kunkel, and P.A. Schwartzkroin (1987)Local circuit interactions between oriens/alveus interneurons and CA1pyramidal cells in hippocampal slices: Electrophysiology and morphol-ogy. J. Neurosci. 7:1979–1993.
Lantos, T.A., T.J. Gorcs, and M. Palkovits (1995) Immunohistochemicalmappings of neuropeptides in the premamillary region of the hypothala-mus in rats. Brain Res. Rev. 20:209–249.
Leranth, C., and M. Frotscher (1986) Synaptic connections of cholecystoki-nin-immunoreactive neurons and terminals in the rat fascia dentata: acombined light and electron microscopic study. J. Comp. Neurol.254:51–64.
Leranth, C., and R. Nitsch (1994) Morphological evidence that hypotha-lamic substance P-containing afferents are capable of filtering thesignal flow in themonkey hippocampal formation. J. Neurosci. 14:4079–4094
Leranth, C., A.J. Malcolm, and M. Frotscher (1990) Afferent and efferentsynaptic connections of somatostatin-immunoreactive neurons in therat fascia dentata. J. Comp. Neurol. 295:111–122.
Liu, H., J.L. Brown, L. Jasmin, J.E. Maggio, S.R. Vigna, P.W. Mantyh, andA.I. Basbaum (1994) Synaptic relationship between substance P andthe substance P receptor: Light and electron microscopic characteriza-tion of the mismatch between neuropeptides and their receptors. Proc.Natl. Acad. Sci. USA 91:1009–1013.
Ljungdahl, A., T. Hokfelt, and G. Nilsson (1978) Distribution of substanceP-like immunoreactivity in the central nervous system of the rat-I. Cellbodies and nerve terminals. Neuroscience 3:861–943.
Magloczky, Z., L. Acsady, and T.F. Freund (1994) Principal cells are thepostsynaptic targets of supramammillary afferents in the hippocampusof the rat. Hippocampus 4:322–334.
Mantyh, P.W., T. Gates C.R. Mantyh and J.E. Maggio (1989) Autoradio-graphic localization and characterization of tachykinin receptor bind-ing sites in the rat brain and peripheral tissues. J Neurosci. 9:258–279.
Mantyh, P.W., C.J. Allen, J.R. Ghilardi, S.D. Rogers, C.R. Mantyh, H. Liu,A.I. Basbaum, S.R. Vigna, and J.E. Maggio (1995) Rapid endocytosis ofa G protein coupled receptor: Substance P-evoked internalization of itsreceptor in the rat striatum in vivo. Proc. Natl. Acad. Sci. USA92:2622–2626.
McBain, C.J., T.J. DiChiara, and J.A. Kauer (1994) Activation of metabo-tropic glutamate receptors differentially affects two classes of hippocam-pal interneurons and potentiates excitatory synaptic transmission. J.Neurosci. 14:4433–4445.
SPR-CONTAINING INTERNEURONS IN THE HIPPOCAMPUS 335

Michelson, H.B., and R.K.S. Wong (1991) Excitatory synaptic responsesmediated by GABAA receptors in the hippocampus. Science 253:1420–1423.
Miettinen, R., A.I. Gulyas, K.G. Baimbridge, D.M. Jacobowitz, and T.F.Freund (1992) Calretinin is present in non-pyramidal cells of the rathippocampus—II. Co-existence with other calcium binding proteinsand GABA. Neuroscience 48:29–43.
Miles, R., K. Toth, A.I. Gulyas, N. Hajos, and T.F. Freund (1996) Distinctfunctional roles for somatic and dendritic inhibition in hippocampus.Neuron 16:815–823.
Morrison, J.H., R. Benoit, P.J. Magistretti, N. Ling, and F.E. Bloom (1982)Immunohistochemical distribution of pro-somatostatin-related pep-tides in hippocampus. Neurosci. Lett. 34:137–142.
Muller, W., and U. Misgeld (1990) Inhibitory role of dentate hilus neuronsin guinea pig hippocampal slice. J. Neurophysiol. 64:46–56.
Nakaya, Y., T. Kaneko, R. Shigemoto, S. Nakanishi, and N. Mizuno (1994)Immunohistochemical localization of substance P receptor in the cen-tral nervous system of the adult rat. J. Comp. Neurol. 347:249–274.
Nunzi, M.G., A. Gorio, F. Milan, T.F. Freund, P. Somogyi, and A.D. Smith(1985) Cholecystokinin-immunoreactive cells form symmetrical synap-tic contacts with pyramidal and nonpyramidal neurons in the hippocam-pus. J. Comp. Neurol. 237:485–505.
Ribak, C.E., (1992) Local circuitry of GABAergic basket cells in the dentategyrus. In Ribak, C.E., Gall, C.M., and Mody I. (eds): The Dentate Gyrusand Its Role in Seizures. Elsevier Science, pp. 29–49.
Ribak, C.E., R. Nitsch, and L. Seress (1990) Proportion of parvalbumin-positive basket cells in the GABAergic innervation of pyramidal andgranule cells of the rat hippocampal formation. J. Comp. Neurol.300:449–61.
Roberts, G.W., P.L. Woodhams, J.M. Polak, and T.J. Crow (1984) Distribu-tion of neuropeptides in the limbic system of the rat: The hippocampus.Neuroscience 11:35–77.
Rogers, J.H. (1989) Two calcium-binding proteins markmany chick sensoryneurons. Neuroscience 31:697–709.
Scharfman, H.E. (1995) Electrophysiological diversity of pyramidal-shapedneurons at the granule cell layer/hilus border of the rat dentate gyrusrecorded in vitro. Hippocampus 5:287–305.
Seress, L., and C. Leranth (1996) Distribution of substance P-immunoreac-tive neurons and fibers in themonkey hippocampal formation. Neurosci-ence 71:633–650.
Shigemoto, R., Y. Nakaya, S. Nomura, R. Ogawa-Meguro, H. Ohishi, T.Kaneko, S. Nakanishi, and N. Mizuno (1993) Immunocytochemicallocalization of rat substance P receptor in striatum. Neurosci. Lett.153:157–160.
Sık, A., M. Penttonen, A. Ylinen, and G. Buzsaki (1995) Hippocampal CA1
interneurons: An in vivo intracellular labeling study. J. Neurosci.15:6651–6665.
Sık, A., M. Penttonen, and G. Buzsaki (1996) Interneurons in the dentategyrus: An in vivo intracellular study. Eur. J. Neurosci., (in press).
Sloviter, R.S., and G. Nilaver (1987) Immunocytochemical localization ofGABA-, cholecystokinin-, vasoactive intestinal polypeptide-, and somato-statin-like immunoreactivity in the area dentata and hippocampus ofthe rat. J. Comp. Neurol. 256:42–60.
Soltesz, I., and M. Deschenes (1993) Low- and high-frequency membranepotential oscillations during theta activity in CA1 and CA3 pyramidalneurons of the rat hippocampus under ketamine-xylazine anesthesia. J.Neurophysiol. 70:97–116.
Somogyi, P., M.G. Nunzi, A. Gorio, and A.D. Smith (1983) A new type ofspecific interneuron in the monkey hippocampus forming synapsesexclusively with the axon initial segments of pyramidal cells. Brain Res.259:137–142.
Somogyi, P., A.J. Hodgson, A.D. Smith, M.G. Nunzi, A. Gorio, and J.Y. Wu(1984) Different populations of GABAergic neurons in the visual cortexand hippocampus of cat contain somatostatin- or cholecystokinin-immunoreactive material. J. Neurosci. 4:2590–2603.
Somogyi, P., T.F. Freund, A.J. Hodgson, J. Somogyi, D. Beroukas, and I.W.Chubb (1985) Identified axo-axonic cells are immunoreactive for GABAin the hippocampus and visual cortex of the cat. Brain Res. 332:143–149.
Soriano, E., and M. Frotscher (1993) GABAergic innervation of the ratfascia dentata:Anovel type of interneuron in the granule cell layer withextensive axonal arborization in the molecular layer. J. Comp. Neurol.334:385–396.
Soriano, E., R. Nitsch, and M. Frotscher (1990) Axo-axonic chandelier cellsin the rat fascia dentata: Golgi–electron microscopy and immunocyto-chemical studies. J. Comp. Neurol. 293:1–25.
Toth, K., and T.F. Freund (1992) Calbindin D28k-containing nonpyramidalcells in the rat hippocampus: Their immunoreactivity for GABA andprojection to the medial septum. Neuroscience 49:793–805.
Toth, K., Z. Borhegyi, and T.F Freund (1993) Postsynaptic targets ofGABAergic hippocampal neurons in the medial septum-diagonal bandof Broca complex. J. Neurosci. 13:3712–3724.
Whittington, M.A., R.D. Traub, and J.G.R. Jeffrey (1995) Synchronizedoscillations in interneuron networks driven by metabotropic glutamatereceptor activation. Nature 373:612–615.
Yanagihara, M., and K. Niimi (1989) Substance P-like immunoreactiveprojection to the hippocampal formation from the posterior hypothala-mus in the cat. Brain Res. Bull. 22:682–694.
Ylinen, A., A. Bragin, Z. Nadasdy, G. Jando, I. Szabo, A. Sık, and G.Buzsaki(1995) Sharp wave-associated high-frequency oscillation (200 Hz) in theintact hippocampus: Network and intracellular mechanisms. J. Neuro-sci. 15:30–46.
336 L. ACSADY ET AL.