impact of land use on arbuscular mycorrhizal fungal communities in...
TRANSCRIPT

Impact of Land Use on Arbuscular Mycorrhizal Fungal Communitiesin Rural Canada
Mulan Dai,a Luke D. Bainard,a Chantal Hamel,a Yantai Gan,a Derek Lynchb
Semiarid Prairie Agricultural Research Centre, Agriculture and Agri-Food Canada, Swift Current, Saskatchewan, Canadaa; Department of Plant and Animal Sciences, Facultyof Agriculture, Dalhousie University, Truro, Nova Scotia, Canadab
The influence of land use on soil bio-resources is largely unknown. We examined the communities of arbuscular mycorrhizal(AM) fungi in wheat-growing cropland, natural areas, and seminatural areas along roads. We sampled the Canadian prairie ex-tensively (317 sites) and sampled 20 sites in the Atlantic maritime ecozone for comparison. The proportions of the different AMfungal taxa in the communities found at these sites varied with land use type and ecozones, based on pyrosequencing of 18SrRNA gene (rDNA) amplicons, but the lists of AM fungal taxa obtained from the different land use types and ecozones were verysimilar. In the prairie, the Glomeraceae family was the most abundant and diverse family of Glomeromycota, followed by theClaroideoglomeraceae, but in the Atlantic maritime ecozone, the Claroideoglomeraceae family was most abundant. In the prai-rie, species richness and Shannon’s diversity index were highest in roadsides, whereas cropland had a higher degree of speciesrichness than roadsides in the Atlantic maritime ecozone. The frequencies of occurrence of the different AM fungal taxa in crop-lands in the prairie and Atlantic maritime ecozones were highly correlated, but the AM fungal communities in these ecozoneshad different structures. We conclude that the AM fungal resources of soils are resilient to disturbance and that the richness ofAM fungi under cropland management has been maintained, despite evidence of a structural shift imposed by this type of landuse. Roadsides in the Canadian prairie are a good repository for the conservation of AM fungal diversity.
Arbuscular mycorrhizal (AM) fungi are a ubiquitous group ofobligate biotrophic fungi that play a key role in the function-
ing and sustainability of agroecosystems (1). These mutualisticfungi associate with the roots of the majority of agricultural plantsand have shown the potential to increase crop productivity. Theprimary function of the symbiosis involves the transfer of photo-synthetic carbon from the host plant to the fungal symbiont inexchange for increased uptake of phosphate and ammonium aswell as other essential mineral nutrients (2, 3). AM fungi alsoprovide other functional benefits to the host plant, including pro-tection from abiotic and biotic stresses (1). For example, there isevidence that these mutualistic fungi can increase the fitness oftheir host plant in harsh environments (4), including under lowsoil fertility (5–7), drought (8, 9), and salinity (10). AM fungi arealso involved in other ecological processes that are critical in agro-ecosystems, such as maintaining soil structure and stability (11)and the cycling of major elements, such as carbon, phosphorus,and nitrogen (2). The beneficial effects and ecological servicesprovided by AM fungi reveal their importance in the efficientfunctioning and sustainability of agroecosystems.
AM fungi share a long history of coevolution with plants invarious ecosystems, resulting in their adaptation to specific natu-ral areas (12, 13). In these areas, highly mutualistic plant-AM fun-gal pairs are stabilized by a positive-feedback loop through whichmutual rewards in the form of soil nutrients and carbon are pref-erentially given by AM fungi and host plants to their symbioticpartners (14). Highly mutualistic plant-AM fungal pairs improvethe performance of an ecosystem, in particular, the efficiency ofnutrient cycling, plant productivity, and the survival of AM fungi.Unfortunately, land management practices often impact the sta-bility and performance of the AM symbiosis, resulting in potentialconsequences to the overall productivity and sustainability ofagroecosystems.
Annual cropping practices deeply transform the plant cover
and soil conditions from their natural state through the use ofheavy machinery and the application of fertilizers and pesticides.As a result, conventional agricultural practices have an impact onAM fungal communities. Monoculture cropping deprives AMfungal taxa that have low compatibility with the crop plant fromhost support and subsequently reduces AM fungal diversity (15,16), while nonhost crops (e.g., canola, rape seed) and fallow treat-ments deprive all AM fungi of an appropriate host plant (17). Soiltillage and the termination of annual crops can cause intense dis-turbance to AM fungal networks and have a negative impact onextraradical hyphal density and the AM root colonization of sub-sequent crops (12, 18). Fertilization is known to strongly impactthe composition, growth, and function of AM fungi (18–21).Overall, agricultural practices have been reported to reduce thediversity and abundance of AM fungi to various degrees, depend-ing on the intensity of crop management (22–24).
The objective of this study was to evaluate the impact of annualcrop production on AM fungal communities in rural Canada bycomparing the relative abundances and compositions of AM fungiin annually cropped land, seminatural areas along roads, and nat-ural areas. Our survey most intensely examined cropland acrossthe edaphoclimatic zones of the rural prairie provinces, where81.5% of all Canadian croplands are located (25), but we alsoincluded some cropland on podzolic soils of the Atlantic maritime
Received 25 April 2013 Accepted 20 August 2013
Published ahead of print 30 August 2013
Address correspondence to Chantal Hamel, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AEM.01333-13.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.01333-13
November 2013 Volume 79 Number 21 Applied and Environmental Microbiology p. 6719–6729 aem.asm.org 6719
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

ecozone. This study provides detailed information on the compo-sition and diversity of indigenous AM fungi in these prime agri-cultural regions and allowed us to test the hypothesis that cropproduction influences the relative abundances and compositionsof AM fungal communities in the landscape of rural Canada. Wealso predicted that roadsides are a repository for the conservationof AM fungal diversity in areas of intensive crop production.
MATERIALS AND METHODSDescription of the surveyed areas. Soil and root samples were taken fromcropland (176 fields of spring wheat or durum wheat [Triticum aestivumL. or Triticum durum L.]), their adjacent roadsides (117 sites), and naturalareas (24 sites). In order to provide good coverage of the AM fungalcommunity of the vast area that is the Canadian prairie at a key stage ofwheat development, i.e., at heading, sampling was performed over 2 years,in 2009 and 2010. Pairing of close-by sampling areas of cropland, road-side, or natural area was performed where possible to ensure that com-parisons made between land use types were not biased by climate or soilconditions. Seventy-two percent of the croplands were paired; 11.2% werepaired with natural areas and 60.8% with roadsides. The distance betweenpaired cropland and roadside areas was less than 100 m, and the distancebetween paired cropland and natural areas was usually less than 100 m andnever exceeded 1 km. Cropland soil at several locations was covered bywheat plants up to the dirt roads and in the absence of roadside or naturalareas. Natural areas are scarce, and to provide power for multivariateanalyses of community structure, four unpaired samples were also takenfrom pristine natural areas, i.e., two sites on municipal land and two sitesin protected parks.
Sampling efforts were focused mainly on covering all the edaphocli-matic zones of the Canadian prairie ecozone (26), known as the brown,dark brown, black, and gray soil zones (27). Samples were also taken frompaired cropland and roadsides at 10 locations in the Atlantic maritimeecozone (26) in Nova Scotia. In the Canadian prairie, samples came fromjust above the U.S. border to the boreal forest over an area spanningapproximately1,450 km between Beaverlodge, Alberta, to the west, and
Brandon, Manitoba, to the east, but the majority of samples were collectedin the province of Saskatchewan.
In the Atlantic maritime ecozone, all samples were taken from soilsclassified as podzol (27). The croplands under organic production in thisecozone were sometimes weedy. Frequent use of perennial hay crops withcomplex plant compositions and the application of animal manure arecharacteristics of the cropping systems sampled in the Atlantic maritimeecozone.
Croplands located on brown, dark brown, and black chernozems ofthe prairie ecozone may have over 100 years of agricultural history, whilecroplands on gray luvisol and dark gray chernozems may have been bro-ken for crop production from natural forest in more recent times (28). Allcropping systems in the prairie province are based on the production ofwheat, which is often grown in crop rotation systems. The most commonrotation crops are canola, pea, lentil, barley, and flax. Organic systems arefertilized with plow-down green-manure crops in occasional semifallowyears. Ninety-nine of the sampling sites in cropland were conventionallymanaged, thus, normally untilled, and 77 were under organic manage-ment. Roadsides were a buffering area between the production field andthe road. They were typically gravelly, shaped as a ditch, and colonized bythe invasive forage grass species crested wheatgrass (Agropyron cristatum[L.] Gaertn) and bromegrass (Bromus inermis Leyss.), which were seededon the edges of newly constructed roads, and by several competitive spe-cies. Some of these species have wide adaptation, but many are found onlyin certain edaphoclimatic zones (see Table S1 in the supplemental mate-rial). The vegetation of most roadsides is cut and harvested as forage inJuly. Natural areas were areas spared from agriculture by their incon-venient location in the landscape, which is most often related to to-pography. Natural areas were usually small virgin patches, but somewere also very large (Fig. 1). The natural areas of different edaphocli-matic zones had very distinct plant communities, in contrast to road-sides, which were dominated by the same invasive forage grassesthroughout the prairie (Table S1).
Soil sampling and processing of samples. At each cropland site, com-posite samples were randomly taken to a depth of 7.5 cm over a samplingarea about 25 m2 in size that was deemed representative of the field, based
FIG 1 (A) Typical field and roadside areas at the fringe of the boreal forest; (B) typical natural vegetation in the semiarid southwest of the Canadian prairie; (C)satellite view showing the organization of the landscape in the Canadian prairie and the importance of roadsides as a repository of biodiversity in the zone ofintense utilization of land for the production of crops. Legal land units, called quarter sections, are distinguishable when they bear different vegetation. The gridroad system is emphasized in an overlay. The arrow indicates a deep coulee on the edge of which native prairie remnants could be found. Satellite photo providedby Kim Hodge (SGIC Ortho-Photography Project 2008 –2011, GC/AAFC) with the permission of Her Majesty the Queen in right of Canada 2009, as representedby the Minister of Agriculture and Agri-Food Canada.
Dai et al.
6720 aem.asm.org Applied and Environmental Microbiology
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

on topography. Thirty cores were taken with a soil probe directly on therow and pooled in a bag. Field edges and accesses were avoided. Stonesand stiff roots prevented the use of the soil probe in most areas along roadsand in natural areas, and composite samples from these areas consisted ofsix soil columns cut out from the top 7.5-cm soil layer with a shovel. Carewas taken to collect all roadside samples of the prairie provinces fromstands or patches of bromegrass. Soil samples were kept on ice in a coolerduring transportation. Sampling sites located outside a 320-km radiusfrom the laboratory in Swift Current were sampled by collaborators andsent by rapid courier. Soil samples were homogenized and freed fromstones by sieving them through a 2-mm mesh sieve in the laboratory.Samples were stored at �23°C prior to DNA extraction.
Molecular analysis. Metagenomic DNA was extracted from 0.5-g soilsamples drawn from each site using an UltraClean soil DNA isolation kit(catalogue no. 12800-100; Mo Bio Laboratories, Inc.) according to themaximum-yield protocol of the manufacturer, and samples were stored at�20°C. The fusion primers constructed with primers AMV4.5NF/AMDGR, adaptors, and the multiplex identifier (MID) (see Table S2 inthe supplemental material) based on those in a 454 sequencing technicalbulletin of Roche (29) were used to amplify the AM fungal 18S rRNA gene(rDNA). Primers AMV4.5NF/AMDGR were previously used successfullyin several studies (30, 31) to amplify sequences from environmental sam-ples of all four AM fungal orders, namely, the Diversisporales, Glomerales,Archaeosporales, and Paraglomerales, as well as other fungi of the Asco-mycota, Basidiomycota, and Chytridiomycota.
Each soil DNA sample was diluted (1:20) and amplified separatelywith the AM fungal primer set AMV4.5NF/AMDGR. In order to decreasevariations in the PCR process, samples were amplified in triplicate (32)using the fusion primer set in a PCR with a 10-�l volume per subsample.Platinum PCR SuperMix (catalogue no. 11306-016; Invitrogen) was usedin the PCRs. The final concentration of the reagent mix per 10-�l volumewas 0.0165 U �l�1Taq DNA polymerase, 1.24 mM MgCl2, 16.5 mM Tris-HCl (pH 8.4), 41.25 mM KCl, 165 �M (each) deoxynucleoside triphos-phate (dNTP), and 0.2 �M (each) primer. Thermal cycling was conductedin a Veriti 96-well fast thermal cycler (Applied Biosystems) with the fol-lowing conditions: 10 min of denaturation at 95°C for the first step; 35cycles of 30 s of denaturation at 94°C, 30 s of annealing at 55°C, and 1 minof elongation at 72°C; and 9 min of final elongation at 74°C. The threePCR products were pooled and purified with a ChargeSwitch PCRclean-up kit (catalogue no. CS12000; Invitrogen). Purified PCR ampli-cons were normalized at 25 ng �l�1 with a Savant DNA 120 SpeedVacconcentrator (Thermo Scientific). The concentration of purified ampli-cons was measured using a NanoDrop 1000 spectrophotometer (ThermoScientific). All amplicons from each sample were barcoded with one ofRoche’s 16 multiplex identifiers (MIDs). The tagged samples were pooledand sent for pyrosequencing, which was performed under contract at theNRC Plant Biotechnology Institute (NRC-PBI, Saskatoon, Saskatchewan,Canada).
Bioinformatic and phylogenetic analysis. Pyrosequencing reads con-taining ambiguous nucleotides (average quality score of �30) (33) or asingle nucleotide mismatch with the PCR primer or reads that were ofatypical length (�230 bp or �250 bp) were removed from the data setusing Mothur version 1.15.0 (34). Sequences belonging to groups otherthan the Glomeromycota were identified by comparison with the Silvaeukaryotic reference for 18S rDNA sequences (http://www.arb-silva.de/)and AM fungal reference sequences obtained from GenBank. All uniquesequences were filtered for reduced computational complexity usingMothur. The average length of the cleaned sequences was 241 bp. Theclean AM fungal sequences were aligned with each other using MUSCLE(35), and alignments were clustered based on 97% similarity into opera-tional taxonomic units (OTUs) using the furthest-neighbor algorithmwith Mothur. In addition, the Shannon diversity index (H=), abundancecoverage estimator (ACE) index, and OTU richness were calculated withMothur. Taxonomic assignment was performed by comparing a repre-
sentative sequence of each OTU to sequences in the GenBank nonredun-dant nucleotide database (36).
Representative OTU sequences and AM fungal reference sequencesfrom GenBank were aligned using MUSCLE (35), and neighbor-joiningphylogenetic reconstruction (37) was used to build the phylogenetic treein MEGA 5 (38). Default parameters were used except that bootstrapreplication was set at 1,000 with the Kimura 2-parameter model (39). Thenomenclature used here was proposed by Redecker et al. (40). The abun-dance of each OTU in a sample is expressed as the number of reads of thatOTU relative to all fungal reads in that sample.
Statistical analysis. Comparison of AM fungal communities betweenpaired samples of cropland and adjacent roadsides or natural areas wasmade by subjecting the paired-site data to Student’s t test using R. Theeffect of land use type on the ACE index and Shannon’s H= was tested byanalysis of variance (ANOVA) (41) for all three biomes, and the signifi-cance of the differences between land use type means was assessed post hocusing Duncan’s test with the Agricolae package in R (42).
The hypothesis that soils in different land use types and ecozones havedistinct AM fungal community structures was tested by subjecting therelative AM fungal OTU abundance data from both paired and unpairedsites to multiresponse permutation procedures (MRPP) in PC-ORD v. 6(43). Nonmetric multidimensional scaling (NMS) analysis (44) was usedin PC-ORD to visualize the whole data. The two most informative dimen-sions of a 3-dimensional solution explaining 59.9% of the variance wereused to construct the NMS ordination graph.
The gplots (45) and RColorBrewer packages (46) were used in R toplot a heatmap showing the proportion of each OTU making up the AMfungal communities found in cropland, along roadsides, and in naturalareas. Spearman correlation analysis was used in JMP v. 3.2.6 to evaluatethe relationship between the relative abundances of AM fungal OTUs andtheir frequencies of occurrence.
Nucleotide sequence accession number. A representative sequence ofeach AM fungal OTU analyzed in this study was deposited in GenBankunder accession numbers KF620139 to KF620260 (OTUs 1 to 122), andtheir sequences are listed in Table S3 in the supplemental material.
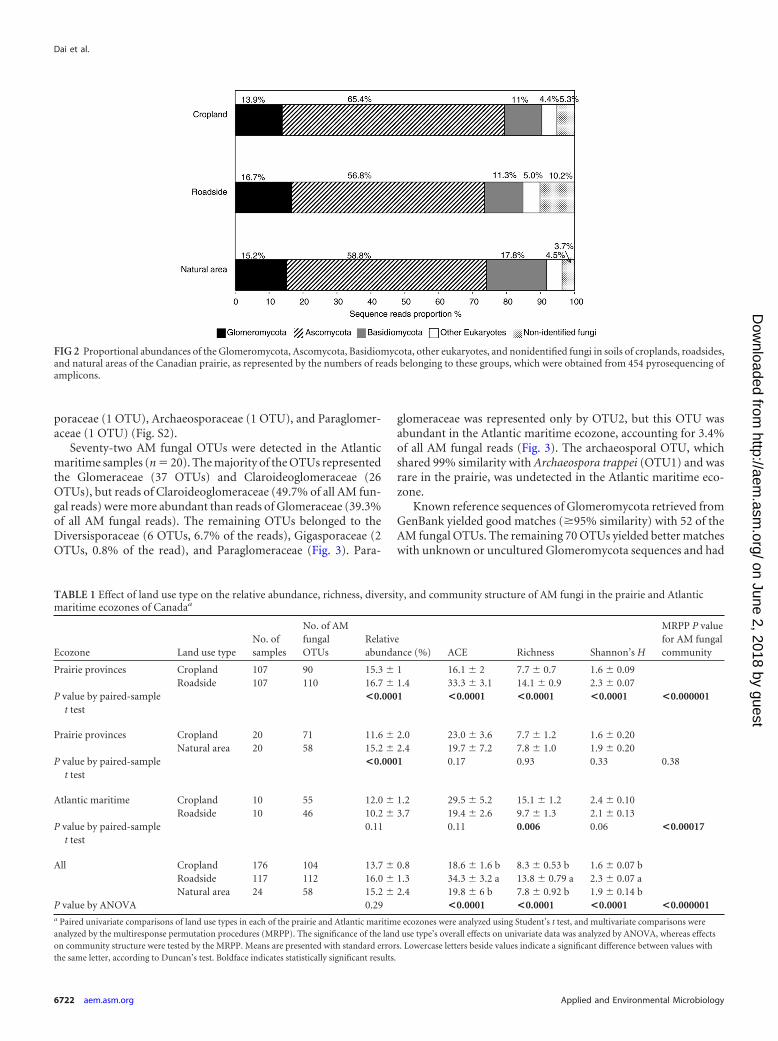
RESULTSMolecular analysis of AM fungi. The pyrosequencing platformproduced an average of 4,213 reads per sample after cleaning.Among land use types, the average numbers of reads per samplewere 5,411 for roadsides, 3,641 for cropland, and 2,318 for naturalareas. The primers used in this study are not specific to AM fungi,and a large proportion of the reads belonged to other fungal phyla.The taxonomic distribution and proportion of all sequences ob-tained in this study are shown in Fig. 2. Sequences belonging to theAscomycota accounted for the highest proportion of reads acrossall three land use types (�55%). Glomeromycota sequences rep-resented the second-largest phylum (13.9 to 16.7%), except innatural areas, where the proportion of sequences of Basidiomy-cota was higher (17.8%). Paired comparisons of land use type inthe prairie revealed higher relative abundances of AM fungal readsfrom roadsides and natural areas than from cropland (P � 0.0001)(Table 1). In contrast, there was no significant difference in therelative abundances of AM fungal reads in cropland and roadsidesamples from the Atlantic maritime region (P � 0.11).
In total, we detected 122 OTUs based on 97% similarity (seeFig. S1 in the supplemental material). In the prairie ecozone (n �317), we detected 120 AM fungal OTUs. The majority of the OTUsand reads belonged to the Glomeraceae (78 OTUs and 47.6% ofAM fungal reads) and Claroideoglomeraceae (30 OTUs and36.9% of AM fungal reads). The remaining OTUs belonged to theDiversisporaceae (9 OTUs and 6.6% of AM fungal reads), Gigas-
Land Use and AM Fungi in Canadian Ecozones
November 2013 Volume 79 Number 21 aem.asm.org 6721
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
poraceae (1 OTU), Archaeosporaceae (1 OTU), and Paraglomer-aceae (1 OTU) (Fig. S2).
Seventy-two AM fungal OTUs were detected in the Atlanticmaritime samples (n � 20). The majority of the OTUs representedthe Glomeraceae (37 OTUs) and Claroideoglomeraceae (26OTUs), but reads of Claroideoglomeraceae (49.7% of all AM fun-gal reads) were more abundant than reads of Glomeraceae (39.3%of all AM fungal reads). The remaining OTUs belonged to theDiversisporaceae (6 OTUs, 6.7% of the reads), Gigasporaceae (2OTUs, 0.8% of the read), and Paraglomeraceae (Fig. 3). Para-
glomeraceae was represented only by OTU2, but this OTU wasabundant in the Atlantic maritime ecozone, accounting for 3.4%of all AM fungal reads (Fig. 3). The archaeosporal OTU, whichshared 99% similarity with Archaeospora trappei (OTU1) and wasrare in the prairie, was undetected in the Atlantic maritime eco-zone.
Known reference sequences of Glomeromycota retrieved fromGenBank yielded good matches (�95% similarity) with 52 of theAM fungal OTUs. The remaining 70 OTUs yielded better matcheswith unknown or uncultured Glomeromycota sequences and had
FIG 2 Proportional abundances of the Glomeromycota, Ascomycota, Basidiomycota, other eukaryotes, and nonidentified fungi in soils of croplands, roadsides,and natural areas of the Canadian prairie, as represented by the numbers of reads belonging to these groups, which were obtained from 454 pyrosequencing ofamplicons.
TABLE 1 Effect of land use type on the relative abundance, richness, diversity, and community structure of AM fungi in the prairie and Atlanticmaritime ecozones of Canadaa
Ecozone Land use typeNo. ofsamples
No. of AMfungalOTUs
Relativeabundance (%) ACE Richness Shannon’s H
MRPP P valuefor AM fungalcommunity
Prairie provinces Cropland 107 90 15.3 � 1 16.1 � 2 7.7 � 0.7 1.6 � 0.09Roadside 107 110 16.7 � 1.4 33.3 � 3.1 14.1 � 0.9 2.3 � 0.07
P value by paired-samplet test
<0.0001 <0.0001 <0.0001 <0.0001 <0.000001
Prairie provinces Cropland 20 71 11.6 � 2.0 23.0 � 3.6 7.7 � 1.2 1.6 � 0.20Natural area 20 58 15.2 � 2.4 19.7 � 7.2 7.8 � 1.0 1.9 � 0.20
P value by paired-samplet test
<0.0001 0.17 0.93 0.33 0.38
Atlantic maritime Cropland 10 55 12.0 � 1.2 29.5 � 5.2 15.1 � 1.2 2.4 � 0.10Roadside 10 46 10.2 � 3.7 19.4 � 2.6 9.7 � 1.3 2.1 � 0.13
P value by paired-samplet test
0.11 0.11 0.006 0.06 <0.00017
All Cropland 176 104 13.7 � 0.8 18.6 � 1.6 b 8.3 � 0.53 b 1.6 � 0.07 bRoadside 117 112 16.0 � 1.3 34.3 � 3.2 a 13.8 � 0.79 a 2.3 � 0.07 aNatural area 24 58 15.2 � 2.4 19.8 � 6 b 7.8 � 0.92 b 1.9 � 0.14 b
P value by ANOVA 0.29 <0.0001 <0.0001 <0.0001 <0.000001a Paired univariate comparisons of land use types in each of the prairie and Atlantic maritime ecozones were analyzed using Student’s t test, and multivariate comparisons wereanalyzed by the multiresponse permutation procedures (MRPP). The significance of the land use type’s overall effects on univariate data was analyzed by ANOVA, whereas effectson community structure were tested by the MRPP. Means are presented with standard errors. Lowercase letters beside values indicate a significant difference between values withthe same letter, according to Duncan’s test. Boldface indicates statistically significant results.
Dai et al.
6722 aem.asm.org Applied and Environmental Microbiology
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
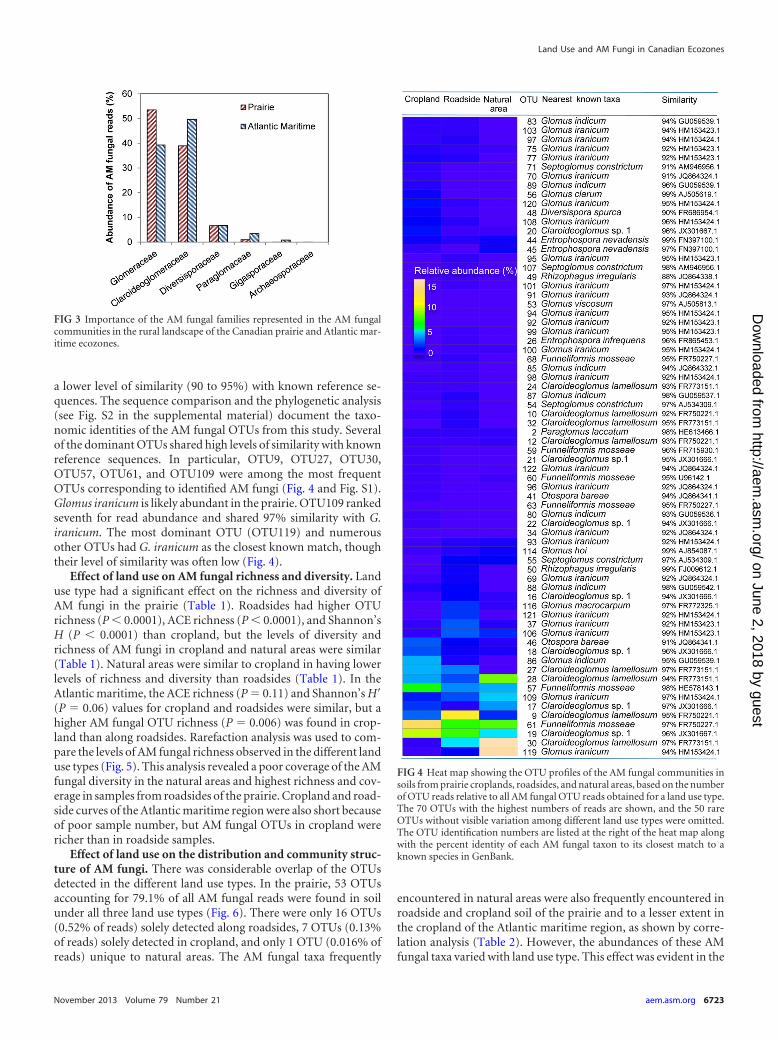
a lower level of similarity (90 to 95%) with known reference se-quences. The sequence comparison and the phylogenetic analysis(see Fig. S2 in the supplemental material) document the taxo-nomic identities of the AM fungal OTUs from this study. Severalof the dominant OTUs shared high levels of similarity with knownreference sequences. In particular, OTU9, OTU27, OTU30,OTU57, OTU61, and OTU109 were among the most frequentOTUs corresponding to identified AM fungi (Fig. 4 and Fig. S1).Glomus iranicum is likely abundant in the prairie. OTU109 rankedseventh for read abundance and shared 97% similarity with G.iranicum. The most dominant OTU (OTU119) and numerousother OTUs had G. iranicum as the closest known match, thoughtheir level of similarity was often low (Fig. 4).
Effect of land use on AM fungal richness and diversity. Landuse type had a significant effect on the richness and diversity ofAM fungi in the prairie (Table 1). Roadsides had higher OTUrichness (P � 0.0001), ACE richness (P � 0.0001), and Shannon’sH (P � 0.0001) than cropland, but the levels of diversity andrichness of AM fungi in cropland and natural areas were similar(Table 1). Natural areas were similar to cropland in having lowerlevels of richness and diversity than roadsides (Table 1). In theAtlantic maritime, the ACE richness (P � 0.11) and Shannon’s H=(P � 0.06) values for cropland and roadsides were similar, but ahigher AM fungal OTU richness (P � 0.006) was found in crop-land than along roadsides. Rarefaction analysis was used to com-pare the levels of AM fungal richness observed in the different landuse types (Fig. 5). This analysis revealed a poor coverage of the AMfungal diversity in the natural areas and highest richness and cov-erage in samples from roadsides of the prairie. Cropland and road-side curves of the Atlantic maritime region were also short becauseof poor sample number, but AM fungal OTUs in cropland werericher than in roadside samples.
Effect of land use on the distribution and community struc-ture of AM fungi. There was considerable overlap of the OTUsdetected in the different land use types. In the prairie, 53 OTUsaccounting for 79.1% of all AM fungal reads were found in soilunder all three land use types (Fig. 6). There were only 16 OTUs(0.52% of reads) solely detected along roadsides, 7 OTUs (0.13%of reads) solely detected in cropland, and only 1 OTU (0.016% ofreads) unique to natural areas. The AM fungal taxa frequently
encountered in natural areas were also frequently encountered inroadside and cropland soil of the prairie and to a lesser extent inthe cropland of the Atlantic maritime region, as shown by corre-lation analysis (Table 2). However, the abundances of these AMfungal taxa varied with land use type. This effect was evident in the
FIG 3 Importance of the AM fungal families represented in the AM fungalcommunities in the rural landscape of the Canadian prairie and Atlantic mar-itime ecozones.
FIG 4 Heat map showing the OTU profiles of the AM fungal communities insoils from prairie croplands, roadsides, and natural areas, based on the numberof OTU reads relative to all AM fungal OTU reads obtained for a land use type.The 70 OTUs with the highest numbers of reads are shown, and the 50 rareOTUs without visible variation among different land use types were omitted.The OTU identification numbers are listed at the right of the heat map alongwith the percent identity of each AM fungal taxon to its closest match to aknown species in GenBank.
Land Use and AM Fungi in Canadian Ecozones
November 2013 Volume 79 Number 21 aem.asm.org 6723
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
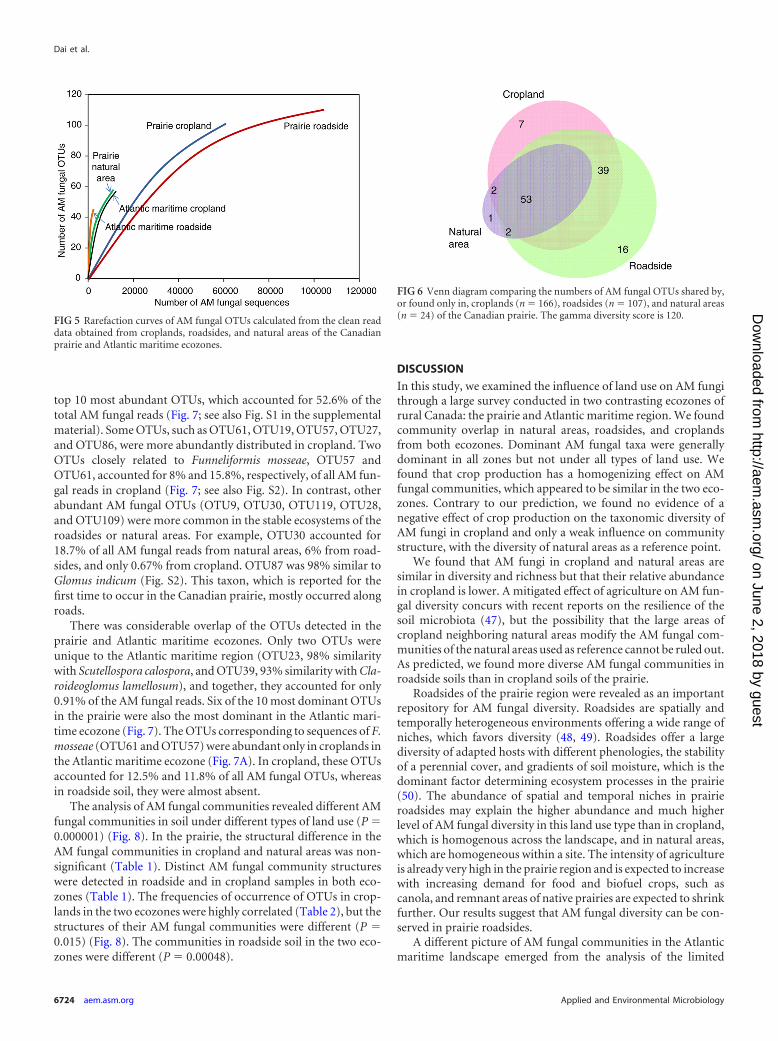
top 10 most abundant OTUs, which accounted for 52.6% of thetotal AM fungal reads (Fig. 7; see also Fig. S1 in the supplementalmaterial). Some OTUs, such as OTU61, OTU19, OTU57, OTU27,and OTU86, were more abundantly distributed in cropland. TwoOTUs closely related to Funneliformis mosseae, OTU57 andOTU61, accounted for 8% and 15.8%, respectively, of all AM fun-gal reads in cropland (Fig. 7; see also Fig. S2). In contrast, otherabundant AM fungal OTUs (OTU9, OTU30, OTU119, OTU28,and OTU109) were more common in the stable ecosystems of theroadsides or natural areas. For example, OTU30 accounted for18.7% of all AM fungal reads from natural areas, 6% from road-sides, and only 0.67% from cropland. OTU87 was 98% similar toGlomus indicum (Fig. S2). This taxon, which is reported for thefirst time to occur in the Canadian prairie, mostly occurred alongroads.
There was considerable overlap of the OTUs detected in theprairie and Atlantic maritime ecozones. Only two OTUs wereunique to the Atlantic maritime region (OTU23, 98% similaritywith Scutellospora calospora, and OTU39, 93% similarity with Cla-roideoglomus lamellosum), and together, they accounted for only0.91% of the AM fungal reads. Six of the 10 most dominant OTUsin the prairie were also the most dominant in the Atlantic mari-time ecozone (Fig. 7). The OTUs corresponding to sequences of F.mosseae (OTU61 and OTU57) were abundant only in croplands inthe Atlantic maritime ecozone (Fig. 7A). In cropland, these OTUsaccounted for 12.5% and 11.8% of all AM fungal OTUs, whereasin roadside soil, they were almost absent.
The analysis of AM fungal communities revealed different AMfungal communities in soil under different types of land use (P �0.000001) (Fig. 8). In the prairie, the structural difference in theAM fungal communities in cropland and natural areas was non-significant (Table 1). Distinct AM fungal community structureswere detected in roadside and in cropland samples in both eco-zones (Table 1). The frequencies of occurrence of OTUs in crop-lands in the two ecozones were highly correlated (Table 2), but thestructures of their AM fungal communities were different (P �0.015) (Fig. 8). The communities in roadside soil in the two eco-zones were different (P � 0.00048).
DISCUSSION
In this study, we examined the influence of land use on AM fungithrough a large survey conducted in two contrasting ecozones ofrural Canada: the prairie and Atlantic maritime region. We foundcommunity overlap in natural areas, roadsides, and croplandsfrom both ecozones. Dominant AM fungal taxa were generallydominant in all zones but not under all types of land use. Wefound that crop production has a homogenizing effect on AMfungal communities, which appeared to be similar in the two eco-zones. Contrary to our prediction, we found no evidence of anegative effect of crop production on the taxonomic diversity ofAM fungi in cropland and only a weak influence on communitystructure, with the diversity of natural areas as a reference point.
We found that AM fungi in cropland and natural areas aresimilar in diversity and richness but that their relative abundancein cropland is lower. A mitigated effect of agriculture on AM fun-gal diversity concurs with recent reports on the resilience of thesoil microbiota (47), but the possibility that the large areas ofcropland neighboring natural areas modify the AM fungal com-munities of the natural areas used as reference cannot be ruled out.As predicted, we found more diverse AM fungal communities inroadside soils than in cropland soils of the prairie.
Roadsides of the prairie region were revealed as an importantrepository for AM fungal diversity. Roadsides are spatially andtemporally heterogeneous environments offering a wide range ofniches, which favors diversity (48, 49). Roadsides offer a largediversity of adapted hosts with different phenologies, the stabilityof a perennial cover, and gradients of soil moisture, which is thedominant factor determining ecosystem processes in the prairie(50). The abundance of spatial and temporal niches in prairieroadsides may explain the higher abundance and much higherlevel of AM fungal diversity in this land use type than in cropland,which is homogenous across the landscape, and in natural areas,which are homogeneous within a site. The intensity of agricultureis already very high in the prairie region and is expected to increasewith increasing demand for food and biofuel crops, such ascanola, and remnant areas of native prairies are expected to shrinkfurther. Our results suggest that AM fungal diversity can be con-served in prairie roadsides.
A different picture of AM fungal communities in the Atlanticmaritime landscape emerged from the analysis of the limited
FIG 5 Rarefaction curves of AM fungal OTUs calculated from the clean readdata obtained from croplands, roadsides, and natural areas of the Canadianprairie and Atlantic maritime ecozones.
FIG 6 Venn diagram comparing the numbers of AM fungal OTUs shared by,or found only in, croplands (n � 166), roadsides (n � 107), and natural areas(n � 24) of the Canadian prairie. The gamma diversity score is 120.
Dai et al.
6724 aem.asm.org Applied and Environmental Microbiology
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
number of samples taken on podzolic soils in this ecozone. Crop-land in the Atlantic maritime hosted richer AM fungal communi-ties than roadsides, disproving both of our hypotheses of a nega-tive effect of agriculture on AM fungal communities and of a rolefor roadsides in the conservation of AM fungal diversity in thisecozone. However, these results suggested that AM fungal diver-sity can be increased by certain cropping practices, as reportedearlier (16, 51). All cropland in the Atlantic maritime ecozonewere on mixed farms under organic management that includedmixed perennial hay crops in their crop rotation system.
Diversity of AM fungal communities in the Canadian land-scape. The influence of land use on the communities of AM fungiin the Canadian landscape is largely unknown. We were surprisedto find similar levels of AM fungal diversity in natural areas and
adjacent croplands of the prairie. Numerous studies have reportednegative effects of crop production on AM fungi (52–55). Soiltillage has a negative impact on AM hyphal network biomass andinfectivity (56, 57) and on the richness of AM fungal communities(58–60), but in the Canadian prairie, soils are rarely tilled (25), asnot tilling is a soil water conservation practice (61, 62). Prairiesoils were commonly tilled 2 decades ago, but tillage was shallow(10 cm) (61, 63) and rarely practiced in the fall. Therefore, theinfluence of tillage on AM fungal communities is likely to be min-imal in the prairie. In the Atlantic maritime ecozone, where soiltillage is commonly practiced and more intensive, AM fungal spe-cies richness was higher in cropland than in roadsides (Table 1),suggesting that tillage is not the main driver of the AM fungaldiversity in croplands. The addition of nitrogen fertilizer to soildecreases soil pH and can affect AM fungal richness in cropland(64, 65), but prairie soils are naturally well buffered, and fertilizersare usually applied parsimoniously since water availability ratherthan soil fertility normally limits yield. For the same reason, thelevels of soil P fertility, which influence AM fungal activity anddevelopment (12, 17, 66–70), may have been similar in croplandand natural areas.
A few generalist AM fungal taxa were very abundant. General-
TABLE 2 Coefficients of correlationa associated with the relationship between the frequencies of occurrence of AM fungal taxa in soils underdifferent types of land use in the prairie and the Atlantic maritime ecozones
Ecozone
r2 for:
Atlantic maritimecropland
Atlantic maritimeroadside
Prairiecropland
Prairieroadside
Prairie naturalareas
Atlantic maritime cropland 1 0.5574 0.8629 0.7452 0.7456Atlantic maritime roadside 0.5574 1 0.5494 0.7072 0.5756Prairie cropland 0.8629 0.5494 1 0.7873 0.8403Prairie roadside 0.7452 0.7072 0.7873 1 0.8300Prairie natural areas 0.7456 0.5756 0.8403 0.8300 1a According to Spearman correlation analysis (P � 0.01; n � 122).
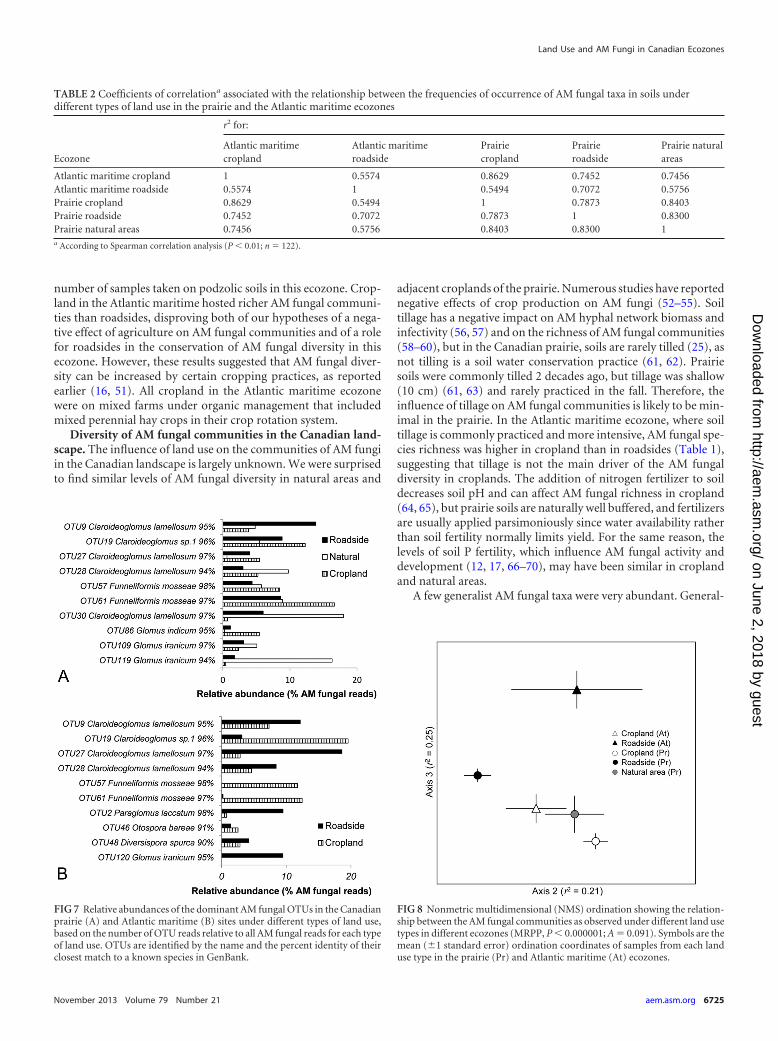
FIG 7 Relative abundances of the dominant AM fungal OTUs in the Canadianprairie (A) and Atlantic maritime (B) sites under different types of land use,based on the number of OTU reads relative to all AM fungal reads for each typeof land use. OTUs are identified by the name and the percent identity of theirclosest match to a known species in GenBank.
FIG 8 Nonmetric multidimensional (NMS) ordination showing the relation-ship between the AM fungal communities as observed under different land usetypes in different ecozones (MRPP, P � 0.000001; A � 0.091). Symbols are themean (�1 standard error) ordination coordinates of samples from each landuse type in the prairie (Pr) and Atlantic maritime (At) ecozones.
Land Use and AM Fungi in Canadian Ecozones
November 2013 Volume 79 Number 21 aem.asm.org 6725
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

ist AM fungi are predicted to be pioneer species with a high dis-persal capacity (71). If we accept high read abundance and fre-quency as evidence of high dispersal capacity, two AM fungal taxacorresponding to F. mosseae correspond to this description. Thesetwo OTUs, OTU61 and OTU57, had very high frequencies andabundances in cropland. F. mosseae is known as a cosmopolitanspecies (72) that is particularly successful in croplands throughoutthe world (73). Although we cannot infer the identities of OTU61and OTU57 based on their high levels of similarity with knownspecies, due to the short lengths of our amplicons, we know fromthe morphological examination of trap cultures from croplandsmade by us (unpublished) and by others (74) that F. mosseae iscommon in cultivated prairie soils. The abundance of these two F.mosseae-like OTUs was a striking characteristic of cropland in theAtlantic maritime ecozone, as these were very rare in adjacentroadside soils in this ecozone (Fig. 7). However, these OTUs weredominant throughout the prairie, suggesting that the AM fungithat they represent are favored by crop production and dry envi-ronments, which agrees with previous reports of the preference ofF. mosseae for semiarid rather than mesic environments (75). Theinfluence of these F. mosseae-like taxa on crop productivity is un-known.
The relative similarity of the AM fungal communities of crop-lands in contrasting ecozones was surprising. The uniformity ofplant cover within croplands may have a homogenizing effect onAM fungal communities. We based this hypothesis on publishedevidence of the selective influence of plants on extraradical AMfungal growth (14) and on the structure of AM fungal communi-ties in soil (76, 77). Host plants have a large influence in shapingAM fungal communities in soils (78, 79). The obvious differencebetween the environments created by the different land use typeswas plant cover. In both ecozones, all croplands were planted withwheat the year of sampling. Although some cropping systems in-clude a phase with a perennial crop, as was the case on the crop-land in the Atlantic maritime ecozone, croplands are normallyunder the influence of plants with short annual life cycles, and theyare usually devoid of living plants during most of the year at theselatitudes.
Within-group variation may explain the low AM fungal speciesrichness and diversity index found in natural areas. Natural areaswere a heterogeneous group. The forms of different plant coverswent from mixed-grass vegetation in the semiarid zone to poplarand willow groves in the parkland to spruce forest in the borealforest, but plant diversity was relatively low in any given naturalarea. The AM fungal communities associated with these differentplant covers may also be variable.
In contrast to the vegetation of natural areas, which was dis-tinct in the edaphoclimatic zones of the prairie, the vegetation ofroadsides was relatively the same everywhere, as they were initiallyplanted with the same persistent grass species, but their plantcover was often diverse within a sampling site. Roadside standsincluded several weedy species adapted to specific edaphoclimaticzones but also plant species with different adaptations to soilmoisture, which grew at different elevations of the roadside. Themain role of roadsides is to drain water off the roads. The temporaldiversity created by the phenology of the plant species making upthe vegetation cover in roadsides is compounded by the practice ofmowing. The heterogeneity of the roadside environment favorsdiversity. The roadside environment also differs from other types
of land use in being less limited by water, a factor that may benefitAM fungal growth (75).
Our observation suggests that cropland is a type of land useconducive to the maintenance of healthy AM fungal communitiesin the Atlantic maritime ecozone, unless OTU61 and OTU57 rep-resent generalist taxa that are harmful. The AM fungal species thatpreferentially associate with crop plants may outcompete lesscompatible species, which may disappear over time (24). The factthat all but one AM fungal OTU found in natural areas was alsofound in cropland and roadside soils (Fig. 6) suggests that few ofthe indigenous AM fungal taxa have been lost due to human ac-tivity, at least in the prairie ecozone. However, the land was not allbroken at the same time in the prairie, and it is possible that cer-tain AM fungal taxa extirpated from lands with a long history ofcrop production still remain in lands that were broken in morerecent times.
AM fungal resource in the Canadian prairie. Our survey ofthe AM fungal communities of the Canadian prairie was exten-sive. It constitutes the first attempt to understand the AM fungalresource hosted in this globally important wheat-producing re-gion (80). This survey and data set constitute a baseline that allowsus to track changes in AM fungal resources that can be caused bythe fortuitous introduction of invasive species through the prac-tice of AM fungal inoculation (71), by other anthropological ac-tivities, or by climate change. Our survey spanned a dry and a wetyear on the prairie, which should be conducive to the detection ofAM fungal taxa with different life history traits and to the captureof diversity.
Overall, the diversity and abundance of the Glomeraceae, rep-resented by four genera, is revealed as a feature of the AM fungalcommunities of the prairie soils, compared to the Atlantic mari-time soils, where Claroideoglomeraceae were more abundant.Claroideoglomeraceae, represented by three genera, were secondto Glomeraceae in terms of diversity and abundance in prairiesoils. This family was particularly abundant in roadsides and nat-ural areas. The scarcity of Paraglomus also appears as a character-istic of the AM fungal communities of prairie soils compared tothose of the Atlantic maritime ecozone, where Paraglomus was adominant OTU.
Our study reports for the first time the presence of AM fungirelated to Glomus indicum in the Canadian prairie. We recentlyreported AM fungi related to Glomus iranicum in cultivated fieldsof the prairie ecozone (81) and now confirm the importance ofthis group. The large number of OTUs clustering with G. iranicumin the phylogenetic analysis and the relative abundances of certainof these OTUs (e.g., OTU109 is the 7th-most-abundant OTU),support the idea that G. iranicum-related taxa are well adapted tothe prairie ecozone. This concurs with the initial discovery of thesetaxa in the semiarid wheat-growing region of southwestern Iran(82). We report for the first time the presence of Archaeosporaceaein the prairie based on the detection of an OTU sharing 99%similarity with Archaeospora trappei. Our observation of onemember of the family Paraglomeraceae sharing 97% of similaritywith Paraglomus brasilianum concurs with the recent report of thepresence of Paraglomus in the Canadian prairie (83), and our ob-servation of a rare sequence clustering with Scutellospora concurswith the earlier report of Scutellospora calospora in prairie crop-land (84). These earlier reports were also based on short sequencesof the 18S rRNA genes, which are of insufficient length for speciesidentification, highlighting the need for AM fungal surveys based
Dai et al.
6726 aem.asm.org Applied and Environmental Microbiology
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

on morphology or for better molecular methods to further im-prove our understanding of the diversity of these important fungiin the prairie.
Conclusions. From this extensive survey of the AM fungal di-versity contained in soils of the Canadian prairie landscape and in20 soils of the Atlantic maritime ecozone, we can conclude thatland use type influences the abundance of a set of AM fungal taxalargely shared by cropland, roadsides, and natural areas of theprairie and the Atlantic maritime ecozones. The attributes of theplant cover and the soil moisture availability level associated withland use appears as the most likely factors shaping the structure ofAM fungal communities in the Canadian landscape. Roadsidesbear a high degree of heterogeneity and offer soil moisture andmultiple niches for the conservation of AM fungal diversity in theprairie region. We found no evidence of a negative impact of cropproduction on the diversity of AM fungal communities in oursurvey, although it does influence the structures of these commu-nities.
ACKNOWLEDGMENTS
This study was funded by the Organic Science Cluster, project A, activity2 (project no. 04340 grant to C. Hamel) and a Ministry of Education Ph.D.scholarship granted by the Chinese Ministry of Education to M. Dai. Wethank Xiaohong Yang and Zhiqing Zhou for support of M. Dai’s study inCanada.
We gratefully acknowledge the assistance of Kira Kotilla, Henry Jen-zen, Newton Lupwayi, Sukhdev Malhi, Guy Lafond, Reynald Lemke, Cyn-thia Grant, numerous seed growers, and organic growers of Saskatchewanfor the provision of samples and sampling sites, Dallas Thomas for bioin-formatics assistance, Marc St. Arnaud for navigation in the landscape, andKeith Hanson and Elijah Atuku for technical assistance.
REFERENCES1. Gianinazzi S, Gollotte A, Binet MN, van Tuinen D, Redecker D, Wipf
D. 2010. Agroecology: the key role of arbuscular mycorrhizas in ecosystemservices. Mycorrhiza 20:519 –530.
2. Fitter AH, Helgason T, Hodge A. 2011. Nutritional exchanges in thearbuscular mycorrhizal symbiosis: implications for sustainable agricul-ture. Fungal Biol. Rev. 25:68 –72.
3. Smith SE, Read DJ. 2008. Mycorrhizal symbiosis. Academic Press, NewYork, NY.
4. Doubková P, Suda J, Sudová R. 2012. The symbiosis with arbuscularmycorrhizal fungi contributes to plant tolerance to serpentine edaphicstress. Soil Biol. Biochem. 44:56 – 64.
5. Hawkins HJ, Johansen A, George E. 2000. Uptake and transport oforganic and inorganic nitrogen by arbuscular mycorrhizal fungi. PlantSoil 226:275–285.
6. Smith SE, Facelli E, Pope S, Smith FA. 2010. Plant performance instressful environments: interpreting new and established knowledge of theroles of arbuscular mycorrhizas. Plant Soil 326:3–20.
7. van der Heijden MGA. 2010. Mycorrhizal fungi reduce nutrient loss frommodel grassland ecosystems. Ecology 91:1163–1171.
8. Birhane E, Sterck FJ, Fetene M, Bongers F, Kuyper TW. 2012. Arbus-cular mycorrhizal fungi enhance photosynthesis, water use efficiency, andgrowth of frankincense seedlings under pulsed water availability condi-tions. Oecologia 169:895–904.
9. Ruth B, Khalvati M, Schmidhalter U. 2011. Quantification of mycorrhi-zal water uptake via high-resolution on-line water content sensors. PlantSoil 342:459 – 468.
10. Garg N, Chandel S. 2011. Effect of mycorrhizal inoculation on growth,nitrogen fixation, and nutrient uptake in Cicer arietinum L. under saltstress. Turk. J. Agr. For. 35:205–214.
11. Rillig MC, Mummey DL. 2006. Mycorrhizas and soil structure. NewPhytol. 171:41–53.
12. Gosling P, Hodge A, Goodlass G, Bending GD. 2006. Arbuscular my-corrhizal fungi and organic farming. Agric. Ecosyst. Environ. 113:17–35.
13. Kahiluoto H, Vestberg M. 1998. The effect of arbuscular mycorrhiza on
biomass production and phosphorus uptake from sparingly solublesources by leek (Allium porrum L.) in Finnish field soils. Biol. Agric. Hor-tic. 16:65– 85.
14. Kiers ET, Duhamel M, Beesetty Y, Mensah JA, Franken O, VerbruggenE, Fellbaum CR, Kowalchuk GA, Hart MM, Bago A, Palmer TM, WestSA, Vandenkoornhuyse P, Jansa J, Bücking H. 2011. Reciprocal rewardsstabilize cooperation in the mycorrhizal symbiosis. Science 333:880 – 882.
15. An ZQ, Hendrix JW, Hershman DE, Ferriss RS, Henson GT. 1993. Theinfluence of crop rotation and soil fumigation on a mycorrhizal fungalcommunity associated with soybean. Mycorrhiza 3:171–182.
16. Oehl F, Sieverding E, Ineichen K, Mäder P, Boller T, Wiemken A. 2003.Impact of land use intensity on the species diversity of arbuscular mycor-rhizal fungi in agroecosystems of Central Europe. Appl. Environ. Micro-biol. 69:2816 –2824.
17. Fester T, Sawers R. 2011. Progress and challenges in agricultural appli-cations of arbuscular mycorrhizal fungi. Crit. Rev. Plant Sci. 30:459 – 470.
18. Anderson EL, Millner PD, Kunishi HM. 1987. Maize root length densityand mycorrhizal infection as influenced by tillage and soil phosphorus. J.Plant Nutr. 10:1349 –1356.
19. Jakobsen I, Nielsen NE. 1983. Vesicular-arbuscular mycorrhiza in field-grown crops. I. Mycorrhizal infection in cereals and peas at various timesand soil depths. New Phytol. 93:401– 413.
20. Santos-González JC, Nallanchakravarthula S, Alström S, Finlay RD.2011. Soil, but not cultivar, shapes the structure of arbuscular mycorrhizalfungal assemblages associated with strawberry. Microb. Ecol. 62:25–35.
21. Wang FY, Tong RJ, Shi Zy Xu XF, He XH. 2011. Inoculations witharbuscular mycorrhizal fungi increase vegetable yields and decreasephoxim concentrations in carrot and green onion and their soils. PLoSOne 6(2):e16949. doi:10.11371/journal.pone.0016949.
22. Helgason T, Daniell TJ, Husband R, Fitter AH, Young JPW. 1998.Ploughing up the wood-wide web? Nature 394:431. doi:10.1038/28764.
23. Hijri I, Sýkorová Z, Oehl F, Ineichen K, Mäder P, Wiemken A, Re-decker D. 2006. Communities of arbuscular mycorrhizal fungi in arablesoils are not necessarily low in diversity. Mol. Ecol. 15:2277–2289.
24. Verbruggen E, Van Der Heijden MGA, Weedon JT, Kowalchuk GA,Rö-Ling WFM. 2012. Community assembly, species richness and nested-ness of arbuscular mycorrhizal fungi in agricultural soils. Mol. Ecol. 21:2341–2353.
25. Huffman T, Eilers W. 2011. Farm land management. 4. Agricultural landuse. In Environmental sustainability of Canadian agriculture: agri-environmental indicator report series, report no. 3. Agriculture andAgri-Food Canada, Ottawa, Ontario, Canada. http://publications.gc.ca/site/archivee-archived.html?url�http://publications.gc.ca/collections/collection_2011/agr/A22-201-2010-eng.pdf.
26. Ecological Stratification Working Group. 1996. A national ecologicalframework for Canada. Minister of Supply and Services Canada, Ottawa,Ontario, Canada. http://sis.agr.gc.ca/cansis/publications/ecostrat/cad_report.pdf.
27. Soil Classification Working Group. 1998. The Canadian system of soilclassification, 3rd ed. Agriculture and Agri-Food Canada Publication1646. NRC Research Press, Ottawa, ON, Canada. http://sis.agr.gc.ca/cansis/publications/manuals/1998-cssc-ed3/cssc3_manual.pdf.
28. Campbell CA, Zentner RP, Janzen HH, Bowren KE. 1990. Crop rotationstudies on the Canadian prairies. Research Branch, Agriculture Canadapublication 1841/E. Research Branch, Agriculture Canada, Ottawa, ON,Canada.
29. Roche. 2009. Technical bulletin. Genome sequencer FLX system. Ampli-con fusion primer design guidelines for GS FLX titanium series Lib-Achemistry. TCB no. 013-2009. Roche, Basel, Switzerland. http://www.igsb.org/uploads/pdf/TCB-09013_AmpliconFusionPrimerDesignGuidelines.pdf.
30. Lumini E, Orgiazzi A, Borriello R, Bonfante P, Bianciotto V. 2010.Disclosing arbuscular mycorrhizal fungal biodiversity in soil through aland-use gradient using a pyrosequencing approach. Environ. Microbiol.12:2165–2179.
31. Sato K, Suyama Y, Saito M, Sugawara K. 2005. A new primer fordiscrimination of arbuscular mycorrhizal fungi with polymerase chainreaction-denature gradient gel electrophoresis. Jpn. Soc. Grassl. Sci. 51:179 –181.
32. Polz MF, Cavanaugh CM. 1998. Bias in template-to-product ratios inmultitemplate PCR. Appl. Environ. Microbiol. 64:3724 –3730.
33. Huse SM, Welch DM, Morrison HG, Sogin ML. 2010. Ironing out the
Land Use and AM Fungi in Canadian Ecozones
November 2013 Volume 79 Number 21 aem.asm.org 6727
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

wrinkles in the rare biosphere through improved OTU clustering. Envi-ron. Microbiol. 12:1889 –1898.
34. Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB,Lesniewski RA, Oakley BB, Parks DH, Robinson CJ, Sahl JW, Stres B,Thallinger GG, Van Horn DJ, Weber CF. 2009. Introducing mothur:open-source, platform-independent, community-supported software fordescribing and comparing microbial communities. Appl. Environ. Micro-biol. 75:7537–7541.
35. Edgar RC. 2004. MUSCLE: multiple sequence alignment with improvedaccuracy and speed, p 728 –729. In . Proceedings of the 2004 IEEE Com-putational Systems Bioinformatics Conference (CSB 2004). The IEEEComputer Society, Washington, DC. doi:10.1109/CSB.2004.1332560.
36. Zhang Z, Schwartz S, Wagner L, Miller W. 2000. A greedy algorithm foraligning DNA sequences. J. Comput. Biol. 7:203–214.
37. Saitou N, Nei M. 1987. The neighbor-joining method: a new method forreconstructing phylogenetic trees. Mol. Biol. Evol. 4:406 – 425.
38. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011.MEGA5: molecular evolutionary genetics analysis using maximum likeli-hood, evolutionary distance, and maximum parsimony methods. Mol.Biol. Evol. 28:2731–2739.
39. Hall BG. 2004. Phylogenetic trees made easy. A how-to manual. SinauerAssociates, Sunderland, MA.
40. Redecker D, Schüßler A, Stockinger H, Stürmer SL, Morton JB, WalkerC. 2013. An evidence-based consensus for the classification of arbuscularmycorrhizal fungi (Glomeromycota). Mycorrhiza doi:10.1007/s00572-013-0486-y.
41. Peck JE. 2010. Multivariate analysis for community ecologists: step-by-step using PC-ORD. MjM Software Design, Gleneden Beach, OR.
42. de Mendiburu F. 2013. Statistical procedures for agricultural research.Package ‘Agricolae,’ version 1.4-4. Comprehensive R Archive Network,Institute for Statistics and Mathematics, Vienna, Austria. http://cran.r-project.org/web/packages/agricolae/agricolae.pdf.
43. McCune B, Grace JB. 2002. Analysis of ecological communities. MjMSoftware Design, Gleneden Beach, OR.
44. Legendre P, Legendre L. 1998. Numerical ecology. Elsevier, Amsterdam,Netherlands.
45. Warnes GR, Bolker B, Bonebakker L, Gentleman R, Liaw WHA, Lumley T,Maechler M, Magnusson A, Moeller S, Schwartz M, Venables B. 2013.Various R programming tools for plotting data. Package ‘gplots,’ version2.11.3. Comprehensive R Archive Network, Institute for Statistics and Math-ematics, Vienna, Austria. http://cran.r-project.org/web/packages/gplots/gplots.pdf.
46. Neuwirth E. 2011. ColorBrewer palettes. Package ‘RColorBrewer,’ ver-sion 1.0-5. Comprehensive R Archive Network, Institute for Statistics andMathematics, Vienna, Austria. http://cran.r-project.org/web/packages/RColorBrewer/RColorBrewer.pdf
47. Postma-Blaauw MB. 2010. Soil biota community structure and abun-dance under agricultural intensification and extensification. Ecology 91:460 – 473.
48. Andersen R, Chapman SJ, Artz RRE. 2013. Microbial communities innatural and disturbed peatlands: a review. Soil Biol. Biochem. 57:979 –994.
49. Richardson PJ, MacDougall AS, Larson DW. 2012. Fine-scale spatialheterogeneity and incoming seed diversity additively determine plant es-tablishment. J. Ecol. 100:939 –949.
50. Hamel C, Schellenberg MP, Hanson K, Wang H. 2007. Evaluation of the“bait-lamina test” to assess soil microfauna feeding activity in mixed grass-land. Appl. Soil Ecol. 36:199 –204.
51. Jansa J, Wiemken A, Frossard E. 2006. The effects of agricultural prac-tices on arbuscular mycorrhizal fungi. Geol. Soc. Lond. Spec. Publ. 266:89 –115. doi:10.1144/GSL.SP.2006.266.01.01.
52. Douds DD, Jr, Millner PD. 1999. Biodiversity of arbuscular mycorrhizalfungi in agroecosystems. Agric. Ecosyst. Environ. 74:77–93.
53. Jansa J, Mozafar A, Banke S, McDonald BA, Frossard E. 2002. Intra- andintersporal diversity of ITS rDNA sequences in Glomus intraradices assessedby cloning and sequencing, and by SSCP analysis. Mycol. Res. 106:670–681.
54. Jumpponen A, Trowbridge J, Mandyam K, Johnson L. 2005. Nitrogenenrichment causes minimal changes in arbuscular mycorrhizal coloniza-tion but shifts community composition— evidence from rDNA data. Biol.Fertil. Soils 41:217. doi:10.1007/s00374-005-0845-8.
55. Oehl F, Sieverding E, Mäder P, Dubois D, Ineichen K, Boller T, WiemkenA. 2004. Impact of long-term conventional and organic farming on the diver-sity of arbuscular mycorrhizal fungi. Oecologia 138:574–583.
56. Dodd J, Boddington C, Rodriguez A, Gonzalez-Chavez C, Mansur I.2000. Mycelium of arbuscular mycorrhizal fungi (AMF) from differentgenera: form, function and detection. Plant Soil 226:131–151.
57. Schnoor TK, Lekberg Y, Rosendahl S, Olsson PA. 2011. Mechanical soildisturbance as a determinant of arbuscular mycorrhizal fungal communi-ties in semi-natural grassland. Mycorrhiza 21:211–220.
58. Alguacil MM, Lumini E, Roldàn A, Salinas-Garcia JR, Bonfante P,Bianciotto V. 2008. The impact of tillage practices on arbuscular my-corrhizal fungal diversity in subtropical crops. Ecol. Appl. 18:527–536.
59. Brito I, Goss MJ, de Carvalho M, Chatagnier O, van Tuinen D. 2012.Impact of tillage system on arbuscular mycorrhiza fungal communities inthe soil under Mediterranean conditions. Soil Till. Res. 121:63– 67.
60. Schnoor TK, Mårtensson LM, Olsson PA. 2011. Soil disturbance altersplant community composition and decreases mycorrhizal carbon alloca-tion in a sandy grassland. Oecologia 167:809 – 819. doi:10.1007/s00442-011-2020-2.
61. Grevers MC, Kirkland JA, De Jong E, Rennie DA. 1986. Soil waterconservation under zero- and conventional tillage systems on the Cana-dian prairies. Soil Till. Res. 8:265–276.
62. Lafond GP, May WE, Stevenson FC, Derksen DA. 2006. Effects of tillagesystems and rotations on crop production for a thin Black Chernozem inthe Canadian Prairies. Soil Till. Res. 89:232–245.
63. Campbell CA, Read DWL, Zentner RP, Leyshon AJ, Ferguson WS.1983. First 12 years of a long-term crop rotation study in southwesternSaskatchewan—yields and quality of grain. Can. J. Plant Sci. 63:91–108.
64. Oehl F, Laczko E, Bogenrieder A, Stahr K, Bösch R, van der Heijden M,Sieverding E. 2010. Soil type and land use intensity determine the com-position of arbuscular mycorrhizal fungal communities. Soil Biol.Biochem. 42:724 –738.
65. Oehl F, Sieverding E, Ineichen K, Mäder P, Wiemken A, Boller T. 2009.Distinct sporulation dynamics of arbuscular mycorrhizal fungal commu-nities from different agroecosystems in long-term microcosms. Agric.Ecosyst. Environ. 134:257–268.
66. Gosling P, Ozaki A, Jones J, Turner M, Rayns F, Bending GD. 2010.Organic management of tilled agricultural soils results in a rapid increasein colonisation potential and spore populations of arbuscular mycorrhizalfungi. Agric. Ecosyst. Environ. 139:273–279.
67. Koide RT. 1991. Tansley review no. 29. Nutrient supply, nutrient demandand plant response to mycorrhizal infection. New Phytol. 117:365–386.
68. Lakshmipathy A, Gowda B, Bagyaraj DJ. 2003. VA mycorrhizal coloni-zation pattern in RET medicinal plants (Mammea suriga, Saraca asoca,Garcinia spp., Embelia ribes and Calamus sp.) in different parts of Karna-taka. Asian J. Microbiol. Biotechnol. Environ. Sci. 5:505–508.
69. Lakshmipathy R, Balakrishna AN, Bagyaraj DJ. 2012. Abundance anddiversity of AM fungi across a gradient of land use intensity and theirseasonal variations in Niligiri Biosphere of the Western Ghats, India. J.Agric. Sci. Technol. 14:903.
70. Van Aarle IM, Plassard C. 2010. Spatial distribution of phosphataseactivity associated with ectomycorrhizal plants is related to soil type. SoilBiol. Biochem. 42:324 –330.
71. Schwartz MW, Hoeksema JD, Gehring CA, Johnson NC, KlironomosJN, Abbott LK, Pringle A. 2006. The promise and the potential conse-quences of the global transport of mycorrhizal fungal inoculum. Ecol.Lett. 9:501–515.
72. Avio L, Cristani CC, Strani P, Giovannetti M. 2009. Genetic and phe-notypic diversity of geographically different isolates of Glomus mosseae.Can. J. Microbiol. 55:242–253.
73. Rosendahl S, McGee P, Morton JB. 2009. Lack of global populationgenetic differentiation in the arbuscular mycorrhizal fungus Glomus mos-seae suggests a recent range expansion which may have coincided with thespread of agriculture. Mol. Ecol. 18:4316 – 4329.
74. Talukdar NC, Germida JJ. 1993. Occurrence and isolation of vesicular-arbuscular mycorrhizae in cropped field soils of Saskatchewan, Canada.Can. J. Microbiol. 39:567–575.
75. Egerton-Warburton LM, Johnson NC, Allen EB. 2007. Mycorrhizalcommunity dynamics following nitrogen fertilization: a cross-site test infive grasslands. Ecol. Monogr. 77:527–544.
76. Fitzsimons MS, Miller RM, Jastrow JD. 2008. Scale-dependent nicheaxes of arbuscular mycorrhizal fungi. Oecologia 158:117–127.
77. Johnson NC, Tilman D, Wedin D. 1992. Plant and soil controls onmycorrhizal fungal communities. Ecology 73:2034 –2042.
Dai et al.
6728 aem.asm.org Applied and Environmental Microbiology
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

78. Al-Yahya’ei MN, Oehl F, Vallino M, Lumini E, Redecker D, WiemkenA, Bonfante P. 2011. Unique arbuscular mycorrhizal fungal communitiesuncovered in date palm plantations and surrounding desert habitats ofsouthern Arabia. Mycorrhiza 21:195–209.
79. Yang C, Hamel C, Schellenberg MP, Perez JC, Berbara RL. 2010.Diversity and functionality of arbuscular mycorrhizal fungi in three plantcommunities in semiarid Grasslands National Park, Canada. Microb.Ecol. 59:724 –733.
80. Curtis BC, Rajaram S, Macpherson HG (ed). 2002. Wheat in the world.Bread wheat improvement and production. FAO, Rome, Italy. http://www.fao.org/docrep/006/Y4011E/Y4011E00.HTM.
81. Dai M, Hamel C, St Arnaud M, He Y, Grant C, Lupwayi N, Janzen H,Malhi S S, Yang X, Zhou Z. 2012. Arbuscular mycorrhizal fungi assem-
blages in Chernozem great groups revealed by massively parallel pyrose-quencing. Can. J. Microbiol. 58:81–92.
82. Błaszkowski J, Kovács GM, Balázs TK, Orłowska E, Sadravi M, WubetT, Buscot F. 2010. Glomus africanum and G. iranicum, two new species ofarbuscular mycorrhizal fungi (Glomeromycota). Mycologia 102:1450 –1462.
83. Ellouze W, Hamel C, Vujanovic V, Gan YT, Bouzid S, St-Arnaud M.2013. Chickpea genotypes shape the soil microbiome and affect the estab-lishment of the subsequent durum wheat crop in the semiarid NorthAmerican Great Plains. Soil Biol. Biochem. 63:129 –141.
84. Ma WK, Siciliano SD, Germida JJ. 2005. A PCR-DGGE method fordetecting arbuscular mycorrhizal fungi in cultivated soils. Soil Biol.Biochem. 37:1589 –1597.
Land Use and AM Fungi in Canadian Ecozones
November 2013 Volume 79 Number 21 aem.asm.org 6729
on June 2, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from