improvement in simple artificial insemination in sheep
TRANSCRIPT

University of Arkansas, FayettevilleScholarWorks@UARK
Theses and Dissertations
8-2017
Improvement in Simple Artificial Insemination inSheep Using Chilled Extended Semen andConsideration of Fertility in EwesMohan AcharyaUniversity of Arkansas, Fayetteville
Follow this and additional works at: http://scholarworks.uark.edu/etd
Part of the Animal Studies Commons, and the Sheep and Goat Science Commons
This Dissertation is brought to you for free and open access by ScholarWorks@UARK. It has been accepted for inclusion in Theses and Dissertations byan authorized administrator of ScholarWorks@UARK. For more information, please contact [email protected], [email protected].
Recommended CitationAcharya, Mohan, "Improvement in Simple Artificial Insemination in Sheep Using Chilled Extended Semen and Consideration ofFertility in Ewes" (2017). Theses and Dissertations. 2502.http://scholarworks.uark.edu/etd/2502

Improvement in Simple Artificial Insemination in Sheep Using Chilled Extended Semen and
Consideration of Fertility in Ewes
A dissertation submitted in partial fulfillment
of the requirement for the degree of
Doctor of Philosophy in Animal Science
by
Mohan Acharya
Tribhuvan University
Bachelor of Veterinary Science and Animal Husbandry, 2010
University of Arkansas
Master of Science in Animal Science, 2014
August 2017
University of Arkansas
This dissertation is approved for recommendation to the Graduate Council.
Rick W. Rorie, Ph.D
Dissertation Director
Joan M. Burke, Ph.D. Kenneth Coffey, Ph.D.
Committee Member Committee Member
Dan. J. Donoghue, Ph.D.
Committee Member

Abstract
Three studies were conducted to improve simple artificial insemination in sheep using
chilled extended semen and evaluate methods to predict fertility in ewes. The first study
evaluated the effect of semen extenders and storage temperature on motility parameters of ram
semen stored for up to 72 h. Single semen collected from each ram was divided into four parts
and extended using four extenders: milk, TRIS, TEST, and CJ-2, diluted to 600 million sperm/ml
and loaded into 0.25 ml straws for storage at 4 and 15° C. Most of the CASA motility parameters
were higher after extension and storage at 4 compared with 15° C (P < 0.05). Ram semen stored
using milk or TRIS based extenders at 4° C maintained similar sperm motility parameters but
both were higher compared with TEST or CJ-2 extenders after storage for up to 3 d (P < 0.05).
The second study evaluated the effects of different combinations of extenders, ethylene glycol
(EG), egg yolk and penicillamine, hypotaurine and epinephrine (PHE) on ram sperm motility
parameters during storage. Semen electroejaculated from each ram was distributed across
treatment combinations consisting of either TRIS or milk extenders supplemented with 5 or 20%
(v/v) egg yolk, ± EG, and ± PHE. Milk extender supplemented with 1% EG, PHE and either 5 or
20% egg yolk can protect sperm cells better than other extender and supplement combination
after storage at 4° C for up to 72 h. The third study was conducted to correlate serum AMH and
lambs born with estimated breeding values determined through National Sheep Improvement
Program for reproductive performance (NLB or number of lambs born; NLW or number of
lambs weaned; MWWT or maternal weaning weight; and the Katahdin index) and WWT or
weaning weight. Additionally, correlation between AMH quartiles and lambs born after fall
versus summer breeding was determined. There was no correlation between serum AMH and
EBVs for NLB, NLW, MWWT, WWT and the Katahdin index in Katahdin ewes (P > 0.05).

Ewes in the lowest quartile (Q1) had lower number of offsprings born from summer breeding
compared with ewes in other AMH quartiles.

Acknowledgements
I would like to express my deepest gratitude and sincere appreciation to my research
advisor Dr. Joan M. Burke. Thank you for all course of studies, research, and training
opportunities you provided during 5 years (MS and PhD). You have thousands of qualities to be
a good advisor.
I would like to thank Dr. Rorie for accepting me as a PhD student from pile of applicants.
I will be missing every morning chats. You have good sense of humor behind quietness. Thank
you for providing opportunities on cattle research studies/practical experiences.
I would acknowledge Dr. Kenneth Coffey, and Dr. Dan J. Donoghue for serving in my
PhD thesis committee and providing valuable suggestions.
My dear parents, Chhabilal Acharya and Sabitra Acharya, my grandfather Dhananjaya
Acharya, a graduate from Banaras Sanskrit University, about a century ago, my big brother
Kamal Acharya (PhD graduate from Norway), my small brother Dinesh Acharya (MSc engineer
graduate from Switzerland), my sisters Manju Acharya and Ranjana Acharya and my beautiful
wife Roshani Sharma Acharya (PhD candidate at University of Arkansas). My achievement
today would not have been possible without the help of USDA-ARS Booneville Folks, Chad
Lee, Erin Wood, Jackie Cherry, Connie Cox, Samuel Tabler, Hannah Toland, Caleb Crelia,
Berlin Howell, and Keren Chapman. Graduate students at the University of Arkansas,
Fayetteville, AR, Saleh A. Alrashedi, Amanda Davis, Fanli Kong, Angella Boyer, Michelle
Thomas, Elizabeth Backes, Brandon Smith, Tony Ryan, Tom Devine, Cristina Turner, Eva wary,
Xheena, Christine Weingartz-Hernandez. Secretaries and staffs at the department, Chris Hansen,
Toby, Linda, Robert Story, Jeannie, Dianna, Connie, Palika, Tes, and TC thank you all help.
I cannot and will not forget Dr. Charles F. Rosenkrans name in here for all his love.

Dedication
I want to dedicate this dissertation, PhD degree and my all course work to my research
advisor Dr. Joan M. Burke. You accepted me as your first Master student as well as first PhD
student. You have thousands of qualities to be a good advisor. Thank you for all the help you did
in this important and productive 5 years.

Table of Contents Page number
Chapter I: Introduction………………………………………………………………………….1
Chapter II: Review of Literature……………………………………………………………….3
a. Common extenders and storage temperatures for extension and cold storage of ram semen……………………………………………………………………………………...3
Early extension of bull semen…………………………………………………………......3
Common semen extenders used in different species……………………………………...3
Common buffers and bases used for storage of ram semen……………………………….4
Milk extender……………………………………………………………………………...6
Comparison of milk with other extender………………………………………………….8
Storage temperatures……………………………………..................................................10
Use of different percentage of egg yolk in the extender medium………………………..11
Low sodium choline based medium (CJ-2)……………………………………………...13
Ethylene glycol…………………………………………………………………………..14
Penicillamine, hypotaurine and epinephrine (PHE)……………………………………..16
b. Anti-Mullerian hormone as an endocrine marker to predict fertility………………18
Anti-Mullerian hormone…………………………………………………………………18
Sexual differentiation in males and females……………………………………………..18
Anti-Mullerian hormone and FSH……………………………………………………….21
Anti-Mullerian hormone studies in ewes………………………………………………...22
Summary and objectives…………………………………………………………………22
References…………………………………………………………………………..........24
Chapter III: Effect of semen extender and storage temperature on ram sperm motility
over time
Abstract………………………………………………………………………………….34

Introduction…………………………………………………………………………......35
Materials and Methods…………………………………………………………………36
Animal Management……………………………………………………………..36
Evaluation of fresh semen by computer assisted sperm analysis (CASA)………36
Preparation of semen extenders………………………………………………….37
Milk extender ……………………………………………………………37
TEST extender…………………………………………………………...37
TRIS extender……………………………………………………………38
CJ-2 extender ……………………………………………………………38
Preparation of semen for storage………………………………………………...38
Evaluation of extended semen using CASA …………………………………….39
Statistical analysis……………………………………………………………......40
Results…………………………………………………………………………………...40
Discussion……………………………………………………………………………….43
References……………………………………………………………………………….48
Tables
Table 1. CASA parameters………………………………………………………50
Table 2. Hourly change in temperature of semen filled straws kept inside
Koolatron cooler maintained at 15° C or laboratory refrigerator at 4° C………..51
Table 3. IVOS machine settings for ram semen motility analysis………………52
Table 4. Effect of temperature on ram sperm motility parameters over 72 h of
storage. Ram sperm motility parameters of extended and stored ram semen diluted
in CJ-2, milk, TEST, and TRIS extenders and stored at 4 or 15° C for 72 h…...53

Table 5. Effect of time of storage on ram sperm motility parameters, CJ-2, milk,
TEST or TRIS and stored at 4 and 15° C for 72 h……………………………….54
Chapter IV: Evaluation of semen extenders for short-term storage of ram semen at 4° C
Abstract………………………………………………………………………………….55
Introduction……………………………………………………………………………..56
Materials and Methods…………………………………………………………………57
Animal Management……………………………………………………………..57
Evaluation of fresh semen by computer assisted sperm analysis (CASA)………58
Experimental treatments…………………………………………………………58
Extenders………………………………………………………………………...59
Milk and TRIS extender…………………………………………………59
Determination of semen extension rate …………………………………………59
Semen extension using extenders ……………………………………………….59
Preparation of PHE ……………………………………………………………...60
Evaluation of extended semen …………………………………………………..61
Statistical analysis ……………………………………………………………….61
Results ………………………………………………………………………...………...62
Discussion……………………………………………………………………………….65
References…………………............................................................................................70
Tables
Table 1. IVOS machine settings for ram semen motility analysis ……………...72
Table 2. CASA parameters……………………………………………………....73
Table 3. Sperm motility parameters after extension using milk or TRIS and stored

at 4° C for 72 h. ………………………………………………………………….74
Table 4. Sperm motility parameters after extension using 5 or 20% (v/v) egg yolk
in milk or TRIS and stored at 4° C for 72 h. …………………………………….75
Table 5. Sperm motility parameters after extending with or without ethylene
glycol in milk or TRIS and stored at 4° C for 72 h. ……………………………. 76
Table 6. Effect of addition of penicillamine, hypotaurine, and epinephrine (PHE)
on sperm motility parameters before evaluation on cold stored ram semen, stored
at 4° C for 72 h.………………………………………………………..................77
Table 7. Sperm motility parameters after extension with or without ethylene
glycol in milk or TRIS with 5 or 20% egg yolk and stored at 4° C for 72 h. Only
significant ethylene glycol × hours interactions (P < 0.05) are expressed……....78
Table 8. Sperm motility parameters after extension using with or without
ethylene glycol (EG) in milk or TRIS extender. Only significant ethylene glycol ×
extender interactions (P < 0.05) are expressed…………………………………..79
Table 9. Sperm motility parameters after extension including 5 or 20% egg yolk
in milk or TRIS extender. Only significant egg yolk (EY) × extender interactions
(P < 0.05) are expressed…………………………………………………………80
Chapter V: Correlation between Anti-Mullerian hormone and estimated breeding value for
reproductive performance in ewes ……………………………………………………………81
Abstract………………...………………………………………………….……………81
Introduction….……………………….…………………………………….…………..82
Materials and Methods ………………………………….…………………………….83
Location …………………………………………………………………………83
Animal management …………………………………………………….………83

Blood sampling and serum collection …………………………………………...83
Anti-Mullerian hormone analysis ………………………………......…………...84
Description of animals and categories…………………………………………...85
Statistical analysis ……………………………………………………………….85
Results …………………………………...……………………………………………...85
Discussion ……………………………………………………...……………………….87
References ………………………...…………………………………………………....92
Table 1. Relationship between AMH quartiles and mean number of lambs born
from different summer and fall breeding. Data expressed in mean ± standard
error (SE)………………………………………………………………………...94
Chapter VI: Conclusions ………………………………… …………………………………...95
Appendix. University of Arkansas Animal Use Protocol (IACUC 15019)…….…...…….…97

1
Chapter I. Introduction
There are many advantages to using artificial insemination (AI) in sheep. In a
conventional breeding program, a ram can mate 50 to 100 ewes per year. However, through the
use of cervical AI, semen collected from a single ram can be used to inseminate over 1000 ewes
during 2 to 3 weeks (Evans and Maxwell, 1987). Faster genetic gain for desirable traits in the
flock is possible through the use of AI technique (Evans and Maxwell, 1987). Artificial
insemination is the cheapest means to spread superior genetic traits of elite rams across long
distances and beyond the limits of live animals (Foote et al., 2002). Artificial insemination
eliminates the need for small producers to keep a breeding ram in a herd and also enables them to
access semen from superior rams (Evans and Maxwell, 1987). For AI, semen quality is examined
on all rams before semen extension and storage, thus eliminating sub-fertile males from the flock
(Foote et al., 2002). When using AI, there is no direct contact between males and females, which
helps to minimize disease transmission during breeding (Evans and Maxwell, 1987). Semen can
be collected from superior rams which may not be able to breed naturally due to injury (Evans
and Maxwell, 1987). Ram-ewe preference during breeding is eliminated in AI (Evans and
Maxwell, 1987). Accurate record keeping is possible through AI, and thus, making it easier
to select desirable traits and minimize undesirable traits from the flock (Evans and Maxwell,
1987). During off-season (sheep are seasonal breeders), ram semen quality is low (Mickelsen et
al., 1981). Artificial insemination enables the collection of high quality semen during the
breeding season and use it to inseminate synchronized ewes during off-season (Evans and
Maxwell, 1987). It is possible to produce only ewe lambs through AI using sex sorted semen
(Foote et al., 2002).
Artificial insemination with frozen thawed semen results in low conception rates because
of poor survival of sperm (Salamon and Maxwell, 2000). A higher conception rate occurs when

2
extended semen is cool stored instead of cryopreserved. Cervical AI conception rate was
compared after extension and storage among various temperatures. Conception rate was higher
with extended semen stored at 4° C for 72 h (70%) or 23° C for 24 h (76%) compared with
cryopreservation (-196° C, 50%; Wusiman et al., 2012). Milk or tris[Hydroxymethyl]
aminomethane (TRIS) based extenders are the most common extenders used for liquid storage of
ram semen (Salamon and Maxwell, 2000). The TEST (N-Tris(hydroxymethyl) methyl-2-
aminoethane sulfonic acid, TES titrated with TRIS) was used in other species but not commonly
used for extension of ram semen (Bolanos et al., 1983; Pope et al., 1991). Very limited studies
are available on choline based (CJ-2) semen extender.
There was evidence of better motility of ram semen after storage at 5° C (O’ Hara et al.,
2010) while others reported better motility after storage at 15° C (Mata-Campuzano et al., 2014).
Conclusions cannot be generalized as those studies differed in types of extenders and storage
duration. The current study was designed to find best semen extenders, storage temperature, and
supplements for liquid storage of ram semen for 3 d.
Anti-Mullerian hormone (AMH) in circulation may reflect the total pool of follicles
within the ovaries, and may be positively correlated with fertility in ewes. An additional
objective of the study includes determining the correlation between serum concentrations of
AMH and EBV for NLB, NLW, MWWT, WWT, and the hair sheep index.

3
Chapter II: Review of literature
a. Common extenders and storage temperatures for extension and cold storage of ram
semen
Early extension of bull semen
Yolk-phosphate was the first most extensively-used semen extender for AI in the US
(Phillips and Lardy, 1940). Salisbury et al. (1941) reported superiority of sodium citrate extender
due to better buffering capacity, and maintaining of sperm motility for a longer period compared
with egg yolk-phosphate extender. During the early 1940’s, bull semen extended with sodium
citrate and stored at 5° C for up to 3 d was used for AI in cattle (Foot et al., 2002). A few years
later, the necessity of semen preservation for long periods developed, especially for dairy cattle;
therefore, a use of a glycerol based semen extender for cryopreservation was initiated (Polge et.
al., 1949). During the 1940’s and 1950’s, AI conception rate in cattle was very low (15%; Foote
et al., 2002). Several studies were conducted in the 1950’s to protect sperm cells from cold shock
and used antibiotics to control disease transmission in order to increase overall AI conception
rate in cattle (Foot and Bratton, 1950). Studies on milk glycerol semen extender for
cryopreservation of bull semen began in the 1950’s (Michajilov, 1950). Caproic acid and
catalase, with 5% egg yolk as antioxidants, were used to preserve bull sperm in New Zealand in
the early 1980’s (Shannon et al., 1984). Use of sugar for successful cryoprotection was initiated
by Polge in 1968 (Polge, 1968). The TRIS egg yolk extender for cryopreservation provided
excellent sperm protection for both cold and frozen storage (Davis et al., 1963), and thereafter,
this medium was used worldwide for semen storage of bull semen (Rehmana, et al., 2013).
Common semen extenders used in different species
Sodium potassium tartrate, sodium sulfate, and peptone were commonly used to extend
and store boar semen between 1920 - 1960 (Foote et al., 2002). After discovery of egg yolk-

4
citrate, and egg yolk-phosphate extenders in the bull, the same extenders were used in the boar
with little modification (Polge, 1956). Frozen storage of boar semen had little success; therefore,
fresh or liquid extended semen is commonly used (Foote et al., 2002).
The most common semen extenders used to extend buck and ram semen is described in
detail (Evans and Maxwell, 1987). The TRIS and citrate are common buffers, fructose or glucose
are used as energy sources, and egg yolk is added to the extender to protect sperm cell
membranes against cold shock (Evans and Maxwell, 1987). Egg yolk causes a negative
interaction with seminal plasma in buck semen; therefore, low egg yolk is preferred for
cryopreservation for goats (Evans and Maxwell, 1987). Sperm extension, extenders, storage
temperature, and AI techniques in sheep and goats has been extensively reviewed by Australian
scientists (Salamon, 1976; Salamon and Maxwell 1995; and Salamon and Maxwell 2000).
Common semen extenders used in poultry are a mixture of phosphates, citrates and/or N,
N-bis(2-hydroxyethyl)-2-aminoethane sulfonic acid (BES), and TES (Donoghue and Wisharttris,
2000). Turkey semen requires oxygen during storage but chicken sperm can survive both in an
aerobic and non-aerobic environment (Donoghue and Wishart, 2000). Artificial insemination in
poultry is mostly from fresh collection. Cryopreservation of poultry semen is limited (Gill et al.,
1999).
Common buffers and bases used for storage of ram semen
Seminal plasma maintains semen pH, but semen pH may fluctuate after extension (Purdy,
2006). In order to maintain sperm cell viability or avoid damage to cells in extended semen and
storage, pH needs to be constant (Purdy, 2006). Buffers are used to minimize pH fluctuation after
extension. Buffering solution for semen storage should have a) pKa between 6 and 8, b)
maximum water solubility, c) minimum passage through cell membranes, d) little interaction

5
with salts, e) dissociation of buffer that is minimally affected by buffer concentration,
temperature, and ionic concentration of medium, f) resists enzymatic and non-enzymatic
digestion, and g) should be prepared from inexpensive materials and easily purified (Good et al.,
1966; Graham et al., 1972). Milk, TRIS, TES, and choline-based extenders meet most of the
above criteria. Appropriate concentrations of these buffers in semen extenders for maintenance
of pH and best survival of sperm was examined. Concentration of TRIS (150 to 550 mM in 50
mM increments) was tested in a medium to compare best post thaw survival of frozen ram semen
(Salamon and Visser, 1972). Spermatozoa tolerated a wide range of TRIS concentrations (250 to
400 mM), although the highest post thaw motility was obtained using 300 mM TRIS.
Raffinose-based extender was the extender of choice before in vitro semen motility was
tested with TRIS based extenders. Visser and Salamon (1973) compared AI conception rate
following cervical AI with frozen-thawed ram semen extended in either TRIS (300 mM TRIS,
27.75 mM glucose, 15% v/v egg yolk, and 5% v/v glycerol) or raffinose (166.5 mM raffinose, 68
mM sodium citrate, 15% v/v egg yolk, and 5% v/v glycerol) based extender. Lambing rates were
similar (40.0 versus 44.8%) for TRIS and raffinose-citrate extender, respectively, but freeze-
thaw sperm motility of frozen ram semen pellets without using thawing solution (pelleted
raffinose in inositol-sodium-citrate solution and pelleted TRIS in TRIS-fructose solution) was
lower in raffinose-citrate extenders compared with TRIS based extender. Thus, TRIS based
extender was recommended by Salamon and Visser (1972) as the extender of choice. A
concentration of 300 mM is still used as a concentration of choice for liquid storage of ram
semen in TRIS extender (Quan et al., 2016).
Studies have also been conducted to determine the best buffer and base combinations for
pH maintenance and sperm cell survival after freeze-thaw of extended semen. Seven different

6
zwitter ion buffers, 3-(Morpholin-4-ium-4-yl)-1-propanesulfonate (MES), 2,2’-(1,4-
Piperazinediyl)diethanesulfonic acid (PIPES), 3-(Morpholin-4-ium-4-yl)-1-propanesulfonate
(MOPS); 2-[Bis(2-hydroxyethyl)amino]-2-(hydroxymethyl)-1,3-propanediol (BES), and N-
Tris(hydroxymethyl) methyl-2-aminoethane sulfonic acid (TES), 2-[4-Hydroxyethyl)-1-
piperazinyl]ethanesulfonic acid (HEPES), and N-[1,3-Dihydroxy-2-(hydroxymethyl)-2-
propanyl]glycine (TRICINE) were used to titrate separately with bases (TRIS, NaOH, and
combination of NaOH and KOH) to pH 7.0 for extension and cryo-storage of bovine semen in a
glycerol-egg yolk medium (Graham et al., 1972). The TES titrated with TRIS better maintained
pH than other buffer-base combinations and maintained semen motility after freezing and
thawing of bovine semen (Graham et al., 1972). Comparison of TRIS (300 mM TRIS, 27.8 mM
glucose, 94.7 mM citric acid, 150 mg/l penicillin, 250 mg/l streptomycin, 0.3% bovine serum
albumin, and 10% (v/v) egg yolk) and TES-TRIS (176 mM TES, 80 mM TRIS, 9 mM glucose,
10 µg/ml gentamicin sulphate, and 10% egg yolk) was done for extension and cold storage (4º C)
of ram semen for 7 d (Quan et al., 2016). However, there were no significant differences in
sperm motility and viability parameters between TRIS and TES-TRIS based extenders. Since
TES is expensive and its inclusion in TRIS based buffer has no advantage for liquid stored ram
semen, its inclusion in TRIS based extender was not justified.
Milk extender
Milk contains lipids, lactose, casein, albumin, globulin, lactoferrins, and several other
compounds. Heating milk to 95° C for 10 minute inactivates lactenin in protein, a compound
toxic to sperm cells (Salamon and Maxwell, 2000). Several studies were done in the past to find
component(s) in milk that protect sperm cells after extension and storage of semen. No
difference in motility or field fertility was found when bovine semen was extended and stored in

7
skim milk (lipid free) or whole milk, suggesting the lipid fraction in milk is not a major factor
that contributes to protection of sperm cells (Almquist et al., 1954). Semen extended in lactose
(not fructose) based extender had higher frozen thaw motility when used with the dialyzable
portion of milk. However, fructose based extender (144 mM fructose, 3% v/v casein, 10 mM K,
2 mM Mg, 0.5 mM Ca, 20 mM mono and disodium phosphate) had higher sperm motility
compared with lactose (equal concentration of fructose replaced by lactose) based extender in
synthetic media (Choong and Wales, 1963). Addition of extra fructose to a 144 mM initial
concentration improved frozen thawed bovine semen motility; thus lactose can be replaced with
fructose without any deleterious effect (Choong and Wales, 1963).
Casein proteins are found in milk; making up to 80% of the total protein in cow and goat
milk. Casein exists in milk as casein micelles (Dalgleish, 1998). The effect of albumin, globulin,
or casein (all 3% w/v) on motility of cold stored bovine semen was compared by adding these
proteins in phosphate buffer solution (mono- and disodium phosphates and 22 mM fructose).
Motility measured after storage of extended bull semen at 5° C for 24 h showed increased
motility in casein but not in albumin or globulin (Choong and Wales, 1963). Based on their
research it was concluded that casein (not albumin or globulin) could help improve semen
motility after cold storage of bovine semen. The mechanism by which casein protein protects
sperm after extension was reviewed by Manjunath et al. (2002). The binder of sperm (BSP)
proteins are present in seminal plasma which deteriorates sperm cells after extension (Manjunath
et al., 2002). In ram semen, casein micelles bound to BSP proteins to decrease cholesterol and
phospholipid loss from the sperm membrane, thus protecting sperm cells after extension (Plante
et al., 2015). Therefore, it could be that casein protein in milk plays a major role in protection of
sperm cells when semen is extended and stored in milk based extenders.

8
Comparison of milk with other extender
Research was conducted TRIS and milk based extenders for better maintenance of ram
sperm cell motility after extension and storage. Motility of ram sperm was higher when extended
and stored at 5 or 20° C for 30 h in a commercial TRIS extender (Minitub, Tiefenbach,
Germany) with 20% v/v egg yolk or TRIS citrate (250 mM TRIS, 69.4 mM fructose, 88.5 g
citric acid, 20% v/v egg yolk) compared with milk (11% non-fat milk powder, 5% v/v egg yolk)
or sodium citrate (112 mM % trisodium citrate, 20% v/v egg yolk) based extenders (Paulenz et
al., 2002). Superiority of TRIS over milk based extender was also reported by Quan et al. (2016)
when milk (10 g skim milk powder, 0.2 g glucose in 80 ml distilled water, and 0.5 g of penicillin
and streptomycin), was compared with TRIS (300 mM TRIS, 27.8 mM glucose, 94.7 mM citric
acid, 150 mg/l penicillin, 250 mg/l streptomycin, 0.3% bovine serum albumin, and 10% (v/v)
egg yolk), and TES-TRIS (176 mM TES, 80 mM TRIS, 9 mM glucose, 10 µg/ml gentamicin
sulphate, and 10% egg yolk) extenders for extension and storage of ram semen at 4º C for 7 d.
Total motility, membrane integrity, and acrosomal status was better when extended and stored
TRIS or TES-TRIS compared with milk extender (Quan et al., 2016). In another study, TRIS
(300 mM TRIS, 27.7 mM fructose, 103 mM citric acid, and 15% v/v egg yolk was dissolved in
100 ml distilled water), sodium citrate (2.9% aqueous solution of trisodium citrate in 20% v/v
egg yolk), milk (11% w/v distilled water with 5% v/v egg yolk), glucose phosphate (108 mM
Na2HPO4, 24 mM KH2PO4, 34 mM glucose, and 2% v/v egg yolk), and commercial proprietary
extender Andromed® (Minitub, Tiefenbach, Germany) were examined for extension and storage
of ram semen at 4º C until no motile spermatozoa were observed. No difference in sperm
motility was observed until the third day between TRIS, sodium citrate and milk based
extenders, but sperm motility was higher than that extended in glucose phosphate, and

9
Andromed® extenders (Gündoğan, 2009). For longer storage (up to 14 d), semen extended in
TRIS maintained better sperm motility compared with milk and other extenders (Gündoğan,
2009). López et al. (1999) compared sodium citrate (112 mM aqueous solution of tri-sodium
citrate mixed in 20% of egg yolk), TRIS (250 mM Tris, 71 mM fructose, 88 mM g citric acid per
1000 ml extender plus 20% egg yolk), or skim milk extender (88 g non-fatty milk powder
dissolved in 800 ml distilled water, 220 ml of mixed solution was dissolved with 12 ml of egg
yolk) for extension and storage of ram semen at 5º C for 7 d. There was no difference in sperm
motility until 24 h of storage, but after 24 h, milk based extender had lower sperm motility
compared with TRIS or sodium citrate based extender (López et al., 1999).
Not all studies reported superiority of TRIS over milk based extender for preservation of
cool stored ram semen. Hollinshead et al. (2004) compared ultra-heat treated milk, TRIS with
10% v/v egg yolk, a proprietary AndroHep® (Minitub, GmbH, Landshut, Germany), and TES-
TRIS 10% v/v egg yolk. Semen stored in ultra-heat treated milk had higher motility, acrosomal
integrity, and greater migration through artificial mucus and at on all times compared with other
extenders (TRIS, AndroHep®, and TES-TRIS; Hollinshead et al., 2004).
Superiority of milk as an extender was also reflected in in vivo studies. There was a
higher vaginal insemination conception rate obtained with semen extended and stored in milk
compared with TRIS based extenders (Paulenz et al., 2003). It could be possible that short term
(< 30 h) storage of semen in milk based extenders will have a higher motility and AI conception
rate compared with TRIS based extenders, and TRIS based extenders may be beneficial for long
term storage (> 24 h). Nevertheless, this is hard to generalize as milk based extenders had low
motility when stored for less than 30 and 24 h in the study reported by Paulenz et al. (2002) and
López et al. (1999), respectively.

10
Storage temperatures
Ram semen extended in sodium citrate (124 mM sodium citrate, 177 mM glucose, 20%
v/v egg yolk, with 6% v/v glycerol added in cryopreserved medium) was stored at 4, 23 or -196°
C (Wusiman et al., 2012). Cervical insemination of ewes with semen stored at 4° C for 72 h
resulted in a similar conception rate compared with semen stored at 23° C for 24 h (69.6% vs
76%), but both were higher than the 50% conception rate for semen cryopreserved and stored at -
196° C (Wusiman et al., 2012). Ram semen quality characteristics were also compared using
milk extenders for storage at 4 or -196° C for 48 h and 4 wk, respectively (Kulaksiz, et al.,
2012). Total motility of sperm cells in milk extender after storage at 4° C for 48 h had a higher
than compared with frozen-thawed ram semen stored at -196° C for 4 wk (47.5 versus 21.2%;
Kulaksiz, et al., 2012). From the two studies above, it can be concluded that ram semen motility
can be better maintained and have a higher AI conception when stored at a low temperature (4°
C) for a short period of time (2 to 3 d) compared with freeze-storage at -196° C.
For cool storage of extended ram semen, different low temperatures have been compared.
Ram semen was extended using milk, TRIS, commercial TRIS or sodium citrate and stored at 5
or 20° C for 30 h (Paulenz et al., 2002). Sperm motility was higher when stored at 5 compared
with 20° C (Paulenz et al., 2002). However, Salamon and Maxwell (2000) stated that 5 or 15° C
was the most suitable temperatures for liquid storage of extended ram semen. To find the best
temperature between 5 or 15° C, ram semen was extended using standard skim milk-egg yolk,
OviPro®, AndroMed®, or INRA 96® for storage at 5 or 15° C. Sperm motility score (0 = non-
motile to 5 = rapid motility) of extended and stored semen at 5° C for 72 h ranged from 2.3 for
skim milk extender, to 3.2 for AndroMed®, whereas those stored at 15° C for a similar time
period ranged from 0 for skim milk based extender to 1.16 for AndroMed® (O’Hara, et al., 2010).

11
In this study, ram semen extended and stored at 5° C had higher motility than that storaged at 15°
C when evaluated at 72 h of storage (O’Hara, et al., 2010). However, the results in this study
were not consistent with other studies in which ram semen was evaluated at 48 h after storage at
5 or 15° C (Mata-Campuzano et al., 2014) using TES-TRIS or commercial INRA96 extenders. It
could be possible that for short time storage (< 48 h) 15° C could be better than 5° C storage.
Consistent with better motility when stored for < 48 h, storage at 15 was better than 5° C with
semen extended in ultra-heat treated milk. Another study compared TRIS 10% v/v egg yolk,
AndroHep®, and TES-TRIS 10% v/v egg yolk (Hollinshead et al., 2004). Spermatozoa stored at
15° C for 6 h migrated a greater distance in artificial cervical mucus than spermatozoa stored at
5° C for 3 d (Hollinshead et al., 2004). Even though semen storage at 5° C was better than at 15°
C for 3 d (O’Hara, et al., 2010), shorter storage (6 h or 48 h) at 15 °C was better than at 5° C.
However, it is hard to make conclusion from these studies, as they differed in type of extenders,
use of supplements, and storage periods.
Use of different percentages of egg yolk in the extender medium
Sperm plasma membrane protects sperm cells from its external environment. Cooling
semen during freezing or cold storage causes massive rearrangements and leakage in sperm
plasma membrane leading to permanent damage to the sperm cell (Holt and North, 1984).
Studies were conducted to find the component of egg yolk and its protective action on sperm
cells during storage of extended semen. Studies between the 1970’s and early 1980’s concluded
that low density lipoprotein in egg yolk protects sperm cells during freezing by making a
spermatozoa-lipoprotein complex (Pace and Graham, 1974; MacDonald and Foulkes, 1981).
Ricker et al. (2006) used purified phosphatidylcholine (PC; soy-PC, or egg-PC) to replace
egg yolk in INRA 82, a commercial semen extender to freeze donkey semen. Total motility,

12
progressive motility, and viability of frozen-thawed semen showed that egg yolk can be replaced
with soy or egg-PC for frozen storage of semen without affecting semen quality. Inclusion of
external lipids (phosphatidylcholine in this case) does not incorporate in sperm membrane but
associates strongly, and this strong attachment remains even after the freeze-thaw process
(Ricker et al., 2006). Thus, low density lipoprotein can be replaced with phosphatidylcholine
without negative effects to maintain sperm motility and viability (Ricker et al., 2006). Thus, the
spermatozoa-lipoprotein complex formation may not be the only mechanism for sperm cell
protection. Purification of phosphatidylcholine either from soy or egg yolk is a very expensive
process, making it less practical.
According to a third hypothesis, bovine seminal plasma contains BSP proteins which are
secreted by seminal vesicles during ejaculation (Manjunath et al., 1988; Desnoyers and
Manjunath, 1992). The BSP proteins modify the sperm membrane and cause cholesterol efflux,
and continuous exposure leads to detrimental effects on the sperm plasma membrane (Manjunath
et al., 2002). When semen extender is supplemented with egg yolk, the low-density fraction
(LDF) of egg yolk binds BSP protein preventing detrimental effect on sperm membranes
(Manjunath et al., 2002).
Ram semen supplemented with egg yolk, or combination of egg yolk and glycerol had
higher sperm subjective motility and membrane integrity compared with no egg yolk (Gil et al.,
2011). Extender supplemented with glycerol but without egg yolk had no protective effect on
cool storage of ram semen (Gil et al., 2011). This shows the importance of egg yolk to protect
extended, cooled sperm cells.
López et al. (1999) used three different extenders: sodium citrate (112 mM aqueous
solution of tri-sodium citrate) with 20% of egg yolk, TRIS with 20% egg yolk, or skim milk with

13
5.5% egg yolk to store ram semen at 5 °C for 7 d. Inclusion of lower percentage of egg yolk in
the milk based extender led to decreased motility, but this was confounded by the difference in
composition of extenders. The importance of increasing egg yolk concentration on resilience of
spermatozoa has been demonstrated. When cooled slowly, concentration of egg yolk (i.e., 10, 15,
and 20% of total volume) in TRIS citrate extender had no effect on motility. However, viability
of ram semen extended in 20% egg yolk was higher compared with 10% egg yolk. When cooled
rapidly or cold shocked, the higher percentage of egg yolk increased motility (Rajabi-Toustani et
al., 2014).
Low density lipoprotein is a fraction of egg yolk that binds bovine seminal plasma (BSP)
protein that prevents detrimental effects on sperm membrane (Manjunath et al., 2002). Inclusion
of 8% LDL extracted from egg yolk improved protection of sperm cells over inclusion of egg
yolk in cryo stored citrate extender (Bashawat et al., 2016). Semen stored in extender containing
8% LDL had the highest freeze-thaw sperm motility (62.66%) compared with 20% egg yolk
(54.18%), or 6 and 10% LDL supplementation (Bashawat et al, 2016). Similarly, 5 and 8% LDL
in TRIS based extender increased forward progressive motility of sperm cells that had been
thawed compared with 3% LDL or egg yolk alone (Gharibi et al., 2014). Progressive forward
motility of sperm cells extended with inclusion of 12% egg yolk or 5% LDL was similar up to 48
h, but was higher after 48 h for semen extended with 5% LDL compared with 12% egg yolk.
Thus, LDL used in an extender medium maintains sperm motility better than using egg yolk for
cooled or frozen storage of ram semen. However, LDL are not commercially available.
Low sodium choline-based medium (CJ2)
Under normal physiological conditions any excess sodium that enters the cell is expelled
by a sodium pump. When oocytes/spermatozoa are slowly frozen in cryoprotectants, water

14
comes out of the cell and sodium diffuses in. The sodium pump is gradually disabled with the
decrease in temperature leading to sodium accumulation inside the cell (Mazur, 1977).
Intracellular sodium accumulation damages cellular integrity and further development of oocytes
(Mazur, 1977; Stachecki and Willadsen, 1998a). It was discovered that choline could be used in
place of sodium as a major cation for cryopreservation of mouse oocytes. Oocytes frozen in
choline based extender had better survival, fertilization, and further development compared with
sodium based extender (Stachecki and Willadsen, 1998b). Similarly, replacing sodium with
choline in a medium for cryopreservation of metaphase II mouse oocytes caused lower pre-
freezing damage (Stachecki et al., 1998b).
Reasonably, choline is a component of the phospholipid bilayer and, therefore, could help
to stabilize cell plasma membrane for better protection of spermatozoa and oocytes. Very limited
studies are available in which choline based (CJ-2) semen extenders were used for extension and
cool storage of semen. Sperm motility parameters were evaluated after extension and cool
storage of bull semen using either caudal epididymal plasma or low sodium medium
supplemented with either a lipid-rich bovine serum albumin or egg yolk. Bull semen extended
and stored at 4° C using CJ-2 egg yolk had increased sperm motility parameters compared with
other semen extenders used (Delgado et al., 2009). Choline based extenders are not used for cool
storage of ram semen. It would be worthwhile to see if beneficial effects of choline based
extenders can be seen after cold storage of ram semen.
Ethylene glycol
Ethylene glycol is a cryoprotectant of choice for freezing embryos because of its low
molecular weight, which enables rapid influx into cells during equilibration and rapid efflux after
thawing (Bautista et al., 1997). Similar effects of ethylene glycol occurs across the plasma
membrane of sperm cells (Hammerstedt and Graham, 1992). Osmotic stress (difference in

15
relative permeability of cryoprotectants and water) is a major factor for spermatozoa damage
during semen cryopreservation (Gao et al., 1997). Sperm cells are sensitive to osmotic stress
during addition or removal of cryoprotectants and this sensitivity changes with the concentration
of cryoprotectants (Gao et al., 1997). Ball et al. (2001) used four different cryoprotectants (1 M;
glycerol, ethylene glycol, dimethylsulfoxide, or propylene glycol) on tyrode albumin lactate-
pyruvate medium (TALP) to evaluate the effect of addition and removal of these cryoprotectants
on osmotic stress after liquid storage of equine semen for 5 min at 22° C. Based on sperm
motility and viability, semen stored in ethylene glycol based TALP medium had the least
osmotic stress compared with other cryoprotectants (Ball et al., 2001).
The protective effect of cryoprotectants at various concentrations (glycerol 5%, ethylene
glycol 3% and 5%, and acetamide 3% and 5%; TRIS fructose media) on ram sperm cells was
measured after extension and freeze storage (Silva et al., 2012). No difference was found in
progressive motility, acrosomal integrity, and mitochondrial membrane integrity between semen
samples stored in ethylene glycol (3 or 5%) compared with 5% glycerol based medium.
However, those parameters were higher than samples extended and freeze-stored in 3 or 5%
acetamide based medium (Silva et al., 2012). In a similar study, cryoprotectants (glycerol 7%,
ethylene glycol 3, 5, or 7%, and DMSO 3, 5 or 7%) in TRIS based medium were examined
to extend and freeze-store ram semen. Semen stored in ethylene glycol (3 or 5%) had similar
progressive motility, viability, and membrane integrity compared with semen cryopreserved in
7% glycerol. The protective effect of these cryoprotectants was better than dimethyl sulfoxide
DMSO (Najafi et al., 2016). Both of the above studies were done in frozen extended semen, and
ethylene glycol or glycerol were found to be better cryoprotectants compared with DMSO or

16
acetamide for cryo storage of ram semen. Ethylene glycol at 3 or 5% can be used for
cryopreservation in a medium for freeze storage of ram semen.
Use of cryoprotectants for cool storage of extended semen were also tested at various
concentrations. Five cryoprotectants (dimethylacetamide, dimethyl sulfoxide, ethylene glycol,
glycerol or propylene glycol) at 2.5, 5 or 10% final concentration were added in TES-TRIS
fructose extender to extend and store ram semen at 4° C for 3 h. Semen stored in medium
containing 2.5% ethylene glycol had higher sperm progressive motility compared with other
cryoprotectants and concentrations used (Vásquez et al., 2013). Similarly, Carrascosa et al.
(2001) used 1.6% final concentration of either glycerol or ethylene glycol for storage of
Chinchila lanigera semen in TES-TRIS sodium citrate fructose medium at 4° C for up to 72 h.
Semen stored in ethylene glycol based medium had higher sperm motility, sperm viability, better
membrane and acrosomal integrity compared with semen stored in glycerol based medium.
Studies on use of ethylene glycol based medium for cold storage of ram semen are rare,
therefore, it would be interesting to see if a beneficial effect can be seen.
Penicillamine, Hypotaurine and Epinephrine (PHE)
Catecholamines (epinephrine), β-amino acid (hypotaurine), and serum albumin stimulate
sperm motility, viability, sperm capacitation, and acrosomal reaction (Bavister et al., 1979;
Meizel et al., 1980; Bavister, 1981). Penicillamine, hypotaurine, and epinephrine have different
modes of action but collectively enhance sperm motility, sperm capacitation, and acrosomal
reaction (Meizel, 1985). Penicillamine is a metal chelator and thus stimulates capacitation and
acrosomal reaction (Meizel, 1985). High K+ concentration could inhibit in vitro acrosomal
reaction, and use of hypotaurine partially reduces Na+, K+-ATPase activity, lowers intracellular
K+ and enhances in vitro fertilization (Meizel, 1985). Epinephrine activates α1-adrenergic

17
receptors and stimulates acrosomal reaction (Meizel, 1985). Leibfried and Bavister (1982)
conducted a series of experiments to determine the effects of epinephrine and hypotaurine on in
vitro fertilization of golden hamsters. Addition of 10 µM of hypotaurine and 1 µM of
epinephrine at the start of incubation in a culture media increased ova penetration by sperm
(71.7%) compared with using hypotaurine alone (6.6%). Addition of hypotaurine is necessary
prior to epinephrine to enable epinephrine to activate spermatozoa for ova penetration (Leibfried
and Bavister, 1982).
Supplementation of the combination penicillamine (10 µl), hypotaurine (1µl),
epinephrine (20 µl; PHE) compared with sodium metabisulfite (2 µM) or individual components
increased bovine sperm penetration when incubated with oocytes. However, the difference was
not seen after 14 h of incubation. It can be concluded that supplementation of PHE decreases
time interval for oocyte penetration by sperm, compared with individually using penicillamine,
hypotaurine or epinephrine (Susko-Parrish, et al., 1990). Miller et al. (1994) compared the effect
of addition of PHE, bovine oviduct epithelial cells (BOEC), or a combination of PHE + BOEC in
in vitro fertilization medium. The PHE (20 µM penicillamine, 10 µM hypotaurine, and 2 µM
epinephrine) was added to TALP fertilization medium. Addition of either PHE (62%) or BOEC
(57%) or the combination (PHE + BOEC) resulted in higher overall cleavage rate and more
embryos compared with no PHE and/or BOEC (Miller et al., 1994).
A study investigated the effect of PHE on sperm cell motility parameters after frozen-
thaw bovine semen. Pearson et al. (2007) compared computer assisted sperm analysis (CASA)
parameters of frozen thawed bovine semen cultured in sperm-TL medium with/without PHE.
Semen was incubated at 39° C and was evaluated every 2 h for 8 h. Semen stored in PHE
supplemented medium had higher progressive motility (PHE, 43.2 vs control, 35.7), path

18
velocity (PHE, 111.5 vs control, 99.4), progressive velocity (PHE, 100.0 vs control, 89.0), and
track speed (PHE, 166.3 vs control, 147.5) compared with no PHE (Pearson et al., 2007). All
these combined studies conclude that supplementation of PHE enhances sperm motility. To our
knowledge, no such studies have been conducted with PHE on cool stored ram semen.
b. Anti-Mullerian hormone as an endocrine marker to predict fertility.
Anti-Mullerian hormone
Anti-Mullerian hormone (AMH; also, known as mullerian inhibiting substance) is a
member of the transforming growth factor-β (TGF-β) family. It is a homodimeric disulfide-
linked glycoprotein, with a molecular weight of 140 kDa (Cate et al., 1986; A La Marca and
Volpe, 2006). Gene coding for AMH resides on short arm of human chromosome 19 (Cohen-
Haguenauer et al., 1987). In males, AMH is produced by sertoli cells as early as during testicular
differentiation (8-9 weeks of gestation in humans), whereas in females the hormone is secreted
by ovarian granulosa cells during late fetal life (36 – 38 weeks in human; Rajpert-De Meyts et
al., 1999).
Sexual differentiation in males and females
During early embryonic development, the genital ridge which arises from mesonephric
cells is bipotent (Wilson and Davies, 2007). This genital ridge develops into testes if a Y
chromosome is present. For this, transcription of Y-specific genes occurs resulting in production
of testes determining factor (Austin and Edwards, 1981). Testis-determining factor is responsible
for initiation of male sex differentiation in human. Absence of Y chromosome gives rise to an
ovary (Austin and Edwards, 1981). In humans, sex cords (Mullerian and Wolffian ducts) are
present within mesonephron, and remain similar in both males and females until seventh week of
gestation. In females, the wolffian duct regresses and the Mullerian duct forms the female

19
reproductive tract. Formation of the female reproductive tract does not depend on any ovarian
products. If genital ridges are removed at early embryonic development (before they develop into
testes or ovaries), a mammal develops into a female regardless of sex chromosomes carried, and
hence, development of female reproductive organs follows a default pathway (Alberts et al.,
2002). In males, two processes occur. First, Leydig cells produce testosterone which maintains
and promotes Wolffian ducts. The Wolffian duct differentiates into male tract and accessory
organs (vas deferens, seminal vesicles, and epididymis). Secondly, sertoli cells in testes secrete
AMH hormone which regresses mullerian duct (Munsterberg and Lovell-Badge, 1991).
In females, granulosa cells produce AMH (Vigier et al., 1984). Production of AMH in the
female fetus starts at third trimester (120 d gestation in sheep; Bezard et al., 1987). Ovarian
AMH hormone messenger RNA (mRNA) is expressed in granulosa cells from preantral and
small antral follicles. Corpora lutea, large antral follicles, oocytes, and primordial follicles lack
AMH hormone expression (Baarends et al., 1995). Weenen et al. (2004) studied the AMH
hormone expression in granulosa cells of human ovaries. Anti-Mullerian hormone protein was
observed in granulosa cells of primary follicles and continued until antral stage follicle (6 – 8
mm). The highest level of AMH was expressed in granulosa cells of preantral and small antral
follicles of less than 4 mm size. Scheetz (2010) analyzed bovine serum AMH concentration of
the ovulatory follicular wave (daily; 6 - 8 d prior to ovulation to the day of ovulation) and found
no difference in an individual during the last 6 - 8 d before ovulation. Therefore, it was
concluded that AMH hormone of an individual remains unchanged during an estrous cycle
(Scheetz, 2010). Anti-Mullerian hormone is highly variable between animals, but remains
unchanged for each animal when measured in 3 month intervals, indicating that this hormone
concentration remains constant in an individual animal over a long period of time (Rico et al.,

20
2009). Minimum variation of AMH occurred during the estrous cycle in 4 to 9 yr old Holstein
dairy cows, an indication of minimum variation with age during the reproductive life span
(Ireland et al., 2010).
About 1 million oocytes are present at birth, and that number decreases to less than half
once girls reach pubertal stage (Visser et al., 2006). With each follicular wave, follicles leave the
pool of primordial follicles and enter into the pool of growing follicles (Visser et al., 2006). After
puberty, a few of these growing follicles are rescued by follicle stimulating hormone (FSH),
while a majority will be lost as a result of atresia (Visser et al., 2006). In uniparous animals,
under the influence of luteinizing hormone (LH), only one among a few FSH-rescued follicles
become a dominant follicle and finally ovulate (Visser et al., 2006). Follicular reserve on the
ovary can be predicted by antral follicle count using an ultrasound (Visser et al., 2006). This
process involves transvaginal ultrasound examination. Transvaginal ultrasound examination for
antral follicle count requires technical expertise and is a time consuming process. Antral follicle
count is positively correlated with circulating AMH and number of morphologically healthy
oocytes in cattle (Ireland et al., 2008). High antral follicle count (> 25 follicles of more than 3
mm size) had 5 times greater number of morphologically healthy oocytes compared with low
antral follicle count (< 15 follicles; Ireland et al., 2008). Assessment of serum AMH
concentrations can be measured to predict follicular reserve in ovary. Positive correlations were
reported between AMH and first service pregnancy rates in dairy cows (Baruselli et al., 2015).
Additionally, dairy cows with lower AMH levels had a higher pregnancy loss between first and
second month of gestation (Baruselli et al., 2015). A positive correlation was also reported
between AMH and pregnancy rate in cows bred at estrus, leading to reinsemination and/or
natural service (Baruselli et al., 2015). In a recent study conducted in our lab, a positive

21
correlation between beef heifer AMH concentrations at first breeding and AI conception rate was
detected, and more cows were likely to conceive earlier in the breeding season when AMH
concentrations were higher (Rorie et al., 2017). In the same study, there was no correlation
between AMH concentrations measured at weaning and AI conception rate at first breeding,
overall pregnancy rate, and estrous response after synchronization (Rorie et al., 2017). Therefore,
based on this study we can conclude that the earliest age to measure AMH concentrations to
predict beef heifer fertility will be at breeding age.
Anti-Mullerian hormone and FSH
Anti-Mullerian hormone limits the number of primordial follicles entering the pool of
growing follicles (Durlinger et al., 2001). Growth of preantral and small antral follicles are FSH
dependent, and presence of AMH decreases FSH responsiveness of the follicle (Durlinger et al.,
2001). A study conducted with mice revealed that AMH deficient mice had normal reproductive
performance initially. However, primordial follicles were depleted at a faster rate compared with
normal mice (Durlinger et al., 2001). It is likely that the group of females with low AMH would
deplete primodial follicles early, resulting in early cessation of ovulation, and a shorter
reproductive life compared with animals with normal AMH level (Durlinger et al., 2001)
Anti-Mullerian hormone studies in ewes
In ewes anti-Mullerian hormone is high during the fetal stage compared with post-natal
concentrations (Bezard et al., 1987). Similarly, plasma AMH concentrations at 40 d of age in
ewe lambs is high compared with 3.6-month old ewes (Lahoz et al., 2012; Torres-Rovira et al.,
2014). Lahoz found vast differences in AMH concentrations between 1 and 5-month-old ewe
lambs, high at 1-month and lower at 5 mo of age (Lahoz, et al., 2016). Ovarian follicular
population, an indicator of fertility, is positively correlated with plasma AMH concentrations in

22
young lambs (Lahoz et al., 2016b). A positive correlation occurred between plasma AMH
concentration in 3.6-month ewe lambs and number of ovulations in response to equine chorionic
gonadotropin (eCG; Lahoz et al., 2012). In Rasa Aragonesa ewe lambs, a plasma AMH
concentration of 97 pg/ml measured at 3.6-months of age was determined as a cutoff value to
predict fertility at first mating (Lahoz et al., 2012). Plasma AMH concentrations higher than 97
pg/ml in prepubertal ewes had higher first mating conception rate compared with lower plasma
AMH concentration (50% vs 15%; Lahoz et al., 2012). High plasma AMH concentrations at 3.5-
months of age (97 pg/ml) were associated with a higher number of total lambs born per ewe until
3.5-years of age compared with ewes with lower AMH concentrations (97 ng/ml; Lahoz et al.,
2016a).
Summary and objective
Frozen storage and AI of ram semen results in low conception rates (Salamon and
Maxwell, 1995). Recommendations are mixed on storage temperature of cooled semen at 5º C or
15º C (O’ Hara et al., 2010; Mata-Campuzano et al., 2014). Differences may be dependent on the
supplements and storage duration. Addition of egg yolk and cryoprotectant prior to cooling, or
supplementation with penicillamine, hypotaurine, and epinephrine (PHE) before evaluation may
maintain or improve semen motility.
i) Therefore, the first objective of the current study was to compare four extenders
(milk, TRIS, TEST or CJ-2), and two temperatures (4 or 15° C) for extension and
storage of ram semen for up to 72 h using an equal amount of egg yolk, fructose, and
gentamicin.

23
ii) Second, to evaluate the effect of combinations of extenders (best two extenders from
objective i), ± ethylene glycol, egg yolk (5 or 20%), and ± PHE (penicillamine,
hypotaurine and epinephrine) on ram sperm motility parameters.
Ewes vary in their reproductive performance. Farmers can maximize their profit if fertile
ewes can be selected during early stages by using endocrine markers. Anti-Mullerian hormone is
produced by granulosa cells of preantral and small antral follicles of 1-3 mm in diameter (Bezard
et al., 1988; Monniaux et al. 2012). Ewe fertility at first mating was positively correlated with
plasma AMH concentrations measured at 3.6 months of age (Lahoz Crespo et al. 2012). High
plasma AMH concentrations at 3.5-month of age (> 97 pg/ml) was associated with a higher
number of lambs born per ewe as adults compared with ewes with lower AMH concentrations (<
97 pg/ml; Lahoz et al., 2016). The current experiment further explores how serum AMH
concentrations are correlated with estimated breeding values (EBV) such as number of lambs
born (NLB), number of lambs weaned (NLW), maternal weaning weight (MWWT), weaning
weight (WWT), and a USA hair index calculated by Sheep Genetics (Armidale, NSW, Australia)
through the National Sheep Improvement Program. Therefore, the objective of the study was to
determine correlations between serum concentrations of AMH and EBV for NLB, NLW,
MWWT, WWT, and the hair sheep index.

24
References
Alberts, B., A. Johnson, and J. Lewis. 2002. Primordial Germ Cells and Sex Determination in
Mammals. In Molecular Biology of the Cells. 4th edition. New York: Garland Science.
Almquist, J., R. J. Flipse, D. L. Thacker. 1954. Diluters for bovine semen. IV. Fertility of bovine
spermatozoa in heated homogenized milk and skim milk. J. Dairy Sci. 37:1303-1307.
Anel, L., M. Alvarez, F. Martinez-Pastor, V. Garcia-Macias, E. Anel, and P. de Paz. 2006.
Improvement strategies in ovine artificial insemination. Reprod. Dom. Anim. 41:30-42.
Anel, L., M. Kaabi, B. Abroug, M. Alvarez, E. Anel, J. C. Boixo, L. F. de la Fuente, P. de Paz.
2005. Factors influencing the success of vaginal and laparascopic artificial insemination
in Churra ewes: A field assay. Theriogenology. 63:1235-1247.
Austin, C. R., and R. G. Edwards (eds.). 1981. Mechanisms of sex differentiation in animals and
man. Academic Press, N. Y.
Baarends, W. M., J. Uilenbroek, P. Kramer, J. W. Hoogerbrugge, E. Van Leeuwen, A. P.
Themmen, J. A. Grootegoed. 1995. Anti-müllerian hormone and anti-müllerian hormone
type II receptor messenger ribonucleic acid expression in rat ovaries during postnatal
development, the estrous cycle, and gonadotropin-induced follicle growth.
Endocrinology, 136:4951-4962.
Ball, B. A., A. T. Vo, and J. Baumber. 2001. Generation of reactive oxygen species by equine
spermatozoa. Am. J. Vet. Res. 62:508-515.
Baruselli, P. S., E. O. S. Batista, L. M. Vieira, A. H. Souza. 2015. Relationship between follicle
population, AMH concentration and fertility in cattle. Anim. Reprod. 12:487-497.
Bashawat, M., M. Moussa, M. R. Al-Merestani, M., S. Destrumelle, D. Tainturier. 2016. The use
of different low-density lipoproteins concentrations for developing a new extender for
Awassi ram’s semen. Sci. J. King Faisal Univ. (Basic Appl. Sci.). 17:21-30.
Bautista, J. A., Y. Takahashi, and H. Kanagawa. 1997. In vitro viability of mouse 8-cell embryos
vitrified in a simple solution of ethylene glycol. Jpn. J. Vet. Res. 45:67-73.
Bavister, B. D. 1981. Substitution of a synthetic polymer for protein in a mammalian gamete
culture system. J. Exp. Zool. 217:45-51.
Bavister, B. D., Chen, A. F. and P. C. Fu. 1979. Catecholamine requirement for hamster sperm
motility in vitro. J. Reprod. Fertil. 56:507-513.
Bezard, J., B. Vigier, D. Tran, P. Mauleon, and N. Josso. 1987. Immunocytochemical study of
anti-Müllerian hormone in sheep ovarian follicles during fetal and post-natal
development. J. Reprod. Fertil. 80:509-516.

25
Bolanos, J. R., J. W. Overstreet, and D. F. Katz. 1983. Human sperm penetration of zona-free
hamster eggs after storage of the semen for 48 hours at 2 degrees C to 5 degrees C. Fertil.
Steril. 39:536-541.
Broer, S. L., F. J. Broekmans, J. S. Laven, and B. C. Fauser, B. C. 2014. Anti-Müllerian
hormone: ovarian reserve testing and its potential clinical implications. Hum. Reprod.
Update. 20:688-701.
Burke, J. M. 2005. Lamb production of Dorper, Katahdin, and St. Croix bred in summer, winter,
or spring in the Southeastern US. Sheep Goat Res. J. 20:51-59.
Campbell, B. K., M. Clinton, and R.Webb. 2012. The role of anti-Müllerian hormone (AMH)
during follicle development in a monovulatory species (sheep). Endocrinology.
153:4533-4543.
Carrascosa, R. E., A. C Martini, M. F. Ponzio, J. M. Busso, A. A. Ponce, and J. L. Lacuara.
2001. Storage of Chinchilla lanigera semen at 4° C for 24 or 72 h with two different
cryoprotectants. Cryobiology. 42:301-306.
Cate, R. L., R. J. Mattaliano, C. Hession, R. Tizard, N. M. Farber, A. Cheung. 1986. Isolation of
the bovine and human genes for Müllerian inhibiting substance and expression of the
human gene in animal cells. Cell. 45:685-698.
Choong, C., and R. Wales. 1963. The use of various diluents for deep-freezing bull spermatozoa.
Aust. J. Biol. Sci. 16:896-904.
Cohen-Haguenauer, O., J. Y. Picard, J. Y. Mattei, S. Serero, N. Van Cong, M. F. N. Tand, D.
Guerrier, M. C. Hors-Cayla, N. Josso, J. Fresal. 1987. Mapping of the gene for anti-
Müllerian hormone to the short arm of human chromosome 19. Cytogenet. Genome Res.
44:2-6.
Dalgleish, D. G. 1998. Casein micelles as colloids: surface structures and stabilities. J. Dairy Sci.
81:3013-3018.
Davis, I. S., R. W. Bratton, and R. H. Foote. 1963. Livability of bovine spermatozoa at 5, -25,
and -85 C in Tris-buffered and citrate-buffered yolk glycerol extenders. J. Diary Sci.
46:333-336.
Delgado, P., T. Lester, and R. Rorie. 2009. Effect of a low-sodium, choline-based diluent on
viability of bovine sperm stored at refrigerator temperatures. Ark. Anim. Sci. Depart.
Rep. 77-79.
Desnoyers, L., and P. Manjunath. 1992. Major proteins of bovine seminal plasma exhibit novel
interactions with phospholipid. J. Biol. Chem. 267:10149-10155.
Donoghue, A. M., and G. J. Wishart. 2000. Storage of poultry semen. Anim. Reprod. Sci.
62:213-232.

26
Durlinger, A. L., M. J. Gruijters, P. Kramer, B. Karels, T. R. Kumar, M. M. M. M. Matzuk, U.
M. Rose, F. H. de Jong. J. T. Uilenbroek, J. A. Grootegoed. 2001. Anti-Mullerian
hormone attenuates the effects of FSH on follicle development in the mouse ovary.
Endocrinology. 142:4891-4899.
Evans, G., W. M. C. Maxwell. 1987. Salamon's Artificial Insemination of Sheep and Goats.
Butterworths, Sydney. 1-7.
Foot, R. H., and R. W Bratton. 1950. The fertility of bovine semen in extenders containing
sulfanilamide, penicillin, streptomycin, and polymyxin. J. Dairy Sci. 33:544-547.
Foote, R. H. 2002. The history of artificial insemination: selection notes and notables. J. Anim.
Sci. 80:1-10.
Foote, R. H., and R. W. Bratton. 1949. The fertility of bovine semen cooled with and without the
addition of citrate-sulfanilamide-yolk extender. J. Dairy Sci. 32:856-861.
Fukui, Y., and E. Roberts. 1978. Further studies on non-surgical intrauterine technique for
artificial insemination in the ewe. Theriogenology. 10:381-391.
Gao, D., P. Mazur, and J. K. Critser. 1997. Fundamental cryobiology of mammalian
spermatozoa. In: Karow A. M., and J. K. Critser (eds.), Reproductive Tissue Banking.
London: Academic Press.
Gharibi, S., A. Niasari-Naslaji, N. Poursasan, and A. A. Moosavi-Movahedi. 2014. Replacement
of salamon with shotor diluent and egg yolk with low density lipoprotein for chilled
storage of ram semen. Iran. J. Vet. Res. 15:279-284.
Gil, J., S. Fierro, O. Bentancur, and J. Olivera‐Muzante. 2011. Chilled storage of ram semen
improves with the addition of egg yolk and glycerol to milk‐based extenders. Reprod.
Dom. Anim. 46:503-507.
Gill, S. P., R. H. Hammerstedt, and R. P. Amann. 1999. Poultry artificial insemination:
Procedures, current status and future needs. In: Proc. Annu. Mtg. Soc. Theriogenology,
Nashville, TN. pp. 353-362.
Good, N. E., G. D. Winget, W. Winter, T. N. Connolly, S. Izawa, and R. M. Singh. 1966.
Hydrogen ion buffers for biological research. Biochemistry. 5:467-477.
Gourley D. D., and R. L. Riese. 1990. Laparoscopic artificial insemination in sheep. Vet. Clin.
North Am. Food Anim. Pract. 6:615-6133.
Graham, E. F., B. Crabo, and K. I. Brown. 1972. Effect of some zwitter ion buffers on the
freezing and storage of spermatozoa I. Bull1. J. Dairy Sci. 55:372-378.

27
Gündoğan, M. 2009. Short term preservation of ram semen with different extenders. Kafkas
Univ. Vet. Fak. Derg. 15:429-435.
Halbert G. W., H. Dobson, J. S. Walton, and B. C. Buckrell. 1990. The structure of the cervical
canal of the ewes. Theriogenology. 33:977-992.
Hammerstedt, R. H., and J. K. Graham. 1992. Cryopreservation of poultry sperm: the enigma of
glycerol. Cryobiology. 29:26-38.
Hollinshead, F.K., J. K. O’Brien, L. Gillan, M. Meyers, M., W. Maxwell, and G. Evans. 2004.
Liquid storage of flow cytometrically sorted ram spermatozoa. Theriogenology. 62:587-
605.
Holt, W. V., and R. North. 1984. Partially irreversible cold‐induced lipid phase transitions in
mammalian sperm plasma membrane domains: Freeze‐fracture study. J. Exp. Zool.
230:473-483.
Ireland, J. J., G. W. Smith, D. Scheetz, F. Jimenez-Krassel, J. K. Folger, J. L. Ireland, F. Mossa,
P. Lonergan, and A. C. Evans. 2010. Does size matter in females? An overview of the
impact of the high variation in the ovarian reserve on ovarian function and fertility, utility
of anti-Müllerian hormone as a diagnostic marker for fertility and causes of variation in
the ovarian reserve in cattle. Reprod. Fertil. Dev. 23:1-14.
Ireland, J., D. Scheetz, F. Jimenez-Krassel, A. Themmen, F. Ward, P. Lonergan, G. W. Smith, G.
I. Perez, A. C. O. Evans, and J. J. Ireland. 2008. Antral follicle count reliably predicts
number of morphologically healthy oocytes and follicles in ovaries of young adult cattle.
Biol. Reprod. 79:1219-1225.
Kershaw, C. M., M. Khalid, M. R. McGowan, K. Ingram, S. Leethongdee, G. Wax, and R. J.
Scaramuzzi. 2005. The anatomy of the sheep cervix and its influence on the transcervical
passage of an inseminating pipette into the uterine lumen. Theriogenology. 64:1225-
1235.
Kulaksiz, R., C. Cebi, and E. Akcay. 2012. The effect of different extenders on the motility and
morphology of ram sperm frozen or stored at 4 degrees C. Turk. J. Vet. Anim. Sci.
36:177-182.
La Marca, A., and A. Volpe. 2006. Anti‐Müllerian hormone (AMH) in female reproduction: is
measurement of circulating AMH a useful tool? Clin. Endocrinol. 64:603-610.
La Marca, A., V. Grisendi, and G. Griesinger. 2013. How much does AMH really vary in normal
women? Int. J. Endocrinol. 2013:1-8.
Lahoz, B., J. L. Alabart, and J. Folch. 2016a. Relationship between Anti-Müllerian hormone
(AMH) and the ovulatory response measured in three-month old ewe lambs with their
reproductive efficiency in adulthood. Asoc. Interprofes. Para el Des. Agr. 112:392-404.

28
Lahoz, B., J. L. Alabart, D. Monniaux, P. Mermillod, J. Folch. 2012. Anti-Mullerian hormone
plasma concentration in prepubertal ewe lambs as a predictor of their fertility at a young
age. BMC Vet. Res. 8:8-18.
Lahoz, B., J. L. Alabart, M. Cocero, D. Monniaux, E. Echegoyen, P. Sánchez, J. Folch. 2014.
Anti-Müllerian hormone concentration in sheep and its dependence of age and
independence of BMP15 genotype: an endocrine predictor to select the best donors for
embryo biotechnologies. Theriogenology. 81:347-357.
Lahoz, B., S. Fabre, J. H. Calvo, L. Drouilhet, P. Sanchez, P. Sarto, E. Echegoyen, J. Folch, D.
Monniaux, J. L. Alabart. 2016 b. Assessing the age-related relationship between plasma
anti-Mullerian hormone (AMH) and ovarian follicle population in prepubertal ewes
(abstract). 18th Int. Cong. Anim. Reprod. Nouzilly, France. 26-30, June 2016.
Leibfried, M. L., and B. D. Bavister. 1982. Effects of epinephrine and hypotaurine on in-vitro
fertilization in the golden hamster. J. Reprod. Fertil. 66:87-93.
López, A., L. Söderquist, and H. Rodriguez-Martinez. 1999. Sperm viability in ram semen
diluted and stored in three different extenders. Acta vet. Scand. 40:1-9.
MacDonald, B. J., and J. A. Foulkes. 1981. A spectrofluorometric investigation, using 1-anilino-
naphthalene-8-sulphonate, of the interaction between washed bovine spermatozoa and
seminal plasma or egg-yolk lipoprotein. J. Reprod. Fertil. 63:407-414.
Manjunath, P., L. Baillargeon, Y. Marcel, N. Seidah, M. Chrétien, and Chapdelaine. 1988.
Diversity of novel proteins in gonadal fluids. In: K. W. Mckerns, M. Chretien (eds.),
Molecular Biology of Brain and Endocrine Peptidergic Systems. New York: Plenum
Press:259-273.
Manjunath, P., V. Nauc, A. Bergeron, and M. Ménard. 2002. Major proteins of bovine seminal
plasma bind to the low-density lipoprotein fraction of hen's egg yolk. Biol. Reprod.
67:1250-1258.
Mata-Campuzano, M., M. Álvarez-Rodríguez, J. Tamayo-Canul, E. López-Urueña, P. de Paz, L.
Anel. 2014. Refrigerated storage of ram sperm in presence of Trolox and GSH
antioxidants: Effect of temperature, extender and storage time. Anim. Reprod. Sci.
151:137-147.
Maxwell, W. M. C. 1984. Current problems and future potential of artificial insemination
programs. In Reproduction in sheep. (ed. R. L. Lindsay and D. T. Pearce). Canberra, A.
C. T.: Australian Academy of Science and Australia Wool Corporation. pp. 291-298.
Maxwell, W. M. C., A. J. Landers, and G. Evans. 1995. Survival and fertility of ram
spermatozoa frozen in pellets, straws and minitubes. Theriogenology. 43:1201-1210.

29
Maxwell, W. M. C., and L. J. Hewitt. 1986. A comparison of vaginal, cervical and intrauterine
insemination of sheep. J. Agric. Sci. Camb. 106:191-193.
Maxwell, W. M. C., L. G. Butler, and H. R. Wilson. 1984. Intra-uterine insemination of ewes
with frozen semen. J. of Agric. Sci. 102:233-235.
Mazur, P. 1977. The role of intracellular freezing in the death of cells cooled at supraoptimal
rates. Cryobiology. 14:251-272.
Meizel, S. 1985. Molecules that initiate or help stimulate the acrosome reaction by their
interaction with the mammalian sperm surface. Dev. Dynam. 174:285-302.
Meizel, S., C. W. Lui, P. K. Working, and R. J. Mrsny. 1980. Taurine and hypotaurine: their
effects on motility, capacitation and the acrosome reaction of hamster sperm in vitro and
their presence in sperm and reproductive tract fluids of several mammals. Dev. Growth
Differ. 22:483-494.
Mickelsen, W. D., L. G. Paisley, J. F. Dahmen. 1981. The effect of season on the scrotal
circumference and sperm motility and morphology in rams. Theriogenology. 16:45-51.
Miller, G. F., D. W. Gliedt, J. M. Rakes, and R. W. Rorie. 1994. Addition of penicillamine,
hypotaurine and epinephrine (PHE) or bovine oviductal epithelial cells (BOEC) alone or
in combination to bovine in vitro fertilization medium increases the subsequent embryo
cleavage rate. Theriogenology. 41:689-696.
Milovanov, V. K. 1964. Artificial insemination of livestock in the U.S.S.R Trans. by A. Birron
and Z. S. Cole. S. Monson, Jerusalem. Tech. Services, U.S. Dept. Commerce,
Washington, DC.
Monniaux, D., L. Drouilhet, C. Rico, A. Estienne, P. Jarrier, J. Touzé, J. Sapa, F. Phocas, J.
Dupont, R. Dalbiès-Tran, and S. Fabre. 2012. Regulation of anti-Müllerian hormone
production in domestic animals. Reprod. Fertil. Dev. 25:1-16.
Munsterberg, A., and R. Lovell-Badge. 1991. Expression of the mouse anti-Mullerian hormone
gene suggests a role in both male and female sexual differentiation. Development.
113:613-624.
Najafi, A., H. Daghigh-Kia, H. V. Dodaran, M. Mehdipour, M. Alvarez-Rodriguez. 2017.
Ethylene glycol, but not DMSO, could replace glycerol inclusion in soybean lecithin-
based extenders in ram sperm cryopreservation. Anim. Reprod. Sci. 177:35-41.
O’Hara, L., J. Hanrahan, L. Richardson, A. Donovan, S. Fair, A. C. Evans, and P. Lonergan.
2010. Effect of storage duration, storage temperature, and diluent on the viability and
fertility of fresh ram sperm. Theriogenology. 73:541-549.
Pace, M., and E. Graham. 1974. Components in egg yolk which protect bovine spermatozoa
during freezing. J. Anim. Sci. 39:1144-1149.

30
Paulenz, H., L. Söderquist, R. Pérez-Pé, and K. Andersen Berg. 2002. Effect of different
extenders and storage temperatures on sperm viability of liquid ram semen.
Theriogenology. 57:823-836.
Paulenz, H., L. Söderquist, T. Ådnøy, K. Soltun, P. Saether, K. Fjellsøy, and K. A. Berg. 2005.
Effect of cervical and vaginal insemination with liquid semen stored at room temperature
on fertility of goats. Anim. Reprod. Sci. 86:109-117.
Paulenz, H., L. Söderquist, T. Ådnøy, O. H. Fossen, and K. A. Berg. 2003. Effect of milk-and
TRIS-based extenders on the fertility of sheep inseminated vaginally once or twice with
liquid semen. Theriogenology. 60:759-766.
Paulenz, H., T. Adnøy, O. Fossen, L. Söderquist, and K. A. Berg. 2002. Effect of deposition site
and sperm number on the fertility of sheep inseminated with liquid semen. Vet. Res.
150:299-302.
Paulenz, H., T. Ådnøy, O. H. Fossen, L. Söderquist. 2010. Effect on field fertility of addition of
gelatine, different dilution rates and storage times of cooled ram semen after vaginal
insemination. Reprod. Domest. Anim. 45:706-710.
Person, C. N., T. D. Lester, M. D. Person, and R. W. Rorie. 2007. Effects of penicillamine,
hypotaurine and epinephrine (PHE) on post-thaw bovine sperm parameters, as measured
by computer-assisted sperm analysis. Ark. Agri. Exp. Sta. Res. Ser. 553:47-50.
Phillips, P. H., and H. A. Lardy. 1940. A yolk-buffer pabulum for preservation of bull semen. J.
Dairy Sci. 23:399-404.
Plante, G., M. F. Lusignan, M. Lafleur, and P. Manjunath. 2015. Interaction of milk proteins and
Binder of Sperm (BSP) proteins from boar, stallion and ram semen. Reprod. Biol.
Endocrinol. 13:92-112.
Polge, C. 1956. The development of artificial insemination service for pigs. Anim. Breed.
24:209-217 (Abstr.).
Polge, C. 1968. Frozen semen and the A.I. Programme in Great Britain. In: Proc. 2nd Tech.
Conf. Artif. Insem. Reprod. NAAB, Columbia, MO. pp 46-51.
Polge, C., A. U. Smith, and A. S. Parkes. 1949. Revival of spermatozoa after vitrification and
dehydration at low temperature. Nature. 164:166.
Pope, C. E., Y. Z. Zhang, and B. L. Dresser. 1991. A simple staining method for evaluating
acrosomal status of cat spermatozoa. J. Zoo Wildl. Med. 22:87-95.
Purdy, P. H. 2006. A review on goat sperm cryopreservation. Small Rumin. Res. 63:215-225.

31
Quan, G. B., G. Q. Wu, Y. J. Wang, D. J. Li, Y. Ma, Q. H. Hong. 2016. Effects of the TRIS,
TES, or skim milk based extender on in vitro parameters of ram spermatozoa during
liquid storage. Small Rumin. Res. 134:14-21.
Rajabi-Toustani, R., M. Roostaei-Ali, and R. Motamedi-Mojdehi. 2014. Effect of different levels
of egg yolk on ram sperm coating and preserving at 5° C. Iran. J. of Vet. Res. 15: 168-
171.
Rajpert-De Meyts, E., N. Jørgensen, N. Græm, J. Muller, R. L. Cate, N. E. Skakkebæk. 1999.
Expression of anti-Mullerian hormone during normal and pathological gonadal
development: Association with differentiation of sertoli and granulosa cells 1. J. Clin.
Endocrinol. Metab. 84: 3836-3844.
Rehmana, F. U., C. Zhao, M. A. Shaha, M. S. Qureshib, and X. Wanga. 2013. Semen extenders
and artificial insemination in ruminants. Veterinaria. 1:1-8.
Ricker, J. V., J. J. Linfor, W. J. Delfino, P. Kysar, E. L. Scholtz, F. Tablin, J. H. Crowe, B. A.
Ball, and S. A. Meyers. 2006. Equine sperm membrane phase behavior: The effects of
lipid-based cryoprotectants. Biol. reprod. 74:359-365.
Rico, C., S. Fabre, C. Médigue, N. di Clemente, F. Clément, M. Bontoux, J. L. Touze, M.
Dupont, E. Briant, B. Remy, J. F. Beckers, and D. Monniaux. 2009. Anti-müllerian
hormone is an endocrine marker of ovarian gonadotropin-responsive follicles and can
help to predict superovulatory responses in the cow. Biol. Reprod. 80:50-59.
Rorie, R., H. Newberry, T. D. Lester, M. Acharya, and C. Hansen. 2016. Anti-müllerian
hormone at weaning and breeding as a predictor of beef heifer fertility. Reprod. Fertl.
Dev. 29:111-112 (Abstr.).
Salamon, S. 1976. Artificial insemination of sheep. Publicity Press, Chippendale. pp 61-64.
Salamon, S., and D. Visser. 1972. Effect of composition of Tris-based diluent and of thawing
solution on survival of ram spermatozoa frozen by the pellet method. Aust. J. Biol. Sci.
25:605-618.
Salamon, S., and W. M. C. Maxwell. 1995. Frozen storage of ram semen I. Processing, freezing,
thawing and fertility after cervical insemination. Anim. Reprod. Sci. 34:185-249.
Salamon, S., and W. M. C. Maxwell. 2000. Storage of ram semen. Anim. Reprod. Sci. 62:77-
111.
Salisbury, G. W., H. K. Fuller, and E. L. Willett. 1941. Preservation of bovine spermatozoa in
yolk-citrate diluent and field results from its use. J. Dairy Sci. 24:905-910.
Salvador, I., M. A. Silvestre, J. Yániz, M. P. Viudes-de-Castro, J. Bernacer, and E. A. Gómez.
2005. Effect of solid storage at 5° C on sperm motility of goat semen by addition of
gelatin. Reprod. Domest. Anim. 40: 381 (abstr.).

32
Scheetz, D. M. 2010. Regulation and role of anti-Mullerian hormone in bovine reproduction
(Doctoral dissertation). Retrieved from UMI ProQuest Database. (UMI No. 1487140).
Seidel, G. E., J. L. Schenk, L. A. Herickhoff, S. P. Doyle, Z. Brink, R. D. Green, D. G. Cran.
1999. Insemination of heifers with sexed sperm. Theriogenology. 52:1407-1420.
Senger P. L. 2003. Pathway to pregnancy and parturation. Pullman, WA. Current Concepts, Inc.
pp 36-56.
Shannon, P., B. Curson, and A. P. Rhodes. 1984. Relationship between total spermatozoa per
insemination and fertility of bovine semen stored in Caprogen at ambient temperature. N.
Z. J. Agric. Res. 27:35-41.
Silva, E., J. Cajueiro, S. Silva, A. Vidal, P. Soares, and M. Guerra. 2012. In vitro evaluation of
ram sperm frozen with glycerol, ethylene glycol or acetamide. Anim. Reprod. Sci.
132:155-158.
Stachecki, J. J., J. Cohen, and S. M. Willadsen. 1998 b. Cryopreservation of unfertilized mouse
oocytes: the effect of replacing sodium with choline in the freezing medium.
Cryobiology. 37:346-354.
Stachecki, J. J., J. Cohen, and S. Willadsen. 1998 a. Detrimental effects of sodium during mouse
oocyte cryopreservation. Biol. of Reprod. 59:395-400.
Susko-Parrish, J. L., M. B. Wheeler, R. L. Ax, N. L. First, and J. J. Parrish. 1990. The effect of
penicillamine, hypotaurine, epinephrine and sodium metabisulfite, on bovine in vitro
fertilization. Theriogenology. 33:333 (Abstr.).
The Baalands. 2017. Katahdin frequently asked question. Available online.
http://www.baalands.com/#!katahdinfaqs/cwoc. Accessed on 24th April 2017.
Torres-Rovira, L., A. Gonzalez-Bulnes, S. Succu, A. Spezzigu, M. E. Manca, G. G. Leoni, G. G.,
M. Sanna, S. Pirino, M. Gallus, S. Naitana, F. Berlinguer. 2014. Predictive value of antral
follicle count and anti-Müllerian hormone for follicle and oocyte developmental
competence during the early prepubertal period in a sheep model. Reprod. Fertil. Dev.
26:1094-1106.
Vanimisetti, H. B., D. R. Notter, and L. A. Kuehn. 2007. Genetic (co) variance components for
ewe productivity traits in Katahdin sheep. J. Anim. Sci. 85:60-80.
Vásquez, J. H., V. H. Núñez, E. A. Florentini, J. M. Gonzales, L. A. Camargo, and M. E.
Valdivia. 2013. Effects of five cryoprotective agents on quality of sheep epididymal
spermatozoa during pre-freezing. Livest. Sci. 152: 94-99.

33
Vigier, B., J. Y. Picard, D. Tran, L. Legeai, and N. Josso. 1984. Production of anti-Mullerian
hormone: another homology between sertoli and granulosa cells. Endocrinology.
114:1315-1320.
Visser, D., and S. Salamon. 1973. Fertility of ram spermatozoa frozen in a TRIS-based diluent.
Aust. J. Biol. Sci. 26:513-515.
Visser, J. A., F. H. de Jong, J. S. Laven, and A. P. Themmen. 2006. Anti-Müllerian hormone: a
new marker for ovarian function. Reproduction. 131:1-9.
Watson, P. F. 1995. Recent developments and concepts in the cryopreservation of spermatozoa
and the assessment of their post-thawing function. Reprod. Fertl. Dev. 7: 871-891.
Weenen, C., J. S. Laven, A. R. Von Bergh, M. Cranfield, N. P. Groome, J. A. Visser, P. Kramer,
B. C. Fauser, and A. P. N. Themmen. 2004. Anti-Mullerian hormone expression pattern
in the human ovary: potential implications for initial and cyclic follicle recruitment. Mol.
Hum. Reprod. 10:77-83.
White, I. G. 1993. Lipids and calcium uptake of sperm in relation to cold shock and preservation:
a review. Reprod. Fertil. Dev. 5:639-658.
Wildeus, S. 2014. Sire breed differences in pregnancy rate of hair sheep ewes following liquid
semen vaginal artificial insemination. J. Anim. Sci. 39-39 (Abstr.).
Wildeus, S., and E. J. Chozu. 2010. Liquid semen vaginal AI in three hair sheep breed during
transition to seasonal breeding. J. Anim. Sci. 88(suppl. 1):34-35 (Abstr.).
Wildeus, S., D. Wright, and D. O’Brien. 2016. Use of liquid semen artificial insemination in
Katahdin sheep in a commercial farm setting. J. Anim. Sci. 95(suppl. 1): 51.
Wilson, C., and D. Davies. 2007. The control of sexual differentiation of the reproductive system
and brain. Reproduction. 133:331-359.
Wusiman, A., Y-P. Wang, K. Ren, G-B. Zhou, X-W. Fu, L. Suo, Z-Q. Fan, L. Wang, and S-E.
Zhu. 2012. Semen storage at 23, 4 or-196° C and its application to artificial insemination
in small-tail Han sheep. Asian J. Anim. Vet. Adv. 7:299-308.
Yániz, J. L., J. A. Mateos, and P. Santolaria. 2011. Zwitterionic buffers preserve ram semen
quality more efficiently than TRIS during storage at 15° C. Small Rumin. Res. 95:54-60.
Yániz, J. L., P. Santolaria, M. A. Marco-Aguado, and F. López-Gatius. 2008. Use of image
analysis to assess the plasma membrane integrity of ram spermatozoa in different
diluents. Theriogenology. 70:192-198.

34
Chapter III
Effect of semen extender and storage temperature on ram sperm motility over time
Abstract
The objective was to determine the effect of semen extender and storage temperature on
motility of ram semen stored up to 72 h. Semen was collected by electroejaculator from 5
Katahdin rams. Each semen collection was divided and extended using four extenders: milk,
TRIS, TEST, and CJ-2, diluted to 600 million sperm/ml, and loaded to 0.25 ml straws for storage
at 4 and 15° C. All extenders were supplemented with 5% (v/v) egg yolk. Sperm motility
parameters were measured at 0, 24, 48, and 72 h of storage using computer an assisted sperm
analysis (CASA) system. Data were analyzed using the PROC MIXED procedure of SAS, with
each semen collection as the experimental unit. Variables in the model included extenders,
temperature, hours (repeated measures) and interactions. Most of the sperm motility parameters
were higher after storage at 4° C compared with 15° C. Ram semen extended using milk or TRIS
and stored at 4° C for 3 d maintained similar sperm motility parameters (P > 0.05). Semen
extended using CJ-2 or TEST extenders and stored at 4° C for 3 d had lower total motility and
path velocity (VAP; P < 0.05) compared with milk or TRIS, while most of the remaining sperm
motility parameters were similar (P > 0.05). For ram semen stored at 15° C for 3 d, TRIS had
higher sperm sperm motility parameters (P < 0.05) compared with other extenders. Milk or
TEST extender maintained intermediate motility, while CJ-2 extender had lowest sperm motility
parameters (P < 0.05) at 15° C. There was an extender × hour interaction (P < 0.05) on track
speed (VCL), path velocity (VAP), progressive velocity (VSL), beat frequency (BCF), lateral
amplitude (ALH), and straightness (STR). Such interactions are due to greater reduction of
sperm motility parameters at 48 and 72 h of storage for CJ-2 or milk extenders compared with
TEST or TRIS extenders. Within 24 h, percentage of progressive motile sperm decreased sharply

35
regardless of extenders or storage temperatures. Progressive motility of spermatozoa is positively
correlated to fertilizing capacity. Therefore, further research is needed to increase progressive
motility.
Keywords: Extenders, ram semen, storage temperature
Introduction
Artificial insemination (AI) of ewes with frozen ram semen often results in low
conception rates (20-30%; Salamon and Maxwell, 1995). Fresh diluted and chilled ram semen is
an alternative to frozen storage (Salamon and Maxwell, 1995). Better motility of ram semen was
found after storage at 4° C for 2 to 3 d compared with frozen storage for several weeks
(Wusiman et al., 2012; Kulaksiz et al., 2012). Cervical AI conception rate was higher when ram
semen extended with sodium citrate was stored at 4 or 23° C for 1 to 3 d compared with several
weeks storage at -196° C (Wusiman et al., 2012). Motility of ram semen was improved when
stored 5° C (O’Hara et al., 2010), or at 15° C (Mata-Campuzano et al., 2014). Differences in
motility among experiments may be due to the type of extender and storage duration.
Milk or tris[Hydroxymethyl]aminomethane (TRIS), based extenders are the most
common extenders used for liquid storage of ram semen (Salamon and Maxwell, 2000). The
TEST (N-[Tris(hydroxymethyl)methyl]-2-aminoethanesulfonic acid i.e., TES titrated with TRIS)
has been evaluated in other species but not rams (Bolanos et al., 1983; Pope et al., 1991). The
CJ-2 extender was initially formulated to overcome harmful effects of sodium for
cryopreservation of oocytes (Mazur, 1977; Stachecki and Willadsen, 1998). Bull semen extended
in CJ-2 egg yolk extender maintained acceptable motility up to 7 d of storage at 4° C (Delgado et
al., 2009. There are no reports for using CJ-2 extender for extension and chilled storage of ram

36
semen. Therefore, the objective of the current study was to compare four different extenders
(milk, TRIS, TEST or CJ-2) stored at two different temperatures (4 or 15° C) for 72 h.
Materials and methods
This trial was conducted at the University of Arkansas, Fayetteville, AR. All animal
procedures were approved by Institutional Animal Care and Use Committee (Protocol # 15019).
Animal management
Five mature Katahdin rams were clinically healthy and raised on mixed tall fescue
(Schedonorus arundinacea) and bermudagrass (Cynodon dactylon) pasture. All rams had access
to water, and mineral salt, and 0.5 kg/d of corn/soybean meal (17% CP, 82% TDN on DM basis)
per ram. Semen was collected by electroejaculation into 15 ml polystyrene tubes, screw capped
and placed immediately into a thermo-flask (Lab-line Instruments Inc., Melrose Park, IL)
containing water maintained at 35° C. Semen samples were transported to the laboratory for
analysis.
Evaluation of fresh semen by computer assisted sperm analysis (CASA)
All semen samples were evaluated within 2 h post collection, using a computer assisted
sperm analysis (CASA; Hamilton-Thorne, IVOS, Beverly MA) with 12 TOX IVOS software.
Sperm motility parameters were analyzed using the default analysis settings recommended by
manufacturer for ram semen. Semen samples were diluted in CJ-2 extender without egg yolk at
1:100 (semen: extender ratio). About 3.5 µl of diluted sample was placed in disposable 20 µm
deep, 2-chambered IVOS slide (Cytonix, Beltsville, MD). At least four fields were scanned, with
30 video frames captured per field at a frame rate of 60 Hz. A minimum of 400 sperm cells were
counted in each sample. Variables measured with CASA were motility (%), progressive motility
(%), track speed (VCL, µm/sec), path velocity (VAP, µm/sec), progressive velocity (VSL,

37
µm/sec), beat frequency (BCF, MHz), lateral amplitude (ALH, µm), linearity (LIN, %), and
straightness (STR, %; Table 1).
Preparation of semen extenders
Milk extender
All the ingredients were from Sigma Company unless otherwise mentioned. Locally
purchased 10 g of organic skim milk powder (NOW Real Food®, organic, non-fat dry milk
powder) was dissolved in a total volume of 95 ml of lukewarm Milli-Q water in a 250-ml beaker.
The solution was constantly stirred for 10 min using a magnetic stirrer while heated at 95° C on a
hot plate (VWR, West Chester, PA model 371). The solution was cooled to room temperature
and then 0.5 g of fructose and 0.5 ml of gentamicin (10 mg/ml; Life Technologies, Gibco®) were
added. The stock solution was stored at 4° C. On the day of semen collection, 5 ml of egg yolk
from USDA certified organic egg was added to the stock solution, mixed thoroughly and
warmed to 35° C.
TEST extender
The TES buffer of 7.5 g was dissolved in 100 ml Milli-Q water in a 250-ml beaker. In
another beaker, 3.93 g of TRIS (tris[Hydroxymethyl] aminomethane) was dissolved in 100 ml
Milli-Q water. The TES solution was titrated with TRIS to a pH of 7.2, using a pH meter (Model
420A, Orion Laboratory Products Division Inc., Boston, MA). Osmolarity of the titrated solution
was 318 mOsmkg-1 determined with a computerized micro-osmometer (Multi-Osmette 2430,
Precision Systems, Natick, MA, USA). One half gram fructose and 0.5 ml of gentamicin (10
mg/ml; Life Technologies, Gibco®) was added to 100 ml of titrated solution. The stock solution
was filtered through a 0.45 µm sterile Gelman Acrodisk filter. The filtered stock solution was

38
stored at 4º C. On the day of semen collection, egg yolk was added to 95 ml of the stock solution
as described for milk extender.
TRIS extender
The extender was prepared by adding 3.634 g of TRIS (tris[Hydroxymethyl]
aminomethane) and 1.99 g of citric acid to 100 ml Milli-Q water in a 250-ml beaker. The pH of
the resulting solution was 7.0 measured using a pH meter (Model 420A, Orion Laboratory
Products Division Inc., Boston, MA). Fructose, gentamicin and egg yolk was added and the
extender was made ready for extension as described for TEST.
CJ-2 extender
Choline extender (CJ-2) was prepared according to the method described by Stachecki et
al. (1998) with modification. Gentamicin was used instead of kanamycin sulfate. Lactic acid,
hemi-calcium salt was substituted for CaCL2*2H2O. Glucose was substituted with fructose, and
phenol red was not added. The pH and osmolarity were 7.55 and 280 mOsm, respectively. To
prepare CJ-2 extender, KCL (0.2 g), KH2PO4 (0.2 g), Na2HPO4 (1.1 g) and MgCl2*6H20 (0.1 g),
was added to a total volume of 1 liter of Milli-Q water and stirred until completely dissolved.
Lactic acid (calcium lactate; 19.259 g) was added very slowly and stirred continuously to prevent
calcium precipitation by K2HPO4. Choline chloride (C5H14NOCl) was added last. The stock
solution was then filtered through a 0.20 µm sterile Gelman Acrodisk filter and stored at 4° C.
Fructose, gentamicin and egg yolk was added and the extender was prepared as described for
TEST.
Preparation of semen for storage
Semen was initially extended to 150 million cells/0.25 ml in each extender. Extended
semen (50 µl aliquots) was then stored in 0.25 ml semen straws (IMV, Paillette Crista, France).

39
However, by initially filling straws with 50 µl respective extender, separating with 50 µl volume
of air, injecting the 50 µl of experimental extended semen, separated by another 50 µl volume of
air, then 50 µl of extender to insulate the extended semen. Total 24 straws were prepared from
single semen collection per ram. The straw was sealed with an electric sealer (H. W. Andersen
Products, Inc., Oyster Bay, NY). Half of the straws were placed in water in a laboratory flask
(Sigma Chemical Co, St. Louis, MO) at 35° C. This flask was then placed in a refrigerator at 4°
C. Remaining straws were similarly placed in anther flask (Sigma Chemical Co, St. Louis, MO)
which was then placed in a cooler (Koolatron PC3, Koolatron, Brantford, Ontario, Canada)
maintained at 15° C. Extended semen was cooled to desired temperature over 2 to 3 h period
(Table 2).
Evaluation of extended semen using CASA
Extended semen was evaluated by IDENT fluorescence method. The IDENT STAIN
(Hoechst 33342) illuminates only DNA containing objects. Minimum and maximum static size
and intensity gates were used to eliminate the chance of counting any other cells. The
illumination intensity for fluorescent illumination was set up between 2050 and 4050. The
chilled extended semen was pre-warmed in a thaw jug maintained at 35° C for 15 sec. The straw
was wiped dry and cut with scissor adjacent to extended semen. Extended semen was removed
with pipet and kept in a well plate. About 3.5 µl of extended semen was pipetted and mixed with
pre-warmed 95 µl of corresponding extender (without egg yolk) in a 0.5 ml Eppendorf tube, then
5 µl of Hoechst 33342 stain (10 µg/ml) was added to the diluted semen and stored in heating
block maintained at 35° C for 15 min before analysis (IVOS IDENT illumination option). For
semen samples diluted using milk extender, 10 µl of Hoechst 33342 stain was added. After 15
min of equilibration with the stain, the sample was analyzed using IVOS system. About 3.5 µl of

40
diluted semen was placed in disposable 20 µm deep, 2-chambered IVOS slide (Cytonix,
Beltsville, MD). The semen analysis procedure was followed as described for fresh semen.
Extended semen was analyzed every 24 h for 7 d. The IVOS machine setting for ram semen
motility analysis is given in a Table 3.
Statistical analysis
Semen motility parameters were analyzed at 24, 48 and 72 h after storage. Data were
analyzed as repeated measures using MIXED procedure of SAS (Littell et al., 1996), with semen
collection of each ram as the experimental unit. Variables in the model included extender,
temperature, hour (as a repeated measures of 24, 48, and 72 h), and all 2 and 3 way interactions.
Non-significant (P > 0.1) interactions were removed from the model. The final model included
extender, temperature × extender, and extender × hour interactions. Data are presented as least
squares means ± standard error. Means separation occurred using PDIFF option of SAS.
Dependent variables measured with IVOS were total motility, progressive motility, VCL, VAP,
VSL, BCF, ALH, LIN, and STR.
Results
Total motility of fresh semen was 48.4%. Total motility decreased to 43.6 ± 2.4, 31.1 ±
2.4, and 27.2 ± 2.4% at 24, 48, and 72 h respectively (P < 0.01). There was a temperature ×
extender interaction (P < 0.01), mainly due to decreased motility of sperm cells stored in CJ-2
and milk based extenders at 15° C. Total motility of sperm cells stored at 4° C in TRIS or milk
was higher compared with TEST or CJ-2 (P < 0.05; Table 4). Extended semen stored at 4° C
tended (P = 0.08) to have a higher sperm motility compared with that stored at 15° C. Motility of
sperm cells stored at 15° C was highest in TRIS compared with other extenders (P < 0.05).

41
Progressive motility of non-extended semen was 35.2%. Progressive motility declined to
8.6 ± 0.7, 5.3 ± 0.7, and 3.2 ± 0.7% at 24, 48, and 72 h, respectively (P < 0.01). There was a
temperature × extender interaction (P < 0.01) mainly due to a decrease in progressive motility of
sperm cells stored in CJ-2 extender at 15° C. Progressive motility of sperm cells stored at 4° C
using TRIS, CJ-2, and milk were similar (P > 0.05) but higher compared with TEST (P < 0.05).
Track speed of fresh semen was 226.3 µm/sec. Mean VCL at 24, 48, and 72 h declined to
116.6 ± 4.2, 95.9 ± 4.2, and 82.1 ± 4.2 µm/sec, respectively (P < 0.01). There was a trend toward
extender × hour interaction on VCL (P = 0.060; Table 5), mainly due to a greater reduction of
VCL on semen stored in CJ-2 and milk extender at 48 and 72 h of storage. A temperature ×
extender interaction on VCL was observed (P < 0.01), mainly due to decreased VCL of sperm
cells extended using CJ-2 or milk and stored at 15° C. Track speed of sperm cells stored at 4° C
was similar across extenders (P > 0.05).
Path velocity of fresh semen was 155.7 µm/sec. Overall VAP at 24, 48 and 72 h was 61.8
± 2.4, 50.4 ± 2.4, and 42.5 ± 2.4 µm/sec, respectively (P < 0.01). An extender × hour interaction
was observed on VAP of sperm cells (P = 0.023), mainly due to a reduction of VAP when stored
in CJ-2 and milk based extenders at 48 and 72 h, respectively. A temperature × extender
interaction on VAP was observed (P < 0.01), mainly due to decreased VAP of sperm cells stored
in CJ-2 or milk and stored at 15° C. Path velocity of sperm cells stored at 4° C using CJ-2, milk,
and TRIS was similar but higher than sperm cells stored in TEST (P < 0.05).
Progressive velocity of fresh sperm cells was 139.3 µm/sec. Overall VSL of sperm cells
at 24, 48, and 72 h declined to 44.5 ± 1.7, 36.1 ± 1.7, 29.1 ± 1.7 µm/sec, respectively (P < 0.01).
An extender × hour interaction was observed for VSL (P = 0.023), mainly due to a reduction of
VSL of sperm cells stored in CJ-2 or milk extenders at 48 and 72 h, respectively. An extender ×

42
temperature interaction was also observed (P < 0.01), mainly due to a decrease of VSL of sperm
cells stored in CJ-2, milk or TEST extenders at 15° C. Progressive velocity of sperm cells stored
at 4° C using CJ-2, milk, and TRIS was similar but higher than sperm cells stored in TEST (P <
0.05).
Beat frequency of fresh sperm was 33.4 MHz. Overall, BCF at 24, 48, and 72 h declined
to 34.4 ± 1.5, 29.3 ± 1.5, and 26.4 ± 1.5 MHZ, respectively (P < 0.01). An extender × hour
interaction was observed on BCF (P = 0.005), mainly due to a reduction of BCF of sperm stored
in CJ-2 or milk extenders at 48 and 72 h, respectively. There was an extender × temperature
interaction (P = 0.01) due to a decrease of BCF of sperm cells stored in CJ-2 or milk based
extenders at 15° C. Beat frequency of sperm cells stored at 4° C was similar across extenders (P
> 0.05).
Lateral amplitude of fresh semen was 6.88 µm. Overall ALH at 24, 48, and 72 h of
storage declined to 6.1 ± 0.2, 5.3 ± 0.2, and 4.8 ± 0.2 µm, respectively (P = 0.011). There was an
extender × temperature interaction (P = 0.03), mainly due to a decrease in ALH in CJ-2 or milk
based extenders at 15° C. Lateral amplitude of sperm cells stored at 4° C was lower in CJ-2 and
milk compared with TEST or TRIS extenders (P < 0.05).
Linearity of fresh semen was 65%. Linearity at 24, 48, and 72 h was 38.3 ± 1.7, 32.7 ±
1.7, and 29.5 ± 1.7 %, respectively (P > 0.05). An extender × hour interaction was observed on
LIN of sperm cells (P = 0.023), mainly due to a reduction of LIN of sperm cells stored in CJ-2 or
milk based extenders at 48 and 72 h, respectively. Temperature × extender interaction on LIN
was observed (P < 0.01), mainly due to a decrease of LIN of sperm cells stored in CJ-2 at 15° C.
Linearity of sperm cells stored at 4° C was similar across extenders (P > 0.05).

43
Straightness of fresh sperm cells was 87%. Overall STR of sperm cells at 24, 48, and 72 h
declined to 66.6 ± 2.8, 50.4 ± 2.4, and 42.5 ± 2.4, MHz, respectively (P = 0.01). An extender ×
hour interaction was observed on STR of sperm cells (P = 0.003), mainly due to a reduction of
STR of sperm cells stored in CJ-2 or milk extenders at 48 and 72 h of storage. There was an
extender × hour interaction, mainly due to a decrease of STR of sperm stored in CJ-2 or milk at
15° C (P < 0.05). Straightness of sperm cells stored at 4° C was similar across extenders (P >
0.05).
Discussion
Effective technologies are needed to move sheep germplasm readily between farms to
facilitate genetic improvement and to allow the use of superior genetic resources. Artificial
insemination enables fast spread of quality genetic traits between farms. Artificial insemination
with frozen ram semen results in low conception rates (20-30%; Salamon and Maxwell, 1995).
Fresh diluted and cold stored semen is an alternative to frozen storage (Salamon and Maxwell,
1995). Ram semen stored at 4° C for a short period (2 to 3 d) of time was more beneficial than
frozen storage for a long-period of time (-196° C for several weeks; Wusiman et al., 2012;
Kulaksiz et al., 2012). Studies on liquid storage of ram semen concluded that semen can
successfully be cold stored at 5 or 15° C for 2 to 3 d (Hollinshead et al., 2004; Mata-Campuzano
et al., 2014). Storage at 5° C maintained viability over storage at 15° C when stored for 72 h (O’
Hara et al., 2010). However, a different result was reported when stored for up to 48 h (Mata-
Campuzano et al., 2014). Use of different extenders, supplements and storage durations in those
studies could have impacted results. In the current study, we compared four different extenders
(milk, TEST, TRIS, and CJ-2) at two storage temperature (5 or 15° C) for extension and storage
of ram semen, using similar concentration of egg yolk, fructose, and gentamycin in each

44
extender. Semen was evaluated before extension and every 24 h up to 72 h. Most of the CASA
motility parameters were higher after storage at 4 compared with 15° C in all the extenders.
However, if ram semen is to be stored at 15° C, then our data from the current study indicates
that semen stored at 15° C for 3 d using TRIS extender had better sperm motility parameters
compared with milk, TEST, or CJ-2 based extenders. Based on results obtained from the current
study, it is suggested that ram semen stored at 4 is superior over storage at 15º C for 3 d.
Milk or TRIS based extenders are the most common extenders used for extension of ram
semen (Lopaz et al., 1999; Paulenz et al., 2002; Hollinshead et al., 2004; Gundogan et al., 2009;
Quan et al., 2016). The TEST based extender was used for extension of semen in other species,
but not commonly used for the extension of ram semen (Bolanos et al., 1983; Pope et al., 1991).
The CJ-2 extender was developed to avoid intercellular damage caused by sodium in sodium
based extenders (Mazur, 1977; Stachecki and Willadsen, 1998a; Stachecki and Willadsen,
1998b). The use of CJ-2 based extender for cool storage of bull semen was tested previously
(Delgado et al., 2009), but to our knowledge it has not been tested for cool storage of ram semen.
In general, the current study shows that ram semen stored at 4° C using milk or TRIS based
extenders maintained similar but better motility parameters compared with storage using TEST
or CJ-2 extenders. There was no difference in most motility parameters using TRIS or milk
based extenders for extension and storage of ram semen at 4° C for 3 d in the current study, these
results are similar to that reported by Gundogan et al. (2009), but contrary to results reported by
others (Paulenz et al., 2002; Hollinshead et al., 2004; Kulaksiz et al., 2012; Quan et al., 2016;
Yaniz et al., 2011). Some studies reported ram semen extended in milk extender maintained
better motility over TRIS (Hollinshead et al., 2004; Wusiman et al., 2012) after storage at 4° C
while others reported better motility using TRIS extender (Paulenz et al., 2002; Lopez et al.,

45
1999; Quan et al., 2016). Differences observed between treatments could be due to use of
different percentage of egg yolk and other supplements in the extender medium. Additionally,
duration of storage differed. Hollinshead et al. (2004) used 6 million spermatozoa in 0.5 ml
extender to store in 5 ml sterile sealed tubes to evaluate after 6 h of storage at 4° C, whereas,
Mata-campuzano et al. (2014) stored at 15° C for 48 h.
In the current study, we used a similar concentration of egg yolk, antibiotics, and fructose
in each of the extenders. Semen was evaluated at a constant interval of 24 h for 3 d. Ram semen
stored using milk or TRIS based extenders maintained similar sperm motility parameters but
better compared with TEST or CJ-2 extenders when stored at 4° C for 3 d.
Milk extender with 5% egg yolk resulted in better conception rate compared with TRIS
extended with 20% egg yolk when ewes were vaginally inseminated with fresh extended semen
(Paulenz et al., 2003). In a preliminary trial, we artificially inseminated 19 Gulf Coast Native
ewes with fresh semen extended in milk with 5% egg yolk or TRIS with 5% egg yolk. The same
percentage of ewes conceived in both groups (milk: 5/9; TRIS: 5/9).
In the current study, there was an extender by storage time interaction in most sperm
motility parameters measured. Motility parameters were similar up to 24 h of storage, but after
that sperm motility declined more rapidly in milk or CJ-2 extenders compared with TEST or
TRIS extender. Such a decrease may be due to a greater reduction at the 15° C temperature using
milk or CJ-2 extenders. Consistent with our results, TEST or TRIS extenders had similar motility
parameters after extension and storage of rabbit semen at 15° C (Roca et al., 2000) for 96 h. A
greater reduction of motility in milk compared with TRIS based extenders in the current study is
similar to that reported by Gundogan et al. (2009), who reported 50% of ram sperm extended in
TRIS up to 4 d compared with similar motility up to only 3 d in milk extender.

46
In the current study, most of sperm motility parameters extended using CJ-2 and milk
decreased sharply after storage at 15° C compared with TRIS or TEST extender. However, such
decreases are not very dramatic after storage at 4° C. It is not known why a greater reduction of
motility of sperm cells was seen after storage using CJ-2 extenders at 15° C. Choline based
extender provided better medium for storage of oocytes compared with traditional sodium
containing medium (Mazur, 1977; Stachecki and Willadsen, 1998a; Stachecki and Willadsen,
1998b). Choline is a component of the phospholipid bilayer and helps in stabilizing cell plasma
membrane for protection of spermatozoa and oocytes. Bull semen extended and cold-stored at 4°
C for 7 d using CJ-2 extender had equal or higher sperm motility parameters compared with
semen stored in synthetic caudal epididymal plasma (Delgado et al., 2009).
In conclusion, most motility parameters were significantly higher after extension and
storage at 4 compared with 15° C. Ram semen stored using milk or TRIS based extenders at 4° C
can maintain similar motility parameters but better compared with TEST or CJ-2 extenders after
storage for 3 d. In the current study, ram semen stored at 15° C for 3 d using TRIS extender had
better sperm motility parameters compared with milk, TEST or CJ-2 extender. Milk and TEST
based extenders had intermediate efficacy for maintaining motility at 15° C. In contrast, CJ-2
based extenders had drastic reduction of most motility parameters after storage at 15° C.
Combined results from both temperatures shows most of the motility parameters were similar up
to 24 h of storage but after that sperm motility declined more rapidly in milk or CJ-2 extenders
compared with TEST or TRIS extender. Such a decrease is due to a greater reduction after 24 h
using milk and CJ-2 extenders and storage at 15° C. Considering sperm progressive motility,
which is also a predictor of in vivo fertility, milk and TRIS extenders stored at 4° C had similar
but higher progressive motility compared with ram sperm extended in CJ-2 and TEST extenders.

47
However, progressive motility of ram sperm declined by 75% when stored at 4° C for 24 h, and
continued to decline over time regardless of extenders used. Further studies can be done to
supplement milk or TRIS based extenders for extension and storage of ram semen at 4° C.

48
References
Bolanos, J. R., J. W. Overstreet, and D. F. Katz. 1983. Human sperm penetration of zona-free
hamster eggs after storage of the semen for 48 hours at 2 degrees C to 5 degrees C. Fertil.
Steril. 39:536-541.
Delgado, P., T. Lester, and R. Rorie. 2009. Effect of a low-sodium, choline-based diluent on
viability of bovine sperm stored at refrigerator temperatures. Ark. Anim. Sci. Depart.
Rep. 77-79.
Gündoğan, M. 2009. Short term preservation of ram semen with different extenders. Kafkas
Univ. Vet. Fak. Derg. 15:429-435.
Hollinshead, F., J. K O’Brien, L. Gillan, M. Meyers, W. M. C. Maxwell, and G. Evans. 2004.
Liquid storage of flow cytometrically sorted ram spermatozoa. Theriogenology, 62:587-
605.
Kulaksiz, R., C. Cebi, and E. Akcay. 2012. The effect of different extenders on the motility and
morphology of ram sperm frozen or stored at 4 degrees C. Turk. J. Vet. Anim. Sci. 36:
177-182.
López, A., L. Söderquist, and H. Rodriguez-Martinez. 1999. Sperm viability in ram semen
diluted and stored in three different extenders. Acta Vet. Scand. 40:1-9.
Mata-Campuzano, M., M. Álvarez-Rodríguez, J. Tamayo-Canul, E. López-Urueña, P. de Paz, L.
Anel, and F. Martinez-Pastor. 2014. Refrigerated storage of ram sperm in presence of
Trolox and GSH antioxidants: Effect of temperature, extender and storage time. Anim.
Reprod. Sci. 151:137-147.
Mazur, P. 1977. The role of intracellular freezing in the death of cells cooled at supraoptimal
rates. Cryobiology. 14:251-272.
O’Hara, L., J. Hanrahan, L. Richardson, A. Donovan, S. Fair, A. C. Evans, and P. Lonergan.
2010. Effect of storage duration, storage temperature, and diluent on the viability and
fertility of fresh ram sperm. Theriogenology. 73:541-549.
Paulenz, H., L. Söderquist, R. Pérez-Pé, and K. Andersen Berg. 2002. Effect of different
extenders and storage temperatures on sperm viability of liquid ram semen.
Theriogenology. 57:823-836.
Paulenz, H., L. Söderquist, T. Ådnøy, O. H. Fossen, and K. A. Berg. 2003. Effect of milk-and
TRIS-based extenders on the fertility of sheep inseminated vaginally once or twice with
liquid semen. Theriogenology. 60:759-766.
Pope, C. E., Y. Z. Zhang, and B. L. Dresser. 1991. A simple staining method for evaluating
acrosomal status of cat spermatozoa. J. Zoo Wildl. Med. 22:87-95.

49
Quan, G. B., G. Q. Wu, Y. J. Wang, D. J. Li, Y. Ma, and Q. H. Hong. 2016. Effects of the Tris,
Tes, or skim milk based extender on in vitro parameters of ram spermatozoa during liquid
storage. Small Rum. Res. 134: 14-21.
Roca, J., S. Martinez, J. M. Vasquez, X. Lucas, I. Parrills, and E. A. Martinez. 2001. Viability
and fertility of rabbit spermatozoa diluted in Tris-buffer extender and stored at 15° C.
Anim. Reprod. Sci. 64:103-112.
Salamon, S., and W. M. C. Maxwell. 1995. Frozen storage of ram semen I. Processing, freezing,
thawing and fertility after cervical insemination. Anim. Reprod. Sci. 34:185-249.
Salamon, S., and W. M. C. Maxwell. 2000. Storage of ram semen. Anim. Reprod. Sci. 62:77-
111.
Stachecki, J. J., J. Cohen, and S. M. Willadsen. 1998 b. Cryopreservation of unfertilized mouse
oocytes: the effect of replacing sodium with choline in the freezing medium.
Cryobiology. 37:346-354.
Stachecki, J. J., J. Cohen, and S. Willadsen. 1998 a. Detrimental effects of sodium during mouse
oocyte cryopreservation. Biol. Reprod. 59:395-400.
Wusiman, A., Y. P. Wang, K. Ren, G. B. Zhou, X. W. Fu, L. Suo, Z. Q. Fan, L. Wang, and S. E.
Zhu. 2012. Semen Storage at 23, 4 or -196° C and its application to artificial
insemination in small-tail Han sheep. Asian J. Anim. Vet. Adv. 7:299-308.
Yániz, J. L., J. A. Mateos, and P. Santolaria. 2011. Zwitterionic buffers preserve ram semen
quality more efficiently than TRIS during storage at 15° C. Small Ruminant Res. 95:54-
60.

50
Table 1. CASA parameters
Parameters Description
Total motility (%) Path velocity ≥ 30 µm/sec and progressive velocity ≥ 15
µm/sec
Progressive motility (%) Path velocity ≥ 50 µm/sec and straightness ≥ 70%
Track speed (VCL, µm/sec) Average velocity measured over the actual point-to-point
track.
Path velocity (VAP, µm/sec) Average velocity of the smoothed (best-fit) sperm path.
Progressive velocity (VSL,
µm/sec)
Average velocity over straight line from the beginning to
the end of the sperm track.
Beat frequency (BCF, MHz) Frequency of sperm head crossing the sperm average path
in either direction.
Lateral amplitude (ALH,
µm)
Mean width of the head oscillation as the sperm swims.
Linearity (LIN, %) Departure of actual sperm path from straight line
(VSL/VCL).
Straightness (STR, %) Departure of average sperm path from a straight line
(VSL/VAP).
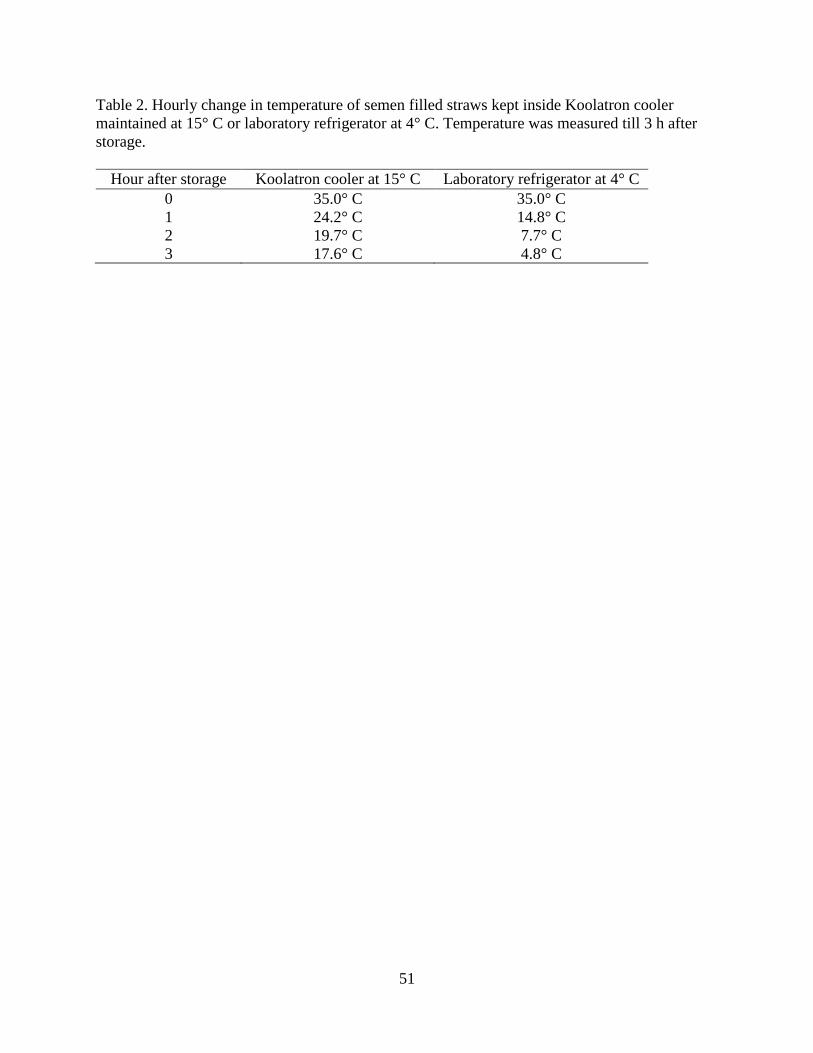
51
Table 2. Hourly change in temperature of semen filled straws kept inside Koolatron cooler
maintained at 15° C or laboratory refrigerator at 4° C. Temperature was measured till 3 h after
storage.
Hour after storage Koolatron cooler at 15° C Laboratory refrigerator at 4° C
0 35.0° C 35.0° C
1 24.2° C 14.8° C
2 19.7° C 7.7° C
3 17.6° C 4.8° C

52
Table 3. IVOS machine settings for ram semen motility analysis
Parameter Setting
Frames per second 60
Number of frames 30
Minimum contrast 60
Minimum size 5
Default cell size 5
Default cell intensity 55
Path velocity (VAP) 75.0
Straightness (STR) 80
Slow cells Static
VAP cut off 21.9
VSL cut off 6.0
Minimum intensity gate 0.25
Maximum intensity gate 1.50
Minimum size gate 0.60
Maximum size gate 8.00
Minimum elongation gate 0
Maximum elongation gate 95
UV fluorescent objective 10×
Target photometer 45-55
Field type Dark
Chamber depth 20

53
Table 4. Effect of temperature on ram sperm motility parameters over 72 h of storage. Ram
sperm motility parameters of extended and stored ram semen diluted in CJ-2, milk, TEST, and
TRIS extenders and stored at 4 or 15° C for 72 h.
Sperm motility Storage Extender treatment
characteristics temperature CJ-2 Milk TEST TRIS SE
Total motility 4° C 30.1b,x 40.6ab,x 30.4b,x 44.5a,x 4.1
15° C 9.1c,y 20.1c,y 41.3b,x 55.2a,x 4.1
Progressive motility 4° C 6.8a,x 7.8a,x 2.8b,x 7.4a,y 1.1
15° C 1.0c,y 4.9b,x 3.8bc,x 11.1a,x 1.1
Track speed 4° C 101.1a,x 116.1a,x 103.4a,x 118.1a,x 6.9
15° C 43.6c,y 66.1b,y 105.5a,x 131.7a,x 6.9
Path velocity 4° C 56.3ab,x 65.5a,x 50.7b,x 60.6ab,x 3.9
15° C 22.9d,y 38.3c,y 51.9b,x 66.2a,x 3.9
Progressive velocity 4° C 42.5a,x 45.2a,x 34.1b,x 42.9a,x 2.9
15° C 17.1c,y 29.6b,y 34.4b,x 46.5a,x 2.9
Beat frequency 4° C 33.3a,x 34.1a,x 32.5a,x 34.7a,x 2.5
15° C 18.2b,y 22.8b,y 31.1a,x 33.5a,x 2.5
Lateral amplitude 4° C 5.1b,x 5.4ab,x 6.6a,x 6.6a,x 0.4
15° C 2.8b,y 3.6b,y 6.4a,x 6.7a,x 0.4
Linearity 4° C 39.2a,x 40.3a,x 35.1a,x 36.8a,x 2.1
15° C 18.4b,y 29.7a,y 33.8a,x 34.7a,x 2.1
Straightness 4° C 65.6a,x 67.2a,x 65.1a,x 67.1a,x 4.5
15° C 32.5c,y 48.3b,y 63.7a,x 63.6a,x 4.5
Data expressed are the least square means and standard error (SE) for sperm motility parameters
taken at 24, 48, and 72 h after storage. abcdValues within the same row with unlike superscripts are different (P < 0.05). xyValues within the same column with unlike superscripts differ (P < 0.05).

54
Table 5. Effect of time of storage on ram sperm motility parameters, CJ-2, milk, TEST or TRIS
and stored at 4 and 15° C for 72 h.
Extenders Time of evaluation
24 h 48 h 72 h SE
Track speed
CJ-2 109.9a,x 59.8b,z 47.2b,z 8.5
Milk 111.2a,x 89.8a,y 72.1b,y 8.5
TEST 110.5a,x 106.2a,xy 96.8a,x 8.5
TRIS 134.6a,x 127.8a,x 112.2a,x 8.5
Path velocity
CJ-2 60.9a,xy 32.9b,z 25.1b,z 4.8
Milk 65.1a,xy 51.1b,y 39.7b,y 4.8
TEST 53.7a,xy 52.1a,y 48.1a,x 4.8
TRIS 67.5a,x 65.6a,x 57.1a,x 4.8
Progressive velocity
CJ-2 46.1a,x 25.3b,z 17.8b,yz 3.5
Milk 49.4a,x 37.1b,y 25.6c,y 3.5
TEST 35.4a,y 34.7a,y 32.8a,xy 3.5
TRIS 47.1a,x 47.3a,x 39.7a,x 3.5
Beat frequency
CJ-2 38.1a,x 22.8b,y 16.4b,y 3.6
Milk 34.4a,x 27.9ab,xy 23.1b,y 3.6
TEST 32.1a,x 31.5a,x 31.8a,x 3.6
TRIS 33.2a,x 35.1a,x 34.4a,x 3.6
Linearity
CJ-2 40.7a,xy 25.9b,y 19.9b,y 3.4
Milk 44.7a,x 33.5ab,xy 26.9b,xy 3.4
TEST 33.2a,y 34.2a,xy 36.6a,x 3.4
TRIS 37.7a,y 37.2a,x 35.5a,x 3.4
Straightness
CJ-2 69.7a,x 43.3b,z 34.2b,z 5.6
Milk 72.1a,x 56.1b,xy 45.5b,yz 5.6
TEST 62.8a,x 64.3a,x 66.1a,x 5.6
TRIS 62.2a,x 68.5a,x 65.4a,x 5.6
Data expressed are the least square means and standard error (SE) for parameters taken at 24, 48,
and 72 h for extender by hour interactions. abcdValues within the same row with unlike superscripts are different (P < 0.05). xyzValues within the same column with unlike superscripts differ (P < 0.05).

55
Chapter IV: Evaluation of semen extenders for short-term storage of ram semen at 4° C
Abstract
The objective of the study is to evaluate the effects of extender, ethylene glycol (EG), egg
yolk and penicillamine, hypotaurine and epinephrine (PHE) on ram sperm progressive motility
during storage. Semen collected from 9 mature rams by electroejaculation was distributed across
treatments consisting of either TRIS or milk extenders supplemented with 5 or 20% (v/v) egg
yolk, ± EG, and ± PHE. For each semen collection, TRIS extender was prepared from 4×
solution so that TRIS, citric acid and fructose concentration were constant at 300, 94.7, 27.8
mM, respectively, regardless of semen dilution factor. A 4× milk extender was also used so that
the extender contained 10% (w/v) milk powder, regardless of semen dilution factor. Semen was
diluted in extender to a final concentration of 300 million sperm/ml in 1.5 ml tubes, and cooled
to 4° C over a 2 to 3 h period. Semen was evaluated initially and daily for 3 d, using computer
assisted sperm analysis. Data were analyzed using the PROC MIXED procedure of SAS, with
ram as the experimental unit. Variables in the model included extenders, supplements, hour
(repeated measures) and interactions. Milk extender maintained better motility parameters over
time compared with TRIS (P < 0.05). Addition of either 5 or 20% egg yolk in the extender had
no effect on motility parameters (P > 0.05). However, milk with 20% egg yolk had the highest
track speed and path velocity followed by TRIS with 20% egg yolk, TRIS with 5% egg yolk, and
milk with 5% egg yolk, respectively (P < 0.05). There was an EG × hour interaction. At 72 h of
storage there was a rapid decline in sperm motility parameters in extenders without EG (P <
0.05). Addition of PHE before semen analysis increased progressive motility (P < 0.05), beat
frequency (P < 0.05), and tended to increase total motility (P = 0.058) and straightness (P =

56
0.078) of sperm cells. Results indicated that milk extender supplemented with 1% EG, PHE and
either 5 or 20% egg yolk can be used to protect ram semen at 4° C for 72 h.
Keywords: cold-storage, ethylene glycol, PHE, ram semen
Introduction
Typically, cryopreservation of ram semen results in poor viability post-thaw (Salamon
and Maxwell, 1995; Salamon and Maxwell, 2000). An alternative to cryopreservation is
temporary storage of ram semen at 4° C (Quan et al., 2016). Milk and TRIS extenders with 5%
egg yolk are commonly used for liquid storage of ram semen (Salamon and Maxwell, 2000;
Camara et al., 2011). However, preliminary studies found that progressive motility of ram sperm
declined up to 75% when stored at 4° C for 24 h, and continued to decline over time when using
extenders supplemented with 5% egg yolk (Acharya et al., 2016). Inclusion of egg yolk in the
extender protects ram sperm cells during cold storage of ram semen (Gil et al., 2011). The
literature is void of studies on optimal percentage of egg yolk to maintain ram sperm motility for
cool storage. If a majority of reduction in progressive motility occurs during the initial cooling
and storage of ram semen, then additional egg yolk and cryoprotectant prior to cooling, and
addition of supplements before evaluation might maintain or improve progressive motility.
Penicillamine, hypotaurine, and epinephrine (PHE) have different mode of action, but
collectively enhance sperm motility, sperm capacitation, and acrosomal reaction (Meizel, 1985).
Addition of PHE in the extender decreased time interval for oocyte penetration by sperm (Susko-
Parrish, et al., 1990). Frozen thawed and cultured bull semen in semen extender with PHE
increased most sperm motility parameters compared with absence of PHE (Pearson et al., 2007).
To our understanding there are no studies which compared the effect of addition of PHE on
sperm motility parameters after cool storage of ram semen.

57
Ethylene glycol (EG) was initially used as a cryoprotectant of choice for freezing
embryos and freeze-storage of ram semen (Bautista et al., 1997). Ethylene glycol causes the least
osmotic stress to sperm cells during cryopreservation compared with other cryoprotectants (Gao
et al., 1997; Ball et al., 2001). Addition of 1.6% EG in the extender medium to extend Chincila
lanigera semen increased sperm motility after storage for 72 h at 4° C compared with no EG
added. Addition of EG in the extender medium for cold storage of ram semen has yet to be
tested. The objective of this study was to evaluate the effects of different combinations of EG,
egg yolk, and PHE in milk or TRIS extenders on ram sperm progressive motility during storage
at 4° C for 72 h.
Materials and methods
The trial was conducted at the University of Arkansas, Fayetteville. All animal
procedures were approved by Institutional Animal Care and Use Committee (Protocol # 15019).
Animal management
Seven Katahdin and 2 Suffolk rams greater than 1 year of age were used for the study.
All rams were clinically healthy and free of internal and external parasites. Katahdin rams were
raised on tall fescue (Schedonorus arundinacea) pasture, whereas Suffolks rams were maintained
on mixed pasture containing white ladino clover (Trifolium repens) and bermudagrass
(Cyanodon dactylon). Animals had free access to drinking water, and mineral mixture and
additionally supplemented with 0.5 kg of sheep ration (17% CP, 82% TDN on DM basis)/ram/d.
Semen was collected by electroejaculation in 15 ml conical polystyrene tubes (Scientific USA,
Orlando, FL), screw capped and placed immediately into a thermo-flask (Lab-line Instruments
Inc., Melrose Park, IL) with water maintained at 30° C. Semen samples were transported to the

58
laboratory for analysis. All semen samples were evaluated within 2 h post collection using
computer assisted sperm analysis (CASA).
Evaluation of fresh semen by computer assisted sperm analysis (CASA)
All semen analysis and extension was conducted in a culture room maintained at 30° C.
A CASA system (Hamilton-Thorne, IVOS, Beverly MA) with 12 TOX IVOS software was used
for semen analysis. Sperm motility parameters were analyzed using default analysis setting
recommended by the manufacturer for ram semen. For analysis, semen samples were diluted in
TRIS extender (without egg yolk) at 1:100 (semen: extender) and 3.5 µl of diluted sample was
placed in a disposable 20 µm deep 2-chambered IVOS slide (Cytonix, Beltsville, MD). At least
four fields were scanned, with 30 video frames captured per field at a frame rate of 60 Hz. A
minimum of 400 sperm cells were counted for each sample analysis. Variables measured
included motility (%), progressive motility (%), track speed (VCL, µm/sec), path velocity (VAP,
µm/sec), progressive velocity (VSL, µm/sec), beat frequency (BCF, MHz), lateral amplitude
(ALH, µm), linearity (LIN, %), and straightness (STR, %).
Experimental treatments
Semen samples with less than 40% total motility were excluded. Semen was distributed
over 8 different treatments, 1) milk + 5% (v/v) egg yolk; 2) milk + 20% (v/v) egg yolk; 3) milk +
5% (v/v) egg yolk + 1% EG; 4) milk + 20% (v/v) egg yolk + 1% EG; 5) TRIS + 5% (v/v) egg
yolk; 6) TRIS + 20% (v/v) egg yolk; 7) TRIS + 5% (v/v) egg yolk + 1% EG; 8) TRIS + 20%
(v/v) egg yolk + 1% EG. Semen was extended at a rate of 300 million sperm cells/ml and stored
in 1.5 ml Eppendorf tubes. Fructose and antibiotics were added at the time of preparation of 4×
milk or 4× TRIS citrate extenders. The PHE 40 µl/1000 µl of extended semen was added 5 min
before evaluation.

59
Extenders
Milk and TRIS extender
For the 4× milk extender, skim milk powder (40g; NOW Real Food®, organic, non-fat
dry milk powder) was dissolved in 100 ml of purified water (Milli-Q) in a 250-ml beaker by
constantly stirring heating to 95° C for 10 min (VWR, West Chester, PA, model 371 hot plate
stirrers). Once cooled, 2 g of fructose (Sigma Chemical Co, St. Louis, MO) and 2 ml of
gentamicin reagent (10 mg/ml; Life Technologies, Gibco®, Waltham, MA) were added.
To prepare the 4× TRIS extender, tris[Hydroxymethyl]aminomethane (TRIS; 14.5 g;
Sigma Chemical Co, St. Louis, MO) and citric acid (8.0g; Sigma Chemical Co, St. Louis, MO)
were dissolved in 100 ml water in a 250-ml beaker. Fructose (2 g; Sigma Chemical Co, St.
Louis, MO) and 2 ml of gentamicin reagent (10 mg/ml; Life Technologies, Gibco®, Waltham,
MA) were added. The stock solution was filtered (Gelman 0.20 µm sterile Acrodisk filter; Sigma
Chemical Co, St. Louis, MO). Once the dilution factor of fresh semen was determined, egg yolk
(5 or 20% v/v), ethylene glycol, and purified water was added to 4× milk or TRIS and mixed
homogenously at 30° C for extension of fresh semen (maintained at 30° C).
Determination of semen extension rate
The concentration of sperm cells in a fresh semen sample determined by CASA analysis
was multiplied by the dilution factor and volume of ejaculate. Semen was extended to 150
million cells/0.25 ml. Each semen sample was diluted with each extender combination.
Semen extension using extenders
A 4× concentration of milk (constituted of milk, fructose, and gentamycin) or TRIS
extender (constituted of TRIS, citric acid, fructose, and gentamycin) was prepared at first. After
determination of semen concentration, the appropriate amount of egg yolk, EG, and purified

60
(Milli-Q) water, and 4× milk or TRIS was added to known concentration of semen. Prepared
extender was added to semen. Each time semen was extended to the constant concentration (300
million sperm cells/ml). For each semen extension, TRIS extender was prepared from 4×
solution so that the TRIS, citric acid and fructose concentration were constant 300, 94.7, 27.8
mM, respectively, regardless of semen dilution factor. A 4× milk extender was also used so that
the extender contained 10% (w/v) milk powder, regardless of semen dilution factor. Both
extenders were supplemented with 50 µg/ml of gentamicin. Egg yolk (5 or 20% v/v) and ± 1%
EG was added at the time of extension. Extended semen samples were stored in 1.5 ml
Eppendorf tubes and gradually cooled to 4° C in a laboratory refrigerator. Semen was cooled to
4° C over a 3 h period. Cooling rate was recorded using Physitemp Model BAT-12 temperature
recorder. Slow cooling rate was achieved by putting Eppendorf tubes in 300 ml laboratory
beaker containing water at 30° C and placing it into a refrigerator.
Preparation of PHE
A penicillamine, hypotaurine and epinephrine (PHE) stock solution was prepared by
adding 5 ml penicillamine stock (2.98 g in 10 ml of 0.8% saline), 5 ml hypotaurine stock (1.09
mg in 0.8% saline) and 4 ml epinephrine stock (165 mg 60% sodium lactate and 50 mg sodium
meta-bisulfite in 50 ml of water; pH was adjusted to 4.0 with 1 N HCL; 1.83 mg epinephrine was
added to 40 ml of this solution in 6 ml 0.8% saline). The final solution was comprised of 20 µl
penicillamine, 10 µl hypotaurine, 2 µl epinephrine, 21 µl sodium meta-bisulfite and 118 µl
sodium lactate. The PHE stock solution was stored in 1.0 ml cryogenic vials (Vangard Cryos,
Sumitomo Bakelite Co., Ltd., Japan) at -20 °C. On the day of semen analysis, 12.5 µl of thawed
PHE stock solution was mixed with 315 µl of diluted semen samples (300 µl of 1× TRIS citrate

61
extender + 15 µl of extended semen) immediately before CASA analysis. Note: the procedure for
PHE preparation was adopted from Rick Rorie (personal communication).
Evaluation of extended semen
Extended semen was evaluated by IDENT fluorescence method every 24 h up to 72 h
(CASA setting; Table 1). Ident stain (Hoechst 33342) was used to illuminate DNA containing
objects. An aliquot of 15 µl of chilled semen was diluted with pre-warmed 300 µl of 1× TRIS
extender (at 35° C) in a 1.5 ml Eppendorf tube. About 12.5 µl of PHE was added to the PHE
treatment samples. The amount of PHE required to stain a given volume of extended semen was
described previously (rate of 40 µl of PHE/ml of diluted semen; Miller et al., 1994; Pearson et
al., 2007). Diluted semen sample was stained (Hoechst 33342) before analysis (IVOS IDENT
illumination option). About 5 µl of stain (10 µg/ml) was added to the diluted semen and
maintained in a heating block at 35° C for 15 min before analysis. For milk egg yolk extender,
10 µl of stain was added. Samples were warmed in a digital heatblock (VWR International,
USA) for 15 min at 35° C. After 15 min of equilibration with stain at 35° C, samples were
analyzed using CASA. About 3.5 µL of extended semen was placed in a disposable 2-chamber,
20 µm deep IVOS slide (Cytonix, Beltsville, MD). At least four fields were scanned, with 30
video frames captured per field at a frame rate of 60 Hz. A minimum of 400 sperm cells were
counted in each sample analysis. Variables measured were total motility, progressive motility,
VCL, VAP, VSL, BCF, ALH, LIN, and STR (Table 2).
Statistical analysis
Data were analyzed as repeated measures using MIXED procedure of SAS (Littell et al.,
1996; SAS Institute, Carey, NC) with ram as the experimental unit. Variables in the model
included extenders (milk or TRIS), 5 or 20% egg yolk, 0 or 1% EG, and 0 or 40 µl PHE/1000 µl

62
of diluted semen and all 2, and 3 way interactions. Non-significant (P > 0.1) interactions were
removed from the model (P > 0.1). The final model included extender, egg yolk, EG, PHE,
ethylene glycol × hours, ethylene glycol × extender, and egg yolk × extender interactions.
An autoregressive covariance structure was used. Data are presented as least square means and
standard error. Means were separated using the PDIFF option of SAS. If complex interactions
were not significant they were removed. Variables analyzed using CASA were motility,
progressive motility, VCL, VAP, VSL, BCF, ALH, LIN, and STR.
Results
Total Motility of fresh sperm cell was 67%. Motility declined to 53.1 ± 1.6, 46.2 ± 1.6,
and 40.7 ± 1.6% after 24, 48, and 72 h after extension and storage (P < 0.01). Motility was
higher with milk compared with TRIS extender (P < 0.01; Table 3). Supplementation of either 5
or 20% egg yolk resulted in similar total motility (P = 0.463; Table 4). Supplementation of EG
increased total motility compared to no addition (P < 0.01; Table 5). Addition of PHE before
semen analysis tended to increase (P < 0.0589; Table 6) total motility compared with no PHE
addition. There was an EG × hour interaction (P < 0.01; Table 7) due to decreased sperm
motility without EG at 72 h.
Progressive motility of fresh sperm cells was 38.6% and declined to 24.4 ± 0.9, 20.1 ±
0.9, and 17.1 ± 0.9 after 24, 48, and 72 h of storage (P < 0.01). There was a trend toward an
extender × EG interaction (P = 0.0793) in that semen extended in milk supplemented with EG
had higher progressive motility compared with TRIS with EG or without EG (P < 0.05; Table 8).
Semen extended using milk + EG tended to have higher sperm progressive motility compared
with milk without EG (P = 0.055). There was an EG × hour interaction (P < 0.05) mainly due to
decreased progressive motility in extenders without EG at 72 h of storage. Sperm progressive

63
motility was similar between extenders with 5 or 20% egg yolk (P = 0.721). Addition of PHE
before semen analysis increased (P < 0.01) sperm progressive motility compared with no PHE
addition.
Track speed of fresh sperm cells was 202.8 µm/sec. Track speed declined to 192.5 ± 5.2,
186.8 ± 5.2, and 174.0 ± 5.2 µm/sec after 24, 48, and 72 h of storage (P < 0.05). There was a
trend toward extender × egg yolk-percentage interaction (P = 0.069; Table 9) on VCL. Storage
of semen in milk + 20% egg yolk had higher VCL compared with milk + 5% egg yolk (P <
0.05), but, was similar between TRIS + 5% egg yolk and TRIS + 20 % egg yolk (P > 0.05).
There was an EG × hour interaction (P = 0.036) mainly due to decreased VCL in extenders
without EG at 72 h of storage. Addition of PHE before semen analysis had no effect on VCL
compared with no PHE addition (P = 0.161).
Path velocity of fresh sperm cells was 130.4 µm/sec. Path velocity declined to 106.3 ±
2.4, 101.5 ± 2.3, and 92.1 ± 2.3 µm/sec after 24, 48, and 72 h of storage (P < 0.05). There was a
trend toward a significance interaction for VAP observed in an extender × egg yolk-percentage
(P = 0.056). Milk + 20% egg yolk had higher VAP compared with milk + 5% egg yolk, but VAP
was similar between TRIS + 5 or 20% egg yolk (P < 0.05). There was an EG × hour interaction
(P < 0.01) mainly due to a decrease in VAP in extenders without EG at 72 h of storage. VAP
was similar whether PHE was added or not before semen analysis (P = 0.276).
Progressive velocity of fresh sperm cells was 109.6µm/sec. Progressive velocity declined
to 82.1 ± 1.7, 78.0 ± 1.7, and 70.7 ± 1.7 µm/sec after 24, 48, and 72 h of storage, respectively (P
< 0.01). An extender × EG interaction was seen on sperm VSL (P < 0.01). Semen stored in milk
with or without EG had a higher VSL compared with TRIS with or without EG (P < 0.05), but
TRIS supplemented with EG had higher VSL compared with TRIS without EG (P < 0.05).

64
There was an EG × hour interaction on sperm VSL mainly due to decreased VSL in extenders
without EG after 72 h of storage (P < 0.01). The concentration of egg yolk (P = 0.163) or PHE
had no effect on VSL (P = 0.541).
Beat frequency of fresh sperm cells was 38.7 MHz. Beat frequency at 24, 48, and 72 h
was 31.9 ± 0.7, 31.0 ± 0.7, and 32 ± 0.7 MHz, respectively (P = 0.614). An extender × EG
interaction was observed on sperm BCF (P < 0.05). Semen extended using milk with or without
EG, and TRIS + EG had similar BCF (P > 0.05), which were higher than TRIS without EG (P <
0.05). BCF was similar between extenders with 5 and 20% egg yolk (P = 0.163). Addition of
PHE compared with no PHE before semen analysis increased BCF (P < 0.01).
Linearity of fresh sperm cells was 55%. Linearity of sperm cells declined to 47 ± 0.9, 45
± 0.9, and 40 ± 0.9% at 24, 48, and 72 h, respectively (P < 0.01). An extender × EG interaction
was observed on LIN (P = 0.006). Linearity of sperm cells was similar after semen storage in
milk with or without EG and TRIS EG extenders (P > 0.05), which were higher compared with
TRIS without EG (P < 0.05). An EG × hour interaction was observed on LIN, mainly due to
decreased LIN of sperm stored in EG non-supplemented extenders at 72 h of storage (P =
0.043). There was no difference in LIN between extenders with 5 or 20% egg yolk (P = 0.185).
Addition of PHE before semen analysis did not influence LIN compared with no PHE addition
(P = 0.267).
Lateral amplitude (ALH) of fresh sperm cells was 6.37 µm. Lateral amplitude of sperm
cells at 24, 48, and 72 h was 7.26 ± 0.1, 7.39 ± 0.1, and 6.92 ± 0.1µm respectively (P = 0.2374).
There was a trend toward an interaction between EG × hour (P < 0.092), mainly due to reduced
ALH of sperm cells stored without EG at 72 h (P < 0.059). Lateral amplitude was higher in
TRIS extender compared with milk (P < 0.01). Sperm ALH was similar between extenders with

65
5 and 20% egg yolk (P = 0.231). Similarly, addition of PHE before semen analysis had no effect
on ALH (P = 0.267).
Straightness of fresh sperm cells was 81%. Straightness of sperm cells declined to 77 ±
1.4, 75 ± 1.4, and 70 ± 1.4% at 24, 48, and 72 h, respectively (P < 0.01). An extender × EG
interaction was observed on STR (P < 0.01) in that TRIS without EG was lower than that of
milk with or without EG and TRIS + EG (P > 0.05). An EG × hour interaction was observed on
sperm STR mainly due to decreased STR in EG non-supplemented extenders at 72 h of storage
(P = 0.027). Straightness was similar in extenders with 5 or 20% egg yolk (P = 0.231). Addition
of PHE before semen analysis tended to increase sperm STR compared with no PHE addition (P
= 0.078).
Discussion
Ram semen extended and stored at 4° C using milk or TRIS maintained better sperm
motility compared with TEST and CJ-2 extender (Acharya et al., 2017; unpublished data).
Storage at 4 maintained better sperm motility parameters compared with storage at 15° C.
(Acharya et al., 2017; unpublished data). Although milk or TRIS extenders maintained better
motility compared with TEST and CJ-2 extenders at 4° C, the overall progressive motility
declined ~ 75% within 24 h of storage and continued to decline over time using extenders
supplemented with 5% egg yolk. If the majority of reduction in progressive motility occurs
during the initial cooling and storage of ram semen, then additional egg yolk and cryoprotectant
at the time of extension or supplementation of PHE before evaluation might maintain or improve
motility of ram semen. In the current study, milk extender maintained better sperm motility
parameters over time compared with TRIS extender. Addition of either 5 or 20% egg yolk in
extenders had no effect on sperm motility parameters. Addition of EG in the extender before

66
storage increased total motility, progressive motility, and BCF of sperm cells compared with no
addition. Addition of PHE before semen analysis increased VSL and BCF, and tended to
increase total motility and STR of sperm cells compared with no PHE addition.
Several studies have been reported on sperm quality between were done in the past to
compare milk and TRIS extenders for extension and chilled storage of ram semen, but with
inconsistent results. Some authors reported superiority of milk over TRIS (Hollinshead et al.,
2004), while others reported TRIS maintained better motility over milk (Quan et al., 2016). Such
inconsistent results could be due to differences in supplements in the extender, storage volume,
storage durations, concentration of sperm cell in the extender, and use of sorted or non-sorted
semen. For example, Quan et al. (2016) used 15 ml extended semen to store in 15 ml plastic vials
at a dilution rate 100 million spermatozoa/ml, while Hollinshead et al. (2004) used sorted semen
for extension and cold storage. Sorted semen was extended at 6 million spermatozoa in 0.5 ml
extender to store in 5 ml sterile sealed tubes to evaluate after 6 h of storage at 4° C (Hollinshead
et al., 2004). Therefore, none of the studies can be used for generalization of superiority of one
extender over other (milk or TRIS). In the current study, ram semen was extended to a
concentration of 300 million sperm cells/ml to match with a standard of 150 million sperm
cells/0.5 ml insemination dose. TRIS, citric acid, and fructose concentrations in the medium was
constant, regardless of semen dilution factor. Similarly, milk extender contained constant amount
of 10% (w/v) milk powder regardless of semen dilution factor. Extended semen was kept in 1.5
ml Eppendorf tubes to store in 300 ml beaker filled with water at 30° C before storage to achieve
slow cooling rate. Semen was evaluated every 24 h up to 72 h of storage at 4° C. Milk
maintained higher sperm motility parameters compared with TRIS extender and with the

67
addition of cryoprotectant and supplements, progressive motility of sperm cell was ~ 60% of the
initial value after storage at 4° C for 72 h.
During the process of cooling extended semen for cold storage, massive rearrangements
and leakage of sperm plasma membrane occurs which could lead to sperm cell death (Holt and
North, 1984). Inclusion of egg yolk in the culture medium protects ram spermatozoa from cell
death (Gil et al., 2011). It is still unclear about the components of egg yolk and mechanism for
protection of ram sperm cells. Inclusion of TRIS + 20% egg yolk better protected sperm cells
against cold shock compared with inclusion of TRIS + 15% egg yolk, and TRIS + 10% egg yolk
(Rajabi-Toustani et al., 2014). Similarly, Lopez et al. (1999) reported better sperm motility after
storage in TRIS or sodium citrate extender with 20% compared with milk and 5.5% egg yolk.
However, the study was confounded as different extenders and different egg yolk percentages
were used (Lopez et al., 1999). In the current study, milk or TRIS extenders were compared
using either 5 or 20% egg yolk in extender medium. There was no difference in motility
parameters using 5 or 20% (v/v) of egg yolk in either of the extender. However, inclusion of
20% egg yolk in milk extender led to a higher VCL and VAP, followed by TRIS + 20% egg
yolk, TRIS + 5% egg yolk and milk 5% egg yolk.
Ethylene glycol is a cryoprotectant of choice for freezing embryos because of its low
molecular weight, which enables rapid influx into cells during equilibration and rapid efflux after
thawing (Bautista et al., 1997). Similar effects of ethylene glycol occurs across plasma
membrane of sperm cells (Hammerstedt and Graham, 1992). Ethylene glycol at 2.5%
concentration in TEST extender effectively protected ram semen stored for 3 h at 4° C compared
with DMSO, propylene glycol, dimethylacetamide, and glycerol at 2.5, 5, and 10 %
concentrations (Vasquez et al., 2013). Inclusion of 1.6% EG in TEST extender was beneficial for

68
extension and storage of Chincila lanigera semen at 4° C for 72 h. Use of EG for extension and
chill storage of ram semen is not available. In the current study, addition of 1% EG in the
extender for extension and storage of ram semen at 4° C for 72 h increased most sperm motility
parameters compared with no EG addition. An EG × hour interaction also occurred in most
sperm motility parameters. Such interactions are due to greater reduction of sperm motility
parameters at 72 h of storage in extenders without EG.
Conception rate was compared using fresh or cool stored ram semen during August in
Midwestern US. Semen was extended using milk extender (with 20% egg yolk and 1% EG) and
inseminated immediately after extension (n = 27) or stored at 4° C for up to 36 h before
insemination (n = 28). The conception rate in ewes inseminated using stored semen was 31%
while fresh extension and insemination resulted 37% conception rate.
Penicillamine, hypotaurine, and epinephrine have different modes of action but
collectively enhance sperm motility, sperm capacitation, and acrosomal reaction (Meizel, 1985).
Addition of PHE in the medium decreased time interval for oocyte penetration by sperm (Susko-
Parrish, et al., 1990). Frozen thawed and cultured bull semen in PHE-added medium increased
most sperm motility parameters compared with no PHE (Pearson et al., 2007). Studies reporting
the effects on PHE on sperm motility parameters of extended ram semen are rare. In the current
study, addition of PHE before semen analysis increased progressive motility and BCF of sperm
cells and tended to increase total motility and STR of sperm cells. In a separate study, we
compared effect of addition or no addition of PHE (40 µl PHE/ 1000 µl extender) in milk
extender before AI. A total of 27 ewes were artificially inseminated with extended, and cold
stored ram semen for 12 to 24 h at 4° C. The PHE was added in extended semen 5 min before AI

69
in PHE group (n = 14) or no PHE addition in control group (n = 13). The conception rate in the
PHE added group was 50% while only 23% conceived in the no PHE group.
In conclusion, milk extender maintained better sperm motility compared with TRIS after
storage at 4° C for up to 72 h. Percentage of egg yolk included in extenders had no effect on
most of the sperm motility parameters. However, inclusion of 20% egg yolk in milk extender led
to a higher VCL and VAP followed by TRIS + 20% egg yolk, TRIS + 5% egg yolk, and milk +
5% egg yolk. At 72 h of storage, sperm cell motility declined sharply in extenders without EG.
Results indicate that milk extender supplemented with 1% EG, 20% egg yolk before storage, and
addition of PHE before analysis is beneficial for liquid storage of ram semen.

70
References
Acharya, M., J. M. Burke, E. Smyth, A. Davis, T. D. Lester, and R. Rorie. 2016. Effect of semen
extender and storage temperature on ram sperm motility over time. J. Anim. Sci.
94(suppl. 1): 53 (Abstr.).
Ball, B. A., A. T. Vo, and J. Baumber. 2001. Generation of reactive oxygen species by equine
spermatozoa. Am. J. Vet. Res. 62:508-515.
Bautista, J. A. N., Takahashi, Y., and H. Kanagawa. 1997. In vitro viability of mouse 8-cell
embryos vitrified in a simple solution of ethylene glycol. Jpn. J. Vet. Res. 45:67-73.
Camara, D. R., M. M. C. Mello-Pinto, L. C. Pinto, O. O. Brasil, J. F. Nunes, and M. M. P.
Guerra. 2011. Effect of reduced glutathione and catalase on the kinematics and
membrane functionality of sperm during liquid storage of ram semen. Small Rum. Res.
100: 44-49.
Desnoyers, L., and P. Manjunath. 1992. Major proteins of bovine seminal plasma exhibit novel
interactions with phospholipid. J. Biol. Chem. 267:10149-10155.
Gao D. Y., P. Mazur, and J. K. Critser. 1997. Fundamental cryobiology of mammalian
spermatozoa. In: Karow A. M., J. K. Critser (eds.) Reproductive tissue banking.
Academic press, San Diego pp 263-327.
Gil, J., S. Fierro, O. Bentancur, and J. Olivera‐Muzante. 2011. Chilled storage of ram semen
improves with the addition of egg yolk and glycerol to milk‐based extenders. Reprod.
Domest. Anim. 46:503-507.
Hammerstedt, R. H., and J. K. Graham. 1992. Cryopreservation of poultry sperm: the enigma of
glycerol. 29:26-38.
Hollinshead, F. K., J. K. O’Brien, L. Gillan, M. Meyers, W. Maxwell, and G. Evans. 2004.
Liquid storage of flow cytometrically sorted ram spermatozoa. Theriogenology. 62:587-
605.
Littell, R. C., G. A. Milliken, W. W. Stroup, and R. D. Wolfinger. 1996. SAS system for Mixed
Models. SAS Institute Inc., Cary, NC. 633 pp.
López, A., L. Söderquist, and H. Rodriguez-Martinez. 1999. Sperm viability in ram semen
diluted and stored in three different extenders. Acta Vet. Scand. 40:1-9.
MacDonald, B., and J. Foulkes. 1981. A spectrofluorometric investigation, using 1-anilino-
naphthalene-8-sulphonate, of the interaction between washed bovine spermatozoa and
seminal plasma or egg-yolk lipoprotein. J. Reprod. Fertil. 63:407-414.

71
Manjunath, P., L. Baillargeon, Y. L. Marcel, N. G. Seidah, M. Chrétien, and A. Chapdelaine.
1988. Diversity of novel proteins in gonadal fluids. In M. Chrétien., K. W. McKerns
(Eds.), Molecular biology of brain and endocrine peptidergic system. Springer US. pp.
259-273.
Meizel, S. 1985. Molecules that initiate or help stimulate the acrosome reaction by their
interaction with the mammalian sperm surface. Dev. Dynam. 174:285-302.
Miller, G. F., D. W. Gliedt, J. M. Rakes, and R. W. Rorie. 1994. Addition of penicillamine,
hypotaurine and epinephrine (PHE) or bovine oviductal epithelial cells (BOEC) alone or
in combination to bovine in vitro fertilization medium increases the subsequent embryo
cleavage rate. Theriogenology. 41:689-696.
Pace, M., and E. Graham. 1974. Components in egg yolk which protect bovine spermatozoa
during freezing. J. Anim. Sci. 39:1144-1149.
Person, C. N., T. D. Lester, M. D. Person, and R. W. Rorie. 2007. Effects of penicillamine,
hypotaurine and epinephrine (PHE) on post-thaw bovine sperm parameters, as measured
by computer-assisted sperm analysis. AAES Research Series. 553:47-50.
Quan, G. B., G. Q. Wu, Y. J. Wang, D. J. Li, Y. Ma, Q. H. Hong. 2016. Effects of the Tris, Tes,
or skim milk based extender on in vitro parameters of ram spermatozoa during liquid
storage. Small Rumin. Res. 134:14-21.
Rajabi-Toustani, R., M. Roostaei-Ali, and R. Motamedi-Mojdehi. 2014. Effect of different levels
of egg yolk on ram sperm coating and preserving at 5° C. Iran. J. Vet. Res. 15:168-171.
Ricker, J. V., J. J. Linfor, W. J. Delfino, P. Kysar, E. L. Scholtz, F. Tablin, J. H. Crowe, B. A.
Ball, and S. A. Meyers. 2006. Equine sperm membrane phase behavior: The effects of
lipid-based cryoprotectants 1. Biol. reprod. 74:359-365.
Salamon, S., and W. M. C. Maxwell. 1995. Frozen storage of ram semen. I. Processing, freezing,
thawing and fertility after cervical insemination. Anim. Reprod. Sci. 37:189-249.
Salamon, S., and W. M. C. Maxwell. 2000. Storage of ram semen. Anim. Reprod. Sci. 62:77-
111.
Susko-Parrish, J. L., M. B. Wheeler, R. L. Ax, N. L. First, and J. J. Parrish. 1990. The effect of
penicillamine, hypotaurine, epinephrine and sodium metabisulfite, on bovine in vitro
fertilization. Theriogenology. 33:333 (Abstr.).
Vásquez, J. H., V. H. Núñez, E. A. Florentini, J. M. Gonzales, L. A. Camargo, and M. E.
Valdivia. 2013. Effects of five cryoprotective agents on quality of sheep epididymal
spermatozoa during pre-freezing. Livest. Sci. 152: 94-9

72
Table 1. IVOS machine settings for ram semen motility analysis
Parameter Setting
Frames per second 60
Number of frames 30
Minimum contrast 60
Minimum size 5
Default cell size 5
Default cell intensity 55
Path velocity (VAP) 75.0
Straightness (STR) 80
Slow cells Static
VAP cut off 21.9
VSL cut off 6.0
Minimum intensity gate 0.25
Maximum intensity gate 1.50
Minimum size gate 0.60
Maximum size gate 8.00
Minimum elongation gate 0
Maximum elongation gate 95
UV fluorescent objective 10×
Target photometer 45-55
Field type Dark
Chamber depth 20

73
Table 2. CASA parameters
Total motility (%) Path velocity ≥ 30 µm/sec and progressive velocity ≥ 15
µm/sec
Progressive motility (%) Path velocity ≥ 50 µm/sec and straightness ≥ 70%
Track speed (VCL, µm/sec) Average velocity measured over the actual point-to-point
track.
Path velocity (VAP, µm/sec) Average velocity of the smoothed (best fit) sperm path.
Progressive velocity (VSL,
µm/sec)
Average velocity over straight line from the beginning to the
end of the sperm track.
Beat frequency (BCF, MHz) Frequency of sperm head crossing the sperm average path in
either direction.
Lateral amplitude (ALH, µm) Mean width of the head oscillation as the sperm swims.
Linearity (LIN, %) Departure of actual sperm path from straight line
(VSL/VCL).
Straightness (STR, %) Departure of average sperm path from a straight line
(VSL/VAP).

74
Table 3. Sperm motility parameters after extension using milk or TRIS and stored at 4° C for 72
h.
Results are expressed as least square means and standard error (SE).
Significant difference was declared at P < 0.05.
Extenders
Sperm characteristics Milk TRIS SE P-value
Total motility 49.7 43.6 1.3 0.0013
Progressive motility 24.5 16.6 0.7 < 0.0001
Track speed 185.6 182.5 4.2 0.60
Path velocity 104.7 95.1 1.9 0.0006
Progressive velocity 83.3 70.6 1.4 < 0.0001
Beat frequency 32.9 30.3 0.6 0.002
Lateral amplitude 6.8 7.4 0.2 0.007
Linearity 45.9 41.6 0.9 0.0009
Straightness 75.5 72.5 1.2 0.07
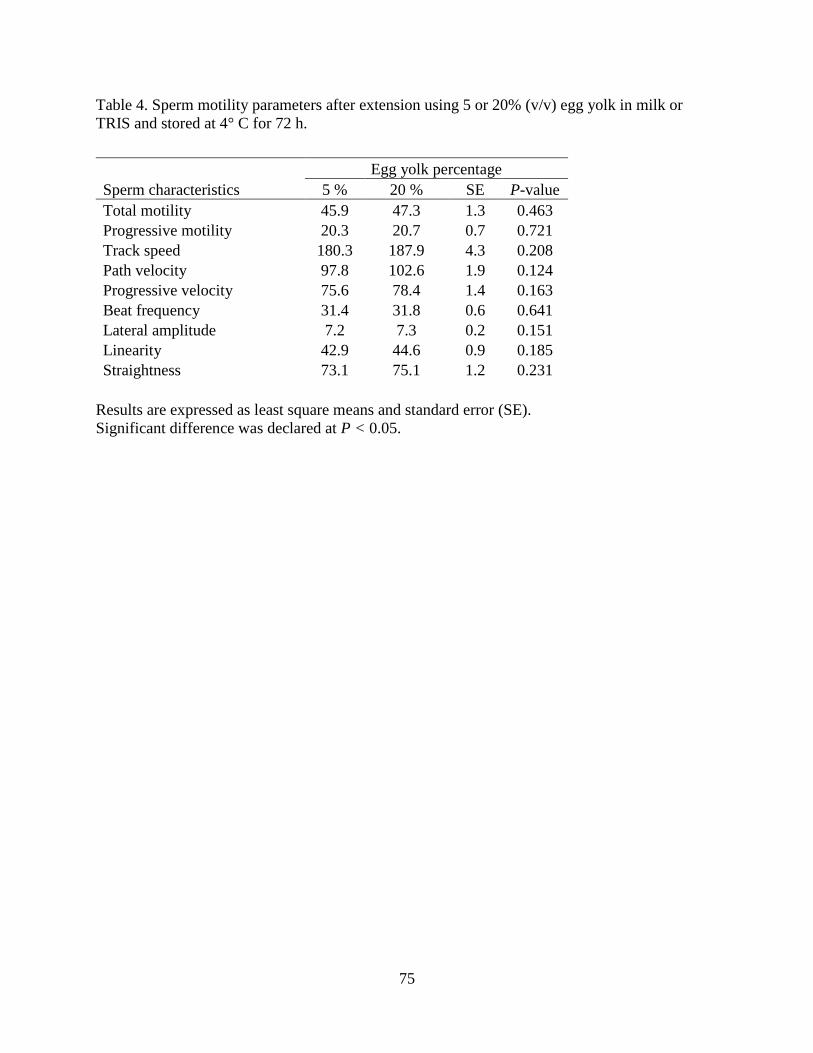
75
Table 4. Sperm motility parameters after extension using 5 or 20% (v/v) egg yolk in milk or
TRIS and stored at 4° C for 72 h.
Results are expressed as least square means and standard error (SE).
Significant difference was declared at P < 0.05.
Egg yolk percentage
Sperm characteristics 5 % 20 % SE P-value
Total motility 45.9 47.3 1.3 0.463
Progressive motility 20.3 20.7 0.7 0.721
Track speed 180.3 187.9 4.3 0.208
Path velocity 97.8 102.6 1.9 0.124
Progressive velocity 75.6 78.4 1.4 0.163
Beat frequency 31.4 31.8 0.6 0.641
Lateral amplitude 7.2 7.3 0.2 0.151
Linearity 42.9 44.6 0.9 0.185
Straightness 73.1 75.1 1.2 0.231

76
Table 5. Sperm motility parameters after extending with or without ethylene glycol in milk or
TRIS and stored at 4° C for 72 h.
Results are expressed as least square means and standard error (SE).
Significant difference was declared at P < 0.05.
Treatments
Sperm characteristics Ethylene glycol Control SE P-value
Total motility 51.2 42.1 1.3 < 0.0001
Progressive motility 23.1 18.1 0.7 < 0.0001
Track speed 184.5 183.6 4.2 0.873
Path velocity 100.2 99.6 1.9 0.826
Progressive velocity 77.5 76.4 1.4 0.589
Beat frequency 32.5 30.6 0.6 0.029
Lateral amplitude 7.2 7.1 0.2 0.831
Linearity 44.3 43.1 0.9 0.345
Straightness 75.5 72.5 1.17 0.06

77
Table 6. Effect of addition of penicillamine, hypotaurine, and epinephrine (PHE) on sperm
motility parameters before evaluation on cold stored ram semen, stored at 4° C for 72 h.
Results are expressed as least square means and standard error (SE).
Significant difference was declared at P < 0.05.
Treatments
Sperm characteristics PHE Control SE P-value
Total motility 48.4 44.8 1.3 0.058
Progressive motility 22.4 18.7 0.7 0.0007
Track speed 179.8 188.3 4.2 0.161
Path velocity 98.4 101.4 1.9 0.276
Progressive velocity 77.5 76.3 1.4 0.541
Beat frequency 32.9 30.2 0.6 0.001
Lateral amplitude 7.1 7.4 0.2 0.152
Linearity 44.5 43.1 0.9 0.267
Straightness 75.5 72.5 1.1 0.078

78
Table 7. Sperm motility parameters after extension with or without ethylene glycol in milk or
TRIS with 5 or 20% egg yolk and stored at 4° C for 72 h. Only significant ethylene glycol ×
hours interactions are expressed.
Results are expressed as least square means and standard error (SE). abValues within the same row followed by different superscripts are different (P < 0.05). xyzValues within the same column with unlike superscripts differ (P < 0.05).
Parameters Hour Ethylene glycol Control SE
Total motility 24 55.4a,x 50.9a,x 2.3
48 48.6a,y 43.4a,y 2.3
72 49.6a,xy 31.7b,z 2.3
Progressive motility 24 26.1a,x 23.1a,x 1.3
48 21.5a,y 18.7a,y 1.3
72 21.5a,y 12.6b,z 1.3
Track speed 24 186a,x 197a,x 7.4
48 182a,x 191a,x 7.4
72 185a,x 162b,y 7.4
Path velocity 24 103.3a,x 109.1a,x 3.3
48 98.4a,x 104.5a,x 3.3
72 99.1a,x 85.2b,y 3.3
Progressive velocity 24 80.3a,x 84.1a,x 2.4
48 75.7a,x 80.2a,x 2.4
72 76.4a,x 65.1b,y 2.4
Linearity 24 46.5a,x 47.2a,x 1.5
48 43.8a,x 45.2a,x 1.5
72 42.8a,x 37.1b,y 1.5
Straightness 24 76.5a,x 77.1a,x 2.1
48 75.1a,x 74.6a,x 2.1
72 72.1a,x 65.7b,y 2.1

79
Table 8. Sperm motility parameters after extension using with or without ethylene glycol (EG)
in milk or TRIS extender. Only significant ethylene glycol × extender interactions are expressed.
abcValues within the same row with unlike superscripts differ (P < 0.05).
Parameters Milk Milk + EG TRIS TRIS + EG SE
Progressive motility 23.8ab 25.9a 13.2c 20.1b 1.1
Progressive velocity 80.8a 85.7a 67.1c 74.1b 2.1
Beat frequency 33.1a 32.9a 28.3b 32.2a 0.8
Linearity 47.1a 44.7a 39.3b 44.1a 1.5
Straightness 76.7a 74.3a 68.2b 76.8a 2.3

80
Table 9. Sperm motility parameters after extension including 5 or 20% egg yolk in milk or TRIS
extender. Only significant egg yolk (EY) × extender interactions are expressed.
abValues within the same row with unlike superscripts differ (P < 0.05).
Parameters Milk, 5% EY Milk, 20% EY TRIS, 5% EY TRIS 20% EY SE
Track speed 176.3b 195.1a 184.2ab 180.8ab 7.4
Path velocity 99.9b 109.4a 95.6b 94.6b 2.7

81
Chapter V: Correlation between anti-Mullerian hormone and estimated breeding values
for reproductive performance in ewes
Abstract
Anti-Mullerian hormone (AMH) in circulation may reflect the total pool of follicles
within the ovaries, and may be positively correlated with fertility in ewes. The objective of this
study was to investigate any correlation between serum AMH concentrations, number of lambs
born, and estimated breeding values determined through the National Sheep Improvement
Program for reproductive performance (NLB or number of lambs born; NLW or number of
lambs weaned; MWWT or maternal weaning weight; and the Katahdin index) and WWT or
weaning weight. Additionally, AMH quartiles (AMH Q1 through Q4, with Q1 the lowest and Q4
the highest, pg/ml) were correlated with mean number of lambs born after fall versus summer
breeding. Blood samples were collected for serum analysis from 163 Katahdin ewes ranging
from 0.5 to 7-yr of age; their breeding and lambing records were obtained from flock data.
Correlation between AMH concentrations and EBVs were determined using PROC CORR and
means generated using least squares means procedure of SAS (SAS Inst. Inc., Cary, NC, USA).
For remaining data, continuous variables were analyzed by PROC GLM and categorical
variables were analyzed by chi-squared, using SAS. Mean serum AMH was 182 pg/ml and
ranged from 0 to 1112 pg/ml. There was no correlation between serum AMH and EBVs for NLB
(-0.04; P = 0.589), NLW (0.06; P = 0.401), MWWT (-0.01; P = 0.872), WWT (0.003; P =
0.964) and the Katahdin index (0.09; P = 0.251). Serum AMH concentration was similar in ewes
in different age category (less than 1, 1 to 2, 2 to 3, 3 to 4, and more than 4-yr of age; P > 0.05).
There was a correlation between EBV for NLB (0.29; P < 0.01) and NLW (0.19; P = 0.013)
with average number of lambs born. Ewes in a lowest AMH quartiles (Q1) had a lower mean
number of offspring born from summer breeding compared with ewes in other AMH quartiles

82
(Q2, Q3, and Q4; P < 0.05). Further study is needed to determine effectiveness of using serum
AMH for selecting ewes for summer (off-season) breeding.
Keywords: Anti-Mullerian hormone, ewes, estimated breeding values, reproductive performance
Introduction
Anti-Mullerian hormone (AMH) is a dimeric glycoprotein that belongs to a member of
the transforming growth factor-β (TGF-β) family (Cate et al. 1986). This hormone is produced
by granulosa cells of preantral and small antral follicles of 1-3 mm diameter (Bezard et al., 1987;
Monniaux et al. 2012). Anti-Mullerian hormone facilitates follicle recruitment and growth of
young follicles (Broer et al. 2014). However, when a follicle increases in size, AMH limits the
number of follicles developing from the gonadotropin responsive stage to the gonadotropin
dependent stage (Campbell, et al., 2012). Although AMH limits the number of ovulatory
follicles, studies have positively correlated plasma AMH with fertility in animals, including
sheep (Lahoz et al., 2012; Lahoz et al., 2016 a, b).
Individual ewes vary in reproductive performance. In the Katahdin breed, some ewe
lambs can lamb at 12-months of age, while others lamb later or may not lamb during their entire
life. Ewes also differ in their prolificacy ranging from singles to quadruplets or more
(Vanimisetti, et al., 2007). Similarly, their ability to conceive out-of-season also varies (Burke,
2005). Estimated breeding values (EBVs) are the mathematical predictions of the genetic merit
of an animal for specific traits. These values are calculated based on an individual animal’s
performance for a specific trait compared with animals within a contemporary group and animals
in other connected flocks. Estimated breeding values are useful to select for specific traits used in
breeding or commercial stock (www.nsip.org).

83
Studies correlating serum AMH concentration with EBV for reproductive performance
(NLB or number of lambs born; NLW or number of lambs weaned; MWWT or maternal
weaning weight; and the Katahdin index) and WWT or weaning weight have not been reported
in ewes. Therefore, this study investigated any correlations between serum AMH, mean number
of lambs born, and EBV for NLB, NLW, MWWT, WWT, and the Katahdin index.
Materials and methods
Location
The experiment was conducted at USDA, Agriculture Research Service (ARS), Dale
Bumpers Small Farms Research Center (DBSFRC) in Booneville, AR, USA. All animal
procedures were approved by Institutional Animal Care and Use Committee (Protocol # 15019).
Animal management
Katahdin ewes were raised at USDA-ARS, DBSFRC in Booneville. A total of 163 ewes
of 0.5 to more than 7-yr of age were used in the study. All the ewes were maintained in a mixed
pasture of tall fescue (Schedonorus arundinacea) and bermudagrass (Cynodon dactylon) and
were supplemented with ewe ration (17% CP, 82% TDN on DM basis). Breeding and lambing
records of the flock were entered into National Sheep Improvement Program (NSIP) between
2001 and present to generate EBV. The EBVs of Katahdin hair sheep were obtained from NSIP
(www.nsip.org) and included were MWWT, WWT, NLB, NLW, and a Katahdin index.
Blood sampling and serum collection
Blood samples were collected July 2015 by jugular venipuncture into 10 ml BD
Vacutainer SS Plus Blood Collection Tubes (Becton, Dickson, Franklin Lakes, NJ). Blood tubes
were inverted to mix several times and allowed to clot for 30 min at room temperature. After
clotting, the tubes were held on ice until centrifuged (Beckman Coulter T J6 refrigerated

84
centrifuge, Fullerton, CA) at 2500 × g for 25 min. Serum was collected and stored in 5 ml
polypropylene tubes at -20° C until analysis.
Anti-Mullerian hormone analysis
Equine and ovine AMH ELISA used in the study is a quantitative three-step sandwich
type of immunoassay (AnshLabs, Webster, TX). Analysis was performed as described by the
manufacturer (www.anshlabs.com). Before analysis, all the kit reagents were warmed to room
temperature and serum samples were thawed overnight at 4° C. Solutions were mixed as
recommended. Calibrators were serially diluted from 13.6 ng/ml to 0.05 ng/ml of AMH. Fifty
microliter of duplicate calibrators, control and serum samples were added to appropriate wells of
the 96-well assay plate. Fifty microliter of AMH assay buffer was added to each well using a
repeater pipette. Plate was shaken at 700 rpm on an orbital microplate shaker (Lab-Line
Instrument Inc. Melrose Park, IL) for 2 h at room temperature. Plate was removed and washed 5
times with wash solution in microplate strip washer (ELP40 Bio-Tek Instruments, New York,
NY). Antibody-Biotin Conjugate RTU (100 µl) was added using a repeater pipette, incubated at
700 rpm on orbital microplate shaker (Lab-Line Instrument Inc. Melrose Park, IL) for 1 h and
washed 5 times with wash solution using the microplate strip washer. Streptavidin-enzyme
conjugate-RTU (100 µl) was added after washing, incubated in the orbital microplate shaker at
700 rpm for 30 min, and washed 5 times using microplate washer. TBM chromogen (100 µl) was
added to each well using a repeated pipette and incubated in orbital microplate shaker at 700 rpm
for 20 min. Finally, 100 µl of stopping solution was added to each well and absorbance was read
using a model 1420-040 Victor3 Multilabel Counter (Perkin Elmer, USA) at 450 nm.

85
Description of animals and categories
A total of 163 ewes aged 0.7 to more than 7-years was categorized into 5 groups: less
than 1 (n = 17), 1 to 2 (n = 57), 2 to 3 (n = 23), 3 to 4 (n = 34), and more than 4-yr of age (n =
32). Lambing data from 2010 to 2015 for ewes used in the current study were obtained from
USDA-ARS sheep flock at Booneville, AR. Ewes were bred either during summer or during
winter. Ewes from first to sixth parity are included and any lambing record from seventh parity
or above are excluded.
Statistical analysis
Regression analysis (bivariate fit) was used to determine the relationship between
absorbance reading and standard concentration of AMH. The resulting regression equation was
used to calculate AMH concentration in each unknown sample within the appropriate assay
plate. Pearson correlation coefficients between AMH and EBV for NLB, NLW, MWWT, WWT,
and Katahdin index were analyzed using the PROC CORR procedure of SAS (version 9.4; SAS
Inst. Inc., Cary, NC, USA). For remaining data, continuous variables were analyzed by PROC
GLM and categorical variables were analyzed by chi-squared using SAS. For categorical data
AMH concentrations were divided into four quartiles: Q1 = 0 – 80 pg/ml, Q2 = 81 – 104 pg/ml,
Q3 = 105 – 248 pg/ml, and Q = 249 – 1112 pg/ml. Data were expressed as least squares means
and standard error. Means were compared using Tukey’s test.
Results
Three microplates were used to run 163 serum samples. The regression equation (R2 =
0.998) used to determine AMH concentration in the first plate was: AMH ng/ml = - 0.034853 +
0.5789461 × absorbance. For plate 2, the regression equation (R2 = 0.99) used to determine
AMH concentration ng/ml serum was = - 0.5929 + 3.9394 × absorbance, and for the third plate,

86
regression equation (R2 = 0.99) used to determine AMH concentration ng/ml was = - 0.059384 +
2.2414964 × absorbance. Duplicate samples within plates were used to calculate intra-assay
variability, while standard curves and control samples in each plates were used to calculate inter-
assay variability. Intra-assay variability of AMH assays ranged from 6.6 to 8.3%. Inter-assay
variability between standards was 4.3%. Serum AMH ranged from 0.00 to 1112 pg/ml, with a
means of 182 pg/ml.
Correlation of EBVs for reproductive performance with serum AMH
Mean NLB, NLW, MWWT, WWT, and Katahdin index values were 0.056 ± 0.09 %,
0.117 ± 0.07 %, 0.365 ± 0.60 lb, 0.641 ± 0.73 lb, and 105 ± 3.14 lb, respectively. There was no
correlation between serum AMH concentrations and EBV for NLB (-0.02; P = 0.793), NLW (-
0.09; P = 0.252), MWWT (-0.02; P = 0.712), WWT (-0.02; P = 0.757), and Katahdin index
(0.072; P = 0.355).
Correlation between EBVs with average number of lambs born
Average number of lambs born were significantly correlated with EBV for NLB (0.29; P
= 0.0002) and NLW (0.19; P = 0.013). However, EBV for WWT (-0.09; P = 0.242), MWWT (-
0.11; P = 0.138), and Katahdin index (0.04; P = 0.547) were not correlated with average number
of lambs born.
Relationship between AMH quartiles and mean number of lambs born from different summer and
fall breeding
The distribution of AMH concentrations was divided into quartiles (AMH Q1 though Q4,
with Q1 the lowest and Q4 the highest pg/ml) for analysis. Anti-Mullerian quartiles with their
concentration range and mean number of lambs born is given Table 1. There was no correlation
between AMH quartiles with mean number of lambs born from fall breeding. However, there

87
was a significant correlation between mean number of lambs born from summer breeding with
AMH quartiles. Ewes in lowest quartile (Q1) had lower number of lambs born from summer
breeding than ewes in other quartiles (Q2, Q3, Q4; P < 0.05).
Influence of ewe age in serum AMH level
Mean AMH concentration in less than 1, 1 to 2, 2 to 3, 3 to 4, and more than 4-year age
was 129 ± 20, 148 ± 10, 159 ± 20, 195 ± 10, 166 ± 10 pg/ml respectively. There was no
difference in serum AMH concentration among age categories (P = 0.157).
Pregnancy and lambing rate of exposed ewes between breeding seasons
Overall pregnancy rate was higher (P < 0.01) in fall bred ewes (78.99%) compared with
summer bred (48.01%). Mean number of lambs born from fall breeding (1.44 ± 0.05) was higher
(P < 0.01) compared with mean number of lambs born from summer breeding (0.78 ± 0.05) per
ewes used for breeding.
Lambing rate per parity
Lambing rate was highest in third parity ewes and lowest in first parity ewes (P < 0.05).
Mean numbers of lambs born in first, second, third, fourth, fifth, and sixth parity are 0.81, 1.06,
1.52, 1.34, 1.09, and 1.33 respectively.
Discussion
Estimated breeding values (EBVs) are the mathematical predictions of the genetic merit
of an animal for specific traits. These values are calculated based on an individual animal’s
performance for a specific trait compared with animals within a contemporary group and animals
in other connected flocks, and also takes into account of progeny and relatives. An expected
progeny difference (EPD) on the other hand is predictor of genetic merit of an animal’s progeny.

88
Mathematically EBV is twice the value of EPD. In the US cattle industry, EPD is more common,
whereas EBV are more common outside US.
The mathematical calculation process for EBV is a very complex process. Data from an
individual lamb is entered into a computerized database. At first, non-genetic and phenotypic
parameters such as number of lambs born, age of lamb, age of dam, weaning weight and more
are adjusted to remove non-genetic effect of performance. Individual performance thus obtained
is compared with average production of a group of contemporary animals. After this, probability
of gene sharing is calculated. For this, an individual’s measurement is compared to the
measurement taken from its relatives. All the records are expressed in relation to their
contemporary groups in order to remove non-genetic difference in average performance. At last,
the EBV calculated is adjusted for the heritability of each trait. The resulting EBV is the best
estimate of the true genetic merit of any individual for a specific genetic trait measured
(www.nsip.org). The accuracy of EBV increases with an increase in data. For example, accuracy
of EBV for a breeding ram with 300 offspring in 5 different flocks will be higher than a ram with
20 offspring in one flock. The EBVs also improves by using a ram in multiple flocks. It is
imperative that a flock use at least two sires within a lambing season and at least 20 off spring
per sire (fewer are accepted within NSIP).
The EBV for Number of lambs born measures genetic potential for prolificacy. The NLB
EBV are expressed in percentage. Average number of lambs born per parity in the Katahdin hair
sheep breed is 2.1. This value is adjusted from all flocks over the country that are enrolled in
NSIP. An ewe with an EBV NLB of 23% means she is expected to lamb 23% more than the
average. Thus, she would be expected to have 2.1 + 0.48 lambs. The 2.1 is the average and 0.48
is 2.1 × 0.23.

89
Number of lambs weaned EBV measures the combined ewe effect on prolificacy and
lamb survival to weaning. The EBV NLW of -5% means a ewe is expected to wean 5% fewer
lambs than the average. If the average number of lambs weaned for Katahdins is 1.9, then she is
expected to wean 1.9 – 0.095 (1.9 × 0.05) lambs per parity or 1.805.
Maternal weaning weight EBV estimates genetic merit for mothering ability. The
MWWT EBV is derived by evaluating lambs that are heavier or lighter than expected based on
WWT EBV of the parents. This is mostly due to increased amount and quality of milk. Ewes
whose lamb grows faster than predicted are assumed to be better milk producers and vice-versa.
The MWWT EBV values are expressed in pound (in US).
Weaning weight EBV estimates preweaning growth potential. Preweaning weights are
taken at 45 to 90 d of age. Katahdin lambs are generally weaned at 90 d of age. The WWT EBV
values are expressed in pounds (in US). Suppose if average 90 d weaning weight of Katahdin
hair sheep is 60 lb; then WWT EBV of +1 of a lamb refers to potential weaning weight of 61 lb.
Katahdin index gives positive emphasis on maternal weaning weight, weaning weight,
and number of lambs weaned EBVs. Also gives small negative emphasis on number of lambs
born. Katahdin hair index = 0.246 × WWT EBV + 2.226 × MWWT EBV + 0.406 × NLW EBV
– 0.035 × NLB EBV. It predicts genetic value for total pounds of lamb marketed by a ewe (in
US). Katahdin hair index of 99 refers to 1 lb below the average whereas 101 refers to 1 lb above
the average.
In terms of link between AMH and EBVs, NLB is probably most directly related as it is
the closest estimate of pregnancy/lambing rate. The EBVs, NLW and MWWT are related to
mothering ability, which physiologically is related to functional hormones post-lambing such as
prolactin. However, EBVs for NLB and NWL are highly correlated. In the current study, EBVs

90
for NLB as well as NLW were correlated with number of lambs. Other EBVs, WWT, MWWT,
and Katahdin index were not correlated with number of lambs born.
In the current study, there was no correlation between serum AMH concentrations and
any of the EBVs. Ewes in the flock were selected for parasite resistance first, then maternal traits
(maternal milk and number of lambs weaned) for several years, but there was still a wide range
of EBV for maternal traits. Accuracy of some of these traits, especially in younger ewes may be
low enough to impact correlations. Other measures of fertility such as lifetime production after
accounting for environmental factors would be ideal but impractical.
Anti-Mullerian hormone is high during the fetal stage compared with postnatal
concentrations (Bezard et al., 1987). Similarly, plasma AMH concentrations in 40 d old ewes is
higher than 3.6-month old ewes (Lahoz et al., 2012; Torres-Rovira et al., 2014). Lahoz found a
vast difference between 1 and 5-month-old ewes, decreasing with age (Lahoz, et al., 2016b).
Serum AMH concentrations of ewes after puberty, at different reproductive age period has not
been reported. In the current study, serum AMH concentrations was similar in different age (less
than one, 1-2, 2-3, 3-4, and more than 4 yr) ewes. No difference in serum AMH concentrations in
different age mature ewes is similar to that reported in cattle (Rico et al., 2009; Ireland et al.,
2010).
Ovarian follicular population, an indicator for fertility is positively correlated with
plasma AMH concentration in young lambs (Lahoz et al., 2016b). Correlation between plasma
AMH concentrations during the prepubertal period and occurrence of ovulation in response to
eCG was reported (Lahoz et al., 2012). Ewes with a high plasma AMH (> 97 pg/ml) had higher
number of lambs born per ewe per year till 3.5 yr age compared with low (< 97 pg/ml) plasma
AMH concentrations measured at 3.5 months of age (Lahoz et al., 2016 a). When ewes were

91
compared by quartiles, based on serum AMH measurement after puberty, AMH hormone
concentration had no effect on mean number of lambs born. Similar results were obtained when
ewes were compared by quartiles, based on serum AMH measurement after puberty with mean
number of lambs born from fall breeding. However, ewes in lowest quartiles (Q1) had lower
number of lambs born from summer breeding than ewes in other quartiles. Low reproductive
performance of ewes in lowest quartiles (Q1) during summer in the current study is similar to the
result we recently reported in cattle (Rorie et al., 2016). In our previous study, we reported AMH
concentration at breeding, heifers in lowest quartiles (Q1) had lower AI pregnancy rate and
conceived at a later estrus cycle in breeding compared with heifers in other quartiles. Anti-
Mullerian hormone measurement after puberty could be an option for selection of ewes for
summer breeding in the Southern US.
In conclusion, serum AMH concentration were similar among age group. There was a
correlation between EBV for NLB and NLW with average number of lambs born. Ewes in the
lowest quartile (Q1) had lower number of offsprings born from summer breeding compared with
ewes in other AMH quartiles (Q2, Q3, and Q4). However, AMH was not useful in predicting
fertility or maternal traits based on EBV of the ewes.

92
References
Bézard, J., B. Vigier, D. Tran, P. Mauleon, and Nathalie Josso. 1988. Anti-Müllerian hormone in
sheep follicles. Reprod. Nutr. Dev. 28:1105-1112.
Broer, S. L., F. J. M. Broekmans, J. S. E. Laven, and B. C. J. M. Fauser. 2014. Anti-Müllerian
hormone: ovarian reserve testing and its potential clinical implications. Hum. Reprod.
Update. 20:688-701.
Burke, J. 2005. Lamb production of Dorper, Katahdin, and St. Croix bred in summer, winter, or
spring in the Southeastern US. Sheep Goat Res. J. 20:51-59.
Campbell, B. K., M. Clinton, and R Webb. 2012. The role of anti-Müllerian hormone (AMH)
during follicle development in a monovulatory species (sheep). Endocrinology 153:4533-
4543.
Cate, R. L., R. J. Mattaliano, C. Hession, R. Tizard, N. M. Farber, A. Cheung, E. G. Ninfa, A. Z.
Frey, D. J. Gash, E. P. Chow, R. A. Fisher, J. M. Bertonis, G. Torres, B. P. Wallner, K. L.
Ramachandran, R. C. Ragin, T. F. Manganaro, D. T. MacLaughlin, and P. K. Donahoe.
1986. Isolation of the bovine and human genes for müllerian inhibiting substance and
expression of the human gene in animal cells. Cell 45:685-698.
Fahmy, M. H. 1986. Preliminary results on fertility, prolificacy, lamb production and carcass
traits of Romanov sheep in Canada. Proc. 3rd World Congr. Genet. Appl. Livest. Prod.
IX:559-564. Lincoln, NE.
Ireland, J. J., G. W. Smith, D. Scheetz, F. Jimenez-Krassel, J. K. Folger, J. L. H. Ireland, F.
Mossa, P. Lonergan, and A. C. O. Evans. 2010. Does size matter in females? An
overview of the impact of the high variation in the ovarian reserve on ovarian function
and fertility, utility of Anti-Mullerian hormone as a diagnostic marker for fertility and
causes of variation in the ovarian reserve in cattle. Reprod. Fertil. dev. 23:1-14.
La Marca, A., V. Grisendi, and G. Griesinger. 2013. How much does AMH really vary in normal
women? Int. J. Endocrinol. 2013:1-8.
Lahoz, B., J. L. Alabart, and J. Folch. 2016a. Relationship between Anti-Müllerian hormone
(AMH) and the ovulatory response measured in three-month old ewe lambs with their
reproductive efficiency in adulthood. Asoc. Interprofes. Para el Des. Agr. 112:392-404.
Lahoz, B., J. L. Alabart, D. Monniaux, P. Mermillod, and J. Folch. 2012. Anti-Mullerian
hormone plasma concentration in prepubertal ewe lambs as a predictor of their fertility at
a young age. BMC Vet. Res. 8:2-9.
Lahoz, B., J. L. Alabart, M. J. Cocero, D. Monniaux, E. Echegoyen, P. Sánchez, and J. Folch.
2014. Anti-Müllerian hormone concentration in sheep and its dependence of age and

93
independence of BMP15 genotype: an endocrine predictor to select the best donors for
embryo biotechnologies. Theriogenology. 81:347-357.
Lahoz, B., S. Fabre, J. H. Calvo, L. Drouilhet, P. Sanchez, P. Sarto, E. Echegoyen, J. Folch, D.
Monniaux, J. L. Alabart. 2016b. Assessing the age-related relationship between plasma
Anti-Mullerian hormone (AMH) and ovarian follicle population in prepubertal ewes.
Published online https://core.ac.uk/download/pdf/71402615.pdf. Accessed on 4th July
2017.
Monniaux, D., L. Drouilhet, C. Rico, A. Estienne, P. Jarrier, J. Touzé, J. Sapa, F. Phocas, J.
Dupont, R. Dalbiès-Tran, and S. Fabre. 2012. Regulation of anti-Müllerian hormone
production in domestic animals. Reprod. Fertil. Dev. 25:1-16.
Rico, C., S. Fabre, C. Médigue, N. di Clemente, F. Clément, M. Bontoux, J. L. Touze, M.
Dupont, E. Briant, B. Remy, J. F. Beckers, and D. Monniaux. 2009. Anti-müllerian
hormone is an endocrine marker of ovarian gonadotropin-responsive follicles and can
help to predict superovulatory responses in the cow. Biol. Reprod. 80:50-59.
Rorie, R., H. Newberry, T. D. Lester, M. Acharya, and C. Hansen. 2016. Anti-müllerian
hormone at weaning and breeding as a predictor of beef heifer fertility. Reprod. Fertl.
Dev. 29:111-112 (Abstr.).
Scheetz, D. 2010. Regulation and role of Anti-Mullerian hormone in bovine reproduction
(Doctoral dissertation). Retrieved from
http://search.proquest.com/openview/9e2ae630eee668c494892306d4d53f0f/1?pq-
origsite=gscholar&cbl=18750&diss=y. Accessed on 4th July 2017.
The Baalands. 2017. Katahdin frequently asked question. Available online.
http://www.baalands.com/#!katahdinfaqs/cwoc. Accessed on 24th April 2017.
Torres-Rovira, L., A. Gonzalez-Bulnes, S. Succu, A. Spezzigu, M. E. Manca, G. G. Leoni, G. G.,
M. Sanna, S. Pirino, M. Gallus, S. Naitana, F. Berlinguer. 2014. Predictive value of antral
follicle count and anti-Müllerian hormone for follicle and oocyte developmental
competence during the early prepubertal period in a sheep model. Reprod. Fertil. Dev.
26:1094-1106.
Vanimisetti, H. B., D. R. Notter, and L. A. Kuehn. 2007. Genetic (co) variance components for
ewe productivity traits in Katahdin sheep. J. Anim. Sci. 85:60-80.

94
Table 1. Relationship between AMH quartiles and mean number of lambs born from different
summer and fall breeding. Data expressed in mean ± standard error (SE).
AMH Quartiles AMH range (pg/ml) Fall Summer Overall
AMHQ 1 0 – 80 1.48 ± 0.11 0.41 ± 0.12b 0.65 ± 0.10
AMHQ 2 81 – 104 1.54 ± 0.11 0.92 ± 0.11a 0.81 ± 0.11
AMHQ 3 105 – 248 1.38 ± 0.11 0.89 ± 0.10a 0.87 ± 0.10
AMHQ 4 249 – 1112 1.43 ± 0.10 0.90 ± 0.10a 0.71 ± 0.11
Means with different superscripts in the same column differ (P < 0.05).

95
Chapter VI: Conclusions
Most of the CASA motility parameters were significantly higher after extension and
storage at 4 compared with 15° C. Ram semen stored using milk or TRIS based extenders at 4° C
can maintain similar motility parameters but better compared with TEST or CJ-2 extenders after
storage for up to 3 d. Ram semen stored at 15° C for 3 d using TRIS extender had better sperm
motility parameters compared with milk, TEST or CJ-2 extender. Milk and TEST based
extenders had intermediate efficacy for maintaining motility at 15° C. In contrast, sperm cells
stored in CJ-2 extenders had drastic reduction of most motility parameters after storage at 15° C.
Combined results from both temperatures shows most of the motility parameters were similar for
up to 24 h of storage in all extenders but after that sperm motility parameters declined more
rapidly in milk or CJ-2 extenders compared with TEST or TRIS extender. Such decrease is due
to greater reduction after 24 h using milk and CJ-2 extenders and storage at 15° C. Considering
sperm progressive motility which is also a predictor for in vivo fertility shows milk and TRIS
extenders stored at 4° C has similar but higher progressive motility compared with ram sperm
extended in CJ-2 and TEST extenders and stored at 4° C. However, progressive motility of ram
sperm declined 75% when stored at 4° C for 24 h, and continued to decline over time irrespective
of extenders used. Further study can be done to supplement different supplements to milk or
TRIS based extenders for extension and storage of ram semen at 4° C.
Milk extender maintained better sperm motility compared with TRIS after storage at 4° C
for up to 72 h when different supplements were added. Addition of 5 or 20% egg yolk on
extenders had no difference on most of the sperm motility parameters. However, inclusion of
20% egg yolk in milk extender had highest track speed, and path velocity followed by TRIS +
20% egg yolk, TRIS + 5% egg yolk, and milk + 5% egg yolk, respectively. At 72 h of storage at

96
4° C, sperm cell motility declined sharply in extenders without ethylene glycol (EG) compared
with extenders supplemented with 1% EG. Results indicate that milk extender supplemented
with 1% EG, 20% egg yolk before storage, and addition of PHE before analysis is beneficial for
liquid storage of ram semen.
Serum AMH concentrations was similar in different aged ewes after puberty. There was a
correlation between EBV for NLB and NLW with average number of lambs born. Ewes in the
lowest quartile (Q1) had lower number of offsprings born from summer breeding compared with
ewes in other AMH quartiles (Q2, Q3, and Q4). However, AMH was not useful in predicting
fertility or maternal traits based on EBV of the ewes.

97