in situ of crop wild relatives: a methodology for -...
TRANSCRIPT

An Integrated European In Situ Management Workplan: Implementing Genetic Reserve and On‐farm Concepts
In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
Tested with case studies for Avena, Beta, Brassica and Prunus
By S. Kell, N. Maxted, L. Frese, J.M. Iriondo, B. Ford‐Lloyd, K. Kristiansen, A. Katsiosis, C. Teeling and F. Branca
http://aegro.jki.bund.de/


AEGRO
An Integrated European In Situ Management Workplan: Implementing Genetic Reserve and On‐farm Concepts
In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
Tested with case studies for Avena, Beta, Brassica and Prunus
By S. Kell, N. Maxted, L. Frese, J.M. Iriondo, B. Ford‐Lloyd, K. Kristiansen, A. Katsiosis, C. Teeling and F. Branca
Front cover: Avena prostrata growing in southern Spain
Photographer: Andreas Katsiotis
Funded by the European Commission’s Community programme on the conservation, characterization, collection and utilization of genetic resources in agriculture as a targeted
action in accordance with Council Regulation (EC) no. 870/2004
Agreement no. 057 AGRI GEN RES
Contract no. AGRI‐2006‐0396


Contents
1.0 Introduction ....................................................................................................................... 3
1.1 Background ..................................................................................................................... 3
1.2 Approaches to in situ CWR conservation ....................................................................... 4
1.3 The genetic reserve concept .......................................................................................... 4
1.4 Identifying priority sites for in situ CWR conservation: developing a common methodology .................................................................................................................. 6
2.0 Methodology for identifying CWR genetic reserve sites for a target crop gene pool ...... 7
2.1 Step 1: Taxon delineation ............................................................................................... 7
2.2 Step 2: Selection of target taxa ...................................................................................... 8
2.3 Step 3: Diversity analysis .............................................................................................. 12
2.4 Step 4: Selection of target sites ................................................................................... 14
3.0 Application of the methodology to four crop case studies ............................................. 17
3.1 Crop case studies: overview and purpose ................................................................... 17
3.2 Application of the methodology in the Avena case study ........................................... 18
3.3 Application of the methodology in the Beta case study .............................................. 21
3.4 Application of the methodology in the Brassica case study ........................................ 25
3.5 Application of the methodology in the Prunus case study .......................................... 28
4.0 Discussion ......................................................................................................................... 32
5.0 Conclusion ........................................................................................................................ 35
References .................................................................................................................................. 36

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
2

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
3
1.0 Introduction
1.1 Background
Crop wild relatives (CWR) are species closely related to crops and are defined by their potential ability to contribute beneficial traits to crops (Maxted et al., 2006); they have been used in plant breeding since the early 20th century and have provided vital genetic diversity—for example, to confer resistance to pests and diseases, improve tolerance to environmental conditions such as extreme temperatures, drought and flooding and to improve nutrition, flavour, colour, texture and handling qualities (Maxted and Kell, 2009). In monetary terms, CWR have contributed significantly to the agricultural and horticultural industries, and to the world economy (Maxted et al., 2008a; Maxted and Kell, 2009).
Today, agricultural production is challenged by climate change (e.g., see Jones et al., 2003; Duveiller et al., 2007; Deryng et al., 2011; Li et al., 2011; Luck et al., 2011). The International Panel on Climate Change (IPCC, 2007) estimates that by 2100, crop yields will be reduced by as much as 40% in some regions unless climate change mitigation is undertaken. Breeders will therefore have to provide varieties able to cope with the impacts of changing growing conditions. Due to the breadth of genetic diversity inherent in CWR populations, which are adapted to a wide range of environmental conditions, they are likely to be needed more than ever before to maintain the adaptability of crops. Thus, CWR are a critical component of plant genetic resources for food and agriculture (PGRFA) and are vital for future food security; however, despite their recognized value, they have historically received relatively little systematic conservation attention.
There are two primary techniques for CWR conservation: in situ, primarily in natural habitats managed as genetic reserves (GRs) (see section 1.3) and ex situ as seed in gene banks. Historically, CWR conservation has focused almost entirely on ex situ collection and storage, but it can be argued that in situ conservation is more appropriate because the genetic diversity inherent in and between wild CWR populations is constantly changing in response to their environment; therefore, CWR populations are a component of natural ecosystems that cannot effectively just be maintained ex situ (Maxted et al., 2008a).
A number of recent initiatives have raised the profile of CWR and put them on the international conservation agenda. However, conservationists and policy‐makers are faced with the difficult challenge of how to conserve the large number of CWR species and the genetic diversity that they contain. If a broad definition of a CWR is used (i.e., all the species in the same genus as a cultivated plant), there are more than 16,000 crop and CWR species in the territories of the EU Member States—13,875 of these are native and at least 2665 are endemic (Kell et al., 2008). CWR are under threat from habitat loss, agricultural intensification, over‐collection, climate change and lack of conservation attention, yet only 9% of PGR accessions in European gene bank collections are CWR (Dias et al., 2011), while most wild populations in situ are not actively monitored and managed in or outside protected areas (PAs) (Maxted et al., 2008b). There is therefore a real challenge to the nature conservation and PGRFA sectors to conserve these valuable resources.
The AEGRO project was initiated to fill some knowledge gaps and to provide a range of practical tools for in situ CWR conservation as a contribution to an integrated EU PGRFA conservation strategy. This report presents a methodology for the development of a CWR in situ conservation strategy for target crop gene pools that can be applied both within Europe and elsewhere in the world. The methodology has been tested and applied in four crop case

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
4
studies (Avena, Beta, Brassica and Prunus) by partners in the AEGRO project and this report presents an analysis of the application of the methodology to each case study with the aim of illustrating how such a generic methodology can be successfully applied across a range of different crop gene pools.
1.2 Approaches to in situ CWR conservation
There are various approaches to achieving the systematic conservation of CWR diversity; however, two distinct but complementary approaches may be characterized as floristic and monographic (Maxted et al., 2011). These approaches relate to the breadth of coverage of the CWR conservation strategy—a floristic approach involves the development of a CWR conservation strategy for CWR diversity that occurs in a defined geographical area (which may be a sub‐national area such as an administrative unit or protected area, a whole country, a supra‐national region, or even the whole world), while a monographic approach is restricted to certain crop gene pools, but like the floristic approach may be carried out at any geographical scale (Maxted et al., 2011).
The floristic approach is comprehensive because it attempts to encompass all CWR diversity that occurs within a geographical unit; however, while being comprehensive for the geographical unit, the full geographic range of an individual taxon may or may not be included, depending on whether it is endemic to that geographical unit. The monographic approach focuses on CWR diversity within target crop gene pools which are usually identified on the basis of their perceived value for food security and/or economic stability. Both approaches will ultimately conclude with the systematic conservation of priority CWR diversity via a network of conservation sites and genetic reserves, with backup in ex situ collections (Maxted et al., 2011).
Whether a floristic or monographic approach is taken is likely to depend on: a) the quantity and quality of existing data and b) the resources available to prepare the conservation strategy. The scope of the parent organization undertaking the conservation may also impact the approach; for example, an international cereal research institute is likely to focus monographically on cereal crops, while a national biodiversity institute is likely to adopt a more floristic approach. It is worth noting that if the goal is to maximize CWR diversity, it is likely that both approaches need to be combined (Maxted et al., 2011). In Europe, a combined floristic and monographic approach is being promoted and taken forward in the context of the EU‐funded project, PGR Secure (www.pgrsecure.org). The development of national CWR conservation strategies (taking the floristic approach) and a European strategy (taking the monographic approach for high priority crop gene pools) will contribute towards an overall European strategy for PGRFA conservation.
1.3 The genetic reserve concept
A genetic reserve is defined as ‘the location, management and monitoring of genetic diversity in natural populations within defined areas designated for long‐term conservation’ (Maxted et al., 1997a). The concept combines in situ conservation with active management and a long‐term approach. The rationale for this type of conservation is that it is a) applicable to all plant species, b) allows for continued evolution and c) allows for multiple‐taxon conservation. Moreover, it conserves the genetic diversity of the target taxon in a dynamic way, as well as its habitat and all existing biotic and abiotic interactions (including humans).

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
5
Several approaches to GR conservation can be identified, each with different aims and strategies, depending on the approach (see Maxted and Kell, 2009). For example, the aims for CWR GRs in Europe are to conserve genetic diversity in the widest range of priority CWR taxa at the European scale; therefore, the aim is to design a network of reserves that adequately and efficiently maintains the genetic diversity of the target taxa. When we talk about ‘adequately’ maintaining the genetic diversity of target taxa, we mean conserving a good representation of the genetic diversity of adaptive and agricultural value present in such taxa. Similarly, by ‘efficiently’ we mean to obtain this goal using the minimum number of GRs.
Coordination with ex situ holdings and crop databases is an important part of the genetic reserve concept (Maxted et al., 1997a). Ex situ seed banks can be a relevant component in the functioning of GRs as they provide a back‐up of genetic diversity in case any catastrophe should occur. Furthermore, they facilitate information exchange, access for breeding and other research, and promote use.
The end point of the methodology described and discussed in this report is the identification of ideal GR sites. However, it is useful to briefly review the steps that follow, once the location of a GR has been decided. Firstly, it is necessary to consider the design of the reserve. Some of the factors to take into account are a) the shape and size of the reserve, b) target taxon population size, c) political and socio‐economical factors and d) the possibility of establishing networks and corridors with other neighbouring reserves. Secondly, a genetic reserve management plan needs to be developed. A management plan is a planning tool that contains a set of prescriptions and interventions to meet the objectives of the GR (Heywood and Dulloo, 2005). The minimum elements of a management plan have been described in detail by Maxted et al. (2008c). Essentially, a management plan is composed of three basic elements: i) assessment of the taxon, population and site, ii) establishment of management targets, and iii) prescription of conservation actions.
Evaluation is an essential component for management success. The purpose is to identify and determine the factors that affect population viability and the maintenance of genetic diversity in the populations. The establishment of clear, precise management targets is needed to obtain and evaluate the desired results and to measure success (Elzinga et al., 1998). Management and monitoring approaches depend on the definition of specific targets (e.g., what status do we want to achieve in the population, or under what circumstances are we going to take a particular action?) Some examples of management targets can be to keep population size over a certain number of individuals, not to allow a decline in population size over a certain percentage in one year, or to keep a specific allele that is key for pathogen tolerance in a gene at a frequency above a certain percentage.
Finally, management prescription includes all types of actions that can be executed on the target population, its habitat and the surrounding community to maintain ecological conditions and processes that are compatible with and necessary for the survival and maintenance of the genetic diversity in the target population (Maxted et al., 2008c). Management prescription also includes monitoring, which is a procedure to assess the effects of the management actions that are implemented. Since as a result of monitoring we may be able to conclude that our management actions are not producing the results we wanted, management prescription must also consider what actions to take when things do not go the way they are supposed to.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
6
1.4 Identifying priority sites for in situ CWR conservation: developing a common methodology
The establishment of GRs for CWR is a priority in order to maintain a broad range of genetic diversity within and between populations; however, with a large number of species to conserve, a systematic approach to the identification of GR sites is needed to maximize resource use. In this report, we present a generic methodology that can be used to prioritize taxa on the basis of their potential use for crop improvement and relative threat status, gather the necessary data to undertake diversity and gap analysis for target taxa, and select the most appropriate CWR GR sites. The methodology is built on those proposed by Maxted et al. (in prep.), Maxted et al. (2008d) and Maxted and Kell (2009), which address floristic and monographic approaches to CWR conservation. Although it may be necessary to adjust parts of the methodology according to the specific biological, ecological and geographical attributes of individual crop complexes, it provides a generic framework for the conservation of any crop gene pool.
There are four basic steps in the methodology: a) taxon delineation, b) selection of target taxa, c) diversity analysis and d) selection of target sites. Within each of the four steps, precise instructions are provided on how to develop the strategy. The end point of the methodology is the identification of ‘ideal’ CWR GR sites. The political and legal steps that need to be taken beyond this point to establish the GRs are not part of the methodology. The next step beyond the methodology for identification of GRs is to make recommendations for site and population management (see Maxted et al., 2008c).
The methodology was first made available to AEGRO project partners via the project intranet as part of the ‘AEGRO Helpdesk’. After testing and further development, the methodology was made available in the public domain (see http://aegro.jki.bund.de/aegro/index.php?id=188) as part of the ‘CWR In Situ Strategy Helpdesk’, which is provided as a guide and information facility for national programmes, research institutes, NGOs, protected area managers or individuals involved in the development of a CWR in situ conservation strategy. The methodology is presented in the next section. For more detailed step by step guidance on implementing the methodology, including a list of data sources, the reader is referred to the Helpdesk.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
7
2.0 Methodology for identifying CWR genetic reserve sites for a target crop gene pool
2.1 Step 1: Taxon delineation
Background
The starting point for a crop gene pool CWR conservation strategy is a list of target taxa; therefore, for the target crop gene pool it is necessary to:
1. Generate a list of taxa that occur in the crop gene pool. Although not all the taxa in the gene pool will necessarily be immediately included in the CWR conservation strategy, the complete list of taxa provides a reference point for future potential conservation actions of lower priority taxa.
2. Generate a list of taxa that occur within the defined geographic range of the conservation strategy (i.e., national, regional or global). These may be both native and introduced, but the conservation strategy is most likely to focus on native species.
To achieve these two steps, online information sources and/or literature (monographs, crop‐specific studies etc.) need to be consulted (see Maxted and Guarino, 2003).
At this stage, it is necessary to adopt an accepted taxonomy to form the basis of the taxon list and the subsequent conservation strategy. The list of taxa should show the accepted taxon name and authority and list primary synonyms with authorities. This is important because different information systems use different accepted taxonomies; therefore, when searching for information on a specific taxon it could be possible to miss important information if synonymy is not taken into account.
Step 1 actions – list the taxa in the crop gene pool
1. Produce a list of all taxa within the crop gene pool, including both accepted names and synonyms with authorities. To achieve this, consult available online information sources and/or consult the literature (monographs, crop‐specific studies etc.).
2. If applicable, list the taxa that occur within the geographic range (i.e., national or regional) of the CWR conservation strategy. Several online data sources can be consulted to provide information on the distribution of the taxa. However, good starting points are the Crop Wild Relative Catalogue for Europe and the Mediterranean, accessible via the Crop Wild Relative Information System (CWRIS – http://www.pgrforum.org/cwris/cwris.asp) and the Global Biodiversity Information Facility (GBIF – http://data.gbif.org/welcome.htm). For national‐level data, the same data sources can be consulted, as well as national Floras and databases, as applicable. To access these online data sources, further information on other sources of national‐level data, and for advice on how to use CWRIS, GBIF and other online information sources, visit the online Helpdesk data sources web page at http://aegro.jki.bund.de/aegro/index.php?id=193.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
8
2.2 Step 2: Selection of target taxa
Background
In general, it is not practical to attempt to actively conserve all the taxa within the crop gene pool due to resource limitations; therefore, we need to prioritize and select taxa from the list that will be proposed for active conservation. Factors that can be used to ascribe ‘value’ and establish conservation priorities include (Maxted et al., 1997a):
Current conservation status
Socio‐economic use
Threat of genetic erosion
Genetic distinctiveness
Ecogeographic distribution
Biological importance
Cultural importance
Cost, feasibility and sustainability
Legislation
Ethical and aesthetic considerations
Priorities of the conservation agency
For CWR, an initial, simple prioritization on the basis of socio‐economic use of the associated crop (a step which will already have been taken in selecting the target crop gene pool) and relative threat has been proposed (e.g., Ford‐Lloyd et al., 2008; Magos Brehm et al., 2008). In addition, Maxted and Kell (2009) proposed that within each crop gene pool, the closest wild relatives should be afforded higher conservation priority over the more distantly related species because these are the taxa that can more easily be used in crop improvement using conventional breeding methods. However, the literature on the taxa within the target crop gene pool should be thoroughly searched to check for cases where a more distantly related taxon has been highlighted as a gene donor (or potential gene donor), and these taxa should also be afforded conservation priority. Of these prioritized taxa, those in most urgent need of conservation action (i.e., those with a very limited geographic range—often rare or endemic taxa—and/or known to be under threat), are given precedence.
This methodology therefore primarily targets the taxa that are most closely related to the crop species (or that have shown promise in crop improvement programs) and that are threatened or have restricted distribution ranges. However, ideally, national and regional in situ networks of CWR GRs should in the long term be expanded to ensure that all taxa of potential importance for crop improvement are actively conserved. In particular, selected populations of the primary and secondary wild relatives that are widespread and common should be actively conserved throughout their range, ensuring that populations representing the extremes of the range (both geographically and topographically) are conserved. Individual populations of these taxa may harbour important genes adapted to particular environmental conditions—genes that may confer important traits to improve crops in the future. Populations of these taxa that already occur within PAs should also be monitored. In many

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
9
cases, if a floristic approach is taken, it is possible to establish a reserve that conserves multiple CWR taxa, which, when possible, has obvious advantages.
There are two stages to the selection of target taxa: a) creation of a level 1 prioritized list based on actual or potential use as gene donors, and b) creation of a level 2 prioritized list based on threat and/or distribution. In this methodology, the two steps are presented sequentially (i.e., the level 2 prioritized list is based on the level 1 prioritized list). The advantage of this approach is that in cases where there is limited information on the distribution of the taxa and/or for gene pools containing a very large number of taxa, the level 1 prioritization narrows the list of taxa down to those that are likely to be most important as gene donors for crop improvement and further information is only sought for that list of taxa. The disadvantage of this approach is that some of the more distantly related taxa in the gene pool that are threatened or have restricted distributions may be missed in the conservation planning process. Therefore, in cases where a gene pool contains a relatively small number of taxa or where distribution data are readily available for all the taxa, it is desirable to undertake the prioritization in the reverse order by collating threat and distribution data on all taxa in the gene pool first, then applying the second level of prioritization based on potential use as gene donors. Using this approach, more distantly related taxa that are threatened or have restricted distributions can be highlighted as a conservation priority on that one criterion, and even though they may still not be given the highest level of priority for immediate conservation action, they may be promoted as candidates for conservation at a later date. Furthermore, if it is not immediately possible to put in place in situ conservation measures for these taxa, they can be earmarked for collection and storage in ex situ collections.
Step 2 actions – a) create a level 1 prioritized taxon list At this stage, the aim is to prioritize the taxon list on the basis of value or potential value as gene donors for crop improvement, as follows:
1. Organize the list of all taxa within the crop gene pool according to their degree of relationship to the crop. To achieve this, search the available literature on the crop complex. Taxa should be organized into a table showing primary, secondary or tertiary wild relatives using one of the following three methods:
a) Where genetic information is available and taxa have been classified using the Gene Pool concept (Harlan and de Wet, 1971), organize the taxa into the table listing those in GP1B as primary wild relatives, those in GP2 as secondary wild relatives and those in GP3 as tertiary wild relatives. The Gene Pool concept works like this:
GP1A: cultivated forms of the crop
GP1B: wild or weedy forms of the crop
GP2: the coenospecies (less closely related species) from which gene transfer to the crop is possible but difficult using conventional breeding techniques
GP3: the species from which gene transfer to the crop is impossible, or if possible, requires sophisticated techniques, such as embryo rescue, somatic fusion or genetic engineering.
b) Where genetic information is not available, if possible, substitute the Gene Pool concept with the Taxon Group concept (Maxted et al., 2006), which provides a proxy for taxon genetic relatedness. Organize the taxa into the table listing those in TG1b as primary wild

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
10
relatives, those in TG2 as secondary wild relatives, and those in TG3 and TG4 as tertiary wild relatives. The Taxon Group concept works like this:
TG1a: crop
TG1b: same species as crop
TG2: same series or section as crop
TG3: same subgenus as crop
TG4: same genus.
c) For crop genera that have not been classified using the Gene Pool concept and not sub‐classified into sections and subgenera, the available information on genetic and/or taxonomic distance must be analysed to make reasoned assumptions about the most closely related taxa.
Whichever system is used, ensure that references are provided to substantiate the assumptions made about taxon relatedness.
2. Select the taxa from the categorized list that occur within the geographical area of the CWR conservation strategy by matching against the geographically defined list of taxa created under Step 1. You will now have two tables, both categorized according to their degree of relationship to the crop—one showing all taxa in the crop genus, the other showing the taxa that occur within the geographical area defined by the CWR conservation strategy.
3. In both tables, flag the taxa that have been identified as gene donors or potential gene donors. To achieve this, consult online information sources, such as GRIN Taxonomy and/or carry out literature searches using online library databases. Specialist libraries can also be consulted, such as those housed in gene banks, botanic gardens and other research institutes. Consult the online Helpdesk data sources page for further information (http://aegro.jki.bund.de/aegro/index.php?id=193). Provide references for any taxa flagged as gene donors or potential gene donors in the table. These may be in the tertiary list, as well and the primary and secondary lists.
4. Make a separate list of the primary and secondary wild relatives and any tertiary wild relatives that have been identified as gene donors or potential gene donors from the list of taxa within the geographical area defined by the CWR conservation strategy. This list (level 1 priority list) now forms the basis for the second level of prioritization under Step 2b.
Step 2 actions – b) create a level 2 prioritized taxon list
At this stage, the aim is to prioritize the level 1 priority taxon list on the basis of threat status. However, as already noted, this level of prioritization can be applied to all taxa listed under Step 1, followed by prioritization on the basis of actual or potential value for crop improvement, depending on the size of the gene pool and on the availability of distribution and/or threat data. Therefore, it is also possible to carry out the prioritization process in reverse order (i.e., by carrying out Step 2b followed by Step 2a).
There are two steps to this selection process:

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
11
1. Select the taxa in the level 1 priority list (or Step 1 list) that are known to be under threat. Where existing evidence is available to show that a taxon in the list is under threat, these taxa should automatically be prioritized for conservation action. To find out whether a taxon is known to be under threat:
Search the IUCN Red List of Threatened Species to find out if any of the taxa are listed.
Where possible, consult national or regional Red Lists to see whether a taxon in the list is considered threatened at national or regional level.
If a taxon is not included in the IUCN Red List, or in a regional or national Red List (i.e., it has not been formally assessed using a set of Red List criteria), it does not mean that it is not threatened. A literature search may reveal important information about the threatened status of a taxon. To achieve this, carry out literature searches using online library databases. Specialist libraries can also be consulted, such as those housed in gene banks, botanic gardens and other research institutes.
Consult the online Helpdesk data sources page (http://aegro.jki.bund.de/aegro/index.php?id=193) for further information and advice on how to carry out these searches.
2. Select the taxa in the level 1 priority list (or Step 1 list) that are not known to be under immediate threat, but are known to have limited distribution ranges. At this stage, a degree of objectivity is required, since there is no clear dividing line between a taxon with a limited distribution range and one with a distribution range that is deemed to enable ‘classification’ of the taxon as one not in immediate need of conservation action, unless very detailed information is already available about the taxa. However, where the range of a taxon is known, the methodology proposed by Ford‐Lloyd et al. (2008, 2009) can be used as a guide when establishing taxon conservation priorities at regional level (e.g., across Europe). Generally speaking, taxa that are known to be endemic to a country or subnational unit or those that occur in only a few countries or subnational units are more likely to be under threat at regional level. Similarly, at national or subnational level, available information must be gathered on the range of the taxa in order to establish which are most likely to be threatened by their limited distribution range. To find the distribution range of the taxa:
At European level, the Crop Wild Relative Catalogue for Europe and the Mediterranean (Kell et al., 2005)—which can be accessed via the Crop Wild Relative Information System (CWRIS – www.pgrforum.org/cwris/cwris.asp)—provides a list of country or subnational occurrences for taxa throughout Europe.
At national or subnational level, a good starting point is GBIF, which provides known taxon occurrences through data provided by organizations managing biodiversity data around the world (e.g., ex situ germplasm collections and herbarium data). For a finer level of detail, search available national databases and literature sources. Taxon specialists, conservation agencies and protected area managers are further important sources of information.
Consult the online Helpdesk data sources page (http://aegro.jki.bund.de/aegro/index.php?id=193) for further information and advice on how to carry out these searches.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
12
After carrying out Step 2, you now have a reduced list of taxa that have been selected on the basis of their value as gene donors and relative threat. This list of target taxa now forms the basis for immediate conservation planning for the crop gene pool.
2.3 Step 3: Diversity analysis
Background
Once the priority list of CWR species has been identified (Step 2), the next step is to collate the available ecogeographic information to assist in further formulation of the CWR conservation strategy1. This involves the collation and analysis of geographic, ecological, environmental and genetic data. These data are predictive and aid the location of the CWR taxonomic (inter‐taxa) and genetic (intra‐taxon) diversity that can then be targeted for conservation. As the goal is to maximize conserved genetic diversity, information on the partitioning of genetic diversity across the ecogeographic distributions of the target taxa is useful for identifying sites or combinations of sites of maximum diversity. However, even with rapidly decreasing costs of analysing genetic diversity, this information may be extremely limited; in which case, analysis of ecological and environmental data associated with the sites at which the populations occur can be used as a proxy for genetic diversity. The culmination of the diversity analysis should be a set of areas with high concentrations of the priority CWR species and populations of CWR taxa containing or thought to contain complementary and/or unique genetic diversity.
Step 3 actions2 – a) carry out an ecogeographic diversity analysis At this stage, the aim is to collate all available ecogeographic data associated with the target taxa. Assuming the taxonomic data has already been collated in Step 1, the remaining data are of three main types:
Geographic
Genetic
Ecological/environmental
1. Gather geographic (distribution) data on the target taxa. Data are of two types—coordinate and descriptive. Ideally, coordinate data should be used for accuracy (however, even coordinate data can sometimes be misleading, depending on the accuracy and quality of the original data). Descriptive data can be converted to coordinate data by consulting gazetteers. A good starting point for the collation of geographic data is GBIF. Data downloaded from GBIF can then be supplemented by consulting a range of other data sources, such as EUNIS (for Europe) and national or specialist taxonomic databases. Consult the data sources page in the Helpdesk for further information on collating distribution data. At this stage in the analysis, issues of data quality have to be taken into account and steps may need to be taken to improve the accuracy of the distribution data to remove any erroneous entries. For example, a criterion used in the AEGRO project is that only population occurrences with geographic coordinates that have two decimal digits
1 Note that if distribution data are readily available for the entire gene pool (i.e., not just the target taxa identified under step 2), diversity analysis can be undertaken for all taxa.
2 Although the actions are presented sequentially, it is not necessary to carry out ecogeographic diversity analysis before undertaking a complementarity analysis.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
13
or more are used in the analysis. Another limitation is that the availability of occurrence data may be very heterogeneous across the range of the target taxon—this needs to be taken into account when making decisions on the selection of target sites (Step 4). Where distribution data are too sketchy or otherwise incomplete or inaccurate, it may be necessary to recommend that a detailed ecogeographic survey is undertaken before further analysis.
2. Gather genetic data on the target taxa. Genetic diversity analysis is only possible where the necessary information already exists or where resources permit the generation of novel genetic diversity information. There are two types of genetic diversity information of interest for the establishment of genetic reserves and for backup in ex situ collections: intra‐population and inter‐population diversity. The precise method of generating genetic diversity information is taxon‐specific. Decisions regarding the type of genetic analysis to undertake can be based on existing studies of related taxa or taxa sharing similar biological attributes. Literature searches can be undertaken to obtain this information, as well as consulting specialist databases and taxon experts.
3. Gather ecological and environmental data on the target taxa. Ecological and environmental data associated with the target taxa can be of two types: actual (i.e., data directly linked to a taxon) or secondary (i.e., data indirectly linked to a taxon via the attributes of the site in which it is found). Actual ecological and environmental data can be sourced by obtaining characterization and evaluation data associated with ex situ accessions, and/or by consulting the available literature on the target taxon—for example, there may be published or grey literature as a result of ecological studies of the taxon or of associated taxa that occur in the same habitats—or by collecting fresh data in the field. Secondary data are obtained by gathering data associated with known locations of a taxon (e.g., climate, soil type, geological substrate, habitat type, altitudinal range and land use). Some of these data are readily available in the form of Geographical Information System (GIS) files, which are overlaid with the distribution data, and from which inferences can be made about the ecological preferences of a taxon. Consult the data sources page for further information on gathering ecological and environmental data.
4. Analyse the ecogeographic data. The data collated is now analysed to build detailed taxon ecogeographic profiles. GIS programs such as ArcGIS or DIVA GIS can be used to create distribution maps overlaid with ecological, environmental and genetic data, and locate complementary genetic reserve locations. The analysis may be simple to complex, depending on availability of data, expertise, time and resources. The data should also be imported into an appropriate information management system from which standard taxon data sheets can be extracted to form the basis of genetic reserve proposals and management plans.
Step 3 actions – b) carry out a complementarity analysis
Complementarity analysis may also be undertaken. This aims to maximize taxonomic diversity conservation in the minimum number of sites and may be useful when dealing with gene pools containing a large number of taxa or for multiple gene pools. The GIS program, DIVA GIS (see Hijmans et al., 2001) is useful for undertaking complementarity analysis and is available for download free of charge.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
14
2.4 Step 4: Selection of target sites
Background In some cases, the range of the target taxon will define the precise site or sites where active in situ conservation is needed. Obviously, a taxon that is known only to occur at one location and is considered a high priority as a potential gene donor, then that single location must be targeted for reserve establishment. Where the geographic range of the target taxon is broader, sites should be selected that represent the widest range of ecogeographic characteristics as possible.
Once the target taxon distribution has been identified and mapped, and diversity analysis undertaken (Step 3), protected area (PA) overlays are used to ascertain whether the target taxon populations occur within the boundaries of an existing PA. CWR, like any other group of wild plant species, are located both within and outside existing PAs; however, the most efficient approach in the first instance (to avoid purchase of sites to establish genetic reserves) is to establish CWR genetic reserves within existing PAs (Maxted et al., 2007). Therefore, the most appropriate PAs (e.g., national parks and heritage sites) within which to locate genetic reserves should be identified.
GIS analysis using PA shape files provides an indication of which PAs contain populations of the target taxa. In addition, this method can be used to predict which PAs contain high concentrations of CWR diversity. To be certain that the populations do exist within the PA(s), it is necessary to confirm their presence before genetic reserve establishment is recommended. This information is not always easy to obtain; however, it may be possible to contact the agency responsible for the management of the PA to see if they have an inventory of taxa available at the site or whether it is possible for site staff to confirm the presence of the taxon. If possible, ground truthing by visiting the site(s) personally should be undertaken. This is of course subject to available time and resources.
Where target taxon populations are found to already occur within existing PAs, these populations should be prioritized for inclusion in the CWR genetic reserve network on the basis that they have already been afforded some degree of protection, even if only by default. However, it is important to stress that even though a target taxon population may occur within the boundaries of a PA, this does not automatically mean that the population is actively conserved. On the contrary, few PAs are established to conserve specific target taxa, and those that have often tend to focus on animal conservation. To conserve the range of genetic diversity inherent in CWR populations, active site management and monitoring is needed (see Iriondo et al., 2011)—some PAs do not even have management plans, and those that do are often limited by financial resources and lack of capacity to put the plan into practice.
In cases where a few to several PAs are found to contain populations of a target taxon, results of the diversity analysis can be used to select sites that best represent the ecogeographic diversity within the target taxon. A further consideration for the selection of PAs is the option for multiple taxa genetic reserves. Analysis of all target taxa within the crop gene pool (and preferably across several crop gene pools) may reveal that some PAs contain populations of more than one taxon. In terms of expediency of resource use, multi‐taxa reserves have obvious advantages over those that only contain a population of one taxon.
Where target taxon populations do not already occur within existing PAs, these populations should also be prioritized for inclusion in the CWR genetic reserve network on the basis that

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
15
they have not already been afforded any degree of protection; especially for rare or threatened species. Obviously, justifying the need for and actually establishing new PAs will involve a significant initial injection of time and resources. Nomination of genetic reserves at the target locations may of course be hindered by a range of socio‐political factors, such as legal issues, land use conflicts, issues of land ownership, or lack of local support. Therefore, if possible a range of alternative sites should be recommended and ranked according to their suitability based on ecogeographic considerations.
Step 4 actions – select and prioritize target sites This is the final stage in the process, leading to the identification of CWR genetic reserves, which should ideally be assigned a priority ranking so that if for some reason a genetic reserve cannot be established at the best site (e.g., due to land use conflict or legal complications), the next best site is promoted, and so on.
1. Ascertain whether one or more populations of the target taxa occur within the boundaries of existing PAs. To achieve this, overlay target taxon distribution data with protected area shape files. A GIS program such as ArcGIS can be used for this purpose. PA data are available free of charge for use for scientific research, which are imported into the GIS program, along with the coordinate data collated under Step 3.
If one or more populations of a target taxon appear to fall within the boundaries of one or more existing PAs, follow up the analysis by verifying the presence of the taxon at the site. This is not always easy or possible, but it is important to try. If it is not possible to visit the site personally, contact the agency responsible for the management of the PA and ask if they have a recent taxon inventory available. If no recent inventory is available, ask if site staff are available to verify the presence of the taxon. If verification is undertaken by a third party it will be necessary to arrange for voucher specimens or at least photographs of the plants to be sent to ensure the correct identification has been made.
2. Select sites for the establishment of CWR genetic reserves. At this stage, there are a number of different variables to take into consideration, depending on the range of the target taxon and other factors:
For taxa that occur in only one location, it is obvious that this location must be recommended for genetic reserve establishment, regardless of whether the population exists within or outside an existing PA.
For taxa that occur in more than one location, the results of the diversity analysis carried out under Step 3 need to be added into the equation. There is no one fixed way of decision‐making at this stage, but first and foremost, since the aim is to conserve the maximum genetic diversity within and between populations of the target taxa as possible, then sites that are most likely to represent that diversity should be selected. For advice on site selection based on diversity analysis, see Maxted et al. (2004, 2008b) and Iriondo et al. (2008).
For taxa that occur both within and outside PAs, existing PAs should ideally be selected for inclusion in the CWR genetic reserve network. However, a balance will have to be met between ecogeographic suitability of sites and feasibility. In other words, if very little intra‐taxon genetic diversity is likely to be conserved by enacting active conservation of populations within existing PAs, it may also be necessary to recommend that new genetic reserves are established for other genetically distinct populations.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
16
When possible, a range of taxa should be assessed all together to try to ascertain whether it is feasible to establish multi‐taxa reserves. When focusing on a crop gene pool conservation strategy, this may not always be possible, depending on whether the target taxa are found at the same locations. The identification of multi‐taxa reserves is most likely to be achieved through the development of national CWR strategies which target a country's entire CWR flora. Therefore, during the development of a crop gene pool in situ
conservation strategy, collaboration with organizations responsible for developing national CWR conservation strategies is advised in countries where the target crop gene pool taxa are to be conserved.
The potential effects of climate change on populations of the target taxa also need to be taken into account. Considerations include the particular vulnerablity of populations in coastal and high altitude areas, whether there is sufficient intra‐population genetic diversity and reproductive success in populations to allow adaptation to new conditions, and whether small, fragmented populations with little migration will be able to colonize new sites (Veteläinen et al., 2007). In the absence of detailed studies on individual target taxa, it will not be possible to predict exactly where sites need to be established because a) we will not know whether populations of a taxon will have the ability to adapt to new conditions at current sites, b) whether populations will have the ability to migrate to new sites, and c) if migration occurs, how quickly it will take place and in what direction. However, greater emphasis on habitat protection to prevent and reduce habitat fragmentation and the establishment of corridors between habitat patches to facilitate range shifts of mobile species is likely to be important for many CWR taxa (Jarvis et al., 2008).
3. Prioritize the selected sites. The main criterion for allocating priorities to sites is the conservation of the maximum genetic diversity possible. When assigning priorities for a particular target taxon, the diversity analysis will form the basis of the priority ranking of sites. When the aim is to conserve multiple taxa within the same sites, a balance has to be met between prioritizing those sites that contain the greatest taxonomic diversity and those that contain less taxonomic diversity, but more genetic diversity specific to particular target taxa. Other factors to take into account when assigning priority ranking to selected sites include: land use, potential development pressures (e.g., sites closer to towns and cities may be less secure), presence of invasive species (particularly on islands), level and quality of site management, legal status, potential conflict with existing site management aims and social unrest. A thorough assessment of all factors, both scientific and socio‐political, must be made and considered when selecting the ideal sites.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
17
3.0 Application of the methodology to four crop case studies
3.1 Crop case studies: overview and purpose
The overarching aim of the AEGRO project is the development and promotion of CWR and landrace conservation strategies as a contribution to an integrated EU agrobiodiversity conservation strategy. To develop and test the CWR in situ conservation strategy methodology and to kick‐start the integrated EU conservation strategy for CWR, four crop case studies were selected for detailed analysis and subsequent conservation strategy planning—Avena sativa (oat), Beta vulgaris subsp. vulgaris (cultivated beets), Brassica spp. (cabbage and other brassicas) and Prunus avium (cherry). The selection of crops was endorsed by experts who are members of the European Cooperative Programme for Plant Genetic Resources (ECPGR) crop networks. Each of these crops is a high priority in economic terms for Europe (Kell et al., 2011), as well as being nutritionally important. Further, in addition to providing a vehicle for illustrating how to achieve in situ CWR conservation, these case studies represent four diverse crops with differing habits, life cycles and reproductive biology—a grain, root and leafy vegetable crop and broad‐leaved tree species; therefore, the lessons learned in conservation planning for these case studies can be applied to other crops sharing similar characteristics within each of these biological and use groups.
For each of the crop case studies, an in situ CWR conservation strategy has been developed which has resulted in the identification of CWR genetic reserve sites for high priority taxa within each crop gene pool throughout their European range, with a more detailed strategy developed at national level for Portugal for the Beta case study. Each strategy has been developed in the framework of the standard protocol presented in section 2.0.
The following sections are an analysis of the application of the methodology for each individual crop case study. The purpose of the analysis is to provide a brief commentary on the practical application of each of the four steps of the methodology with the aims of:
Revealing different perspectives on the application of the protocol by each of the crop case study experts;
Investigating ways in which the application of the individual steps may differ between the four crop groups;
Scrutinizing the methodology to confirm its applicability to a range of crop groups;
Refining the methodology to ensure that it is widely applicable to any crop gene pool and easily understood by all those involved in CWR in situ conservation strategy planning.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
18
3.2 Application of the methodology in the Avena case study
Introduction to the Avena case study
The cultivated oats (Avena sativa and A. byzantina) rank sixth among cereals in world production. Oats are more nutritious in some respects, than other EU grown cereals. The oil (8% in the seed), protein (16–18% in groats) and beta‐glucan content of oat grains are unique in composition and quality amongst cereal grains. They are also an intrinsically low input cereal crop, since their nutrient requirements are less than those for wheat or corn and are among the least demanding in regard to suitable soil type for growing. World oat production has halved in the last 30 years and now stands at about 25 million tonnes (22.7 million tonnes in 2008). While more than 75% of oat grain production is used as animal feed, human consumption of oats has more than doubled during the last decade due to its nutritional effects.
The genus Avena L. belongs to the tribe Aveneae and is comprised of a polyploid series of wild, weedy and cultivated series at the diploid (2n=2x=14), tetraploid (2n=4x=28) and hexaploid (2n=6x=42) levels. Cultivated types are present at all ploidy levels (A. strigosa at the diploid level, A. abyssinica at the tetraploid level, A. sativa and A. byzantina at the hexaploid level). All Avena species are autogamous annuals, except for A. macrostachya, which is a perennial, allogamous autotetraploid (Baum and Rajhathy, 1976), and are distributed mostly in the northern hemisphere and mainly around the Mediterranean Sea and the Canary Islands. The highest diversity is found in northern Africa (Tunisia, Algeria and Morocco), southern Spain and Sicily (Italy). All major biological species are found in Morocco except A. canariensis, A. ventricosa, A. insularis and A. macrostachya. A. agadiriana, A. atlantica and A. maroccana are endemic to Morocco. A. canariensis is unique to the Canary Islands, A. damascena to Syria and Morocco and A. insularis to Tunisia and Sicily (Italy). Avena murphyi, A. longiglumis, A. prostrata are found in restricted areas in southern Spain. Finally, A. ventricosa is found in Cyprus, while A. insularis was first discovered in Sicily (Figure 1).
The classification of Avena species has been approached either by the biological or the taxonomic concept. Baum’s monograph (1977) (taxonomic approach), based on micromorphological characters, divides Avena species into seven sections and 27 species, and this classification was later revised to also include the newly discovered species and increase the number to 31 (Legget and Thomas, 1995). The biological classification by Ladizinsky (1988) on the other hand includes only 14 species and nine synonyms. However, under this classification the hexaploids A. sterilis, A. fatua, A. byzantina, A. hybrida, A. atherantha, A. occidentalis, A. trichophylla and A. sativa are classified as A. sativa. Online databases differ in the numbers of species that are presented (e.g., in EURISCO 43 species are identified, in GBIF 71 species, in GRIN Taxonomy 83 species). It was decided in the present study to adopt the biological species classification with modifications.
European genebanks hold 34,512 accessions of Avena (EURISCO Catalogue, 2011), 162 of which are tetraploids and 194 diploids (excluding A. strigosa). Thus, wild species represent only about 1% of the total ex situ collections of Avena species, highlighting an urgent need for surveying populations and identifying sites for their in situ conservation, as well as an increase in sampling for back‐up in ex situ collections.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
19
Figure 1. Avena species distribution
Analysis of the application of the methodology in the Avena case study
Step 1: Taxon delineation
The taxonomy of Avena is either based on the morphological (Baum, 1977) or the biological species (Ladizinsky, 1988) concept. Thirty‐three species are described using the morphological concept and only fourteen using the biological concept. At the diploid level A and C genome species are present, at the tetraploid AB (AA') and AC, and at the hexaploid ACD (ACA''). All species at the hexaploid level are included in the primary oat gene pool. Most of those species are well represented in the genebanks (2456 accessions of A. sterilis, A. fatua and A. ludoviaciana) and have been used for breeding purposes. The secondary gene pool comprises three tetraploid AC genome species: A. murphyi, A. marocanna and A. insularis (totalling 58 accessions). Finally, the tertiary gene pool includes all diploid species (194 accessions, excluding A. strigosa entries) as well as the AB (AA') genome tetraploids (395 accessions).
During the Avena experts meeting in Athens (March 26–27, 2008) taxonomy and species geographical distribution within Europe was extensively discussed, in order to prioritize and select the target taxa (see Step 2).
Step 2: Selection of target taxa
Prioritization of species for in situ conservation can only be accomplished if genomic and species relationships are known. In recent years, molecular techniques have been used to reveal these relationships (Drossou et al., 2004; Nikoloudakis and Katsiotis, 2008; Nikoloudakis et al. 2008). Furthermore, gene introgression and ease of crosses between wild and cultivated material also needs to be considered. Gene flow using the secondary gene pool is partly restricted since the F1 hybrids are pentaploids and backcrossing to the cultivated parent is

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
20
needed. Also, restricted distribution and threat of specific species was taken into consideration, as well as presence of species within the boundaries of the EU.
Selection of target taxa was based on all of the above‐mentioned factors and the following species were included (in no specific order):
1. Avena murphyi (AC genome tetraploid). This species shows weaker genetic ties with the common oat than Avena insularis but is still a member of the common oat secondary gene pool.
2. Avena insularis (AC genome tetraploid). This is a recently discovered species which appears to be the tetraploid progenitor of all the hexaploid oat types, including the common oat.
3. Avena ventricosa (Cv genome diploid, present in Cyprus). Molecular data revealed that it could be the C genome donor of the hexaploids.
4. Avena hirtula (As genome diploid). This species is the wild progenitor of the slim oat, A. strigosa, present in south Europe.
5. Avena longiglumis (Al genome diploid). This species is widespread from the Middle East, through north Africa to southern Spain and is the possible donor of the A genome to the evolution of the hexaploids.
Step 3: Diversity analysis
For the diversity analysis an updated version of the European Avena DataBase (EADB) was used, containing 23,889 accessions (duplicates excluded) which is the representation of 34 Avena taxa in 81 countries. However, a number of obstacles were encountered, such as non‐consistent taxonomy, misspellings of species names, wrong coordinates and above all the poor representation of the prioritized species in ex situ collections. In the EURISCO database, which contains the EADB, there are 16 accessions of A. murphyi, seven accessions of A. insularis, 21 accessions of A. ventricosa (19 of which have been obtained through the collection organized by AEGRO in collaboration with the Avena Working Group of ECPGR), 30 accessions of A. hirtula (seven of which have been obtained through the collection organized by AEGRO in collaboration with the Avena Working Group of ECPGR, and four are duplicates), and 15 accessions of A. longiglumis; most of them rarely include coordinates. Furthermore, identification of Avena species is difficult and within species genetic variation can be revealed only by using molecular markers. In our study, ten RAPD primers were used to screen genetic diversity of A. ventricosa (34 individual plants originating from 14 sites), A. insularis (19 individual plants from five sites) and A. murphyi (27 individual plants from 15 different sites). Genetic similarities among entries within A. ventricosa ranged from 0.72 to 0.99, for A. insularis from 0.80 to 0.95, and for A. murphyi from 0.75 to 0.96.
Step 4: Selection of target sites
In order to identify and select candidate sites for in situ conservation several criteria had to be fulfilled. One of the most important criteria is to recommend a site with the highest possible amount of within species genetic diversity and a good population size. Unfortunately, the information provided by the databases for the prioritized species is restricted with incomplete passport data. It was thus decided to visit three geographic areas (Cyprus, Sicily in Italy, and Andalusia in Spain) where all of the prioritized species are present, in order to survey the population sizes and propose conservation sites. The second most important criterion was to

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
21
identify areas with the prioritized species lying within already protected sites (Natura 2000, National Parks, etc.). Cyprus was visited in 2009, and Sicily (Italy) and Andalusia (Spain) were visited in 2010. In Cyprus, three species (A. ventricosa, A. hirtula and the non‐prioritized A. eriantha) were surveyed and seeds were collected. Three sites with good sized populations were identified and selected as conservation sites; two sites are under Natura 2000 and the third one is a National Park protected by the Forest law of Cyprus. In Sicily A. insularis was collected and surveyed. This is a recently discovered species (Ladizinsky, 1998) and thus additional sites with A. insularis were recorded for the first time. Two sites (West Lake Comunelli and Mountain Giase) monitored by the Regione Siciliana, Assessorato Agricolotura e Foreste, with restricted activities, had good sized populations of A. insularis and are proposed as conservation sites. Finally, the other three prioritized species, A. murphyi, A. longiglumis and A. hirtula, are present in southwest Spain. Sites were surveyed and good sized populations of A. longiglumis and A. hirtula were found within the boundaries of the National Park of Donaña and the National Park of la Breña y Marismas del Barbate. All sites with good sized populations of A. murphyi are privately owned pasture land, although they are found inside or very close to the boundaries of two Natura 2000 sites—Los Alcornocales and Estrecho. In most cases scattered A. murphyi plants are found within fields because there is not proper management. In order to conserve this species, action needs to be taken at the regional government level by informing farmers about proper management of their fields and monitoring the population sizes.
3.3 Application of the methodology in the Beta case study
Introduction to the Beta case study
Sugar beet covered 2.1 million hectares with the EU 25 and provided approximately 1.8% of the value of agricultural production output. The reform of the EU sugar market foresees a price reduction at producer level from 631.90 €/t in 2006/2007 to 404.40 €/t from 2010/2011 onwards (European Commission, 2006). As a result of this policy sugar beet production will concentrate even more around processing plants (Isermeyer et al., 2005). In Switzerland a maximal share of 25% beets in the farmer’s crop rotation system is considered ecologically sound (Anonymous, 2010). However, because the choice of economically interesting crops is limited to a few, a relatively wide rotation system composed of winter canola, winter wheat, sugar beet and corn or winter barley may still cause yield losses in sugar beet stands by cyst nematode (Heterodera schachtii) attack or Rhizoctonia solani root rot. There is therefore a constant need for breeding of resistant varieties. The traits required by farmers depend on the specific growing conditions and vary widely. The wild relatives of sugar beet harbour the appropriate genetic variation (Frese et al., 2001a); they have been used to develop resistant varieties in the past and will be needed as donors of novel genetic variation in future to cope with resistance breaking pathotypes as well pest and diseases not yet known.
The gene pool as defined by Harlan and de Wet (1971) consists of the genus Beta (section Beta with three species and section Corollinae with four species and two hybrid species), and the genus Patellifolia with three species. The centre of diversity of Beta section Beta and the genus Patellifolia is located in Europe while Beta section Corollinae mainly occurs in Turkey and the eastern Caucasus area, except for Beta nana, which is endemic to Greece. Occurrences of Beta and Patellifolia species can be encountered in areas adjacent to Europe. All wild species are thus native to Europe and Turkey. The crop species, Beta vulgaris subsp.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
22
vulgaris was domesticated in Europe and is composed of the four cultivar groups, sugar, fodder, garden, and leaf beet and is nowadays distributed worldwide.
European genebanks hold 7965 accessions of Beta / Patellifolia of which 1767 accessions are grouped into the sample status category ‘wild’ (EURISCO Catalogue, 2010). The fraction of wild beet species accessions managed ex situ is 22.2%, which is a much higher percentage when compared with total PGRFA holdings in European genebanks—c. 9% (Dias et al., 2011).
Sugar beet breeding scientists and geneticists working in the public research sector as well as commercial sugar beet breeders are the main users of Beta / Patellifolia germplasm and the main driving force. The systematic collection of genetic resources of wild and cultivated beets started with the IBPGR funded Beta plant exploration in Greece conducted by Crombie, Cortessi and Denton in 1979. Between 1979 and 1993 various institutions conducted germplasm acquisition missions to capture genetic variation required for breeding of resistant varieties. In total, 1513 accessions were collected (Doney et al., 1995). Several collecting missions followed to close geographic gaps in germplasm holdings (Frese, 2000; El Bahloul et al., 2009). As most of the geographic distribution area of wild beets has been sampled since 1979, genebanks will phase out wild beet collecting activities.
At the last meeting of the ECPGR working group on Beta (Frese et al., 2009), the conservation objectives were directed towards in situ management of wild beets for four reasons. First, evolution of useful novel genetic variation for resistance will not occur in seed samples frozen in genebanks. Second, all species of Beta section Corollinae and all Patellifolia species are hard‐seeded—their regeneration requires hand labour and is thus expensive. Third, all species of Beta section Corollinae are not well adapted to the environmental conditions of genebank locations. The least adapted plants do not reach flowering stage and do not contribute to the next generation. This kind of site‐specific selection causes loss of genetic variation at the first reproduction cycle. Fourth, specific occurrences of Corollinae species such as Beta lomatogona in Azerbaijan and Iran (Frese, 2001b), B. macrorhiza in the Daghestan region (Frese, 2000) and B. nana on the Taygetos mountain in Greece (Frese et al., 2009) are already known to be threatened. There is therefore an urgent need to organize and establish an in situ management programme for wild beets which is complemented by ex situ backup collections.
Analysis of the application of the methodology to the Beta case study
Step 1: Taxon delineation
The beet genepool consists of two genera: Beta and Patellifolia. The genus Beta is divided into section Beta with three species and two subspecies, the section Corollinae is composed of three basic species and two hybrid species as well as B. nana (the only species of the former section Nanae). The genus Patellifolia encompasses three species. This information was available beforehand (Frese, 2010) and no further information sources had to be used to produce the species list.
Due to the detailed expert knowledge on the geographic distribution of the species, Step 1(2) was readily achieved. It was known which taxa exist within Europe and Turkey (i.e., within the geographic range of the European level of the Beta case study). For the national level case studies, the identification of taxa that occur in Portugal (three Beta and two Patellifolia species) and Germany (one Beta species) was straightforward.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
23
Step 2: Selection of target taxa
The species were organized according to their degree of relationship to cultivated beets (Step 2a(1)). Step 2a(2) was skipped as all taxa occur in Europe, as was (3) and (4) since all wild species are either known as potential donors of useful genes or have been used already in crop enhancement programmes.
It was not possible to create the priority level 2 list (Step 2b) on the grounds of IUCN Red List information. None of the Beta and Patellifolia species were Red Listed at the time of producing the Beta case study. Instead Step 2b(2) was performed and eight species were included in the priority list of the European level Beta crop case study.
Step 3: Diversity analysis
The implementation of Step 3 in a systematic way was found to be more difficult. It would have required additional scientific project staff which was not available in the AEGRO project. Sufficiently detailed information required for the implementation of Step 1 could be found in the International Data Base for Beta (IDBB – http://eadb.bafz.de/CCDB_PHP/idbb/). However, this system holds information on genebank accessions only and excludes distribution data recorded for ‘observations’ in the terminology of GBIF. GBIF, GRIN Taxonomy and IDBB data were collated and the data processed. The main obstacles to a straightforward use of these data were:
Many different taxonomic names were provided by GBIF whereby the term ‘name’ here describes any string indicating a Beta / Patellifolia species. The processing of the taxonomic information started with the collation of distinct strings in a table which was expanded by taxonomic names found in other information systems and in literature. Obvious typing mistakes such as in “Beta vulgaris var. macrorrhiza Stev.” were corrected after checking literature. Then the information contained in the string was partitioned and transferred to the appropriate descriptors such genus “Beta”, species “vulgaris”, varietas “macrorhiza”, varietas author name “Steven”. At the end of this process a table of 701 rows existed containing the original and the processed taxonomic information. This table was then used to build the three taxonomic systems offered to users of CWRIS PLIS.
The geographic information was processed in a similar way. However, the amount of data was much higher and the processing was very time consuming. The following ciphers provide an idea of the amount of work: 51,289 entries of which 24,626 are considered distinct entries; 10,096 sites descriptions of which 7413 had geographic coordinates in different formats. As for the taxonomic data the geographic data had to be normalized. The processing capacity per day is about 200 entries.
This part of the work was supported by a biolology/ bioinformatic student for one month who mainly assisted in collating data sources and in building the software tool “site converter”. One month was far from sufficient and more importantly, the production of useful datasets requires detailed expert knowledge and experience. Hence, the majority of the data processing work was done by the permanently employed scientific staff.
The normalized and harmonized data were later used to locate collecting sites or observation sites within Natura 2000 protected areas.
Step 3(2) (gather genetic data on the target taxa) was used to establish the geographic pattern of genetic diversity within species. This was achieved by referring to publications on genetic
In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
24
marker studies (e.g., Letschert, 1993; Villain, 2007; Panella et al., 2010). Steps 3(3) and (4) were skipped knowing that the University of Madrid (Universidad Rey Juan Carlos) would provide thematic data, GIS tools and GIS knowledge required for a systematic ecogeographic analysis at a later stage of the AEGRO project.
Step 4: Selection of target sites
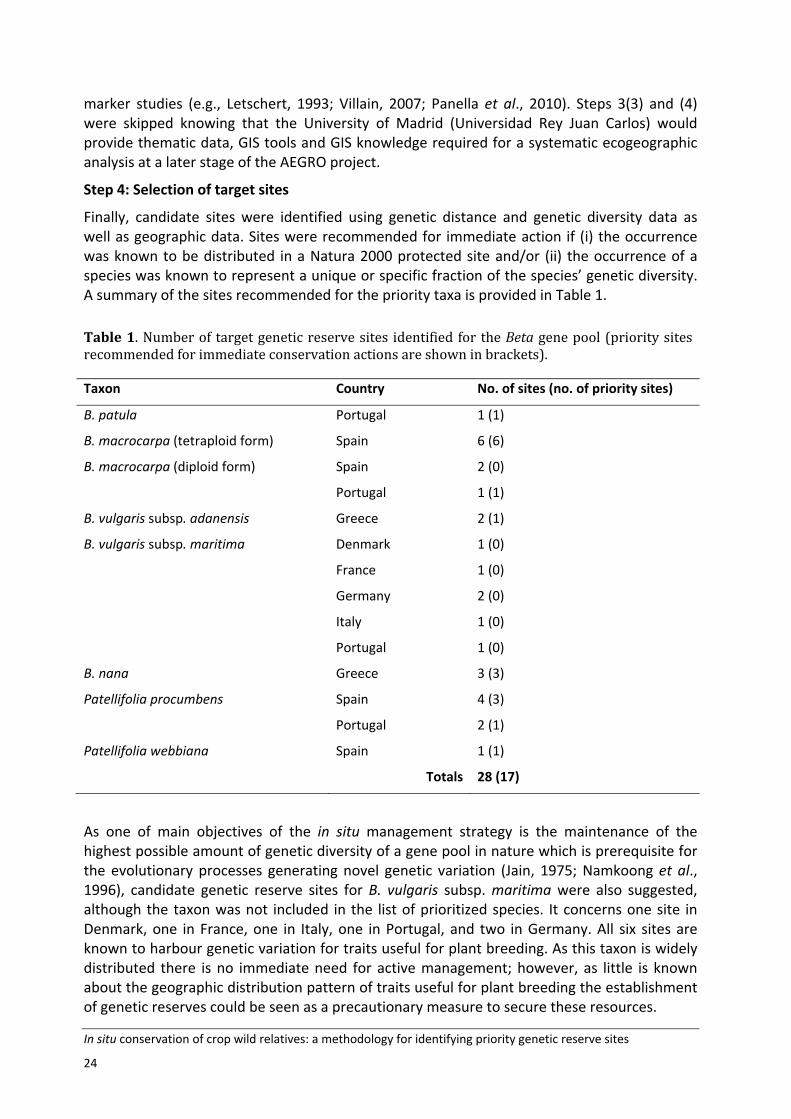
Finally, candidate sites were identified using genetic distance and genetic diversity data as well as geographic data. Sites were recommended for immediate action if (i) the occurrence was known to be distributed in a Natura 2000 protected site and/or (ii) the occurrence of a species was known to represent a unique or specific fraction of the species’ genetic diversity. A summary of the sites recommended for the priority taxa is provided in Table 1.
Table 1. Number of target genetic reserve sites identified for the Beta gene pool (priority sites recommended for immediate conservation actions are shown in brackets).
Taxon Country No. of sites (no. of priority sites)
B. patula Portugal 1 (1)
B. macrocarpa (tetraploid form) Spain 6 (6)
B. macrocarpa (diploid form) Spain 2 (0)
Portugal 1 (1)
B. vulgaris subsp. adanensis Greece 2 (1)
B. vulgaris subsp. maritima Denmark 1 (0)
France 1 (0)
Germany 2 (0)
Italy 1 (0)
Portugal 1 (0)
B. nana Greece 3 (3)
Patellifolia procumbens Spain 4 (3)
Portugal 2 (1)
Patellifolia webbiana Spain 1 (1)
Totals 28 (17)
As one of main objectives of the in situ management strategy is the maintenance of the highest possible amount of genetic diversity of a gene pool in nature which is prerequisite for the evolutionary processes generating novel genetic variation (Jain, 1975; Namkoong et al., 1996), candidate genetic reserve sites for B. vulgaris subsp. maritima were also suggested, although the taxon was not included in the list of prioritized species. It concerns one site in Denmark, one in France, one in Italy, one in Portugal, and two in Germany. All six sites are known to harbour genetic variation for traits useful for plant breeding. As this taxon is widely distributed there is no immediate need for active management; however, as little is known about the geographic distribution pattern of traits useful for plant breeding the establishment of genetic reserves could be seen as a precautionary measure to secure these resources.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
25
Additional comments about the application of the methodology
After the CWRIS AEGRO PLIS for Beta had been established it took about one week of work in total to search for occurrences and sites. This work was performed by a person knowing the crop and CWR in detail. A student was employed for one month to compile taxonomic names from existing databases, to complete this list from literature sources and harmonize the names. This kind of preparatory work took three months.
3.4 Application of the methodology in the Brassica gene pool
Introduction to the Brassica case study
The genus Brassica consists of annual, biennial and perennial species (the latter can be suffrutescent or grow as shrubs). Severe self‐incompatibility has been reported in several species (e.g., Glemin et al., 2005; Edh et al., 2009). For some Mediterranean species reports of conservation strategies have been made (Noël et al., 2010). Most species closely related to one of the economically most important brassica crops, B. oleracea, are found around the Mediterranean basin; thus, only Mediterranean species have been selected as target species in the Brassica case study, as suggested by the ECPGR Brassica Working Group (Branca and Bas, 2009). These species often grow at altitudes below 1000 m and are frequently found in coastal regions. Several species grow on limestone cliffs and rocks, in screes, gorges, quarries, scrubs or waste places.
Brassica contains major crop species such as B. napus (oilseed rape), B. oleracea (cabbage, kale, Brussels sprout, broccoli etc.) and B. rapa (turnip), as well as the minor economically important species like B. carinata (Ethiopian or Abyssinian mustard), B. juncea (Indian or Chinese mustard) and B. nigra (black mustard) (FitzJohn et al., 2007). B. nigra, B. oleracea and B. rapa are the ancestral parents of the other three mentioned brassica crops according to the triangle of U (U, 1935). According to Warwick and Sauder (2005), B. oleracea and B. rapa are closely related, whereas B. nigra is more distantly related to the two other species. Further, a number of species in the tertiary gene pool are wild‐harvested and used locally as vegetables or edible/industrial oils. Brassica crops are genetically very diverse but by including wild relatives in breeding programs, cultivars improved for a large number of specific characteristics can be developed. Warwick et al. (2009) mention male sterility, resistance to diseases, C3‐C4 photosynthetic activity, as well as tolerance to adverse abiotic conditions as potential traits that could be transferred from wild relatives to improve brassica crops. They also list wild Brassicaceae species as sources of agronomic traits such as chemical metabolites.
Brassica is included in the tribe Brassiceae that contains 50 genera and ca. 240 species; however, the inclusion of the number of genera in Brassiceae is debated (Warwick and Sauder, 2005). Sub‐tribal divisions have also been suggested but this is rather problematic (Warwick and Sauder, 2005). Based on three molecular data sets, Warwick and Sauder (2005) concluded that the genus Brassica is not monophyletic; thus, some species in other genera are more closely related to brassica crops than other Brassica species. Within the Euro‐Mediterranean region, there are 137 species within genera in the tribe Brassiceae that contain at least one species that has produced hybrids with brassica crops (Table 2).
FitzJohn et al. (2007) noted that out of 21 Brassica species included in hybridization experiments with crop brassicas, B. balearica was the only one not producing hybrids, and for that species only a single experiment was performed. Warwick et al. (2009) also review inter‐specific and inter‐generic hybridizations. From these two review papers, species of Capsella,

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
26
Coincya, Crambe, Diplotaxis, Enarthrocarpus, Eruca, Erucastrum, Hirschfeldia, Moricandia, Ochryphragmus, Physaria, Raphanus, Rapistrum, Rorippa, Sinapidendron and Sinapis have produced hybrids after hybridizations with crop brassicas. In some of the hybridization experiments, in vitro culture of immature embryos was necessary to obtain hybrids—hybrids with further genera (Arabidopsis, Armoracia, Barbarea, Camelina, Descurainia and Trachystoma) have been produced by protoplast fusion (Warwick et al., 2009).
Table 2. Brassiceae species found within the Euro‐Mediterranean region in genera in which at least one species has produced hybrids with brassica crops (FitzJohn et al., 2007; Warwick et al., 2010). Occurrences compiled from Kell et al. (2005).
Genus Within Europe and Macaronesia Outside Europe but within the Mediterranean region
Brassica balearica, barrelieri, cadmea, cossoniana, cretica, elongata, incana, gravinae, hilarionis, incana, insularis, juncea, macrocarpa, maurorum, montana, nigra, nivalis, oxyrrhina, procumbens, rapa, repanda, rupestris, souliei, tournefortii, villosa
deflexa, desertii, desnottesii, dimorpha, loncholoma, setulosa, spinescens
Capsella bursa‐pastoris, grandiflora, heegeri, orientalis, rubella
Crambe abyssinica, arborea, cordifolia, filiformis, fruticosa, gigantea, gomerae, hispanica, laevigata, maritima, microcarpa, pritzelii, scaberrima, scoparia, strigosa, sventenii, tataria
alutacea, armena, aspera, auch‐eri, gibberosa, glaberrima, gran‐diflora, juncea, koktebelica, kra‐likii, orientalis, parviflora, steveni‐ania,
Diplotaxis
catholica, erucoides, gomez‐campoi, harra, ibi‐censis, ilorcitana, muralis, parvula, siettiana, sii‐folia, tenuifolia, vicentina, viminea, virgata, wirt‐geni
acris, assurgens, berthautii, cretacea, duveyrieriana, ollivieri, pitardiana, simplex, tenuisiliqua
Eruca pinnatifida, vesicaria
Erucastrum canariense, cardaminoides, gallicum, nasturtiifolium, palustre, varium, virgatum
arabicum, armoracioides, brevirostre, elatum, ifniense, leucanthum, littoreum, rifanum
Raphanus raphanistrum, sativus rapinastrum
Rorippa amphibia, anceps, armoracioides, astyla, austri‐aca, barbareoides, erythrocaulis, hispanica, hun‐garica, islandica, kerneri, lippizensis, palustris, prolifera, pyrenaica, sylvestris, thracica, valdes‐bermejoi
aurea, dogadovae, hayanica, indica, integrifolia
Sinapis alba, arvensis allioni
It has been possible to make successful crosses between species and genera with different chromosome numbers and a large proportion of attempted combinations have been successful even under natural conditions. The gene pool of Brassica is therefore very large and

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
27
all Brassica species seem to be within the primary gene pool of at least one of the brassica crops. Further, a number of other Brassicaceae genera could be included in the primary gene pool of brassica crops; for example, genera successfully hybridized with brassica crops include Diplotaxis, Eruca, Erucastrum, Raphanus, Rapistrum, Rorippa and Sinapis.
Analysis of the application of the methodology in the Brassica case study
Step 1: Taxon delineation
Brassica crops include several species (e.g., B. juncea, B. napus, B. oleracea, B. rapa, B. carinata and B. nigra) and by consulting the literature—especially FitzJohn et al. (2007) and Warwick et al. (2009)—it is found that the number of genera and species in the crop gene pool is very large and may consist of the whole tribe Brassiceae (50 genera and 240 species). Further, the number of synonyms is also very large. Warwick et al. (2009) list approximately 900 taxa epithets within Brassiceae and the Crop Wild Relative Catalogue for Europe and the Mediterranean includes 250 Brassica epithets.
In this case study, only species within the genus Brassica were selected as they are the ones that are most closely related to the target crop, B. oleracea. Within the EU, 26 Brassica species occur and including the EU and Mediterranean area, a further seven species occur.
For determination of relevant taxa, CWRIS, GBIF, Snogerup et al. (1990), Kell et al. (2005), FitzJohn et al. (2007) and Warwick et al. (2009) were consulted.
Step 2: Selection of target taxa
Very little and scattered information of relevance with respect to the use of specific species in breeding programs of brassica crops is available, although Warwick et al. (2009) report a list of characters in specific genera and species that might be of interest in future breeding. Within Europe a number of species could be selected as target taxa due to limited distribution areas; these are: B. cretica subsp. cretica, B. cretica subsp. laconica, B. incana, B. insularis, B. montana, B. balearica, B. bourgeaui, B. cadmea, B. gravinae, B. hilarionis, B. macrocarpa, B. nivalis, B. procumbens and B. tyrrhena. As examples for the case study, B. cretica subsp. cretica, B. cretica subsp. laconica, B. incana, B. insularis, B. montana were selected. The main reasons for the selection were limited distribution areas and available literature with respect to genetic diversity.
Step 3: Diversity analysis
CWRIS‐AEGRO, GBIF and Snogerup et al. (1990) were used to source distribution data of the selected target species. Within GBIF some of the occurrences of some species had obvious wrong coordinates (e.g., occurrences of B. bourgeaui and B. balearica were placed in the Atlantic and Mediterranean Sea, respectively). Thus, other occurrences may also be questionable. Snogerup et al. (1990) was mainly used for area descriptions and data from the Danish Metrological Institute are used for climate data. For some taxa, papers describing distribution and diversity within and between populations also included relevant ecogeographic and genetic data. Snogerup et al. (1990) include morphological descriptions of the species and thus also data indicating genetic variation within species.
Step 4: Selection of target sites
The very thoroughly but relatively old paper by Snogerup et al. (1990) indicates interesting target sites. Already at that time some interesting and large sites were threatened by urbanization and introgression from garden plants. More recent papers indicate limited gene

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
28
flow between populations for several species and self‐incompatibility; thus, many sites have to be selected to secure a large proportion of the genetic diversity.
For each of the selected target taxa the number of target sites are here given in parentheses: B. cretica subsp. cretica (3), B. cretica subsp. laconica (4), B. incana (4), B. insularis (3) and B. montana (5).
3.5 Application of the methodology in the Prunus case study
Introduction to the Prunus case study
Prunus avium has great economic importance as a crop and in its wild form also has value for crop improvement and for breeders of forest, fruit and ornamental trees. The wood of wild cherry has an attractive pattern, and is second only in value to walnut (CABI, 2005). It is hard, close‐grained, solid and durable (Hedrick, 1915). For several centuries, it dominated the style of furniture making (CABI, 2005) and this trade is still the main user of the timber. As a noble hardwood, it is still valued today for panelling and cabinet making (Diaz et al., 2007), but in Britain, wild cherry has variable demand and is thought inferior compared to other hardwoods such as oak and ash (Nicoll, 1993).
P. avium is increasingly planted in Europe, both in afforestation of abandoned farmland and enrichment of forests, due to its fast growth (rotation of 50 to 60 years) (Santi et al., 1998; Kleinschmit et al., 2006). Kobliha (2002) described the beginnings of a wild cherry breeding program for Czech forestry for example, where progeny and clonal tests, seed orchards and clone archives were established.
P. avium also has medicinal value—it can be used as an antitussive (prevents or relieves cough), astringent, diuretic, and tonic (PFAF, 2008). Other uses include: alcoholic beverages, honey, digestive aid, sedative, antihistamine, haemorrhoid treatment, bladder infection relief, rheumatism treatment and antispasmodic drugs (Vivero et al., 2001). P. avium stems have been used by herbalists in Europe for diuretic and astringent purposes; other medicinal uses include treatment for cystitis, nephritis and urinary retention, and arthritic problems, notably gout (NHM, 2007).
Wild cherry is native to Europe, western Asia and northern Africa, and has been introduced into many countries (Vivero et al., 2001). Its latitudinal range is from approximately 30°– 61° N and is thought to have originated in the Caucasus and surrounding area (Faust and Suranyi, 1997) with the centre of origin being Asia with Europe as a secondary centre (Watkins, 1995). Sub‐fossil evidence indicates that it is native to northwest and central Europe (Russell, 2003), and fossil remains in Britain confirm the existence of preglacial populations, natural populations having established in the southern half of Britain around 8000 years ago (White, 1995), after oak and ash. It is found growing freely almost throughout Britain (Rackham, 1980) but wild populations have a scattered distribution and extended natural populations are rare (Russell, 2003).
P. avium occurs abundantly on forest slopes of southern, central and western Europe (OECD, 2006), has long grown spontaneously over a wide area, and may have been domesticated in several widely separated regions (Hedrick, 1915). In their study of eight Prunus species in Andalusia, Vivero et al. (2001) suggested that the Iberian Peninsula was probably a refugium for the species and for this reason, the scattered populations are considered relict and declining; it is common in the north of the Peninsula and rarer in the south.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
29
P. avium can be found growing across the temperate zones of Europe with a maximum altitude of about 1900 m reported in France; in Britain it is essentially a lowland species (Russell, 2003) and in Central Europe it is generally found at lower altitudes (Gomory, 2004). In Andalusia, Vivero et al. (2001) found scattered populations of wild cherry over several mountains at elevations of 1300–1800 m. Due to summer drought in this region, it is found in wet ravines of mountain ranges where the effects of the drought are lessened. A combination of already long, dry summers and increasing temperatures may further restrict the range of the species (Vivero et al., 2001).
Reproduction in P. avium occurs asexually via suckering, and sexually through insect‐mediated pollination and subsequent seed dispersal by birds and mammals (Vaughan et al., 2007). It is a typical pioneer plant favouring plenty of light and little competition (Bratberg and Vestrheim, 2004) and is a colonizing species of early forest successional stages due to its capacity for rapid vegetative propagation (Gomory, 2004) and when large gaps appear, such as after disturbance.
This species’ asexual reproduction is a complementary adaptational strategy; it does not reproduce easily from seed (Holtken and Gregorius, 2006) and seed dispersal is somewhat limited (Vaughan et al., 2007), despite its importance in gene flow. Pollination is entomophilous and seeds are ingested and dispersed by animals (Mariette et al., 1997), especially birds, such as pigeons, thrushes, starlings and jays, and by small mammals (Russell, 2003).
A working group for Prunus was established in 1983, emanating from the European Cooperative Programme for Crop Genetic Resources Networks (ECP/GR)3. A paper by Zanetto et al. (2002) details the Group’s activities over the previous 20 years, describing Prunus genetic resources management and the establishment of the European Prunus Collection. One of the Group’s first objectives was to produce a database documenting Prunus accessions of European collections. The European Prunus database (EPDB) is maintained by the National Institute for Agronomical Research (INRA) in Bordeaux, France. The broader Prunus National Network in France includes 13 partners under the direction of the Genetic Resources Office, BRG (Bureau des Ressources Génétiques), including INRA, the Conservatoire Botanique National Méditerranéen de Porquerolles, Centre Technique Interprofessionnel des Fruits et Légumes (CTIFL) and Association Française pour la Conservation des Espèces Végétales (AFCEV). The Prunus European Network consists of 32 Member States implied within the framework of the ECPGR, of which Unité de Recherches sur les Espèces Fruitières (UREF) is in charge of the database development and management (Dosba and Zanetto, 2004).
Analysis of the application of the methodology in the Prunus case study
Step 1: Taxon delineation
The initial part of this step was followed in that a formal taxonomy was selected and a list of taxa within the genus Prunus was compiled as part of an MS Access database produced by C. Germeier (AEGRO workpackage 10). The genus Prunus is large and complex and estimated to contain between 200 (NHM, 2007) and 430 species (Campbell et al., 2002). However, little agreement could be found regarding the taxonomic position of Prunus avium. In older classifications, where emphasis was placed upon fruit or stone instead of flower
3 Now the European Cooperative Program for Plant Genetic Resources (ECPGR – www.ecpgr.cgiar.org/).

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
30
characteristics, both sweet and sour cherries were located in the genus Cerasus; however, the ‘true’ cherries are now considered by most botanists to be in the genus Prunus (Webster, 1996). An exception to this is in countries such as Russia and China where P. avium is known as Cerasus avium. However, a general confusion over the habit of suckering arises; the Flora of China (eFloras, 2003) describes C. avium, with P. avium L. as its synonym, but as a “tree to 25 m tall, producing no suckers”.
Some closely related species were considered for more in‐depth study at this stage but, due to taxonomic complexity, as indicated above, the decision was taken to continue working on Prunus avium only, as defined in the AEGRO workplan.
Step 2: Selection of target taxa
As the taxon focused upon in this workpackage, the prioritization of wild Prunus avium for active conservation can be explained as follows:
P. avium is one of three parent species of the commercially valuable sweet cherry and for this reason is also under pressure from introduction of genetic material from cultivated forms.
It is a European native and maintains a broad distribution across the wider continent.
The species has a long and complex socio‐economic and cultural history.
It is a valuable component of mixed broadleaved woodland and is an important element of biodiversity in its habitat—particularly for insects.
The taxon under consideration in this case is not under immediate threat in Europe, except by way of the increased introduction of cultivated material in the south of Spain, this species has been Red Listed as Vulnerable (VU) (Gavira, 2006).
Step 3: Diversity analysis
The same process as that outlined in Step 3 of the methodology was used but with some differences. Passport data were downloaded from GBIF, EURISCO, and PC‐GRIN. When downloading information from GBIF, many synonyms were already included within a Prunus avium search, but when downloading from EURISCO, it was also necessary to include Cerasus avium specifically. The initial Prunus database, which contained 129,788 records, was queried to exclude cultivated forms and non‐European countries; taxa were also included when found to be synonymous with Prunus avium. Data were then extracted from the initial database and transferred to a new database specifically for Prunus avium.
An initial distribution map was produced to highlight any outliers, which were then checked and amended. Some of the database records were only partially complete and approximately 400 were without coordinates. When only partial location data are available, it is still possible to allocate a set of coordinates, to a certain accuracy, using a variety of methods.
A range of webpages was used to georeference those accessions without coordinates (see Table 2). Initially, approximate coordinates were obtained using a bulk geocoder, such as GPS Visualizer (Schneider, 2009). These were then used to locate the site in Google Earth (Google, 2009). If this was incorrect, and also to confirm the location, the National Geospatial‐Intelligence Agency (NGA) (2009) website was used to check how many places had the same name; if more than one place existed, the information for the next administrative level was used.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
31
Table 2. Sources used for georeferencing incomplete accessions
Source URL
BatchGeo http://batchgeo.com/
Falling Rain www.fallingrain.com/world/
GeoNames (GNA) www.geonames.org/
Google Earth earth.google.co.uk
Google translate http://translate.google.co.uk/?hl=en#
GPS Visualizer www.gpsvisualizer.com/
Herbaria United (collector information) herbariaunited.org/
Ordnance Survey www.ordnancesurvey.co.uk/
Gazetteer for Scotland www.scottish‐places.info/
Wikipedia http://en.wikipedia.org/wiki/Main_Page
There were some intrinsic errors in the data which meant that in some cases, coordinates could not be confidently applied—some records did not have any information below country level and for others the site converter had wrongly applied location information—so these records were manually georeferenced in the same way as the other accessions. Altitudes were calculated using coordinate data through the GPS Visualizer website (Schneider, 2009) and then a selection of these were cross‐checked using Google Earth (Google, 2009). In order to remove duplicates, a query was written to remove records which had the same coordinates along with the same collection date.
Coordinates were extracted from the database and entered into a GIS; the data were analysed based primarily on their geographic distribution using ArcGIS v 9.3 (ESRI, 2009). Points were imported from a text file and plotted, and a mask was produced to delimit the area of Europe under consideration on a map of the world; this was used to display all maps. DIVA GIS (Hijmans et al., 2009) was also used to map and analyse spatial data, providing a predicted species distribution based on the climatic envelope of the point data provided. The coordinate system used was WSGS84 and all data were converted to this before mapping. Further observational and historical records have been incorporated into the database and a sub‐sample of the resulting approximately 18,000 accessions were selected using the R programming language (R Development Core Team, 2010) to obtain a more spatially balanced sample for use in species distribution modelling.
The species distribution modelling work is in addition to the suggested methodology, but is a direct result of the problems of using historical data. A niche‐based model provides an approximation of the species’ niche. When, as in this project, only incomplete occurrence data are available, a model that only requires partial occurrence information is very valuable (Graham et al., 2004). Historical collections only record locations where the species has been observed and contain no information on absences (Mateo et al., 2010). Although hard to find, reliable absence data can be used to identify the effects of any constraints of environmental variables; restricting model predictions, preventing inclusion of sites where it is known that the species does not occur (Jiménez‐Valverde et al., 2008). It is also possible to incorporate

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
32
pseudo‐absences or target group background points when using presence‐only methods (Mateo et al., 2010). The data were then combined with environmental variables that describe some of the influences on the species in that environment. These samples of the realised niche (where a species lives) are a portion of fundamental niche (where a species could live), actually occupied, and may be smaller than its fundamental niche due to human or biotic factors). Combined with the model, samples were used to predict environmental suitability as a function of the examined environmental variables (Phillips et al., 2006). Information on environmental variables was obtained from a variety of sources, such as the European Environment Agency (EEA, 2010), the National Geosphysical Data Centre of the US (NGDC, 2011) and Geonetwork (Geotnetwork, 2011); raster files were downloaded and manipulated for use in the modelling software. Despite being geographically biased to certain countries and species, historical and museum data are still the most useful source of information in the majority of cases and with broad temporal coverage, species distribution models can be used to infer changes over time, particularly with changing climate conditions (Newbold, 2010).
In terms of how the diversity analysis informs the conservation strategy, the biased nature of a subset selected from the Prunus database highlights the need for a more detailed modelling approach in order to fully incorporate ecogeographic diversity.
Step 4: Selection of target sites
In the case of Prunus avium, protected sites containing this species have already been the subject of genetic research in particular and were already known to the group (AEGRO); for the initial recommendations for reserves, therefore, these locations were obvious first choices for genetic reserves, notwithstanding the fact they had not been selected using the current protocol.
The outcomes of the Prunus case study are two‐fold. Firstly, using SDM, sites can be selected as genetic reserves for P. avium and given priority because they have the optimum combination of suitable variables affecting the species’ distribution, then potential distribution of this species can be modelled with a view to predicting sites for introduction, where other factors are favourable for the long‐term conservation of this (and potentially incorporating other) species (e.g., as influenced by climate or land use change). Secondly, the predictor variables influencing the distribution of Prunus avium can be elucidated, apropos of the samples selected.
SDM provides an approach that can help extend the usefulness of direct observation and improve our interpretation and understanding of species distributions. In addition to final suggestions made for reserve site selection, an important outcome of this study is the recognition of the need for data quality checking, and the importance of acknowledging the limitations of the chosen variables and field data, and their spatial attributes.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
33
4.0 Discussion
In this report, we have proposed a methodology for the identification of genetic reserves for a target crop gene pool which addresses the need to conserve the maximum range of genetic diversity in the highest priority taxa in terms of their known or potential value for crop improvement and relative threat. This approach has now been applied to the gene pools of four crops important for food security and economic stability in Europe—Avena sativa (oat), Beta vulgaris subsp. vulgaris (cultivated beets), Brassica spp. (cabbage and other brassicas) and Prunus avium (cherry). The methodology has also been applied outside of Europe for a number of globally important crops (see Maxted and Kell, 2009). We have found that the model is relevant to a range of crop gene pools; however, it is clear that its application will necessarily be slightly adapted according to the different cases to which it is applied.
With regard to target taxon selection, firstly, the number of species in the gene pool may affect the chosen method because when dealing with a large number of species it is not always possible to obtain distribution data for all taxa in the gene pool, depending on the availability of data and resources for the actual research. For example, Brassica is a large genus compared with Avena, Beta and Patellifolia and species in a number of closely allied genera may also be important gene donors to brassica crops. This means that when dealing with a relatively large crop gene pool, it is likely that the target taxa will initially be selected on the basis of their degree of relationship to the crop and known potential as gene donors rather than on relative distribution or threat. On the other hand, when dealing with smaller gene pools such as sugar beet and oats (which are also well‐studied gene pools), both relative threat and potential as gene donors can be used in parallel to select target taxa. Secondly, knowledge of the genetic relationship between taxa varies between crop gene pools and this means that in cases where genetic information is lacking, it is not possible to apply the Gene Pool (GP) concept. For example, there is better knowledge about the genetic relationship between taxa in the beet and oat gene pools than for the brassica gene pool. In the absence of genetic data, the Taxon Group concept can however be applied as a proxy for the GP concept. Thirdly, knowledge of the breeding potential of species varies between crop gene pools; for example, there is better knowledge for the beet and oat gene pools than for the brassica gene pool. This implies that in cases where there is good knowledge of breeding potential, target taxa can be selected on the basis of their potential use for crop improvement with a degree of confidence, while for gene pools where this knowledge is lacking, important taxa with as yet unknown potential could be missed in conservation planning. This highlights the importance of using relative threat as a criterion for target taxon selection as taxa with limited distribution ranges or otherwise threatened but with unknown potential as gene donors will still be included in the conservation strategy.
The number of crops in the gene pool is also likely to affect the selection of target taxa if a multi‐crop approach is taken. For example, the genus Brassica contains several crops and therefore the delineation of the gene pools of the different crops will vary. While one taxon may be a primary wild relative of one brassica crop, it may be a secondary or tertiary wild relative of another. However, this does not affect the application of the methodology as it is based on the premise of developing an in situ conservation strategy for a single crop gene pool. If multiple crops are targeted at the same time, whether or not these species are within the same genus, the methodology has to be applied to individual crop gene pools independently of one another. However, if dealing with multiple crops in the same genus or

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
34
closely allied genera, it will be beneficial to synthesize the results of each gene pool analysis to identify overlapping target taxa and sites. This approach was already taken to some degree in the AEGRO project under Workpackage 9 as a detailed ecogeographic analysis of the beet gene pool was undertaken and then the results were synthesized to identify multi‐taxa reserves, including target taxa from the other crop gene pools that were the subject of this project (see Parra‐Quijano et al., 2011).
The means of selecting target sites also varies from one gene pool to another, depending on existing knowledge of intra‐ and inter‐specific genetic diversity of target taxa and of the localities of target species (e.g., population size, threats and suitability of the site to establish a GR). Obviously, the better the knowledge base, the more sound a basis there is for conservation planning. However, even without genetic diversity knowledge, the methodology can be applied by using other types of ecogeographic data as a proxy for genetic data; for example, this has been successfully demonstrated in the Beta crop case study (see Parra‐Quijano et al., 2011). Furthermore, both genetic knowledge and ecogeographic analysis can be used in tandem to identify target sites—this is likely to result in the most comprehensive in situ conservation strategy plan for a target crop gene pool.
The application of the methodology also highlighted the difficulty of dealing with taxonomic data (i.e., different nomenclature in different information systems) and occurrence data (e.g., lack of coordinates, problems of data quality, and evenness of data availability or knowledge of distribution across Europe); however, these are challenges that we face in conservation planning in general and are not specific to this model. Means of overcoming these problems were applied in this project at different stages. Firstly, the site converter software was developed to improve the usability of descriptive location data and applied to distribution data collated for all of the crop case studies. Secondly, in the Prunus case study, spatially balanced sampling was used to reduce the effect of the bias in the data; this was followed by species distribution modelling to interpolate new data points within the range of the known distribution data. Thirdly, a method of improving coordinate data accuracy was applied in the Beta case study by filtering out coordinates with less than two decimal digits.
The Avena case study also highlighted the importance of fresh data collection through field visits to carry out population surveys. While this is not always possible when planning genetic reserve conservation, it is highly recommended when resources permit.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
35
5.0 Conclusion
Crop wild relatives contain a wide pool of genetic diversity that is important to maintain for its use in plant breeding for crop improvement. The highest priority CWR for food security are not adequately conserved, either in situ or ex situ, and we cannot rely only on ex situ conservation of these resources as it does not maintain the evolutionary process of adaptation found in wild populations—this can only be achieved by managing in situ populations in CWR genetic reserves. The value of CWR for food security and the need for their conservation has recently been placed firmly on the international conservation agenda, but due to the large number of taxa that exist, coupled with limited resources, a means of setting priorities for their conservation is needed. Further, as CWR have largely been neglected by the conservation community, we face the challenge of identifying where and how to conserve them in situ. A logical and systematic framework for CWR conservation is needed that is applicable to any country or region and to any crop gene pool. This may involve both the floristic and monographic approaches, but in order to conserve the maximum range of genetic diversity in the highest priority crop gene pools for global, regional and local food security, a crop gene pool approach is needed that can be applied in tandem with the floristic approach at national level.
In this report, we have presented a methodology for the identification of genetic reserves for a target crop gene pool which addresses the need to conserve the maximum range of genetic diversity in the highest priority taxa in terms of their known or potential value for crop improvement and relative threat. By applying the methodology to the gene pools of a range of different crop types—a cereal, leafy vegetable, root crop and broad‐leaved tree, it has been possible to a) reveal different perspectives on its application by several experts, b) investigate ways in which the application of the individual steps may differ between different crop groups, c) scrutinize the methodology to confirm its applicability to a range of crop groups and d) refine the methodology to ensure that it is widely applicable to any crop gene pool and easily understood by all those involved in CWR in situ conservation strategy planning. Most importantly, it was found that the methodology is relevant to all the crop gene pools it was applied to, although the approach to each step varied slightly according to the specific characteristics of the gene pool (e.g., number of species) and the extent and quality of data available (e.g., distribution, genetic diversity and potential use).
The methodology can now be applied to develop conservation strategies for more priority crop gene pools with the aim of eventually ensuring that the genetic diversity that we may rely on in years to come is secured in a network of national, regional and global CWR genetic reserves. In Europe, this work is being taken forward in the context of the EU Framework 7 project, PGR Secure. The case studies themselves have resulted in the identification of target genetic reserves for four crop gene pools which are socio‐economically important in Europe. The next step will be to put in place the practical and legal requirements to establish the sites and ensure their long‐term sustainability.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
36
References
Baum, B.R. (1977) Oats, Wild and Cultivated: A Monograph of the Genus Avena L. (Poaceae). Biosystematics Research Institute, Canada Department of Agriculture. 463 pp.
Baum, B.R. and Rajhathy, T. (1976) A study of Avena macrostachya. Canandian Journal of Botany 54, 2434–2439.
Branca, F. and Bas, N. (2009) Working group on Brassica. In: Astley, D., Bas, N., Branca, F., Daunay, M.C., Díez, M.J., Keller, J., Van Dooijeweert, W., van Treuren, R., Maggioni, L. and Lipman, E. (compilers), Report of a Vegetables Network, Second Meeting, 26–28 June 2007, Olomouc, Czech Republic. Bioversity International, Rome, Italy. Pp. 14–19.
Bratberg, E. and Vestrheim, S. (2004) Sweet cherries in Norway. In: Dille, L.L. (ed.), Nordic Gene Resources 2004. Nordic Council of Ministers, Copenhagen, 22–23.
CABI (2005) Prunus avium. Forestry Compendium. CAB International, Wallingford, UK.
Campbell, C.S., Kellogg, E.A., Stevens, P.F. and Donoghue, M.J. (2002) Plant Systematics: a Phylogenetic Approach (2nd ed.). Sinauer associates, Inc., Massachusetts, USA.
Deryng, D., Sacks, W.J., Barford, C.C. and Ramankutty, N. (2011) Simulating the effects of climate and agricultural management practices on global crop yield. Global Biogeochemical Cycles, 25, GB2006, doi:10.1029/2009GB003765.
Dias, S., Dulloo, M.E. and Arnaud, E. (2011) The role of EURISCO in promoting use of agricultural biodiversity. In: Maxted, N., Dulloo, M.E., Ford‐Lloyd, B.V., Frese, L., Iriondo, J.M. and Pinheiro de Carvalho, M.A.A. (eds.) Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces. CAB International, Wallingford.
Diaz, R., Zas, R. and Fernandez‐Lopez, J. (2007) Genetic variation of Prunus avium in susceptibility to cherry leaf spot (Blumeriella jaapii) in spatially heterogeneous infected seed orchards. Annals of Forest Science 64, 21–30.
Doney, D.L., Ford‐Lloyd, B.V., Frese, L. and Tan, A. (1995) Scientists worldwide rally to rescue the native beet of the Mediterranean. Diversity 11(1 and 2), 124–125.
Dosba, F. and Zanetto, A. (2004) The Prunus European Cooperative Programme for Genetic Resources: a networking actitvity for the European Prunus Database and the challenge for European collections. Journal of Fruit and Ornamental Plant Research 12, 77–85.
Drossou, A., Katsiotis, A., Leggett, J.M., Loukas, M. and Tsakas, S. (2004) Genome and species relationships in genus Avena based on RAPD and AFLP molecular markers. Theoretical and Applied Genetics 109, 48–54.
Duveiller, E., Singh, R.P. and Nicol, J.M. (2007) The challenges of maintaining wheat productivity: pests, diseases, and potential epidemics. Euphytica 157, 417–430.
Edh, K., Widen, B. and Ceplitis, A. (2009) Molecular population genetics of the SRK and SCR self‐incompability genes in the wild species Brassica cretica (Brassicaceae). Genetics 181, 985–995.
EEA (2011) Data and Maps [web document]. European Environment Agency. Available at: http://www.eea.europa.eu/data‐and‐maps/ (accessed 10 March 2010).

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
37
eFloras (2003) Flora of China Vol. 9. Rosaceae, Subfam. Prunoideae [web document]. http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=20512 (accessed 24 June 2011).
El Bahloul, Y., Van Cutsem, P. and Sadiki, M. (2009) Survey of wild Beta genetic resources. In: L. Frese and L. Maggioni (compilers), Report of a Working Group on Beta and World Beta Network, Third Joint Meeting, 8–11 March 2006, Puerto de la Cruz, Tenerife, Spain. Pp. 105–106.
Elzinga, C.L., Salzer, D.W. and Willoughby, J.W. (1998) Measuring and Monitoring Plant Populations. BLM Technical Reference 1730–1. Bureau of Land Management, Denver, Colorado.
ESRI (2009) Overview. What is GIS? [web document]. Environmental Systems Research Institute. http://www.esri.com/what‐is‐gis/index.html (accessed 24 June 2011).
EURISCO Catalogue (2010) http://eurisco.ecpgr.org (accessed 08 July 2010)
EURISCO Catalogue (2011) http://eurisco.ecpgr.org (accessed 01 February 2011)
European Commission (2006) The European Sugar Sector. A long‐term competitive future.
Faust, M. and Surányi D. (1997) Origin and dissemination of cherry. Horticultural Reviews 19, 263–317.
FitzJohn, R.G., Armstrong, T.T., Newstrom‐Lloyd, L.E., Wilton, A.D. and Cochrane, M. (2007) Hybridization within Brassica and allied genera: evaluation of potential for transgene escape. Euphytica 158, 209–230.
Ford‐Lloyd, B.V., Maxted, N. and Kell, S.P. (2008) Establishing conservation priorities for crop wild relatives. In: Maxted, N., Ford‐Lloyd, B.V., Kell, S.P., Iriondo, J., Dulloo, E. and Turok, J. (eds.), Crop Wild Relative Conservation and Use. CABI Publishing, Wallingford. Pp. 110–119.
Ford‐Lloyd, B.V., Maxted, N. and Kell, S. (2009) Prioritization of wild Beta species for conservation: the PGR Forum experience. In: L. Frese and L. Maggioni (compilers), Report of a Working Group on Beta and World Beta Network, Third Joint Meeting, 8–11 March 2006, Puerto de la Cruz, Tenerife, Spain. Pp. 27–30.
Frese, L. (2000) Expedition in den Kaukasus: Sammelreise zu den Vorfahren der Zuckerrübe Forschungs Report 1. Pp. 13–17.
Frese, L. (2010) Conservation and access to sugarbeet germplasm. Sugar Tech 12 (3/4), 207–219.
Frese, L., Desprez, B. and Ziegler, D. (2001a) Potential of genetic resources and breeding strategies for base‐broadening in Beta. In: Cooper, H.D., Spillane, C. and Hodgkin, T. (eds.), Broadening the Genetic Base of Crop Production. FAO, IBPRGI jointly with CABI Publishing, Rome, Italy. Pp. 295–309.
Frese, L., Akbarov, Z., Burenin, V.I., Arjmand, M.N. and Hajiev, V. (2001b) Plant exploration in the Talysch Mountains of Azerbaijan and Iran. Plant Genetic Resources Newsletter 126, 21–26.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
38
Frese, L., Maggioni, L. and Lipman, E. (eds.) (2009) Report of the ECPGR Beta Working Group and World Beta Network. Third joint meeting 8–11 March 2006, Puerto de la Cruz, Tenerife, Spain. Bioversity International, Rome, Italy.
Gavira, O.N. (2006) Análisis de un corredor florístico entre los Parques Naturales Sierra de Grazalema y Sierra de las Nieves (Cádiz y Málaga, España). Anales de Biología 28, 45–72.
Geonetwork (2011) Dataset Collections [web document]. http://www.fao.org/geonetwork/srv/en/main.home (accessed 11 August 2009).
Glemin S., Petit C., Maurice S. and Mignot A. (2008) Consequences of low mate availability in the rare self‐incompatible species Brassica insularis. Conservation Biology 22, 216–221.
Gomory, D. (2004) Mutual links of demographic and genetic processes in a wild cherry population during the colonization. Biologia 59(4), 493–500.
Google (2009) Google Earth [web document]. http://earth.google.co.uk/ (accessed 10 February 2008).
Graham, C. H., Ferrier, S., Huettman, F., Moritz, C. and Peterson, A.T. (2004) New developments in museum‐based informatics and application in biodiversity analysis, Trends in Ecology and Evolution 19, 497–503.
Harlan J. and de Wet J. (1971) Towards a rational classification of cultivated plants. Taxon 20, 509–517.
Hedrick, U.P. (1915) The Cherries of New York. J.B. Lyon Company Albany, Albany, USA.
Heywood, V.H. and Dulloo, E. (2005) In Situ Conservation of Wild Plant Species: a Critical Global Review of Good Practices. IPGRI Technical Bulletin No. 11.
Hijmans, R.J., Guarino, L., Cruz, M. and Rojas, E. (2001) Computer tools for spatial analysis of plant genetic resources data: 1. DIVA‐GIS. Plant Genetic Resources Newsletter 127, 15–19.
Hijmans, R., Guarino, L., Mathur, P. and Jarvis, A. (2009) DIVA‐GIS: Geographic Information System for Biodiversity Research [web document]. http://www.diva‐gis.org/ (accessed 24 June 2011).
Holtken, A.M. and Gregorius, H.R. (2006) Detecting local establishment strategies of wild cherry (Prunus avium). BMC Ecology 6, 13.
IPCC (2007) Fourth Assessment Report Climate Change 2007: Synthesis Report. Intergovernmental Panel on Climate Change, Geneva, Switzerland.
Iriondo, J.M., Maxted, N. and Dulloo, M.E. (eds.) (2008) Conserving Plant Genetic Diversity in Protected Areas. CABI Publishing, Wallingford, UK. Pp. 212.
Iriondo, J.M., Maxted, N., Kell, S.P., Ford‐Lloyd, B., Lara‐Romero, C., Labokas, J. and Magos Brehm, J. (2011) Identifying quality standards for genetic reserve conservation of CWR. In: Maxted, N., Dulloo, M.E., Ford‐Lloyd, B.V., Frese, L., Iriondo, J.M. and Pinheiro de Carvalho, M.A.A. (eds.) Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces. CAB International, Wallingford, UK.
Isermeyer, F., Gocht, A., Kleinhanß, W., Küpker, B., Offermann, F., Osterburg, B., Riedel J. and Sommer, U. (2005) Vergleichende Analyse verschiedener Vorschläge zur Reform der

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
39
Zuckermarktordnung. Eine Studie im Auftrag des Bundesministeriums für Verbraucherschutz, Ernährung und Landwirtschaft. Braunschweig, im April 2005.
Jain, S.K. (1975) Genetic reserves. In: Frankel, O.H. and Hawkes, J.G. (eds.), Crop Genetic Resources for Today and Tomorrow. International Biological Programme 2, Cambridge University Press. Cambridge. Pp. 379–396.
Jarvis, A., Lane, A. and Hijmans, R.J. (2008) The effect of climate change on crop wild relatives. Agriculture, Ecosystems and Environment 126(1–2), 13–23.
Jones, P.D., Lister, D.H., Jaggard, K.W. and Pidgeon, J.D. (2003) Future climate change impact on the productivity of sugar beet (Beta vulgaris L.) in Europe. Climatic Change 58, 93–108.
Kell, S.P., Knüpffer, H., Jury, S.L., Maxted, N., Ford‐Lloyd, B.V. (2005) Catalogue of Crop Wild Relatives for Europe and the Mediterranean. Available online via the Crop Wild Relative Information System (CWRIS – http://www.pgrforum.org/cwris.htm) and on CD‐ROM. University of Birmingham, UK.
Kell, S.P., Knüpffer, H., Jury, S.L., Ford‐Lloyd, B.V. and Maxted, N. (2008) Crops and wild relatives of the Euro‐Mediterranean region: making and using a conservation catalogue. In: Maxted, N., Ford‐Lloyd, B.V., Kell, S.P., Iriondo, J., Dulloo, E. and Turok, J. (eds.) Crop Wild Relative Conservation and Use. CABI Publishing, Wallingford. Pp. 69–109.
Kell, S.P., Maxted, N. and Bilz, M. (2011) European crop wild relative threat assessment: knowledge gained and lessons learnt. In: Maxted, N., Dulloo, M.E., Ford‐Lloyd, B.V., Frese, L., Iriondo, J.M. and Pinheiro de Carvalho, M.A.A. (eds.) Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces. CAB International, Wallingford, UK.
Kleinschmit, J., Stephan, R. and Wagner, I. (2006) Wild fruit trees (Prunus avium, Malus sylvestris and Pyrus pyraster) genetic resources conservation strategy [web document]. http://www.bioversityinternational.org/Networks/Euforgen/networks/Scattered_Broadleaves/NHStrategies/WildFruitConsStrategy.htm (accessed 14 March 2008).
Kobliha, J. (2002) Wild cherry (Prunus avium L.) breeding program aimed at the use of this tree in the Czech forestry. Journal of Forest Science 48(5), 202–218.
Ladizinsky, G. (1988) Biological species and wild genetic resources in Avena. In: Mattsson, B. and Lyhagen, R. (eds.), Proceeding of the 3rd International Oat Conference, Lund, Sweden. Pp 76–86.
Ladizinsky, G. (1998) A new species of Avena from Sicily, possibly the tetraploid progenitor of hexaploid oats. Genetic Resources and Crop Evolution 45, 263–269.
Leggett, J.M. and Thomas, H. (1995) Oat evolution and cytogenetics. In: Welch, R.W. (ed.), The Oat Crop: Production and Utilization. Chapman and Hall, London. Pp.121–149.
Letschert, J.P.W. (1993) Beta section Beta: biogeographical patterns of variation and taxonomy. 1–155. Wageningen Agricultural University Papers, 93–1.
Li, X., Takahashi, T., Suzuki, N. and Kaiser, H.M. (2011) The impact of climate change on maize yields in the United States and China. Agricultural Systems 104, 348–353.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
40
Luck, J., Spackmand, M., Freemand, A. ,Trębicki, P., Griffiths, W., Finlay, K. and Chakraborty, S. (2011) Climate change and diseases of food crops. Plant Pathology 60, 113–121.
Magos Brehm, J., Maxted, N., Ford‐Lloyd, B.V. and Martins‐Loução, M.A. (2008) National inventories of crop wild relatives and wild harvested plants: case‐study for Portugal. Genetic Resources and Crop Evolution 55, 779–796.
Mariette, S., Lefranc, M., Legrand, P., Taneyhill, D., Frascaria‐Lacoste, N. and Machon, N. (1997) Genetic variability in wild cherry populations in France. Effects of colonizing processes. Theoretical and Applied Genetics 94, 904–908.
Mateo, R.G., Croat, T.B., Felicisimo, A.M. and Munoz, J. (2010) Profile or group discrimintaive techniques? Generating reliable species distribution models using pseudo‐absences and target‐group absences from natural history collections. Diversity and Distributions 16, 94–94.
Maxted, N. and Guarino, L. (2003) Planning plant genetic conservation. In: Smith, R.D., Dickie, J.B., Linington, S.H., Pritchard, H.W. and Probert, R.J. (eds.), Seed Conservation: turning science into practice. Royal Botanic Gardens, Kew. Pp. 37–78.
Maxted, N. and Kell, S. (2009) Establishment of a Network for the In Situ Conservation of Crop Wild Relatives: Status and Needs. Background Study Paper No. 39. Commission on Genetic Resources for Food and Agriculture. Food and Agriculture Organization of the United Nations, Rome, Italy. 211 pp.
Maxted, N., Hawkes, J.G., Ford‐Lloyd, B.V. and Williams, J.T. (1997a) A practical model for in situ genetic conservation. In: Maxted, N., Ford‐Lloyd, B.V. and Hawkes, J.G. (eds.), Plant Genetic Conservation: the In Situ Approach. Chapman and Hall, London. Pp. 339–367.
Maxted, N., Mabuza‐Dlamini, P., Moss, H., Padulosi, S., Jarvis, A. and Guarino, L. (2004) An Ecogeographic Survey: African Vigna. Systematic and Ecogeographic Studies of Crop Genepools 10. IPGRI, Rome. Pp. 1–468.
Maxted, N., Scholten, M.A., Codd, R. and Ford‐Lloyd, B.V. (2007) Creation and use of a national inventory of crop wild relatives. Biological Conservation 140, 142–159.
Maxted, N., Ford‐Lloyd, B.V., Jury, S.L., Kell, S.P. and Scholten, M.A. (2006) Towards a definition of a crop wild relative. Biodiversity and Conservation 15(8), 2673–2685.
Maxted, N., Ford‐Lloyd, B.V. and Kell, S.P. (2008a) Crop wild relatives: establishing the context. In: Maxted, N., Ford‐Lloyd, B.V., Kell, S.P., Iriondo, J., Dulloo, E. and Turok, J. (eds.), Crop Wild Relative Conservation and Use. CABI Publishing, Wallingford. Pp. 3–30.
Maxted, N., Iriondo, J., Dulloo, E., and Lane, A. (2008b) Introduction: the integration of PGR conservation with protected area management. In: Iriondo, J.M., Maxted, N. and Dulloo, E. (eds.), Plant Genetic Population Management. CABI Publishing, Wallingford, UK. Pp. 1–22.
Maxted, N., Iriondo, J.M., De Hond, L., Dulloo, E., Lefèvre, F., Asdal, Å., Kell, S.P. and Guarino, L. (2008c) Genetic reserve management. In: Iriondo, J.M., Maxted, N. and Dulloo, E. (eds.), Conserving Plant Genetic Diversity in Protected Areas. CABI Publishing, Wallingford, UK. Pp. 65–87.

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
41
Maxted, N., Dulloo, E., Ford‐Lloyd, B.V., Iriondo, J. and Jarvis, A. (2008d) Genetic gap analysis: a tool for more effective genetic conservation assessment. Diversity and Distributions 14, 1018–1030.
Maxted, N., Kell, S. and Magos Brehm, J. (2011) Options to promote food security: on‐farm management and in situ conservation of plant genetic resources for food and agriculture. Food and Agriculture Organization of the United Nations, Rome, Italy.
Maxted, N., Kell. S.P., Magos Brehm, J. and Ford‐Lloyd, B.V. (in prep.) Systematic strategies for crop wild relative conservation.
Namkoong, G., Boyle, T., Gregorius, H.‐R., Joly, H., Savolainen, O., Ratnam, W. and Young, A. (1996) Testing Criteria and Indicators for Assessing the Sustainability of Forest Management: Genetic Criteria and Indicators. CIFOR Working Paper No 10.
Newbold, T. (2010) Applications and limitations of museum data for conservation and ecology, with particular attention to species distribution models. Progress in Physical Geography 34(1), 3–22.
NGA (2009) OGC Viewer GNS Search [web document]. National Geospatial‐Intelligence Agency. http://geonames.nga.mil/ggmaviewer/MainFrameSet.asp (accessed 21 August 2009).
NGDC (2011) Data and Information [web document]. National Geophysical Data Centre. http://www.ngdc.noaa.gov/ (accessed 10 March 2009).
NHM (2007) Seeds of Trade [web document]. Natural History Museum. http://www.nhm.ac.uk/jdsml/nature‐online/seeds‐of‐trade/ (accessed 10 August 2009).
Nicoll, F.J. (1993) Genetic improvement of cherry for farm woodlands. Quarterly Journal of Forestry 87, 187–194.
Nikoloudakis, N. and Katsiotis, A. (2007) The origin of the C‐genome and cytoplasm of Avena polyploids. Theoretical and Applied Genetics 117, 273–281.
Nikoloudakis, N., Skaracis, G. and Katsiotis, A. (2008) Evolutionary insights inferred by molecular analysis of the ITS1‐5.8S‐ITS2 and IGS Avena sp. sequences. Molecular Phylogenetics and Evolution 46, 102–115.
Noël, F., Maurice, S., Mignot, A., Glemin, S., Carbonell, D., Justy, F., Guyot, I., Olivieri, I. and Petit, C. (2010) Interaction of climate, demography and genetics: a ten‐year study of Brassica insularis, a narrow endemic Mediterranean species. Conservation Genetics 11, 509–526.
OECD (2006) Safety Assessment of Transgenic Organisms. OECD Consensus Documents Vol. 1. Organization for Economic Co‐operation and Development, OECD Publishing, Paris, France.
Panella, L., Fenwick, A.L., Hellier, B., Frese, L. and Richards, C.M. (2010) Genetic diversity within and among occurrences of Beta nana. Proceedings of the 72nd IIRB Congress, 22–23/06/2010, Copenhagen, Denmark.
Parra‐Quijano, M., Iriondo, J.M., Frese, L. and Torres, E. (2011) Spatial and ecogeographic approaches for selecting genetic reserves. In: Maxted, N., Dulloo, M.E., Ford‐Lloyd, B.V., Frese, L., Iriondo, J.M. and Pinheiro de Carvalho, M.A.A. (eds.), Agrobiodiversity

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
42
Conservation: Securing the Diversity of Crop Wild Relatives and Landraces. CAB International, Wallingford, UK.
PFAF (2008) Prunus avium L. [web document]. Plants For A Future http://www.pfaf.org/database/plants.php?Prunus+avium (accessed 20 October 2008).
Phillips, S.J., Anderson, R.P. and Schapire, R.E. (2006) Maximum entropy modeling of species geographic distributions. Ecological Modelling 190, 231–259.
R Development Core Team (2010) The R Project for Statistical Computing [web document]. http://www.r‐project.org/ (accessed 10 May 2010).
Rackham, O. (1980) Ancient Woodland: its History, Vegetation and Uses in England. Edward Arnold, London.
Russell, K. (2003) EUFORGEN Technical Guidelines for Genetic Conservation and Use for Wild Cherry (Prunus avium). International Plant Genetics Resources Institute, Rome, Italy. 6 pp.
Santi, F., Muranty, H., Dufour, J. and Paques, L.E. (1998) Genetic parameters and selection in a multisite wild cherry clonal test. Silvae Genetica 47, 2–3.
Schneider, A. (2009) GPS Visualizer: Do‐It‐Yourself Mapping [web document]. http://www.gpsvisualizer.com/ (accessed 21 October 2009).
Snogerup, S., Gustafsson, M. and von Bothmer, R. (1990) Brassica sect. Brassica (Brassicaceae) I. Taxonomy and variation. Willdenowia 19, 271–365.
U, N. (1935) Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Japanese Journal of Botany 7, 389–452.
Vaughan, S.P., Cottrell, J.E., Moodley, D.J., Connolly, T. and Russell, K. (2007) Distribution and fine‐scale spatial‐genetic structure in British wild cherry (Prunus avium). Heredity 98, 274–283.
Veteläinen, M., Helgadóttir, Á. and Weibull, J. (compilers) (2007) Climatic change and genetic resources in northern Europe. Report of a Workshop, 18–19 September 2006, Rovaniemi, Finland. Bioversity International, Rome, Italy. http://www.ecpgr.cgiar.org/publications/publication/issue/climatic_change_and_genetic_resources_in_northern_europe.html (accessed 09 May 2011).
Villain, S. (2007) Histoire evolutive de la section Beta. Mise en évidence des phénomènes d’hybridation et de spéciation au sein de la section dans le bassin méditerranéen. These pour obtenir le grade de docteur en sciences de l’Université de Lille 1. Spécialité: sciences de la vie. Présentée par Sarah Villain le 13 décembre 2007.
Vivero, J.L., Hernandez‐Bermejo, J.E. and Prados Ligero, J. (2001) Conservation strategies and management guidelines for wild Prunus genetic resources in Andalusia, Spain. Genetic Resources and Crop Evolution 48(5), 533–546.
Warwick, S.U. and Sauder, C.A. (2005) Phylogeny of tribe Brassiceae (Brassicaceae) based on chloroplast restriction site polymorphism and nuclear ribosomal internal transcribed spaver and chloroplast trnL intron sequences. Canadian Journal of Botany 83, 467–483.
Warwick, S.I., Francis, A. and Gugel, R.K. (2009) Guide to wild germplasm of Brassica and allied crops (tribe Brassicaceae, Brassicaceae) (3rd edition).

In situ conservation of crop wild relatives: a methodology for identifying priority genetic reserve sites
43
Http://www.brassica.info/info/publications/guide‐wild‐germplasm.php (accessed 02 June 2011)
Watkins, R. (1995) Cherry, plum, peach, apricot and almond. In: Smartt, J. and Simmonds, N.W. (eds.), Evolution of Crop Plants (2nd ed.). Longman, London. Pp. 423–429.
Webster, A.D. (1996) The taxonomic classification of sweet and sour cherries and a brief history of their cultivation. In: Webster, A.D. and Looney, N.E. (eds.), Cherries: Crop Physiology, Production and Uses. CAB International, Wallingford. Pp. 3–24.
White, J. (1995) Forest and Woodland Trees in Britain. Oxford University Press, Oxford.
Zanetto, A., Maggioni, L., Tobutt, K.R. and Dosba, F. (2002) Prunus genetic resources in Europe: achievement and perspectives of a networking activity. Genetic Resources and Crop Evolution 49, 331–337.