induction and modulation of apoptosis in neonatal monocytes by polyunsaturated fatty acids
TRANSCRIPT

www.elsevier.com/locate/jpedsurg
Induction and modulation of apoptosis in neonatalmonocytes by polyunsaturated fatty acidsB
Brian Sweeneya,*, Prem Puria, Denis J. Reena,b
aChildren’s Research Centre, University College Dublin, Our Lady’s Hospital for Sick Children, Dublin K1H 8L1, IrelandbConway Institute of Biomolecular and Biomedical Research, University College Dublin,
Our Lady’s Hospital for Sick Children, Dublin K1H 8L1, Ireland
0022-3468/$ – see front matter D 2007
doi:10.1016/j.jpedsurg.2006.12.024
B Supported by a Health Research
ship Grant.
* Corresponding author. Children’
Ottawa, Ontario, Canada K1H 8L1. Tel
613 7384297.
E-mail address: [email protected]
Index words:Monocytes;
Apoptosis;
Human;
Lipid mediators;
Inflammation;
Fatty acids
Abstract Polyunsaturated fatty acids (PUFAs), known modulators of the immune response, are the
source of essential fatty acids in total parenteral nutrition–dependent patients. Critically ill infants on
TPN have an increased incidence of sepsis, and lipid emulsions depress various immune functions.
Recent studies have demonstrated that PUFAs induce apoptosis in various tissue cells in vitro and ex
vivo. The susceptibility of neonatal monocytes, as major early effector cells in the host response to
sepsis, to PUFA-mediated apoptosis and the mechanisms associated with PUFA-induced apoptosis were
investigated. Both n-3 and n-6 PUFAs induced rapid, dose-dependent cell death in purified monocytes.
Polyunsaturated fatty acids induced significant activation of upstream caspases 8 and 9 as well as
caspase 3. The PUFA treatment resulted in a 4-fold increase in oxidative stress and a loss of monocyte
mitochondrial potential compared with carrier controls (P b .05). The addition of cyclosporin, which
blocks the development of mitochondrial transition pores, completely abolished the proapoptotic effects
of PUFAs. Although Trolox (Sigma Aldrich) reduced PUFA-induced intracellular oxidative stress in
neonatal monocytes, apoptosis was not blocked by this potent antioxidant. The data identify PUFAs as
potent inducers of monocyte apoptosis, which can occur independently of the induction of oxidative
stress, by using a mitochondrial dependent pathway. The TPN-dependent infant may be particularly
sensitive to such PUFA effects, having a relatively poor capacity to both use and clear PUFAs.
D 2007 Elsevier Inc. All rights reserved.
The introduction of lipid emulsions for total parenteral
nutrition (TPN) has been a major breakthrough in medicine.
The most frequently used lipid formula in TPN is based on
Elsevier Inc. All rights reserved.
Board Research Training Fellow-
s Hospital of Eastern Ontario,
.: +1 613 7377600x2196; fax: +1
.ca (B. Sweeney).
soybean oil and contains predominantly long-chain fatty
acids of the n-6 series (linoleic acid) [1]. More recently, fish
oil–based lipid emulsions, which are rich in fatty acids of the
n-3 polyunsaturated fatty acids (PUFAs) (a-linolenic acid),
have been used [2]. Furthermore, it has been demonstrated
that the use of soybean oil–derived lipid formula results in
significant elevation of n-6 PUFAs such as arachidonic acid
(AA), whereas the use of fish oil–based lipid formula results
in an elevation of n-3 PUFAs such as docosahexaenoic acid
(DHA) and eicosapentaenoic acid (EPA) [3]. The n-3 and n-6
fatty acids have long been recognized to have potent effects
Journal of Pediatric Surgery (2007) 42, 620–628

1 Dr. Jaydeep Sarma, Welcome Trust Research Fellow, University
of Edinburgh Medical School, Edinburgh, Scotland.
Induction and modulation of apoptosis in neonatal monocytes 621
as modulators of the immune response and have been shown
to have differential effects on monocyte, lymphocyte, and
neutrophil function [4,5]. In addition to extensive in vitro
and ex vivo laboratory animal work, some human studies
have been conducted assessing various immune parameters
after dietary supplementation with n-3 PUFAs. This work
has been extensively reviewed [4], and some benefits of fish
oils have been shown in rheumatoid arthritis, psoriasis,
inflammatory bowel disease, systemic lupus erythematosus,
and post–renal transplantation patients [6]. Strikingly,
critically ill neonates on TPN have an increased incidence
of sepsis [7,8]; and additional studies have shown that lipid
emulsions depress monocyte chemotaxis [9], phagocytosis
[10], as well as complement [11] and cytokine production
[12]. Monocytes are recognized as major effector cells of the
immune system, playing a central role in the initiation,
development, and outcome of the innate immune response.
Monocyte survival is exquisitely regulated in vivo by various
pro- and antiapoptotic signals, survival being promoted
during the inflammatory response and diminished when
surplus to requirements or in the presence of recognized
immunosuppressive agents such as glucocorticoids [13].
Monocyte apoptosis has emerged as a central regulatory
event in hematopoiesis and inflammation [14], and some of
the elements of the complex system regulating the tight
control of monocyte survival are gradually emerging.
Regulation of monocyte apoptosis may be an important
homeostatic mechanism for controlling the number of
monocytes available to respond to infection, wound healing,
and tumor growth.
Recent studies have demonstrated that PUFAs are
capable of inducing apoptosis in vitro and ex vivo [15]. In
a rodent feeding model, Garrido et al have shown that
dietary supplementation with PUFAs results in depletion of
cellular antioxidants such as glutathione, thus rendering
them more susceptible to oxidative stress [16].
How PUFAs mediate cell death has yet to be fully
elucidated. Several potential mechanisms have been pro-
posed to explain their immunomodulatory effects, including
their effects on eicosanoid formation, signal transduction,
gene expression, and lipid peroxidation [17]. It is known
that lipid peroxidation and the metabolites of this process
are associated with increased oxidative stress [18]. Oxida-
tive stress, in turn, has been implicated as a significant
mediator of apoptosis [19]. Reactive oxygen species (ROS)
resulting from oxidative stress have been implicated in the
regulation of Fas-mediated monocyte apoptosis. This form
of monocyte apoptosis is associated with increased intra-
cellular levels of ROS and can be blocked with the
antioxidant n-acetylcysteine [14]. Synthesizing these data,
there is considerable evidence to support the concept that
agents that have a proapoptotic effect on monocytes may
have a potentially anti-inflammatory effect.
The aim of this study was to gain mechanistic insights
into PUFA-regulated apoptosis in monocytes in a human
neonatal model by determining upstream intracytoplasmic
events in apoptosis, specifically, the development of
oxidative stress, the loss of mitochondrial inner membrane
potential, and the activation of initiator and effector
caspases. If PUFAs have apoptogenic effects, identifying
the mechanisms whereby they may modulate the immune
response could have important implications for management
of infants on TPN. The TPN-dependent infant may be
particularly sensitive to such PUFA effects, having a
relatively poor capacity to both use and clear PUFAs [8].
1. Materials and methods
1.1. Monocyte isolation by negative selection
Umbilical cord blood monocytes were isolated by an
immunomagnetic bead negative selection method (Miltenyi
Biotech, Germany), using a modification of a protocol
provided by Dr Jaydeep Sarma1. Density gradient isolated
cord blood mononuclear cells (peripheral blood mononu-
clear cells; PBMCs), derived from acid citrate dextrose
anticoagulated whole blood, were washed twice in cation-
free Hanks balanced salt solution and counted. The PBMCs
were resuspended in an isolation buffer composed of cation-
free Hanks balanced salt solution, 4% autologous platelet
free plasma (APFP), and EDTA (250 lmol/L) at a volume
of 40 lL of isolation buffer per 107 PBMCs. Added to this
were 40 lL of APFP and 20 lL of hapten antibody cocktail
per 107 PBMCs. This mixture was incubated for 5 minutes
at 48C. The cells were then washed twice in the isolation
buffer before resuspension in 10 lL of isolation buffer,
40 lL of APFP, and 20 lL of antihapten microbeads per 107
cells. In addition, 10 lL per 107 cells of anti–glycophorin A,
anti-CD15, and anti-CD61 microbeads was added to remove
contaminating erythroblasts, neutrophils, and platelets,
respectively. The mixture was then incubated at 48C for
15 minutes. The cells were washed before resuspension in
500 lL of isolation buffer. The mixture was then passed
through a primed LS+/VS+ isolation column in a magnetic
field. The resulting negatively selected monocytes were
washed once in RPMI and 10% APFP. Their purity was
routinely greater than 85% as determined flow cytometri-
cally using CD45, CD14, and CD41 monoclonal antibodies.
The viability of freshly isolated negatively selected mono-
cytes was greater than 90% on ethidium bromide/acridine
orange staining. Purified monocytes were then incubated at
a concentration of 106/mL in RPMI 1640 and either 1% or
10% APFP in a 5% CO2 incubator at 378C in 24- or 96-well
ultralow attachment polystyrene flat-bottomed plates.
1.2. Preparation of PUFAs
Stock solutions of the n-3 PUFAs DHA and EPA and the
n-6 PUFA AAwere made up in a nitrogen atmosphere using
an Atmosbag (Sigma Aldrich) in 100% ethanol and were

Fig. 1 Cell death in monocytes treated with PUFAs. Negatively
selected blood monocytes were cultured for 2 hours in RPMI
and 1% APFP with carrier control or various concentrations of
the PUFAs AA, DHA, and EPA. The treated cells were then
harvested and triple stained with annexin V, CD14, and PI to
quantify the degree of cell death by flow cytometry as determined
by measuring annexin V positivity. Data are mean F 1 SD for
4 separate experiments (*P b .05 indicates a significant difference
from control).
B. Sweeney et al.622
stored in the dark at �208C to prevent peroxidation The
stocks were diluted at least 1000-fold in complete medium
to the test culture concentrations. Carrier controls consisted
of the addition of 0.1% ethanol in the absence of fatty acids.
They had no influence on cell viability at the concentration
used for all experiments. The doses of PUFAs used ranged
from 0 to 100 lmol/L and were based on previously
published cell line data [15,20].
1.3. Detection and quantification of monocytecell death
Cell death of monocytes was measured essentially
according to the protocol of Schmidt et al [13]. After the
designated culture period, carrier control– or fatty acid–
treated monocytes were harvested with ice-cold phosphate-
buffered saline (PBS), washed and resuspended in annexin
V binding buffer, and triple labeled with annexin V
fluorescein isothiocyanate (FITC), CD14 phycoerythrin
(PE), and propidium iodide (PI). Ten thousand events were
acquired on 3 channels of a FACscan flow cytometer
(Becton Dickinson, Oxford, UK) and data were analyzed
with CellQUEST software (Becton Dickinson, Oxford, UK).
Cell death was quantified by measuring the number of
apoptotic annexin V–positive and primarily or secondarily
necrotic PI-positive events.
1.4. Detection of active caspases 8 and 9in monocytes
CaspaTag caspase activity kits (Intergen, UK) were used
to detect active caspases 8 and 9 through the use of specific
carboxyfluorescein-labeled caspase substrates as per the
manufacturer’s protocol. Briefly, 300-lL aliquots of carrier
control– and PUFA-treated cells were incubated for 1 hour
in the dark at 378C with 10 lL of 30� working dilution of
the peptide substrates FAM-LETD-FMK or FAM-LEHD-
FMK to measure active caspase 8 or 9, respectively. The
cells were then washed twice in the wash buffer provided by
the manufacturer before flow cytometric analysis. The
resulting frequency histogram of number of events (y-axis)
versus FL1 fluorescein intensity (x-axis) was used to
determine the level of caspase activation.
1.5. Detection of caspase 3
An FITC-conjugated rabbit antiactive caspase 3 antibody
was used to detect active caspase 3 in monocytes after 2 hours
of culture in RPMI 1640 and 1% APFP as per the
manufacturer’s protocol (Pharmingen, CA). Briefly, carrier
control– or PUFA-treated cord monocytes were washed in
PBS and then fixed and permeabilized for 20 minutes at room
temperature, washed, and resuspended in Perm/Wash Buffer
(Pharmingen). Cells were then stainedwith antiactive caspase
3 antibodies for 1 hour at room temperature in the dark. The
permeabilized cells were then washed and resuspended in
Perm/Wash Buffer and analyzed by flow cytometry.
1.6. Determination of oxidative stress inmonocytes
Umbilical cord blood PBMCs were labeled with dichlor-
ofluorescin diacetate (DCFH-DA) (5 lmol/L) for 30 minutes
in an incubator, and intracellular oxidative stress of
monocytes was measured as described by Bass et al [21].
Briefly, DCFH-DA diffuses into the cell and is deacetylated
by intracellular esterases to form 2V, 7V-DCFH, which is
trapped within the cell. In the presence of hydrogen
peroxide, it is oxidized to become highly fluorescent.
Cumene hydroperoxide (CUHP) (50 lmol/L) was used as
a positive control of oxidative stress as described by others
[22]. Cells were also labeled with aCD14 PE antibody for
15 minutes before acquisition of 50,000 events on the flow
cytometer. Live monocytes were gated based on their CD14
and forward/side scatter characteristics.
1.7. Measurement of mitochondrial membranepotential by flow cytometry
To evaluate mitochondrial membrane potential (DCm),
the cationic lipophilic fluorochrome 3,3V-dihexyloxacarbo-cyanine iodide (DiOC6) was used as previously described
[23]. Briefly, umbilical cord blood PBMCs, at a concentra-
tion of 106/mL, were incubated for 20 minutes at 378C with

Induction and modulation of apoptosis in neonatal monocytes 623
DiOC6 (50 nmol/L) before 60 minutes with carrier control
or PUFA treatment. Cells were then washed in PBS before
flow cytometric analysis of gated live monocytes.
1.8. Statistics
The Mann-Whitney nonparametric U test was used to
compare the results of carrier control treatment with the
individual PUFA treatments using statistical analysis soft-
ware package (Instat, GraphPad Software, CA). P values
less than .05 were considered significant.
2. Results
2.1. PUFAs induce rapid cell death in monocytes
Purified umbilical cord blood monocytes were incubated
for 2 hours with carrier control or individual fatty acids. A
concentration-dependent significant increase (P b .05) in
PUFA-induced cell death was observed for each of the
Fig. 2 Polyunsaturated fatty acids induce caspase 8, 9, and 3 activation
C, D), caspase 9 (E, F, G, H), and caspase 3 (I, J, K, L) activation in
(100 lmol/L). These results are representative of 3 separate experiment
respective treatment is shown in the top right of each panel.
3 PUFAs under study over the concentration range 25 to
100 lmol/L (Fig. 1). At the highest PUFA concentration
(100 lmol/L), a significant proportion of monocytes were
PI positive (data not shown), suggesting that either primary
or secondary necrosis had taken place. Apoptosis and
necrosis have been reported to occur side by side in
response to an identical stimulus [24]. However, PI uptake
in the nucleus, traditionally considered an archetypal marker
of necrosis, has also been reported to be taken up at an early
stage in the apoptotic process, thus limiting its ability to
distinguish between apoptosis and necrosis [25]. Therefore,
to confirm that monocyte apoptosis was associated with
exposure to PUFAs, alternative markers of apoptotic cell
death were investigated. Upstream caspases 8 and 9 and the
effector caspase (caspase 3) activities were measured by
flow cytometry. Incubation of monocytes with PUFAs for
90 minutes resulted in activation of both caspase 8 and
caspase 9 (Fig. 2). These cells were also significantly PI
positive, suggesting that they were secondarily necrotic
(data not shown). This conclusion is supported by the recent
in monocytes. Flow cytometric histogram data of caspase 8 (A, B,
cord blood monocytes treated with control, AA, DHA, and EPA
s. The percentage of monocytes positive for each caspase after the
B. Sweeney et al.624
work by Denecker et al [26] who have demonstrated that
apoptotic cells in culture release activated caspases into
culture supernatants, whereas necrotic cells release unpro-
cessed caspases. Our data clearly showed massive caspase
activation, indicating that apoptosis was the dominant mode
of cell death in this model.
2.2. PUFAs activate caspase 3 in monocytes
Having determined that PUFA treatment resulted in
upstream caspase activation, it was decided to determine if
the downstream effector caspase (caspase 3) was also
activated. The level of active caspase 3 in cord monocytes
was 76 F 15%, 85 F 15%, 73 F 7% and 8 F 3% with AA,
DHA, EPA, and carrier control treatments, respectively
(Fig. 2), indicating that the total caspase cascade system
was activated.
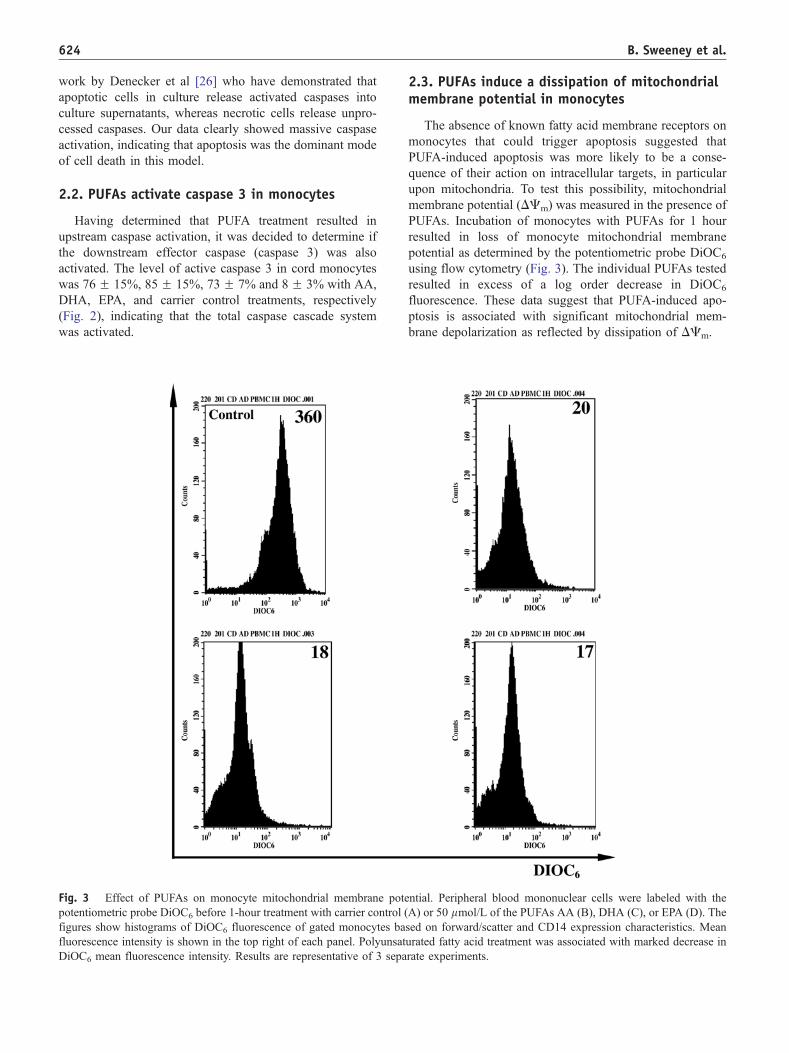
Fig. 3 Effect of PUFAs on monocyte mitochondrial membrane pote
potentiometric probe DiOC6 before 1-hour treatment with carrier control (
figures show histograms of DiOC6 fluorescence of gated monocytes ba
fluorescence intensity is shown in the top right of each panel. Polyunsat
DiOC6 mean fluorescence intensity. Results are representative of 3 sepa
2.3. PUFAs induce a dissipation of mitochondrialmembrane potential in monocytes
The absence of known fatty acid membrane receptors on
monocytes that could trigger apoptosis suggested that
PUFA-induced apoptosis was more likely to be a conse-
quence of their action on intracellular targets, in particular
upon mitochondria. To test this possibility, mitochondrial
membrane potential (DCm) was measured in the presence of
PUFAs. Incubation of monocytes with PUFAs for 1 hour
resulted in loss of monocyte mitochondrial membrane
potential as determined by the potentiometric probe DiOC6
using flow cytometry (Fig. 3). The individual PUFAs tested
resulted in excess of a log order decrease in DiOC6
fluorescence. These data suggest that PUFA-induced apo-
ptosis is associated with significant mitochondrial mem-
brane depolarization as reflected by dissipation of DCm.
ntial. Peripheral blood mononuclear cells were labeled with the
A) or 50 lmol/L of the PUFAs AA (B), DHA (C), or EPA (D). The
sed on forward/scatter and CD14 expression characteristics. Mean
urated fatty acid treatment was associated with marked decrease in
rate experiments.

Fig. 4 Cyclosporin A inhibition of PUFA-induced apoptosis in
monocytes. Negatively selected cord blood monocytes were
incubated in 1% APFP in the presence or absence of CsA
(50lmol/L) for 30 minutes before the addition of carrier control
or one of the PUFAs AA, DHA, or EPA (100 lmol/L). After
2 hours of further incubation, the cells were harvested and triple
stained with annexin V, CD14, and PI to quantify the degree of cell
death by flow cytometry. Cyclosporin A significantly inhibited
PUFA-mediated apoptosis in all cases (*P b .05). Data are mean F1 SD for 3 separate experiments.
Fig. 5 Fatty acids induce oxidative stress in monocytes.
Peripheral blood mononuclear cells were labeled with DCFH-DA
(5 lmol/L) before the addition of carrier control or the fatty acids
AA, DHA, or EPA (50 lmol/L) for 30 minutes. Cumene
hydroperoxide (50 lmol/L) was used as a positive control for
induction of oxidative stress [19]. Cells were also labeled with CD14
to enable flow cytometric identification of monocytes by flow
cytometry. The fluorescence of the deacetylated probe is expressed
relative to carrier control fluorescence for each of the test fatty acids
and cumene hydroperoxide (CUHP). Data are mean F 1 SD for
4 separate experiments. All the fatty acids as well as CUHP induced
significant oxidative stress relative to carrier control (*P b .05).
Induction and modulation of apoptosis in neonatal monocytes 625
2.4. Cyclosporin A protects against PUFA-mediatedapoptosis in monocytes
One of the many biological activities of cyclosporin A
(CsA) is its function as a potent inhibitor of loss of DCm
[27]. The effect of CsA on PUFA-induced monocyte
apoptosis was therefore measured. Negatively selected
monocytes, in RPMI 1640 and 1% APFP, were pretreated
with CsA (50 lmol/L) for 15 minutes before the addition of
carrier control or PUFA (100 lmol/L) treatments. This dose
of CsA has been reported to prevent apoptosis in a model of
ceramide-mediated apoptosis in the human monocytic U937
cell line [28]. After a further 2 hours of incubation, the cells
were harvested; and cell death was measured as already
described (Fig. 4). Carrier control treatment was associated
with 16 F 1.4% cell death compared with 66 F 5.7%, 76 F8.5%, and 47 F 4% cell death with AA, DHA, and EPA
treatment, respectively. However, CsA pretreatment com-
pletely blocked PUFA-mediated cell death and reduced the
level of death down to that of carrier control treatment.
These results provided further evidence for an effect of
PUFAs on mitochondrial integrity.
2.5. PUFAs induce oxidative stress in monocytes
The apoptogenic effects of PUFAs via mitochondrial
membrane perturbation and caspase activation are likely
to be a consequence of their potent oxidizing effects on
cellular substrates. Oxidation of DCFH was used to measure
PUFA-induced intracellular oxidative changes in human
monocytes. Polyunsaturated fatty acids induced significant
substrate oxidation in neonatal monocytes (P b .05)
compared with carrier control–treated cells (Fig. 5). The
fatty acid treatment was associated with approximately
a 4-fold increase in oxidative stress compared with
carrier control.
B. Sweeney et al.626
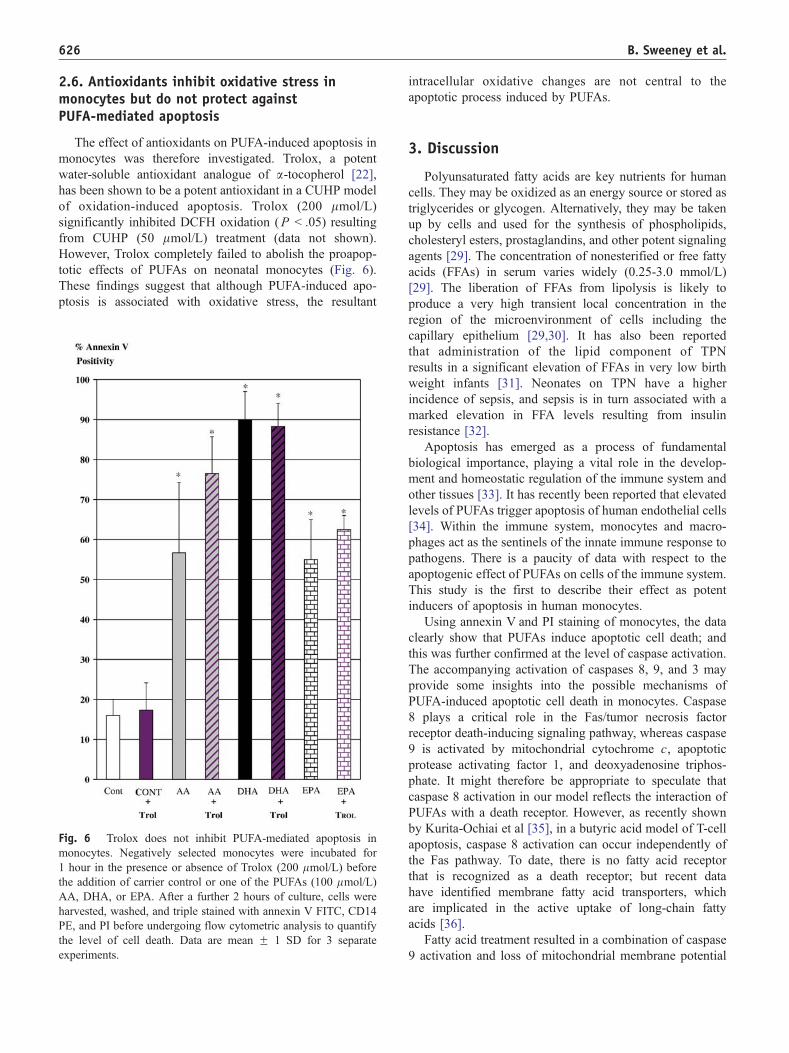
2.6. Antioxidants inhibit oxidative stress inmonocytes but do not protect againstPUFA-mediated apoptosis
The effect of antioxidants on PUFA-induced apoptosis in
monocytes was therefore investigated. Trolox, a potent
water-soluble antioxidant analogue of a-tocopherol [22],
has been shown to be a potent antioxidant in a CUHP model
of oxidation-induced apoptosis. Trolox (200 lmol/L)
significantly inhibited DCFH oxidation (P b .05) resulting
from CUHP (50 lmol/L) treatment (data not shown).
However, Trolox completely failed to abolish the proapop-
totic effects of PUFAs on neonatal monocytes (Fig. 6).
These findings suggest that although PUFA-induced apo-
ptosis is associated with oxidative stress, the resultant
Fig. 6 Trolox does not inhibit PUFA-mediated apoptosis in
monocytes. Negatively selected monocytes were incubated for
1 hour in the presence or absence of Trolox (200 lmol/L) before
the addition of carrier control or one of the PUFAs (100 lmol/L)
AA, DHA, or EPA. After a further 2 hours of culture, cells were
harvested, washed, and triple stained with annexin V FITC, CD14
PE, and PI before undergoing flow cytometric analysis to quantify
the level of cell death. Data are mean F 1 SD for 3 separate
experiments.
intracellular oxidative changes are not central to the
apoptotic process induced by PUFAs.
3. Discussion
Polyunsaturated fatty acids are key nutrients for human
cells. They may be oxidized as an energy source or stored as
triglycerides or glycogen. Alternatively, they may be taken
up by cells and used for the synthesis of phospholipids,
cholesteryl esters, prostaglandins, and other potent signaling
agents [29]. The concentration of nonesterified or free fatty
acids (FFAs) in serum varies widely (0.25-3.0 mmol/L)
[29]. The liberation of FFAs from lipolysis is likely to
produce a very high transient local concentration in the
region of the microenvironment of cells including the
capillary epithelium [29,30]. It has also been reported
that administration of the lipid component of TPN
results in a significant elevation of FFAs in very low birth
weight infants [31]. Neonates on TPN have a higher
incidence of sepsis, and sepsis is in turn associated with a
marked elevation in FFA levels resulting from insulin
resistance [32].
Apoptosis has emerged as a process of fundamental
biological importance, playing a vital role in the develop-
ment and homeostatic regulation of the immune system and
other tissues [33]. It has recently been reported that elevated
levels of PUFAs trigger apoptosis of human endothelial cells
[34]. Within the immune system, monocytes and macro-
phages act as the sentinels of the innate immune response to
pathogens. There is a paucity of data with respect to the
apoptogenic effect of PUFAs on cells of the immune system.
This study is the first to describe their effect as potent
inducers of apoptosis in human monocytes.
Using annexin V and PI staining of monocytes, the data
clearly show that PUFAs induce apoptotic cell death; and
this was further confirmed at the level of caspase activation.
The accompanying activation of caspases 8, 9, and 3 may
provide some insights into the possible mechanisms of
PUFA-induced apoptotic cell death in monocytes. Caspase
8 plays a critical role in the Fas/tumor necrosis factor
receptor death-inducing signaling pathway, whereas caspase
9 is activated by mitochondrial cytochrome c, apoptotic
protease activating factor 1, and deoxyadenosine triphos-
phate. It might therefore be appropriate to speculate that
caspase 8 activation in our model reflects the interaction of
PUFAs with a death receptor. However, as recently shown
by Kurita-Ochiai et al [35], in a butyric acid model of T-cell
apoptosis, caspase 8 activation can occur independently of
the Fas pathway. To date, there is no fatty acid receptor
that is recognized as a death receptor; but recent data
have identified membrane fatty acid transporters, which
are implicated in the active uptake of long-chain fatty
acids [36].
Fatty acid treatment resulted in a combination of caspase
9 activation and loss of mitochondrial membrane potential

Induction and modulation of apoptosis in neonatal monocytes 627
in our model. In addition, the use of CsA completely
blocked fatty acid–induced apoptosis. These data suggest
that the PUFAs mediate apoptosis, at least in part, through
the mitochondrion. This conjecture is consistent with other
work that has recently demonstrated CsA-sensitive apopto-
sis induced by fatty acids in various cells [27,37,38].
Our finding that PUFAs induce significant oxidative
stress in monocytes is not surprising in light of published
data that have long recognized PUFAs as a potential source
of oxidative stress [39]. By definition, they possess
multiple double bonds, which are readily susceptible to
peroxidation; and the lipoperoxides resulting from this
process are potential mediators of apoptosis [18]. Interest-
ingly, defective mitochondria have also been identified as a
major and primary source of free radicals, which can lead
cells into a death cascade [40]. In addition, modulation of
the oxidation state of the cell has been shown to have
dramatic effects on the level of apoptosis in various
models. For example, apoptosis induced by tumor necrosis
factor a results in a rapid rise in intracellular ROS; and
inhibition of this stress using n-acetylcysteine as an
antioxidant has been shown to inhibit apoptosis in this
model [41]. Recently, Arita et al showed that PUFAs
induce oxidative stress that was associated with significant
apoptosis after 6 hours of treatment in HL-60 cells and
found that dimethyl sulphoxide, which has antioxidant
properties, could be protective against the proapoptotic
effects of fatty acid treatment [42]. Therefore, it was
surprising that Trolox, a potent antioxidant, was not
protective in our model. However, Finstad et al were
unable to inhibit EPA-induced apoptosis in the HL-60
monocytic cell line using either a-tocopherol or ascorbic
acid (50-100 lmol/L) [43]. They argue that their findings
support the concept that PUFAs may regulate cellular
processes such as apoptosis independently of lipid perox-
ide/oxidative stress pathways, which is consistent with our
findings in primary human monocytes.
As previously stated, neonates receiving TPN are prone
to developing an elevation of FFA levels [31], especially in
the presence of sepsis [32]. Whether this in turn can result in
monocyte apoptosis, despite the incorporation of antiox-
idants into lipid emulsions, is a critical question that needs
to be addressed. Oxidative stress–independent induction of
apoptosis by PUFAs as demonstrated in this study is
potentially yet another mechanism whereby these nutrients
may modulate the immune response.
Further in vivo studies are needed to explore the
relevance of our findings in the context of TPN-dependent
neonates. In vivo, there is evidence that patients on TPN
have levels of the PUFAs AA, DHA, and EPA in the range
of approximately 100 to 200 lmol/L [3]. However, in
blood, the solubilization and transport of PUFAs are made
possible by their binding to albumin. This results, in the
physiological setting, in FFA levels of 5 to 50 nmol/L [44].
We did not determine the FFA level in our model system;
but it is likely to have been considerably higher than this
because we cultured using 1% plasma, whereas in vivo, the
neonatal plasma concentration is approximately 50%. As
yet, there are no in vivo data that indicate that there is
increased monocyte apoptosis occurring in neonates on
TPN. Our laboratory did consider conducting such experi-
ments, but current monocyte isolation techniques require a
significant volume of blood to isolate sufficient mono-
cytes to conduct these experiments, which we currently
cannot justify either ethically or clinically. We are planning,
however, to perform in vitro coincubation experi-
ments of purified umbilical cord monocytes with both
physiological and pharmacological levels of commercially
available lipid emulsions to determine if they induce
monocyte apoptosis.
References
[1] Miles EA, Calder PC. Modulation of immune function by dietary fatty
acids. Proc Nutr Soc 1998;57:277-92.
[2] Garnacho-Montero J, Ortiz-Leyba C, Garnacho-Montero MC, et al.
Effects of three intravenous lipid emulsions on the survival
and mononuclear phagocyte function of septic rats. Nutrition
2002;18:751-4.
[3] Mayer K, Fegbeutel C, Hattar K, et al. Omega-3 vs omega-6 lipid
emulsions exert differential influence on neutrophils in septic shock
patients: impact on plasma fatty acids and lipid mediator generation.
Intensive Care Med 2003;29:1472-81.
[4] Calder PC. N-3 polyunsaturated fatty acids and immune cell function.
Adv Enzyme Regul 1997;37:197-237.
[5] Calder PC. Effects of fatty acids and dietary lipids on cells of the
immune system. Proc Nutr Soc 1996;55:127-50.
[6] Blok WL, Deslypere JP, Demacker PN, et al. Pro- and anti-
inflammatory cytokines in healthy volunteers fed various doses of
fish oil for 1 year. Eur J Clin Invest 1997;27:1003-8.
[7] Okada Y, Klein NJ, van Saene HK, et al. Bactericidal activity against
coagulase-negative staphylococci is impaired in infants receiving
long-term parenteral nutrition. Ann Surg 2000;231:276-81.
[8] Fischer GW, Hunter KW, Wilson SR, et al. Diminished bacterial
defences with intralipid. Lancet 1980;2:819-20.
[9] Garnacho Montero J, Shou J, Ortiz Leyba C, et al. Lipids and immune
function. Nutr Hosp 1996;11:230-7.
[10] de Pablo MA, Alvarez de Cienfuegos G. Modulatory effects of dietary
lipids on immune system functions. Immunol Cell Biol 2000;78:31-9.
[11] Konig D, Berg A, Weinstock C, et al. Essential fatty acids, immune
function, and exercise. Exerc Immunol Rev 1997;3:1 -31.
[12] Wallace FA, Yaqoob P, Miles EA, et al. Dietary fat influences the
production of Th1- but not Th2-derived cytokines. Lipids
1999;34(Suppl):S141.
[13] Schmidt M, Pauels HG, Lugering N, et al. Glucocorticoids induce
apoptosis in human monocytes: potential role of IL-1 beta. J Immunol
1999;163:3484-90.
[14] Um HD, Orenstein JM, Wahl SM. Fas mediates apoptosis in human
monocytes by a reactive oxygen intermediate dependent pathway.
J Immunol 1996;156:3469 -77.
[15] Diep QN, Intengan HD, Schiffrin EL. Endothelin-1 attenuates omega
3 fatty acid–induced apoptosis by inhibition of caspase 3. Hyperten-
sion 2000;35:287 -91.
[16] Garrido A, Garrido F, Guerra R, et al. Ingestion of high doses of fish
oil increases the susceptibility of cellular membranes to the induction
of oxidative stress. Lipids 1989;24:833-5.
[17] Field CJ, Johnson IR, Schley PD. Nutrients and their role in host
resistance to infection. J Leukoc Biol 2002;71:16 -32.

B. Sweeney et al.628
[18] Avula CP, Fernandes G. Modulation of antioxidant enzymes and
apoptosis in mice by dietary lipids and treadmill exercise. J Clin
Immunol 1999;19:35 -44.
[19] Buttke TM, Sandstrom PA. Oxidative stress as a mediator of
apoptosis. Immunol Today 1994;15:7 -10.
[20] Finstad HS, Myhrstad MC, Heimli H, et al. Multiplication and death
type of leukemia cell lines exposed to very long chain polyunsaturated
fatty acids. Leukemia 1998;12:921-9.
[21] Bass DA, Parce JW, Dechatelet LR, et al. Flow cytometric studies of
oxidative product formation by neutrophils: a graded response to
membrane stimulation. J Immunol 1983;130:1910 -7.
[22] Makrigiorgos GM, Kassis AI, Mahmood A, et al. Novel fluorescein-
based flow cytometric method for detection of lipid peroxidation. Free
Radic Biol Med 1997;22:93-100.
[23] Dinsdale D, Zhuang J, Cohen GM. Redistribution of cytochrome c
precedes the caspase-dependent formation of ultracondensed mito-
chondria, with a reduced inner membrane potential, in apoptotic
monocytes. Am J Pathol 1999;155:607-18.
[24] Leist M, Single B, Castoldi AF, et al. Intracellular adenosine
triphosphate (ATP) concentration: a switch in the decision between
apoptosis and necrosis. J Exp Med 1997;185:1481 -6.
[25] Lizard G, Fournel S, Genestier L, et al. Kinetics of plasma membrane
and mitochondrial alterations in cells undergoing apoptosis. Cytom-
etry 1995;21:275 -83.
[26] Denecker G, Vercammen D, Steemans M, et al. Death receptor–
induced apoptotic and necrotic cell death: differential role of caspases
and mitochondria. Cell Death Differ 2001;8:829-40.
[27] Kong JY, Rabkin SW. Palmitate-induced apoptosis in cardiomyocytes
is mediated through alterations in mitochondria: prevention by
cyclosporin A. Biochim Biophys Acta 2000;1485:45 -55.
[28] Nardini M, Leonardi F, Scaccini C, et al. Modulation of ceramide-
induced NF-kappaB binding activity and apoptotic response by caffeic
acid in U937 cells: comparison with other antioxidants. Free Radic
Biol Med 2001;30:722-33.
[29] Hamilton J, Kamp F. How are free fatty acids transported in
membranes? Is it by proteins or by free diffusion through the lipids?
Diabetes 1999;48:2255-69.
[30] Cistola DP, Small DM. Fatty acid distribution in systems modeling the
normal and diabetic human circulation. A 13C nuclear magnetic
resonance study. J Clin Invest 1991;87:1431-41.
[31] Ruben S, Kleinfeld AM, Richeiri GV, et al. Serum levels of unbound
free fatty acids. II: The effect of intralipid administration in premature
infants. J Am Coll Nutr 1997;16:85-7.
[32] Andersen SK, Gjedsted J, Christiansen C, et al. The roles of insulin
and hyperglycemia in sepsis pathogenesis. J Leukoc Biol 2004;
75:413-21.
[33] Hetts SW. To die or not to die: an overview of apoptosis and its role in
disease. JAMA 1998;279:300-7.
[34] Artwohl M, Roden M, Waldhausl W, et al. Free fatty acids trigger
apoptosis and inhibit cell cycle progression in human vascular
endothelial cells. FASEB J 2004;18:146-8.
[35] Kurita-Ochiai T, Ochiai K, Fukushima K. Butyric acid–
induced T-cell apoptosis is mediated by caspase-8 and -9 activa-
tion in a Fas-independent manner. Clin Diagn Lab Immunol 2001;8:
325 -32.
[36] Kruidering M, Evan GI. Caspase-8 in apoptosis: the beginning of btheendQ? IUBMB Life 2000;50:85 -90.
[37] Kashiwagi A, Kanno T, Arita K, et al. Suppression of T(3)- and fatty
acid–induced membrane permeability transition by L-carnitine. Comp
Biochem Physiol B Biochem Mol Biol 2001;130:411 -8.
[38] Wu D, Cederbaum AI. Cyclosporine A protects against arachidonic
acid toxicity in rat hepatocytes: role of CYP2E1 and mitochondria.
Hepatology 2002;35:1420-30.
[39] Begin ME, Ells G, Horrobin DF. Polyunsaturated fatty acid–induced
cytotoxicity against tumor cells and its relationship to lipid perox-
idation. J Natl Cancer Inst 1988;80:188 -94.
[40] Rustin P, von Kleist-Retzow JC, Vajo Z, et al. For debate: defective
mitochondria, free radicals, cell death, aging-reality or myth-ochon-
dria? Mech Ageing Dev 2000;114:201-6.
[41] Larrick JW, Wright SC. Cytotoxic mechanism of tumor necrosis
factor–alpha. FASEB J 1990;4:3215 -23.
[42] Arita K, Kobuchi H, Utsumi T, et al. Mechanism of apoptosis in
HL-60 cells induced by n-3 and n-6 polyunsaturated fatty acids.
Biochem Pharmacol 2001;62:821-8.
[43] Finstad HS, Drevon CA, Kulseth MA, et al. Cell proliferation,
apoptosis and accumulation of lipid droplets in U937-1 cells incubated
with eicosapentaenoic acid. Biochem J 1998;336:451-9.
[44] Abumrad N, Coburn C, Ibrahimi A. Membrane proteins implicated in
long-chain fatty acid uptake by mammalian cells: CD36, FATP and
FABPm. Biochim Biophys Acta 1999;1441:4-13.