induction of increased collagen and elastin biosynthesis...
TRANSCRIPT

Induction of Increased Collagenand Elastin Biosynthesis inCopper-Deficient Pig Aorta
Kenneth E. Hill and Jeffrey M. Davidson
Young pigs raised on a copper-deficient diet develop severe abnormalities of con-nective tissue due to defective cross-linking of collagen and elastin. They eventuallysuccumb to anemia and cardiovascular damage, the latter apparently due to thedefective connective tissue metabolism. We evaluated the effects of nutritional cop-per deficiency upon collagen and elastin synthesis using short-term explant culturesof the medial portion of four successive segments of the descending aorta from 110-day-old pigs raised on a copper-deficient diet. Collagen synthesis was evaluated bycollagenase susceptibility, and elastin synthesis was quantified by immunoprecipita-tion with an antiporcine-elastin antiserum. In the normal developing aorta, elastinsynthesis was maximal in the upper thoracic aorta, while levels of collagen synthesiswere highest in the lower abdominal aorta. Both activities subsided by 110 dayspostpartum. Compared with controls, the copper-deficient group showed: 1) histo-pathologic changes confined to the luminal half of the thoracic aorta; 2) a 1.3- to 1.6-fold increase in cellularity along the entire length of the organ; 3) a 1.3- to 2.4-foldincrease in relative collagen synthesis, the greatest change occurring in the thoracicportion; 4) a 3- to 4-fold increase of relative elastin synthesis in the thoracic aorta, theabdominal aorta remaining unchanged; 5) 4- to 10-fold increases in collagen produc-tion; and 6) a greater than 15-fold increase in elastin production by the tissue of thethoracic aorta. Although copper deficiency produced a complex set of alterations, thecombination of defective cross-linking, hypertension, and increased cardiac outputappeared to be responsible for the hyperplasia of vascular smooth muscle and astriking induction of connective tissue production. These effects may represent anauthentic compensatory mechanism.(Arteriosclerosis 6:98-104, January/February 1986)
Collagen and elastin are the connective tissueproteins responsible for the structural integrity of
the aorta.12 They impart the resiliency and tensilestrength required to withstand the hydrostatic pres-sures exerted by the heart. Dietary copper directlyinfluences connective tissue biosynthesis due to itsrequirement in lysyl oxidase activity.3 This enzymeplays a major role in the cross-linking of collagen a-chains, collagen molecules, and tropoelastin mole-cules by catalyzing oxidative deamination of lysyl
From the Department of Pathology, University of Utah School ofMedicine and Veteran's Administration Medical Center, Salt LakeCity, Utah.
Address for reprints: Jeffrey M. Davidson, Ph.D., Research Ser-vice, 151C, Veterans Administration Medical Center, Salt LakeCity, Utah 84148.
This work was supported by the Veterans Administration, theMontana Heart Association, and by a gift from R. J. ReynoldsIndustries.
Received November 13,1984; revision accepted July 11,1985.
residues which then condense to form characteristic,covalent cross-links.4 Previous studies have shownthat removal of copper from the diet of various juve-nile animals can produce a diseased state in partresembling lathyrism but accompanied by anemiaand cardiac hypertrophy.5 The structural abnormali-ties associated with copper deficiency are pro-nounced in many connective tissues including theaorta and coronary arteries. These organs becomefragile and prone to dissecting aneurysms. There isgross anatomical and histological damage to the tu-nica media, and the biochemical composition of thetissue is altered.6
Nutritional copper deficiency has been extensivelystudied with respect to collagen and elastin accumu-lation.7"10 Altered cross-linking affects collagentypes,11 amino acid composition,12 mechanical prop-erties,5 glycosaminoglycan content,1314 and colla-gen degradation in the vascular wall.15 Little isknown, however, about the way in which connectivetissue cells respond to alterations in the mechanical
98
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

INDUCTION OF COLLAGEN AND ELASTIN SYNTHESIS Hill and Davidson 99
properties of the extracellular matrix. The combinedeffects of copper deficiency on hematopoiesis andconnective tissue cross-linking provide a complex,but intriguing, form of vascular disease.
We have recently shown that in the newborn pig,collagen synthesis and concentration progressivelyincrease in the aorta distal from the heart, while elas-tin synthesis and accumulation follow the oppositetrend.18 The presence of these biosynthetic gradi-ents determines the regional physical properties ofthe vascular wall. Recent studies have further indi-cated that the shape of these biosynthetic gradientsis a function of developmental age and that syntheticactivity subsides to basal levels by 110 days (David-son JM, Hill KE, and Alford J, unpublished results).However, it is not known how altered cross-linking oranemia-induced hypertension affects these patternsof synthetic activity.
In this study, we describe the altered patterns ofconnective tissue biosynthesis in the copper-defi-cient aorta and the relationship of these changes toposition along and within the vessel. We demon-strate that the tissues of the arterial wall responddramatically to the effects of copper deficiency, bywhat appears to be compensatory hypertrophy andincreased connective tissue biosynthesis by the vas-cular wall.
MaterialsAnimals
We obtained 3-day-old pigs from the Departmentof Animal Science, Brigham Young University, andstudied three normal and four experimental animals.The experimental group was placed on a copper-deficient diet,17 while the control animals were fed asimilar but copper-supplemented diet. The animalswere maintained for 15 weeks on their respectiveregimens and then sacrificed by an intravenous in-jection of 50 mg of sodium barbital per kilogram ofanimal weight.
Isolation of Aortic Segments
The experimental protocol was similar to previous-ly published procedures.16 The aortas were removedfrom the conus arteriosus to the iliac bifurcation bysterile dissection, and the tissues were further dis-sected free of fat, lymph nodes, and adventitia. Foursegments were analyzed for biosynthesis: A, conusarteriosus to the upper midpoint of the thoracic aorta;B, upper midpoint to the diaphragm; C, diaphragm tothe lower midpoint of the abdominal aorta; D, lowermidpoint to the iliac bifurcation.
Hlstologlcal Procedures
Tissue strips were fixed in 10% buffered formalinand processed by standard methodologies. Hema-toxylin and eosin, Masson's trichrome, Verhoeff-VanGieson, and alcian blue stains were used to estimatecellularity and discriminate among collagen, elastic
tissue, and glycosaminoglycans, respectively. Nu-clei were counted by using a microscope equippedwith a 1 cm2 grid in a x i o ocular to estimate celldensity.
DNA Content
Strips of each segment were weighed, lyophilized,minced in 0.05 N NaOH, and extracted overnight at4° C. The DNA content of the tissue extract wasdetermined by a fluorometric assay.18 Data were nor-malized to the wet weight of the tissue. Parallel de-terminations were done by extraction in 8 M guani-dinium-HCI, precipitation with perchloric acid (1.0M), and a diphenylamine assay, as previously de-scribed.19
Biosynthetic Labeling of Collagen and Elastln
Tissues were minced and incubated in Dulbecco'smodified Eagle's medium containing 20 /xCi/ml 3H-proline (New England Nuclear, > 100 Ci/mmol) for 3hours at 37° C on a gyratory shaker bath.16 Labelingwas terminated by aspirating the media and rinsingwith ice-cold phosphate-buffered saline. Tissue wasresuspended in 3 to 5 ml of 0.5 M acetic acid contain-ing 1 fig/rr\\ pepstatin and 10 mM iodoacetic acid.Samples were then homogenized and extractedovernight at 4° C. Homogenates were dialyzed firstagainst 10 volumes and then extensively against 0.5M acetic acid. The initial dialysate was saved forestimation of praline pool size.1820 The sampleswere clarified by centrifugation and the supernatantswere used for biosynthetic quantification.
Quantification of Protein Synthesis
Collagen synthesis was determined by the amountof radioactivity released into trichloroacetic (TCA)-soluble material after limited digestion with highlypurified bacterial collagenase.21 Elastin synthesiswas estimated by immunoprecipitation of the radiola-beled material with a antiporcine-a-elastin anti-serum.1622 Absolute rates of protein synthesis wereestimated from the specific activity of dialyzable freepraline as determined by amino acid analysis andliquid scintillation counting.1620
Results
Anatomic and Hlstologlcal Differences
The gross appearance of the copper-deficient aor-tas was markedly different from that of aortas fromnormal animals. The entire organ was paler, waxenin appearance, thicker, and more friable. The differ-ences were more apparent in the thoracic area thanin the abdominal area and were consistent with pre-viously published histological results.23'24
The most striking morphologic feature was the in-crease in the thickness and the diameter of the tho-racic aorta, almost twice the normal size. The ab-dominal aorta did not show any noticeable distortion
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

100 ARTERIOSCLEROSIS VOL 6, No 1, JANUARY/FEBRUARY 1986
Table 1. Cellularlty
Analysis
of Aortic Segments
Thoracic
A
segments
B
Abdominal
C
segments
D
Biochemical (cells per gram tissue wet weight (x 106))Copper-deficient 493 ±16 521 ±64Normal 376±46 332±15Change* +1.3 +1.6p <0.05 <0.05
Histologic (nuclei per mm2)Copper-deficient 16.3 ±2.1Normal 10.8 ±0.8Change* +1.5
346±16225±19
+ 1.5<0.01
325 ±33230±13
+ 1.4<0.05
11.5±0.38.4±0.2
+ 1.4
Values are means ± SEM. Probability factor (p) was determined by one-tailed f test. Segment A =conus arteriosus to the upper midpoint of the thoracic aorta; Segment B = upper midpoint to thediaphragm; Segment C = diaphragm to the lower midpoint of the abdominal aorta; Segment D = lowermidpoint to the iliac bifurcation.
'Copper-deficient divided by normal values.
when compared with normal tissue (data not shown).The copper-deficient aorta showed a significant in-crease in the number of medial smooth muscle cellsper unit area throughout the length of the organ(Table 1). Subjectively, the ratio of connective tissuematrix to cells appeared to be lower in the thoracicaorta and higher in the abdominal portion.
Histochemical staining of the normal aorta showeda regular staining pattern throughout the length andwidth of the organ; that is, collagen, elastin, and gly-cosaminoglycans were evenly distributed within themedia. As shown in previous studies,2324 copper-deficient thoracic tissue showed that the elastic la-mellae of the internal, luminal portion were split orfragmented in appearance, and gtycosaminoglycanswere increased and appeared to be pooled in theinterfibrillar spaces.13 The outer portion of the mediawas less affected than the inner, and the abdominalsegments did not show any extensive histologicaldamage (data not shown).
Cellularlty
Comparison of the guanidine-diphenylamine andfluorimetric DNA assays showed the former methodto give slightly lower and more variable results thanaccepted literature values in connective tissue. Con-sequently, the fluorimetric data were utilized as thebasis for determining cellularity.
Based on the DNA content per wet weight, thenormal aorta showed similar cell densities in Seg-ments A through D, averaging about 3 x 108 cells/gwet weight or 0.2% (Table 1). In contrast, the aver-age number of cells per unit weight was at least 50%greater in the copper-deficient aortas. Microscopicenumeration of cell nuclei corresponded to the val-ues derived from biochemical analysis (Table 1).
Collagen and Elastin Biosynthesis
Explanted aortic segments were incubated understandardized conditions which gave linear incorpor-ation of 3H-proline with time, and the tissues were
extracted with 0.5 M acetic acid, which solubilizedover 90% of incorporated radioactivity. The TCA-insoluble proteins were then analyzed for relativecollagen and elastin synthesis.
Figure 1 presents the biochemical data from nor-mal and copper-deficient tissues. The normal group(white bars) showed uniform relative collagen syn-thesis and absolute production (synthesis correctedfor precursor pool specific activity) along the longitu-dinal axis of the organ. In contrast, the copper-defi-cient group (dark bars) showed a prominent de-creasing gradient of collagen synthesis fromSegments A to D (12.20% ± 1.03% to 5.57% ±0.48%). The greatest variation from normal was theelevated synthesis in the upper thoracic aorta. Netproduction of collagen per cell increased dramatical-ly, being over 10-fold higher in the copper-deficientthoracic segments.
Relative elastin synthesis in normal 110-day pigsexhibited a shallow, decreasing gradient which wasparalleled in the absolute production of elastin. Thecopper-deficient aortas had a much steeper gradientof synthesis (17.0% ± 1.56% to 2.5% ± 0.1%).Relative synthetic activity increased fourfold in Seg-ment A, and Segment B had a threefold increase,while the abdominal aorta (Segments C and D) re-mained similar to the normal. The net production ofelastin per cell was markedly increased in the thorac-ic aorta and elevated in the entire organ.
Figure 1 also illustrates the total protein productionof the four segments. Production progressively in-creased in the more distal aortic segments in controlanimals, but a similar protein production profile in theexperimental animals was considerably higher.
To determine the actual biosynthetic output of theaortic tissue, the production per cell was multipliedby the cell density of the tissue. Table 2 shows theaverage total protein, collagen, and elastin produc-tion of copper-deficient aortas compared to normalaortas. These data combine the effects of increasedcellular production and cellularity. Thus, the net pro-duction of collagen increased markedly (> 15-fold) in
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

INDUCTION OF COLLAGEN AND ELASTIN SYNTHESIS Hill and Davidson 101
IX (/>
Effects of Copper Deficiencyon Segmental Collagen and Eiastin Production
Collagen Synthesis
Collagen Production
Eiastin Synthesis
Eiastin Production
Total Protein Production
..i
< £
1600
1200 _
1 800_a
400_ 1Segment: A
Anenosis \
BDiaphragm
1hc
1h
D
- ^ Illiac" ^ Bifurcation
Rgure 1. Connective tissue protein biosynthesis in copper-deficient and normal aortas.Collagen synthesis and production, eiastin synthesis and production, and total protein pro-duction are compared between the copper-deficient (•) and normal (a) pig aortas. Thediagram of the aorta corresponds to the segmental dissection of the intact organ. Error barsrepresent SEM, and statistical significance (f test) is shown by asterisks (*** = p < 0.001;" = p < 0.05; * = p<0 .1 ) .
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from
102 ARTERIOSCLEROSIS VOL 6, No 1, JANUARY/FEBRUARY 1986
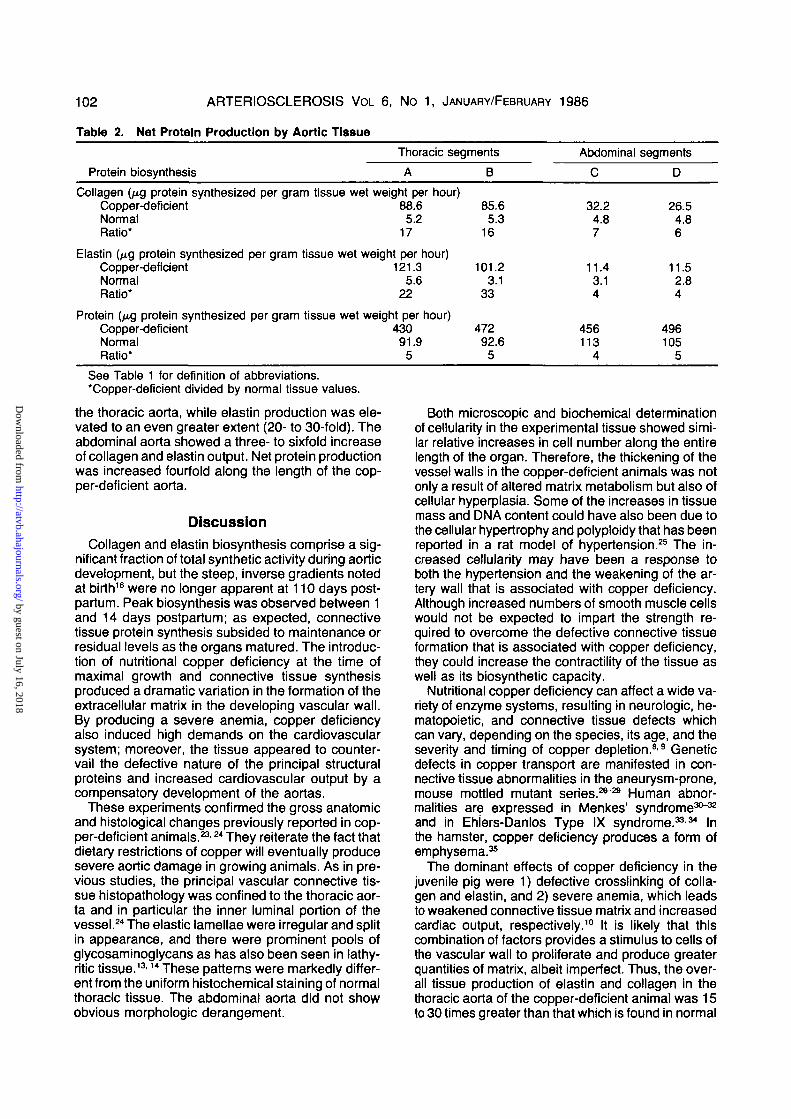
Table 2.
Protein
Net Protein
biosynthesis
Production by Aortic Tissue
Thoracic
A
segments
B
Abdominal
C
segments
D
Collagen (fig protein synthesized per gram tissue wet weight per hour)Copper-deficient 88.6 85.6Normal 5.2 5.3Ratio* 17 16
Elastin (^g protein synthesized per gram tissue wet weight per hour)Copper-deficient 121.3 101.2Normal 5.6 3.1Ratio* 22 33
Protein (^g protein synthesized per gram tissue wet weight per hour)Copper-deficient 430 472Normal 91.9 92.6Ratio* 5 5
32.24.87
11.43.14
456113
4
26.54.86
11.52.84
496105
5See Table 1 for definition of abbreviations.'Copper-deficient divided by normal tissue values.
the thoracic aorta, while elastin production was ele-vated to an even greater extent (20- to 30-fold). Theabdominal aorta showed a three- to sixfold increaseof collagen and elastin output. Net protein productionwas increased fourfold along the length of the cop-per-deficient aorta.
DiscussionCollagen and elastin biosynthesis comprise a sig-
nificant fraction of total synthetic activity during aorticdevelopment, but the steep, inverse gradients notedat birth16 were no longer apparent at 110 days post-partum. Peak biosynthesis was observed between 1and 14 days postpartum; as expected, connectivetissue protein synthesis subsided to maintenance orresidual levels as the organs matured. The introduc-tion of nutritional copper deficiency at the time ofmaximal growth and connective tissue synthesisproduced a dramatic variation in the formation of theextracellular matrix in the developing vascular wall.By producing a severe anemia, copper deficiencyalso induced high demands on the cardiovascularsystem; moreover, the tissue appeared to counter-vail the defective nature of the principal structuralproteins and increased cardiovascular output by acompensatory development of the aortas.
These experiments confirmed the gross anatomicand histological changes previously reported in cop-per-deficient animals.23'24 They reiterate the fact thatdietary restrictions of copper will eventually producesevere aortic damage in growing animals. As in pre-vious studies, the principal vascular connective tis-sue histopathology was confined to the thoracic aor-ta and in particular the inner luminal portion of thevessel.24 The elastic lamellae were irregular and splitin appearance, and there were prominent pools ofglycosaminoglycans as has also been seen in lathy-ritic tissue.13-14 These patterns were markedly differ-ent from the uniform histochemical staining of normalthoracic tissue. The abdominal aorta did not showobvious morphologic derangement.
Both microscopic and biochemical determinationof cellularity in the experimental tissue showed simi-lar relative increases in cell number along the entirelength of the organ. Therefore, the thickening of thevessel walls in the copper-deficient animals was notonly a result of altered matrix metabolism but also ofcellular hyperplasia. Some of the increases in tissuemass and DNA content could have also been due tothe cellular hypertrophy and polyploidy that has beenreported in a rat model of hypertension.25 The in-creased cellularity may have been a response toboth the hypertension and the weakening of the ar-tery wall that is associated with copper deficiency.Although increased numbers of smooth muscle cellswould not be expected to impart the strength re-quired to overcome the defective connective tissueformation that is associated with copper deficiency,they could increase the contractility of the tissue aswell as its biosynthetic capacity.
Nutritional copper deficiency can affect a wide va-riety of enzyme systems, resulting in neurologic, he-matopoietjc, and connective tissue defects whichcan vary, depending on the species, its age, and theseverity and timing of copper depletion.89 Geneticdefects in copper transport are manifested in con-nective tissue abnormalities in the aneurysm-prone,mouse mottled mutant series.2*"29 Human abnor-malities are expressed in Menkes' syndrome30"32
and in Ehiers-Danlos Type IX syndrome.3334 Inthe hamster, copper deficiency produces a form ofemphysema.35
The dominant effects of copper deficiency in thejuvenile pig were 1) defective crosslinking of colla-gen and elastin, and 2) severe anemia, which leadsto weakened connective tissue matrix and increasedcardiac output, respectively.10 It is likely that thiscombination of factors provides a stimulus to cells ofthe vascular wall to proliferate and produce greaterquantities of matrix, albeit imperfect. Thus, the over-all tissue production of elastin and collagen in thethoracic aorta of the copper-deficient animal was 15to 30 times greater than that which is found in normal
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

INDUCTION OF COLLAGEN AND ELASTIN SYNTHESIS Hill and Davidson 103
animals. Increased production appears to be a com-pensatory response by the tissues to the alteredproperties of the extracellular matrix.
A limited examination of the relative increases inboth Type I collagen mRNA and elastin mRNA hassupported the conclusions of this study (Alford J, HillKE, and Davidson JM, unpublished results), butmore precise quantitation is required to determinewhether the absolute, as well as relative, contents ofthese transcripts have been amplified by vascularcells from the copper-deficient pig.
Both relative and absolute production of collagenand elastin were markedly stimulated in the copper-deficient tissue, and the greatest effects were in theupper thoracic segment, nearest the heart. This in-crease was great enough to reverse the proximodis-tal gradient of collagen synthesis and steepen thegradient of elastin gene expression to levels ap-proaching those of the newborn animal.
Nevertheless, the extent to which collagen andelastin production were stimulated was obviouslygreater than the increases in contents of these pro-teins. Extensive degradation of newly synthesizedsoluble precursors could have accounted for this dis-crepancy. Previous studies in the copper-deficientchick have shown that soluble elastin is inefficientlyconverted to an insoluble form,36 and more recentstudies indicate a very high rate of turnover for tro-poelastin which fails to be cross-linked.37 Furtherstudies should be able to establish the extent of rapidproteolysis in this experimental system.
These results show that the responses of the cop-per-deficient porcine aorta were localized within cer-tain regions of the vessel. There was an apparentinduction of collagen and elastin gene expression,since the synthesis and production were markedlydifferent from a normal developing vascular tissue. Agreatly increased cell density and an elevated netproduction of connective tissue matrix were reflec-tive of the tissues' compensatory response to vascu-lar damage. Despite the pleiotropic consequences ofcopper deficiency, this model may be instructive inunderstanding the response of vascular tissue to in-creased physiologic and mechanical stress.
AcknowledgmentsWe thank the Learning Resource Service for illustrations, Mary
Ann McDonnell for supplying pig tissues, Richard Phelps andDavid Pinnick for technical assistance, Trudy Childs and Gwene-vere Shaw for manuscript preparation, and Lawrence B. Sand-berg for advice and critical commentary.
References1. Grant RL. Content and distribution of aortic collagen, elastin
and carbohydrate in different species. J Atherosclerosis Res1967;7:463-472
2. Wollnaky H, Glagov S. Comparison of abdominal and tho-racic aortic medial structure in mammals. Circ Res1969;25:677-686
3. Slegel RC, Martin GR. Collagen-crosslinking. Enzymaticsynthesis of lysine-derived aldehydes and the production ofcrossllnked components. J Biol Chem 1970:245:1653-1658
4. Sandberg LB, Davidson JM. Elastin and its gene. In: MeamMTW, ed. Peptide and protein reviews, vol 3. New York:Marcel Dekker, 1984:169-226
5. Walsman J, Cancllla P, Coulson W. Cardiovascular studieson copper-deficient swine. II. Mechanical properties of theaorta. Lab Invest 1969:21:548-554
6. Coulson WF, Carries WWH. Cardiovascular studies on cop-per-deficient swine; XIII. The effect of chronic copper defi-ciency on the cardiovascular system of miniature pigs. LabInvest 1962;1t:1316-1321
7. Smith D, Carries W. Biosynthesis of soluble elastin by pigaortic tissue in vitro. J Biol Chem 1973;248:8157-8161
8. Carnes W. Role of copper in connective tissue metabolism.Fed Proc 1971:30:995-1000
9. Klvlrlkko K, Pettonen L Abnormalities in copper metabo-lism and disturbances in the synthesis of collagen and elastin.Med Biol 1982;60:45-48
10. Welssman N, Shields G, Carries W. Cardiovascular studieson copper-deficient swine; IV. Content and solubility of theaortic elastin, collagen and hexosamine. J Biol Chem1963:238:3115-3118
11. Dawson R, Milne G, Williams R. Changes in the collagen ofrat heart in copper-defldency-induced cardiac hypertrophy.Cardiovasc Res 1982;16:559-565
12. Smith DW, Welssman N, Carnes W. Cardiovascular studieson copper-deficient swine; XII. Partial purification of a solubleprotein resembling elastin. Biochem Biophys Res Comm1968;31:309
13. Linker A, Coulson W, Carnes W. Cardiovascular studies oncopper-deficient swine; VI. The mucoporysacchartde compo-sition of aorta and cartilage. J Biol Chem 1964;239:1690-1693
14. Pasquall-Ronchettl I, Bressan GM, Fornlerl C, Baccaranl-Corrtri M, Volpln D. Elastic fiber-associated glycosaminogly-cans in B-aminopronitrile-induced lathyrism. Exp Mol Pathol1984;40:235-245
15. Vater CA, Harris ED, Slegel RC. Native cross-links in colla-gen fibrils induced resistance to human synovial collagenase.Biochem J 1979:181:639-645
16. Davidson J, Hill K, Mason M, Giro M. Longitudinal gradientsof collagen and elastin gene expression in the porcine aorta. JBiol Chem 1984:260:1901-1908
17. Sandberg LB, Wort TB. Production and isolation of solubleelastin from copper-deficient swine. Methods Enzymol1982:82:657-665
18. Hlnegsrdner RT. An improved fluorometric assay for DNA.Anal Biochem 1971:39:197-201
19. Burton K. A study of the conditions and mechanisms of thediphenylamine reaction for the colormetric estimation ofdeoxyribonucleic add. Biochem J 1956:62:315-323
20. Shlbahara S, Davidson JM, Smith K, Crystal R. Modulationof tropoelastin production and elastin messenger ribonucleicadd activity in developing sheep lung. Biochemistry1981:20:6577-6584
21. Peterkofsky B, Chojkler M, Bateman J. Determination ofcollagen synthesis in tissue and cell culture systems. In:Fuefhmayr H, ed. Immunochemistry of the extracellular ma-trix, Florida: CRD Press, 1982;19-^9
22. Giro MG, Hill K, Sandberg L, Davidson J. Quantitation ofelastin production in cultured vascular smooth muscle cellsby a sensitive and spedfic enzyme-linked immunoassay. Col-lagen Rel Res 1984;4:24-34
23. Shields GS, Coulson W, Klmball D, Carnes W, CartwrlghtG, Wlntrobe M. Lesions caused by copper depletion in thechicken. Am J Pathol 1962;41:603-617
24. Kadar A, Joos A, Jelllnek H. Studies on copper metabolism;XXXII. Cardiovascular lesions in copper-defident swine.Light and electron microscopic study. Paroi Arterielle — Arte-rial Wall, T 1977;4:45-55
25. Owens GK, Rablnovltch PS, Schwartz SM. Smooth musclecell hypertrophy versus hyperplasia in hypertension. ProcNatl Acad Sd USA 1981;78:7759-7763
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

104 ARTERIOSCLEROSIS VOL 6, No 1, JANUARY/FEBRUARY 1986
26. Rowe DW, McGoodwin EB, Martin GR, Grahn D. De-creased lysl oxidase activity in the aneurysm-prone, mottledmouse. J Biol Chem 1977;252:939-942
27. Rowe DW, McGoodwin EB, Martin GR, Sussman MD,Grahn D, Faris B, Franzblau C. A sex-linked defect in thecross-linking of collagen and elastin associated with the mot-tled locus in mice. J Exp Med 1974;139:180-194
28. Starcher B, Madaras JA, Fisk D, Perry EF, Hill CH. Abnor-mal cellular copper metabolism in the blotchy mouse. J Nutr1978;108:1229-1233
29. Starcher BC, Madaras JA, Tepper AS. Lysyl oxidase defi-ciency in lung and fibroblasts from mice with hereditary em-physema. Biochem Biophys ResCommun 1977;78:706-712
30. Goka TJ, Stevenson RE, Hefferan PM, Howell RR. Menkesdisease: A biochemical abnormality in cultured human fibro-blasts. Proc Natl Acad Sci USA 1976;73:604-606
31. Camakaris J, Danks DM, Ackland L, Cartwright E, BorgerP, Cotton RGH. Altered copper metabolism in cultured cellsfrom human Menkes' syndrome and mottled mouse mutants.
Biochem Genet 1980;18:117-13132. Procopis P, Camakaris J, Danks DM. A mild form of
Menkes steely hair syndrome. J Pediatr 1981;98:97-9933. Kuivaniiemi H, Peltonen L, Palotie A, Kaitila I, Kivirikko K.
Abnormal copper metabolism and deficient lysyl oxidase ac-tivity in a heritable connective tissue disorder. J Clin Invest1982:69:730-733
34. MacFarlane JD, Hollister DW, Weaver DD, Brand KD, Luz-zatti L, Biegel AA. A new Ehlers-Danlos syndrome with skel-etal dysplasia (abstr). Am J Hum Genet 1980;32:118A
35. Soskel NT, Watanabe S, Sandberg LB. Mechanisms of lunginjury in the copper-deficient hamster model of emphysema.Chest 1984;85:70s-73s
36. Buckingham K, Heng-Khoo CS, Dubick M, et al. Copperdeficiency and elastin metabolism in avian lung. Proc SocExp Biol Med 1981:166:310-319
37. Romero N, Tinker D, Himebaugh M, Hyde D, Rucker RB.Tropoelastin degradation in copper-deficient and lathryticchick aorta (abstr). Fed Proc 1985;44:663A
Index Terms: aorta • copper deficiency • collagen • elastin • crosslinking • protein synthesis • pig
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from

K E Hill and J M DavidsonInduction of increased collagen and elastin biosynthesis in copper-deficient pig aorta.
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1986 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleArteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.6.1.981986;6:98-104Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/6/1/98World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document.Permissions and Rights Question and AnswerFurther information about this process is available in theis being requested is located, click Request Permissions in the middle column of the Web page under Services.Clearance Center, not the Editorial Office. Once the online version of the published article for which permission
can be obtained via RightsLink, a service of the CopyrightArteriosclerosis, Thrombosis, and Vascular Biology Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 16, 2018http://atvb.ahajournals.org/
Dow
nloaded from