infection of differentiated epithelial cells by viruses of
TRANSCRIPT

University of Veterinary Medicine Hannover
Institute of Virology
Infection of differentiated epithelial cells by
viruses of the bovine respiratory disease complex
THESIS
Submitted in partial fulfillment of the requirements for the degree
-Doctor rerum naturalium-
(Dr. rer. nat.)
awarded by the University of Veterinary Medicine Hannover
by
Jana Kirchhoff
(Herzberg am Harz)
Hannover, Germany 2014

Supervisor: Prof. Dr. Georg Herrler
Advisory Committee: Prof. Dr. Georg Herrler
Prof. Dr. Christiane Pfarrer
Prof. Dr. Evgeni Ponimaskin
1st Evaluation: Prof. Dr. Georg Herrler
Institute of Virology
University of Veterinary Medicine Hannover, Foundation
Prof. Dr. Christiane Pfarrer
Department of Anatomy
University of Veterinary Medicine Hannover, Foundation
Prof. Dr. Evgeni Ponimaskin
Institute for Cellular Neurophysiology
Hannover Medical School
2nd Evaluation: Prof. Dr. Michael Veit
Institute of Virology
Free University Berlin
Date of oral exam: 22st April 2014
This work was financed by a grant of Deutsche Forschungsgemeinschaft (SFB 587,
TPA1).

To my family


Contents
List of Abbreviations ................................................................................................. I
List of Figures .......................................................................................................... III
Abstract ..................................................................................................................... V
Zusammenfassung ................................................................................................. VII
1 Introduction ........................................................................................................ 1
1.1 Bovine respiratory disease complex .............................................................. 1
1.1.1 Infection of cattle ..................................................................................... 1
1.1.2 Infection of other ruminant species ......................................................... 3
1.2 Respiratory syncytial virus (RSV) .................................................................. 4
1.2.1 Epidemiology .......................................................................................... 4
1.2.2 Clinical signs and Pathology ................................................................... 4
1.2.3 Vaccination ............................................................................................. 5
1.2.4 Taxonomy ............................................................................................... 6
1.2.5 Virus structure ......................................................................................... 6
1.2.6 Viral life cycle .......................................................................................... 8
1.3 Bovine parainfluenza virus type 3 (BPIV3) .................................................... 9
1.4 Bovine herpesvirus type 1 (BHV-1) ............................................................. 11
1.4.1 Epidemiology ........................................................................................ 11
1.4.2 Clinical signs and Pathology ................................................................. 11
1.4.3 Vaccination ........................................................................................... 12
1.4.4 Taxonomy ............................................................................................. 13
1.4.5 Virus structure ....................................................................................... 13
1.4.6 Viral life cycle ........................................................................................ 14
1.5 The respiratory epithelial barrier .................................................................. 16
1.5.1 Basic functions of the respiratory epithelium ......................................... 16
1.5.2 Polarized virus entry ............................................................................. 17
1.5.3 Cultures for differentiated respiratory epithelial cells ............................. 18
1.6 Aim of the study ........................................................................................... 20

2 Publication I ...................................................................................................... 23
Three viruses of the bovine respiratory disease complex apply different strategies
to initiate infection
3 Publication II ..................................................................................................... 25
Infection of differentiated airway epithelial cells from caprine lungs by viruses of the
bovine respiratory disease complex
4 Discussion ........................................................................................................ 27
4.1 In vitro cultures for differentiated respiratory epithelial cells ........................ 27
4.2 Infection strategies of bovine parainfluenza virus type 3 ............................. 29
4.2.1 Infection of well-differentiated bovine airway epithelial cells ................. 29
4.2.2 Infection of well-differentiated caprine airway epithelial cells ................ 31
4.3 Infection strategies of bovine herpesvirus type 1 ......................................... 32
4.3.1 Infection of well-differentiated bovine airway epithelial cells ................. 32
4.3.2 Infection of well-differentiated caprine airway epithelial cells ................ 34
4.4 Infection strategies of bovine respiratory syncytial virus .............................. 35
4.4.1 Infection of well-differentiated bovine airway epithelial cells ................. 35
4.4.2 Infection of well-differentiated caprine airway epithelial cells ................ 38
4.5 Summary and Outlook ................................................................................. 40
4.5.1 Strategies of viruses of the BRDC to infect the bovine host .................. 40
4.5.2 Goats as a virus reservoir ..................................................................... 41
5 References ........................................................................................................ 43
6 Appendix ........................................................................................................... 62
7 Acknowledgements ......................................................................................... 63

I List of Abbreviations
List of Abbreviations
ALI Air-liquid interface
BAEC Bovine airway epithelial cells
BHV-1 Bovine herpesvirus type 1
BPIV3 Bovine parainfluenza virus type 3
BRD Bovine respiratory disease
BRDC Bovine respiratory disease complex
BRSV Bovine respiratory syncytial virus
BSA Bovine serum albumin
CapHV-1 Caprine herpesvirus type 1
CO2 Carbon dioxide
CPE Cytopathic effect
CRSV Caprine respiratory syncytial virus
Cy3 Indocarbocyanine
DAPI 4’,6’-Diamidino-2-phenylindole
DCs Dendritic cells
DMEM Dulbecco’s modified Eagle medium
dpi days post infection
et al. et alii (and others)
FFU Focus-forming units
Fig. Figure
FITC Fluorescine isothiocyanate
h hour
hpi hours post infection
HRSV Human respiratory syncytial virus
IFN Interferon
L liter
M Molarity, - molar
MDBK Madin-Darby bovine kidney
MDCK Madin-Darby canine kidney

II List of Abbreviations
min minute
MOI Multiplicity of infection
mRNA messenger RNA
NA Neuraminidase
PBS Phosphate buffered saline
PCLS Precision-cut lung slices
PFA Paraformaldehyde
pH potentia Hydrogenii
TEER Transepithelial electrical resistance
TTF-1 Thyroid transcription factor-1
ZO-1 Zonulae occludentes-1 protein
°C degree Celsius
α anti (antibodies) or alpha (sialic acids)
µ micro (gram or liter for example)
Ω ohm

III List of Figures
List of Figures
Figure 1-1 Factors contributing to the bovine respiratory disease complex ................ 2
Figure 1-2 Structure of respiratory syncytial virus ....................................................... 7
Figure 1-3 Schematic drawing of an Alphaherpesvirus particle ................................ 14
Figure 1-4 composition of the respiratory epithelium ................................................ 16
Figure 1-5 ALI culture system. .................................................................................. 19
Figure 1-6 Light microscopic (A) and confocal (B) picture of a typical PCLS ............ 20

IV -
-

V Abstract
Abstract
Infection of differentiated epithelial cells by viruses of the bovine
respiratory disease complex
Jana Kirchhoff
Respiratory tract infections of cattle are often associated with the bovine respiratory
disease complex (BRDC). BRDC is a multifactorial disease and several factors like
stress or reduced immunity contribute to the severity of disease. Bovine respiratory
syncytial virus (BRSV), bovine parainfluenza virus type 3 (BPIV3) and bovine
herpesvirus type 1 (BHV-1) are important viral pathogens associated with this
disease complex.
The aim of the thesis was to analyze the susceptibility of bovine airway epithelial
cells (BAEC) to the three viruses of the BRDC and the different viral strategies
concerning entry and release. For this purpose, two culture systems for well-
differentiated BAEC were used, which were previously established in our institute:
the air-liquid interface (ALI) system, where filter-grown respiratory BAEC undergo
mucociliary differentiation and precision-cut lung slices (PCLS), where cells are
maintained in their original structural conformation and stay vital for more than one
week. When the viruses were applied to these culture systems, different infection
strategies were observed: BPIV3 efficiently infected ciliated epithelial cells and
replicated to high titers. Entry and release were a) sialic acid-dependent and b)
localized to the apical side of the ALI cultures. In contrast, BAEC were largely
resistant to infection by BHV-1, which could be overcome by injury of the cell
monolayer or destruction of tight junctions. Moreover, BHV-1 infection was primarily
directed to basal cells. BRSV infected the respiratory epithelial cells inefficiently; in
contrast to BHV-1, this infection was not enhanced when the integrity of the epithelial
barrier was destroyed. Subepithelial cells and pneumocytes type I and II appeared to
be the target of infection by this virus.

VI Abstract
Furthermore, non-bovine ruminants such as goats can be considered as a target for
viruses of BRDC and therefore may serve as a virus reservoir as well. To address
this question, a culture system for differentiated caprine epithelial cells, caprine
PCLS, was established for the first time. When the three viruses of the BRDC were
applied to the cultures, it was observed that the infection pattern was similar to that
reported for bovine cultures. While infection of caprine cells by BPIV3 and BRSV was
as efficient as that observed for bovine cells, infection of BHV-1 required a tenfold
higher dose of infectious virus in comparison to the bovine culture system.
Nevertheless, BHV-1 displayed the same tropism for basal cell. These results
indicate that non-bovine ruminants are also important hosts for viruses of the BRDC
and should be considered in the development of control strategies.

VII Zusammenfassung
Zusammenfassung
Infektion differenzierter Epithelzellen durch Viren des Rindergrippe-
Komplexes
Jana Kirchhoff
Die Enzootische Bronchopneumonie (oder Rindergrippe-Komplex) ist eine
Infektionskrankheit des Respirationstraktes bei Rindern, die multifaktoriellen
Ursprungs ist. Neben zahlreichen viralen und bakteriellen Erregern tragen auch der
Immunstatus des Tieres sowie Umweltbedingungen zum Schweregrad der
Erkrankung bei. Wichtige Viren sind das bovine respiratorische Synzytialvirus
(BRSV), das bovine Parainfluenzavirus 3 (BPIV3) sowie das bovine Herpesvirus 1
(BHV-1).
Das Ziel dieser These war es, die Empfänglichkeit von bovinen primären
Lungenepithelzellen gegenüber den obig genannten Viren zu untersuchen sowie
deren Strategien zum Ein- und Austritt aus dem Epithelgewebe zu analysieren. Um
dieses Ziel zu erreichen, wurden zwei Kultursysteme für primäre respiratorische
Epithelzellen genutzt, die zuvor in unserer Arbeitsgruppe etabliert worden waren:
Zum einen handelt es sich hierbei um das Air-liquid interface (ALI)-System, in dem
Zellen aus der Rinderlunge isoliert und anschließend auf Filterkultureinsätzen zur
Ausdifferenzierung gebracht werden. Als Vergleichskultur wurden
Lungenpräzisionsschnitte (PCLS; precision-cut lung slices) verwendet. Diese
Schnitte haben den Vorteil, dass die respiratorischen Epithelzellen im ursprünglichen
Gewebeverband verbleiben und zudem für mindestens eine Woche vital sind.
Infektionsversuche zeigten, dass die drei Viren unterschiedliche Infektionsstrategien
verfolgen: BPIV3 infizierte die zilientragenden Epithelzellen sehr effizient und
replizierte zu hohen Titern. Des Weiteren ist die Infektion lokal ausgeprägt und war
abhängig von der Gegenwart von Sialinsäuren auf der Zelloberfläche. Im Gegensatz
dazu waren die Epithelzellen resistent gegenüber einer Infektion mit BHV-1. Diese
Resistenz konnte jedoch überwunden werden, wenn den Viren Zugang zur basalen

VIII Zusammenfassung
Seite des Epithels verschafft wurde, z.B. durch chemische oder manuelle Eingriffe.
Zudem konnte gezeigt werden, dass nicht die differenzierten Epithelzellen selbst
infiziert wurden, sondern die Basalzellen, die als Vorläuferzellen der differenzierten
Epithelzellen fungieren. Wie schon in vorherigen Studien beschrieben wurde, hatte
das Flimmerepithel nur eine geringe Empfänglichkeit gegenüber einer BRSV-
Infektion; diese Unempfänglichkeit konnte nicht durch die oben bei BHV-1
beschriebenen Eingriffe am Epithel überwunden werden. Durch Infektion der
Lungenpräzisionsschnitte wurde deutlich, dass subepitheliale Zellen sehr effizient
durch BRSV infiziert wurden; bei diesen Zellen könnte es sich um dendritische Zellen
handeln. Zudem stellte sich heraus, dass Typ-I- und Typ-II-Pneumozyten sehr
empfänglich für eine BRSV-Infektion sind.
Es wird angenommen, dass auch kleine Wiederkäuer wie Ziegen durch Viren des
Rindergrippe-Komplexes infiziert werden und somit als Virusquelle dienen können.
Um diese Thematik zu untersuchen, wurde ein Kultursystem für enddifferenzierte
Epithelzellen aus der Ziegenlunge etabliert: caprine Lungenpräzisionsschnitte.
Infektionsversuche mit den obig genannten Viren zeigten, dass die
Infektionsstrategien ähnlich waren wie im bovinen System sowie gleiche Zielzellen
infiziert wurden. Zudem wurde deutlich, dass die Infektion mit BPIV3 und BRSV
ähnlich effizient war wie in den bovinen Kultursystemen; die Infektion mit BHV-1
benötigte jedoch einen zehnfach höheren Titer. Auch hier waren die Basalzellen Ziel
der Infektion. Diese Ergebnisse zeigen auf, dass auch nicht-bovine Wiederkäuer als
Reservoir für Viren des Rindergrippe-Komplexes dienen können und somit bei
Interventionsmaßnahmen berücksichtigt werden sollten.

Introduction 1
1 Introduction
1.1 Bovine respiratory disease complex
1.1.1 Infection of cattle
Virus infections of the respiratory tract are the most frequent cause of viral diseases
worldwide. They range from mild infections of the upper respiratory tract such as
common colds to severe lower respiratory tract infections like life-threatening
pneumonia (Garibaldi et al., 1985). Bovine respiratory disease complex (BRDC) is a
common term for respiratory disease in cattle. Other names are “shipping fever” and
“enzootic bronchopneumonia”. BRDC is considered to be the major cause of
morbidity and mortality within feedlot and dairy cattle (Yates, 1982; Taylor et al.,
2010; Iglseder et al., 2011).
Bovine respiratory disease (BRD) has a multifactorial origin; therefore several factors
contribute to the severity of disease (Figure 1-1). Among viruses, bovine respiratory
syncytial virus (BRSV), bovine parainfluenza virus type 3 (BPIV 3) and bovine
herpesvirus type 1 (BHV-1) are causative agents (Cusack et al., 2003; Ellis, 2009).
These were identified by immunohistochemistry (Fulton et al., 2000; Juarez Barranco
et al., 2003) or serological surveys (Durham et al., 1991; Fulton et al., 2000;
Hagglund et al., 2007). According to these reports, viral infection can result in
disease without bacterial co-infection, but the most common scenario is that viruses
predispose to subsequent bacterial invasion. This is facilitated due to the viral ability
to decrease the clearance system of the lung (Lopez et al., 1976; Jones, 1983;
Cusack et al., 2003). The bacterial pathogens which are associated with disease are
Pasteurella multocida, Mannheimia haemolytica, Mycoplasma bovis and Histophilus
somni (Allen et al., 1991; Gagea et al., 2006; Rice et al., 2007; Srikumaran et al.,
2007; Confer, 2009). These were identified by isolation from animals suffering from
BRD (Harris and Janzen, 1989; Welsh et al., 2004; Gagea et al., 2006; Fulton et al.,
2009) or by serological examinations (Martin et al., 1989; Booker et al., 1999).
Further factors, which predispose to disease are the special anatomy of the bovine

Introduction 2
lung and the immune status of the host which for example might be impaired in
young animals due to deficient colostral transfer (Müller, 2012). Also, environmental
stressors are considered to be an important issue. One main stressor is
transportation to different places e.g. auction markets (Pinchak et al., 2004;
Sanderson et al., 2008). Others are weaning prior to sale, crowding and introduction
of new animals into a herd. Also, poor housing conditions have an important effect on
BRD (Sanderson et al., 2008; Ohlson et al., 2010; Taylor et al., 2010). How and to
what extent these factors contribute to the disease is widely debated (Taylor et al.,
2010).
Figure 1-1 Factors contributing to the bovine respiratory disease complex (see text for references).
BRD causes high economic losses worldwide; it was estimated for the US that BRD-
associated costs for the cattle industry amount to about 1 billion dollars per year
(Griffin, 1997; McVey, 2009). Costs are not only due to death losses but also include
treatment costs, reduction in feed efficiency, reproductive performance and milk
production. Expenses easily accumulate because reinfection is common (Van der
Poel et al., 1993; Griffin, 1997; Jim, 2009; Fulton and Confer, 2012). Diagnosis of
disease is difficult due to its multifactorial nature. Characteristic symptoms are

Introduction 3
elevated body temperature, respiratory signs, low appetite and depression (Allen et
al., 1991; Barbour et al., 1997; Booker et al., 1999).
1.1.2 Infection of other ruminant species
Pneumonia is also a serious disease in other ruminant species like sheep and goats
(Ackermann and Brogden, 2000). Many aspects concerning epidemiology,
pathogenesis and clinical signs are comparable to the outcomes in cattle populations
(Dungworth, 1993). Accordingly, infection of goats and sheep is caused by similar
pathogens as observed in the bovine counterpart. Alphaherpesviruses for instance
can be isolated from a wide variety of ruminant species and many of them share
antigenic properties (Yates, 1982). Especially caprine herpesvirus 1 (CapHV-1) and
BHV-1 exhibit a close relationship (Nixon et al., 1988; Whetstone and Evermann,
1988), also bovine and caprine RSV (CRSV), which have been suggested to belong
to one subgroup (Duncan and Potgieter, 1993; Alansari et al., 1999; Eleraky et al.,
2003), share antigenic relationship. As far as bacterial pathogens are concerned, M.
haemolytica is considered to be an important agent contributing to the severity of
disease (Brogden et al., 1998). Therefore, concerns about BRD eradication schemes
came up because it seems likely that viruses of BRDC can cross the species barrier.
Since respiratory viruses are transmitted directly through aerosols or by close contact
among infected animals, interspecies transmission is common as ruminants are often
kept in close contact to each other. Accordingly, antibodies to BRSV, BHV-1 and
BPIV3 were found in goats and sheep (Fulton et al., 1982; Elazhary et al., 1984; Van
der Poel et al., 1995; Yener et al., 2005). Moreover, sheep and goats developed
clinical signs after experimental infection with BHV-1 and BRSV (Lehmkuhl and
Cutlip, 1979; Engels et al., 1992; Meehan et al., 1994; Six et al., 2001). Also BPIV3
was found to infect sheep and to induce clinical signs (Woods et al., 1965; Hore,
1966; Stevenson and Hore, 1970; Rudolph et al., 2007). For goats, data on BPIV3
infection are rare (Yener et al., 2005). Based on these studies, sheep and goats have
been designated as a reservoir for viruses of BRD (Van der Poel et al., 1995; Thiry et
al., 2006). Further research addressing this question is important to analyze whether
virus isolates cocirculate in ruminant populations and whether cross-transmission of
these viruses is common (Geraghty et al., 1998).

Introduction 4
1.2 Respiratory syncytial virus (RSV)
1.2.1 Epidemiology
Bovine respiratory syncytial virus (BRSV) is the most common cause of respiratory
disease in cattle, especially in calves (Stott and Taylor, 1985). It was first isolated in
Europe in the early 1970s (Paccaud and Jacquier, 1970). BRSV is distributed all over
the world, and virus was isolated in different countries from both, beef and dairy
cattle (Smith et al., 1975; Lehmkuhl and Gough, 1977; Van Vuuren, 1990; Gagea et
al., 2006; Hagglund et al., 2006; Bidokhti et al., 2009; Saa et al., 2012). Infections
normally occur in autumn and wintertime (Stott et al., 1980; Van der Poel et al.,
1993), with a seroprevalence of 28-95% (Lehmkuhl and Gough, 1977; Collins et al.,
1988; Van Vuuren, 1990; Van der Poel et al., 1994; Elvander, 1996; Hagglund et al.,
2006; Saa et al., 2012), but can also take place in summer (Gershwin, 2007; Sacco
et al., 2012). Morbidity is high (60-80%) and mortality can increase up to 20% (Baker
et al., 1986; Elvander, 1996; Valarcher and Taylor, 2007). Especially young animals
are under high risk of getting infected (Kimman et al., 1988; Hagglund et al., 2006)
and infection even occurs under the protection of maternal antibodies (Van der Poel
et al., 1994). Though calves are under higher risk of getting severe clinical signs
(Stott et al., 1980; Kimman et al., 1988), adult animals are also at risk mainly in naïve
herds (Van der Poel et al., 1994). Reinfection is common but severity is reduced in
these cases (Kimman et al., 1987). It is not completely understood how the virus can
survive in a cattle population (Stott et al., 1980), but there is some evidence that
BRSV can persist in its host (Van der Poel et al., 1993; De Jong et al., 1996;
Valarcher et al., 2001).
1.2.2 Clinical signs and Pathology
BRSV is transmitted by aerosols or upon direct contact (Mars et al., 1999). The
incubation time takes between 2-5 days. The course of disease can be either
asymptomatic, restricted to the upper respiratory tract or spread to the lower airways
as well (Verhoeff et al., 1984). Clinical symptoms associated with mild cases are
cough and ocular and nasal discharge. Severe cases result in depression,
hyperthermia, anorexia and polypnea (Verhoeff et al., 1984). Microscopic lesions

Introduction 5
include necrotic and apoptotic epithelial cells, also occurrence of syncytia in the
bronchiolar or alveolar epithelium and infiltration of mononuclear cells is
characteristic (Viuff et al., 1996; Viuff et al., 2002; Brodersen, 2010).
BRSV predominantly infects respiratory epithelial cells but was also found to replicate
in type II pneumocytes (Viuff et al., 1996; Viuff et al., 2002). In vitro studies
demonstrated that RSV in general causes little or no cytopathic effect in epithelial cell
cultures; therefore, it is assumed that the pathology results from the host immune
response which is triggered by a skewed Th2 response (Hussell et al., 1997;
Kristjansson et al., 2005; Bueno et al., 2008; Antonis et al., 2010). In this context,
eosinophils, neutrophils and lymphocytes are activated which produce pro-
inflammatory cytokines and chemokines leading to destruction of the epithelium
(Valarcher and Taylor, 2007).
1.2.3 Vaccination
Vaccine design for prevention of BRSV and human RSV (HRSV) infection is still a
challenging task. For HRSV, formalin-inactivated vaccines were produced in the
1960s, but they were not only unable to prevent RSV infection but rather intensified
the clinical response upon natural infection. Until now, there is no approved vaccine
available (Kim et al., 1969; Prince et al., 1986; Gershwin et al., 1998; Schreiber et al.,
2000). As far as BRSV is concerned, there are several vaccines on the market, killed
as well as modified live virus vaccines. But also for the bovine counterpart, infections
combined with severe disease symptoms were observed in vaccinated animals
(Schreiber et al., 2000; Antonis et al., 2003). Successful vaccination in calves at the
age of 2 to 6 months is hampered by the presence of maternal antibodies (Valarcher
and Taylor, 2007). Furthermore, protection is not long-lasting (Sacco et al., 2012).
Efforts to develop improved vaccines are ongoing. Modified live vaccines lacking e.g.
the NS2 or NS1 genes appeared to be promising candidates (Valarcher et al., 2003)
but never got to the stage approved vaccines. Problems concerning vaccine design
include the heterogeneity within the RSV genomes since there is no proofreading
mechanism during virus replication (Valarcher et al., 2000; Brodersen, 2010). So far,
there is only one medication available for high risk patients; Palivizumab is a

Introduction 6
humanized monoclonal antibody, which prevents infection by binding to the F protein
of HRSV or reduces the severity of disease in already infected infants (Malley et al.,
1998; Singleton et al., 2003).
1.2.4 Taxonomy
Bovine respiratory syncytial virus (BRSV) belongs to the family Paramyxoviridae
within the order Mononegavirales. Among paramyxoviruses, it is classified in the
subfamily Pneumovirinae and the genus Pneumovirus (ICTV, 2012b). Other
pneumoviruses are ovine RSV (ORSV), caprine RSV (CRSV) (Lehmkuhl et al., 1980)
and pneumonia virus of mice (PVM). CRSV is most closely related to BRSV. A
characteristic feature of all pneumoviruses is the presence of two non-structural
proteins (Smith et al., 1979; Trudel et al., 1989).
BRSV strains are assigned to either of four antigenic subgroups: A, AB, B and
untyped (Schrijver et al., 1996). Within an antigenic subtype, the genetic variability of
the G protein between strains is less than 15 % (Prozzi et al., 1997). In contrast,
HRSV has been divided into two subgroups (Anderson et al., 1985; Cristina et al.,
1990) which exhibit a greater variability (Collins et al., 1990).
BRSV and HRSV are closely related and similar in their pathogenesis and
epidemiology (Sacco et al., 2012). Therefore, research about the bovine counterpart
also provides information about the human virus and vice versa (Van der Poel et al.,
1994; Valarcher and Taylor, 2007). Nevertheless, the tropism of BRSV and HRSV in
vivo is restricted to the natural host. The F and the NS proteins are major
determinants of the host range (Bossert and Conzelmann, 2002; Schlender et al.,
2003).
1.2.5 Virus structure
BRSV particles are surrounded by an envelope which is derived from the plasma
membrane of the host cell. The genome consists of single-stranded, non-segmented
RNA in a negative orientation. It comprises ~15,000 nucleotides and sequentially
encodes for 11 proteins which are translated from a gradient of transcription which

Introduction 7
decreases from the mRNA for the NS proteins to the mRNA for the L protein
(Valarcher and Taylor, 2007).
Figure 1-2 Structure of respiratory syncytial virus; adapted from Valarcher and Taylor (2007).
RSV encodes for three different surface glycoproteins which are incorporated as
separate spikes into the envelope (see Figure 1-2): the glycoprotein (G), the fusion
protein (F) and the small hydrophobic protein (SH). The glycoprotein G is a type II
membrane protein which is heavily glycosylated. It serves as an attachment protein;
blocking of G abolishes the virus binding (Levine et al., 1987). G is unique to RSV,
and its structure differs from those of other paramyxovirus attachment proteins
(Wertz et al., 1985; Langedijk et al., 1996). Apart from its attachment function, it is
the target of neutralizing antibodies (Valarcher and Taylor, 2007). The F protein
contributes to virus attachment. In addition, it has fusion activity inducing the fusion of
the viral membrane with the host cell membrane and the formation of syncytia
between adjacent cells. It is a type I membrane protein and highly conserved among
BRSV isolates (Valarcher et al., 2000). It is synthesized as a precursor protein F0
and proteolytic cleavage at two multibasic furin cleavage sites generates a fusion-

Introduction 8
active heterodimer made up from an F1 and an F2 subunit which are connected by a
disulphide bridge (Gonzalez-Reyes et al., 2001; Zimmer et al., 2001). The process
goes along with the removal of a small intervening peptide with the length of 27
amino acids (pep 27) (Zimmer et al., 2001; Zimmer et al., 2003), which is released as
a virokinin that can induce bronchoconstriction (Zimmer et al., 2003) and eosinophil
recruitment (Valarcher and Taylor, 2007). The role of the SH protein in virus
replication is not known (Karger et al., 2001). The envelope of the virus surrounds a
helical nucleocapsid which comprises the genomic RNA. It is associated with the
nucleoprotein (N), which protects against degradation by RNases (Samal et al.,
1991; Mallipeddi et al., 1996), the phosphoprotein (P), which regulates transcription
and replication of the viral genome (Alansari and Potgieter, 1994; Valarcher and
Taylor, 2007), and a viral RNA-dependent polymerase protein (L) (Yunus et al.,
1998). In addition to the structural proteins, there are two non-structural proteins NS1
and NS2, which are important for counteracting the cellular interferon activity
(Bossert and Conzelmann, 2002; Bossert et al., 2003). Additionally, RSV has three
different matrix proteins; the non-glycosylated M protein is associated with the inner
surface of the viral envelope and is important for virion assembly (Collins et al., 2001;
Valarcher and Taylor, 2007).The M2 mRNA encodes for 2 proteins, M2-1 and M2-2
(Collins et al., 1990); M2-1 serves as an elongation factor and M2-2 has regulatory
functions in the transcription of the viral RNA (Hardy and Wertz, 1998; Bermingham
and Collins, 1999).
Virions are pleomorphic and can exhibit a spherical or filamentous shape. The
spherical type ranges in its size between 100 and 350 nm in diameter and the
filamentous type has a length up to 5 µm (Trudel et al., 1989).
1.2.6 Viral life cycle
Attachment of RSV is mediated by the surface glycoprotein G which binds to
glycosaminoglycans (Krusat and Streckert, 1997) as well as by F, which binds to
both glycosaminoglycans and a specific protein receptor (Feldman et al., 2000;
Techaarpornkul et al., 2002). The glycoprotein F plays a crucial role in cell entry by
inducing fusion of the viral envelope with the cellular membrane; recently nucleolin

Introduction 9
was identified as a putative receptor for HRSV (Tayyari et al., 2011). So far, no
receptors for BRSV have been identified. After fusion, the genome and the
nucleocapsids are released into the cytoplasm. Subsequently, the polymerase
transcribes the viral RNA which results in the generation of subgenomic RNAs. After
a sufficient amount of protein has been translated from the mRNAs, full length RNAs
are transcribed which are associated with the N, P and L proteins to form
ribonucleoproteins (RNPs). RNPs are enveloped by a budding process. The M
protein plays a crucial role in the assembly of the virions (Valarcher and Taylor,
2007). Budding occurs either directly at the host plasma membrane or at the
membrane of cytoplasmic vesicles (Arslanagic et al., 1996). In vitro studies showed
that HRSV buds at the apical surface of differentiated human respiratory epithelia
cells (Zhang et al., 2002), which was also observed for BRSV (Valarcher and Taylor,
2007).
1.3 Bovine parainfluenza virus type 3 (BPIV3)
Bovine parainfluenza virus type 3 was first isolated in 1959 in the United States
(Reisinger et al., 1959). It belongs to the genus Respirovirus within the subfamily
Paramyxovirinae and the family Paramyxoviridae (ICTV, 2012b). BPIV3 can be
divided into 2 genotypes (Chanock et al., 2001; Horwood et al., 2008). It is closely
related to human parainfluenza virus type 3 (HPIV3) (Bailly et al., 2000); additional
members of the genus are human parainfluenza virus type 1 (HPIV1) and Sendai
virus (SeV). Like RSV, PIV3 is an enveloped negative-stranded and non-segmented
RNA virus. The pleomorphic virion is 150 to 200 nm in size (Chanock et al., 2001).
The nucleocapsid is surrounded by a lipid envelope and consists of the viral genome,
the nucleoprotein (N), the phosphoprotein (P) and a large RNA polymerase (L). In
addition to the structural proteins, the genome encodes for the non-structural proteins
C, D and V, the functions of which are not well known but are assumed to inhibit IFN-
α and β activity (Durbin et al., 1999; Chanock et al., 2001). The matrix protein (M)
mediates the interaction of the nucleocapsid with the envelope and is responsible for
maturation of the virion and for budding (Chanock et al., 2001; Lamb and Kolakofsky,

Introduction 10
2001). PIV3 has two surface glycoproteins; the fusion protein F mediates cell entry
by inducing fusion of the viral envelope with the cellular membrane. This process is
pH-independent and requires the cooperation with the hemagglutinin-neuraminidase
(HN) (Heminway et al., 1994; Lamb and Kolakofsky, 2001; Takimoto et al., 2002).
Primary attachment occurs via HN which binds to (and cleaves) sialic acid-containing
cellular glycoconjugates on host cells (Scheid et al., 1972; Markwell and Paulson,
1980; Chanock et al., 2001). Replication occurs in the cytoplasm and budding at the
cytoplasmic membrane. It has been shown for HPIV3 that virus entry and release
occurs via the apical surface of polarized A549 cells (Bose et al., 2001) and of
human differentiated airway epithelia cells (Zhang et al., 2005). Although most
infections are mild or subclinical, it is considered to be the second most common
cause of severe respiratory tract infections in children and calves (Glezen and
Denny, 1973; Haanes et al., 1997). Transmission occurs via droplet infection. PIV3
predominantly infects the epithelial cells of the upper respiratory tract and only in
severe cases enters the lower airways (Chanock et al., 2001). Infection of the lower
respiratory tract may result in bronchitis, bronchiolitis or alveolitis. Microscopic lesions
include loss of ciliated cells, inclusion bodies and syncytia formation (Craighead and
Brennan, 1968; Bryson et al., 1983). As in the case of RSV, disease mainly occurs in
the winter months (Stott et al., 1980). Vaccines which are available on the market are
most often combination vaccines which also include other respiratory viruses or
bacteria (Salt et al., 2007; Ellis, 2010). There exist inactivated vaccines as well as
modified live vaccines which are still under improvement (Matsuoka et al., 1966; Salt
et al., 2007; Vangeel et al., 2009). Obstacles in vaccine design are similar but maybe
not as pronounced as those for RSV, e.g. inhibition due to maternal antibodies in
young animals (Vangeel et al., 2009) or boosting of the host immune response (Ellis,
2010).

Introduction 11
1.4 Bovine herpesvirus type 1 (BHV-1)
1.4.1 Epidemiology
Bovine herpesvirus was first isolated in 1955 (Madin et al., 1956). It is the etiological
agent of two major clinical cattle diseases: infectious bovine rhinotracheitis (IBR)
(Schroeder and Moys, 1954) as well as infectious pustular vulvovaginitis (IPV)/
balanoposthitis (IPB) (Kendrick et al., 1958; McKercher, 1964; van Oirschot, 1995).
Outbreaks can occur in the absence of acute infection and are caused by
maintenance of latent virus within a cattle herd and reactivation after specific stimuli
(Kaashoek et al., 1996; Ellis, 2009). Morbidity is high and can approach 100% in
unvaccinated herds; mortality rates range between 1% and 10% (Yates, 1982). In
recent years, eradication programs were initiated to eliminate BHV-1 from several
countries like Denmark, Switzerland and Austria (Ackermann and Engels, 2006).
Intervention strategies applied are mainly based on the detection and elimination of
latently infected animals within a herd.
1.4.2 Clinical signs and Pathology
Transmission occurs via aerosols at short distance or direct nose-to-nose contact in
case of respiratory infections (Mars et al., 1999; Mars et al., 2000); genital forms are
transmitted through mating and virus contaminated semen (Kupferschmied et al.,
1986). BHV-1 has an incubation period of 2-7 days (Yates, 1982). Clinical symptoms
are fever, anorexia, coughing, nasal discharge, conjunctivitis and dyspnea (Jericho
and Darcel, 1978; Jones and Chowdhury, 2007), but most infections remain
subclinical (Curtis et al., 1966; Hage et al., 1998). In fatal cases abortion and
systemic disease can occur, especially in newborn animals (Curtis et al. 1966, Bryan
1994, Muylkens 2007). The primary targets of BHV-1 are mucosal surfaces in the
upper airways (or genital tract) which results in massive cell destruction due to
programmed cell death and necrosis. Erosions and ulcers within the mouth, nose and
trachea are common (Curtis et al., 1966; Jericho and Darcel, 1978; Yates, 1982;
Muylkens et al., 2007). The cytopathic effect by BHV-1 causes epithelial damage
comprising cell ballooning and intranuclear inclusion bodies (Curtis et al., 1966;
Jericho and Darcel, 1978; Bryan et al., 1994). Respiratory disease lasts up to 10

Introduction 12
days (Curtis et al., 1966; Yates, 1982). Production of antibodies can be observed
between 8 and 12 days post-infection (Rouse and Babiuk 1978). After infection, virus
is released at high titers being responsible for the rapid transmission within a cattle
population (Yates, 1982). Once infected, BHV-1 causes lifelong latency (Stevens
1978). Latency is mainly established in ganglionic neurons, particularly the trigeminal
ganglion, but is assumed to take place also in tonsils and lymph nodes (Mweene et
al., 1996; Winkler et al., 1999; Winkler et al., 2000). Viremia is possible, therefore,
BHV-1 can also enter other organs and tissues and thus lead to atypical clinical
symptoms (Bryan et al., 1994; Kaashoek et al., 1998). BHV-1 plays a role in the
initiation of BRD as it can suppress the host immune system after infection. This
implies downregulation of IFN-α (Henderson et al., 2005), induction of apoptosis in
leukocytes, especially CD4+ T cells (Devireddy and Jones, 1999; Winkler et al., 1999)
as well as downregulation of MHC I molecules. Therefore, antigen presentation is
suppressed and the removal of virus-infected cells is hampered (Nataraj et al., 1997;
Koppers-Lalic et al., 2005). The impairment of the immune system in turn clears the
way for secondary bacterial infections which contribute to severe lower respiratory
tract infections (Jericho and Langford, 1978; Yates, 1982). Important bacterial
pathogens which are associated with BHV-1 are M. haemolytica or P. multocida
(Allan and Msolla, 1980; Ellis, 2009).
1.4.3 Vaccination
There exist several commercially available vaccines, either killed whole virus or
modified live attenuated vaccines (van Drunen Littel-van den Hurk, 2006), but they
can lead to disease in young calves due to immunosuppression and subsequent
induction of BRD (Jones and Chowdhury, 2007). Thus, so called vaccine outbreaks
have occurred in herds of vaccinated feedlot cattle (Bryan et al., 1994; van Drunen
Littel-van den Hurk et al., 2001). In recent years, many research efforts have been
directed to modified live virus vaccines and gE- and Us9- deletion mutants have been
suggested to be the most promising candidates (Kaashoek et al., 1998; Chowdhury
et al., 1999; Muylkens et al., 2006; Butchi et al., 2007).

Introduction 13
1.4.4 Taxonomy
BHV-1 belongs to the order Herpesvirales within the family Herpesviridae and the
subfamily Alphaherpesvirinae (ICTV, 2012a). Viruses belonging to the subfamily
Alphaherpesvirinae cause similar respiratory tract infections in a wide variety of
species. All members share a large host range, short replication cycles and the ability
to establish latent infections. Within the Alphaherpesvirinae, BHV-1 is classified in the
genus Varicellovirus. This genus consists of several viruses which are closely
related. Apart from BHV-1, the most prominent ones within this genus are Varicella
zoster virus (human herpesvirus 3), which causes chickenpox in humans and
pseudorabies virus (suid herpesvirus 1), which is responsible for Aujesky’s disease
(Roizman and Knipe, 2001; Roizman and Pellett, 2001).
On the basis of genomic and antigenetic surveys, three different subtypes of BHV-1
have been identified, BHV-1.1, BHV-1.2a and BHV-1.2b (Metzler et al., 1985).
Subtype 1.1 isolates were associated with IBR and found in the respiratory tract and
aborted fetuses. Subtype 1.2a can be associated with the respiratory and genital
tract and is considered to be the etiologic agent for IPV/IPB, IBR and abortions.
Subtype 1.2b is the causative agent of IPV/IPB, mild respiratory disease but has no
abortigenic potential (Miller et al., 1991; van Oirschot, 1995; D'Arce et al., 2002). Up
to now, the subtype tropism of BHV-1 for either the respiratory or the genital tract is
not supported by molecular (Muylkens et al., 2007) or experimental data (Steukers et
al., 2011). All subtypes exhibit more than 95% homology when compared by DNA
sequence analysis (Seal et al., 1985; Engels et al., 1986).
1.4.5 Virus structure
Herpesviruses are large (135 kb), linear double-stranded DNA viruses. The core is
surrounded by an icosahedral nucleocapsid which has a diameter of 100 to 110 nm
and is composed of 150 hexamers and 12 pentamers. The envelope consists of a
lipid bilayer with more than a dozen of glycoproteins (see Figure 1-3); among them
the glycoproteins gB, gC and gD are present in large amounts. Nucleocapsid and
envelope are connected by a protein-rich tegument. The size of a virus particle varies
between 120 and 300 nm in diameter (Roizman and Pellett, 2001). The genome of all

Introduction 14
members belonging to the genus Varicellovirus is divided into one long unique unit
(UL) and one short unique unit (US), which are flanked by repeat sequences. The
genome consists of a set of genes which are under temporal regulation; therefore,
one distinguishes between immediate early (IE), early (E) and late (L) genes, which
encode more than 70 proteins (Wirth et al., 1989). Herpesviruses are equipped with a
DNA polymerase that has proofreading activity (Ellis, 2009).
Figure 1-3 Schematic drawing of an Alphaherpesvirus particle; adapted from Flint et al. (2004).
1.4.6 Viral life cycle
Alphaherpesviruses like BHV-1, herpes simplex virus (HSV-1) or pseudorabies virus
utilize the glycoproteins gB and/or gC for primary attachment to heparan sulfate
structures (Liang et al., 1991; Okazaki et al., 1991). To mediate entry, the virions bind
via the surface glycoprotein gD to one or more of three known receptors (nectin-1,
herpesvirus entry mediator HVEM, 3-O sulfated heparan sulfate) (Johnson and
Spear, 1989; Montgomery et al., 1996; Geraghty et al., 1998; Shukla et al., 1999;
Campadelli-Fiume et al., 2000). Nectin-1 (also known as herpesvirus entry mediator

Introduction 15
C or poliovirus receptor-related-1) is a member of a family of cellular adhesion
molecules which are characterized by three extracellular immunoglobulin-like
domains (Ogita and Takai, 2006). It is ubiquitously expressed and can be found as a
protein of the adherens junctions (Rikitake et al., 2012). HVEM belongs to the tumor
necrosis factor family (Montgomery et al., 1996). Up to now, only nectin-1 was
identified as an entry receptor for BHV-1 (Geraghty et al., 1998). Due to the broad
tissue distribution of this receptor, BHV-1 infects many different cell types in the
respiratory tract as well as in other organs (Yates, 1982; Ellis, 2009). After receptor
binding, fusion is accomplished by interaction of the surface glycoproteins
gB/gD/gH/gL (Liang et al., 1995; Meyer et al., 1998; Gerdts et al., 2000; Pertel et al.,
2001). Once the fusion process has taken place, the capsids are transported to the
nucleus via the microtubuli. This process is up to know not well understood (Antinone
et al., 2006; Muylkens et al., 2007). After entry into the nucleus, the genome is
assumed to circularize (Garber et al., 1993). Within the nucleus, the transcription of
viral genes and the replication of the viral DNA takes place. Gene expression is
divided into three phases: immediate early (IE), early (E) and late (L) (Wirth et al.
1989). Immediate early gene expression starts after release of the genome from the
capsid and mRNAs for regulatory proteins are transcribed which initiate expression of
early genes by the host cell RNA polymerase. The expression of late genes is
delayed and starts after DNA replication to build structural proteins (Roizman and
Knipe, 2001; Schynts et al., 2001). Viral capsids are formed which contain the viral
DNA. They acquire a first envelope from the inner nuclear membrane and are
released from the nucleus into the cytoplasm by fusion with the outer nuclear
membrane. From the cytoplasm, they bud into a trans-Golgi compartment to be
finally released from the cell (Granzow et al., 2001; Muylkens et al., 2007).
To establish latency, herpesviruses enter sensory neurons via direct cell-to-cell
spread. Spread to neighboring cells is accomplished by the interaction of gD; deletion
of gD abolishes cell-to-cell spread (Fehler et al., 1992). Viral gene expression and
generation of infectious virus are arrested, so that only viral genomes are detectable
during this phase (Schang and Jones, 1997; Jones, 1998). Reactivation from latency
is induced by different stimuli like high levels of corticosteroids or suppression of the

Introduction 16
immune system; subsequently, gene expression in the neurons is reactivated and
secretion of infectious virus occurs from nasal or ocular regions (Sheffy and Davies,
1972; Rock et al., 1992; Jones, 2003; Jones et al., 2006b).
1.5 The respiratory epithelial barrier
1.5.1 Basic functions of the respiratory epithelium
The respiratory airways are common sites of infection (Greenberg, 2002; Sloots et
al., 2008), because they are in close contact with the environment and therefore a
target for airborne transmission of microorganisms (Vareille et al., 2011). The airway
epithelium which lines the respiratory tract represents the first barrier to infection. It is
composed of different cell types e.g. ciliated cells and goblet cells as well as basal
cells which serve as progenitor cells for the differentiated epithelial cells (Figure 1-4).
The goblet cells continuously secrete mucins which form the mucus barrier on the
surface of the respiratory epithelium. The ciliary activity of specialized cells transports
the mucus – and entrapped foreign material – out of the lung; therefore, ciliated cells
are an important part of the clearance system of an organism.
Figure 1-4 composition of the respiratory epithelium; adapted from Flint et al. (2004).

Introduction 17
The epithelial cells are connected to adjacent cells via cell-cell junctions. These
include tight junctions, adherens junctions, gap junctions and desmosomes (Roche et
al., 1993; Davies and Garrod, 1997). Cell-cell contacts form a mechanical barrier
which divides the epithelium into an apical, surface-orientated domain and a
basolateral domain which faces the internal environment. Due to the expression of
different proteins and enzymes on each domain, the epithelium obtains its polarity.
An ionic gradient is built across the epithelium which allows the directional transport
of molecules (Davies and Garrod, 1997). Especially the tight junctions are essential
to preserve the epithelial integrity (Roche et al., 1993).
1.5.2 Polarized virus entry
Cell-cell junctions serve as a barrier to viral entry into and egress from the respiratory
tissue. Viruses usually enter the epithelium through either the apical or basolateral
side. Apical entry may be hampered by the mucin-rich layer which covers the ciliated
epithelium throughout the respiratory tract. The mucus consists not only of mucins,
but also of antiviral substances like IFNs and nitric oxide being part of the innate
immune response (Nicholas et al., 2006; Rose and Voynow, 2006). It is estimated
that about 90% of particles which enter the respiratory tract are removed by the
mucociliary clearance system (Voynow and Rubin, 2009; Vareille et al., 2011). There
are several functions which help airway epithelial cells in their fight against invading
pathogens; for instance they express pattern recognition receptors like toll-like
receptors (TLRs) which recognize viruses and subsequently induce the onset of the
immune response (Diamond et al., 2000; Akira, 2009). Upon activation, they release
cytokines and chemokines in the submucosal environment to recruit immune cells to
the site of infection (McNamara et al., 2004; McNamara et al., 2005). Also the
basolateral membrane consists of several proteins which serve as interaction
partners for respiratory viruses (Bergelson, 2003; Bergelson, 2009). Therefore, the
question arises how these viruses can cross the epithelia barrier to successful infect
their host.
Many research efforts have been made in recent years to understand how viruses
can cross the epithelial barrier. Some viruses directly bind to the luminal surface. An

Introduction 18
example of this strategy are human and avian influenza viruses (Matrosovich et al.,
2004; Thompson et al., 2006) as well as HPIV3 (Zhang et al., 2005) or SeV
(Villenave et al., 2010). Also for HRSV, it was shown that it infects differentiated
human air-liquid interface cultures from the apical side (Zhang et al., 2002; Wright et
al., 2005; Villenave et al., 2012). Other viruses enter epithelial cells from the
basolateral surface. One example is measles virus (MV), which was found to
preferentially enter well-differentiated human airway epithelial cells from the
basolateral side (Sinn et al., 2002). Also human adenovirus predominantly infects
from the basolateral side (Lam et al., 2011; Lutschg et al., 2011) as well as vaccinia
virus (Vermeer et al., 2007).
1.5.3 Cultures for differentiated respiratory epithelial cells
As mentioned above, the primary target cells for many respiratory viruses are cells of
the respiratory epithelium. Differentiated respiratory epithelial cells are an interesting
tool to analyze virus infections since they resemble the in vivo situation more closely
than immortalized cell lines do. Two successful culture systems for differentiated
respiratory epithelial cells are the air-liquid interface system and precision-cut lung
slices. Respiratory epithelial cells can be cultured at the air-liquid interface to
generate a pseudostratified and differentiated epithelium, which closely resembles
the morphology of a respiratory epithelium in vivo. For ALI cultures, cells are seeded
on a filter membrane (Figure 1-5 A) and the medium initially covers the apical and
basolateral compartment to establish a confluent monolayer. Afterwards, the medium
in the upper chamber is removed and cells require about 3-4 weeks to grow in the
presence of retinoic acid to a multilayered and differentiated epithelium. Figure 1-5
shows characteristic features of such an epithelium like cilia development (B, upper
panel), multiple layers of nuclei (B, lower panel) and the development of tight
junctions (C). Integrity of epithelia can be evaluated by a stable transepithelial
electrical resistance (TEER). Fig. 1-5 D shows a typical time course of the TEER of
an ALI culture with three phases: 1) increase to a peak indicating polarization, 2)
phase of decrease to a lower value and (3) subsequent increase to a stable level.
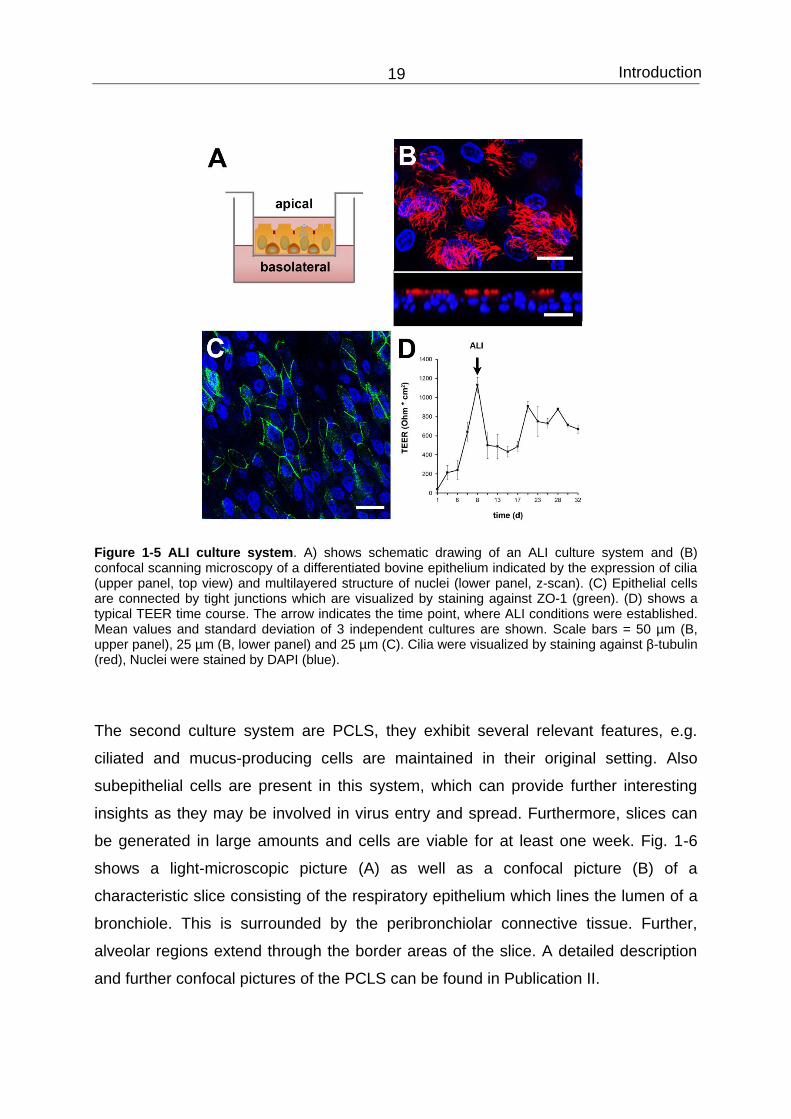
Introduction 19
Figure 1-5 ALI culture system. A) shows schematic drawing of an ALI culture system and (B) confocal scanning microscopy of a differentiated bovine epithelium indicated by the expression of cilia (upper panel, top view) and multilayered structure of nuclei (lower panel, z-scan). (C) Epithelial cells are connected by tight junctions which are visualized by staining against ZO-1 (green). (D) shows a typical TEER time course. The arrow indicates the time point, where ALI conditions were established. Mean values and standard deviation of 3 independent cultures are shown. Scale bars = 50 µm (B, upper panel), 25 µm (B, lower panel) and 25 µm (C). Cilia were visualized by staining against β-tubulin (red), Nuclei were stained by DAPI (blue).
The second culture system are PCLS, they exhibit several relevant features, e.g.
ciliated and mucus-producing cells are maintained in their original setting. Also
subepithelial cells are present in this system, which can provide further interesting
insights as they may be involved in virus entry and spread. Furthermore, slices can
be generated in large amounts and cells are viable for at least one week. Fig. 1-6
shows a light-microscopic picture (A) as well as a confocal picture (B) of a
characteristic slice consisting of the respiratory epithelium which lines the lumen of a
bronchiole. This is surrounded by the peribronchiolar connective tissue. Further,
alveolar regions extend through the border areas of the slice. A detailed description
and further confocal pictures of the PCLS can be found in Publication II.

Introduction 20
Figure 1-6 Light microscopic (A) and confocal (B) picture of a typical PCLS, scale bar = 100 µm.
1.6 Aim of the study
The aim of the thesis was to analyze strategies of respiratory viruses of the BRDC to
infect the bovine host. This problem was addressed with the help of ex vivo/ in vitro
respiratory epithelial cell cultures which exhibit in vivo characteristics of respiratory
epithelial cells more closely than immortalized cell lines do. These culture systems
have been established in our laboratory: The air-liquid interface system where
respiratory epithelial cells are grown on filter supports to establish a pseudostratified,
differentiated epithelium and precision-cut lung slices where slices are obtained from
the lobus accessorius after having stabilized the airways by low-melting point
agarose. In PCLS, the differentiated epithelial cells are maintained in their original
setting. Both culture systems showed that differentiated respiratory epithelial cells
were susceptible to infection by bovine parainfluenza virus type 3 (BPIV3) but not to
bovine respiratory syncytial virus (BRSV). These interesting observations were
subject to further detailed investigations within this thesis. In this context, another
important virus of the BRDC, bovine herpesvirus type 1 (BHV-1) was included into
the study. For BHV-1, it is not known which cells are the primary target cells within
the respiratory epithelium. Also for the human counterpart, herpes simplex virus type
1, it is not completely understood how the virus enters the respiratory airways and
which cells are the main target of infection. Relating to this question, it should be
determined whether infection by BHV-1 and BRSV is triggered by mechanical or

Introduction 21
chemical intervention of the epithelium. Further investigations should compare the
growth characteristics of the viruses. In the case of BRSV, further efforts addressed
the identification of primary target cells (Publication I).
Since it was observed that viruses of the BRDC can cross the species barrier, it is of
interest to analyze whether viruses can also infect in vitro cultures of non-bovine
origin. Since goats are considered as an important source of transmission, it was a
further aim of the thesis to establish PCLS of caprine lungs and to analyze them for
susceptibility to the three bovine viruses. Investigations like these are important to
understand interspecies transmission of viruses among ruminants (Publication II).

Introduction 22

Publication I 23
2 Publication I:
Three viruses of the bovine respiratory disease complex
apply different strategies to initiate infection
Jana Kirchhoff1, Sabine Uhlenbruck1, Katherina Goris1, Günther M. Keil2, Georg
Herrler1
1Institute of Virology, University of Veterinary Medicine Hannover, Hannover, Germany
2Institute of Molecular Biology, Friedrich-Loeffler-Institut, Greifswald - Insel Riems, Germany
Veterinary Research 2014, 45(1):20
DOI: 10.1186/1297-9716-45-20.
Authors’ contributions
JK and GH conceived and designed the experiments; JK, SU (technical assistance)
and KG performed the experiments; JK, SU, GK, KG and GH analyzed the data; GK
contributed reagents/materials/analysis tools; JK and GH wrote the paper. All authors
read and approved the final manuscript.
The extent of contribution from Jana Kirchhoff to this article:
A) Performance of experiments: 80%
B) Analysis of experiments: 90%
C) Writing of the paper: 80%

Publication I 24
Abstract
Bovine respiratory disease complex (BRDC) is the major cause of serious respiratory
tract infections in calves. The disease is multifactorial, with either stress or reduced
immunity allowing several pathogens to emerge. We investigated the susceptibility of
bovine airway epithelial cells (BAEC) to infection by the three major viruses
associated with the BRDC: bovine respiratory syncytial virus (BRSV), bovine
herpesvirus type 1 (BHV-1) and bovine parainfluenza virus type 3 (BPIV3). For this
purpose, two culture systems for well-differentiated BAEC were used: the air-liquid
interface (ALI) system, where filter-grown BAEC differentiate into a pseudostratified
respiratory epithelium and precision-cut lung slices (PCLS) where BAEC are
maintained in the original tissue organisation. Comparative infection studies
demonstrated that entry and release of BPIV3 occurred specifically via the apical
membrane with ciliated cells being the major target cells. By contrast, airway
epithelial cells were largely resistant to infection by BHV-1. When the epithelial
barrier was abolished by opening tight junctions or by injuring the cell monolayer,
BHV-1 infected mainly basal cells. Respiratory epithelial cells were also refractory to
infection by BRSV. However, this virus infected neither differentiated epithelial cells
nor basal cells when the integrity of the epithelial barrier was destroyed. In contrast to
cells of the airway epithelium, subepithelial cells were susceptible to infection by
BRSV. Altogether, these results indicate that the three viruses of the same disease
complex follow different strategies to interact with the airway epithelium. Possible
entry mechanisms are discussed.
Keywords
Bovine respiratory disease complex, bovine respiratory syncytial virus, bovine
herpesvirus type 1, bovine parainfluenza virus type 3, air-liquid interface, precision-
cut lung slices.

Publication II 25
3 Publication II:
Infection of differentiated airway epithelial cells from
caprine lungs by viruses of the bovine respiratory disease
complex
Jana Kirchhoff1, Sabine Uhlenbruck1, Günther M Keil2, Christel Schwegmann-
Wessels1, Martin Ganter3, Georg Herrler1
1Institute of Virology, University of Veterinary Medicine Hannover, Hannover, Germany
2Institute of Molecular Biology, Friedrich-Loeffler-Institut, Greifswald – Insel Riems, Germany
3Clinic for Swine and Small Ruminants, Forensic Medicine and Ambulatory Service, University of Veterinary Medicine, Hannover, Germany
Veterinary Microbiology 2014, 170:58-64
DOI: 10.1016/j.vetmic.2014.01.038.
Authors’ contributions
JK, MG and GH conceived and designed the experiments; JK and SU (technical
assistance) performed the experiments; JK, SU, GMK, CSW and GH analyzed the
data; GMK, CSW and MG contributed reagents/materials/analysis tools; JK and GH
wrote the paper. All authors read and approved the final manuscript.
The extent of contribution from Jana Kirchhoff to this article:
A) Performance of experiments: 80%
B) Analysis of experiments: 95%
C) Writing of the paper: 90%

Publication II 26
Abstract
Bovine respiratory syncytial virus (BRSV), bovine parainfluenza virus type 3 (BPIV3)
and bovine herpesvirus type 1 (BHV-1) are important pathogens associated with the
bovine respiratory disease complex (BRDC). Non-bovine ruminants such as goats
may also be infected and serve as a virus reservoir to be considered in the
development of control strategies. To evaluate the susceptibility of caprine airway
epithelial cells to infection by viruses of BRDC, we established a culture system for
differentiated caprine epithelial cells. For this purpose, we generated precision-cut
lung slices (PCLS), in which cells are retained in their original structural configuration
and remain viable for more than a week. The three bovine viruses were found to
preferentially infect different cell types. Ciliated epithelial cells were the major target
cells of BPIV3, whereas BHV-1 preferred basal cells. Cells infected by BRSV were
detected in submucosal cell layers. This spectrum of susceptible cells is the same as
that reported recently for infected bovine PCLS. While infection of caprine cells by
BRSV and BPIV3 was as efficient as that reported for bovine cells, infection of
caprine cells by BHV-1 required a tenfold higher dose of infectious virus as compared
to infection of bovine airway cells. These results support the notion that non-bovine
ruminants may serve as a reservoir for viruses of BRDC and introduce a culture
system to analyze virus infection of differentiated airway epithelial cells from the
caprine lung.
Keywords
Bovine respiratory disease complex, Goat respiratory disease, Bovine respiratory
syncytial virus, Bovine herpesvirus type 1, Bovine parainfluenza virus type 3,
Precision-cut lung slices

Discussion 27
4 Discussion
4.1 In vitro cultures for differentiated respiratory epithelial cells
Culture systems for primary bronchial epithelial cells were widely used within the last
decade to study replication strategies of respiratory viruses. When cultured under air-
liquid interface (ALI) conditions, cells undergo mucociliary differentiation to generate
a pseudostratified epithelium including ciliated cells, secretory cells and progenitor
cells; they closely resemble the protective epithelial barrier pathogens have to face
when they invade a host. Therefore, they mimic the in vivo situation better than
immortalized cell lines do; even cell lines like MDCK cells which grow as polarized
cells when seeded on filter membranes are not directly applicable to the natural
situation since they do not resemble the phenotype of differentiated epithelia.
Nevertheless, so far, protocols for culturing differentiated epithelial cells are often a
challenge and it is not trivial to transfer them from one species to another. Until now,
ALI cultures have been widely used to study entry strategies of human respiratory
viruses, e.g. it was found that human influenza viruses cause localized infection with
an initial preference for non-ciliated cells (Matrosovich et al., 2004; Ibricevic et al.,
2006). Human adenoviruses have been reported to preferentially enter the epithelium
from the basolateral side (Pickles et al., 1998; Lam et al., 2011) as it was also
observed for measles virus (Sinn et al., 2002). Moreover, several studies have been
performed on the entry and infectivity of HRSV (Zhang et al., 2002; Villenave et al.,
2011; Villenave et al., 2012) with different outcomes regarding the susceptibility of
cells (see chapter 4.4). In summary, studies on well-differentiated ALI cultures helped
to elucidate infection strategies of respiratory viruses in terms of entry pathways,
targeted cell types, tropism, receptor-expression, cytopathogenesis and immune
response of the epithelium. Furthermore, the application of this culture system to
cells from species other than human has been successful in the case of cattle (Goris
et al., 2009), swine (Lam et al., 2011; Bateman et al., 2013) and mouse (Ibricevic et
al., 2006); but in general the establishment of ALI cultures for other species is still a
challenging task. A big advantage of cultures grown under ALI conditions is the
formation of two compartments, so that viruses can be applied to the apical and/or

Discussion 28
basolateral side. Disadvantages are the long differentiation time and the cost-
intensive maintenance of the cultures. Morphological analyses showed that
differentiated human airway epithelial cultures closely resemble the airway epithelium
in vivo (Karp et al., 2002). Another study compared cells grown under ALI conditions
with cells derived from tracheal and bronchial brushings via microarray analysis and
concluded that the transcriptional profile was similar (Pezzulo et al., 2011).
Interestingly, it was also found in that study that cells isolated from tracheal and
bronchial regions were similar in their transcriptional profile. A more recent
development are ALI cultures which are co-cultured with e.g. dendritic cells or
macrophages. Such culture systems allow to address questions concerning the
influence of immune cells on viral entry or cytopathogenesis (Lutschg et al., 2011).
The involvement of immune cells on virus infection can also be studied with PCLS.
Slices of 200-300 µm thickness can be prepared from the lower respiratory tract. The
advantage of this culture system is that the differentiated epithelial cells are
maintained in their original setting and the above mentioned inclusion of other cell
types like immune cells and other cells of the peribronchiolar and alveolar regions.
Further interesting features are the high yield of slices which can be produced from
one lung, the inexpensive maintenance for more than a week, easy monitoring of the
ciliary activity and the possibility to induce bronchoconstriction. However, this culture
system is less convenient to study polarized entry of viruses since there is no direct
access to either side of the polarized cells. PCLS have been widely used to address
physiological and pharmacological questions (Martin et al., 2001; Vietmeier et al.,
2007; Henjakovic et al., 2008), whereas infection studies are only emerging (Ebsen
et al., 2002; Goris et al., 2009; Abd El Rahman et al., 2010; Punyadarsaniya et al.,
2011; Meng et al., 2013). Previously two different in vitro airway systems for well-
differentiated bovine epithelial cells have been established at our institute, ALI
cultures and PCLS (Goris et al., 2009), and infection studies with these culture
systems were extended during this thesis (Kirchhoff et al., 2014a). Furthermore, a
culture system for caprine bronchial epithelial cells was established to study the
influence of other ruminant species on virus transmission (Kirchhoff et al., 2014b).
The application of two culture systems, ALI cultures and PCLS, has the advantage

Discussion 29
that (i) results can be confirmed by different approaches to exclude artifacts and (ii)
results can complement each other by making use of the strength of each system.
4.2 Infection strategies of bovine parainfluenza virus type 3
4.2.1 Infection of well-differentiated bovine airway epithelial cells
When BPIV3 was applied to the apical surface of airway epithelial cells the virus was
able to efficiently infect the epithelium. Ciliated cells were observed to be the primary
target of infection as indicated by co-staining of β-tubulin. Within the ALI system, only
ciliated cells were infected, whereas other cell types, e.g. mucus-producing cells
were not targeted. The same result was obtained when virus was applied to PCLS.
Here, the viral antigen also colocalized with β-tubulin expression (Goris et al., 2009;
Kirchhoff et al., 2014a). Titration of the supernatants of BPIV3 infected ALI cultures
on MDBK cells revealed that BPIV3 replicated to high titers which peaked at 96 h pi.
and that infected BAEC remain highly productive during the course of the
experiment. Moreover, progeny virus was released exclusively from the apical
surface; no virus release into the basolateral compartment was observed. It is
therefore proposed that BPIV3 causes a localized infection with entry and release
being restricted to the apical side of the respiratory epithelium. These conclusions
are supported by the findings that no infection occurred when the inverted,
basolateral side of the filter membrane was exposed to the virus (data not shown).
Virus entry and release from the apical side of well-differentiated human cultures
have also been reported for the human counterpart of BPIV3, HPIV3 (Zhang et al.,
2005). These results are consistent with previous studies performed with HPIV1,
Sendai virus (SeV), HRSV as well as human and avian influenza viruses (Zhang et
al., 2002; Thompson et al., 2006; Bartlett et al., 2008; Villenave et al., 2010).
Cytopathic effects were not detected after one week; these observations go in line
with the finding that there was not decline in TEER. Therefore, infection by BPIV3
does not affect the integrity of the epithelium. The preference for the apical side may
be explained by the expression of receptors. It has been shown for influenza viruses
that sialic acids present on the apical surface of respiratory epithelia are crucial for

Discussion 30
entry into target cells (Matrosovich et al., 2004; Ibricevic et al., 2006; Thompson et
al., 2006). Glycoconjugates of ciliated cells have been reported to contain both, α2-6-
linked and α2-3-linked sialic acid residues whereas α2-6 linked sialic acids are
predominantly expressed on non-ciliated cells. Both types are well-known receptor
determinants for avian and human strains, respectively (Matrosovich et al., 2004;
Ibricevic et al., 2006; Thompson et al., 2006). Also for some coronaviruses, ciliated
cells are supposed to be the main entry port (Dijkman et al., 2013). Prior
neuraminidase treatment of PCLS abolished infection of BAEC by BPIV3; thus, sialic
acids present on the cell surface are also crucial for BPIV3 entry into bovine epithelial
cells. It has been reported that infection of human ALI cultures by HPIV3 is sensitive
to pretreatment with a neuraminidase that is specific for α2-6-linked sialic acid
residues (Zhang et al., 2005). By contrast, two studies that employed glycan array
analyses showed that HPIV3 preferred α2-3-linked sialic acids (Suzuki et al., 2001;
Amonsen et al., 2007). Therefore, further studies on well-differentiated airway
epithelial cells are necessary to specify the linkage type of the sialic acids required.
Up to now, studies were mostly performed with plant lectins, which bind to specific
linkage types of sialic acids. However, those plant lectins only recognize a small
subset of sialylated N-glycans. Comparative studies with influenza viruses have
shown that the spectrum of sialoglycans preferentially recognized by the viral
hemagglutinin may be quite different from that bound by plant lectins. Therefore, it
has been questioned whether plant lectins are a reliable marker for detection of
potential receptors for influenza viruses in general (Sauer et al., 2014). Because of
these considerations, plant lectins may also not provide the correct information about
the presence of receptors for BPIV3. Therefore, there is a need for more research
directed to the entry mechanism of BPIV3. For instance, the role of the
neuraminidase as well as the expression of sialic acids on bovine cultures has to be
analyzed in more detail in the future.
Apart from the susceptibility of ciliated cells of well-differentiated BAEC to BPIV3
infection, immortalized cell lines as well as non-polar BAEC were also found to be
readily infected by this virus. Interestingly, polarized BAEC, which were in a pre-
differentiating status, were refractory to infection (personal observations; J. Klein,

Discussion 31
Master thesis, University of Veterinary Medicine Hannover). Also Zhang et al. (2005)
observed a reduced susceptibility to infection during the differentiation phase and
concluded that there might be other, sialic acid-independent, pathways being
involved in the infection of non-polarized or immortalized cell lines. Studies on an
immortalized alveolar cell line (A549) showed that HPIV3 was able to bind to both,
sialic acid and heparan sulfate (HS) residues (Bose and Banerjee, 2002).
Concordantly, HS-moieties were found on the surface of non-polarized human airway
epithelial cells, but were restricted to basal cells when cultures were fully
differentiated (Zhang et al., 2005).
In PCLS, when BPIV3 infection was monitored over a period of one week, a strong
cytopathic effect (CPE) was observed as indicated by ciliated cells shedding into the
lumen of the bronchioli. As mentioned above, there was no visible BPIV3-mediated
CPE observed in infected ALI cultures after one week. Since it is suggested for
BPIV3 that viral clearance is mediated via an interaction of innate and adaptive
immunity (Ellis, 2010) which targets both, the virus and the infected cell, the lack of
cytopathology within the ALI cultures might be due to the lack of immune cells which
respond to the infection and therefore contribute to destruction of the epithelium. In
summary, an entry strategy is proposed where BPIV3 enters the epithelium via the
apical side by the use of sialic acid as receptor determinants and then replicates
within the ciliated cells. Spread of virus to adjacent cells is achieved by virus released
from the apical membranes and subsequent movement due to the ciliary activity of
BAEC.
4.2.2 Infection of well-differentiated caprine airway epithelial cells
As observed for the bovine cultures, also caprine PCLS were highly susceptible to
BPIV3. Co-staining of viral antigen and β-tubulin revealed that exclusively ciliated
cells were infected, which were shed into the lumen of the bronchioles 5 days after
infection. Occasionally, also infection of alveolar epithelial cells was observed,
consistent with the spread of virus to the lower airways in severe cases of BPIV3
infection. Therefore, it is assumed, that the course of infection by this virus is
comparable in the bovine and caprine lung. The pathogenic role of BPIV3 has been

Discussion 32
reported to be similar in cattle and sheep (Dungworth, 1993), but so far, there exist
only few studies about experimental infections of goats (Afshar and Terlecki, 1979).
In that study, BPIV3 was recovered from nasal, tracheal and bronchial regions up to
6 days after infection and the authors detected BPIV3-specific antibodies as well as
characteristic histological lesions. However, goats did not develop clinical signs.
Moreover, serological surveys based on indirect immunofluorescence assays
revealed antibodies against BPIV3 in goats (Fulton et al., 1982; Elazhary et al., 1984;
Trahman and Singh, 1990; Yesilbag and Gungor, 2009). Furthermore, Yener et al.
(2005) analyzed paraffin sections of pneumonic caprine lungs and detected BPIV3-
specific antigen in the majority of cases. Virus-positive staining was predominantly
found in the cytoplasm of bronchiolar luminal epithelial cells and type II pneumocytes.
Thus, it appears likely that BPIV3 plays a role in caprine pneumonia; however, since
data on BPIV3 infection of goats is rare, further in vivo and in vitro studies are
needed to clarify the role of goats in BPIV3 transmission.
4.3 Infection strategies of bovine herpesvirus type 1
4.3.1 Infection of well-differentiated bovine airway epithelial cells
When BHV-1 was applied to the apical side of ALI cultures, infection was detectable
only occasionally even when the viral titer was increased 100-fold compared to the
titer applied in the case of BPIV3. In contrast, when cell-cell contacts were disrupted
by mechanical injury or depletion of calcium, viral entry was strongly enhanced. Also,
when BHV-1 was applied to the basolateral surface of the filter-grown cells, the
epithelial cells were susceptible to infection. Thus, BHV-1 needs to get access to the
basal side to successfully invade the epithelium. Furthermore, infection studies with
the ALI cultures revealed that nonlumenal airway epithelial cells were infected by
BHV-1. Immunostaining showed that these cells were basal cells, which serve as
progenitor cells for the differentiated epithelial cells. Spread of infection occurred in a
plaquewise manner. These findings were confirmed with the PCLS, where also basal
cells were found to be the primary site of replication. Recently, Steukers et al. (2011)
established bovine organ cultures derived from the upper respiratory tract of cattle

Discussion 33
and found that BHV-1 can invade the mucosa across the basement membrane when
an infectious dose of 107 TCID50/mL was used. As stated above, occasional infection
from the apical side was also observed within this study when ALI cultures were
infected with a comparable titer. However, this titer is 100-fold higher compared to
that which was used to establish successful infection from the basal side, as well as
100-fold higher than the titer which is required for successful infection by BPIV3 from
the apical side. Numerous studies were performed to analyze the entry mechanism
of HSV-1. Studies with MDCK cells grown under air-liquid interface conditions
revealed similar entry strategies with HSV-1 being able to infect the polarized
epithelium after disrupting the tight junctions or injuring the cell monolayer
(Schelhaas et al., 2003; Marozin et al., 2004). However, other studies proposed
HSV-1 entry from both sides (Sears et al., 1991; Tran et al., 2000) or exclusively via
the apical membrane (Galen et al., 2006). Indeed, monolayer cultures do not
resemble the morphology of a differentiated epithelium and therefore are not suitable
to identify the cell type which is targeted by a virus. This can only be achieved with
cultures of well-differentiated cells. In this context two studies which were conducted
on primary keratinocyte cultures are of special interest: Here it was found that
successful infection by HSV-1 requires access to the basal side (Schelhaas et al.,
2003) and it was therefore concluded that viral replication is initiated in the basal cells
by passing through microabrasions within the uppermost layers (Visalli et al., 1997).
Thus, also for BHV-1 it appears likely that the virus enters through small lesions
within the epithelium (Tran et al., 2000; Yoon and Spear, 2002; Schelhaas et al.,
2003; Marozin et al., 2004), replicates within the basal cell layer and then either
reaches blood vessels via cell-to-cell spread to cause viremia or enters sensory
neurons to establish lifelong latency. This model is consistent with the cell-to-cell
spread observed within the PCLS when the incubation time was increased up to five
days. In general, it cannot be ruled out that alphaherpesvirus entry comprises
different routes and involves multiple receptors. The most prominent one is nectin-1,
a member of the immunoglobulin superfamily (Geraghty et al., 1998; Spear et al.,
2000), which colocalizes to adherent junctions of epithelial cells (Takai and
Nakanishi, 2003) but was also reported to be expressed on the apical side of the

Discussion 34
epithelium (Schelhaas et al., 2003; Marozin et al., 2004). However, since the
distribution of nectin-1 differs depending on the study and the cell system used,
further entry receptors like HVEM may play a role (Montgomery et al., 1996). So far,
only nectin-1 was shown to be important in the entry process of BHV-1 (Geraghty et
al., 1998). Due to the complex infection cycle of alphaherpesviruses in general, one
cannot exclude that studies on polarized and differentiated epithelia mimic different
steps in herpesvirus entry and spread, e.g. invasion through the mucosa upon
primary infection and/or infection from the basolateral side and subsequent shedding
after reactivation (Geraghty et al., 1998). Which receptors and entry pathways are
explicitly used by BHV-1 remains to be elucidated.
4.3.2 Infection of well-differentiated caprine airway epithelial cells
In comparison to BPIV3 and BRSV, BHV-1 infected caprine PCLS very inefficiently.
When the virus was applied to the caprine slices with the same infectious dose as in
the bovine system, there was no infection visible. Only when the viral titer was
increased tenfold, infection occurred; basal cells were the primary site of infection as
it was also observed in the bovine system. Moreover, the disruption of tight junctions
facilitates viral entry. The group of alphaherpesviruses consists of different ruminant
viruses which are closely related. Therefore, interspecies transmission is considered
to be likely and was observed for almost all of them (Thiry et al., 2006). Accordingly,
it has been concluded that also non-bovine ruminant species have to be considered
as a reservoir for BHV-1 (Engels and Ackermann, 1996). Natural BHV-1 infections
have been recognized in goats, sheep and swine as indicated by virus isolation and
serological surveys (Fulton et al., 1982; Elazhary et al., 1984; Engels et al., 1992;
Clark et al., 1993). Especially BHV-1 and caprine HV-1 (CapHV-1) exhibit a close
relationship (Nixon et al., 1988; Whetstone and Evermann, 1988) and experimental
infections revealed that cross-infection is possible with higher susceptibility of goats
for BHV-1 than calves for CapHV-1 (Engels and Ackermann, 1996; Six et al., 2001).
In comparison, one study (Hage et al., 1997) analysed BHV-1 transmission from
experimentally infected sheep and concluded that sheep do not play a major role in
viral spread. For goats, so far such studies have not been conducted, but will be an
important step in the analysis of virus transmission. The fact that BHV-1 is less

Discussion 35
infectious for goats when compared to BPIV3 and BRSV might be due to the fact that
there exists also a species-specific component. Alphaherpesviruses contain several
different surface glycoproteins; some of them are closely related but others are very
distinct within members of the same genus. One example is glycoprotein C, which is
involved in the primary attachment to the host cell and considered to be the most
host-specific surface glycoprotein (Ackermann et al., 1986; Engels et al., 1992).
4.4 Infection strategies of bovine respiratory syncytial virus
4.4.1 Infection of well-differentiated bovine airway epithelial cells
When BRSV was applied to the apical surface of ALI cultures, infection was
predominantly found in ciliated cells and only occasionally in non-ciliated cells. In
comparison to BPIV3, the infection level was low since GFP-expression was
detected in only a few, non-contiguous cells when equal amounts of virus were
applied. Concordantly, titers of infectious progeny virus were up to 600 fold lower
than those observed for BPIV3. Moreover, progeny virus was released exclusively
via the apical membrane; therefore, results from ALI cultures indicate that also BRSV
causes localized infections in the respiratory tract. No syncytia formation was
observed and no cytopathogenic effect. The infection pattern resembles that reported
for HRSV by Zhang et al. (2002) where HRSV was applied to well-differentiated
human airway epithelial cells and basically the same mechanism of infection was
observed; but infection levels were high in comparison to the titers which were
observed in this thesis. However, these outcomes result from an unusual high viral
titer which was applied to the human ALI cultures. Contrary to that study and in
accordance with results obtained in this thesis, Wright et al. (2005) merely observed
low replication of HRSV in the airway epithelium of human nasopharyngeal mucosa,
and in studies by Villenave et al. (2012), RSV infection of pediatric bronchial
epithelial cells was only present in a subset of ciliated cells. Therefore, the authors
concluded that the entry receptor is not expressed on all ciliated cells and needs to
be identified in the future. Since disruption of tight junctions as well as wounding of
the epithelium did not enhance infection, it appears likely that BRSV applies other

Discussion 36
entry strategies than herpesviruses do. One explanation might be the upregulation of
a potential receptor molecule. In this context, environmental stressors may have an
influence which renders the epithelium susceptible to infection. Since severe BRSV
infections predominantly occur in young cattle (Kimman et al., 1988; Hagglund et al.,
2006), stressful events within the first month of live like weaning, transport, or
introduction into new herds can make the animals susceptible. Also co-infection with
other viruses or bacteria of the BRDC may predispose them to disease. Bacteria like
P. multocida, M. haemolytica, M. bovis and H. somni may contribute to severe
respiratory tract infections (Virtala et al., 1996; Dabo et al., 2007; Booker et al.,
2008). Gershwin et al. (2005) showed that experimental co-infection of calves with H.
somni and BRSV results in a more severe disease as compared to infection with
either of the pathogens alone. In this study it has been suggested that viral infection
causes immunosuppression of the host which leads to increased colonization of
commensal bacteria; however an inverse order of events cannot be excluded:
primary attachment of bacteria may also facilitate subsequent viral entry into the
respiratory tract. This model was also proposed by Nguyen et al. (2010) who showed
that HRSV infection was enhanced in primary human epithelial cells after treatment
with a synthetic bacterial lipopeptide. So far, such experimental co-infection studies
are rare and should be performed in the future both in vivo and in vitro. Especially
well-differentiated respiratory epithelial cell cultures are a useful tool to address such
questions. Work on BRSV infection is still hampered by the fact that the entry
receptor is not known to date. Recently, the core protein nucleolin was proposed as a
receptor for HRSV (Tayyari et al., 2011). Nucleolin has the capacity to shuttle
between the nucleus and the surface especially in cells which undergo mitosis
(Ginisty et al., 1999). A two-step process of entry has been proposed with initial
attachment to cellular proteoglycans via the G protein and subsequent fusion of the
viral envelope with the cellular membrane mediated by a specific receptor, e.g.
nucleolin (Mastrangelo and Hegele, 2013). Since nucleolin is ubiquitously expressed,
it cannot explain the tropism of RSV for the respiratory tract; therefore, it appears
likely that other co-receptors are involved in the entry of RSV. Identification of these

Discussion 37
co-receptors will be helpful to understand the limited susceptibility of bovine airway
epithelial cells to BRSV.
When BRSV was applied to PCLS, infection was observed in subepithelial cells and
only occasionally within the epithelium. Unfortunately, antibodies for the bovine
system are limited and those who are available displayed a high background. Thus,
so far it was not possible to identify the type of infected cells within this region.
Nevertheless, the morphological appearance suggests that at least some of them
might be dendritic cells (DCs). Staining with an antibody against CD86, which was
shown to be expressed on bovine dendritic cells (Yamakawa et al., 2008) indicated
some colocalization with BRSV-infected cells (data not shown). Since there exist
more than one subtype of dendritic cells in the bovine lung, more immune markers
have to be included in the analysis to truly identify the targeted cell type. DCs are
antigen-presenting cells which take up antigen and migrate to lymph nodes to induce
a specific immune response (Banchereau and Steinman, 1998); however viruses
may also use them as a “Trojan horse” to migrate to other regions of the body
(Yewdell and Brooke, 2012), either to establish persistence or to cause viremia. For
bovine DCs, it has indeed been shown that they can be infected by BRSV (Werling et
al., 2002) as well as it has been reported for human DCs and HRSV (Jones et al.,
2006a; Hobson and Everard, 2008; Everard et al., 2009). Involvement of dendritic
cells in the uptake of viruses was proposed for other viruses as well, e.g. measles
virus (Sinn et al., 2002). As this virus was found to preferentially infect well-
differentiated respiratory epithelial cells from the basolateral side, it has been
suggested that immune cells like macrophages or DCs may take up the virus from
the lumen and transport it to lymph nodes or the bloodstream (Sinn et al., 2002;
Mesman et al., 2012). Also for HIV-1, it has been reported that DCs are involved in
viral entry and spread (Geijtenbeek et al., 2000). Therefore, the possibility has to be
considered that also RSV exploits immune cells like DCs to overcome the epithelial
barrier.
When the incubation time of BRSV was increased to one week, spread of infected
cells into the alveolar epithelium was observed. Staining with a marker against TTF-1

Discussion 38
showed that type II pneumocytes were targeted. These cells are cuboidal in shape
and are typically located at the intersections of two or more alveoli. Thyroid-
transcription factor (TTF)-1 has regulatory functions in the transcription of genes in
the thyroid and lung, e.g. expression of surfactant gene and Clara cell secretory
protein. Originally, it was used as a marker for lung adenocarcinoma (Lau et al.,
2002). A positive staining for TTF-1 has been reported for type II pneumocytes and
Clara cells; e.g. in the lungs of sheep infected with sheep pox virus (Beytut, 2010) or
in a study where pigs were experimentally infected with porcine reproductive and
respiratory syndrome virus (PRRSV) (Balka et al., 2013). Moreover, also TTF-1-
negative cells within the alveolar epithelium were infected. These cells had a
flattened morphology being characteristic for type I pneumocytes. Thus, it is
concluded that BRSV can infect both, type I and type II pneumocytes. Replication of
BRSV in type I and type II pneumocytes was found in histological sections of calves
experimentally infected with BRSV (Viuff et al., 2002). Moreover, this group showed
that severe clinical outcomes were correlated to spread of BRSV to deeper regions of
the lung, e.g. the alveoli, and concluded that both, the host immune response and
deficient clearance of the virus with a subsequent spread to alveoli can lead to
severe BRSV infection. Infection of alveolar epithelial cells within PCLS is a striking
feature and may help to understand the strategies of BRSV to establish infection in
the upper and lower respiratory tract as well as to identify possible entry pathways in
the future.
4.4.2 Infection of well-differentiated caprine airway epithelial cells
When BRSV was applied to the caprine PCLS, subepithelial cells were found to be
the primary target of infection as described above for the bovine system. When the
titer was increased tenfold, only occasionally epithelial cells became infected and
were identified as ciliated cells by staining with a marker against β-tubulin. This
refractory behaviour against BRSV infection could not be overcome by increasing the
incubation time to 5 days; under these conditions, cells rather spread to the alveolar
regions of the slice. Within the alveolar regions both, cells with a cuboidal and those
with a flattened morphology were identified as being infected; thus it is concluded
that, like in the bovine system, type I and type II pneumocytes were infected. To

Discussion 39
confirm these observations, antibody staining with the marker of TTF-1 would be the
next step. Unfortunately, this marker was obtained at the very end of the thesis,
where infection studies on caprine slices had already been finished. Thus, these
staining procedures could not be included in the thesis but will be done afterwards.
However, it appears likely that BRSV infects the same cell types in the bovine and in
the caprine system, therefore, cross- infection of goats may be a characteristic
feature of BRSV.
Antibodies to BRSV were found in goats and to a lesser extent in sheep (Elazhary et
al., 1984; Van der Poel et al., 1995; Langedijk et al., 1996; Yesilbag and Gungor,
2009). Van der Poel et al. (1995) performed an RSV-specific ELISA with sera from
different animal species and humans which all had prior contact to cattle. In this
study, it has been observed that more than 50% of the sera were BRSV-positive with
the strongest impact being directed to goats. In contrast, samples of human and
sheep were not positive. Antibody prevalence against BRSV was also found in
serological examinations of goats in North-Western Turkey (Yesilbag and Gungor,
2009) with values similar to those observed for cattle; thus, BRSV is considered to be
the most common cause of respiratory infections within this region. Prevalence of
BRSV antibodies in goats was also reported in studies from Canada (Elazhary et al.,
1984) and USA (Fulton et al., 1982).
Based on immunological and nucleotide sequence data on the G-protein, the most
variable protein among RSV isolates, it has been suggested that there exist at least
two ruminant RSV subgroups, one is represented by strains isolated from cattle and
goat and the other one by RSV isolated from sheep (Duncan and Potgieter, 1993;
Alansari et al., 1999; Eleraky et al., 2003). Experimental infection of sheep resulted in
mild clinical signs and BRSV could be isolated from the lungs of all infected animals.
Accordingly, antigen was mainly detected in the bronchiolar and alveolar epithelium,
but the overall amount of BRSV-positive cells was low (Meehan et al., 1994).
Interestingly, this group infected ovine lung slice organ cultures with BRSV at a titer
of 106 CCID50/mL and could not find antigen in any of the lung slices even after 72 h.
In comparison, slices were positive for BPIV3 when the virus was applied at similar

Discussion 40
titers, and infection was observed within the bronchiolar epithelium and occasionally
within the alveolar regions. Another group (Lehmkuhl and Cutlip, 1979) infected
lambs and also detected mild clinical symptoms, necrotic lesions and
seroconversion. Moreover, virus was recovered from 2 of 5 animals. Unfortunately,
information about experimental infection of goats is rare (Fulton et al., 1982).
It has been suggested that BRSV can survive in cattle populations since outbreaks
occur in the absence of initial infection or detection of clinical signs. So far, it is not
known, if BRSV can somehow persist in its host, although there is some evidence
that the virus can persist in e.g. lymph nodes (Van der Poel et al., 1993; De Jong et
al., 1996; Valarcher et al., 2001). Therefore, it is also possible that BRSV is
transmitted from other sources, e.g. goats and reintroduced into cattle populations.
4.5 Summary and Outlook
4.5.1 Strategies of viruses of the BRDC to infect the bovine host
Viruses of the BRDC are causative agents of respiratory disease in cattle and
especially young calves. In this thesis, the entry strategies of three bovine viruses
associated with the BRDC - BPIV3, BRSV and BHV-1 - were analyzed. In summary,
it can be concluded that the viruses display different strategies to infect their bovine
host: BPIV3 enters the cell via sialic acid containing glycoconjugates located on the
apical side of the respiratory epithelium, replicates to high titers in the ciliated
epithelial cells and progeny virus is shed into the lumen of the airways being
characteristic for a localized infection. Spread of BPIV3 to adjacent cells is
accomplished via the ciliary activity. BHV-1 is supposed to enter through small
lesions into the epithelium and primarily replicates within the basal cells, the
progenitor cells of the respiratory epithelium. Then the virus can either be released
from the apical side to be shed into the environment during acute infection or
establish a persistent infection in neurons. BRSV is likely to enter the epithelium from
the apical surface via a putative receptor molecule which is enhanced after specific
stimuli or may be taken up from immune cells like dendritic cells to cross the

Discussion 41
epithelial barrier. The potential of BRSV to somehow persist in its host remains
speculative. Since BRDC is a multifactorial disease, future research should include
co-infection studies with other viruses or bacteria to analyze, whether the
susceptibility of cells to BRSV can be enhanced under these conditions. Also, the
finding that pneumocytes are readily infected by BRSV may help to understand the
infection strategy of BRSV in the future.
Since respiratory viruses are primarily transmitted through the upper airways
including mouth, pharynx and trachea, it would be useful to establish well-
differentiated epithelial cell cultures from the uppermost airways like the nose to
clarify whether the replication of the viruses, especially BRSV, is enhanced in those
regions of the respiratory tract.
Taken together, analyses of the entry strategies of viruses of the BRDC are important
to understand the pathogenicity of these viruses and will help to create new
intervention strategies. Moreover, such studies will also help to design new or
improve existing vaccines in the future.
4.5.2 Goats as a virus reservoir
Pneumonia is considered to be an important disease in small ruminant species like
sheep and goats (Ackermann and Brogden, 2000; Yener et al., 2005) and it has been
suggested that viruses of the BRDC are causative agents in the naturally occurring
caprine pneumonia (Van der Poel et al., 1995; Thiry et al., 2006). Therefore, studies
as they were conducted in this thesis are important to analyze the role of goats as a
reservoir of viruses of the BRDC. Thus, one aim of the thesis was to establish
caprine PCLS and to infect these with viruses of the BRDC. In summary, it was found
that BPIV3 efficiently infected the epithelial cells while BRSV was mostly found in
submucosal cell layers. The infection efficiency of these viruses was similar to that
seen in bovine PCLS. In contrast, BHV-1 was much less efficient in infecting caprine
cells, but displayed the same tropism to basal cells as seen in bovine samples. The
thesis describes for the first time the preparation of caprine PCLS and demonstrates
that these slices can represent a first step in investigating pathogen dynamics among
co-housed closely related species. It can be concluded that in vitro studies are a

Discussion 42
useful ex vivo tool to investigate such interactions. A next consequent step is to carry
out infection studies of goats in vivo to close gaps in terms of infectivity, antibody-
production, clinical outcomes and shedding of viruses. Furthermore, transmission
studies have to be performed to analyze whether virus can be transmitted to
seronegative cattle. Also for small ruminant species like goats, it has been suggested
that viruses of the BRDC can increase the susceptibility to secondary bacterial
infections with e.g. M. haemolytica. Therefore, also in this context it would be
interesting to perform co-infections in order to see if they enhance the susceptibility
of caprine PCLS to BRSV. Studies like these are important to understand the role of
goats in the BRDC and are an essential step for vaccine design.

References 43
5 References
Abd El Rahman S., Winter C., El-Kenawy A., Neumann U. and Herrler G. (2010): Differential sensitivity of well-differentiated avian respiratory epithelial cells to infection by different strains of infectious bronchitis virus. J Virol 84(17): 8949-8952.
Ackermann M. and Engels M. (2006): Pro and contra IBR-eradication. Vet Microbiol 113(3-4): 293-302.
Ackermann M., Metzler A. E., Mc Donagh H., Bruckner L., Müller H. K. and Kihm U. (1986): Stellen nichtbovine Paarhufer ein IBR-Virus-Reservoir da? I. BHV-1 and CapHV-1-Infektions- und Reaktivierungsversuche an Ziegen, Virustyp-Spezifizität der humoralen Antikörper und Charakterisierung der viralen Antigene. Schweiz Arch Tierheilkd 128: 557-573.
Ackermann M. R. and Brogden K. A. (2000): Response of the ruminant respiratory tract to Mannheimia (Pasteurella) haemolytica. Microbes Infect 2(9): 1079-1088.
Afshar A. and Terlecki S. (1979): Experimental infection of goats with parainfluenza virus type 3. Zentralbl Veterinarmed B 26(8): 641-651.
Akira S. (2009): Pathogen recognition by innate immunity and its signaling. Proc Jpn Acad Ser B Phys Biol Sci 85(4): 143-156.
Alansari H., Duncan R. B., Baker J. C. and Potgieter L. N. (1999): Analysis of ruminant respiratory syncytial virus isolates by RNAse protection of the G glycoprotein transcripts. J Vet Diagn Invest 11(3): 215-220.
Alansari H. and Potgieter L. N. (1994): Molecular cloning and sequence analysis of the phosphoprotein, nucleocapsid protein, matrix protein and 22K (M2) protein of the ovine respiratory syncytial virus. J Gen Virol 75 ( Pt 12): 3597-3601.
Allan E. M. and Msolla P. M. (1980): Scanning electron microscopy of the tracheal epithelium of calves inoculated with bovine herpesvirus 1. Res Vet Sci 29(3): 325-327.
Allen J. W., Viel L., Bateman K. G., Rosendal S., Shewen P. E. and Physick-Sheard P. (1991): The microbial flora of the respiratory tract in feedlot calves: associations between nasopharyngeal and bronchoalveolar lavage cultures. Can J Vet Res 55(4): 341-346.
Amonsen M., Smith D. F., Cummings R. D. and Air G. M. (2007): Human parainfluenza viruses hPIV1 and hPIV3 bind oligosaccharides with alpha2-3-linked sialic acids that are distinct from those bound by H5 avian influenza virus hemagglutinin. J Virol 81(15): 8341-8345.
Anderson L. J., Hierholzer J. C., Tsou C., Hendry R. M., Fernie B. F., Stone Y. and McIntosh K. (1985): Antigenic characterization of respiratory syncytial virus strains with monoclonal antibodies. J Infect Dis 151(4): 626-633.
Antinone S. E., Shubeita G. T., Coller K. E., Lee J. I., Haverlock-Moyns S., Gross S. P. and Smith G. A. (2006): The Herpesvirus capsid surface protein, VP26, and the majority of the tegument proteins are dispensable for capsid transport toward the nucleus. J Virol 80(11): 5494-5498.
Antonis A. F., de Jong M. C., van der Poel W. H., van der Most R. G., Stockhofe-Zurwieden N., Kimman T. and Schrijver R. S. (2010): Age-dependent differences in the pathogenesis of bovine respiratory syncytial virus infections related to the development of natural immunocompetence. J Gen Virol 91(Pt 10): 2497-2506.

References 44
Antonis A. F., Schrijver R. S., Daus F., Steverink P. J., Stockhofe N., Hensen E. J., Langedijk J. P. and van der Most R. G. (2003): Vaccine-induced immunopathology during bovine respiratory syncytial virus infection: exploring the parameters of pathogenesis. J Virol 77(22): 12067-12073.
Arslanagic E., Matsumoto M., Suzuki K., Nerome K., Tsutsumi H. and Hung T. (1996): Maturation of respiratory syncytial virus within HEp-2 cell cytoplasm. Acta Virol 40(4): 209-214.
Bailly J. E., McAuliffe J. M., Skiadopoulos M. H., Collins P. L. and Murphy B. R. (2000): Sequence determination and molecular analysis of two strains of bovine parainfluenza virus type 3 that are attenuated for primates. Virus Genes 20(2): 173-182.
Baker J. C., Werdin R. E., Ames T. R., Markham R. J. and Larson V. L. (1986): Study on the etiologic role of bovine respiratory syncytial virus in pneumonia of dairy calves. J Am Vet Med Assoc 189(1): 66-70.
Balka G., Ladinig A., Ritzmann M., Saalmuller A., Gerner W., Kaser T., Jakab C., Rusvai M. and Weissenbock H. (2013): Immunohistochemical characterization of type II pneumocyte proliferation after challenge with type i porcine reproductive and respiratory syndrome virus. J Comp Pathol 149(2-3): 322-330.
Banchereau J. and Steinman R. M. (1998): Dendritic cells and the control of immunity. Nature 392(6673): 245-252.
Barbour E. K., Nabbut N. H., Hamadeh S. K. and Al-Nakhli H. M. (1997): Bacterial identity and characteristics in healthy and unhealthy respiratory tracts of sheep and calves. Vet Res Commun 21(6): 421-430.
Bartlett E. J., Hennessey M., Skiadopoulos M. H., Schmidt A. C., Collins P. L., Murphy B. R. and Pickles R. J. (2008): Role of interferon in the replication of human parainfluenza virus type 1 wild type and mutant viruses in human ciliated airway epithelium. J Virol 82(16): 8059-8070.
Bateman A. C., Karasin A. I. and Olsen C. W. (2013): Differentiated swine airway epithelial cell cultures for the investigation of influenza A virus infection and replication. Influenza Other Respir Viruses 7(2): 139-150.
Bergelson J. M. (2003): Virus interactions with mucosal surfaces: alternative receptors, alternative pathways. Curr Opin Microbiol 6(4): 386-391.
Bergelson J. M. (2009): Intercellular junctional proteins as receptors and barriers to virus infection and spread. Cell Host Microbe 5(6): 517-521.
Bermingham A. and Collins P. L. (1999): The M2-2 protein of human respiratory syncytial virus is a regulatory factor involved in the balance between RNA replication and transcription. Proc Natl Acad Sci U S A 96(20): 11259-11264.
Beytut E. (2010): Sheep pox virus induces proliferation of type II pneumocytes in the lungs. J Comp Pathol 143(2-3): 132-141.
Bidokhti M. R., Traven M., Fall N., Emanuelson U. and Alenius S. (2009): Reduced likelihood of bovine coronavirus and bovine respiratory syncytial virus infection on organic compared to conventional dairy farms. Vet J 182(3): 436-440.
Booker C. W., Abutarbush S. M., Morley P. S., Jim G. K., Pittman T. J., Schunicht O. C., Perrett T., Wildman B. K., Fenton R. K., Guichon P. T. and Janzen E. D. (2008): Microbiological and histopathological findings in cases of fatal bovine respiratory disease of feedlot cattle in Western Canada. Can Vet J 49(5): 473-481.

References 45
Booker C. W., Guichon P. T., Jim G. K., Schunicht O. C., Harland R. J. and Morley P. S. (1999): Seroepidemiology of undifferentiated fever in feedlot calves in western Canada. Can Vet J 40(1): 40-48.
Bose S. and Banerjee A. K. (2002): Role of heparan sulfate in human parainfluenza virus type 3 infection. Virology 298(1): 73-83.
Bose S., Malur A. and Banerjee A. K. (2001): Polarity of human parainfluenza virus type 3 infection in polarized human lung epithelial A549 cells: role of microfilament and microtubule. J Virol 75(4): 1984-1989.
Bossert B. and Conzelmann K. K. (2002): Respiratory syncytial virus (RSV) nonstructural (NS) proteins as host range determinants: a chimeric bovine RSV with NS genes from human RSV is attenuated in interferon-competent bovine cells. J Virol 76(9): 4287-4293.
Bossert B., Marozin S. and Conzelmann K. K. (2003): Nonstructural proteins NS1 and NS2 of bovine respiratory syncytial virus block activation of interferon regulatory factor 3. J Virol 77(16): 8661-8668.
Brodersen B. W. (2010): Bovine respiratory syncytial virus. Vet Clin North Am Food Anim Pract 26(2): 323-333.
Brogden K. A., Lehmkuhl H. D. and Cutlip R. C. (1998): Pasteurella haemolytica complicated respiratory infections in sheep and goats. Vet Res 29(3-4): 233-254.
Bryan L. A., Fenton R. A., Misra V. and Haines D. M. (1994): Fatal, generalized bovine herpesvirus type-1 infection associated with a modified-live infectious bovine rhinotracheitis parainfluenza-3 vaccine administered to neonatal calves. Can Vet J 35(4): 223-228.
Bryson D. G., McNulty M. S., McCracken R. M. and Cush P. F. (1983): Ultrastructural features of experimental parainfluenza type 3 virus pneumonia in calves. J Comp Pathol 93(3): 397-414.
Bueno S. M., Gonzalez P. A., Cautivo K. M., Mora J. E., Leiva E. D., Tobar H. E., Fennelly G. J., Eugenin E. A., Jacobs W. R., Jr., Riedel C. A. and Kalergis A. M. (2008): Protective T cell immunity against respiratory syncytial virus is efficiently induced by recombinant BCG. Proc Natl Acad Sci U S A 105(52): 20822-20827.
Butchi N. B., Jones C., Perez S., Doster A. and Chowdhury S. I. (2007): Envelope protein Us9 is required for the anterograde transport of bovine herpesvirus type 1 from trigeminal ganglia to nose and eye upon reactivation. J Neurovirol 13(4): 384-388.
Campadelli-Fiume G., Cocchi F., Menotti L. and Lopez M. (2000): The novel receptors that mediate the entry of herpes simplex viruses and animal alphaherpesviruses into cells. Rev Med Virol 10(5): 305-319.
Chanock R. M., Murphy B. R. and Collins P. L. (2001). Parainfluenza viruses. Fields Virology. D. M. Knipe and P. Howley. Philadelphia, PA, Lippincott/ The Williams & Wilkins Co. 1: 1341-1380.
Chowdhury S. I., Ross C. S., Lee B. J., Hall V. and Chu H. J. (1999): Construction and characterization of a glycoprotein E gene-deleted bovine herpesvirus type 1 recombinant. Am J Vet Res 60(2): 227-232.
Clark R. K., Whetstone C. A., Castro A. E., Jorgensen M. M., Jensen J. F. and Jessup D. A. (1993): Restriction endonuclease analysis of herpesvirus isolated from two peninsular bighorn sheep (Ovis Canadensis Cremnobates). J. Wildlife Disease 29: 50-56.

References 46
Collins J. K., Teegarden R. M., MacVean D. W., Salman, Smith G. H. and Frank G. R. (1988): Prevalence and specificity of antibodies to bovine respiratory syncytial virus in sera from feedlot and range cattle. Am J Vet Res 49(8): 1316-1319.
Collins P. L., Chanock R. M. and Murphy B. R. (2001). Respiratory syncytial virus. Fields Virology. D. M. Knipe and P. Howley. Philadelphia, PA, Lippincott/ The Williams & Wilkins Co. 1.
Collins P. L., Hill M. G. and Johnson P. R. (1990): The two open reading frames of the 22K mRNA of human respiratory syncytial virus: sequence comparison of antigenic subgroups A and B and expression in vitro. J Gen Virol 71 (12): 3015-3020.
Confer A. W. (2009): Update on bacterial pathogenesis in BRD. Anim Health Res Rev 10(2): 145-148.
Craighead J. E. and Brennan B. J. (1968): Cytopathic effects of parainfluenza virus Type 3 in organ cultures of human respiratory tract tissue. Am J Pathol 52(2): 287-300.
Cristina J., Lopez J. A., Albo C., Garcia-Barreno B., Garcia J., Melero J. A. and Portela A. (1990): Analysis of genetic variability in human respiratory syncytial virus by the RNase A mismatch cleavage method: subtype divergence and heterogeneity. Virology 174(1): 126-134.
Curtis R. A., Van Dreumel A. A. and Ditchfield J. (1966): Infectious bovine rhinotracheitis--clinical, pathological, and virological aspects. Can Vet J 7(8): 161-168.
Cusack P. M., McMeniman N. and Lean I. J. (2003): The medicine and epidemiology of bovine respiratory disease in feedlots. Aust Vet J 81(8): 480-487.
D'Arce R. C., Almeida R. S., Silva T. C., Franco A. C., Spilki F., Roehe P. M. and Arns C. W. (2002): Restriction endonuclease and monoclonal antibody analysis of Brazilian isolates of bovine herpesviruses types 1 and 5. Vet Microbiol 88(4): 315-324.
Dabo S. M., Taylor J. D. and Confer A. W. (2007): Pasteurella multocida and bovine respiratory disease. Anim Health Res Rev 8(2): 129-150.
Davies J. A. and Garrod D. R. (1997): Molecular aspects of the epithelial phenotype. Bioessays 19(8): 699-704.
De Jong M. C., van der Poel W. H., Kramps J. A., Brand A. and van Oirschot J. T. (1996): Quantitative investigation of population persistence and recurrent outbreaks of bovine respiratory syncytial virus on dairy farms. Am J Vet Res 57(5): 628-633.
Devireddy L. R. and Jones C. J. (1999): Activation of caspases and p53 by bovine herpesvirus 1 infection results in programmed cell death and efficient virus release. J Virol 73(5): 3778-3788.
Diamond G., Legarda D. and Ryan L. K. (2000): The innate immune response of the respiratory epithelium. Immunol Rev 173: 27-38.
Dijkman R., Jebbink M. F., Koekkoek S. M., Deijs M., Jonsdottir H. R., Molenkamp R., Ieven M., Goossens H., Thiel V. and van der Hoek L. (2013): Isolation and characterization of current human coronavirus strains in primary human epithelial cell cultures reveal differences in target cell tropism. J Virol 87(11): 6081-6090.
Duncan R. B., Jr. and Potgieter L. N. (1993): Antigenic diversity of respiratory syncytial viruses and its implication for immunoprophylaxis in ruminants. Vet Microbiol 37(3-4): 319-341.
Dungworth D. L. (1993). Infectious diseases of the respiratory system. Pathology of Domestic Animals. K. V. F. Jubb, P. C. Kennedy and N. Palmer. San Diego, CA, Elsevier Science. 2: 613-615.

References 47
Durbin A. P., McAuliffe J. M., Collins P. L. and Murphy B. R. (1999): Mutations in the C, D, and V open reading frames of human parainfluenza virus type 3 attenuate replication in rodents and primates. Virology 261(2): 319-330.
Durham P. J., Hassard L. E. and Van Donkersgoed J. (1991): Serological studies of infectious bovine rhinotracheitis, parainfluenza 3, bovine viral diarrhea, and bovine respiratory syncytial viruses in calves following entry to a bull test station. Can Vet J 32(7): 427-429.
Ebsen M., Mogilevski G., Anhenn O., Maiworm V., Theegarten D., Schwarze J. and Morgenroth K. (2002): Infection of murine precision cut lung slices (PCLS) with respiratory syncytial virus (RSV) and chlamydophila pneumoniae using the Krumdieck technique. Pathol Res Pract 198(11): 747-753.
Elazhary M. A., Silim A. and Dea S. (1984): Prevalence of antibodies to bovine respiratory syncytial virus, bovine viral diarrhea virus, bovine herpesvirus-1, and bovine parainfluenza-3 virus in sheep and goats in Quebec. Am J Vet Res 45(8): 1660-1662.
Eleraky N. Z., Kania S. A., Evermann J. F. and Potgieter L. N. (2003): Comparison of targeting F and G protein genes to detect bovine and ovine respiratory syncytial viruses. J Vet Diagn Invest 15(3): 277-280.
Ellis J. A. (2009): Update on viral pathogenesis in BRD. Anim Health Res Rev 10(2): 149-153.
Ellis J. A. (2010): Bovine parainfluenza-3 virus. Vet Clin North Am Food Anim Pract 26(3): 575-593.
Elvander M. (1996): Severe respiratory disease in dairy cows caused by infection with bovine respiratory syncytial virus. Vet Rec 138(5): 101-105.
Engels M. and Ackermann M. (1996): Pathogenesis of ruminant herpesvirus infections. Vet Microbiol 53(1-2): 3-15.
Engels M., Giuliani C., Wild P., Beck T. M., Loepfe E. and Wyler R. (1986): The genome of bovine herpesvirus 1 (BHV-1) strains exhibiting a neuropathogenic potential compared to known BHV-1 strains by restriction site mapping and cross-hybridization. Virus Res 6(1): 57-73.
Engels M., Palatini M., Metzler A. E., Probst U., Kihm U. and Ackermann M. (1992): Interactions of bovine and caprine herpesviruses with the natural and the foreign hosts. Vet Microbiol 33(1-4): 69-78.
Everard M., Ugonna K., Coleman C. and Plant K. (2009): Respiratory syncytial virus and persistent infection of dendritic cells. Pediatr Infect Dis J 28(5): 450; author reply 450-451.
Fehler F., Herrmann J. M., Saalmuller A., Mettenleiter T. C. and Keil G. M. (1992): Glycoprotein IV of bovine herpesvirus 1-expressing cell line complements and rescues a conditionally lethal viral mutant. J Virol 66(2): 831-839.
Feldman S. A., Audet S. and Beeler J. A. (2000): The fusion glycoprotein of human respiratory syncytial virus facilitates virus attachment and infectivity via an interaction with cellular heparan sulfate. J Virol 74(14): 6442-6447.
Flint S. J., Enquist L. W., Racaniello V. R. and Skalka A. M. (2004). Principles of virology: molecular biology, pathogenesis, and control of animal viruses. Washington, DC, ASM Press.
Fulton R. W., Blood K. S., Panciera R. J., Payton M. E., Ridpath J. F., Confer A. W., Saliki J. T., Burge L. T., Welsh R. D., Johnson B. J. and Reck A. (2009): Lung pathology and infectious agents in fatal feedlot pneumonias and relationship with mortality, disease onset, and treatments. J Vet Diagn Invest 21(4): 464-477.

References 48
Fulton R. W. and Confer A. W. (2012): Laboratory test descriptions for bovine respiratory disease diagnosis and their strengths and weaknesses: gold standards for diagnosis, do they exist? Can Vet J 53(7): 754-761.
Fulton R. W., Downing M. M. and Hagstad H. V. (1982): Prevalence of bovine herpesvirus-1, bovine oral diarrhea, parainfluenza-3, bovine adenoviruses-3 and -7, and goat respiratory syncytial viral antibodies in goats. Am J Vet Res 43(8): 1454-1457.
Fulton R. W., Purdy C. W., Confer A. W., Saliki J. T., Loan R. W., Briggs R. E. and Burge L. J. (2000): Bovine viral diarrhea viral infections in feeder calves with respiratory disease: interactions with Pasteurella spp., parainfluenza-3 virus, and bovine respiratory syncytial virus. Can J Vet Res 64(3): 151-159.
Gagea M. I., Bateman K. G., van Dreumel T., McEwen B. J., Carman S., Archambault M., Shanahan R. A. and Caswell J. L. (2006): Diseases and pathogens associated with mortality in Ontario beef feedlots. J Vet Diagn Invest 18(1): 18-28.
Galen B., Cheshenko N., Tuyama A., Ramratnam B. and Herold B. C. (2006): Access to nectin favors herpes simplex virus infection at the apical surface of polarized human epithelial cells. J Virol 80(24): 12209-12218.
Garber D. A., Beverley S. M. and Coen D. M. (1993): Demonstration of circularization of herpes simplex virus DNA following infection using pulsed field gel electrophoresis. Virology 197(1): 459-462.
Garibaldi R. A., Brodine S., Matsumiya S. and Coleman M. (1985): Evidence for the non-infectious etiology of early postoperative fever. Infect Control 6(7): 273-277.
Geijtenbeek T. B., Kwon D. S., Torensma R., van Vliet S. J., van Duijnhoven G. C., Middel J., Cornelissen I. L., Nottet H. S., KewalRamani V. N., Littman D. R., Figdor C. G. and van Kooyk Y. (2000): DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 100(5): 587-597.
Geraghty R. J., Krummenacher C., Cohen G. H., Eisenberg R. J. and Spear P. G. (1998): Entry of alphaherpesviruses mediated by poliovirus receptor-related protein 1 and poliovirus receptor. Science 280(5369): 1618-1620.
Gerdts V., Beyer J., Lomniczi B. and Mettenleiter T. C. (2000): Pseudorabies virus expressing bovine herpesvirus 1 glycoprotein B exhibits altered neurotropism and increased neurovirulence. J Virol 74(2): 817-827.
Gershwin L. J. (2007): Bovine respiratory syncytial virus infection: immunopathogenic mechanisms. Anim Health Res Rev 8(2): 207-213.
Gershwin L. J., Berghaus L. J., Arnold K., Anderson M. L. and Corbeil L. B. (2005): Immune mechanisms of pathogenetic synergy in concurrent bovine pulmonary infection with Haemophilus somnus and bovine respiratory syncytial virus. Vet Immunol Immunopathol 107(1-2): 119-130.
Gershwin L. J., Schelegle E. S., Gunther R. A., Anderson M. L., Woolums A. R., Larochelle D. R., Boyle G. A., Friebertshauser K. E. and Singer R. S. (1998): A bovine model of vaccine enhanced respiratory syncytial virus pathophysiology. Vaccine 16(11-12): 1225-1236.
Ginisty H., Sicard H., Roger B. and Bouvet P. (1999): Structure and functions of nucleolin. J Cell Sci 112 ( Pt 6): 761-772.
Glezen P. and Denny F. W. (1973): Epidemiology of acute lower respiratory disease in children. N Engl J Med 288(10): 498-505.

References 49
Gonzalez-Reyes L., Ruiz-Arguello M. B., Garcia-Barreno B., Calder L., Lopez J. A., Albar J. P., Skehel J. J., Wiley D. C. and Melero J. A. (2001): Cleavage of the human respiratory syncytial virus fusion protein at two distinct sites is required for activation of membrane fusion. Proc Natl Acad Sci U S A 98(17): 9859-9864.
Goris K., Uhlenbruck S., Schwegmann-Wessels C., Kohl W., Niedorf F., Stern M., Hewicker-Trautwein M., Bals R., Taylor G., Braun A., Bicker G., Kietzmann M. and Herrler G. (2009): Differential sensitivity of differentiated epithelial cells to respiratory viruses reveals different viral strategies of host infection. J Virol 83(4): 1962-1968.
Granzow H., Klupp B. G., Fuchs W., Veits J., Osterrieder N. and Mettenleiter T. C. (2001): Egress of alphaherpesviruses: comparative ultrastructural study. J Virol 75(8): 3675-3684.
Greenberg S. B. (2002): Respiratory viral infections in adults. Curr Opin Pulm Med 8(3): 201-208.
Griffin D. (1997): Economic impact associated with respiratory disease in beef cattle. Vet Clin North Am Food Anim Pract 13(3): 367-377.
Haanes E. J., Guimond P. and Wardley R. (1997): The bovine parainfluenza virus type-3 (BPIV-3) hemagglutinin/neuraminidase glycoprotein expressed in baculovirus protects calves against experimental BPIV-3 challenge. Vaccine 15(6-7): 730-738.
Hage J. J., Schukken Y. H., Dijkstra T., Barkema H. W., van Valkengoed P. H. and Wentink G. H. (1998): Milk production and reproduction during a subclinical bovine herpesvirus 1 infection on a dairy farm. Prev Vet Med 34(2-3): 97-106.
Hage J. J., Vellema P., Schukken Y. H., Barkema H. W., Rijsewijk F. A., van Oirschot J. T. and Wentink G. H. (1997): Sheep do not have a major role in bovine herpesvirus 1 transmission. Vet Microbiol 57(1): 41-54.
Hagglund S., Hjort M., Graham D. A., Ohagen P., Tornquist M. and Alenius S. (2007): A six-year study on respiratory viral infections in a bull testing facility. Vet J 173(3): 585-593.
Hagglund S., Svensson C., Emanuelson U., Valarcher J. F. and Alenius S. (2006): Dynamics of virus infections involved in the bovine respiratory disease complex in Swedish dairy herds. Vet J 172(2): 320-328.
Hardy R. W. and Wertz G. W. (1998): The product of the respiratory syncytial virus M2 gene ORF1 enhances readthrough of intergenic junctions during viral transcription. J Virol 72(1): 520-526.
Harris F. W. and Janzen E. D. (1989): The Haemophilus somnus disease complex (Hemophilosis): A review. Can Vet J 30(10): 816-822.
Heminway B. R., Yu Y. and Galinski M. S. (1994): Paramyxovirus mediated cell fusion requires co-expression of both the fusion and hemagglutinin-neuraminidase glycoproteins. Virus Res 31(1): 1-16.
Henderson G., Zhang Y. and Jones C. (2005): The Bovine herpesvirus 1 gene encoding infected cell protein 0 (bICP0) can inhibit interferon-dependent transcription in the absence of other viral genes. J Gen Virol 86(Pt 10): 2697-2702.
Henjakovic M., Martin C., Hoymann H. G., Sewald K., Ressmeyer A. R., Dassow C., Pohlmann G., Krug N., Uhlig S. and Braun A. (2008): Ex vivo lung function measurements in precision-cut lung slices (PCLS) from chemical allergen-sensitized mice represent a suitable alternative to in vivo studies. Toxicol Sci 106(2): 444-453.

References 50
Hobson L. and Everard M. L. (2008): Persistent of respiratory syncytial virus in human dendritic cells and influence of nitric oxide. Clin Exp Immunol 151(2): 359-366.
Hore D. E. (1966): Isolation of ovine strains of parainfluenza virus serologically related to type 3. Vet Rec 79(16): 466-467.
Horwood P. F., Gravel J. L. and Mahony T. J. (2008): Identification of two distinct bovine parainfluenza virus type 3 genotypes. J Gen Virol 89(Pt 7): 1643-1648.
Hussell T., Baldwin C. J., O'Garra A. and Openshaw P. J. (1997): CD8+ T cells control Th2-driven pathology during pulmonary respiratory syncytial virus infection. Eur J Immunol 27(12): 3341-3349.
Ibricevic A., Pekosz A., Walter M. J., Newby C., Battaile J. T., Brown E. G., Holtzman M. J. and Brody S. L. (2006): Influenza virus receptor specificity and cell tropism in mouse and human airway epithelial cells. J Virol 80(15): 7469-7480.
ICTV (2012a). "International Committee on Taxonomy of Viruses, Classification Herpesvirales." Retrieved 25.02.2014, from http://ictvonline.org/virusTaxonomy.asp.
ICTV (2012b). "International Committee on Taxonomy of Viruses, Classification Mononegavirales." Retrieved 26.02.2014, from http://ictvonline.org/virusTaxonomy.asp.
Iglseder A., Franz S., Benetka V., Mostl K., Latif M., Walk K. and Baumgartner W. (2011): [Diagnosis of bovine viral respiratory diseases]. Schweiz Arch Tierheilkd 153(10): 441-447.
Jericho K. W. and Darcel C. Q. (1978): Response of the respiratory tract of calves kept at controlled climatic conditions to bovine Herpesvirus 1 in aerosol. Can J Comp Med 42(2): 156-167.
Jericho K. W. and Langford E. V. (1978): Pneumonia in calves produced with aerosols of bovine herpesvirus 1 and Pasteurella haemolytica. Can J Comp Med 42(3): 269-277.
Jim K. (2009): Impact of bovine respiratory disease (BRD) from the perspective of the Canadian beef producer. Anim Health Res Rev 10(2): 109-110.
Johnson R. M. and Spear P. G. (1989): Herpes simplex virus glycoprotein D mediates interference with herpes simplex virus infection. J Virol 63(2): 819-827.
Jones A., Morton I., Hobson L., Evans G. S. and Everard M. L. (2006a): Differentiation and immune function of human dendritic cells following infection by respiratory syncytial virus. Clin Exp Immunol 143(3): 513-522.
Jones C. (1998): Alphaherpesvirus latency: its role in disease and survival of the virus in nature. Adv Virus Res 51: 81-133.
Jones C. (2003): Herpes simplex virus type 1 and bovine herpesvirus 1 latency. Clin Microbiol Rev 16(1): 79-95.
Jones C. and Chowdhury S. (2007): A review of the biology of bovine herpesvirus type 1 (BHV-1), its role as a cofactor in the bovine respiratory disease complex and development of improved vaccines. Anim Health Res Rev 8(2): 187-205.
Jones C., Geiser V., Henderson G., Jiang Y., Meyer F., Perez S. and Zhang Y. (2006b): Functional analysis of bovine herpesvirus 1 (BHV-1) genes expressed during latency. Vet Microbiol 113(3-4): 199-210.

References 51
Jones C. D. (1983): Mucociliary clearance from the calf lung. Can J Comp Med 47(3): 265-269.
Juarez Barranco F., Trigo Tavera F. J., Chavez Gris G. and Vargas Garcia R. E. (2003): Viral participation in respiratory disease in feedlot cattle, as identified by immunohistochemistry. Veterinaria Mexico 34: 1-12.
Kaashoek M. J., Rijsewijk F. A. and Oirschot J. T. (1996): Persistence of antibodies against bovine herpesvirus 1 and virus reactivation two to three years after infection. Vet Microbiol 53(1-2): 103-110.
Kaashoek M. J., Rijsewijk F. A., Ruuls R. C., Keil G. M., Thiry E., Pastoret P. P. and Van Oirschot J. T. (1998): Virulence, immunogenicity and reactivation of bovine herpesvirus 1 mutants with a deletion in the gC, gG, gI, gE, or in both the gI and gE gene. Vaccine 16(8): 802-809.
Karger A., Schmidt U. and Buchholz U. J. (2001): Recombinant bovine respiratory syncytial virus with deletions of the G or SH genes: G and F proteins bind heparin. J Gen Virol 82(Pt 3): 631-640.
Karp P. H., Moninger T. O., Weber S. P., Nesselhauf T. S., Launspach J. L., Zabner J. and Welsh M. J. (2002): An in vitro model of differentiated human airway epithelia. Methods for establishing primary cultures. Methods Mol Biol 188: 115-137.
Kendrick J. W., Gillespie J. H. and McEntee K. (1958): Infectious pustular vulvovaginitis of cattle. Cornell Vet 48(4): 458-495.
Kim H. W., Canchola J. G., Brandt C. D., Pyles G., Chanock R. M., Jensen K. and Parrott R. H. (1969): Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am J Epidemiol 89(4): 422-434.
Kimman T. G., Westenbrink F., Schreuder B. E. and Straver P. J. (1987): Local and systemic antibody response to bovine respiratory syncytial virus infection and reinfection in calves with and without maternal antibodies. J Clin Microbiol 25(6): 1097-1106.
Kimman T. G., Zimmer G. M., Westenbrink F., Mars J. and van Leeuwen E. (1988): Epidemiological study of bovine respiratory syncytial virus infections in calves: influence of maternal antibodies on the outcome of disease. Vet Rec 123(4): 104-109.
Kirchhoff J., Uhlenbruck S., Goris K., Keil G. M. and Herrler G. (2014a): Three viruses of the bovine respiratory disease complex apply different strategies to initiate infection. Vet Res 45(1): 20.
Kirchhoff J., Uhlenbruck S., Keil G. M., Schwegmann-Wessels C., Ganter M. and Herrler G. (2014b): Infection of differentiated airway epithelial cells from caprine lungs by viruses of the bovine respiratory disease complex. Vet Microbiol.
Koppers-Lalic D., Reits E. A., Ressing M. E., Lipinska A. D., Abele R., Koch J., Marcondes Rezende M., Admiraal P., van Leeuwen D., Bienkowska-Szewczyk K., Mettenleiter T. C., Rijsewijk F. A., Tampe R., Neefjes J. and Wiertz E. J. (2005): Varicelloviruses avoid T cell recognition by UL49.5-mediated inactivation of the transporter associated with antigen processing. Proc Natl Acad Sci U S A 102(14): 5144-5149.
Kristjansson S., Bjarnarson S. P., Wennergren G., Palsdottir A. H., Arnadottir T., Haraldsson A. and Jonsdottir I. (2005): Respiratory syncytial virus and other respiratory viruses during the first 3 months of life promote a local TH2-like response. J Allergy Clin Immunol 116(4): 805-811.
Krusat T. and Streckert H. J. (1997): Heparin-dependent attachment of respiratory syncytial virus (RSV) to host cells. Arch Virol 142(6): 1247-1254.

References 52
Kupferschmied H. U., Kihm U., Bachmann P., Muller K. H. and Ackermann M. (1986): Transmission of IBR/IPV virus in bovine semen: A case report. Theriogenology 25(3): 439-443.
Lam E., Ramke M., Groos S., Warnecke G. and Heim A. (2011): A differentiated porcine bronchial epithelial cell culture model for studying human adenovirus tropism and virulence. J Virol Methods 178(1-2): 117-123.
Lamb R. A. and Kolakofsky D. (2001). Paramyxoviridae: the virus and their replication. Fields Virology. D. M. Knipe and P. Howley. Philadelphia, Lippincott/ The Williams & Wilkins Co. 1: 1305-1340.
Langedijk J. P., Schaaper W. M., Meloen R. H. and van Oirschot J. T. (1996): Proposed three-dimensional model for the attachment protein G of respiratory syncytial virus. J Gen Virol 77 ( Pt 6): 1249-1257.
Lau S. K., Luthringer D. J. and Eisen R. N. (2002): Thyroid transcription factor-1: a review. Appl Immunohistochem Mol Morphol 10(2): 97-102.
Lehmkuhl H. D. and Cutlip R. C. (1979): Experimentally induced respiratory syncytial viral infection in lambs. Am J Vet Res 40(4): 512-544.
Lehmkuhl H. D. and Gough P. M. (1977): Investigation of causative agents of bovine respiratory tract disease in a beef cow-calf herd with an early weaning program. Am J Vet Res 38(11): 1717-1720.
Lehmkuhl H. D., Smith M. H. and Cutlip R. C. (1980): Morphogenesis and structure of caprine respiratory syncytial virus. Arch Virol 65(3-4): 269-276.
Levine S., Klaiber-Franco R. and Paradiso P. R. (1987): Demonstration that glycoprotein G is the attachment protein of respiratory syncytial virus. J Gen Virol 68 ( Pt 9): 2521-2524.
Liang X., Pyne C., Li Y., Babiuk L. A. and Kowalski J. (1995): Delineation of the essential function of bovine herpesvirus 1 gD: an indication for the modulatory role of gD in virus entry. Virology 207(2): 429-441.
Liang X. P., Babiuk L. A., van Drunen Littel-van den Hurk S., Fitzpatrick D. R. and Zamb T. J. (1991): Bovine herpesvirus 1 attachment to permissive cells is mediated by its major glycoproteins gI, gIII, and gIV. J Virol 65(3): 1124-1132.
Lopez A., Thomson R. G. and Savan M. (1976): The pulmonary clearance of Pasteurella hemolytica in calves infected with bovine parainfluenza-3 virus. Can J Comp Med 40(4): 385-391.
Lutschg V., Boucke K., Hemmi S. and Greber U. F. (2011): Chemotactic antiviral cytokines promote infectious apical entry of human adenovirus into polarized epithelial cells. Nat Commun 2: 391.
Madin S. H., McKercher D. G. and York C. J. (1956): Isolation of the infectious bovine rhinotracheitis virus. Science 124(3225): 721-722.
Malley R., DeVincenzo J., Ramilo O., Dennehy P. H., Meissner H. C., Gruber W. C., Sanchez P. J., Jafri H., Balsley J., Carlin D., Buckingham S., Vernacchio L. and Ambrosino D. M. (1998): Reduction of respiratory syncytial virus (RSV) in tracheal aspirates in intubated infants by use of humanized monoclonal antibody to RSV F protein. J Infect Dis 178(6): 1555-1561.
Mallipeddi S. K., Lupiani B. and Samal S. K. (1996): Mapping the domains on the phosphoprotein of bovine respiratory syncytial virus required for N-P interaction using a two-hybrid system. J Gen Virol 77 ( Pt 5): 1019-1023.

References 53
Markwell M. A. and Paulson J. C. (1980): Sendai virus utilizes specific sialyloligosaccharides as host cell receptor determinants. Proc Natl Acad Sci U S A 77(10): 5693-5697.
Marozin S., Prank U. and Sodeik B. (2004): Herpes simplex virus type 1 infection of polarized epithelial cells requires microtubules and access to receptors present at cell-cell contact sites. J Gen Virol 85(Pt 4): 775-786.
Mars M. H., Bruschke C. J. and van Oirschot J. T. (1999): Airborne transmission of BHV1, BRSV, and BVDV among cattle is possible under experimental conditions. Vet Microbiol 66(3): 197-207.
Mars M. H., de Jong M. C., van Maanen C., Hage J. J. and van Oirschot J. T. (2000): Airborne transmission of bovine herpesvirus 1 infections in calves under field conditions. Vet Microbiol 76(1): 1-13.
Martin C., Uhlig S. and Ullrich V. (2001): Cytokine-induced bronchoconstriction in precision-cut lung slices is dependent upon cyclooxygenase-2 and thromboxane receptor activation. Am J Respir Cell Mol Biol 24(2): 139-145.
Martin S. W., Bateman K. G., Shewen P. E., Rosendal S. and Bohac J. E. (1989): The frequency, distribution and effects of antibodies, to seven putative respiratory pathogens, on respiratory disease and weight gain in feedlot calves in Ontario. Can J Vet Res 53(3): 355-362.
Mastrangelo P. and Hegele R. G. (2013): RSV fusion: time for a new model. Viruses 5(3): 873-885.
Matrosovich M. N., Matrosovich T. Y., Gray T., Roberts N. A. and Klenk H. D. (2004): Human and avian influenza viruses target different cell types in cultures of human airway epithelium. Proc Natl Acad Sci U S A 101(13): 4620-4624.
Matsuoka T., Gale C., Ose E. E. and Berkman R. N. (1966): Calfhood immunization against an experimental Parainfluenza and Pasteurella challenge. Can J Comp Med Vet Sci 30(8): 228-232.
McKercher P. D. (1964): A Comparison of the Viruses of Infectious Bovine Rhinotracheitis (IBR), Infectious Pustular Vulvovaginitis (IPV), and Rinderpest: Part I. Studies of Antigenic Relationships. Can J Comp Med Vet Sci 28(4): 77-88.
McNamara P. S., Flanagan B. F., Hart C. A. and Smyth R. L. (2005): Production of chemokines in the lungs of infants with severe respiratory syncytial virus bronchiolitis. J Infect Dis 191(8): 1225-1232.
McNamara P. S., Flanagan B. F., Selby A. M., Hart C. A. and Smyth R. L. (2004): Pro- and anti-inflammatory responses in respiratory syncytial virus bronchiolitis. Eur Respir J 23(1): 106-112.
McVey D. S. (2009): BRD research needs in the next 10-20 years. Anim Health Res Rev 10(2): 165-167.
Meehan J. T., Cutlip R. C., Lehmkuhl H. D., Kluge J. P. and Ackermann M. R. (1994): Infected cell types in ovine lung following exposure to bovine respiratory syncytial virus. Vet Pathol 31(2): 229-236.
Meng F., Punyadarsaniya D., Uhlenbruck S., Hennig-Pauka I., Schwegmann-Wessels C., Ren X., Durrwald R. and Herrler G. (2013): Replication characteristics of swine influenza viruses in precision-cut lung slices reflect the virulence properties of the viruses. Vet Res 44: 110.
Mesman A. W., de Vries R. D., McQuaid S., Duprex W. P., de Swart R. L. and Geijtenbeek T. B. (2012): A prominent role for DC-SIGN+ dendritic cells in initiation and dissemination of measles virus infection in non-human primates. PLoS One 7(12): e49573.

References 54
Metzler A. E., Matile H., Gassmann U., Engels M. and Wyler R. (1985): European isolates of bovine herpesvirus 1: a comparison of restriction endonuclease sites, polypeptides, and reactivity with monoclonal antibodies. Arch Virol 85(1-2): 57-69.
Meyer G., Hanon E., Georlette D., Pastoret P. P. and Thiry E. (1998): Bovine herpesvirus type 1 glycoprotein H is essential for penetration and propagation in cell culture. J Gen Virol 79 ( Pt 8): 1983-1987.
Miller J. M., Whetstone C. A. and Van der Maaten M. J. (1991): Abortifacient property of bovine herpesvirus type 1 isolates that represent three subtypes determined by restriction endonuclease analysis of viral DNA. Am J Vet Res 52(3): 458-461.
Montgomery R. I., Warner M. S., Lum B. J. and Spear P. G. (1996): Herpes simplex virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor family. Cell 87(3): 427-436.
Müller K. E. (2012): Bedeutung von Stressoren im Komplex der Enzootischen Bronchopneumonie der Kälber und Jungrinder. Tierärztliche Umschau 67: 55-61.
Muylkens B., Meurens F., Schynts F., Farnir F., Pourchet A., Bardiau M., Gogev S., Thiry J., Cuisenaire A., Vanderplasschen A. and Thiry E. (2006): Intraspecific bovine herpesvirus 1 recombinants carrying glycoprotein E deletion as a vaccine marker are virulent in cattle. J Gen Virol 87(Pt 8): 2149-2154.
Muylkens B., Thiry J., Kirten P., Schynts F. and Thiry E. (2007): Bovine herpesvirus 1 infection and infectious bovine rhinotracheitis. Vet Res 38(2): 181-209.
Mweene A. S., Okazaki K. and Kida H. (1996): Detection of viral genome in non-neural tissues of cattle experimentally infected with bovine herpesvirus 1. Jpn J Vet Res 44(3): 165-174.
Nataraj C., Eidmann S., Hariharan M. J., Sur J. H., Perry G. A. and Srikumaran S. (1997): Bovine herpesvirus 1 downregulates the expression of bovine MHC class I molecules. Viral Immunol 10(1): 21-34.
Nguyen D. T., de Witte L., Ludlow M., Yuksel S., Wiesmuller K. H., Geijtenbeek T. B., Osterhaus A. D. and de Swart R. L. (2010): The synthetic bacterial lipopeptide Pam3CSK4 modulates respiratory syncytial virus infection independent of TLR activation. PLoS Pathog 6(8): e1001049.
Nicholas B., Skipp P., Mould R., Rennard S., Davies D. E., O'Connor C. D. and Djukanovic R. (2006): Shotgun proteomic analysis of human-induced sputum. Proteomics 6(15): 4390-4401.
Nixon P., Edwards S. and White H. (1988): Serological comparisons of antigenically related herpesviruses in cattle, red deer and goats. Vet Res Commun 12(4-5): 355-362.
Ogita H. and Takai Y. (2006): Nectins and nectin-like molecules: roles in cell adhesion, polarization, movement, and proliferation. IUBMB Life 58(5-6): 334-343.
Ohlson A., Heuer C., Lockhart C., Traven M., Emanuelson U. and Alenius S. (2010): Risk factors for seropositivity to bovine coronavirus and bovine respiratory syncytial virus in dairy herds. Vet Rec 167(6): 201-206.
Okazaki K., Matsuzaki T., Sugahara Y., Okada J., Hasebe M., Iwamura Y., Ohnishi M., Kanno T., Shimizu M., Honda E. and et al. (1991): BHV-1 adsorption is mediated by the interaction of glycoprotein gIII with heparinlike moiety on the cell surface. Virology 181(2): 666-670.
Paccaud M. F. and Jacquier C. (1970): A respiratory syncytial virus of bovine origin. Arch Gesamte Virusforsch 30(4): 327-342.

References 55
Pertel P. E., Fridberg A., Parish M. L. and Spear P. G. (2001): Cell fusion induced by herpes simplex virus glycoproteins gB, gD, and gH-gL requires a gD receptor but not necessarily heparan sulfate. Virology 279(1): 313-324.
Pezzulo A. A., Starner T. D., Scheetz T. E., Traver G. L., Tilley A. E., Harvey B. G., Crystal R. G., McCray P. B., Jr. and Zabner J. (2011): The air-liquid interface and use of primary cell cultures are important to recapitulate the transcriptional profile of in vivo airway epithelia. Am J Physiol Lung Cell Mol Physiol 300(1): L25-31.
Pickles R. J., McCarty D., Matsui H., Hart P. J., Randell S. H. and Boucher R. C. (1998): Limited entry of adenovirus vectors into well-differentiated airway epithelium is responsible for inefficient gene transfer. J Virol 72(7): 6014-6023.
Pinchak W. E., Tolleson D. R., McCloy M., Hunt L. J., Gill R. J., Ansley R. J. and Bevers S. J. (2004): Morbidity effects on productivity and profitability of stocker cattle grazing in the Southern Plains. J Anim Sci 82(9): 2773-2779.
Prince G. A., Hemming V. G. and Chanock R. M. (1986): The use of purified immunoglobulin in the therapy of respiratory syncytial virus infection. Pediatr Infect Dis 5(3 Suppl): S201-203.
Prozzi D., Walravens K., Langedijk J. P., Daus F., Kramps J. A. and Letesson J. J. (1997): Antigenic and molecular analyses of the variability of bovine respiratory syncytial virus G glycoprotein. J Gen Virol 78 ( Pt 2): 359-366.
Punyadarsaniya D., Liang C. H., Winter C., Petersen H., Rautenschlein S., Hennig-Pauka I., Schwegmann-Wessels C., Wu C. Y., Wong C. H. and Herrler G. (2011): Infection of differentiated porcine airway epithelial cells by influenza virus: differential susceptibility to infection by porcine and avian viruses. PLoS One 6(12): e28429.
Reisinger R. C., Heddleston K. L. and Manthei C. A. (1959): A myxovirus (SF-4) associated with shipping fever of cattle. J Am Vet Med Assoc 135(3): 147-152.
Rice J. A., Carrasco-Medina L., Hodgins D. C. and Shewen P. E. (2007): Mannheimia haemolytica and bovine respiratory disease. Anim Health Res Rev 8(2): 117-128.
Rikitake Y., Mandai K. and Takai Y. (2012): The role of nectins in different types of cell-cell adhesion. J Cell Sci 125(Pt 16): 3713-3722.
Roche W. R., Montefort S., Baker J. and Holgate S. T. (1993): Cell adhesion molecules and the bronchial epithelium. Am Rev Respir Dis 148(6 Pt 2): S79-82.
Rock D., Lokensgard J., Lewis T. and Kutish G. (1992): Characterization of dexamethasone-induced reactivation of latent bovine herpesvirus 1. J Virol 66(4): 2484-2490.
Roizman B. and Knipe D. M. (2001). Herpes Simplex Virus and their replication. Fields Virology. D. M. Knipe and P. Howley. Philadelphia, PA, Lippincott/ The Williams & Wilkins Co. 1: 2399-2461.
Roizman B. and Pellett P. E. (2001). The family Herpesviridae: A brief introduction. Fields Virology. D. M. Knipe and P. Howley. Philadelphia. PA, Lippincott/ The Williams & Wilkins Co. 1: 2381-2398.
Rose M. C. and Voynow J. A. (2006): Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol Rev 86(1): 245-278.
Rudolph K. M., Hunter D. L., Rimler R. B., Cassirer E. F., Foreyt W. J., DeLong W. J., Weiser G. C. and Ward A. C. (2007): Microorganisms associated with a pneumonic epizootic in Rocky Mountain bighorn sheep (Ovis canadensis canadensis). J Zoo Wildl Med 38(4): 548-558.

References 56
Saa L. R., Perea A., Jara D. V., Arenas A. J., Garcia-Bocanegra I., Borge C. and Carbonero A. (2012): Prevalence of and risk factors for bovine respiratory syncytial virus (BRSV) infection in non-vaccinated dairy and dual-purpose cattle herds in Ecuador. Trop Anim Health Prod 44(7): 1423-1427.
Sacco R. E., McGill J. L., Palmer M. V., Lippolis J. D., Reinhardt T. A. and Nonnecke B. J. (2012): Neonatal calf infection with respiratory syncytial virus: drawing parallels to the disease in human infants. Viruses 4(12): 3731-3753.
Salt J. S., Thevasagayam S. J., Wiseman A. and Peters A. R. (2007): Efficacy of a quadrivalent vaccine against respiratory diseases caused by BHV-1, PI3V, BVDV and BRSV in experimentally infected calves. Vet J 174(3): 616-626.
Samal S. K., Zamora M., McPhillips T. H. and Mohanty S. B. (1991): Molecular cloning and sequence analysis of bovine respiratory syncytial virus mRNA encoding the major nucleocapsid protein. Virology 180(1): 453-456.
Sanderson M. W., Dargatz D. A. and Wagner B. A. (2008): Risk factors for initial respiratory disease in United States' feedlots based on producer-collected daily morbidity counts. Can Vet J 49(4): 373-378.
Sauer A. K., Liang C. H., Stech J., Peeters B., Quéré P., Schwegmann-Wessels C., Wu C. Y., Wong C. H. and Herrler G. (2014): Characterization of the Sialic Acid Binding Activity of Influenza A Viruses Using Soluble Variants of the H7 and H9 Hemagglutinins. PLos One.
Schang L. M. and Jones C. (1997): Analysis of bovine herpesvirus 1 transcripts during a primary infection of trigeminal ganglia of cattle. J Virol 71(9): 6786-6795.
Scheid A., Caliguiri L. A., Compans R. W. and Choppin P. W. (1972): Isolation of paramyxovirus glycoproteins. Association of both hemagglutinating and neuraminidase activities with the larger SV5 glycoprotein. Virology 50(3): 640-652.
Schelhaas M., Jansen M., Haase I. and Knebel-Morsdorf D. (2003): Herpes simplex virus type 1 exhibits a tropism for basal entry in polarized epithelial cells. J Gen Virol 84(Pt 9): 2473-2484.
Schlender J., Zimmer G., Herrler G. and Conzelmann K. K. (2003): Respiratory syncytial virus (RSV) fusion protein subunit F2, not attachment protein G, determines the specificity of RSV infection. J Virol 77(8): 4609-4616.
Schreiber P., Matheise J. P., Dessy F., Heimann M., Letesson J. J., Coppe P. and Collard A. (2000): High mortality rate associated with bovine respiratory syncytial virus (BRSV) infection in Belgian white blue calves previously vaccinated with an inactivated BRSV vaccine. J Vet Med B Infect Dis Vet Public Health 47(7): 535-550.
Schrijver R. S., Daus F., Kramps J. A., Langedijk J. P., Buijs R., Middel W. G., Taylor G., Furze J., Huyben M. W. and van Oirschot J. T. (1996): Subgrouping of bovine respiratory syncytial virus strains detected in lung tissue. Vet Microbiol 53(3-4): 253-260.
Schroeder R. J. and Moys M. D. (1954): An acute upper respiratory infection of dairy cattle. J Am Vet Med Assoc 125(933): 471-472.
Schynts F., Lemaire M., Ros C., Belak S. and Thiry E. (2001): Establishment of latency associated with glycoprotein E (gE) seroconversion after bovine herpesvirus 1 infection in calves with high levels of passive antibodies lacking gE antibodies. Vet Microbiol 82(3): 211-222.
Seal B. S., St Jeor S. C. and Taylor R. E. (1985): Restriction endonuclease analysis of bovine herpesvirus 1 DNA and nucleic acid homology between isolates. J Gen Virol 66 ( Pt 12): 2787-2792.

References 57
Sears A. E., McGwire B. S. and Roizman B. (1991): Infection of polarized MDCK cells with herpes simplex virus 1: two asymmetrically distributed cell receptors interact with different viral proteins. Proc Natl Acad Sci U S A 88(12): 5087-5091.
Sheffy B. E. and Davies D. H. (1972): Reactivation of a bovine herpesvirus after corticosteroid treatment. Proc Soc Exp Biol Med 140(3): 974-976.
Shukla D., Liu J., Blaiklock P., Shworak N. W., Bai X., Esko J. D., Cohen G. H., Eisenberg R. J., Rosenberg R. D. and Spear P. G. (1999): A novel role for 3-O-sulfated heparan sulfate in herpes simplex virus 1 entry. Cell 99(1): 13-22.
Singleton R., Dooley L., Bruden D., Raelson S. and Butler J. C. (2003): Impact of palivizumab prophylaxis on respiratory syncytial virus hospitalizations in high risk Alaska Native infants. Pediatr Infect Dis J 22(6): 540-545.
Sinn P. L., Williams G., Vongpunsawad S., Cattaneo R. and McCray P. B., Jr. (2002): Measles virus preferentially transduces the basolateral surface of well-differentiated human airway epithelia. J Virol 76(5): 2403-2409.
Six A., Banks M., Engels M., Bascunana C. R. and Ackermann M. (2001): Latency and reactivation of bovine herpesvirus 1 (BHV-1) in goats and of caprine herpesvirus 1 (CapHV-1) in calves. Arch Virol 146(7): 1325-1335.
Sloots T. P., Whiley D. M., Lambert S. B. and Nissen M. D. (2008): Emerging respiratory agents: new viruses for old diseases? J Clin Virol 42(3): 233-243.
Smith M. H., Frey M. L. and Dierks R. E. (1975): Isolation, characterization, and pathogenicity studies of a bovine respiratory syncytial virus. Arch Virol 47(3): 237-247.
Smith M. H., Lemkuhl H. D. and Phillips S. M. I. (1979): Isolation and characterization of respiratory syncytial virus from goats. American Association of
Veterinary Laboratory Diagnosticians 22: 259-269.
Spear P. G., Eisenberg R. J. and Cohen G. H. (2000): Three classes of cell surface receptors for alphaherpesvirus entry. Virology 275(1): 1-8.
Srikumaran S., Kelling C. L. and Ambagala A. (2007): Immune evasion by pathogens of bovine respiratory disease complex. Anim Health Res Rev 8(2): 215-229.
Steukers L., Vandekerckhove A. P., Van den Broeck W., Glorieux S. and Nauwynck H. J. (2011): Comparative analysis of replication characteristics of BoHV-1 subtypes in bovine respiratory and genital mucosa explants: a phylogenetic enlightenment. Vet Res 42: 33.
Stevenson R. G. and Hore D. E. (1970): Comparative pathology of lambs and calves infected with parainfluenza virus type 3. J Comp Pathol 80(4): 613-618.
Stott E. J. and Taylor G. (1985): Respiratory syncytial virus. Brief review. Arch Virol 84(1-2): 1-52.
Stott E. J., Thomas L. H., Collins A. P., Crouch S., Jebbett J., Smith G. S., Luther P. D. and Caswell R. (1980): A survey of virus infections of the respiratory tract of cattle and their association with disease. J Hyg (Lond) 85(2): 257-270.
Suzuki T., Ikeda K., Koyama N., Hosokawa C., Kogure T., Takahashi T., Jwa Hidari K. I., Miyamoto D., Tanaka K. and Suzuki Y. (2001): Inhibition of human parainfluenza virus type 1 sialidase by analogs of 2-deoxy-2,3-didehydro-N-acetylneuraminic acid. Glycoconj J 18(4): 331-337.

References 58
Takai Y. and Nakanishi H. (2003): Nectin and afadin: novel organizers of intercellular junctions. J Cell Sci 116(Pt 1): 17-27.
Takimoto T., Taylor G. L., Connaris H. C., Crennell S. J. and Portner A. (2002): Role of the hemagglutinin-neuraminidase protein in the mechanism of paramyxovirus-cell membrane fusion. J Virol 76(24): 13028-13033.
Taylor J. D., Fulton R. W., Lehenbauer T. W., Step D. L. and Confer A. W. (2010): The epidemiology of bovine respiratory disease: What is the evidence for predisposing factors? Can Vet J 51(10): 1095-1102.
Tayyari F., Marchant D., Moraes T. J., Duan W., Mastrangelo P. and Hegele R. G. (2011): Identification of nucleolin as a cellular receptor for human respiratory syncytial virus. Nat Med 17(9): 1132-1135.
Techaarpornkul S., Collins P. L. and Peeples M. E. (2002): Respiratory syncytial virus with the fusion protein as its only viral glycoprotein is less dependent on cellular glycosaminoglycans for attachment than complete virus. Virology 294(2): 296-304.
Thiry J., Keuser V., Muylkens B., Meurens F., Gogev S., Vanderplasschen A. and Thiry E. (2006): Ruminant alphaherpesviruses related to bovine herpesvirus 1. Vet Res 37(2): 169-190.
Thompson C. I., Barclay W. S., Zambon M. C. and Pickles R. J. (2006): Infection of human airway epithelium by human and avian strains of influenza a virus. J Virol 80(16): 8060-8068.
Trahman T. and Singh B. (1990): Incidence and pathology of viral pneumonia in goats. Indian J Anim Sci 60: 1159-1162.
Tran L. C., Kissner J. M., Westerman L. E. and Sears A. E. (2000): A herpes simplex virus 1 recombinant lacking the glycoprotein G coding sequences is defective in entry through apical surfaces of polarized epithelial cells in culture and in vivo. Proc Natl Acad Sci U S A 97(4): 1818-1822.
Trudel M., Nadon F., Simard C., Belanger F., Alain R., Seguin C. and Lussier G. (1989): Comparison of caprine, human and bovine strains of respiratory syncytial virus. Arch Virol 107(1-2): 141-149.
Valarcher J. F., Bourhy H., Lavenu A., Bourges-Abella N., Roth M., Andreoletti O., Ave P. and Schelcher F. (2001): Persistent infection of B lymphocytes by bovine respiratory syncytial virus. Virology 291(1): 55-67.
Valarcher J. F., Furze J., Wyld S., Cook R., Conzelmann K. K. and Taylor G. (2003): Role of alpha/beta interferons in the attenuation and immunogenicity of recombinant bovine respiratory syncytial viruses lacking NS proteins. J Virol 77(15): 8426-8439.
Valarcher J. F., Schelcher F. and Bourhy H. (2000): Evolution of bovine respiratory syncytial virus. J Virol 74(22): 10714-10728.
Valarcher J. F. and Taylor G. (2007): Bovine respiratory syncytial virus infection. Vet Res 38(2): 153-180.
Van der Poel W. H., Brand A., Kramps J. A. and Van Oirschot J. T. (1994): Respiratory syncytial virus infections in human beings and in cattle. J Infect 29(2): 215-228.
Van der Poel W. H., Kramps J. A., Middel W. G., Van Oirschot J. T. and Brand A. (1993): Dynamics of bovine respiratory syncytial virus infections: a longitudinal epidemiological study in dairy herds. Arch Virol 133(3-4): 309-321.

References 59
Van der Poel W. H., Langedijk J. P., Kramps J. A., Middel W. G., Brand A. and Van Oirschot J. T. (1995): Bovine respiratory syncytial virus antibodies in non-bovine species. Arch Virol 140(9): 1549-1555.
van Drunen Littel-van den Hurk S. (2006): Rationale and perspectives on the success of vaccination against bovine herpesvirus-1. Vet Microbiol 113(3-4): 275-282.
van Drunen Littel-van den Hurk S., Myers D., Doig P. A., Karvonen B., Habermehl M., Babiuk L. A., Jelinski M., Van Donkersgoed J., Schlesinger K. and Rinehart C. (2001): Identification of a mutant bovine herpesvirus-1 (BHV-1) in post-arrival outbreaks of IBR in feedlot calves and protection with conventional vaccination. Can J Vet Res 65(2): 81-88.
van Oirschot J. T. (1995): Bovine herpesvirus 1 in semen of bulls and the risk of transmission: a brief review. Vet Q 17(1): 29-33.
Van Vuuren M. (1990): Serological studies of bovine respiratory syncytial virus in feedlot cattle in South Africa. J S Afr Vet Assoc 61(4): 168-169.
Vangeel I., Ioannou F., Riegler L., Salt J. S. and Harmeyer S. S. (2009): Efficacy of an intranasal modified live bovine respiratory syncytial virus and temperature-sensitive parainfluenza type 3 virus vaccine in 3-week-old calves experimentally challenged with PI3V. Vet J 179(1): 101-108.
Vareille M., Kieninger E., Edwards M. R. and Regamey N. (2011): The airway epithelium: soldier in the fight against respiratory viruses. Clin Microbiol Rev 24(1): 210-229.
Verhoeff J., Van der Ban M. and van Nieuwstadt A. P. (1984): Bovine respiratory syncytial virus infections in young dairy cattle: clinical and haematological findings. Vet Rec 114(1): 9-12.
Vermeer P. D., McHugh J., Rokhlina T., Vermeer D. W., Zabner J. and Welsh M. J. (2007): Vaccinia virus entry, exit, and interaction with differentiated human airway epithelia. J Virol 81(18): 9891-9899.
Vietmeier J., Niedorf F., Baumer W., Martin C., Deegen E., Ohnesorge B. and Kietzmann M. (2007): Reactivity of equine airways--a study on precision-cut lung slices. Vet Res Commun 31(5): 611-619.
Villenave R., O'Donoghue D., Thavagnanam S., Touzelet O., Skibinski G., Heaney L. G., McKaigue J. P., Coyle P. V., Shields M. D. and Power U. F. (2011): Differential cytopathogenesis of respiratory syncytial virus prototypic and clinical isolates in primary pediatric bronchial epithelial cells. Virol J 8: 43.
Villenave R., Thavagnanam S., Sarlang S., Parker J., Douglas I., Skibinski G., Heaney L. G., McKaigue J. P., Coyle P. V., Shields M. D. and Power U. F. (2012): In vitro modeling of respiratory syncytial virus infection of pediatric bronchial epithelium, the primary target of infection in vivo. Proc Natl Acad Sci U S A 109(13): 5040-5045.
Villenave R., Touzelet O., Thavagnanam S., Sarlang S., Parker J., Skibinski G., Heaney L. G., McKaigue J. P., Coyle P. V., Shields M. D. and Power U. F. (2010): Cytopathogenesis of Sendai virus in well-differentiated primary pediatric bronchial epithelial cells. J Virol 84(22): 11718-11728.
Virtala A. M., Mechor G. D., Grohn Y. T., Erb H. N. and Dubovi E. J. (1996): Epidemiologic and pathologic characteristics of respiratory tract disease in dairy heifers during the first three months of life. J Am Vet Med Assoc 208(12): 2035-2042.
Visalli R. J., Courtney R. J. and Meyers C. (1997): Infection and replication of herpes simplex virus type 1 in an organotypic epithelial culture system. Virology 230(2): 236-243.

References 60
Viuff B., Tjornehoj K., Larsen L. E., Rontved C. M., Uttenthal A., Ronsholt L. and Alexandersen S. (2002): Replication and clearance of respiratory syncytial virus: apoptosis is an important pathway of virus clearance after experimental infection with bovine respiratory syncytial virus. Am J Pathol 161(6): 2195-2207.
Viuff B., Uttenthal A., Tegtmeier C. and Alexandersen S. (1996): Sites of replication of bovine respiratory syncytial virus in naturally infected calves as determined by in situ hybridization. Vet Pathol 33(4): 383-390.
Voynow J. A. and Rubin B. K. (2009): Mucins, mucus, and sputum. Chest 135(2): 505-512.
Welsh R. D., Dye L. B., Payton M. E. and Confer A. W. (2004): Isolation and antimicrobial susceptibilities of bacterial pathogens from bovine pneumonia: 1994--2002. J Vet Diagn Invest 16(5): 426-431.
Werling D., Collins R. A., Taylor G. and Howard C. J. (2002): Cytokine responses of bovine dendritic cells and T cells following exposure to live or inactivated bovine respiratory syncytial virus. J Leukoc Biol 72(2): 297-304.
Wertz G. W., Collins P. L., Huang Y., Gruber C., Levine S. and Ball L. A. (1985): Nucleotide sequence of the G protein gene of human respiratory syncytial virus reveals an unusual type of viral membrane protein. Proc Natl Acad Sci U S A 82(12): 4075-4079.
Whetstone C. A. and Evermann J. F. (1988): Characterization of bovine herpesviruses isolated from six sheep and four goats by restriction endonuclease analysis and radioimmunoprecipitation. Am J Vet Res 49(6): 781-785.
Winkler M. T., Doster A. and Jones C. (1999): Bovine herpesvirus 1 can infect CD4(+) T lymphocytes and induce programmed cell death during acute infection of cattle. J Virol 73(10): 8657-8668.
Winkler M. T., Doster A. and Jones C. (2000): Persistence and reactivation of bovine herpesvirus 1 in the tonsils of latently infected calves. J Virol 74(11): 5337-5346.
Wirth U. V., Gunkel K., Engels M. and Schwyzer M. (1989): Spatial and temporal distribution of bovine herpesvirus 1 transcripts. J Virol 63(11): 4882-4889.
Woods G. T., Sibinovic K. and Marquis G. (1965): Experimental Exposure of Calves, Lambs, and Colostrum-Deprived Pigs to Bovine Myxovirus Parainfluenza-3. Am J Vet Res 26: 52-56.
Wright P. F., Ikizler M. R., Gonzales R. A., Carroll K. N., Johnson J. E. and Werkhaven J. A. (2005): Growth of respiratory syncytial virus in primary epithelial cells from the human respiratory tract. J Virol 79(13): 8651-8654.
Yamakawa Y., Pennelegion C., Willcocks S., Stalker A., Machugh N., Burt D., Coffey T. J. and Werling D. (2008): Identification and functional characterization of a bovine orthologue to DC-SIGN. J Leukoc Biol 83(6): 1396-1403.
Yates W. D. (1982): A review of infectious bovine rhinotracheitis, shipping fever pneumonia and viral-bacterial synergism in respiratory disease of cattle. Can J Comp Med 46(3): 225-263.
Yener Z., Saglam Y. S., Timurkaan N. and Ilhan F. (2005): Immunohistochemical detection of parainfluenza type 3 virus antigens in paraffin sections of pneumonic caprine lungs. J Vet Med A Physiol Pathol Clin Med 52(6): 268-271.
Yesilbag K. and Gungor B. (2009): Antibody prevalence against respiratory viruses in sheep and goats in North-Western Turkey. Trop Anim Health Prod 41(4): 421-425.

References 61
Yewdell J. W. and Brooke C. B. (2012): Monocytes, viruses and metaphors: hanging the Trojan horse. Cell Cycle 11(9): 1748-1749.
Yoon M. and Spear P. G. (2002): Disruption of adherens junctions liberates nectin-1 to serve as receptor for herpes simplex virus and pseudorabies virus entry. J Virol 76(14): 7203-7208.
Yunus A. S., Collins P. L. and Samal S. K. (1998): Sequence analysis of a functional polymerase (L) gene of bovine respiratory syncytial virus: determination of minimal trans-acting requirements for RNA replication. J Gen Virol 79 ( Pt 9): 2231-2238.
Zhang L., Bukreyev A., Thompson C. I., Watson B., Peeples M. E., Collins P. L. and Pickles R. J. (2005): Infection of ciliated cells by human parainfluenza virus type 3 in an in vitro model of human airway epithelium. J Virol 79(2): 1113-1124.
Zhang L., Peeples M. E., Boucher R. C., Collins P. L. and Pickles R. J. (2002): Respiratory syncytial virus infection of human airway epithelial cells is polarized, specific to ciliated cells, and without obvious cytopathology. J Virol 76(11): 5654-5666.
Zimmer G., Budz L. and Herrler G. (2001): Proteolytic activation of respiratory syncytial virus fusion protein. Cleavage at two furin consensus sequences. J Biol Chem 276(34): 31642-31650.
Zimmer G., Rohn M., McGregor G. P., Schemann M., Conzelmann K. K. and Herrler G. (2003): Virokinin, a bioactive peptide of the tachykinin family, is released from the fusion protein of bovine respiratory syncytial virus. J Biol Chem 278(47): 46854-46861.

Appendix 62
6 Appendix
Parts of the thesis have already been published or communicated.
Publications:
Kirchhoff, J., Uhlenbruck, S., Goris, K., Keil, G.M., Herrler, G. Three viruses of the
bovine respiratory disease complex apply different strategies to initiate infection. Vet.
Res. 45(1):20.
Kirchhoff, J., Uhlenbruck, S, Keil, G.M., Schwegmann-Wessels, C., Ganter, M and
Herrler, G. Infection of differentiated airway epithelial cells from caprine lungs by
viruses of the bovine respiratory disease complex. Vet. Microbiol. 170:58-64
Oral presentation:
Kirchhoff, J., Uhlenbruck, S., Keil, G.M. and Herrler, G. Different strategies of host
infection by bovine respiratory viruses. 5th International Congress of Virology, Lyon,
France, 11-14 September 2013.
Poster presentation (selected):
Kirchhoff, J., Uhlenbruck, S., Goris, K. and Herrler, G. Different strategies of host
infection by viruses of the bovine respiratory disease complex. 8th International RSV
Symposium, New Mexico, USA, 27-30 September 2012.
Best Poster Award, 5th Graduate School Day 23/24, November 2012, HGNI

Acknowledgements 63
7 Acknowledgements
I am particularly grateful to Prof. Dr. Georg Herrler for providing this very interesting
topic. Thank you for your continuous advice, encouragement and support throughout
my research and Ph.D. study.
Prof. Dr. Christiane Pfarrer and Prof. Dr. Evgeni Ponimaskin I would like to thank for
interesting discussions and helpful remarks during our meetings and for evaluation of
this thesis. Also, I thank Prof. Dr. Michael Veit for external review of this work.
Furthermore, I thank Prof. Martin Ganter for the collaboration during the goat project,
Prof. Günther Keil for providing recombinant BHV-1, Prof. Klaus Conzelmann for
providing recombinant BRSV and Dr. Wiebke Köhl for generating BRSV-GFP.
I am very grateful to Sabine Uhlenbruck for the assistance and all the good time we
had especially during our excursions. In this context, I also like to thank Günter
Thiem and Holger Mosch for the “shuttle-service”.
I would also like to thank Markus Hoffmann for his help, for sharing his incredible
knowledge and to be patience itself.
Also I like to thank all recent and former members of our institute, in particular Anna,
Ann-Kathrin, Anne, Christel, Christine, Fandan, Yang-Wei, Katarina, Karim, Mareike,
Martina, Moni, Nadine, Nai-Huei, Sarah, Sandra B., Sandra S. (especially), Trust and
Tim. Working with all of you was inspiring and family-like.
At the end I like to thank Tjardo and my family ….for everything; I can´t put into
words.