influence of two daphnia species on summer phytoplankton
TRANSCRIPT

Journal of Plankton Research Vol.19 no.8 pp.1025-1044, 1997
Influence of two Daphnia species on summer phytoplanktonassemblages from eutrophic lakes
Peter H.Kasprzak2 and Richard C.Lathrop1
University ofWisconsin, Madison, Center for Limnology, 680 North Park Street,Madison, W153706, USA
'Present address: Wisconsin Department of Natural Resources, 1350 FemriteDrive, Monona, WI53716, USA2To whom correspondence should be addressed at: Institute of FreshwaterEcology and Inland Fisheries, Miiggelseedamm 310, D-12587 Berlin, FRG
Abstract We conducted grazing experiments to test whether larger-bodied Daphnia puticaria have adifferent effect from smaller-bodied Daphnia galeata mendotae on the composition of summer algalassemblages in eutrophic lakes. Three separate cubitainer experiments were run for 5 days in a repli-cated factorial design utilizing two algal community types and the two Daphnia species. Inorganicphosphorus and nitrogen were added to prevent nutrient limitation of the algae. Both edible andinedible size fractions of chlorophyll a increased in cubitainers without Daphnia spp. Grazer additionusually resulted in a reduction in edible chlorophyll; reductions were greater in D.pulicana cubi-tainers. Grazing by Daphnia spp. on presumed inedible chlorophyll was variable. Algal size was notalways a good predictor of grazeability. The results of this study indicate that D.pulicaria, because ofits greater filtration potential and ability to ingest larger particles, provides a stronger control oninedible-sized algae when compared to equal numerical densities of D.g.mendotae. However, Aphan-izomenon increased as a response to heavy grazing pressure by D.pulicaria on other algal species. Thissuggests that biomanipulation efforts that promote large-bodied Daphnia may not produce desirableresults if nutrient inputs remain high.
Introduction
Inorganic nutrients are of primary importance to the production and biomass ofphytoplankton in standing fresh waters (Ohle, 1953; Rodhe, 1958; Vollenweider,1976; Schindler, 1977). Herbivorous grazing, in contrast, is considered to be oneof the major loss processes of phytoplankton (e.g. Uhlmann, 1954; Nauwerk,1963; McKauley and Kalff, 1981; Pace, 1984; McQueen etaL, 1986; Carpenter andKitchell, 1988; Cyr and Pace, 1992). However, more recent investigations haveshown that the interactions between nutrients, phytoplankton and grazers arelargely influenced by food web structure (Benndorf, 1988; Gulati et al, 1990;Carpenter and Kitchell, 1993; Schindler et al, 1993). Moreover, this relationshipis modified by levels of nutrient loading and in-lake concentrations (Olsen andWillen, 1980; Benndorf, 1990; Elser and Goldman, 1991; Vanni et al, 1992;Koschel et al, 1993). Depending on the degree of nutrient limitation and thestructure of the phytoplankton and herbivore community, grazing may haveeither an inhibiting or an enhancing effect on the algae (Bergquist and Carpen-ter, 1986). Nevertheless, zooplankton filtration rates of ^0.3-0.4 day-1 have beenshown to reduce the biomass in many phytoplankton communities (Reynolds,1984a). Daphnia spp. were often found to be the most powerful filter feederswithin the community of herbivorous zooplankton (Morgan et al, 1980; Sterner,1989; Vanni and Temte, 1990).
© Oxford University Press 1025
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

P.H.Kaspnak and R.CLathrop
Food-uptake mechanisms in planktonic animals are diverse, spanning therange from highly selective to non-selective feeders (Joergensen, 1966). Daphniais considered to be non-selective (Kasprzak etai, 1986) but, even in this genus, anumber of factors modify the resource-consumer relationship (Knoechel andHoltby, 1986a; Lampert, 1987). Gliwicz (1977) has shown that if particles arebeyond the range of availability, a selection is imposed, even on obligate filterfeeders like Daphnia. Nevertheless, whether a particle can be filtered from theambient suspension and subsequently ingested is first a function of its size (Burns,1968). Particles £20 (xm are usually particularly ingestible, whereas those >50-60p,m are not. For the remaining size range of particles (20-50 |xm), ingestibilitylargely depends on the shape of the algae (greatest axial linear dimension;Reynolds, 1984a).
Several authors have shown that filamentous algae, especially blue-green fila-ments, and large colonies of algae such as Microcystis, tend to impair the filtrationpotential of Daphnia (e.g. Webster and Peters, 1978; Gliwicz and Siedlar, 1980;Thompson et ai, 1982; Pearl, 1988). Therefore, Daphnia is not able to reverse thedominance of an established population of larger Microcystis or filamentous blue-green algae (Davidowicz et al, 1988). On the other hand, investigations of Daphniagut contents have sometimes revealed surprisingly large particles, indicating thatthese animals are also able to graze on the so-called 'canopy species' (Horn, 1981;Vyhnalek, 1983; Sommer et al, 1986; Sarnelle, 1992; Epp, 19%). However, differ-ences in grazing potential between small and large Daphnia may be significant.
Most of the experimental work on phytoplankton-grazing interactions wascarried out in oligotrophic or mesotrophic systems (e.g. Lehman and Sandgren,1985; Bergquist and Carpenter, 1986; Elser and Goldman, 1991). This paper,however, concentrates on the grazing influence of Daphnia pulicaria and Daphniagaleata mendotae on summer phytoplankton assemblages of eutrophic lakes.Both Daphnia species are found in many northern eutrophic US lakes. Their rela-tive dominance during the spring clear-water phase and into the early summermonths appears to be controlled by the degree of planktivory (Luecke et al., 1992;Rudstam et al, 1993; Lathrop et al, 1996). While both species have similar cara-pace length, D.pulicaria, with a much greater body mass, only dominates whenplanktivory is low. Promoting the dominance of D.pulicaria (and the similarDaphnia pulex) is often the desired outcome of biomanipulation to reduce thesummer algal problems in lakes (Shapiro et al, 1975; Benndorf, 1988; Gulati etal, 1990; Kitchell, 1992). Our interest was in evaluating the differential grazingimpacts of D.pulicaria and D.g.mendotae on phytoplankton as they develop fromthe clear-water phase into the summer bloom. Special emphasis was placed onthose groups or genera of algae that are considered resistant to grazing. Themajority of these algae may not be avoided except on the basis of size. If they aresmall enough relative to the size of grazers, they are liable to be consumed.
Method
The experimental design followed an idea developed by Lehman and Sandgren(1985). Three separate grazing experiments (Clt Q, C3) were carried out in 20 1
1026
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnia on phytoplankton assemblages
translucent plastic bags (cubitainers) suspended in the littoral zone of LakeMendota at a depth of approximately one-half of incident solar radiation andwater temperatures of ~25°C. Each experiment ran for 5 days in the summer of19% (Table I). Six eutrophic lakes in southeastern Wisconsin, USA, served assources for natural phytoplankton assemblages, D.pulicaria (DpuT) andD.g.mendotae (Dgm). In all three experiments, Pike Lake was selected as a sourceof filamentous algae dominated by Aphanizomenon. Lake Marie, Lake Wingraand Silver Lake were chosen because their phytoplankton was characterized bynon-filamentous forms. Daphnia spp. was taken from Pike Lake, Lake Mendotaand Lake Ripley.
This phytoplankton was exposed to Daphnia grazing at three different stagesin the summer algal development: (i) middle of the clear-water phase; (ii) tran-sition stage between the clear-water phase and summer bloom; (iii) and summerbloom, respectively. In order to remove most of the crustacean zooplankton, thetest water was filtered through a 350 jim screen. Microscopic analysis confirmedthat the composition of phytoplankton was not changed by screening.
To prevent nutrient limitation of the phytoplankton and to exclude the influ-ence of their various mechanisms of nutrient recycling due to grazing, thecubitainers were supplemented with dissolved inorganic phosphorus (DIP,KH2PO4) and nitrogen (DIN, NH4NO3). The nitrogen addition amounted to 300u,g N I"1. The phosphorus addition was 100 \ig P H in experiment Q, and 200 mg
T«We L Schematic representation of the treatments
Exp. no.
c,
Q
c3
Date
15-20.06.%
30.06-0.5.07.96
21-26.08.96
27.08-01.09.96
Phytoplanktonsource lake
Pike Lake
Lake Marie
Pike Lake
Lake Wingra
Pike Lake
Silver Lake
Daphnia source lake
D.pulicaria:Lake MendotaD.g.mendotae:Pike Lake
D.pulicaria:Lake MendotaD.g.mendotae:Pike Lake
D.pulicaria:Lake MendotaD.g.mendotae.Pike Lake
Treatments
C,/Pike/NC,/Pike/N/Dg/TjC,/Pike/N/Dpu/
Q/Marie/NC|/Marie/N/£)gmC,/Marie/N/Dpu/
Q/PikeTNC/Pike/N/Dg/n(VPike/N/Dpu/
Cj/Wingra/NQ/Wingra/N/DgmCj/Wingra/N/Dpu/
CyPike/NCyPike/N/DgmCi/Pike/NIDpul
CySilver/NCySilver/N/Og/nCj/Silver/N/Dpu/
N, nutrients added; Dgm, D.g.mendotae added; Dpul, D.pulicaria added. All experiments were run intriplicates.
1027
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

PJLKaspizak and UGLathrop
P h1 in Cj and C3. DIP and DIN analyses were performed at the State Labora-tory of Hygiene in Madison using widely accepted procedures.
In order to obtain a sufficiently high grazing rate, a Daphnia concentration of-40-50 ind. H was intended. For that reason, the density of the particular popu-lation in each of the Daphnia source lakes was estimated 1 day prior to the experi-ment using a 30 cm diameter Wisconsin plankton net of 350 jim mesh size. Thenext day, an appropriate number of animals was sampled by vertical net hauls,put into a bucket, gently stirred and then added to the cubitainers. Microscopicinspection showed that there were only a few copepods and other cladocerans inthe Daphnia inocula.
After the termination of the three experiments, all cubitainers were individu-ally processed. Daphnia were separated from the water using the same 350 u,mnet, fixed with a buffered formalin-sugar mixture, and stored for later counting.A phytoplankton sample fixed with glutaraldehyde was also taken.
To measure the chlorophyll (Chi) a concentration of the samples, an appropri-ate volume of water was filtered through glass fiber filters (Gelman Scientific) andcold-extracted in methanol for at least 2 weeks. The measurement was madeusing a Turner Model 450 fluorometer. Calibration was performed usingAnacystis nidulans Chi a extracts (Sigma Chemical Co.). Pheopigments were esti-mated following acidification of the samples. Using a screen, edible Chi a was esti-mated in the particle fraction <35 u,m, whereas total Chi a was determined in theuntreated sample. Supposedly inedible Chi a was calculated by subtracting ediblefrom total Chi a.
For the Daphnia treatments, the volume containing -100 individuals wascounted and their size measured. In the nutrient-only treatments, triplicate 25 mlsamples were counted to confirm the low crustacean densities. Daphnia bodymass was calculated according to published length-dry weight relationships(Lynch et ai, 1986).
D.g.mendotae: DW (u-g) = 5.48 L (mm)2300
D.pulicaria: DW (u,g) = 10.67 L (mm)2-093
The Daphnia filtration potential in each cubitainer was calculated using amodel developed by Peters and Downing (1984). The mean maximum size of par-ticles that can be ingested by either D.pulicaria (MPP) or D.g.mendotae (MPG)was calculated according to Burns (1968).
The abundance and size structure (greatest axial linear dimension, GALD;Lehman and Sandgren, 1985) of the phytoplankton were determined using aninverted microscope following the technique of Utermdhl (1958). To get a goodestimate of the number and size distribution of the phytoplankton communityassemblage, the GALD of individual cells, colonies, and filaments was usuallydetermined until the standard error was <10% of the mean GALD. The sizedistribution of algae was estimated in a sample taken from the source lake at thebeginning of the experiment and in the cubitainers at the end of each of theexperiments. Algal identification followed Prescott (1970).
1028
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnia on phytoplankf on assemblages
Results
Nutrients
Excess concentrations of DIP and DIN after the termination of experiment Qindicated that nutrient limitation did not occur (Table II). Concentration differ-ences between nutrient-alone and nutrient-grazer treatments were relativelysmall, indicating that recycling due to grazing and/or uptake by different phyto-plankton biomass were negligible in comparison to the amount of nutrientsadded. In experiment Q, about two-thirds of the added DIP and again one-thirdof the DIN were taken up by the Pike Lake seston. DIP and DIN were unde-tectable in C2/Wingra nutrient-alone treatments, indicating that nutrient limit-ation may have occurred by the end of the experiment. In both Q/Pike andQ/Wingra cubitainers with Daphnia, measurable amounts of DIP and DINremained, most likely as a result of recycling due to grazing or less uptake as aconsequence of lower phytoplankton biomass. In experiment C3, the sameamount of DIP and DIN as in treatment Q was added, but no nutrient measure-ments were made at the end of the experiment.
Daphnia
The number of Daphnia found in the cubitainers of experiments Q , C2 andCyPike was similar within the triplicates as well as between the experimental sets(Figure 1, Table III). However, the abundance of Daphnia spp. in C^Silver was-25 ind. H, which was much lower than intended.
While the numerical densities of the two Daphnia species added to thecubitainers were relatively similar, D.pulicaria biomasses were always muchlarger because of that species' greater weight per unit length. Because individual
Table IL Nutrient addition and concentration in the cubitainer experiments (u.g H)
No. of experiment/nutrient
PhosphorusC,/PikeC|/MarieCVPikeCyWingraCj/PikeCifSilvei
NitrogenC,/PikeC,/MarieCj/PikeCVWingraCyPikeCj/Silver
Add
200200100100200200
300300300300300300
Nut
10413617<2
--
162160160<2
--
Nut/Dp
1041283728
--
137144227103--
Nut/Dg
127154345
--
20419122862
--
add, amount of added nutrients; Nut, concentration in the nutrient treatment; Nut/Dp, concentrationin the Dfidicaria treatment; Nut/Dg, concentration in the D.g.mendotae treatment. Phosphorus wasadded as KH2PO4, nitrogen was added as NH^JC^.
1029
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

PJLKasprzak and R.CLathrop
Pike/Dp Pike/Dg Marie/Dp Marie/Dg
Fig. L Concentration, mean size, individual biomass and community filtration potential of D.pulicaria(Dp) and D.g.mendotae (Dg), respectively, in experiment Q. Sources of the algal assemblages werePike Lake and Lake Marie. Vertical bars are 95% confidence limits.
biomass is one of the major factors in the filtration rate model used, the filtrationpotential in Dgm cubitainers was lower than in Dpul treatments.
Chlorophyll a
In the nutrient-only treatments, nutrient addition was followed by a strongincrease in Chi a relative to concentrations found in each lake at the beginningof the experiments (Figure 2). This response was especially pronounced for edibleChi a, but the inedible fraction also increased to varying degrees. Other thanthese general patterns, the increase in inedible Chi a in CVWingra/N was especi-ally pronounced. In addition, the relatively low concentration of edible Chi a inthis experiment was most likely due to contamination of these cubitainers withD.g.mendotae. With the exception of Q/Wingra/N (Daphnia contamination),there was a clear increase in the edible/inedible Chi a ratio in Q and Q, whereaslittle change was found in C3 (Figure 3).
1030
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022
Influence of two Daphnia on phytoplankton assemblages
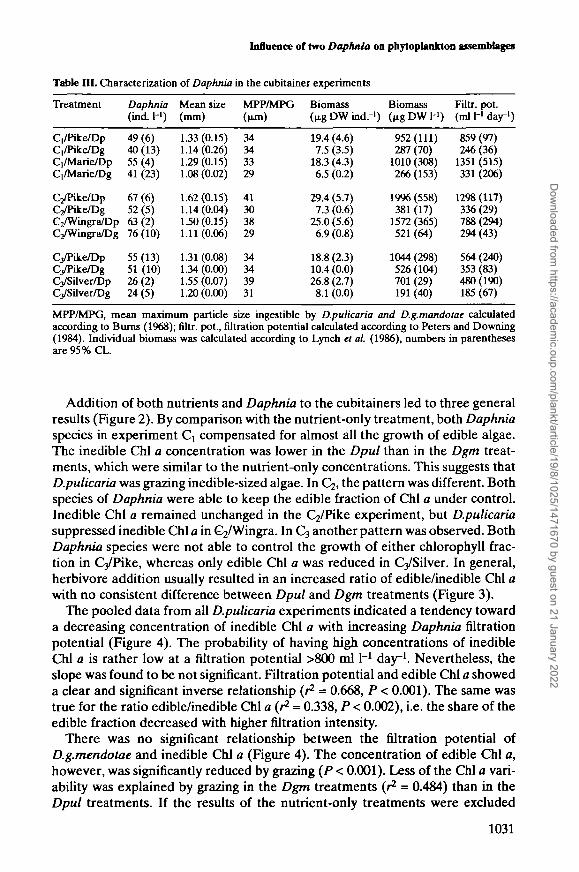
Table IIL Characterization of Daphnia in the cubitainer experiments
Treatment
Q/Pike/DpC,/Pike/DgQ/Marie/DpCi/Marie/Dg
Q/Pike/DpQ/Pike/DgCyWingra/Dp(VWingra/Dg
C3/PikeyDpC3/Pike/DgC3/Silver/DpCySilver/Dg
Daphnia(ind. 1-')
49(6)40(13)55(4)41(23)
67(6)52(5)63(2)76 (10)
55 (13)51 (10)26(2)24(5)
Mean size(mm)
133 (0.15)1.14 (0.26)1.29 (0.15)1.08 (0.02)
1.62 (0.15)1.14 (0.04)130 (0.15)1.11 (0.06)
1.31 (0.08)1.34(0.00)1.55 (0.07)1.20 (0.00)
MPP/MPG((im)
34343329
41303829
34343931
Biomass(tig DW ind."1)
19.4 (4.6)7.5 (3.5)
183 (4.3)63 (0.2)
29.4 (5.7)73 (0.6)
25.0 (5.6)6.9 (0.8)
18.8 (2.3)10.4 (0.0)26.8 (2.7)8.1 (0.0)
BiomassGigDWF)
952(111)287(70)
1010 (308)266 (153)
1996 (558)381 (17)
1572 (365)521 (64)
1044(298)526 (104)701 (29)191 (40)
Filtr. pot.(ml H day1)
859(97)246 (36)
1351 (515)331 (206)
1298 (117)336 (29)788(294)294 (43)
564 (240)353 (83)480(190)185 (67)
MPP/MPG, mean maximum particle size ingestible by D.pulicaria and D.g.mandotae calculatedaccording to Burns (1968); filtr. pot., filtration potential calculated according to Peters and Downing(1984). Individual biomass was calculated according to Lynch et al. (1986), numbers in parenthesesare 95% CL.
Addition of both nutrients and Daphnia to the cubitainers led to three generalresults (Figure 2). By comparison with the nutrient-only treatment, both Daphniaspecies in experiment Q compensated for almost all the growth of edible algae.The inedible Chi a concentration was lower in the Dpul than in the Dgm treat-ments, which were similar to the nutrient-only concentrations. This suggests thatD.pulicaria was grazing inedible-sized algae. In Q, the pattern was different. Bothspecies of Daphnia were able to keep the edible fraction of Chi a under control.Inedible Chi a remained unchanged in the Q/Pike experiment, but D.pulicariasuppressed inedible ChJ a in GyWingra. In C3 another pattern was observed. BothDaphnia species were not able to control the growth of either chlorophyll frac-tion in C3/Pike, whereas only edible Chi a was reduced in C^Silver. In general,herbivore addition usually resulted in an increased ratio of edible/inedible Chi awith no consistent difference between Dpul and Dgm treatments (Figure 3).
The pooled data from all D.pulicaria experiments indicated a tendency towarda decreasing concentration of inedible Chi a with increasing Daphnia filtrationpotential (Figure 4). The probability of having high concentrations of inedibleChi a is rather low at a filtration potential >800 ml h1 day"1. Nevertheless, theslope was found to be not significant. Filtration potential and edible ChJ a showeda clear and significant inverse relationship (r2 = 0.668, P < 0.001). The same wastrue for the ratio edible/inedible Chi a (r2 = 0.338, P < 0.002), i.e. the share of theedible fraction decreased with higher filtration intensity.
There was no significant relationship between the filtration potential ofD.g.mendotae and inedible Chi a (Figure 4). The concentration of edible Chi a,however, was significantly reduced by grazing (P < 0.001). Less of the Chi a vari-ability was explained by grazing in the Dgm treatments (r2 = 0.484) than in theDpul treatments. If the results of the nutrient-only treatments were excluded
1031
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

PJLKasprzak and R.CLathrop
I•§
40
30
20
10-
0- 1 n
I11I rjb1 M
C1
11
30
20
10
0 i n
•11B n
C1
inPike N Dp Dg Marie N Dp Og
4 0
30
20-
10-
0 in
I
II • i t
C2 60
40
20
0 In I .ftC2
1Pike N Dp Dg Wingra N Dp Dg
30
20
10
0I n n
Pike
C3
N Dp DgTreatment
30
20
10
I n I1
C3
1• edibleD inedible
Silver N Dp DgTreatment
Fig. 2. Concentrations of edible and inedible Chi a in experiments C,, Q and C3. Sources of the algalassemblages were Pike Lake, Lake Marie, Lake Wingra and Silver Lake. N, nutrient-only treatments;Dp, Dg, nutrient addition and D.pulicaria or D.g.mendotac addition, respectively Vertical bars are95% confidence limits.
from the regression analysis of Dgm treatments, no significant relationship dueto increased filtration potential was found. Grazing by D.g.mendotae significantlychanged the ratio of edible/inedible Chi a (r2 = 0.3856, P < 0.001).
Algal groups
Some examples of the response of different groups and genera of phytoplanktonto nutrient addition and grazing are summarized in Figures 5 and 6. The sizedistributions of Microcystis colonies in the source lake sample at the beginningof the experiment and in the nutrient-only treatment of CyWingra at the end ofthe experiment were completely different from the mean maximum particle sizes
1032
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnia on phytopbnkton assemblages
O
20%
Pike N Dp Dg Marie N Dp Dg
100%
Pike N Dp Dg
20%
Wingra N Dp Dg
100%
Pike N Dp DgTreatment
Silver N Dp DgTreatment
• edible chla @ inedible chla
Pig. 3. Percentage of edible and inedible Chi a in experiments C, and C3. The *-axis indicates thesource lakes of the phytoplankton assemblages and the treatments. N, nutrients; Dp, nutrients plusD.pulicaria; Dg, nutrient plus D.g.mendotae.
(Burns, 1968) ingestible by D.pulicaria (MPP) or D.g.mendotae (MPG; Figure 5).On the other hand, there was a clear overlap between the size of the colonies andMPP and MPG, respectively, in the Daphnia treatments, indicating that parts ofthe algal population should have been grazeable. Compared to the lake sample,Microcystis concentrations in experiment Q increased as a response to nutrientaddition when no grazers were added. It seems that by grazing, D.pulicaria wasable to compensate for part of this growth, although the differences between thetwo treatments were not significant. However, algal densities were greater in theDgm treatments than in the nutrient-alone treatments. The difference in algaldensities between the Dgm and the Dpul treatments was even more striking.While D.pulicaria suppressed the growth of Microcystis, D.g.mendotae enhancedits growth.
In experiment CySilver, the situation differed (Figure 5). The mean size ofMicrocystis colonies in the lake sample and in the nutrient-only treatments wassmaller than in both Daphnia treatments. Moreover, there were remarkabledifferences in colony size distribution. Only Microcystis in the Dgm treatmentsand MPG were found to overlap. Although variability was high, nutrient addition
1033
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

P.H.Kasprzak and R.GLathrop
Daphnia pulicaria
500 1000 1500 2000filtration potential [ml/1 d]
60
40
20
Daphnia galeata mendotae
040
1 | 30
£ 20
§ 10
0
8
inedible chl-a
edible chl-a
edible/inedible chl-a
100 200 300 400 500filtration potential [ml/1 d]
Fig. 4. Concentration of inedible and edible Chi a and the ratio of both fractions as influenced by thefiltration potential of both D.pulicaria and D.g.mendotae.
again enhanced the growth of the alga, but this time grazing by both D.pulicariaand D.g.mendotae totally compensated for the increase.
Aphanizomenon in Cj/Pike had shorter filaments in both Daphnia treatmentsthan in the lake sample (Figure 5). MPP and MPG overlapped very little with thesize of the algae in the grazer-containing cubitainers. Nutrient addition had no sig-nificant effect on the growth of Aphanizomenon. Daphnia pulicaria was able todecrease the abundance of the Aphanizomenon by -50% of that of the nutrient-only treatments. Daphnia g.mendotae did not influence Aphanizomenon densities.
In Q/Wingra, a similar pattern in size distribution of Aphanizomenon, MPPand MPG was found. Nevertheless, the results in abundance were completelydifferent (Figure 5). Nutrient addition alone had a negative effect on Aphani-zomenon; the species virtually disappeared in the cubitainers. In combinationwith D.pulicaria, however, Aphanizomenon clearly increased in density. Theinfluence of D.g.mendotae was again insignificant.
The size distributions of Ankyra, MPP and MPG in Ci/Pike obviously over-lapped (Figure 6). Moreover, the algae in the grazer cubitainers were muchsmaller compared to those in the nutrient-only treatments. Nutrient additioncaused no effect on the abundance of Ankyra, whereas Daphnia grazing clearlystimulated the alga. Daphnia pulicaria had the strongest enhancing effect.
1034
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnia on phytoplankton assemblages
The size distribution of gelatinous green algae, MPP and MPG overlappedcompletely, as shown by the example of Q/Wingra (Figure 6). Nutrient additionshowed an insignificant increase in algal concentration. Both D.pulicaria as wellas D.g.mendotae decreased algal densities
The size distributions of Melosira, MPP and MPG are completely different(Figure 6). Nutrient addition caused remarkable growth. Only D.pulicariadecreased algal densities. The same pattern was true for Fragilaria in Q/Wingra,except that the reduction by D.g.mendotae was also significant (Figure 6).
Discussion
The addition of nutrients was always followed by an increase in the concentrationof edible Chi a, which is in good agreement with many other field and laboratoryinvestigations (Figure 2; e.g. Schindler and Fee, 1974; Lund and Reynolds, 1982;Koschel and Scheffler, 1985; Bergquist and Carpenter, 1986; Harris, 1986;Schindler, 1990). However, this may not apply to all populations of all species atall times. Species not limited by nutrients will not react to an increased supply(Olsen and Willen, 1980; Vyhnalek, 1983; Lehman and Sandgren, 1985). Nutri-ents also stimulated growth of inedible Chi a, though the enhancement wasusually less pronounced. Larger phytoplankton tend to grow at a lower rate,perhaps because of a lower surface to volume ratio (Banse, 1976; McCauley andBriand, 1979; Reynolds, 1984b). This factor may explain why some investigatorsfound inedible phytoplankton not to benefit greatly from increasing nutrient con-centrations (Watson and McCauley, 1988; Watson et al, 1992). The short dura-tion of the experiments and the restriction of water movement to within thecubitainers may have enhanced this tendency by providing better growing con-ditions to small algae and by selective sedimentation in disfavor of large phyto-plankton (Bloesch and Buergi, 1989).
The increase in inedible Chi a showed remarkable differences between thethree single experiments (Figures 2 and 3). One reason may have been the sizeand the composition of the inoculum. The results indicated a tendency towardshigher concentrations of inedible Chi a when the percentage in the lake sample(source of the inoculum) was high as well.
Another aspect is the seasonal development of natural phytoplankton com-munities in eutrophic lakes. According to the 'PEG-modeP of phytoplankton suc-cession, the period immediately after the clear-water phase is commonlycharacterized by colonial green algae followed in many cases by filamentouscyanobacteria (Sommer et al, 1986). Both of these algal groups are known to berelatively non-digestible, non-ingestible, or both (Porter, 1976; Gliwicz andSiedlar, 1980). The inoculum of Q/Wingra (Figure 6), for instance, had thehighest abundance of gelatinous green algae found in all experiments. Theyresponded to nutrient addition but were also able to withstand high grazing pres-sure. Aphanizomenon was below detection level in the nutrient treatment, butgrew well in the presence of grazers (Figure 5).
For edible Chi a, nutrient addition in the presence of grazers led to the expectedresults in experiments Q and Q (Figures 2 and 3). Daphnia pulicaria and
1035
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

P.H.Kasprzak and R-CLathrop
200 150
lake N Dp Dgtreatment
lake NS Dp MPP Dg MPG
400
lake N Dp MPP Dg MPG
Dg
lake N Dp MPP Dg MPG
N Dp Dgtreatment
Figs 5 and 6. Size distribution (greatest axial linear dimension, GALD) and abundance developmentof groups and genera in the cubitainer experiments. Lake, size distribution in the source lake sampleat the beginning of the experiment; N, size distribution in the nutrient-only treatments at the end ofthe experiment; Dp, size distribution in the nutrients plus D.pulicaria treatments at the end of theexperiment; Dg, size distribution in the nutrient plus D.g.mendotae treatments at the end of theexperiment Vertical bars indicate 1.5 times the standard deviation of the mean. MPP and MPG, meanmaximum ingestible particle size by D.pulicaria and D.g.mendotae, respectively, calculated accordingto Burns (1968). Vertical bars indicate 95% confidence limits.
D.g.mendotae grazed heavily on this fraction. However, this was not true forC3/Pike or, to some extent, for C^Silver. Here again, seasonal differences in thecomposition of edible phytoplankton may explain the differences. Daphniagrazing may cause rapid shifts within the phytoplankton community, resulting inthe dominance of ingestible but non-digestible algae (Kerfoot et aL, 1988).
The influence of Daphnia grazing on inedible Chi a showed almost all combin-ations of possible results. Nevertheless, there seems to be a seasonal trend in the
1036
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnia on phytoplankton assemblages
60T
lake N Dp MPP Dg MPG
lake N Dp Dgtreatment
lake N Dp MPP Dg MPG
lake N Dp Dgtreatment
1000
lake N Dp MPP Dg MPG
lake N Dp Dgtreatment
lake N Dp Dgtreatment
results (Figures 2 and 3). Experiment Q occurred during the clear-water phase.At that time, inedible algae did not grow very much. Consequently, there was notmuch available to Daphnia, although some difference was found between theDpul and Dgm treatment. Later, during experiment Q, which was run near theend of the clear-water phase, inedible algae played a much bigger role, but at leastD.pulicaria was able to offset some of the algal growth. Finally, in experiment C3,which was carried out in the middle of the summer bloom, both daphnids stimu-lated the development of inedible Chi a, possibly through the elimination of com-petition from smaller, faster growing algae. This time-dependent development isalso in agreement with the existing literature (Pace, 1984; Buergi et ai, 1985;Lehman and Sandgren, 1985; Bergquist and Carpenter, 1986; Sommer etaL, 1986;Elser and McKay, 1989; Vanni and Temte, 1990).
1037
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

PJLKaspnak and R.GLathrop
As a general result of this investigation, D.pulicaria was found to have astronger negative influence on the concentration of edible Chi a and on theedible/inedible ratio. A smaller effect was found for D.g.medotae, although theconcentration of animals and their length were similar to the D.pulicaria treat-ments. Therefore, it remains uncertain whether this difference was a property ofthe species or simply a consequence of biomass and allometry (Figures 2-4).
As shown by the length-dry weight relationships, D.g.mendotae at a given sizehas a much smaller individual biomass than D.pulicaria (Lynch et al, 1986).Because biomass is a major factor in the filtration rate model used, higher filtra-tion potential was calculated for the Dpul treatments (Peters and Downing,1984). Although filtration of planktonic animals was found to be notoriously vari-able, size-dependent rate models may provide a basis for comparison (Peters,1983; Knoechel and Holtby, 1986a). However, Lampert (1988) has shownremarkable differences between model calculations and in situ measurements ofcommunity filtration rates (see also Cyr and Pace, 1993).
Burns (1969a), using a suspension of different sized plastic beads, found a verysimilar distribution of particle sizes ingested by both species. Morgan et al. (1980)indicated a maximum ingestible particle size of 35 and 40 u.m for D.g.mendotaeand D.pulicaria, respectively. Kasprzak et al. (1986) found no size-selectivegrazing of D.pulicaria in a mixture of big and small Chlamydomonas geitleri.Burns (1969b), however, refers to higher specific filtration rates in D.g.mendotae.Because she found almost no differences in size-specific body weight between thetwo species, the same is true for the absolute clearance rate. In eutrophic LakeMendota, substantially lower phytoplankton biomass and clearer water occurredin years when D.pulicaria and not D.g.mendotae dominated the crustacean plank-ton (Lathrop and Carpenter, 1992; Lathrop et al, 1996).
Body size is generally considered to be a good predictor of the maximum par-ticle size ingested as well as the filtration rate of cladocerans (Geller, 1975; Hoppand Horn, 1984; Pace, 1984; Schoenberg and Carlson, 1984; Knoechel and Holtby,1986b; Sterner, 1989). The upper limits of ingestible particle size for eitherspecies, calculated according to Burns (1968), indicate maximum differences of 5|im in experiment Q, 10 jim in C2 and 8 jim in C3. Especially in Q, D.pulicariacould take particles larger than 35 fun, the supposed threshold of inedibility,whereas the maximum size ingestible by D.g.mendotae was always lower thanthis. Therefore, different abilities to ingest larger particles may explain at leastsome of the differences found.
Another factor is the metabolic rate, as influenced by the above-mentionedsize-dependent biomass difference in both species. Given a body size of 1 mm,D.g.mendotae has a dry weight of 5.5 u-g compared to 10.7 u.g in D.pulicaria.According to Konoplev et al (1978; quoted in Peters, 1983), this is equivalent toa difference in metabolic rate of ~ 1.71. Because metabolism is in close correlationwith food uptake (Lampert, 1977), the higher metabolic demands in D.pulicariamay be another reason for its greater influence on phytoplankton.
Nevertheless, Reynolds (1984a) indicated community filtration rates >0.3 day1
to be critical for grazeable phytoplankton. Daphnia g.mendotae clearly exceedsthis level; therefore, the missing relationship between filtration potential and Chi
1038
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnia on phytoplankton assemblages
a concentration within the group of grazing treatments (-100-480 ml H day1) isstill somewhat surprising. One reason might be that some small grazeable algaecan grow much faster than 0.3 day1.
The different results of Daphnia grazing concerning the fractions of edible andinedible Chi a are not necessarily reflected at the level of genera and groups(Figures 5 and 6). Compared to copepods, Daphnia is considered to be a non-selective grazer (Joergensen, 1966; DeMott and Moxter, 1991). Therefore, one ofthe major prerequisites making an alga grazeable and ingestible to this group ofanimals is size (Sterner, 1989). Other factors are also important (reviewed byLampert, 1987).
The experiments in Q/Wingra indicate that D.pulicaria had little effect onMicrocystis growth, whereas D.g.mendotae enhanced the development. On thecontrary, the results of CySilver show a strong inhibition (Figure 5). Microcystisis considered to be difficult to ingest because the colonies are usually too big andtoxic strains sometimes inhibit filtration (Thompson etaL, 1982; Fulton and Pearl,1987; Jungmann and Benndorf, 1994). However, Ferguson et al. (1982) occasion-ally found colonies up to a size of 60 u.m in the gut of D.hyalina. As shown inFigure 5, an overlap occurred between the maximum particle size ingested byD.pulicaria (MPP) as well as D.g.mendotae (MPG); therefore, grazing may havesuppressed algal growth. The differences between the outcome of both experi-ments may be explained in terms of nutrient competition (Tilman, 1981). Micro-cystis from Lake Wingra faced a lack of nutrients, as indicated by both theintensive uptake and high Chi a concentrations. The opposite was true for Micro-cystis in Silver Lake (Table II). Consequently, grazing at different levels of nutri-ent limitation may have quite different results.
A similar pattern was found for Aphanizomenon (Figure 5). In Q/Pike, D.puli-caria was able to decrease the concentration significantly (see Epp, 1996).However, nutrient-only addition in CyWingra resulted in a decreased abundanceof Aphanizomenon, whereas in the presence of D.pulicaria an increased concen-tration was found.
For Ankyra, nutrient addition was not sufficient for net growth (Figure 6). BothDaphnia species stimulated Ankyra, although the cell size indicated the speciesto be ingestible. Reynolds et al. (1982), however, found no grazing resistance inthis genus.
Colonial green algae are considered to be ingestible, but non-digestible due totheir gelatinous capsule (Porter, 1976). However, it is unclear whether there is afunctional relationship between the thickness of the mucilage layer and thedegree of protection. The results of our experiments indicate the group to be sus-ceptible to grazing, but it is also able to sustain rather high numbers (Figure 6).
Melosira and Fragilaria were surprisingly inhibited in both Daphnia treat-ments, although there was almost no overlap between the size distribution of thealgae and MPP and MPG, respectively (Figure 6). Nevertheless, Horn (1981) andVyhnalek (1983) found both diatoms in the gut of Daphnia. Dawidowicz et al.(1988) observed Daphnia breaking filaments of blue-green algae in order toingest them, so a similar breaking of the diatoms may have occurred in our experi-ments.
1039
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

P.H.Kaspriak and R.CLalhrop
In summary, nutrient addition always had an enhancing effect on the concen-trations of Chi a, although the edible and inedible fractions reacted differently.The influence of D.pulicaria and D.g.mendotae was not uniform, but indicated aseasonal change in the interaction between phytoplankton and grazers. Daphniapulicaria had a bigger influence on both fractions of Chi a than did D.g.mendo-tae. The results concerning the interaction of both Daphnia species and differentgroups and genera of phytoplankton reflect a variable picture. They indicate thatat least under the conditions of these experiments, remarkably large algae can behandled by Daphnia. On the other hand, species which are considered to begrazeable increased in abundance at rather high filtration potentials.
With respect to biomanipulation as a tool in water quality management, ourresults indicate that large-bodied Daphnia are desirable because of their greaterinfluence on the edible fraction of phytoplankton. Moreover, because themaximum size of particles that can be handled by these larger Daphnia is a func-tion of its body size, the amount of inedible algae being grazed may also increase.Therefore, the duration of the spring clear-water period may be extended becauselarge-bodied D.pulicaria effectively graze those algae typical of the transitionstage between the clear-water phase and the beginning of the summer bloom. Ourresults further indicate that the direct influence of Daphnia grazing on the com-munity structure of summer phytoplankton is rather small. However, the indirectinfluence of large-bodied grazers such as D.pulicaria may be large. Because large-bodied grazers have much greater filtration rates than equal numerical densitiesof small-bodied grazers, small, fast-growing algae are eliminated. Then largeslow-growing algae such as Aphanizomenon can develop especially when nutri-ent supply rates are high. This association between D.pulicaria (or D.pulex) andAphanizomenon has been reported in many eutrophic lakes (Hrbacek, 1964;Lynch, 1980; Ganf, 1983; Andersson and Cronberg, 1984; Pechar and Fort, 1991).Consequently, biomanipulation may not produce substantial improvements inwater clarity in lakes unless nutrient control is also implemented (Reynolds, 1994;Benndorf, 1995; Lathrop et al, 1996).
Acknowledgements
Both authors thank Steve Carpenter and an anonymous reviewer for many helpfulcomments on the draft. This research was funded in part by a grant given to P.H.K.by the German Research Foundation (# Kal206/l-l). Field and laboratory supportwere provided by the North Temperate Lakes Long-Term Ecological Researchprogram and the Wisconsin Department of Natural Recourses. As senior author,I am grateful to the Center for Limnology of the University of Wisconsin, Madison,and its people for accommodating and hosting me for almost a year. My specialthanks to Steve Carpenter for accepting me as a visiting scientist and for a numberof new insights into the field of limnology. Very many thanks to Richard Lathrop,to whom I became friend, for his help in developing and conducting the researchprogram, and for many other little problems that had to be settled during my stayin the USA. Last, but not least, my thanks to Martha Wessels and Sara Mockertfor their help in the field and for teaching me a lively English: 'hang a Louie!'
1040
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnia on phytoplankton assemblages
ReferencesAndersson,G. and Cronberg,G. (1984) Aphanizomenon flos-aquae and large Daphnia: an interesting
plankton association in hypereutrophic water. In Bosheim.S. and Nicholls,M. (eds), Nordisk Lim-nologsymposium, Interactions Between Trophic Levels in Freshwater. Norsk limnologforening,Oslo, Norway, pp. 63-76.
Banse.K. (1976) Rates of growth, respiration and photosynthesis of unicellular algae as related to cellsize—a review. / PhycoL, 12,135-140.
Bergquist,A.M. and Carpenter.S.R. (1986) Limnetic herbivory: effects on phytoplankton and primaryproduction. Ecology, 67,1351-1360.
BenndorfJ. (1988) Objectives and unsolved problems in ecotechnology and biomanipulation: Apreface. Limnologica (Berlin), 19, 5-8.
BenndorfJ. (1990) Conditions for effective biomanipulation; conclusions derived from whole-lakeexperiments in Europe. Hydrobiologia, 200/201,187-203.
BenndorfJ. (1995) Possibilities and limits for controlling eutrophication by biomanipulation. Int. Rev.Ces. HydrobioL, 80, 519-534.
BloeschJ. and Buergi,H.R. (1989) Changes in phytoplankton and zooplankton biomass and compo-sition reflected by sedimentation. LimnoL Oceanogr., 34,113-122.
Buergi,H.R., Weber,P. and Bachmann,H. (1985) Seasonal variations in the trophic structure of phyto-and zooplankton communities in lakes in different trophic states. Schweiz. Z HydroL, 47,197-224.
Burns.C.W. (1968) The relationship between body size of filter-feeding cladocera and the maximumsize of particle ingested. LimnoL Oceanogr., 13, 675-678.
Burns,C.W. (1969a) Particle size and sedimentation in the feeding behavior of two species of Daphnia.Umnol. Oceanogr, 14, 392-^02.
Bums.C.W. (1969b) Relation between filtering rate, temperature, and body size in four species ofDaphnia. LimnoL Oceanogr., 14, 693-700.
Carpenter.S.R. and KitchellJ.F. (1988) Consumer control of lake productivity. BioScience, 38,764-769.
Carpenter.S.R. and KitchellJ.F. (1993) Cascading trophic interactions. In Carpenter.S.R. andKitchellJ.F. (eds), The Trophic Cascade in Lakes. Cambridge University Press, pp. 1-14.
Cyr,H. and Pace,M.L. (1992) Grazing by zooplankton and its relation to community structure. Can.J. Fish. AquaL Set, 49,1455-1465.
Cyr,H. and Pace,M.L. (1993) Allometric theory: extrapolations from individuals to communities.Ecology, 74, 1234-1245.
Dawidowicz.P., Gliwicz,Z.M. and Gulati,R.D. (1988) Can Daphnia prevent an algal bloom in hyper-trophic lakes? A laboratory test. Limnologica (Berlin), 19, 21-26.
DeMott.W.R. and Moxter,F. (1991) Foraging on cyanobacteria by copepods: Responses to chemicaldefenses and resource abundance. Ecology, 72,1820-1843.
ElserJJ. and Goldman.C.R. (1991) Zooplankton effects on phytoplankton in lakes of contrastingtrophic status. LimnoL Oceanogr., 36, 64-90.
ElserJJ. and MacKay,N.A. (1989) Experimental evaluation of effects of zooplankton biomass andsize distribution on algal biomass and productivity in three nutrient limited lakes. Arch. HydrobioL,114,481^*96.
Epp.G.T. (1996) Grazing on filamentous cyanobacteria by Daphnia pulicaria. LimnoL Oceanogr, 41,560-567.
Ferguson,AJ.D., ThompsonJ.M. and Reynolds.C.S. (1982) Structure and dynamics of zooplanktoncommunities maintained in a closed system, with special reference to algal food supply. J. Plank-ton Res., 4,523-543.
Fulton,R.S. and Pearl,H.W. (1987) Effects of colonial morphology on utilization of algal resourcesduring blue-green algal (Microcystis aeruginosa) bloom. LimnoL Oceanogr, 32, 634-644.
Ganf.G.C. (1983) An ecological relationship between Aphanizomenon and Daphnia pulex. AusL J.Mar. Freshwater Res., 34, 755-773.
Geller,W. (1975) Die Nahrungsaufnahme von Daphnia pulex in Abhangigkeit von der Futter-konzentration, der Temperatur, der KorpergrOfSe und dem Hungerzustand der Tiere. Arch. Hydro-bioL, 48(SuppL), 47-107.
Gliwicz^-M. (1977) Food size selection and seasonal succession of filter feeding zooplankton in aeutrophic lake. Ekol Pol., 25,179-225.
Gliwicz»Z.M. and SiedlarJ2- (1980) Food size limitation and algae interfering with food collection inDaphnia. Arch. HydrobioL, 88,155-177.
Gulati,R.D., Lammens,E.H.R.R., Meijer,M.-L. and van DonkJE. (eds) (1990) Biomanipulation: Toolfor Water Management Hydrobiologia, 200/201, 628 pp.
1041
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

RiLKaspnak and R.CLathrop
Harris.G.P. (1986) Phytoplankton Ecology. Structure, Function and Fluctuation. Chapman and Hall,London, pp. 275-290.
Hopp.I. and Horn.W. (1984) Untersuchungen uber die AbhSngigkeit der Filtrier- und Grazingratevon der KOrpergosse planktischer Cladoceren, dargestellt am Beispiel von Daphnia hyalina(Leydig). Limnologica (Berlin), 15,143-148.
Horn,W. (1981) Phytoplankton losses due to zooplankton grazing in a drinking water reservoir. IntRev. Ges. Hydrobiol., 66, 787-810.
HrbacekJ. (1964) Contribution to the ecology of water-bloom-forming blue-gTeen algae: Aphani-zomenon flos-aquac and Microcystis aeruginosa. Verh. Int. Ver. Limnol., 15, 837-846.
Joergensen,C.B. (1966) Biology of Suspension Feeding. Pergamon Press, Oxford, 357 pp.Jungmann£>. and BenndorfJ. (1994) Toxicity of a compound extracted from laboratory and natural
Microcystis spp., and the role of microcystins. Freshwater Biol., 32,13-20.Kasprzak,P., Vyhnalek.V. and Straskraba.M. (1986) Feeding and food selection in Daphnia pulicaria
(Crustacea; Cladocera). Limnologica (Berlin), 17, 309-323.Kerfoot.W.C, Levitan.C. and DeMott.W.R. (1988) Dap/inia-phytoplankton interactions: Density-
dependent shifts in resource quality. Ecology, 69,1806-1825.Kitchell J.F. (1992) Introduction: The rationale and goals for food web management in Lake Mendota.
In KitchellJ.F. (ed.), Food Web ManagemenL A Case Study of Lake Mendota. Springer-Verlag, NewYork, pp. 1-5.
Knoechel,R- and Holtby,L.B. (1986a) Construction and validation of a body-length-based model forthe prediction of cladoceran community filtering rates. UmnoL Oceanogr., 31,1-16.
Knoechel,R- and Holtby,L.B. (1986b) Cladoceran filtering rate: body length relationships for bacterialand large algal particles. Limnol Oceanogr., 31,195-200.
Koschel,R. and Scheffler.W. (1985) The primary production. In Casper,SJ. (ed.), Lake Strechlin. ATemperate Oligotrophic Lake. WJunk Publishers, Dordrecht, pp. 287-322.
Koschel̂ R., Kasprzak,P., KrienitzJL. and RonnebergerJD. (1993) Long-term effects of reduced nutri-ent loading and food-web manipulation on plankton in a stratified Baltic hardwater lake (LakeHaussee, Germany, Feldberg). Verh. Int Ver. Limnol, 26, 647-651.
Lampert.W. (1977) Studies on the carbon balance of Daphnia pulex De Geer as related to environ-mental conditions. II. The dependence of carbon assimilation on animal size, food concentrationand diet species. Arch. Hydrobiol, 48, 310-335.
Lampert.W. (1987) Feeding and nutrition in Daphnia. Mem. 1st Ital Idrobiol., 45,143-192.Lampert,W. (1988) The relationship between zooplankton biomass and grazing: A review. Limno-
logica (Berlin), 19,11-20.Lathrop,R.C. and Carpenter.S.R. (1992) Zooplankton and their relationship to phytoplankton. In
KitchellJJ7. (ed.), Food Web Management. A Case Study of Lake Mendota. Springer-Verlag, NewYork, pp. 127-150.
Lathrop,R.C, Carpenter.S.R. and Rudstam.LG. (1996) Water quality in Lake Mendota since 1900:response to differing levels of nutrients and herbivory. Can. J. Fish. Aquat Set, 53,2250-2261.
LehmanJ.T. and Sandgren.C.D. (1985) Species-specific rates of growth and grazing losses amongfreshwater algae. UmnoL Oceanogr., 30,34-46.
Luecke.G, Rudstam,L.G. and Allen,Y. (1990) Lnterannual pattern of planktivory 1987-89: Analysisof vertebrate and invertebrate planlrtivores. In Kitchell^.F. (ed.), Food Web Management A CaseStudy of Lake Mendota. Springer-Verlag, Berlin, pp. 277c301.
LundJ.W.G. and Reynolds.C.S. (1982) The development and operation of large limnetic enclosuresin Blelham Tarn, English Lake District, and their contribution to phytoplankton ecology. Prog.PhycoL Res., 1,1-65.
Lynch,M. (1980) Aphanizomenon blooms: alternate control and cultivation by Daphnia pulex. InKerfoot.W.C. (ed.), Evolution and Ecology of Zooplankton Communities. University Press of NewEngland, Hanover, NH, pp. 299-304.
Lynch>t, Weider^LJ. and Lampert.W. (1986) Measurement of the carbon balance in Daphnia.Limnol Oceanogr., 31,17-33.
McCauley,E. and Briand,F. (1979) Zooplankton grazing and phytoplankton species richness: Fieldtest of the predation hypothesis. Limnol Oceanogr., 24, 243-252.
McCauley,E. and KalffJ. (1981) Empirical relationships between phytoplankton and zooplankton inlakes. Can. J. Fish. Aquat Set, 38,458-463.
McQueenJ) J., PostJ.R. and Mills,E.L (1986) Trophic relationships in freshwater pelagic ecosystems.Can. J. Fish. Aquat Set, 43,1571-1581.
Morgan,N.C. et al. (1980) Secondary production. In Le CrenJE.D. and Lowe-McConnel,R.H. (eds),The Functioning of Freshwater Ecosystems. Cambridge University Press, Cambridge pp. 247-340.
1042
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

Influence of two Daphnln on phytoplankton assemblages
Nauwerk,A. (1962) Die Beziehungen zwischen dem Zooplankton und dem Phytoplankton im SeeErken. Synth Bot. Ups., 17,163 pp.
Ohle.W. (1953) Phosphor als Initialfaktor der Seeneutrophierung. Jahrb. Wasser, 20,11-23.Olsen .P. and WillenJE. (1980) Phytoplankton response to sewage reduction in Vaettem, a large oligo-
trophic lake in central Sweden. Arch. HydrobioL, 89,171-188.Pace,M. (1984) Zooplankton community structure, but not biomass, influences the phosphorus-
chlorophyll a ratio. Can. J. Fish. AquaL Sci., 41,1089-1096.Pearl,H. (1988) Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. LimnoL
Oceanogr., 33,823-847.Pechar,L. and FottJ. (1991) On the occurrence of Aphanizomenon flos-aquae var.flos-aquae in fish
ponds. InL Rev. Ges. HydrobioL, 76, 57-66.Peters,R.H. (1983) The Ecological Implication of Body Size. Cambridge Studies in Ecology. Cam-
bridge University Press, Cambridge, 329 pp.Peters^RH. and DowningJ.A. (1984) Empirical analysis of zooplankton filtering and feeding rates.
LimnoL Oceanogr., 29, 763-784.Porter,K.G. (1976) Enhancement of algal growth and productivity by grazing zooplankton. Science,
192,1332-1334.Prescott.G.W. (1970) The Freshwater Algae. W.Brown Co., Dubuque, IA, 348 pp.Reynolds,C.S. (1984a) The Ecology of Freshwater Phytoplankton. Cambridge Studies in Ecology.
Cambridge University Press, Cambridge, 384 pp.Reynolds.C.S. (1984b) Phytoplankton periodicity: the interactions of form, function and environ-
mental variability. Freshwater Biol., 14,111-142.Reynolds.C.S. (1994) The ecological basis of successful biomanipulation of aquatic communities.
Arch. HydrobioL, 130,1-33.Reynolds.C.S., ThompsonJ.M., Ferguson.AJ.D. and Wiseman.S.W. (1982) Loss processes in the
population dynamics of phytoplankton maintained in a closed system. /. Plankton Res., 4,561-600.
Rodhe.W. (1958) Primarproduktion und Seentyp. Verh. InL Ver. LimnoL, 13,121-141.Rudstam,L.G., Lathrop.R.C. and Carpenter,S.R. (1993) The rise and fall of a dominant planktivore:
Direct and indirect effects on zooplankton. Ecology, 74, 303-319.Sarnelle.O. (1992) Nutrient enrichment and grazer effects on phytoplankton in lakes. Ecology, 73,
551-560.Schindler.D.W. (1977) Evolution of phosphorus limitation in lakes: natural mechanisms compensate
for deficiencies of nitrogen and carbon in eutrophied lakes. Science, 195, 260-262.Schindler,D.W. (1990) Experimental perturbation of whole lakes as tests of hypotheses concerning
ecosystem structure and function. Oikos, 57, 25-41.Schindler.D.W. and FeeJEJ. (1974) Experimental lakes area: Whole-lake experiments in eutrophica-
tion. / Fish. Res. Board Can., 31,937-953.Schindler.D.E., KitchelU.F., He,X., Carpenter.S.R., HodgsonJ.R. and Kottingham.K.L. (1993) Food
web structure and phosphorus cycling in lakes. Trans. Am. Fish. Soc, 122, 765-772.Schoenberg,S.A. and Carlson^R.E. (1984) Direct and indirect effects of zooplankton grazing on
phytoplankton in a hypertrophic lake. Oikos, 42, 291-302.ShapiroJ., Lamarra.V. and Lynch,M. (1975) Biomanipulation: an ecosystem approach to lake restora-
tion. In Brezonik,P.L. and FoxJ.L. (eds), Proceedings of a Symposium on Water Quality Manage-ment Through Biological ControL University of Florida, Gainesville, FL, pp. 85-96.
Sommer.U., Gliwicz,Z.M., Lampert.W. and Duncan^A. (1986) The PEG-model of seasonal succes-sion of planktonic events in fresh waters. Arch. HydrobioL, 106,433-471.
Stemer,R.W. (1989) The role of grazers in phytoplankton succession. In Sommer.U. (ed.), PlanktonEcology. Succession in Plankton Communities. Springer-Verlag, Berlin, pp. 107-170.
ThompsonJ.M., Ferguson.AJ.D. and Reynolds.C.S. (1982) Natural filtration rates of zooplank-ton in a closed system: The derivation of a community grazing index. / Plankton Res., 4,545-560.
Tilman J). (1981) Tests of resource competition theory using four species of Lake Michigan algae.Ecology, 62, 802-815.
UhlmannJ). (1954) Zur Kenntnis der naturlichen Nahmngswahl von Daphnia magna und Daphniapulex. Z. Fisch., 3, 449-478.
UtennOhl JL (1958) Zur VervoUkommnung der quantitativen Phytoplanktonmethodik. Mia. InL Ver.LimnoL, 9,1-38.
VanniMJ. and Temte J. (1990) Seasonal patterns of grazing and nutrient Limitation of phytoplanktonin a eutrophic lake. LimnoL Oceanogr., 35,697-709.
1043
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022

PJLKasprzafc and R.CLathrop
Vanni,MJ., TemteJ., Allen.Y., Dodds.R., Howard^.R, Leavit.P.R. and Luecke.C. (1992) Herbivory,nutrients, and phytoplankton dynamics in Lake Mendota, 1987-89. In Kitchell J.F. (ed.), Food WebManagement. A Case Study of Lake Mendota. Springer-Verlag, New York, pp. 243-273.
Vollenweider.R.A. (1976) Advances in denning critical loading levels for phosphorus in lake eutroph-ication. Mem. 1st. hal. Idrobiol., 33,53-83.
VyhnaJek.V. (1983) Effects of filter-feeding zooplankton on phytoplankton in fish ponds. Int. Rev.Ges. Hydrobiol., 68, 397-410.
Watson.S. and McCauley,E. (1988) Contrasting patterns of net- and nanoplankton production andbiomass among lakes. Can. J. Fish. Aquat. 5c/., 45, 915-920.
Watson.S., McCauley,E. and DowningJ.A. (1992) Sigmoid relationship between phosphorus, algalbiomass, and algal community structure. Can. J. Fish. Aquat. Sci., 49, 2605-2610.
Webster,K.E. and Peters.R.H. (1978) Some size-dependent inhibitors of larger dadoceran filterers infilamentous suspension. LimnoL Oceanogr., 23,1238-1245.
Received on December 4, 1996; accepted on April 9, 1997
1044
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/8/1025/1471670 by guest on 21 January 2022